Download presentation
Presentation is loading. Please wait.

1
酶促反应动力学 是研究酶促反应的速率以及影响此速率的各种因素的科学。

2
一、

3
影响酶促反应速度的因素 二.底物浓度对酶促反应速度的影响 在低底物浓度时, 反应速度与底物浓度成正比,表现为一级反应特征。
1903年,Henri 用蔗糖酶水解蔗糖的实验 在低底物浓度时, 反应速度与底物浓度成正比,表现为一级反应特征。 当底物浓度达到一定值,反应速度达到最大值(Vmax),此时再增加底物浓度,反应速度不再增加,表现为零级反应。
,此时再增加底物浓度,反应速度不再增加,表现为零级反应。")
4
(一)酶与底物的中间络合物学说(Henri和Wurtz)
该学说认为当酶催化某一化学反应时,酶首先和底物结合生成中间复合物(ES),然后生成产物(P),并释放出酶。 S+E ES P+E
,然后生成产物(P),并释放出酶。 S+E ES P+E.")
5
(二)酶促反应的动力学方程式 S+E ES P+E 米氏方程
Ks k S+E ES P+E 1913年前后,Michaelis和Menten在前人工作的基础上,假定 S+E ES 快速建立平衡,底物浓度远远大于酶浓度,ES分解产物的逆反应忽略不计,推导出下列方程: 米氏方程

6
1.米氏方程的推导 在Michaelis和Menten的酶促反应动力学基础上,1925年,Briggs和Haldane提出酶反应分两步进行,即“稳态平衡”理论: 所谓稳态是指反应进行一段时间后,系统的复合物ES浓度,由零逐渐增加到一定数值,在一定时间内,尽管底物浓度和产物浓度不断地变化.复合物ES也在不断地生成和分解,但是当反应系统中ES的生成速率和ES的分解速率相等时,络合物ES浓度保持不变的这种反应状态称为稳态,即: 第一步: 第二步: ES的浓度与(1)(2)都有关系 [E] 为酶的总浓度 [ES] 为中产物浓度 [E]-[ES] 为游离酶浓度 [S] 为底物浓度 由于[S][E]所以 [S]-[ES][S] (1) (2) ES的形成速度为: (3) (4) ES的分解速度:
(2)都有关系. [E] 为酶的总浓度. [ES] 为中产物浓度. [E]-[ES] 为游离酶浓度. [S] 为底物浓度. 由于[S][E]所以. [S]-[ES][S] (1) (2) ES的形成速度为: (3) (4) ES的分解速度:")
7
米氏方程 在稳态下,ES的生成速率和ES的分解速率相等,即[ES]保持动态平衡: 因为酶反应速率 与[ES]成正比,即:
由干反应系统中[S]>>[E],当[S]很高时所有的酶都被底物所饱和形成ES,即[E]=[ES],酶促反应达到最大速率 ,则: 米氏方程
![米氏方程 在稳态下,ES的生成速率和ES的分解速率相等,即[ES]保持动态平衡: 因为酶反应速率 与[ES]成正比,即:](http://slidesplayer.com/slide/11177878/60/images/7/%E7%B1%B3%E6%B0%8F%E6%96%B9%E7%A8%8B+%E5%9C%A8%E7%A8%B3%E6%80%81%E4%B8%8B%EF%BC%8CES%E7%9A%84%E7%94%9F%E6%88%90%E9%80%9F%E7%8E%87%E5%92%8CES%E7%9A%84%E5%88%86%E8%A7%A3%E9%80%9F%E7%8E%87%E7%9B%B8%E7%AD%89%EF%BC%8C%E5%8D%B3%5BES%5D%E4%BF%9D%E6%8C%81%E5%8A%A8%E6%80%81%E5%B9%B3%E8%A1%A1%EF%BC%9A+%E5%9B%A0%E4%B8%BA%E9%85%B6%E5%8F%8D%E5%BA%94%E9%80%9F%E7%8E%87+%E4%B8%8E%5BES%5D%E6%88%90%E6%AD%A3%E6%AF%94%EF%BC%8C%E5%8D%B3%EF%BC%9A.jpg "由干反应系统中[S]>>[E],当[S]很高时所有的酶都被底物所饱和形成ES,即[E]=[ES],酶促反应达到最大速率 ,则: 米氏方程.")
8
当[S]Km时,v=Vmax/Km[S]
米氏常数是当酶反应速率达到最大反应速率的一半时的底物浓度。 因此,米氏常数的单位为mol/L。
![当[S]Km时,v=Vmax/Km[S]](http://slidesplayer.com/slide/11177878/60/images/8/%E5%BD%93%5BS%5D%EF%80%BC%EF%80%BCKm%E6%97%B6%EF%BC%8Cv%3DVmax%2FKm%5BS%5D.jpg "米氏常数是当酶反应速率达到最大反应速率的一半时的底物浓度。 因此,米氏常数的单位为mol/L。")
9
2.动力学参数的意义 (1)米氏常数的意义 ①Km是酶的一个重要的特征常数。
只与酶的性质有关,而与其浓度无关。Km值只是在固定的底物,一定的温度和pH条件下,一定的缓冲体系中测定的,不同条件下具有不同的Km值。 Km值可用来鉴别酶。 ②Km值可以判断酶的专一性和天然底物。 有的酶可作用于几种底物,因此有几个 Km 值,同一种酶有几种底物就有几个Km值,其中Km值最小的底物一般称为该酶的最适底物或天然底物。1/Km值可近似地表示酶对底物亲和力的大小, 1/Km值大表示亲和程度大,酶的催化活性低; 1/Km值小表示亲和程度大,酶的催化活性高。 可判断酶的专一性。

10
特例: Km=K2/K1=Ks(在K3K2时)
所以1/Km表示形成ES的趋势大小 特例: Km=K2/K1=Ks(在K3K2时) ③Km与Ks Km不等于Ks 。在K3<<K1、K2时, Km看作Ks,也只有此时1/Km 才可以近似表示酶与底物结合的难易程度。
 ③Km与Ks Km不等于Ks 。在K3<<K1、K2时, Km看作Ks,也只有此时1/Km 才可以近似表示酶与底物结合的难易程度。")
11
④若已知某个酶的Km值,可以计算在某一个底物浓度时,反应速率相当于最大反应速率Vmax的百分率。
如:[S]=3Km时,根据: 则: ⑤Km可以帮助推断某一反应的方向和途径

12
A.判断可逆反应方向的效率,了解其主要催化方向
肌酸+ATP 磷酸肌酸+ADP Km (mol/L) B.可推断连锁反应的限速步骤 Km ( mol/L) 酶 酶 酶 Km小的酶促反应比较占优势
 B.可推断连锁反应的限速步骤. Km ( mol/L) 酶 酶 酶 Km小的酶促反应比较占优势.")
13
丙酮酸 Km=1.7x10-5mol/L 乳酸 Km=1.3x10-3 mol/L 乙酰CoA Km=1.0x10-3mol/L 乙醛
乳酸脱氢酶 丙酮酸脱氢酶 乙酰CoA 丙酮酸脱羧酶 Km小的酶促反应比较占优势 乙醛

14
(2)Vmax和 的意义 在一定酶浓度下,酶对特定底物的Vmax也是一个常数。同一种酶对不同底物的Vmax 也不同,pH、温度和离子强度等因素也影响Vmax 的数值。 当[S] 很大时, 成线性关系,而直线的斜率为 ,为一级反应速率常数,它表示当酶被底物饱和时每秒钟每个酶分子转换底物的分子数,这个常数又叫做转换数(简称TN),通称为催化常数 。 值越大,表示酶的催化效率越高。
,通称为催化常数 。 值越大,表示酶的催化效率越高。")
15
米氏方程

17
3. 利用作图法测定Km和Vmax Km = + V Vmax [S] Vmax
![3. 利用作图法测定Km和Vmax 1 Km 1 1 = + V Vmax [S] Vmax](http://slidesplayer.com/slide/11177878/60/images/17/3.+%E5%88%A9%E7%94%A8%E4%BD%9C%E5%9B%BE%E6%B3%95%E6%B5%8B%E5%AE%9AKm%E5%92%8CVmax+1+Km+1+1+%EF%82%BE%EF%82%BE%EF%82%BE+%3D+%EF%82%BE%EF%82%BE+%EF%82%B4+%EF%82%BE%EF%82%BE%EF%82%BE+%2B+%EF%82%BE%EF%82%BE+V+Vmax+%5BS%5D+Vmax.jpg "3. 利用作图法测定Km和Vmax 1 Km 1 1 = + V Vmax [S] Vmax")
18
a.双倒数作图法 (Lineweaver-Burk)
斜率=Km/Vmax -1/Km 1/Vmax

19
b. v-v/[S]作图法 (Eadie-Hofstee)
将米氏方程改写: Vmax Vmax/Km
![b. v-v/[S]作图法 (Eadie-Hofstee)](http://slidesplayer.com/slide/11177878/60/images/19/b.+v-v%2F%5BS%5D%E4%BD%9C%E5%9B%BE%E6%B3%95+%28Eadie-Hofstee%29.jpg "将米氏方程改写: Vmax. Vmax/Km.")
20
c.Hanes-Woolf作图法 Km = + V Vmax [S] Vmax
![c.Hanes-Woolf作图法 1 Km 1 1 = + V Vmax [S] Vmax](http://slidesplayer.com/slide/11177878/60/images/20/c.Hanes-Woolf%E4%BD%9C%E5%9B%BE%E6%B3%95+1+Km+1+1+%EF%82%BE%EF%82%BE%EF%82%BE+%3D+%EF%82%BE%EF%82%BE+%EF%82%B4+%EF%82%BE%EF%82%BE%EF%82%BE+%2B+%EF%82%BE%EF%82%BE+V+Vmax+%5BS%5D+Vmax.jpg "c.Hanes-Woolf作图法 1 Km 1 1 = + V Vmax [S] Vmax")
21
d. Eisenthal和Cornish-Bowden直接线性作图
将米氏方程改写为:

22
(三)多底物的酶促反应动力学 1.酶促反应按参加的底物数目来分 2.按动力学机制来分
分单底物、双底物和三底物反应,最重要的是双底物反应。 2.按动力学机制来分 (1)序列反应——底物的结合和产物的释放有一定的顺序,产物不能在底物完全结合前释放。A和B两底物均结合到酶上,然后反应产生两产物P和Q。 序列反应分两类:有序反应和随机反应
序列反应——底物的结合和产物的释放有一定的顺序,产物不能在底物完全结合前释放。A和B两底物均结合到酶上,然后反应产生两产物P和Q。 序列反应分两类:有序反应和随机反应.")
23
①有序反应——底物按照一定的顺序与酶结合,形成三元复合物,然后按照一定的顺序释放反应的产物。
需要NAD或NADP的脱氢酶属于这种类型,这些脱氢酶的一般反应为: 优先底物A 随后底物B 随后产物Q 优先产物P

24
以乙醇脱氢酶为例: 底物B 底物A AEB PEQ 产物Q 产物P

25
②随机反应——两底物随机地与酶结合,形成三元复合物,产物的释放也是随机的。

26
以肌酸激酶为例: 肌酸(Cr) + ATP 磷酸肌酸+ADP
 + ATP 磷酸肌酸+ADP")
27
(2)乒乓反应——酶与领先底物A的反应产物(P)在酶与第二个底物B反应之前释放出来,结果酶E转变为一种修饰酶形式E’,然后再与底物B反应生成第二个产物Q,同时释放出未被修饰的酶E。
氨基转移酶属于这类作用机制

28
谷氨酸:天冬氨酸氨基转移酶

29
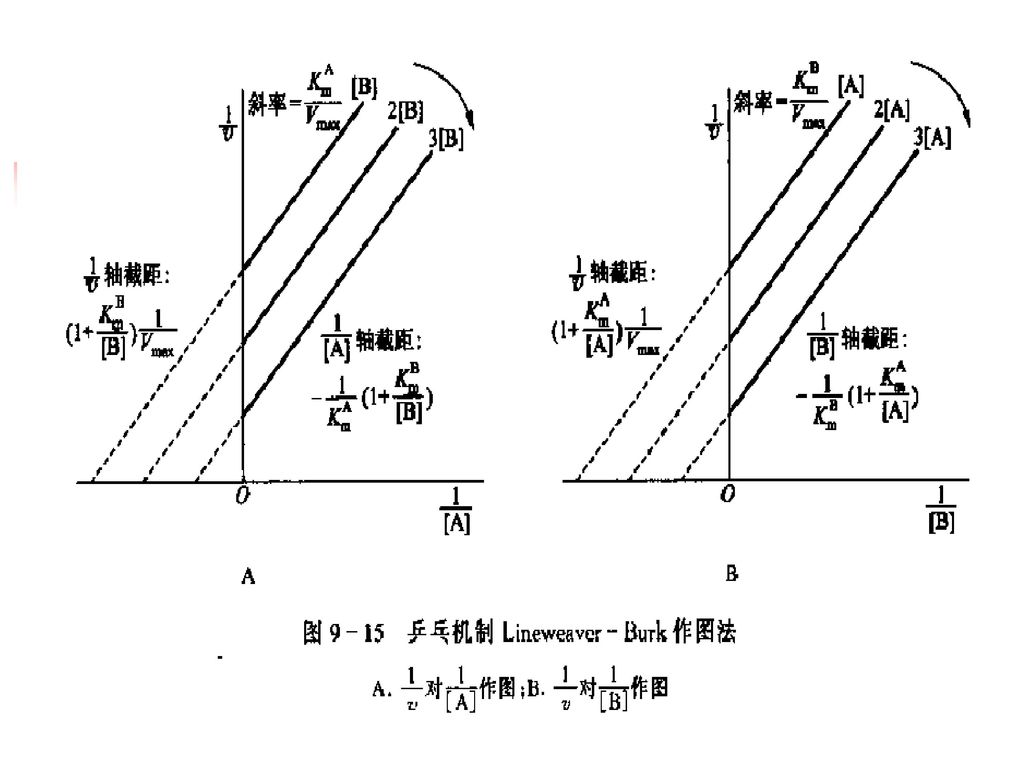
双底物反应的动力学方程 (1)乒乓机制的动力学方程和动力学图 对上式取双倒数,得下式:
乒乓机制的动力学方程和动力学图 对上式取双倒数,得下式:")
31
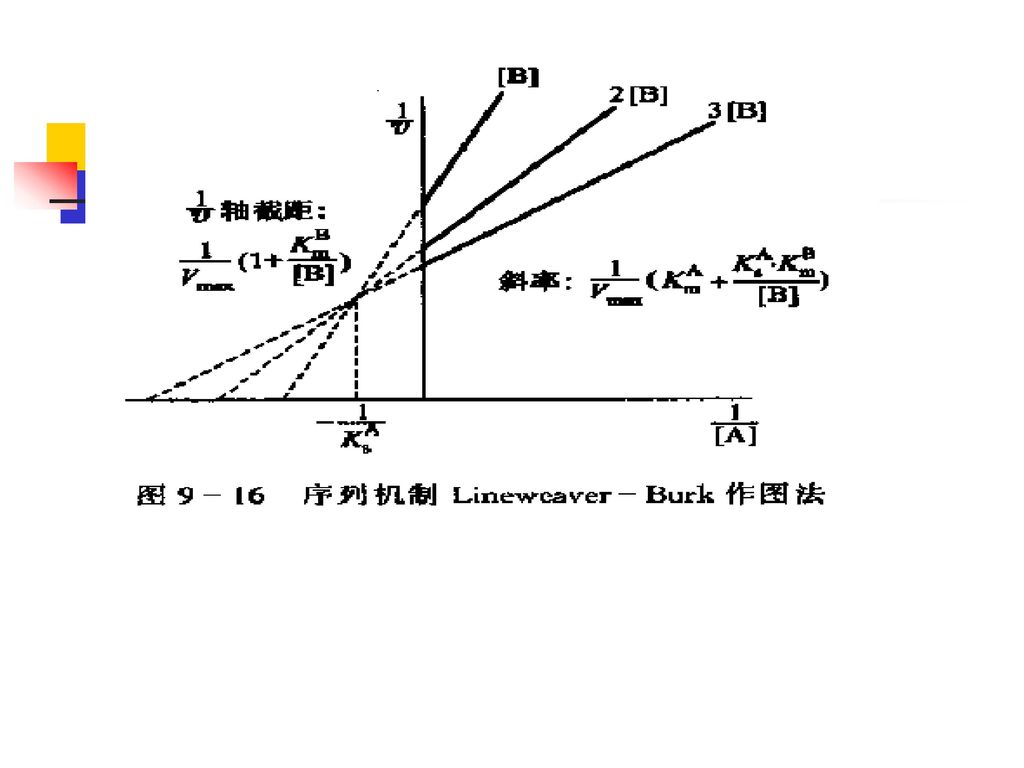
(2)序列机制的动力学方程和动力学图 对上式取双倒数,得下式:
序列机制的动力学方程和动力学图 对上式取双倒数,得下式:")
33
三.酶的抑制作用 抑制作用与抑制剂 (抑制剂对酶反应速度的影响)
凡使酶的活性降低或丧失,但并不引起酶蛋白变性的作用称为抑制作用(inhibition)。 能够引起抑制作用的化合物则称为抑制剂(inhibitor)。(抑制剂不同于变性剂) (一) 抑制程度的表示方法 加入抑制剂后的反应速率 不加抑制剂时的反应速率
。 能够引起抑制作用的化合物则称为抑制剂(inhibitor)。(抑制剂不同于变性剂) (一) 抑制程度的表示方法. 加入抑制剂后的反应速率. 不加抑制剂时的反应速率.")
34
(二) 抑制作用的类型 包括不可逆抑制作用(irreversible inhibition)和可逆抑制作用(reversible inhibition)。 1. 不可逆抑制作用(irreversible inhibition) 抑制剂与酶蛋白中的必需基团以共价键形式结合,引起酶的永久性失活,不能用透析或超滤等物理方法除去抑制剂而恢复酶活性。 专一性不可逆抑制作用:这类抑制剂只作用于与酶活性部位有关的氨基酸残基或一类酶。 非专一性不可逆抑制作用:这类抑制剂作用于酶分子上不同的基团或作用于几类不同的酶。如:酰化剂酸酐和磺酰氯等可使酶蛋白的-OH、SH、NH2等发生酰化。

35
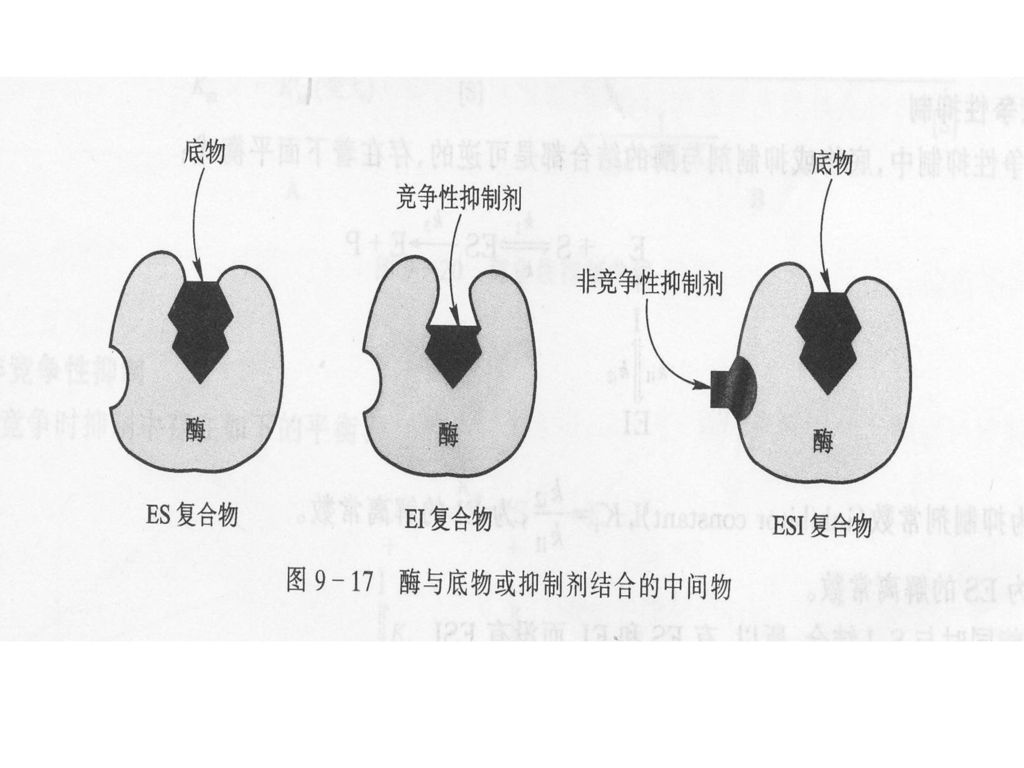
竞争性抑制(competitive inhibition) 非竞争性抑制(noncompetitive inhibition )
2. 可逆抑制作用(reversible inhibition) 抑制剂与酶蛋白以非共价键方式结合,引起酶活性降低或暂时性丧失。抑制剂可以通过透析等方法被除去,并且能部分或全部恢复酶的活性。根椐抑制剂与酶结合的情况,又可以分为三类 竞争性抑制(competitive inhibition) 非竞争性抑制(noncompetitive inhibition ) 反竞争性抑制(uncompetitive inhibition )
 抑制剂与酶蛋白以非共价键方式结合,引起酶活性降低或暂时性丧失。抑制剂可以通过透析等方法被除去,并且能部分或全部恢复酶的活性。根椐抑制剂与酶结合的情况,又可以分为三类. 竞争性抑制(competitive inhibition) 非竞争性抑制(noncompetitive inhibition ) 反竞争性抑制(uncompetitive inhibition )")
36
(1)竞争性抑制(competitive inhibition)
某些抑制剂的化学结构与底物相似,因而能与底物竞争与酶活性中心结合。当抑制剂与活性中心结合后,底物被排斥在反应中心之外,其结果是酶促反应被抑制了。 竞争性抑制通常可以通过增大底物浓度, 即提高底物的竞争能力来消除。

37
竟争性抑制作用的例子:

38
(2) 非竟争性抑制(noncompetitive inhibition )
酶可同时与底物及抑制剂结合,即底物和抑制剂没有竞争作用。酶与抑制剂结合后,还可与底物结合;酶与底物结合后,也可再结合抑制剂,但是三元的中间产物不能进一步分解为产物,所以酶活性降低。 如某些金属离子(Cu2+、Ag+、Hg+、Pb2+等)通常能与酶分子的调控部位中的-SH基团作用,改变酶的空间构象,引起非竞争性抑制;EDTA结合金属离子引起的抑制作用也属于非竞争性抑制。 非竞争性抑制剂与酶活性中心以外的基团结合。这类抑制作用不会因提高底物浓度而减弱
通常能与酶分子的调控部位中的-SH基团作用,改变酶的空间构象,引起非竞争性抑制;EDTA结合金属离子引起的抑制作用也属于非竞争性抑制。 非竞争性抑制剂与酶活性中心以外的基团结合。这类抑制作用不会因提高底物浓度而减弱.")
40
(3) 反竞争性抑制(uncompetitive inhibition )
酶只有与底物结合后才与抑制剂结合,从而导致酶活性下降。这类抑制作用最不重要。

41
(三)可逆抑制作用和不可逆抑制作用的鉴别
1.物理方法 用透析、超滤或凝胶过滤等物理方法区别。 2.动力学方法 ①在测定酶活力的系统中加入一定量的抑制剂,然后测定不同酶浓度的反应初速率。

42
②在测定酶活力的系统中加入不同浓度的抑制剂,然后测定不同酶浓度的反应初速率。

43
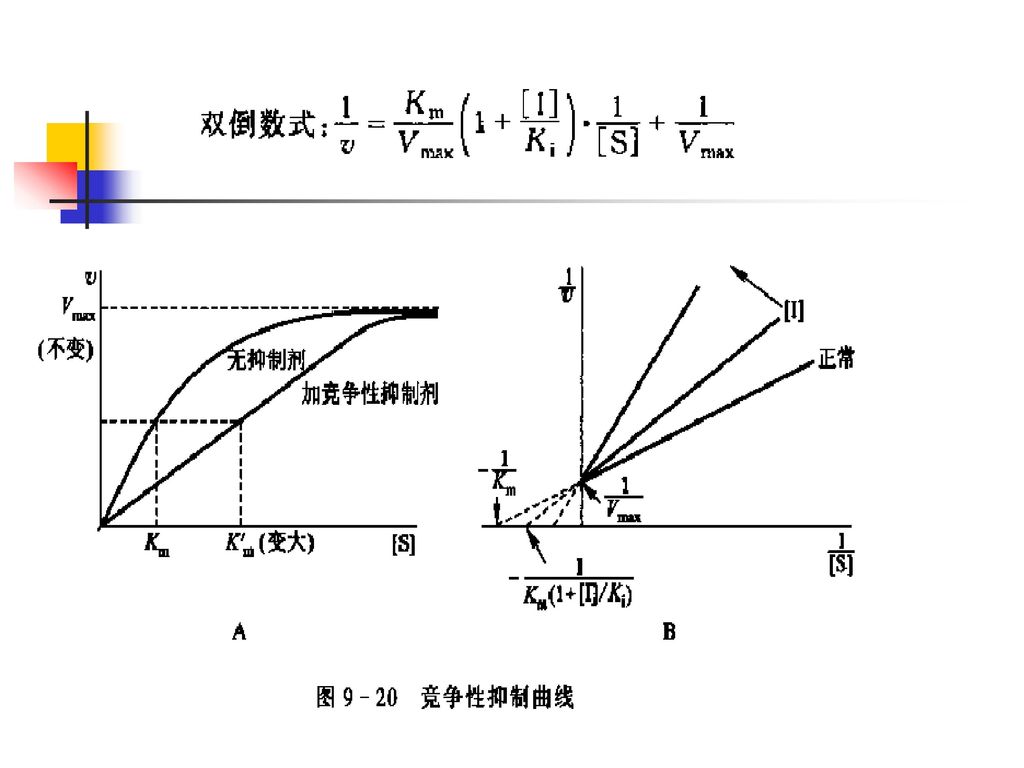
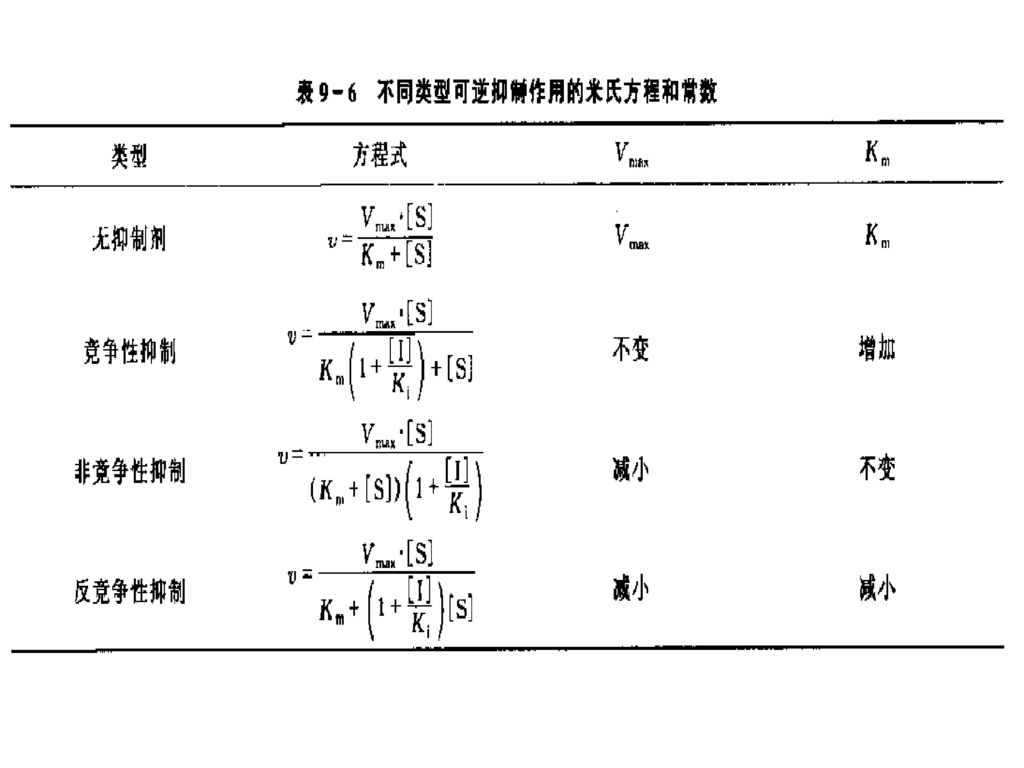
(四)可逆抑制作用动力学 1. 竞争性抑制 加入竞争性抑制剂后,Km 变大,酶促最大反应速度不变。
可逆抑制作用动力学 1. 竞争性抑制 加入竞争性抑制剂后,Km 变大,酶促最大反应速度不变。")
45
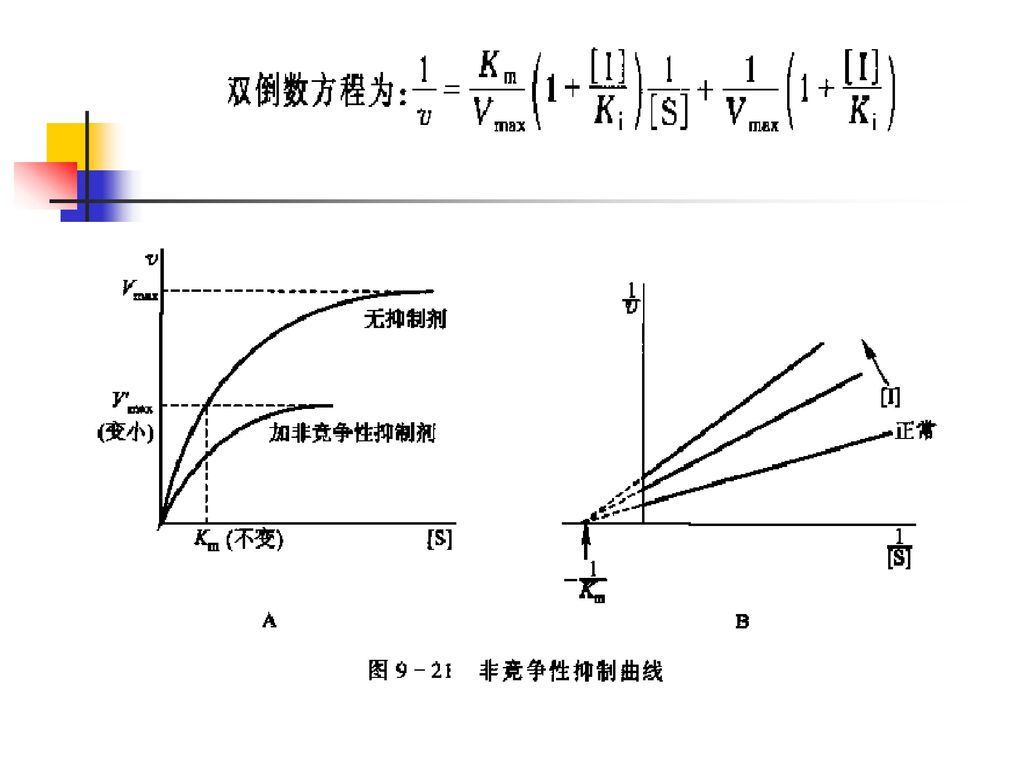
2.非竞争性抑制 加入非竞争性抑制剂后,Km 不变,而Vmax减小。

47
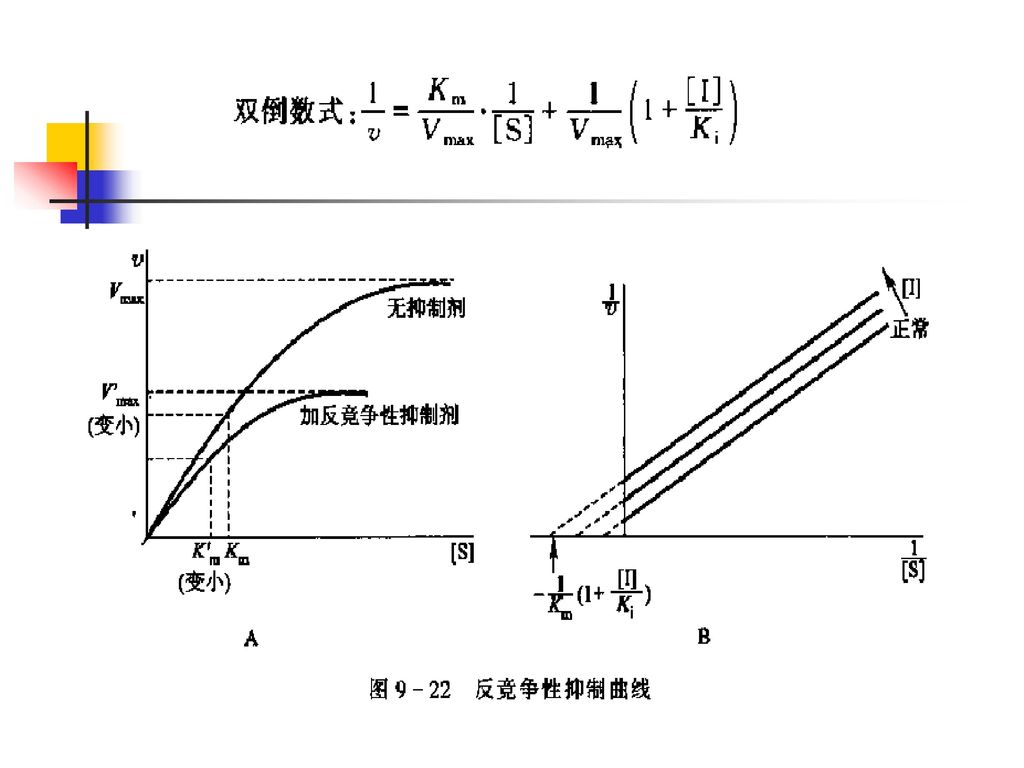
3.反竞争性抑制 加入反竞争性抑制剂,使Km和Vmax均减小

50
(五)一些重要的抑制剂 1.不可逆抑制剂 (1)非专一性不可逆抑制剂 ①有机磷化合物—与酶活性直接有关的丝氨酸上的-OH牢固地结合,从而抑制某些蛋白酶或酯酶。(DFP、敌百虫、敌敌畏、农药1605等)
 .")
51
这类化合物又叫神经毒剂 抑制 胆碱酯酶 乙酰胆碱 乙酸+胆碱 可用解磷定(碘化醛肟甲基吡啶)或氯磷定(氯化醛肟甲基吡啶)解毒
或氯磷定(氯化醛肟甲基吡啶)解毒")
52
②有机汞、有机砷化合物—与酶蛋白上的-SH作用,从而抑制含-SH酶的活性。这类抑制作用可加入过量的巯基化合物(如半胱氨酸或还原型谷胱甘肽GSH)解除。
如,对氯汞苯甲酸 有机砷化合物如路易斯毒气作用机制

53
③重金属盐——如Ag+、Cu2+ 、Hg2+、Pb2+、Fe3+等在高浓度时能使大多数酶失活,低浓度时起抑制作用,加入金属螯合剂EDTA、半胱氨酸等可以解除。
④烷化剂——如碘乙酸、碘乙酰胺、2,4-二硝基氟苯等能使酶蛋白中的 –SH、-NH2、-OH、-COOH、咪唑基等发生烷基化, 使酶失活。 ⑤氰化物、硫化物、CO—与含铁卟啉的酶中的Fe3+结合,使酶失活。 ⑥抗菌素类药物,与糖肽转肽酶活性部位Ser-OH共价结合,使酶失活,该酶在细菌细胞壁合成中使肽聚糖链交联,使细菌细胞壁合成受到阻碍,从而起到抗菌作用。

54
,该酶 (2)专一性不可逆抑制剂 ① Ks型不可逆抑制剂 这类抑制机可分为Ks型和Kcat型两大类
这类抑制剂与酶的底物结构类似,可以和相应的酶结 合,同时还带有一个活泼的化学基团,能与酶活性中心基 团反应进行化学修饰,从而抑制酶活性。因抑制是通过对 酶的亲和力来对酶进行修饰标记的,又叫亲和标记试剂 (affnity labeling reagent)。 由于这类抑制剂的活泼基团也可以修饰酶分子其他部 位的同一基团,因此其专一性有一定的限度。这取决于抑 制剂与活性部位必需基团在反应前形成非共价络合物的解 离常数以及非活性部位同类基团形成非共价络合物的解离 常数之比,即Ks的比值,故这类抑制剂称为Ks型不可逆抑 制剂。 ,该酶
。 由于这类抑制剂的活泼基团也可以修饰酶分子其他部. 位的同一基团,因此其专一性有一定的限度。这取决于抑. 制剂与活性部位必需基团在反应前形成非共价络合物的解. 离常数以及非活性部位同类基团形成非共价络合物的解离. 常数之比,即Ks的比值,故这类抑制剂称为Ks型不可逆抑. 制剂。 ,该酶.")
55
例如胰蛋白酶:活性中心有一个催化必需基团His57。
底物 底物类似物

56
例如β-卤代-D-Ala是细菌中丙氨酸消旋酶(AR)的Kcat型不可逆抑制剂,属于以磷酸吡哆醛为辅酶的酶类的自杀性底物。
Kcat型抑制剂不但具有与天然底物类似的结构,而且本身也是酶的底物。还有一个潜伏的反应基团,当酶对这类抑制剂进行催化反应时,该潜伏反应基团被暴露或活化,又作用于酶活性中心的有关基团,使酶不可拟失活。即底物需经过酶催化之后,才能形成酶的不可逆抑制剂,因此,称之为“自杀性底物”(suicide substrates)。 例如β-卤代-D-Ala是细菌中丙氨酸消旋酶(AR)的Kcat型不可逆抑制剂,属于以磷酸吡哆醛为辅酶的酶类的自杀性底物。 AR L-Ala D-Ala
。 例如β-卤代-D-Ala是细菌中丙氨酸消旋酶(AR)的Kcat型不可逆抑制剂,属于以磷酸吡哆醛为辅酶的酶类的自杀性底物。 AR. L-Ala D-Ala.")
57
磷酸吡哆醛

58
2.可逆抑制剂 在可逆抑制剂中最重要的是竞争性抑制剂。大多数竞争性抑制剂与酶催化的天然代谢物在结构上十分相似,能选择性地抑制病菌或癌细胞在代谢过程中的某些酶,而具有抗菌和抗癌作用。这类抑制剂可称为抗代谢物或代谢类似物。例如: ①5′-氟尿嘧啶(5′- FU)抑制胸腺嘧啶核苷酸合成酶。 尿嘧啶脱氧核糖核苷酸 胸腺嘧啶核苷酸 胸腺嘧啶核苷酸合成酶 DNA
抑制胸腺嘧啶核苷酸合成酶。 尿嘧啶脱氧核糖核苷酸 胸腺嘧啶核苷酸. 胸腺嘧啶核苷酸合成酶. DNA.")
59
②磺胺类药物:以对氨基苯磺酰胺为例说明磺胺药物的作用机制。
人可利用外源叶酸,而细菌则不能! 叶酸(外源) 对氨基苯磺酰胺 二氢叶酸还原酶 抑制 二氢叶酸合成酶 对氨基苯甲酸 二氢叶酸还原酶 二氢叶酸 四氢叶酸 嘌呤核苷酸合成途径中的辅酶
 对氨基苯磺酰胺. 二氢叶酸还原酶. 抑制. 二氢叶酸合成酶. 对氨基苯甲酸. 二氢叶酸还原酶. 二氢叶酸. 四氢叶酸. 嘌呤核苷酸合成途径中的辅酶.")
60
③过渡态类似物 腺嘌呤核苷——小牛肠腺苷脱氨酶 腺苷五磷酸——腺苷酸激酶(催化ATP+AMP 2ADP)
草酸——乳酸脱氢酶、草酰乙酸脱羧酶、丙酮酸羧化酶 (乳酸、草酰乙酸、丙酮酸共有的过渡态是烯醇式丙酮酸,草酸是烯醇式丙酮酸的结构类似物)
")
61
四. 温度对酶反应的影响 一方面是温度升高,酶促反应速度加快(温度系数Q10:反应温度提高10 C,其反应速度与原来的反应速度之比)。
另一方面,温度升高,酶的高级结构将发生变化或变性,导致酶活性降低甚至丧失。 因此大多数酶都有一个最适温度。 在最适温度(optimum temperature )条件下,反应速度最大。 酶对温度的耐受力与其存在状态有关。
条件下,反应速度最大。 酶对温度的耐受力与其存在状态有关。")
62
五. pH 的影响 在一定的pH 下, 酶具有最大的催化活性,通常称此pH 为最适 pH(optimum pH)。
各种酶在一定条件下都有其特定的最适pH,因此最适pH是酶的特性之一。但酶的最适pH不是一个常数,受许多因素影响,随底物种类和浓度、缓冲液种类和浓度的不同而改变,因此最适pH只有在一定条件下才有意意义。、大多数酶的最适pH在5-8之间。动物体的酶多在pH 之间,植物及微生物中的酶多在pH4.5~6.5左右。

63
pH影响酶活力的原因有以下几个方面: (1)过酸或过碱可以使酶的空间结构破坏。引起酶构象的改变,酶活性丧失。
(2)当pH改变不很剧烈时,酶虽然末变性,但活力受到影响。pH影响了底物的解离状态,或使底物不能和酶结合,或者结合后不能生成产物; pH影响酶分子活性部位上有关基团的解离,从而影响与底物的结合或催化,使酶活性降低;也可能影响到中间络合物ES的解离状态,不利于催化生成产物。 (3) pH影响维持酶分子空间结构的有关基团解离,从而影响了酶活性部位的构象,进而影响酶的活性。 由于酶活力受pH的影响很大,因此在酶的分离纯化时要选择酶的稳定pH,通常在某一pH缓冲液中进行。
当pH改变不很剧烈时,酶虽然末变性,但活力受到影响。pH影响了底物的解离状态,或使底物不能和酶结合,或者结合后不能生成产物; pH影响酶分子活性部位上有关基团的解离,从而影响与底物的结合或催化,使酶活性降低;也可能影响到中间络合物ES的解离状态,不利于催化生成产物。 (3) pH影响维持酶分子空间结构的有关基团解离,从而影响了酶活性部位的构象,进而影响酶的活性。 由于酶活力受pH的影响很大,因此在酶的分离纯化时要选择酶的稳定pH,通常在某一pH缓冲液中进行。")
64
六.酶浓度对酶反应速度的影响 Vmax=K3[E] [E] 酶浓度对反应速度的影响
![六.酶浓度对酶反应速度的影响 Vmax=K3[E] [E] 酶浓度对反应速度的影响](http://slidesplayer.com/slide/11177878/60/images/64/%E5%85%AD.%E9%85%B6%E6%B5%93%E5%BA%A6%E5%AF%B9%E9%85%B6%E5%8F%8D%E5%BA%94%E9%80%9F%E5%BA%A6%E7%9A%84%E5%BD%B1%E5%93%8D+Vmax%3DK3%5BE%5D+%EF%81%AE+%5BE%5D+%E9%85%B6%E6%B5%93%E5%BA%A6%E5%AF%B9%E5%8F%8D%E5%BA%94%E9%80%9F%E5%BA%A6%E7%9A%84%E5%BD%B1%E5%93%8D.jpg "六.酶浓度对酶反应速度的影响 Vmax=K3[E] [E] 酶浓度对反应速度的影响")
65
凡能提高酶活性的物质,都称为激活剂(activator)。
七.激活剂对酶反应速度的影响 凡能提高酶活性的物质,都称为激活剂(activator)。 (1)无机离子:金属离子(K+ Na+ Mg2+ Zn2+ Fe2+ Ca2+)、阴离子(Cl- Br-)、氢离子 (2)小分子有机化合物:某些还原剂(如半胱氨酸、GSH等)、乙二胺四乙酸(EDTA) (3)某些酶类:酶原激活过程中的酶类
。 (1)无机离子:金属离子(K+ Na+ Mg2+ Zn2+ Fe2+ Ca2+)、阴离子(Cl- Br-)、氢离子. (2)小分子有机化合物:某些还原剂(如半胱氨酸、GSH等)、乙二胺四乙酸(EDTA) (3)某些酶类:酶原激活过程中的酶类.")
Similar presentations






 氧化分解供应能量 转化为其他物质.>")
 二、脂肪.>")


 蔡哲瑜(4BS ) 詹君瑞(4BS ) 指導老師 : 潘文彬 老師>")









