Download presentation
Presentation is loading. Please wait.

1
第7章 种内和种间关系 主要的种内相互作用有竞争、自相残杀、性别关系、领域性和社会等级等。
而主要的种间相互作用是竞争、捕食、寄生和互利共生。偶尔,种间相互作用对一方没有影响,而对另一方或有益(偏利共生)或有害(偏害共生)。 第7章 种内和种间关系
或有害(偏害共生)。 第7章 种内和种间关系.")
2
7.1 种内关系 存在于生物种群内部个体间的互相关系称为种内关系。 同种个体间发生的竞争叫做种内竞争。
通过降低拥挤种群个体的适合度,它即可影响基础过程如繁殖力和死亡率,进而调节种群大小,还可使个体产生行为适应来克服或应付竞争,如扩散和领域性。

3
7.1.1 密度效应 7.1.1.1 最后产量衡值法则 Y= ×d = Ki
Donald对三叶草密度和产量的关系作了一系列研究后发现,不管初始播种密度如何,在一定范围内,当条件相同时,植物的最后产量差不多总是一样的。 最后产量衡值法则可表示为: Y= ×d = Ki 其中: 表示植物个体平均重量; d为密度; Y为单位面积产量; Ki是一常数。 三叶草

4
7.1.1.2 —3/2自疏法则 随着播种密度的提高,种内竞争不仅影响到植株生长发育的速度,也影响到植株的存活率。
同样在年龄相等的固着性动物群体中,竞争个体不能逃避,竞争结果典型的也是使较少量的较大个体存活下来。 这一过程叫做自疏。 自疏(self thinning)与-3/2定律: 自疏——指同种植物因种群密度而引起种群个体死亡而密度减少的过程。 –3/2定律——植物种群自疏过程中,其个体平均重量与种群密度成-3/2直 线斜率的变化。 W=Cd-3/2 logw=logc-3/2logd W~平均单株重量 C~为常数 d~种群密度 (植物个体重量与密度说:密度降低,重量增大)
与-3/2定律: 自疏——指同种植物因种群密度而引起种群个体死亡而密度减少的过程。 –3/2定律——植物种群自疏过程中,其个体平均重量与种群密度成-3/2直 线斜率的变化。 W=Cd-3/2 logw=logc-3/2logd W~平均单株重量 C~为常数 d~种群密度 (植物个体重量与密度说:密度降低,重量增大)")
5
7.1.2 性别生态学 研究物种内部性别关系的类型、动态及环境因素对性别的影响是性别生态学(ecology of sex)。
外形与狗、狼相近,体型比狼小,体长100厘米左右,体重10余千克。体毛红棕色或灰棕色,杂有少量具黑褐色毛尖的针毛,腹色较浅。四肢较短。耳短,端部圆钝。尾较长。额部隆起,鼻长,吻部短而宽。全身被毛较短,尾毛略长,尾型粗大,尾端黑色。 豺为典型的山地动物,栖息于山地草原、亚高山草甸及山地疏林中。多结群营游猎生活,性警觉,嗅觉很发达,晨昏活动最频繁。十分凶残,喜追逐,发现猎物后聚集在一起进行围猎,主要捕食狍、麝、羊类等中型有蹄动物。秋季交配,冬季产仔,怀孕期约60天,每胎3~4仔。 由于适应环境自然选择的结果,雌雄外表一致难区分,往往在繁殖期间雄性依靠尿味识别雌性进行交配繁殖。

6
7.1.2.1 两性细胞结合与有性生殖 性别生态学与两个重要的生物学问题有关 一、两性细胞的结合和亲代投入问题
亲代投入是指花费于生产后代和抚养后代的能量 和物质资源。 两性细胞的结合有自体受精和异体受精两种方式。 自体受精指雌雄配子由同一个体产生。 兼备产生雌雄配子的动植物是雌雄同体的,但雌 雄同体的并不一定都是自体受精的。 自我兼容可以视为是防止缺少异体受精的一种保 险措施。 另一种极端是:有些植物有花,但从不开花(是 闭花受精),仅能通过自体受精而生殖。 一个物种可能采取一种或多种受精策略。 孟德尔性状遗传 豌豆闭花受精
,仅能通过自体受精而生殖。 一个物种可能采取一种或多种受精策略。 孟德尔性状遗传 豌豆闭花受精.")
7
二、寻找为什么大多数生物都营有性繁殖的答案
因为无性繁殖较有性生殖在进化选择上有下列重要优越性: ①可迅速增殖,占领暂时性新栖息地; ②母体所产的后代都带有母体的整个基因组,因此给下代复制的基因组是有 性繁殖的两倍。 有性繁殖要在进化选择上处于有利地位,必须使之获得利益超过所偿付的减 数分裂价、基因重组价和交配价。 一般认为,有性繁殖是对生存在多变和易遭不测环境下的一种适应性。因为 有性生殖混合或重组了双亲的基因组,导致产生遗传上易变的配子,并转而 产生遗传上易变的后代。遗传新物质的产生,使受自然选择作用的种群的遗 传变异保持高水平,使种群在不良环境下至少能保证少数个体生存下来,并 获得繁殖后代的机会。 美国生态家T.H.Hamilton提出了一种假说:营有性繁殖的物种之间的竞争和 捕食者—猎物间相互作用是使有性繁殖持续保持的重要因素。

8
7.1.2.2 性比 性比通常以种群中雄体对雌体的相对数来表示,如雌雄个体数相等,性比即为1:1。性比也可以用雄体占种群总数的比列来表示,如雌雄数相等,其比例为0.5。大多数生物种群的性比倾向于1:1。这种倾向的进化原因叫做Fisher氏性比理论。如果母体偏向于生产性别较少的后代,母体的适合度就比较高。这就是稀少型有利的例子。Fisher氏性比理论认为,雌雄两体应该有相等投入,这是稀少型有利的结果。 在拥挤的条件下,雌体间的局域资源竞争很紧张,因而产出雌仔并离开家区是很有利的,这样可使生殖成效有更高的提高机遇。在同胞姊妹间存在交配竞争的情况下,母体如果产同样数量雄仔和雌仔就会形成浪费,因而性比偏于雌,这叫做局域交配竞争,该现象在许多无脊椎动物中出现。

9
性比(Sex ratio)是反映种群中雄性个体(♂)和雌性个体(♀)比例的参数。受精卵的♂与♀比例,大致是50:50,这是第一性比,幼体成长到性成熟这段时间里,由于种种原因,♂与♀的比例变化,至个体开始性成熟为止,♂与♀的比例叫做第二性比,此后,还会有成熟的个体性比叫第三性比。 研究年龄结构和性比的意义:有助于了解种群的发展趋势,预测种群的兴衰;有助于指导生产或合理开发生物资源;对于人口年龄结构的研究,则是经济计划的依据;也可用来计算依靠比例。

10
7.1.2.3 性选择 1.植物的选择受精 选择受精(selective fertilization)是指具有特定遗传基础的精核与卵细胞优先受精的现象。 选择受精主要表现为生理生化和遗传上的特征,包括自交不亲和性、远缘杂交、不亲和性、多个花粉精核间的竞争等现象。 植物的选择受精的生物学意义: (1)可保证最适应的两性细胞的高度融合,从而增强后代的存活能力; (2)限制异种之间的自由交配,使种间生殖隔离,从而保证各个种的相对稳定性。
可保证最适应的两性细胞的高度融合,从而增强后代的存活能力; (2)限制异种之间的自由交配,使种间生殖隔离,从而保证各个种的相对稳定性。")
11
2.动物的性选择 (1)雌性动物的婚配选择: 在两性间对后代投入的差别较大,为接近高投入性别(一般是此雌
性者)低投入性别(一般是雄性)者之间的竞争也就越激烈;高投入性别者的更加挑剔,也必然从低投入性别者那里获得更好的代价。简言之,雄性应该是有进攻性的,雌性应该是挑剔性的。雌性精心选择那些携带最好基因型的雄性个体交配,来获得高质量的后代,提高其繁殖成效。为此,雌性动物往往对雄性个体有敏锐的洞察力,特别对色彩和声音有较高的鉴别力。此外,对雄性的体态、行为特征(如争斗、给饵等)等也有一定的鉴别力,从中择优选择,才能保证后代健康。 (2)性选择可能通过两条途径产生性内选择,即通过同性成员间的配偶竞争(性内选择intrasexual selection ),或通过偏爱异性的某个独特特征(性间选择intersexual selection ),或者两条途径兼而有之。性内选择可以解释打斗武器的发生,如雄性哺乳动物的鹿角、洞角、獠牙、大犬齿。另一方面, 性间选择对极乐鸟、孔雀等雄鸟明显无用的软 件,如奢侈的尾和头羽等提供了解释。 让步赛 理论认为,拥有质量好的大尾(或其他奢侈的 特征),表明拥有者必须有好的基因,而弱个 体不可能忍受这种能量消耗,也加大了奢侈特 征者被捕食的敏感性。可供选择的Fisher氏私 奔模型认为,雄性这种诱惑性特征开始恣意的 雌性所选择,并将继续进化,如果雌性基因对 挑选特征(如选大尾)编码,雄性也会对该特 征(如尾的大小)编码,形成明显的雌雄二形(sexual dimorphism)现象。
低投入性别(一般是雄性)者之间的竞争也就越激烈;高投入性别者的更加挑剔,也必然从低投入性别者那里获得更好的代价。简言之,雄性应该是有进攻性的,雌性应该是挑剔性的。雌性精心选择那些携带最好基因型的雄性个体交配,来获得高质量的后代,提高其繁殖成效。为此,雌性动物往往对雄性个体有敏锐的洞察力,特别对色彩和声音有较高的鉴别力。此外,对雄性的体态、行为特征(如争斗、给饵等)等也有一定的鉴别力,从中择优选择,才能保证后代健康。 (2)性选择可能通过两条途径产生性内选择,即通过同性成员间的配偶竞争(性内选择intrasexual selection ),或通过偏爱异性的某个独特特征(性间选择intersexual selection ),或者两条途径兼而有之。性内选择可以解释打斗武器的发生,如雄性哺乳动物的鹿角、洞角、獠牙、大犬齿。另一方面, 性间选择对极乐鸟、孔雀等雄鸟明显无用的软. 件,如奢侈的尾和头羽等提供了解释。 让步赛. 理论认为,拥有质量好的大尾(或其他奢侈的. 特征),表明拥有者必须有好的基因,而弱个. 体不可能忍受这种能量消耗,也加大了奢侈特. 征者被捕食的敏感性。可供选择的Fisher氏私. 奔模型认为,雄性这种诱惑性特征开始恣意的. 雌性所选择,并将继续进化,如果雌性基因对. 挑选特征(如选大尾)编码,雄性也会对该特. 征(如尾的大小)编码,形成明显的雌雄二形(sexual dimorphism)现象。")
13
7.1.2.4 植物的性别系统 同株异花:有些植物种的个体具有雌雄两类花,雄花产生花粉,雌花产生胚珠。
雌雄异株:雌雄花分别长在不同的植株上。 银杏,银杏科,银杏属。落叶乔木,又名公孙树、鸭脚子、白果树等。树干高大,雌雄异株,现世界上权存一属一种,为中国特有树种。从外部形态上也可以区分出来。雄株笔直,雌株宽大 。

14
7.1.2.5 动物的婚配制度 婚配制度是指种群内婚配的种种类型,包括婚配的数目、配偶持续时间以及对后代的抚育等。
单配制(monogamy) :出现在一雄与一雌结成配偶对,或者只在生殖季节,或者保持到有一个死亡。 一雄多雌制(polygamy) :是最普遍的婚配制度。一雄多雌出现在一个雄体与数个或许多雌体交配时。 决定动物婚配制度的主要生态因素可能是资源的分布,主要是食物和营巢地在空间和时间上的分布情况。
 :出现在一雄与一雌结成配偶对,或者只在生殖季节,或者保持到有一个死亡。 一雄多雌制(polygamy) :是最普遍的婚配制度。一雄多雌出现在一个雄体与数个或许多雌体交配时。 决定动物婚配制度的主要生态因素可能是资源的分布,主要是食物和营巢地在空间和时间上的分布情况。")
15
表 婚配制度的生态学分类 (引自 Smith, 1980)
单配偶制 雌雄两性都只能独占一个异性 一雄多制 雄性占有多个雌性 保卫资源型一雄多雌制 雄性间接控制雌性,通过占有最重要资源-食物 保卫雌性型一雄多雌制 雄性直接控制雌性,由于雌性有集群性 雌性优势型一雄多雌制 在繁殖季节中,雄性集中格斗,雌性选择雄性配偶 暴发性繁殖集会 雌雄两性的繁殖期短促而同步,性比接近1,性选择低 求偶集会(lek) 雄性在集会中为争夺优势地位而进行直接竞争 高速多窝型一雄多雌制 在短期中生产多窝,雌雄两性各自分别伏窝孵卵 一雌多雄制 雌性占有多个雄性 保卫资源型一雌多雄制 雌性间接控制雄性,通过占有最重要资源-食物 雌性控制型一雌多雄制 雌性不保卫雄性所需资源,但雌性之间相互作用抑制某些雌性与雄性交配。 表 婚配制度的生态学分类 (引自 Smith, 1980)
 雄性在集会中为争夺优势地位而进行直接竞争. 高速多窝型一雄多雌制. 在短期中生产多窝,雌雄两性各自分别伏窝孵卵. 一雌多雄制. 雌性占有多个雄性. 保卫资源型一雌多雄制. 雌性间接控制雄性,通过占有最重要资源-食物. 雌性控制型一雌多雄制. 雌性不保卫雄性所需资源,但雌性之间相互作用抑制某些雌性与雄性交配。 表 婚配制度的生态学分类 (引自 Smith, 1980)")
16
7.1.3 领域性和社会等级 领域(territory) :是指由个体,家庭或其他社群单位所占据的并积极保卫不让同种其他成员侵入的空间。
保卫领域的方式有很多,如以鸣叫、气味标志或特异的姿势向入侵者宣告其领域范围,或以威胁、直接进攻驱赶入侵者等,这些行为称为领域行为。 具领域性(territoriality)的种类在脊椎动物中最多,尤其是鸟兽。 东北虎、非洲狮具有领域行为,可达十几平方千米。 动物生存需要一定的空间,这个空间就是动物生存的领域,在这个领域中,动物可以取食、繁殖、抚育后代。领域虽然没有明确的界限,但是领域的占有者却熟知它的边界。领域的占有者通常使用姿态、气味、鸣叫等方式来警告周围的动物,以保护自己的领域。
的种类在脊椎动物中最多,尤其是鸟兽。 东北虎、非洲狮具有领域行为,可达十几平方千米。 动物生存需要一定的空间,这个空间就是动物生存的领域,在这个领域中,动物可以取食、繁殖、抚育后代。领域虽然没有明确的界限,但是领域的占有者却熟知它的边界。领域的占有者通常使用姿态、气味、鸣叫等方式来警告周围的动物,以保护自己的领域。")
17
在动物领域性的研究中,总结出以下几条规律:
1、领域面积随其占有者的体重而扩大,领域大小必须以能保证供 应足够的食物资源为前提,动物越大,需要资源越多,领域面积 也就越大。 2、领域面积受食物品质的影响,食肉动物的领域面积较同等体重的食草动物 大,且体重越大,这种差别也越大。 3、领域面积和行为往往随生活史,尤其是繁殖节律而变化。 社会等级(social hierarchy) :是指动物种群中各个动物的地位具有一定顺序的等级现象。 等级形成的基础是支配行为,或称支配-从属关系。 社会等级的优越性包括优势个体在食物、栖所、配偶选择中均有优先权 。 领域性和社会等级是两类重要的社会性行为,与种群调节有密切联系。
 :是指动物种群中各个动物的地位具有一定顺序的等级现象。 等级形成的基础是支配行为,或称支配-从属关系。 社会等级的优越性包括优势个体在食物、栖所、配偶选择中均有优先权 。 领域性和社会等级是两类重要的社会性行为,与种群调节有密切联系。")
18
7.1.4 他感作用 他感作用:也称作异株克生,通常指一种植物通过向体外分泌代谢过程中的化学物质,对其他植物产生直接或间接的影响。
这种作用是生存斗争的一种特殊形式,种间、种内关系都有此现象。 他感作用中植物分泌物称克生物质,对克生物质的提取、分离和鉴定已做了许多工作。 他感作用具有重要的生态学意义: 1、对农林业生产和管理具有重要意义。 2、他感作用对植物群落的种类组成有重要影响,是造成种类成分对群落的选择性以及某种植物的出现引起另一类消退的主要原因之一。 3、他感作用是引起植物群落演替的重要内在因素之一。

19
7.2 种间关系 种间关系包括竞争、捕食、互利共生等,是构成生物群落的基础。
所以种间关系的研究是种群生态学与群落生态学之间的界面,其研究内容主要包括两个方面: ①两个或多个物种在种群动态上的互相影响,即相互动态; ②彼此在进化过程和方向上的相互作用,即协同进化。

20
7.2.1 种间竞争 种间竞争:是指两物种或更多物种共同利用同样的有限资源时产生的相互竞争作用。

21
7.2.1.1 竞争类型及其一般特征 竞争关系是指两物种或更多物种共同利用同样的有限资源时而产生的相互竞争作用。
1)竞争排斥原理(Principle of competitive exclusion) 在一个稳定的环境内,两个以上受资源限制的、但具有相同资源利用方式的种,不能长期共存在一起,也即完全的竞争者不能共存。 2)竞争的类型 竞争有两种作用方式,或仅通过损耗有限的资源(利用性竞争),而个体不直接相互作用,或通过竞争个体间直接的相互作用(干扰性竞争)。他感作用也是一种典型的相互干扰性竞争。 利用竞争是指一种生物所利用的资源对另一种生物来说也非常重要,即两种生物同时竞争利用同一种资源。 干扰竞争是指生物借助行为排除另一种生物,使其得不到资源。
竞争排斥原理(Principle of competitive exclusion) 在一个稳定的环境内,两个以上受资源限制的、但具有相同资源利用方式的种,不能长期共存在一起,也即完全的竞争者不能共存。 2)竞争的类型. 竞争有两种作用方式,或仅通过损耗有限的资源(利用性竞争),而个体不直接相互作用,或通过竞争个体间直接的相互作用(干扰性竞争)。他感作用也是一种典型的相互干扰性竞争。 利用竞争是指一种生物所利用的资源对另一种生物来说也非常重要,即两种生物同时竞争利用同一种资源。 干扰竞争是指生物借助行为排除另一种生物,使其得不到资源。")
22
3)竞争的一个共同特点 竞争结果的不对称性是种间竞争的一个共同特点。一个体的竞争代价常远高于另一个体。竞争杀死失败者是很普通的,或通过掠夺资源(使他们丧失资源)或通过干扰(直接杀伤或毒害它们)。竞争不对称的例子大大超过对称性结果的例子。 种间竞争的另一个共同特点是对一种资源的竞争,能影响对另一种资源的竞争结果。例如植物间的竞争,冠层中占优势的植物,减少了竞争对手进行光合作用所需的阳光辐射。这种对阳光的竞争也影响植物根部吸收营养物质和水分的能力。也就是说,在植物的种间竞争中,根竞争与枝竞争之间有相互作用。

23
4)竞争模型——Lotka-Volterra模型
式中:α是种2的一个个体对种1的阻碍系数(竞争系数) β是种1的一个个体对种2的阻碍系数 两物种的竞争结局从理论上讲可有以下三种: (1) 种1胜而种2被排除; (2) 种2胜而种1被排除; (3) 两种共存。
 β是种1的一个个体对种2的阻碍系数. 两物种的竞争结局从理论上讲可有以下三种: (1) 种1胜而种2被排除; (2) 种2胜而种1被排除; (3) 两种共存。")
24
(a) N1取胜, N2灭亡 (b) N2取胜, N1灭亡
(c) 不稳定共存(两种都有可能取胜) (d)稳定的平衡(两种共存)
 不稳定共存(两种都有可能取胜) (d)稳定的平衡(两种共存)")
25
7.2.1.2生态位理论 生态位(niche):是生态学中的一个重要概念,指物种在生物群落或生态系统中的地位和角色。
由Grinell 在1917年最早提出,Elton在1927年再定义,Hutchinson提出n-维生态位的概念。 J.Grinnell认为生态位是一个种所占有的微环境,强调的是空间生态位的概念。C.Elton将生态位看作是“物种在生物群落或生态系统中的地位与功能作用”。G.E.Hutchinson提出n-维生态位的概念,使生态位理论取得明显进展。另外,Hutchinson还提出了基础生态位与实际生态位的概念。 美国学者R.H.Whittaker认为,生态位是每个种在一定生境的群落中都有不同于其他种的自己的时间、空间位置,也包括在生物群落中的功能地位,并指出生态位的概念与生境和分布区的概念是不同的。 生境是指生物生存的周围环境,分布区是指种分布的地理范围,生态位则说明在一个生物群落中某个种群的功能地位。 竞争物种在资源利用分化上的临界阈值叫做极限相似性。

26
栖息地(habitat): 有机体所处的物理环境。栖息地一般包括许多生态位并支持许多物种。
生态位空间(niche space) Hutchinson(1958) 提出 影响有机体的每个条件,和有机体能够利用的每个资源都可以被认为是一个轴或维(dimension),在此轴或维上,可以定义有机体将出现的一个范围。同时考虑一系列这样的维,就可以得到有机体生态位的一个增强了的定义图,称为生态位空间。 基础生态位:物种能够栖息的理论上的最大空间。 实际生态位:物种能够占据的生态位空间。(由竞争和捕食胁迫造成,互利共生可扩大实际生态位)。
 Hutchinson(1958) 提出. 影响有机体的每个条件,和有机体能够利用的每个资源都可以被认为是一个轴或维(dimension),在此轴或维上,可以定义有机体将出现的一个范围。同时考虑一系列这样的维,就可以得到有机体生态位的一个增强了的定义图,称为生态位空间。 基础生态位:物种能够栖息的理论上的最大空间。 实际生态位:物种能够占据的生态位空间。(由竞争和捕食胁迫造成,互利共生可扩大实际生态位)。")
27
生态位分化 三个共存物种的资源利用曲线。 图生态位狭,相互重叠少; (b) 图生态位宽,相互重叠多。 d 为曲线峰值间的距离,w为曲线的标准差
 图生态位宽,相互重叠多。 d 为曲线峰值间的距离,w为曲线的标准差")
28
竞争排斥(Competive exclusion)原理陈述,共存只能在物种生态位分化的稳定,均匀环境中发生。如在以下情况下,则不会出现竞争排斥:
①不稳定的环境中,物种之间不能达到平衡; ②栖息在没有资源竞争的环境中的物种之间; ③在一物种被排挤掉之前,环境的改变使竞争方向改变。 竞争排斥原理的反对派认为,许多亲缘很近的种类往往在同一栖息地中生活在一起,进行野外采集工作的每一个人,都会感到在同一地方栖息着许多昆虫,那么怎样来协调实验培养中近缘种经常被排挤掉和自然群落中有许多物种共存的事实呢?在回答这个问题中有两种简单的观点。一种认为,自然界中很少见到竞争,即物种很少为有限资源而竞争。因此,在自然群落中没有必要去找竞争排斥的证据。另一种观点认为,自然界中。竞争十分普遍,它是群落中支配物种进化发展的主要因素。那么,自然界中,竞争究竟是普遍的还是少见的?许多研究工作集中在亲缘接近的种类,并认为这些种类间最可能有激烈竞争。

29
拉克相信,进一步深入研究,能发现5-7种之间的区别。
拉克研究了同一水流取食,同一峭壁上营巢的两种鸬鸶(Ptalacrocorax Carbo和P.aristotelis),经过深入的观察证明,后者主要以水上层自由游泳的鱼类和沙鳗为食;前者主要捕食底栖的比目鱼和底栖无脊椎动物。换言之,它们的营养生态位是有区别的,拉克还对英国的全部鸣禽同亲缘接近的鸟类进行了分析,结果如下: 3 18或更多 4 5 2 5-7 地理 隔离 栖息地隔离 食性隔离 因个体大小不同而分离 因不同越冬地区而分离 明显的生态位重叠 鸟对数 所有这些实事说明,那些在同样的环境中能够共存的近缘种, 其生态位有明显区别的,竞争由此而减少。 拉克相信,进一步深入研究,能发现5-7种之间的区别。
,经过深入的观察证明,后者主要以水上层自由游泳的鱼类和沙鳗为食;前者主要捕食底栖的比目鱼和底栖无脊椎动物。换言之,它们的营养生态位是有区别的,拉克还对英国的全部鸣禽同亲缘接近的鸟类进行了分析,结果如下: 3. 18或更多 地理 隔离. 栖息地隔离. 食性隔离. 因个体大小不同而分离. 因不同越冬地区而分离. 明显的生态位重叠. 鸟对数. 所有这些实事说明,那些在同样的环境中能够共存的近缘种, 其生态位有明显区别的,竞争由此而减少。 拉克相信,进一步深入研究,能发现5-7种之间的区别。")
30
7.2.1.3 竞争释放和性状替换 在缺乏竞争者时,物种会扩张其实际生态位。这种竞争释放(Competive release)可认为是在野外竞争作用的证据。 如:在北以色列,两种沙鼠(Gerbillus allenbyi和Meriones tristrami)在一定范围内重叠。在重叠处,G.allebyi只发现在非沙性土中,而在只有G.allenbyi的地方它既占据沙性土也占据非沙性土。似乎在没有M.tristrami的情况下,G.allenbyi能够扩张其生态位。 偶尔,竞争产生的生态位收缩会导致形态性 状变化,叫做性状替换( Character displacement),就是指两个亲缘关系密切 的种类若在异域性分中,它们的特征往往很 相似,甚至难以区别。但在同域性分布中, 它们之间的区别就明显,彼此之间必须出 现明显的生态分离。
在一定范围内重叠。在重叠处,G.allebyi只发现在非沙性土中,而在只有G.allenbyi的地方它既占据沙性土也占据非沙性土。似乎在没有M.tristrami的情况下,G.allenbyi能够扩张其生态位。 偶尔,竞争产生的生态位收缩会导致形态性. 状变化,叫做性状替换( Character. displacement),就是指两个亲缘关系密切. 的种类若在异域性分中,它们的特征往往很. 相似,甚至难以区别。但在同域性分布中, 它们之间的区别就明显,彼此之间必须出. 现明显的生态分离。")
31
7.2.1.4 种间竞争与空间和时间异质性 竞争者之间经常变化的竞争平衡最初是作为对“浮游生物悖论”现象的解释而提出的。该现象指的是在海上层结构简单的生境中持续存在大量浮游生物种类。日周期性和季节性持续变化的环境,伴随着温度、光、氧气和营养物的变化,会排斥达成任何种间平衡。 在许多环境中会不可预测地产生缝隙,起因于极端天气或死亡。在斑块生境中,许多情况下竞争优势者取决于哪个个体最先占在该地点,即优先权效果。 斑块=生境斑块:局部种群所占据的空间区域。 异质种群:通过个体扩散相互联系的局部种群的集合。 局部种群:同一个种的,并且以很高的概率相互作用的个体的集合。

32
7.2.2 捕食作用 捕食(predation):可定义为一种生物摄取其他生物个体的全部或部分为食,前者称为捕食者(predator),后者称为被食者(prey)。 这一广泛的定义包括:1、“典型的捕食”,他们在袭击猎物后迅速杀死而食之; 2、食草,他们逐渐杀死对象生物,且只消费对象个体的一部分; 3、寄生,它们与单一对象个体(寄主)有密切关系,通常生活在寄生的组织中。昆虫中的拟寄生者(Parasitoids),例如寄生蜂,它们与真寄生虫者(如血吸虫)的区别是总要杀死其宿主。 4、同类相食(Cannibalism),也是捕食现象的一种特例,只不过是捕食者与被食者是同一特种而已。 捕食者也可分为以植物组织为食的食草动物(herbivores) ,以动物组织为食的食肉动物(carnivores)以及以动植物两者为食的杂食动物(omnivores) 。 同时两种类型的被捕食者都有保护自己的身体结构设置和对策,植物主要利用化学防御,而动物则形成一系列行为对策。 捕食者的食物变化很大,一些捕食者是食物选择性非常强的特化种,仅摄取一种类型的猎物,而另一些是泛化种,可以吃几种类型的猎物。 草食性动物一般比食肉动物更加特化(或是只吃一种类型食物的单食者),或是以少数几种食物为食的寡食者,它们集中摄取具有相似防御性化学物质的很少几种植物。而草食性动物中的泛化种(或广食者),可通过避免取食毒性更大的部分或个体,而以一定范围的植物种类为食。动植物寄生者都是特化种。
有密切关系,通常生活在寄生的组织中。昆虫中的拟寄生者(Parasitoids),例如寄生蜂,它们与真寄生虫者(如血吸虫)的区别是总要杀死其宿主。 4、同类相食(Cannibalism),也是捕食现象的一种特例,只不过是捕食者与被食者是同一特种而已。 捕食者也可分为以植物组织为食的食草动物(herbivores) ,以动物组织为食的食肉动物(carnivores)以及以动植物两者为食的杂食动物(omnivores) 。 同时两种类型的被捕食者都有保护自己的身体结构设置和对策,植物主要利用化学防御,而动物则形成一系列行为对策。 捕食者的食物变化很大,一些捕食者是食物选择性非常强的特化种,仅摄取一种类型的猎物,而另一些是泛化种,可以吃几种类型的猎物。 草食性动物一般比食肉动物更加特化(或是只吃一种类型食物的单食者),或是以少数几种食物为食的寡食者,它们集中摄取具有相似防御性化学物质的很少几种植物。而草食性动物中的泛化种(或广食者),可通过避免取食毒性更大的部分或个体,而以一定范围的植物种类为食。动植物寄生者都是特化种。")
33
7.2.2.1 捕食者与猎物 捕食者与猎物的相互关系是经过长期的协同进化逐步形成的。捕食者进化了一整套适应性特征如锐齿、利爪、尖喙、毒牙等工具,诱饵追击、集体围猎等方式,以更有力地捕食猎物。另一方面。猎物也形成了一系列行为对策,如保护色、警戒色、拟态、假死、快跑、集体抵御等以逃避被捕食。自然选择对于捕食者在于提高发现、捕获和取食猎物的效率,而对于猎物在于提高逃避,防止被捕食的效率,显然这两种选择是对立的。 协同进化(coevolution) 一个物种的性状作为对另一物种性状的反应而进化,而后一物种的这一性状本身又是作为对前一物种性状的反应而进化的。 捕食者与猎物协同进化的结果常常是使有害的“负作用” 减弱。
 一个物种的性状作为对另一物种性状的反应而进化,而后一物种的这一性状本身又是作为对前一物种性状的反应而进化的。 捕食者与猎物协同进化的结果常常是使有害的 负作用 减弱。")
34
(1)捕食者与猎物的协同进化
捕食者与猎物的协同进化")
36
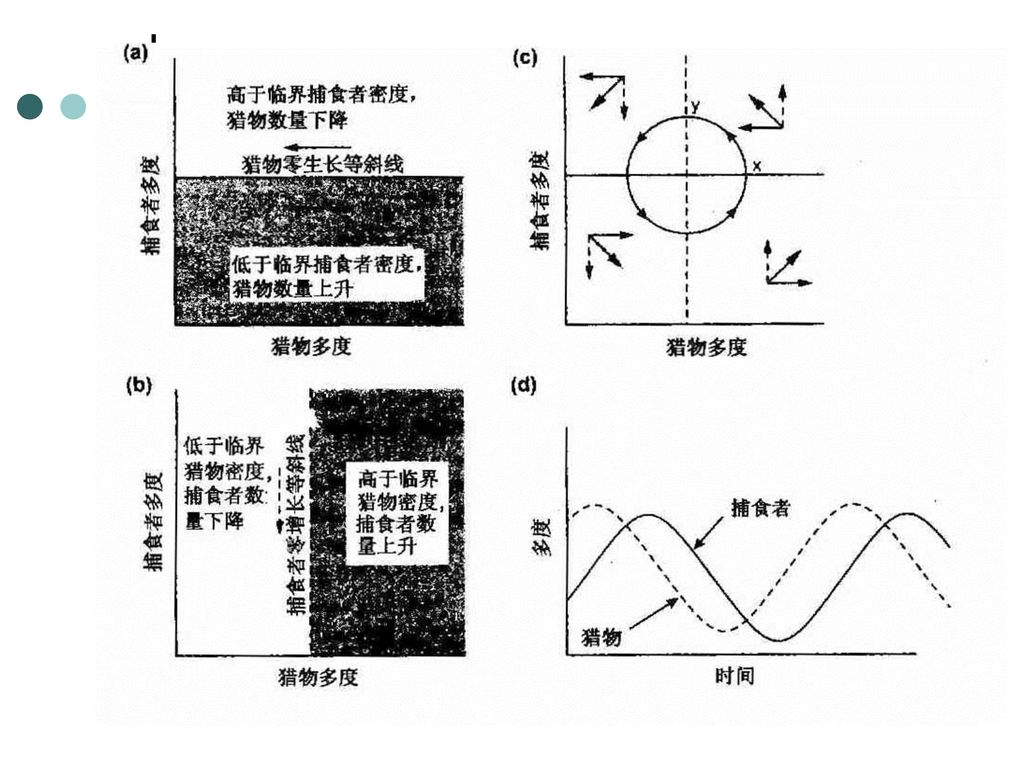
(2)猎物—捕食者的简单模型 该方程是以第二和第三营养级, 特别是小型动物捕食关系为基础的。 式中:N是猎物密度; a是猎物的内禀增长率; α为猎物与捕食者相遇被吃掉的几率; P是捕食者密度, b为捕食者的内禀增长率; Β是捕食者攻击效率(捕食能力)的测度。
的测度。")
39
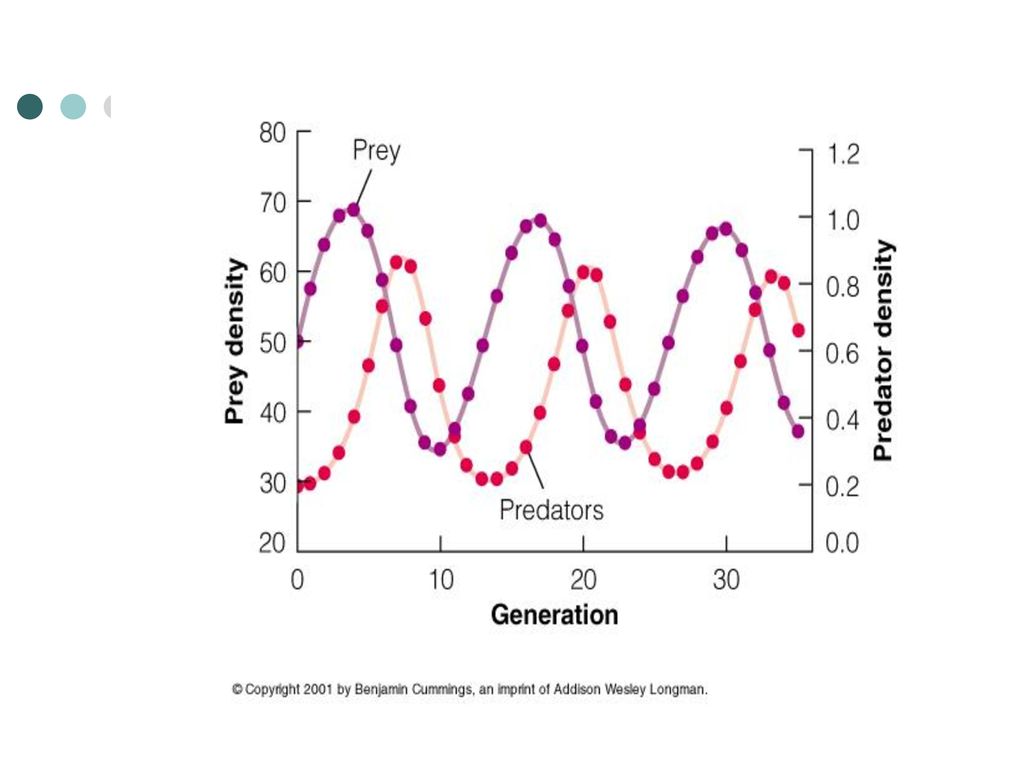
兔子与猞猁 的种群震荡

40
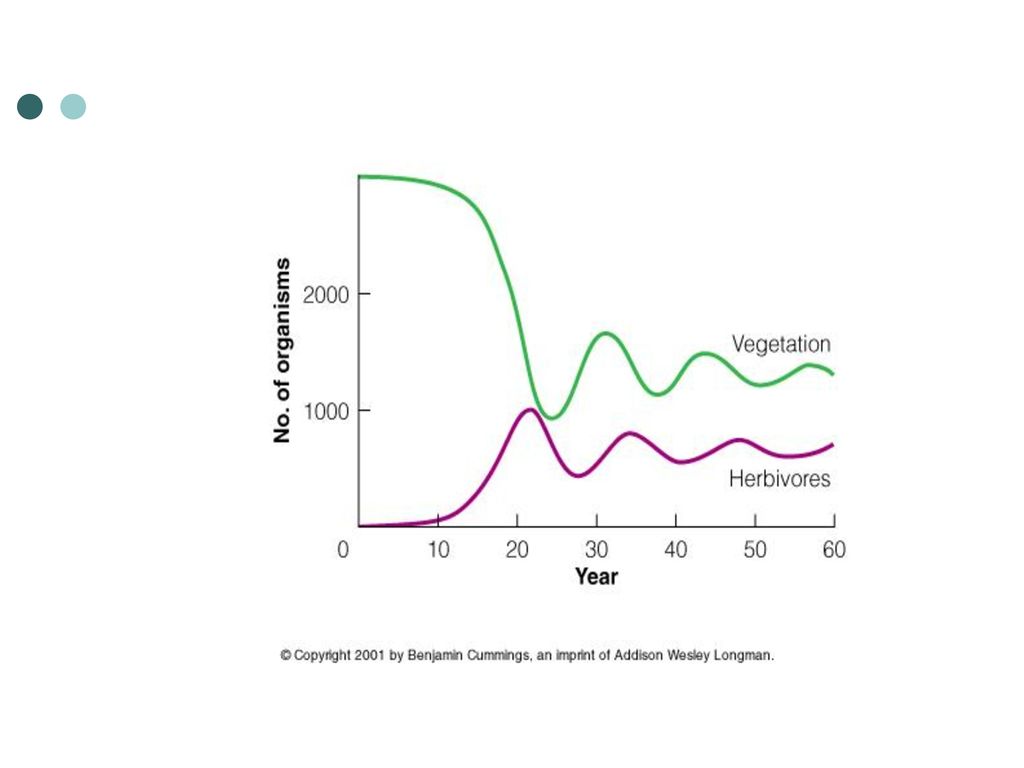
(3)自然界中捕食者对猎物种群大小的影响 目前有两种主要观点: ①任一捕食者的作用,只占猎物种总死亡率的很小一部分,因此去除捕食者对猎物种仅有微弱影响。如许多捕食者捕食田鼠,蛇仅是捕食者之一,所以去除蛇对田鼠种群数量影响不大。 ②捕食者只是利用了对象种中超出环境所能支持的部分个体,所以对最终猎物种群大小没有影响。这时限制猎物种群的主要因素不是捕食者数量,而是其他因素 。

41
捕食对猎物数量影响的一些例子: 实验 结果 1 将鸭捕食者(狐狸、浣熊、獾和条纹鼬)从营巢区域除去 2 一些区域去除狐狸,另一些区域去除小型食肉动物 3 澳洲野犬的控制和排除 1 鸭营巢密度增加300%,筑巢成功率增加50% 2 北美野兔数量在去除狐狸的情况下增加300%,但在去除小型食肉动物的情况下数量没有变化 3 十多种中型哺乳动物数量增加,野猪数量大增

42
7.2.2.2 食草作用 食草是广义捕食的一种类型。其特点是植物不能逃避被食,而动物对植物的危害只是使部分机体受损害,留下部分能够再生。
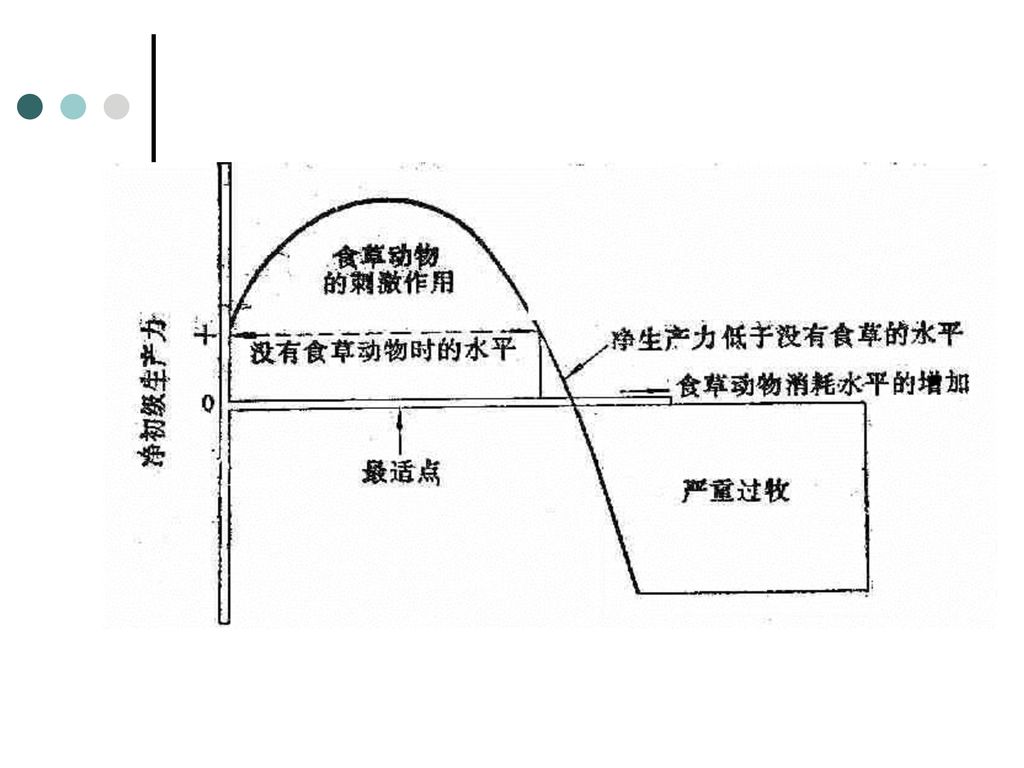
植物被“捕食”而受损害的程度随损害部位、植物发育阶段的不同而异。 1) 协同进化 植物的补偿作用(生长补偿) 与防卫反应(机械防御或化学防御) 植物主要以两种方式来保护自己免遭捕食:①毒性与差的味道; ②防御结构。 植物—食草动物系统也称为放牧系统。在放牧系统中,食草者与植物之间具有复杂的相互关系,简单认为食草动物的牧食会降低草场生产力是错误的。过度放牧也会破坏草原群落。
 协同进化. 植物的补偿作用(生长补偿) 与防卫反应(机械防御或化学防御) 植物主要以两种方式来保护自己免遭捕食:①毒性与差的味道; ②防御结构。 植物—食草动物系统也称为放牧系统。在放牧系统中,食草者与植物之间具有复杂的相互关系,简单认为食草动物的牧食会降低草场生产力是错误的。过度放牧也会破坏草原群落。")
43
2)植物与食植动物的种群模型 其中:V为植物密度;H为食草动物密度; r1—植物内禀增长率;K—未放牧时植物最大密度; d1—在植被稀少时,动物的牧食效率(寻觅效率); d2—在植被变稀时的动物繁殖能力; a—当草场被啃平时,动物的下降率; C1—每头食草动物最大取食率; C2—当草地高密度时对动物下降状况的改善率;

46
7.2.3 寄生与共生 竞争、捕食、寄生、偏害都属于种间负相互作用。
负相互作用使受影响的种群的增长率降低,但不意味着有害。会增加自然选择率,产生新的适应。 图示: 菟丝子寄生在豆科植物上

47
7.2.3.1 寄生 寄生:是指一个种(寄生物)寄居于另外一个种(寄生)体内或体表、靠寄主体液、组织或已消化物质获取营养而生存。
寄生物的分类: 1 微寄生物,在寄主体内或体表繁殖; 2 大寄生物,在寄主体内或表面生长,但不繁殖。在动物中,寄生蠕虫特别重要,而昆虫是植物的主要大寄生物,尽管其他植物也可能是重要的大寄生物。 3 拟寄生物(也叫做重寄生物),包括一大类昆虫大寄生物(主要是寄生蜂和蝇),它们在昆虫寄主身上或体内产卵,通常导致寄主死亡。 大多数寄生物是食生物者,仅在活组织上生活,但一些寄生物在其寄主死后仍能继续存活在寄主上,如丝光绿蝇和引起植物幼苗腐烂的植物真菌,这些称做食尸动物。
,包括一大类昆虫大寄生物(主要是寄生蜂和蝇),它们在昆虫寄主身上或体内产卵,通常导致寄主死亡。 大多数寄生物是食生物者,仅在活组织上生活,但一些寄生物在其寄主死后仍能继续存活在寄主上,如丝光绿蝇和引起植物幼苗腐烂的植物真菌,这些称做食尸动物。")
48
1)寄生物与寄主的相互适应与协同进化 寄生物:神经系统、感觉器官退化,生殖系统发达,复杂的生活史 寄主:产生免疫反应,动物的整理行为、迁移。 进化结果:危害逐渐减弱。
寄生物与寄主的相互适应与协同进化 寄生物:神经系统、感觉器官退化,生殖系统发达,复杂的生活史 寄主:产生免疫反应,动物的整理行为、迁移。 进化结果:危害逐渐减弱。")
49
脊椎动物被微寄生物感染后会产生强烈的免疫反应,这种反应有两种明显成分:
①细胞免疫反应,吞噬细胞(如白血细胞——T淋巴细胞)攻击并吞没病原体细胞, ②B-细胞免疫反应,以特定蛋白(或抗体)的产生为基础,由B淋巴细胞结合到病原体表面。 许多脊椎动物具备整理毛或羽的行为,有效地去除了外寄生物。 植物对病原体还有另一种反应——局部细胞死亡。 寄生物与寄主的协同进化,常常使有害物质的“负作用”减弱,甚至演变为互利共生的关系。
攻击并吞没病原体细胞, ②B-细胞免疫反应,以特定蛋白(或抗体)的产生为基础,由B淋巴细胞结合到病原体表面。 许多脊椎动物具备整理毛或羽的行为,有效地去除了外寄生物。 植物对病原体还有另一种反应——局部细胞死亡。 寄生物与寄主的协同进化,常常使有害物质的 负作用 减弱,甚至演变为互利共生的关系。")
50
寄主密度的增加加剧了寄生物与寄主的接触,为寄生物广泛扩散和传播创造了有利条件,使寄主种群发生流行病并大量死亡。
刚孵出的杜鹃鸟将巢中的其他蛋驱除 2)寄生物与寄主种群相互动态 寄主密度的增加加剧了寄生物与寄主的接触,为寄生物广泛扩散和传播创造了有利条件,使寄主种群发生流行病并大量死亡。 3)社会性寄生物 社会性寄生物不像真寄生物那样摄取寄主组织,而是通过强迫其寄主动物为其提供食物或其他利益而获利,如鸟类的窝寄生。种内窝寄生可发现于一些种类,特别在鸭中很普遍。种间窝寄生包括欧洲的大杜鹃和北美的褐头牛鹂,它们将蛋下在其他种鸟的巢中。
寄生物与寄主种群相互动态. 寄主密度的增加加剧了寄生物与寄主的接触,为寄生物广泛扩散和传播创造了有利条件,使寄主种群发生流行病并大量死亡。 3)社会性寄生物. 社会性寄生物不像真寄生物那样摄取寄主组织,而是通过强迫其寄主动物为其提供食物或其他利益而获利,如鸟类的窝寄生。种内窝寄生可发现于一些种类,特别在鸭中很普遍。种间窝寄生包括欧洲的大杜鹃和北美的褐头牛鹂,它们将蛋下在其他种鸟的巢中。")
51
4)寄生关系的起源和进化 寄生和互利共生都是历史上形成的生物之间在空间上和食物上的联系。寄生关系的起源有三条途径: 1、由空间联系发展到食物联系。先有简单的共栖,再过渡到宿主体上,进而进到体内共栖,不同程度的共栖为发展营养联系建立基础。食物联系可能开始时只是一种对一方有利另一方无害的偏利共生。进一步发展可能出现一方依赖于另一方体液来维持生活,即发展为寄生关系;也可能双方彼此利用代谢产物,发展为互利共生关系。 2、通过捕食过渡到寄生。在自然界中尚保存过渡的痕迹。例如欧洲蛭纲中的平扁舌蛭(Glossiphonia complanata)是营自由生活的捕食者,整个地吞食小无脊椎动物;黄蛭(naemopis)在吞食小动物时和平扁舌蛭一样,但对大动物的攻击和吸血是暂时性的。居住在灌木丛中的山蛭(Hameadipsa)不再采用其它营养方式,而只依赖不时吸吮宿主血液的专性、暂时性寄生生活。尺蠖鱼蛭(Piscicola geometrica)的全部生命活动均在鱼的体躯上,只在繁殖期离开宿主。 3、第三条途径是未来的宿生物偶然的潜入体内,虽然寄主体内是暂时的生活地点,但对寄生物十分有利,成功为兼性寄生物。寄生关系从上述共栖、捕食和偶然寄生三条途径产生后,可以往不同的方向演化。寄生物和宿主的协同进化,常是使有害的“负作用”减弱,甚至于演变为互利共生关系。宿主和寄生物的协同进化可能有三种模式: 1、相互攻击性模式 2、精明寄生性模式 3、早期互利型模式,宿主和寄生物都进化产生互利的特征,以致能加强相互间的持续存在,这是一种由寄生向互利共生的模式发展。
是营自由生活的捕食者,整个地吞食小无脊椎动物;黄蛭(naemopis)在吞食小动物时和平扁舌蛭一样,但对大动物的攻击和吸血是暂时性的。居住在灌木丛中的山蛭(Hameadipsa)不再采用其它营养方式,而只依赖不时吸吮宿主血液的专性、暂时性寄生生活。尺蠖鱼蛭(Piscicola geometrica)的全部生命活动均在鱼的体躯上,只在繁殖期离开宿主。 3、第三条途径是未来的宿生物偶然的潜入体内,虽然寄主体内是暂时的生活地点,但对寄生物十分有利,成功为兼性寄生物。寄生关系从上述共栖、捕食和偶然寄生三条途径产生后,可以往不同的方向演化。寄生物和宿主的协同进化,常是使有害的 负作用 减弱,甚至于演变为互利共生关系。宿主和寄生物的协同进化可能有三种模式: 1、相互攻击性模式. 2、精明寄生性模式. 3、早期互利型模式,宿主和寄生物都进化产生互利的特征,以致能加强相互间的持续存在,这是一种由寄生向互利共生的模式发展。")
52
7.2.3.2 偏利共生 1、原始合作(protocooperation) :两个生物种群生活在一起,彼此都有所得,但两者之间不存在依赖关系。如鸵鸟视觉敏锐,斑马嗅觉灵敏,它们常生活在一起,对发现天敌有利。又如海葵附着于寄居蟹匿居的贝壳上,其刺丝胞可保护寄居蟹,寄居蟹的活动又扩大了海葵的觅食范围。鱼利用强有力的吸盘吸附在鲨鱼等大型鱼类的体表,它吃大鱼吃剩的食物,还靠大鱼扩大自己的活动范围。。 海葵与寄居蟹 2、偏利共生(commensalism):亦称共栖,两个不同物种的个体间发生一种对对方有利而对另一方无害的关系,称为偏利共生。两种都能独立生存的生物以一定的关系生活在一起的现象。如一种色彩鲜艳的双锯鱼常在海葵的触手间游动,受到海葵的保护,而其他种类的小鱼若靠近海葵,就会被其触手抓住并被吃掉的。 双锯鱼与海葵
 :两个生物种群生活在一起,彼此都有所得,但两者之间不存在依赖关系。如鸵鸟视觉敏锐,斑马嗅觉灵敏,它们常生活在一起,对发现天敌有利。又如海葵附着于寄居蟹匿居的贝壳上,其刺丝胞可保护寄居蟹,寄居蟹的活动又扩大了海葵的觅食范围。鱼利用强有力的吸盘吸附在鲨鱼等大型鱼类的体表,它吃大鱼吃剩的食物,还靠大鱼扩大自己的活动范围。。 海葵与寄居蟹. 2、偏利共生(commensalism):亦称共栖,两个不同物种的个体间发生一种对对方有利而对另一方无害的关系,称为偏利共生。两种都能独立生存的生物以一定的关系生活在一起的现象。如一种色彩鲜艳的双锯鱼常在海葵的触手间游动,受到海葵的保护,而其他种类的小鱼若靠近海葵,就会被其触手抓住并被吃掉的。 双锯鱼与海葵.")
53
7.2.3.3 互利共生 互利共生:是两个生物种群生活在一起,相互依赖,相互得益,
其结果使得两个种群都发展得更好。 互利共生发生在生活需要极不相同的生物之间。 1、专性互利共生和兼性互利共生 专性互利共生:指永久性成对组合的生物,其中一方或双方不可能独立生活。 互利共生现象多数属于兼性互利共生,共生者可能不互相依赖着共存,仅是机会性互利共生。 2、传粉和种子散布 自然界中普遍存在的一种植物与动物之间的互利共生存在于有花植物与传粉动物之间。为了与种群中其他个体交换基因,异型杂交植物需要将其花粉转移到另一同种植物的柱头上,并接受同株植物个体的花粉。 传粉者:可能是昆虫、鸟、蝙蝠或小型哺乳动物在植物间传递花粉。啮齿动物、蝙蝠、鸟类和蚂蚁都是重要的种子传播者。 3、防御性互利共生 有一些互利共生为其中一方提供对捕食者或竞争者的防御。蚂蚁-植物互利共生很普遍。 4、动物组织或细胞内的共生性互利共生 一些白蚁还拥有可固定空气中氮的细菌,因为木中氮含量很低,这是有价值的。 藻类与地衣

54
本章到此结束, 谢谢!

Similar presentations

















口-资源-环境动力模型 2007.>")



