Download presentation
Presentation is loading. Please wait.

1
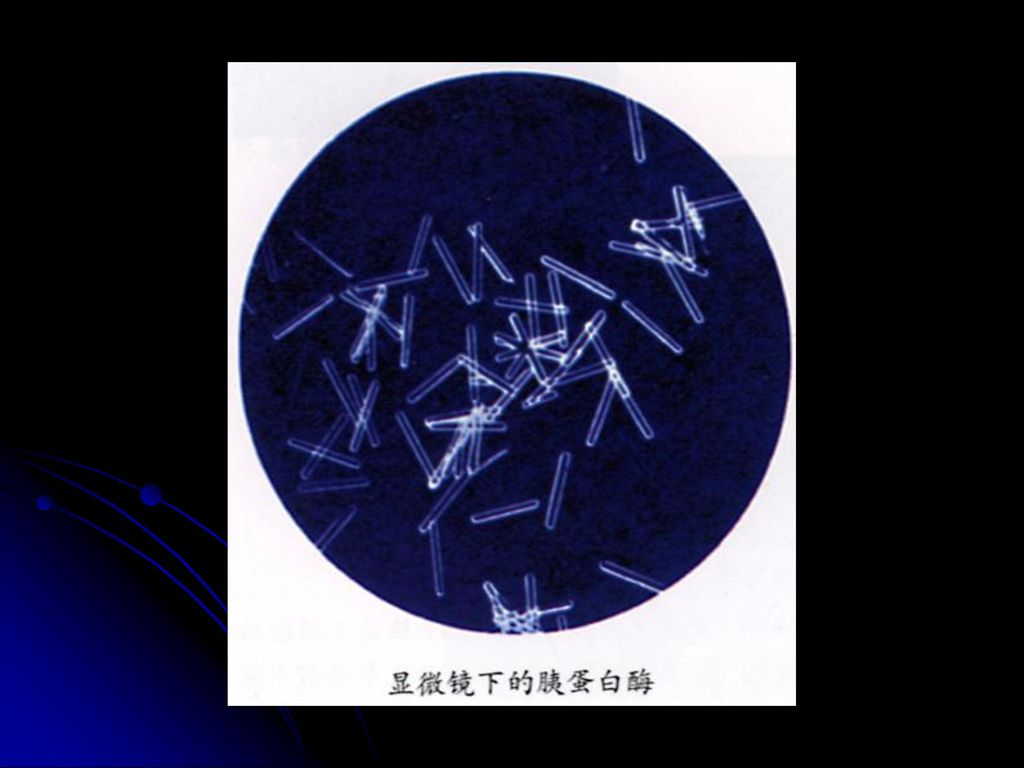
酶 胃蛋白酶

2
第一节 酶的概念 一、酶是生物催化剂 (一)酶的定义 (二)酶的化学本质—蛋白质(核酸)
第一节 酶的概念 一、酶是生物催化剂 (一)酶的定义 酶是由活细胞产生的具有催化功能的蛋白质(核酸),亦称生物催化剂。 (二)酶的化学本质—蛋白质(核酸) 酶的化学本质除有催化活性的RNA之外几乎都是蛋白质。 绝大多数酶都是蛋白质。 1983年发现某些RNA分子具有催化活性,对有催化 活性的RNA称为核酶 酶催化的生物化学反应,称为酶促反应 在酶的催化下发生化学变化的物质,称为底物
酶的定义. 酶是由活细胞产生的具有催化功能的蛋白质(核酸),亦称生物催化剂。 (二)酶的化学本质—蛋白质(核酸) 酶的化学本质除有催化活性的RNA之外几乎都是蛋白质。 绝大多数酶都是蛋白质。 1983年发现某些RNA分子具有催化活性,对有催化 活性的RNA称为核酶. 酶催化的生物化学反应,称为酶促反应. 在酶的催化下发生化学变化的物质,称为底物.")
4
二、酶的作用特点 (一)与一般催化剂的相同点 1.反应前后质与量不变,且用量少。 2.缩短到达平衡的时间,不改变平衡点。
3.只能催化本来进行的反应。 4.降低反应所需活化能。

5
活化能:在一定温度下1mol底物全部进入活化态所需要的自由能,单位为kJ/mol.

6
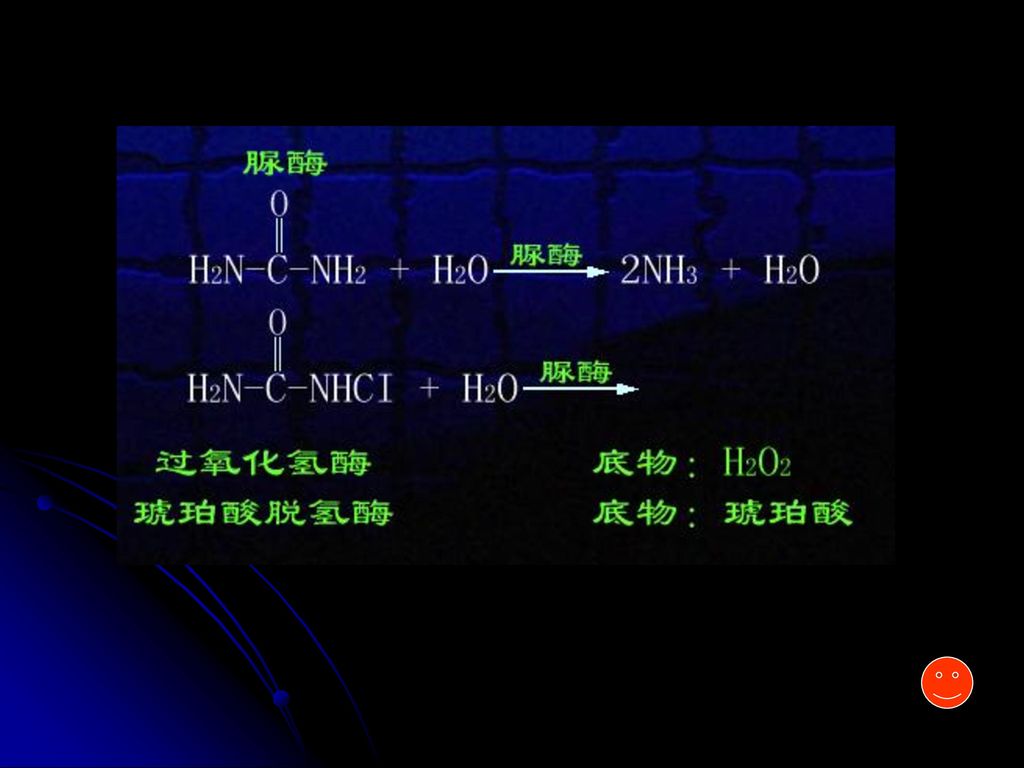
1mol/L过氧化氢酶1秒钟可分解105mol/L底物 1mol/L铁离子1秒钟可分解10-5 mol/L底物
(二)与一般催化剂的不同点 1.高效 比一般催化剂高106—1013倍 1mol/L过氧化氢酶1秒钟可分解105mol/L底物 1mol/L铁离子1秒钟可分解10-5 mol/L底物 (75.4—48.9—8.4kJ/mol) 2.高度专一 酶对催化的反应和反应物有严格的选择性 3.条件温和(酶易失活) 引起蛋白质变性的因素都能使酶失活 4.活性能调节、控制 5.常需要辅助因子
与一般催化剂的不同点. 1.高效 比一般催化剂高106—1013倍. 1mol/L过氧化氢酶1秒钟可分解105mol/L底物. 1mol/L铁离子1秒钟可分解10-5 mol/L底物. (75.4—48.9—8.4kJ/mol) 2.高度专一 酶对催化的反应和反应物有严格的选择性. 3.条件温和(酶易失活) 引起蛋白质变性的因素都能使酶失活. 4.活性能调节、控制. 5.常需要辅助因子.")
7
三、酶的分布 酶分布在所有的细胞和组织中,相 对隔离,各自发挥作用。

8
四、酶的专一性 酶对底物和催化的反应有严格的选择性。 一种酶仅能作用于一种底物或结构上相似的一类物质,促进发生一定的化学反应。 绝对专一
结构专一 键专一 相对专一 专一性 基团专一 旋光异构 立体(异构)专一 顺反异构
专一. 顺反异构.")
9
1.绝对专一 一种酶只作用于一种底物。 2.相对专一 一种酶能够作用于结构上类似的一系列化合物。 3.立体异构专一性

10
第二节 酶的命名与分类 一、酶的分类 二、酶的命名

11
第三节 酶的组成与结构 一、酶的组成 二、几个重要的辅助因子 三、酶的结构 (一)辅酶、辅基 (二)金属离子
第三节 酶的组成与结构 一、酶的组成 二、几个重要的辅助因子 (一)辅酶、辅基 (二)金属离子 三、酶的结构 (一)酶蛋白与蛋白质相同,都有一定的空间结构 (二)根据酶蛋白结构上的特点,酶分为: (三)酶的活性中心 (一)含义 (二)形成 (三)酶原
辅酶、辅基. (二)金属离子. 三、酶的结构. (一)酶蛋白与蛋白质相同,都有一定的空间结构. (二)根据酶蛋白结构上的特点,酶分为: (三)酶的活性中心. (一)含义. (二)形成. (三)酶原.")
13
(1)键专一:只要求合适的化学键,对键两端的基团并无严格要求。
(2)基团专一:不但要求一定的化学键,还要求键一端的基团是一定的。 a-葡萄糖苷酶 a-是糖苷键 糖苷键的一端是葡萄糖 底物:蔗糖、麦芽糖
基团专一:不但要求一定的化学键,还要求键一端的基团是一定的。 a-葡萄糖苷酶. a-是糖苷键. 糖苷键的一端是葡萄糖. 底物:蔗糖、麦芽糖.")
14
(1)旋光异构专一性: (2)顺反异构专一性:
旋光异构专一性: (2)顺反异构专一性:")
15
根据国际生化协会酶命名委员会的规定,每一个酶都用四个打点隔开的数字编号,编号前冠以EC(酶学委员会缩写),四个数字依次表示该酶应属的大类、亚类、亚亚类及酶的顺序号,这种编码一种酶的四个数字即是酶的标码。
,四个数字依次表示该酶应属的大类、亚类、亚亚类及酶的顺序号,这种编码一种酶的四个数字即是酶的标码。")
16
(一)习惯命名 根据 作用底物(S):淀粉酶 蛋白酶 反应性质 :脱氢酶 转氨酶 两者结合 :乳酸脱氢酶 谷丙转氨酶 来源 :胃蛋白酶 木瓜蛋白酶 (二)系统命名 命名原则 底物1:底物2 反应性质 酶 “L-乳酸:NAD+氧化还原酶”—乳酸脱氢酶 国际酶学委员会建议双命名法 系统名 习惯名

17
据化学组成: 1.单纯蛋白酶类: 只是由氨基酸组成,此外不含其他成分,其活性仅仅决定于它的蛋白质结构。
2.结合蛋白酶类:另一些酶如转氨酶、碳酸酐酶、脱氢酶等均属于结合蛋白质。这些酶其蛋白质部分称之为酶蛋白,非蛋白质部分称为辅因子(或辅助因子)。酶蛋白与辅因子单独存在时,均无催化活力,或活性很弱,结合在一起组成全酶后,才表现有明显的催化作用。 全酶 = 酶蛋白 + 辅因子(金属离子、辅酶、辅基) (有性)(无活性)(无活性) 辅酶、辅基往往是由维生素参与形成的小分子有机物
。酶蛋白与辅因子单独存在时,均无催化活力,或活性很弱,结合在一起组成全酶后,才表现有明显的催化作用。 全酶 = 酶蛋白 + 辅因子(金属离子、辅酶、辅基) (有性)(无活性)(无活性) 辅酶、辅基往往是由维生素参与形成的小分子有机物.")
18
维生素: 维生素是维持机体生命活动不可缺少的一类小分子化合物,它既不是生物体构成成分,也不是能量物质,之所以对生命活动如此重要,是因为维生素是辅酶或辅基的组成成分,参与体内代谢过程。 维生素的特点: 1、种类多 2、需要量少 3、常常需要从食物中获得 4、大部分充当辅酶 维生素 脂溶性维生素 水溶性维生素

19
脂溶性维生素: A D E K 维生素 硫辛酸(氧化型) 水溶性维生素: Vc VB: B1 B2 B3(泛酸) B5(PP) B12(氰钴胺素) B6 (吡哆醇/醛/胺) B7 (生物素 ) B11(叶酸) 硫辛酸(还原型)
 B6 (吡哆醇/醛/胺) B7 (生物素 ) B11(叶酸) 硫辛酸(还原型)")
20
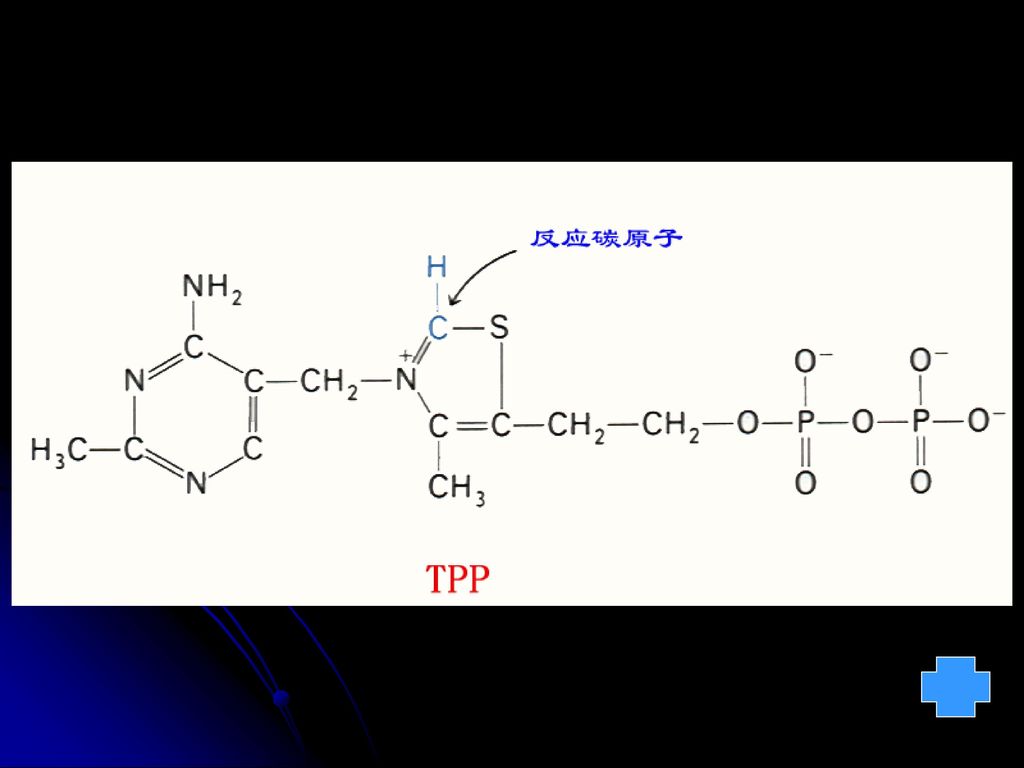
一、维生素B1 —硫胺(素)(抗神经炎V、抗脚气病V)
缺乏症:1. 脚气病 2. 中枢神经和肠胃患糖代谢失常 NH2 H3C CH2 N —C—CH3 HC C—CH2CH2OH S Cl P P 焦磷酸硫胺素(TTP) 硫胺素(B1) 功能: 以辅酶方式参加糖的分解代谢。TPP是脱羧酶、脱氢酶的辅酶。 分布:在米糠、麦麸、黄豆、酵母、瘦肉等食物中含量最丰富。
 硫胺素(B1) 功能: 以辅酶方式参加糖的分解代谢。TPP是脱羧酶、脱氢酶的辅酶。 分布:在米糠、麦麸、黄豆、酵母、瘦肉等食物中含量最丰富。")
21
二 维生素B2(又称核黄素) 1、维生素B2的生理功能: 是作为递氢辅酶,参与生物氧化作用。 2、维生素B2每人每天需要量:
儿童0.6mg,成人1.6mg。动物体内不能合成维生素B2。过量则排出。 3、膳食中长期缺乏: 口角炎、舌炎、唇炎、眼睑炎,角膜血管增生等症状。 4、分布: 广泛存在于动、植物中。在酵母、肝肾、蛋黄、奶及大豆中含量丰富。

22
三 泛酸(VB3)(遍多酸) 1、结构: 是β-丙氨酸与α,γ-二羟-β,β-二甲基丁酸结合而成的化合物。 2、泛酸的生物功能:
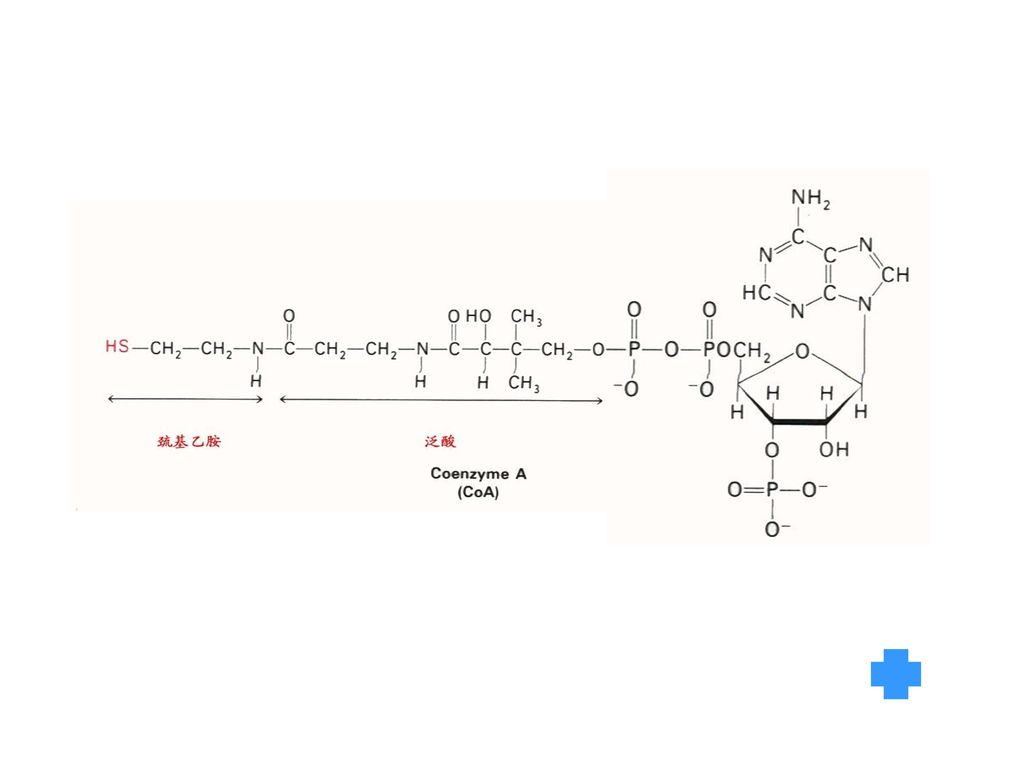
是以CoA形式参加代谢,是酰基的载体,是体内酰化酶的辅酶,对糖、脂、蛋白质代谢过程中的乙酰基转移有重要作用。 3、成人每天需要量为5—10mg,一般膳食的泛酸含量丰富。 未发现典型的缺乏症。

23
四 维生素PP(VB5)(抗癞皮病V) 包括尼克酸(烟酸)和尼克酰胺。尼克酰胺为维生素B5的化学名
N COOH N CONH2 尼克酸 尼克酰胺 分布:广泛存在于自然界,以酵母、花生、谷类、豆类、肉类和动物肝中含量丰富。

24
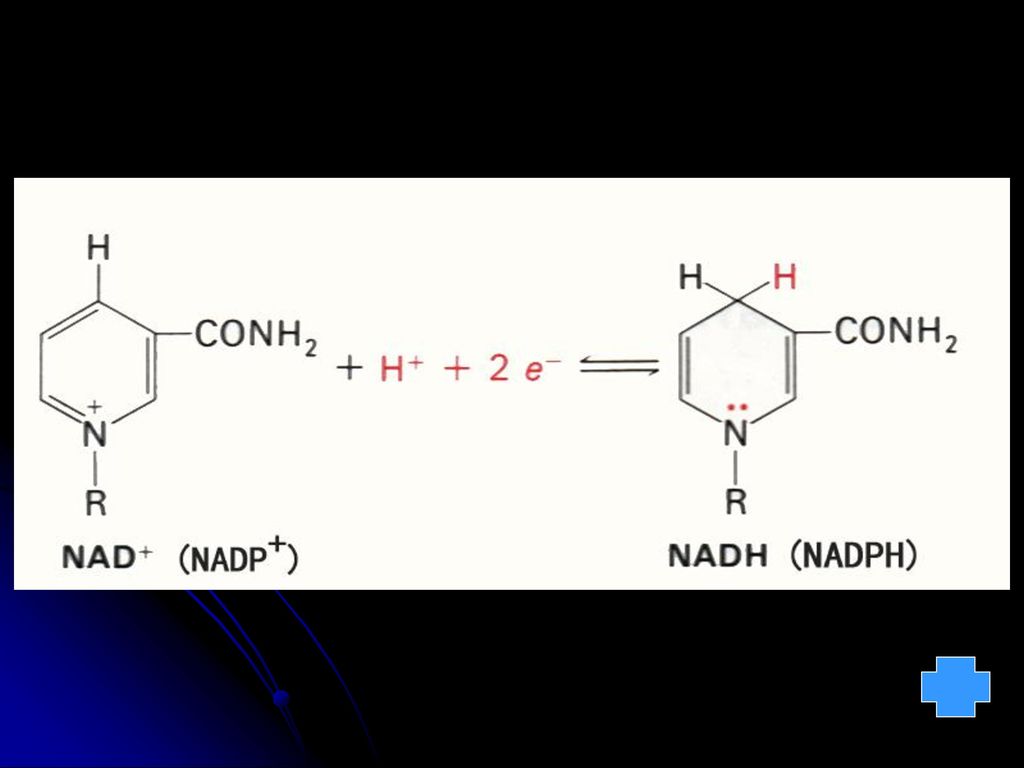
功能:以NAD+或NADP+形式作为脱氢酶的辅酶而起到递氢体的作用。
CONH2 R N CONH2 R H +2H -2H NAD(P)+ +2H -2H NAD(P)H + H+ 缺乏症:膳食中长期缺乏维生素PP所引起的疾病为对称性皮炎,又叫赖皮病。在狗生黑舌病。 Trp可转变为尼克酰胺,以玉米为主食易患缺乏症。
+ +2H. -2H. NAD(P)H + H+ 缺乏症:膳食中长期缺乏维生素PP所引起的疾病为对称性皮炎,又叫赖皮病。在狗生黑舌病。 Trp可转变为尼克酰胺,以玉米为主食易患缺乏症。")
25
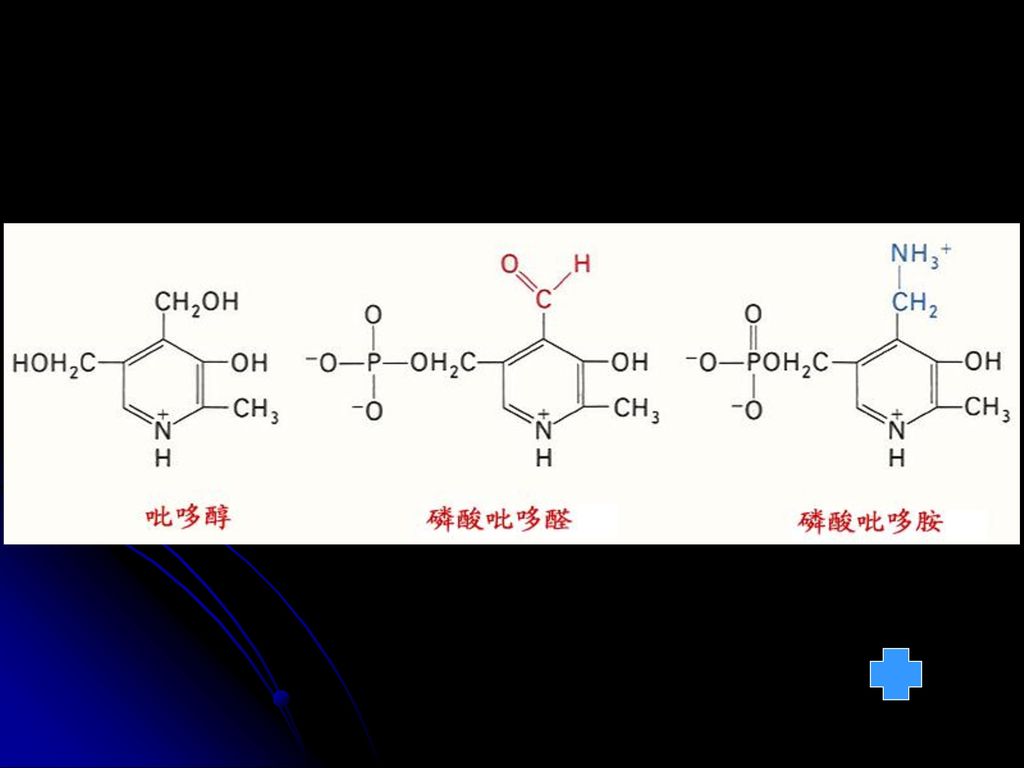
五 维生素B6 维生素B6又称吡哆素 包括吡哆醇、吡哆醛、吡哆胺。 吡哆胺 吡哆醛 吡哆醇 N CH2OH HO H3C N CH2OH
CH2NH2 HO H3C 吡哆胺 吡哆醛 吡哆醇

26
功能:作为辅酶参加多种代谢反应,包括脱羧、转氨、氨基酸内消旋、Trp代谢、含硫氨基酸的脱硫、羟基氨基酸的代谢和氨基酸的脱水等。
N CH2O— CHO HO H3C P (磷酸吡哆醛,PLP) 功能:作为辅酶参加多种代谢反应,包括脱羧、转氨、氨基酸内消旋、Trp代谢、含硫氨基酸的脱硫、羟基氨基酸的代谢和氨基酸的脱水等。 缺乏症:导致皮肤、中枢神经系统和造血机构的损害,可以引发皮炎。 分布:动植物中分布很广,谷类外皮含量尤为丰富。同时肠道细菌也能合成供人体需要
 功能:作为辅酶参加多种代谢反应,包括脱羧、转氨、氨基酸内消旋、Trp代谢、含硫氨基酸的脱硫、羟基氨基酸的代谢和氨基酸的脱水等。 缺乏症:导致皮肤、中枢神经系统和造血机构的损害,可以引发皮炎。 分布:动植物中分布很广,谷类外皮含量尤为丰富。同时肠道细菌也能合成供人体需要.")
27
六 生物素(VB7)(为含硫维生素) 功能:生物素是多种羧化酶的辅酶,在CO2固定反应中起重要作用。 缺乏症:人体一般不会发生生物素缺乏。
人类缺少生物素可能导致皮炎、肌肉疼痛、感觉过敏、怠倦、厌食、轻度贫血等。 分布:肝、肾、蛋黄、酵母、蔬菜和谷类中都含有。同时肠道细菌也能合成供人体需要

28
七 叶酸—(VB11) 由蝶呤啶、对氨基苯甲酸与L-谷氨酸连接而成
叶酸的5、6、7、8位置,在NADPH2存在下,可被还原成四氢叶酸(FH4或THFA)。四氢叶酸的N5 和N10位可与多种一碳单位结合作为它们的载体。 功能:THFA是转一碳基团酶系的辅酶。 缺乏症:叶酸缺乏时,红细胞的发育受到影响,造成巨红细胞性贫血症。 分布:肝、酵母、蔬菜中都含有。同时人肠道细菌也能合成,故一般不发生缺乏症。
。四氢叶酸的N5 和N10位可与多种一碳单位结合作为它们的载体。 功能:THFA是转一碳基团酶系的辅酶。 缺乏症:叶酸缺乏时,红细胞的发育受到影响,造成巨红细胞性贫血症。 分布:肝、酵母、蔬菜中都含有。同时人肠道细菌也能合成,故一般不发生缺乏症。")
29
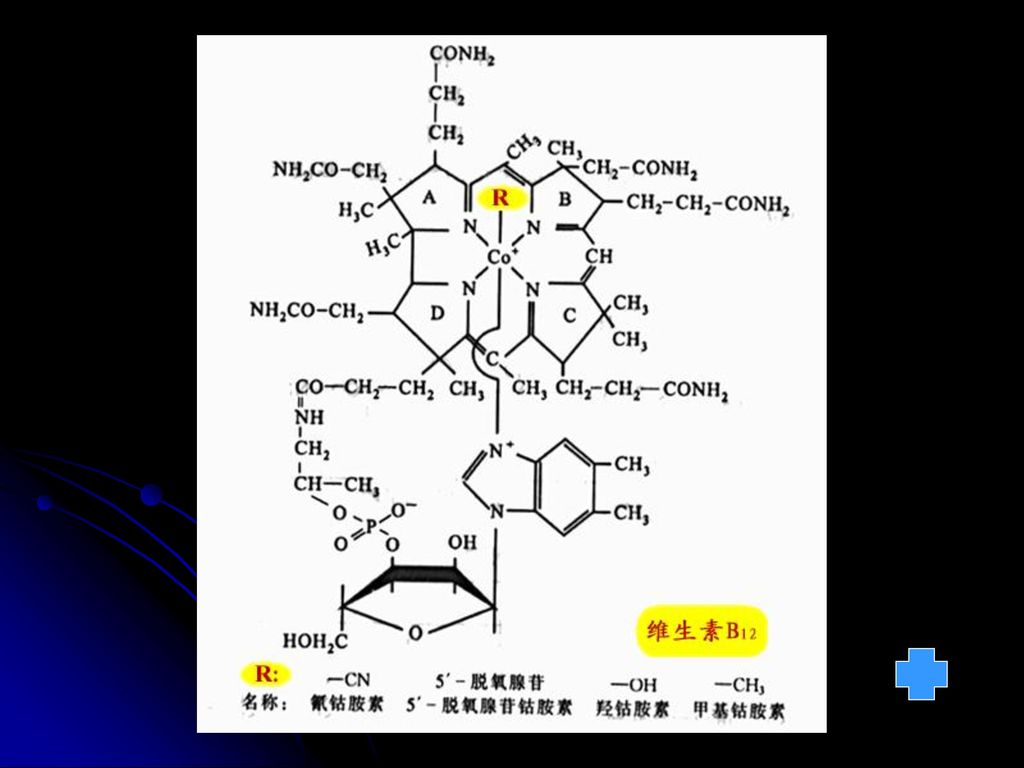
八 维生素B12(氰钴胺素) 维生素B12是含钴的化合物,又称钴胺素。 在自然界中只有微生物能合成维生素B12。
功能:1.促进甲基转移作用; 2.维持-SH的还原型状态 缺乏症: 1. 儿童及幼龄动物发育不良 2. 消化道上皮组织细胞失常 3. 造血器官功能失常,不能正常产生血红细胞,导致恶性贫血。 分布:动物性食品,特别是肉类和肝中含量丰富,人和动物的肠道细菌都能合成,一般不会缺少。

30
九 维生素C 维生素C能防治坏血病,又称抗坏血酸. 功能: 促进各种支持组织及细胞间粘合物的形成。是脯氨酸羟化酶的辅酶。
2. 对生物氧化有重要作用。 缺乏症: 坏血病,毛细血管易出血和齿、骨发育不全或退化

31
水溶性维生素

32
脂溶性V 十 维生素A 维生素A只存在于动物性食物中,包括A1 和 A2两种。A1即视黄醇,主要存在于咸水鱼的肝脏;A2即3-脱氢视黄醇,主要存在于淡水鱼肝脏。在高等植物和动物中普遍存在的β-胡萝卜素可转变为维生素A。 功能: 1、维生素A促进年幼动物生长,有助于动物生殖和泌乳。 2、维持上皮组织的健康及正常视觉, 缺乏症: 1、产生干眼病(眼结膜炎) 2、发生夜盲症
 2、发生夜盲症.")
33
十一 维生素D 维生素D具有抗佝偻病作用,又称抗佝偻病维生素。已确知有4种,即维生素D2、D3、D4、D5,均为类固醇衍生物,其中D2和D3较为重要。 功能:调节钙、磷代谢,维持血液正常的钙、磷浓度,从而促进钙化,使牙齿、骨骼发育正常。 缺乏症:维生素D摄食不足,不能维持钙的平衡,儿童骨骼发育不良,产生佝偻病。孕妇和授乳妇人的脱钙作用严重时导致骨质疏松症,患者骨骼易折,牙齿易脱落。

34
机体只能从胆汁排出过多的维生素D,维生素D如摄食过量则会中毒。
早期症状为:乏力、疲倦、恶心、头痛、腹泻等。 较严重时:引起软组织(包括血管、心肌、肺、肾、皮肤等)的钙化,导致重大病患。
的钙化,导致重大病患。")
35
十二 维生素E 维生素E又称生育酚或抗不育维生素,已知有8种,其中4种(α、β、γ、δ-生育酚)较为重要,α-生育酚的效价最高。动物组织的维生素E都是从食物中取得的。 缺乏症: 1. 生殖系统的上皮细胞毁坏,雄性睾丸退化,不产生精子,雌性流产或胎儿被溶化吸收。 肌肉(包括心肌)萎缩,形态改变,代谢反常。 3. 血胆固醇水平增高,红细胞破坏,发生贫血。 维生素E摄食过量无毒性。
萎缩,形态改变,代谢反常。 3. 血胆固醇水平增高,红细胞破坏,发生贫血。 维生素E摄食过量无毒性。")
36
十三 维生素K 维生素K是一类能促进血液凝固的萘醌衍生物。1929年发现。有K1、K2、K3三种,K1、K2为天然产物,K3为人工合成品。

37
种类 化学本质 功能 VA 不饱和一元醇 缺乏得干眼病、夜盲症 VD 类固醇衍生物 与Ca、P代谢有关,佝偻病
种类 化学本质 功能 VA 不饱和一元醇 缺乏得干眼病、夜盲症 VD 类固醇衍生物 与Ca、P代谢有关,佝偻病 VE 生育酚 与生育有关,抗氧化 VK 奈醌衍生物 促进血液凝固

38
(一)辅酶、辅基 根据它们与蛋白质结合的松紧程度不同分: 1.辅酶:指与酶蛋白结合比较松弛的小分子有机物质,通过透析方法可以除去。
如:辅酶Ⅰ 和辅酶Ⅱ 等。 2.辅基:是以共价键和酶蛋白结合,结合的较紧密,不能通过透析法除去,需要经过一定的化学处理才能与酶蛋白分开。 如:细胞色素氧化酶中的铁卟啉等

39
1.TPP:焦磷酸硫胺素(前体:硫胺素,含B1)
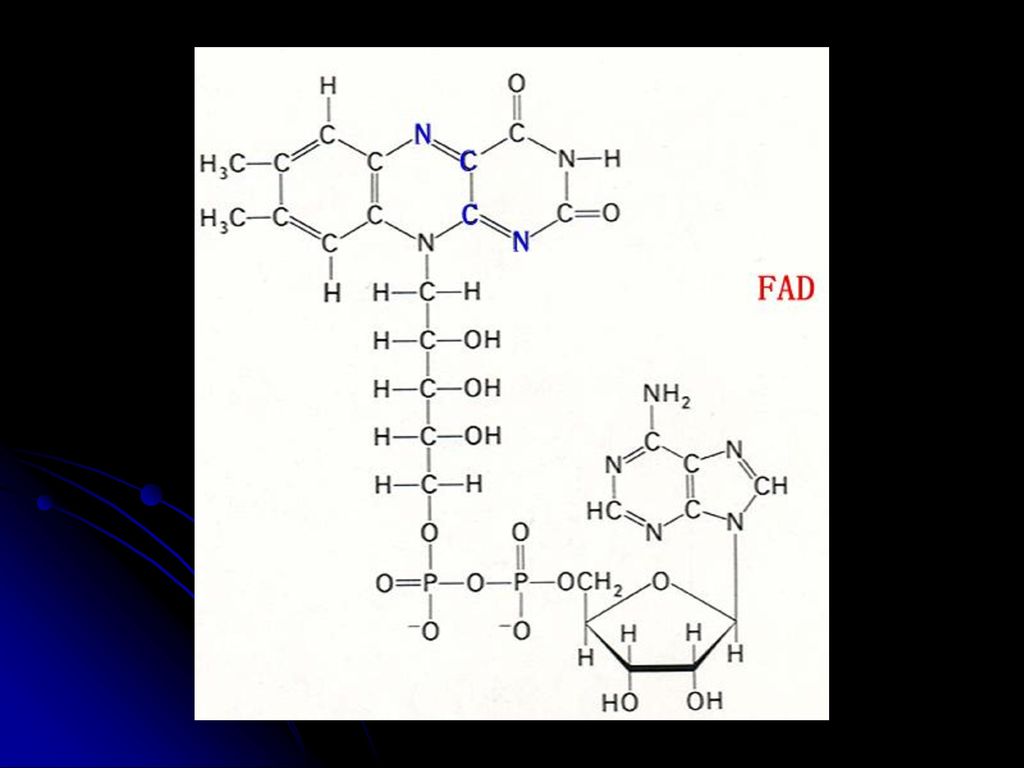
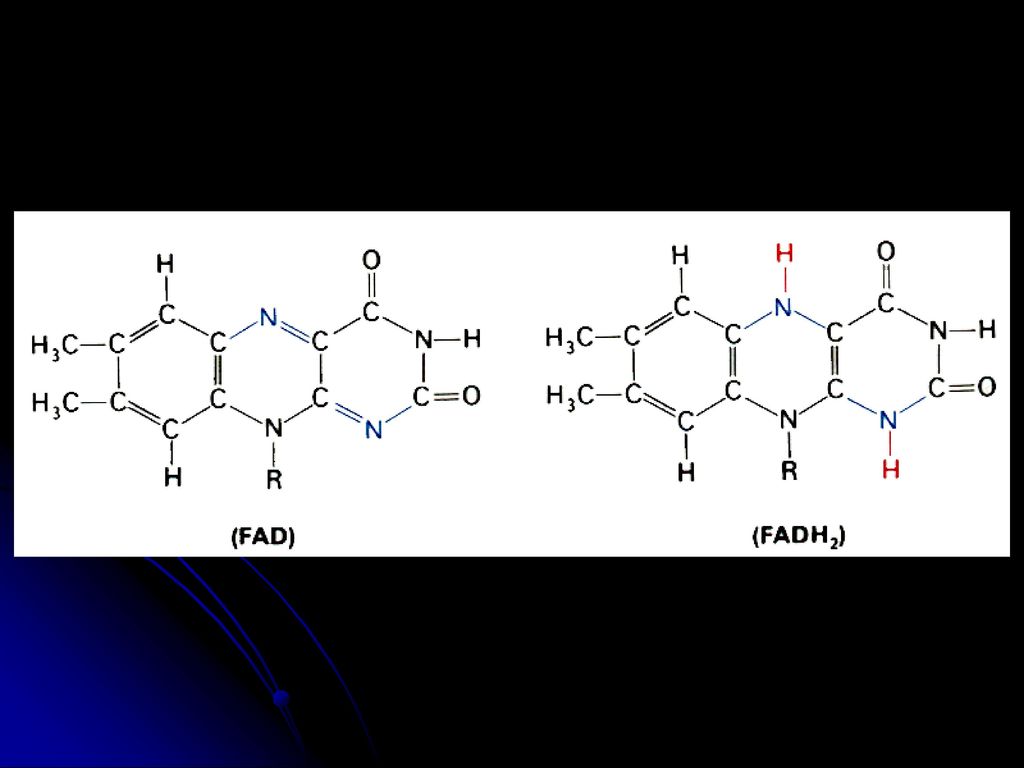
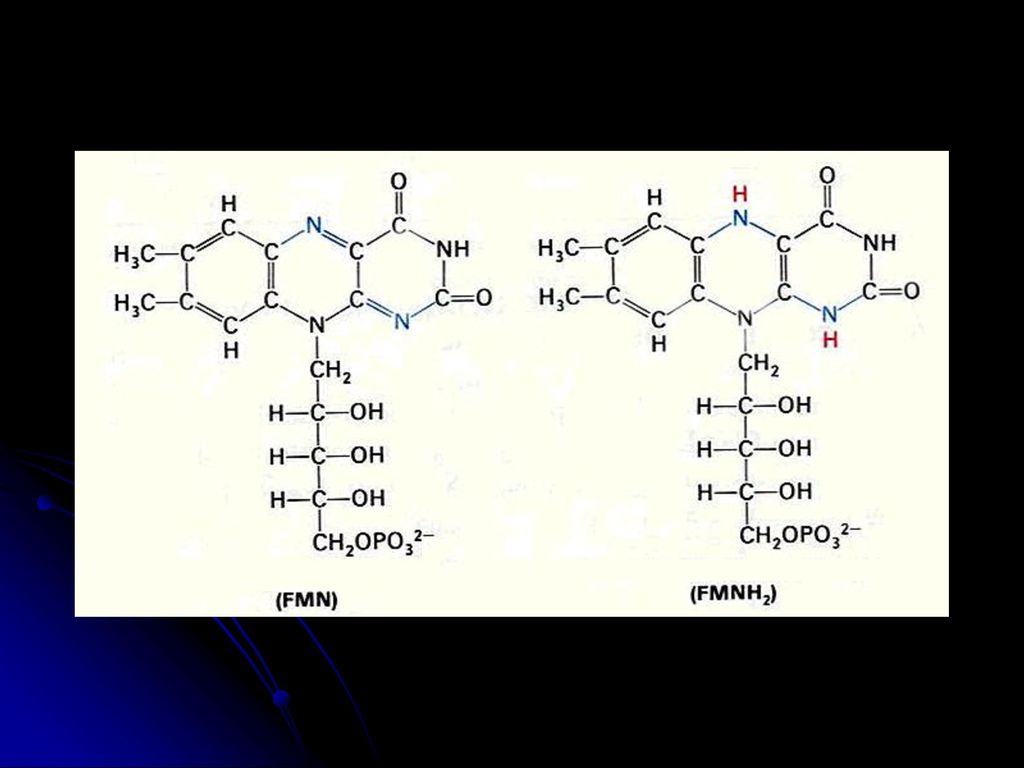
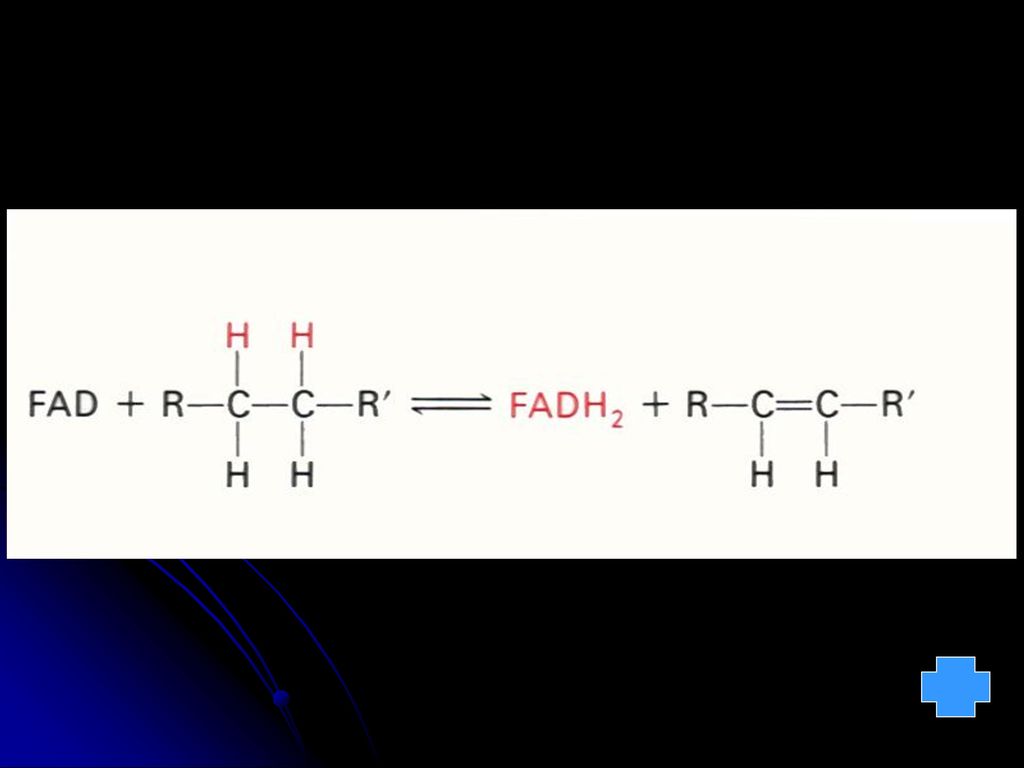
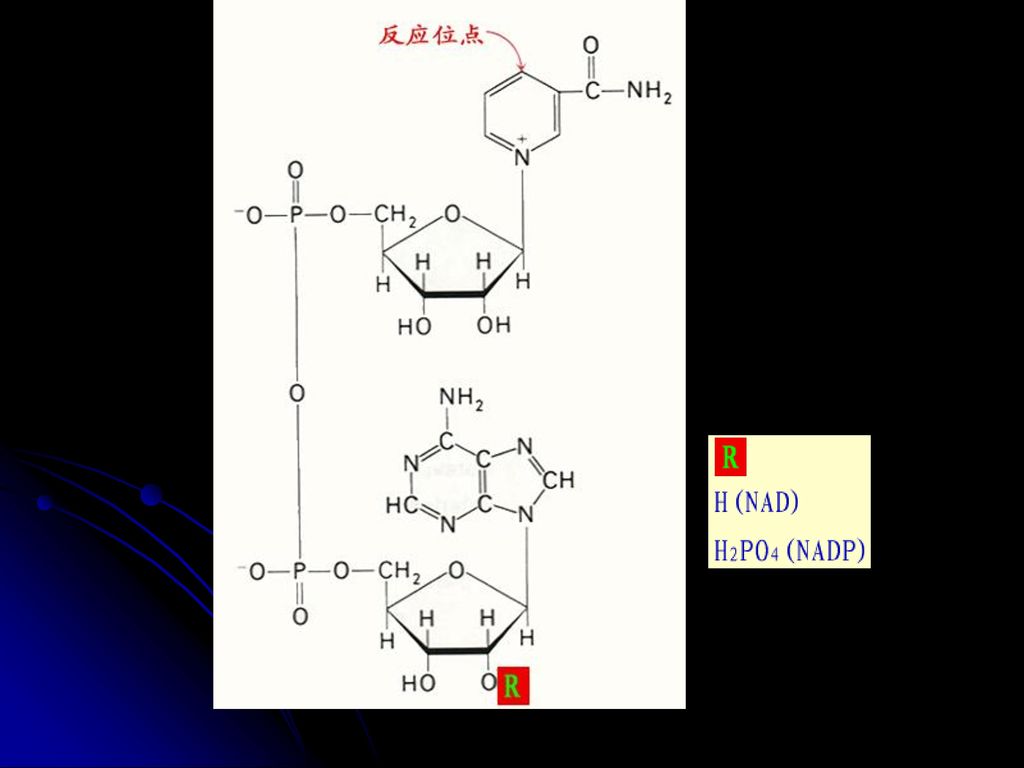
羧化酶的辅酶 与a-酮酸氧化脱羧有关 2.FMN FAD(核黄素的衍生物,含B2) 氧化还原酶的辅基(传递电子和质子) FMN FMNH2 FAD FADH2 3.NAD NADP(烟酰胺和烟酸衍生物,含B5) 脱氢酶的辅酶(传递电子和质子) 亦称:辅酶Ⅰ 和辅酶Ⅱ NAD++(NADP+) NADH(NADPH)+H+ 见图 见图 见图
 氧化还原酶的辅基(传递电子和质子) FMN FMNH2. FAD FADH2. 3.NAD NADP(烟酰胺和烟酸衍生物,含B5) 脱氢酶的辅酶(传递电子和质子) 亦称:辅酶Ⅰ 和辅酶Ⅱ. NAD++(NADP+) NADH(NADPH)+H+ 见图. 见图. 见图.")
47
4.CoASH(泛酸,含B3) 见图 酰基转移酶的辅酶(酰基载体) 5.磷酸吡哆醛(胺)(含B6) 见图 转氨和脱羧过程中的辅酶
转氨:通过醛、胺转化 6.氰钴胺素(B12) 亦称辅酶B12 变位酶的辅酶,也参与甲基及 其它一碳单位的转移反应 见图 见图
 亦称辅酶B12. 变位酶的辅酶,也参与甲基及. 其它一碳单位的转移反应. 见图. 见图.")
51
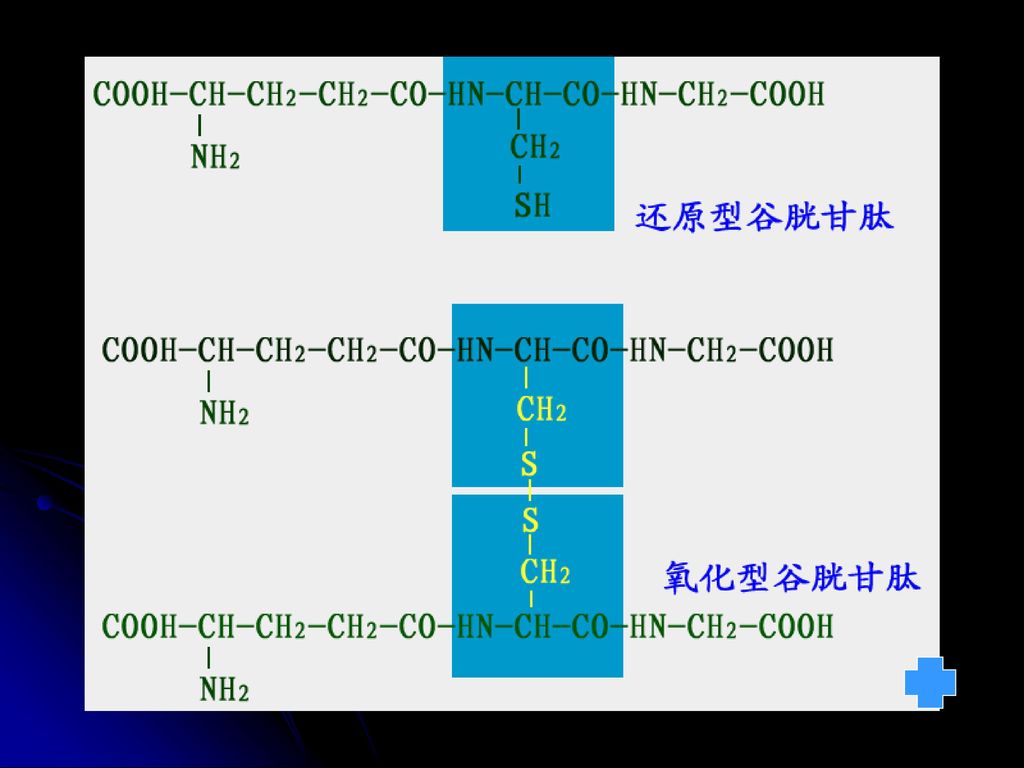
见图 见图 见图 7.硫辛酸 丙酮酸和α-酮戊二酸脱氢酶复合体的辅酶,在氧化脱羧过程中起着传递氢和酰基的作用 8.GSH 递氢
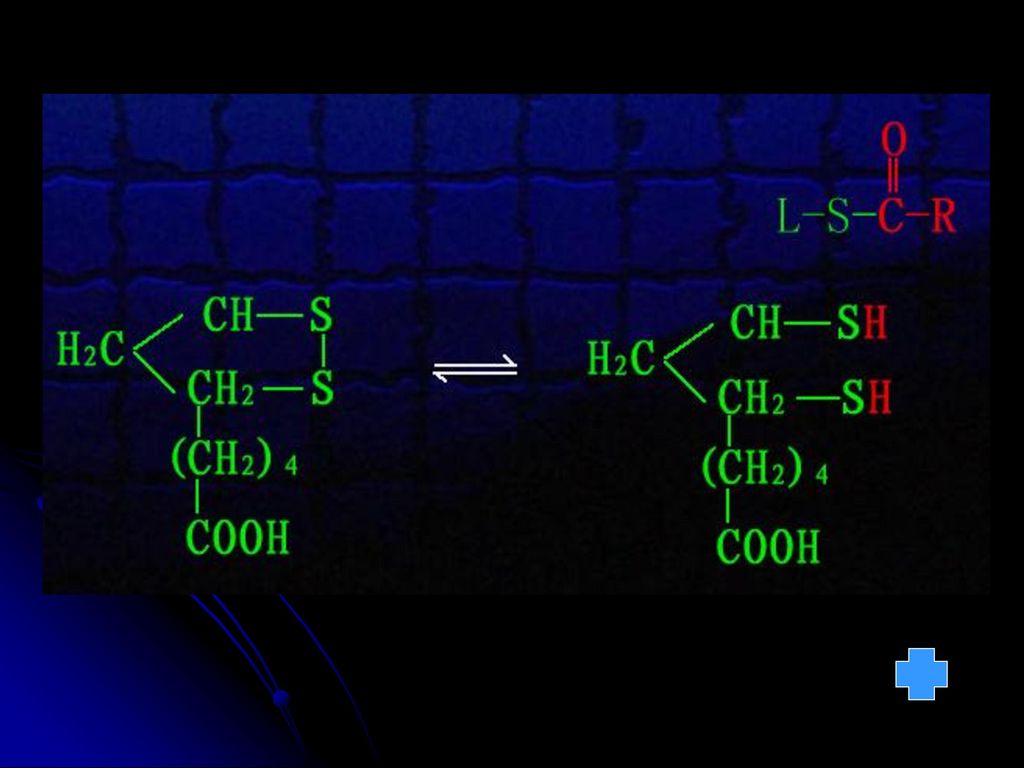
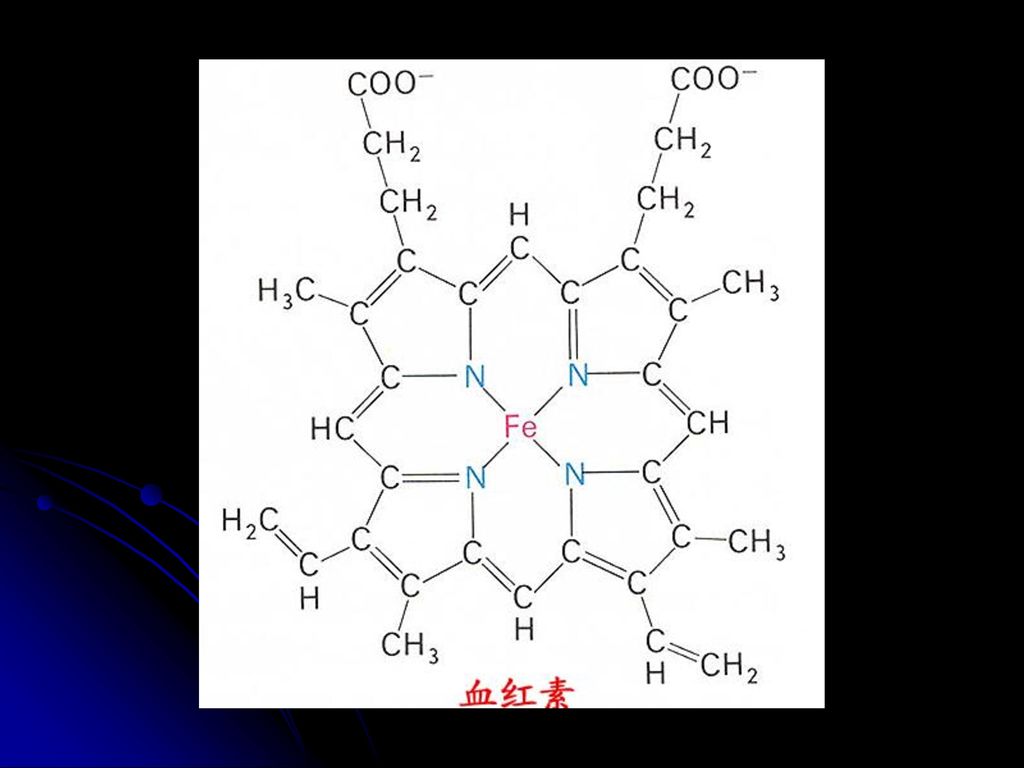
2GSH GSSG 9.生物素(含B7) 许多羧化酶的辅酶,与专一性的酶蛋白结合而参与羧化反应 10.四氢叶酸(FH4) 转一碳单位的辅酶(甲基、亚甲基、次甲基、甲酰基等 11.铁卟啉(血红素) 递电子 Fe Fe2+ 见图 见图 见图
 许多羧化酶的辅酶,与专一性的酶蛋白结合而参与羧化反应. 10.四氢叶酸(FH4) 转一碳单位的辅酶(甲基、亚甲基、次甲基、甲酰基等. 11.铁卟啉(血红素) 递电子. Fe3+ Fe2+ 见图. 见图. 见图.")
55
维生素及辅酶类型

56
(二)金属离子 分类: (据结合程度不同) 作用: 1.金属酶:酶蛋白与金属离子结合紧密。主要是一些过渡金属离子。
2.金属激酶:金属离子与酶的结合一般较松散。在溶液中,酶与这类离子结合而被激活。主要是一些碱金属离子或碱土金属离子 作用: 1.活性中心的组成成分 如:多酚氧化酶中的酮 2.在E与S之间起作用 如:羧肽酶中的锌 3.稳定结构

57
1.单体酶: 一条多肽链组成的酶,种类较少,多是催化水解反应的酶。分子量 1.3万—3.5万。 2.寡聚酶: 由两个或两个以上的亚基组成的酶,分子量3.5万—几百万。相当数量的寡聚酶是调节酶。 3.多酶复合物: 功能相关的几个酶靠非共价键彼此嵌合而成的聚合体,所有反应依次连接,有利于一系列反应的连续进行。分子量几百万以上。

58
四、酶的结构组成 一、酶的结构 结合部位 活性中心 必需基团 催化部位 酶的结构 活性中心以外的必需基团 其它部分

59
(一)概念:酶的活性中心是指结合底物和将底物转化为产物的区域,通常是相隔很远的氨基酸残基形成的三维实体。
二.酶的活性基团及活性中心 (一)概念:酶的活性中心是指结合底物和将底物转化为产物的区域,通常是相隔很远的氨基酸残基形成的三维实体。 结合部位 中心内必需基团 活性中心 催化部位 调控基团 中心外必需基团
概念:酶的活性中心是指结合底物和将底物转化为产物的区域,通常是相隔很远的氨基酸残基形成的三维实体。 结合部位. 中心内必需基团. 活性中心. 催化部位. 调控基团. 中心外必需基团.")
60
1、结合部位 酶分子中与底物结合,使底物与酶的一定构象形成复合物的基团。 酶的结合基团决定酶反应的专一性。 2、催化部位 酶分子中催化底物发生化学反应并将其转变为产物的基团。 催化基团决定酶所催化反应的性质,同时也是决定反应的高效性。 3、调控基团 酶分子中一些可与其他分子发生某种程度的结合并引起酶分子空间构象的变化,对酶起激活或抑制作用的基团。

61
一些酶活性中心的基团

62
酶的活性中心示意图

63
第四节、酶的催化作用机理 一. 酶作用专一性机理 2.诱导契合学说—酶受底物诱导而变形 二. 酶作用高效性机理 三.影响酶高效性的因素
1.锁钥学说 2.诱导契合学说—酶受底物诱导而变形 二. 酶作用高效性机理 中间产物学说 三.影响酶高效性的因素 1.邻近定向效应 2.底物的形变和诱导契合 3.亲核催化/亲电催化(共价催化) 4.酸碱催化 5.微环境的影响
 4.酸碱催化. 5.微环境的影响.")
64
1、锁钥学说(1890): 认为整个酶分子的天然构象是具有刚性结构的,酶表面具有特定的形状。酶与底物的结合如同一把钥匙对一把锁一样。
此学说可以较好的解释酶的立体异构专一性;但不能解释:酶的多底物现象、酶对正反方向的催化等。

65
锁钥学说

66
2、诱导契合学说(1964年 ) 该学说认为酶的活性部位并不是和底物的形状正好互补的,而是在酶和底物结合的过程中,底物分子或酶分子,有时是两者的构象同时发生了一定的变化后才互补的,这时催化基团的位置也正好在所催化底物键的断裂和即将生成键的适当位置。这个动态的辨认过程称为诱导契合。
 该学说认为酶的活性部位并不是和底物的形状正好互补的,而是在酶和底物结合的过程中,底物分子或酶分子,有时是两者的构象同时发生了一定的变化后才互补的,这时催化基团的位置也正好在所催化底物键的断裂和即将生成键的适当位置。这个动态的辨认过程称为诱导契合。")
67
诱导契合学说 动画

68
诱导契合机制 酶与底物靠近 定向 酶与底物相互诱导变形 契合形成中间产物 产物脱离 A.靠近定向 定向 靠近 电性吸引疏水作用 底物 酶

69
B.诱导契合 活性中心催化基团进行催化 诱导 互补性结构变化 契合 能否契合— 专一性的由来

70
C.产物脱离 酶复原-催化剂

71
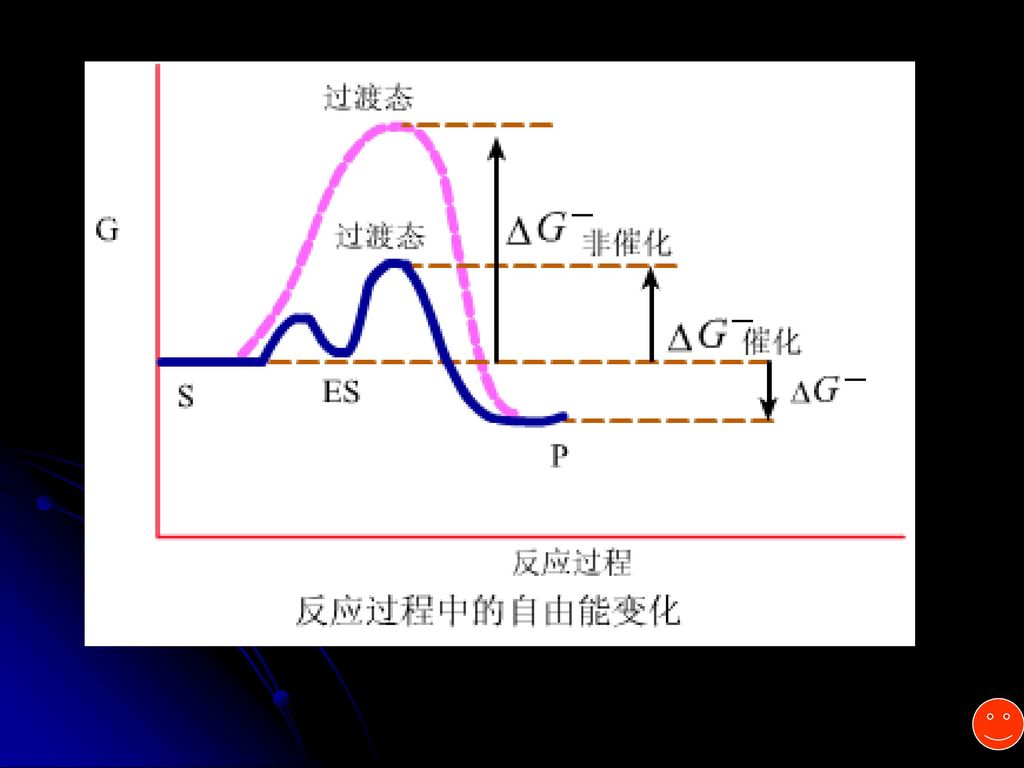
中间产物学说(过渡态学说) 非酶促反应 S P 酶促反应 S+E ES P+E 反应分两步走,活化能大大降低,因此反应容易进行。
S+E ES ES* EP P+E 过渡态 底物—酶中间物 产物—酶中间物 酶作用的实质在于降低反应活化能

73
1、 邻近效应、 定向效应 邻近效应:在酶促反应中,由于酶和底物分子之间的亲和性,底物分子有向酶的活性中心靠近的趋势,最终结合到酶的活性中心,使底物在酶活性中心的有效浓度大大增加的效应。 定向效应:当专一性底物向酶活性中心靠近时,会诱导酶分子构象发生改变,使酶活性中心的相关基团和底物的反应基团正确定向排列,同时使反应基团之间的分子轨道以正确方向严格定位,使酶促反应易于进行。 以上两种效应使酶具有高效率和专一性特点。

74
邻近定向效应

75
普通化学反应—随机碰撞(受浓度、碰撞角度影响)
相当于——社会上的自由恋爱 酶的活性中心—相当于“婚姻介绍所” 邻近作用提高了酶活性(“婚介”)中心的底物浓度(—非婚男女集中) 定向作用缩短了底物与催化基团间(“男女”)的距离 提高反应速度(成功率)108倍
中心的底物浓度(—非婚男女集中) 定向作用缩短了底物与催化基团间( 男女 )的距离. 提高反应速度(成功率)108倍.")
76
2.底物的形变和诱导契合 当酶与底物靠近时,不仅酶构象受底物作用而变化,底物分子也常常受酶作用而变化,也就是酶使底物分子中的敏感键发生“变形”,从而促使底物中的敏感键更易于破裂。因而更容易形成一个互相契合的酶—底的复合物而提高催化效率。

77
酶 底物 同时形变 产物 底物分子发生形变

78
3、酸碱催化 酸碱催化是通过瞬时的向反应物提供质子或从反应物接受质子以稳定过渡态,加速反应的一类催化机制。 酶分子中可以作为广义酸、碱的基团
广义酸基团 (质子供体) 广义碱基团(质子受体)
 广义碱基团(质子受体)")
79
4、共价催化 催化剂通过与底物形成反应活性很高的共价过渡产物,使反应活化能降低,从而提高反应速度的过程,称为共价催化。
它包括两种类型:亲核催化和亲电催化 亲核基团: His 的咪唑基,Cys 的硫基,Asp 的羧基,Ser 的羟基等; 亲电子基团:H+ 、Mg2+、 Mn2+ 、Fe3+ 某些辅酶,如焦磷酸硫胺素和磷酸吡哆醛等也可以参与共价催化作用。

80
5、活性部位微环境的影响(酶活性中心是低介电区域)
酶活性中心处于一个非极性环境中,其介电常数较在水介质(极性介质)中的介电常数低,在非极性环境中两个带电基团之间的静电作用比在极性环境中显著增高。从而有利于同底物的结合。
中的介电常数低,在非极性环境中两个带电基团之间的静电作用比在极性环境中显著增高。从而有利于同底物的结合。")
81
酶催化作用机理: 综上所述: 酶与底物结合时,由于酶的变形(诱导契合)或底物变形使二者相互适合,并依靠离子键、氢键、范德华力的作用和水的影响,结合成中间产物,在酶分子的非极性区域内,由于酶与底物的邻近、定向,使二者可以通过亲核\亲电催化、一般酸\碱催化或金属离子催化方式进行多元催化,从而大大降低反应所需的活化能,使酶促反应迅速进行。
或底物变形使二者相互适合,并依靠离子键、氢键、范德华力的作用和水的影响,结合成中间产物,在酶分子的非极性区域内,由于酶与底物的邻近、定向,使二者可以通过亲核\亲电催化、一般酸\碱催化或金属离子催化方式进行多元催化,从而大大降低反应所需的活化能,使酶促反应迅速进行。")
82
四、酶原与酶原的激活 (一)概念: 酶原——酶在生物体内首先合成出来的无活性前体。
酶原的激活——酶原必须在一定的条件下去掉一个或几个特殊的肽键,从而使酶的构象发生一定的变化,才有活性,这一过程称为酶原激活。 (二)激活的本质或实质 酶的活性中心形成或暴露的过程 意义:避免自身受损伤
激活的本质或实质. 酶的活性中心形成或暴露的过程. 意义:避免自身受损伤.")
83
胰蛋白酶的激活 动画 缬 缬 缬 缬 为什么不直接以酶形式存在呢?保护正常组织不受伤害 肠激酶(激活作用) 缬 天 赖 异 甘 组 S 丝
胰蛋白酶原 胰蛋白酶的激活 动画 活性中心 缬 天 赖 异 甘 组 S 丝 胰蛋白酶 缬 缬 为什么不直接以酶形式存在呢?保护正常组织不受伤害

84
第四节、影响酶促反应速度的因素 1.底物浓度对酶促反应速度的影响 在低底物浓度时: 反应速度与底物浓度成正比,表现为一级反应特征。

85
随着底物浓度的增高 反应速度不再成正比例加速; 反应为混合级反应。

86
当底物浓度达到一定值 反应速度达到最大值(Vmax),此时再增加底物浓度,反应速度不再增加,表现为零级反应。
,此时再增加底物浓度,反应速度不再增加,表现为零级反应。")
87
两者的关系可用中间中间产物学说来解释。

88
酶与底物的中间络合物学说 S+E ES P+E 底物浓度与酶反应速度关系的数学表达式: 米氏方程 基础:中间产物学说 米氏常数

89
前 提 ★ S与E形成中间产物,且整个反应速度取决于 ES P+E ★ 产物浓度接近为0(初速度) ★ [S]>>[E]
★ 反应达到平衡
![前 提 ★ S与E形成中间产物,且整个反应速度取决于 ES P+E ★ 产物浓度接近为0(初速度) ★ [S]>>[E]](http://slidesplayer.com/slide/11183160/60/images/89/%E5%89%8D+%E6%8F%90+%E2%98%85+S%E4%B8%8EE%E5%BD%A2%E6%88%90%E4%B8%AD%E9%97%B4%E4%BA%A7%E7%89%A9%EF%BC%8C%E4%B8%94%E6%95%B4%E4%B8%AA%E5%8F%8D%E5%BA%94%E9%80%9F%E5%BA%A6%E5%8F%96%E5%86%B3%E4%BA%8E+ES+P%2BE+%E2%98%85+%E4%BA%A7%E7%89%A9%E6%B5%93%E5%BA%A6%E6%8E%A5%E8%BF%91%E4%B8%BA0%EF%BC%88%E5%88%9D%E9%80%9F%E5%BA%A6%EF%BC%89+%E2%98%85+%5BS%5D%3E%3E%5BE%5D.jpg "★ 反应达到平衡.")
90
Vmax[S] 米氏常数Km Km + [S] 米氏常数 :当v=1/2 Vmax 时的底物浓度 Km=[S]
Km值:是当酶促反应速度达到最大反应速度一半时的底物浓度,它的单位是mol/L,与底物浓度的单位一样。
![Vmax[S] 米氏常数Km Km + [S] 米氏常数 :当v=1/2 Vmax 时的底物浓度 Km=[S]](http://slidesplayer.com/slide/11183160/60/images/90/Vmax%5BS%5D+%E7%B1%B3%E6%B0%8F%E5%B8%B8%E6%95%B0Km+Km+%2B+%5BS%5D+%E7%B1%B3%E6%B0%8F%E5%B8%B8%E6%95%B0+%EF%BC%9A%E5%BD%93v%3D1%2F2+Vmax+%E6%97%B6%E7%9A%84%E5%BA%95%E7%89%A9%E6%B5%93%E5%BA%A6+Km%3D%5BS%5D.jpg "Km值:是当酶促反应速度达到最大反应速度一半时的底物浓度,它的单位是mol/L,与底物浓度的单位一样。")
91
关于米氏常数Km的几点说明: a.不同的酶具有不同Km值,它是酶的一个重要的特征物理常数,只与酶的性质有关,而与其浓度无关。
b.Km值只是在固定的底物,一定的温度和pH条件下,一定的缓冲体系中测定的,不同条件下具有不同的Km值。 C.一般情况下,1/Km可以近似地表示酶对底物的亲和力大小, 1/Km愈大,表明亲和力愈大。 (同一种酶有几种底物就有几个Km值,其中Km值最小的底物一般称为该酶的最适底物或天然底物)
")
92
米氏常数的求法 双倒数作图法 1 Km 1 1 斜率=Km/Vmax 1/Vmax = +
V Vmax [S] Vmax

93
2. pH对酶促反应速度的影响 酶反应速度最大时的溶液pH,称为酶的最适pH pH值影响酶活力的原因有以下几点 影响酶分子构象的稳定性。
影响酶分子(包括辅因子)极性基团的解离状态,使其荷电性发生变化 c. 影响底物分子的解离状态
极性基团的解离状态,使其荷电性发生变化. c. 影响底物分子的解离状态.")
94
3. 温度的影响 使酶促反应速度达最大时的温度称为酶的最适温度。 温度对酶促反应速度的影响:
a.一方面是温度升高,酶促反应速度加快(温度系数Q10:反应温度提高10 C,其反应速度与原来的反应速度之比。 Q10多为1-2 ) 。 b.另一方面,温度升高,酶的高级结构将发生变化或变性,导致酶活性降低甚至丧失,反应速度很快下降。
 。 b.另一方面,温度升高,酶的高级结构将发生变化或变性,导致酶活性降低甚至丧失,反应速度很快下降。")
95
4.酶浓度对酶反应速度的影响 在一定条件下酶促反应的速度与酶的浓度成正比。
当底物浓度大大超过酶浓度时,反应达到最大速度。如果此时增加酶的浓度可增加反应速度,酶促反应的速度与酶的浓度成正比关系。

96
5.激活剂对酶反应速度的影响 凡能提高酶活性的物质,都称为激活剂
(1)无机离子:金属离子(K+ Na+ Mg2+ Zn2+ Fe2+ Ca2+)、阴离子(Cl- Br-)、氢离子 (2)简单有机分子:某些还原剂、乙二胺四乙酸(EDTA) (3)具有蛋白质性质的大分子物质 主要是激活酶原 无活性的酶原 有活性的酶 激活作用
无机离子:金属离子(K+ Na+ Mg2+ Zn2+ Fe2+ Ca2+)、阴离子(Cl- Br-)、氢离子. (2)简单有机分子:某些还原剂、乙二胺四乙酸(EDTA) (3)具有蛋白质性质的大分子物质. 主要是激活酶原. 无活性的酶原 有活性的酶. 激活作用.")
97
6. 抑制剂对酶反应速度的影响 (1)抑制作用与抑制剂 (2)抑制作用的类型 (3)可逆抑制作用的动力学特征 (4)一些重要的抑制剂
抑制作用与抑制剂 (2)抑制作用的类型 (3)可逆抑制作用的动力学特征 (4)一些重要的抑制剂")
98
(1)抑制作用与抑制剂 凡使酶的活性降低或丧失,但并不引起酶蛋白变性的作用称为抑制作用。主要是由于酶的必需基团化学性质的改变而引起的。 (抑制作用不同于失活作用) 能够引起抑制作用的化合物则称为抑制剂。 (抑制剂不同于变性剂)
")
99
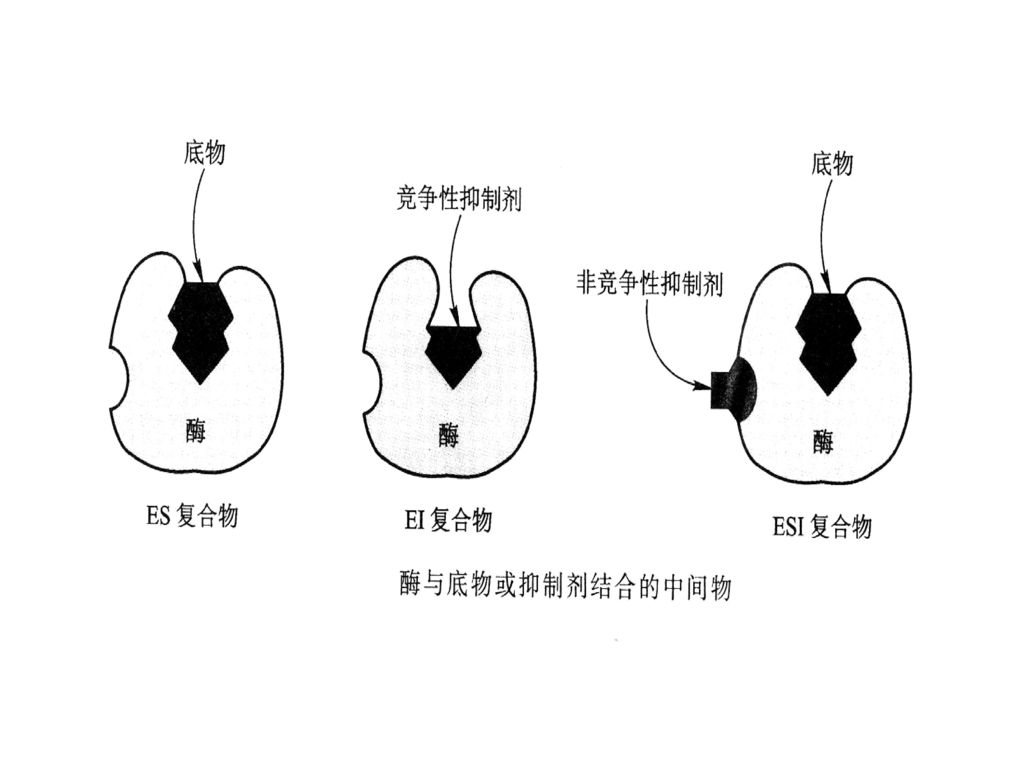
竞争性抑制 非竞争性抑制 (2) 抑制作用的类型 a. 不可逆抑制作用 专一性不可逆抑制作用 b. 可逆抑制作用 非专一性不可逆抑制作用
反竞争性抑制

100
定义:抑制剂与酶的活性中心的功能基团共价结合而抑制酶的活性,不能用透析或超滤等物理方法除去抑制剂而恢复酶活性。
不可逆抑制作用(修饰抑制) 定义:抑制剂与酶的活性中心的功能基团共价结合而抑制酶的活性,不能用透析或超滤等物理方法除去抑制剂而恢复酶活性。 专一性不可逆抑制作用:这类抑制剂只作用于与酶活性部位有关的氨基酸残基或一类酶。 非专一性不可逆抑制作用:这类抑制剂作用于酶分子上一类或几类不同的基团或作用于几类不同的酶。 如:酰化剂酸酐和磺酰氯等可使酶蛋白的-OH、SH、NH2等发生酰化而使酶失活。
 定义:抑制剂与酶的活性中心的功能基团共价结合而抑制酶的活性,不能用透析或超滤等物理方法除去抑制剂而恢复酶活性。 专一性不可逆抑制作用:这类抑制剂只作用于与酶活性部位有关的氨基酸残基或一类酶。 非专一性不可逆抑制作用:这类抑制剂作用于酶分子上一类或几类不同的基团或作用于几类不同的酶。 如:酰化剂酸酐和磺酰氯等可使酶蛋白的-OH、SH、NH2等发生酰化而使酶失活。")
101
抑制剂与酶蛋白以非共价方式结合,引起酶活性暂时性丧失。抑制剂可以通过透析等物理方法被除去,并且能部分或全部恢复酶的活性。
b. 可逆抑制作用 抑制剂与酶蛋白以非共价方式结合,引起酶活性暂时性丧失。抑制剂可以通过透析等物理方法被除去,并且能部分或全部恢复酶的活性。 根椐抑制剂与酶结合的情况,又可以分为三类 b1 竟争性抑制 b2 非竟争性抑制 b3 反竞争性抑制

102
b1竟争性抑制 某些抑制剂的化学结构与底物相似,因而能与底物竟争与酶活性中心结合。当抑制剂与活性中心结合后,底物被排斥在反应中心之外,其结果是酶促反应被抑制了。 S 竟争性抑制通常可以通过增大底物浓度,即提高底物的竞争能力来消除。

103
例:丙二酸和戊二酸对琥珀酸脱氢酶结合 但不能催化脱氢

104
竞争性可逆抑制图示 I

105
b2 非竟争性抑制 酶可同时与底物及抑制剂结合,即底物和抑制剂没有竞争作用。酶与抑制剂结合后,还可与底物结合;酶与底物结合后,也可再结合抑制剂,但是三元的中间产物不能进一步分解为产物,所以酶活性降低。 非竞争性抑制剂与酶活性中心以外的基团结合。这类抑制作用不会因提高底物浓度而减弱

106
非竞争性可逆抑制图示 非竞争

108
b3 反竞争性抑制 酶只有与底物结合后才与抑制剂结合,形成的三元中间产物不能进一步分解为产物,所以酶活性降低。这类抑制作用最不重要。
反竞争性抑制作用常见于多底物反应中,而在单底物反应中比较少见。

109
(3)可逆抑制作用的动力学 a. 竞争性抑制 加入竞争性抑制剂后,Km 变大,酶促最大反应速度Vmax不变。 1/v 1/[s]
无抑制剂 1/[s]
![(3)可逆抑制作用的动力学 a. 竞争性抑制 加入竞争性抑制剂后,Km 变大,酶促最大反应速度Vmax不变。 1/v 1/[s]](http://slidesplayer.com/slide/11183160/60/images/109/%EF%BC%883%EF%BC%89%E5%8F%AF%E9%80%86%E6%8A%91%E5%88%B6%E4%BD%9C%E7%94%A8%E7%9A%84%E5%8A%A8%E5%8A%9B%E5%AD%A6+a.+%E7%AB%9E%E4%BA%89%E6%80%A7%E6%8A%91%E5%88%B6+%E5%8A%A0%E5%85%A5%E7%AB%9E%E4%BA%89%E6%80%A7%E6%8A%91%E5%88%B6%E5%89%82%E5%90%8E%EF%BC%8CKm+%E5%8F%98%E5%A4%A7%EF%BC%8C%E9%85%B6%E4%BF%83%E6%9C%80%E5%A4%A7%E5%8F%8D%E5%BA%94%E9%80%9F%E5%BA%A6Vmax%E4%B8%8D%E5%8F%98%E3%80%82+1%2Fv+1%2F%5Bs%5D.jpg "无抑制剂. 1/[s]")
110
b.非竞争性抑制 1/v 非竞争性抑制剂 加入非竞争性抑制剂后,Km 不变,而Vmax减小。 1/Vmax 无抑制剂 -1/km 1/[s]
![b.非竞争性抑制 1/v 非竞争性抑制剂 加入非竞争性抑制剂后,Km 不变,而Vmax减小。 1/Vmax 无抑制剂 -1/km 1/[s]](http://slidesplayer.com/slide/11183160/60/images/110/b.%E9%9D%9E%E7%AB%9E%E4%BA%89%E6%80%A7%E6%8A%91%E5%88%B6+1%2Fv+%E9%9D%9E%E7%AB%9E%E4%BA%89%E6%80%A7%E6%8A%91%E5%88%B6%E5%89%82+%E5%8A%A0%E5%85%A5%E9%9D%9E%E7%AB%9E%E4%BA%89%E6%80%A7%E6%8A%91%E5%88%B6%E5%89%82%E5%90%8E%EF%BC%8CKm+%E4%B8%8D%E5%8F%98%EF%BC%8C%E8%80%8CVmax%E5%87%8F%E5%B0%8F%E3%80%82+1%2FVmax+%E6%97%A0%E6%8A%91%E5%88%B6%E5%89%82+-1%2Fkm+1%2F%5Bs%5D.jpg "b.非竞争性抑制 1/v 非竞争性抑制剂 加入非竞争性抑制剂后,Km 不变,而Vmax减小。 1/Vmax 无抑制剂 -1/km 1/[s]")
111
C.反竞争性抑制 加入反竞争性抑制剂,使 Km和Vmax均减小 1/ [I]增加 1/[S] -1/km(1+[I]/ki)
无抑制剂 1/[S] -1/km(1+[I]/ki)
![C.反竞争性抑制 加入反竞争性抑制剂,使 Km和Vmax均减小 1/ [I]增加 1/[S] -1/km(1+[I]/ki)](http://slidesplayer.com/slide/11183160/60/images/111/C.%E5%8F%8D%E7%AB%9E%E4%BA%89%E6%80%A7%E6%8A%91%E5%88%B6+%E5%8A%A0%E5%85%A5%E5%8F%8D%E7%AB%9E%E4%BA%89%E6%80%A7%E6%8A%91%E5%88%B6%E5%89%82%EF%BC%8C%E4%BD%BF+Km%E5%92%8CVmax%E5%9D%87%E5%87%8F%E5%B0%8F+1%2F%EF%81%AE+%5BI%5D%E5%A2%9E%E5%8A%A0+1%2F%5BS%5D+-1%2Fkm%281%2B%5BI%5D%2Fki%29.jpg "无抑制剂. 1/[S] -1/km(1+[I]/ki)")
112
可逆抑制的动力学比较 抑制类型 Vmax Km 无抑制剂 Vmax Km 竞争性抑制 不变 变大 非竞争性抑制 变小 不变
竞争性抑制 不变 变大 非竞争性抑制 变小 不变 反竞争性抑制 变小 变小

113
(4)一些重要的抑制剂 a.不可逆抑制剂 1.有机磷化合物—与酶活性直接有关的丝氨酸上的-OH牢固地结合,从而抑制某些蛋白酶或酯酶。(敌百虫、敌敌畏、农药1605等) 2.有机汞、有机砷化合物—与酶蛋白上的-SH作用,从而抑制含-SH酶的活性。 (对氯汞苯甲酸)
")
114
3.氰化物、硫化物和CO—这类物质能与酶中的金属离子形成较为稳定的络合物,使酶的活性受到抑制。
4.重金属—Ag、Cu、Hg等盐能使大多数酶失活,加入EDTA可以除去。 5.烷化剂—这一类试剂中往往含一个活泼的卤素原子,如碘乙酸、碘乙酰胺和2,4-二硝基氟苯等,使酶蛋白中的 –SH、-NH2、-OH等发生烷基化, 失活。 6.青霉素—抗菌素类药物,与糖肽转肽酶活性部位Ser-OH共价结合,使酶失活。 “自杀性底物”

115
b. 可逆抑制剂 在可逆抑制剂中最重要的是竞争性抑制剂。 如:磺胺药、氨基叶酸等。用增加底物浓度的方法可以减弱抑制作用。

116
第六节、别构酶 同工酶 诱导酶 抗体酶 核酶 一、别构酶 (一)别构酶的概念和特点 1.概念: 别构酶也称变构酶,它是代谢过程中的关键酶。通过效应物(调节物)和酶的别构中心的结合来调节其活性,从而调节酶反应速度和代谢过程。
和酶的别构中心的结合来调节其活性,从而调节酶反应速度和代谢过程。 .")
117
2.别构酶的基本特点: (1)由多个亚基组成的寡聚酶 (2)具有四级结构
(3)除了有可以结合底物的酶的活性中心外,还有可以结合调节物的别构中心(有时也称调节中心)。这两个中心可能位于同一亚基上,也可能分别位于不同亚基上。 (4)每个别构酶分子可以有一个以上的活性部位和调节部位,因此可以结合 一个以上的底物分子和调节物分子。 别构酶的活性中心负责酶对底物的结合与催化,别构中心则负责调节酶反应速度。
除了有可以结合底物的酶的活性中心外,还有可以结合调节物的别构中心(有时也称调节中心)。这两个中心可能位于同一亚基上,也可能分别位于不同亚基上。 (4)每个别构酶分子可以有一个以上的活性部位和调节部位,因此可以结合 一个以上的底物分子和调节物分子。 别构酶的活性中心负责酶对底物的结合与催化,别构中心则负责调节酶反应速度。")
118
3.几个相关概念 别构效应:调节物(或效应物)与酶分子上的别构中心结合后,诱导出或稳定住酶分子的某种构象,使酶活性中心对底物的结合和催化作用受到影响,从而调节酶反应速度及代谢过程,此效应即为酶的别构效应 同促效应(同种协同效应):它的调节物分子就是底物分子,这种酶分子上有两个以上的底物结合中心,其调节作用取决于酶是有多少个底物结合中心被占据。 异促效应:这种别构酶除了与底物分子作用外,还可与其他的调节物分子结合,它的调节物分子不是底物分子。 多数别构酶兼有同促效应和异促效应。
:它的调节物分子就是底物分子,这种酶分子上有两个以上的底物结合中心,其调节作用取决于酶是有多少个底物结合中心被占据。 异促效应:这种别构酶除了与底物分子作用外,还可与其他的调节物分子结合,它的调节物分子不是底物分子。 多数别构酶兼有同促效应和异促效应。")
119
(二)别构酶的动力学及别构酶对酶反应速度的调节
1.大多数别构酶具有正协同效应 正协同效应:酶分子结合一分子底物或效应物后,酶的构象发生变化,这种新的构象有利于后续分子与酶的结合,大大促进后续分子与酶的亲合性) 其初速度与底物浓度的关系呈S形的v-[S]曲线。 当底物浓度发生很小的变化时,别构酶就极大地控制着反应速度。在正协同效应中使得酶反应速度对底物浓度的变化极为敏感。
 其初速度与底物浓度的关系呈S形的v-[S]曲线。 当底物浓度发生很小的变化时,别构酶就极大地控制着反应速度。在正协同效应中使得酶反应速度对底物浓度的变化极为敏感。")
120
2.另一类别构酶具有负协同效应,其动力学曲线在表现上与双曲线相似,但意义不同
正 [s] 具有负协同效应的酶在底物浓度较低的范围内酶活力上升快,但再继续下去,底物浓度虽有较大的提高,但反应速度升高却较小。使得酶反应速度对底物浓度的变化不敏感

121
(三)别构酶调节酶活性的机理 1.协同模型或对称模型(MWC模型)(1965)
1.别构酶是由确定数目的亚基组成的寡聚酶,各亚基占有相等的地位,因此每个别构酶都有一个对称轴。 2.每一个亚基对一种配体(或调节物)只有一个结合位点。 3.每种亚基有两种构象状态,一种为有利于结合底物或调节物的松弛型构象(R型)另一种为不利于底物或调节物结合的紧张型构象(T型) 各亚基在同一时间内均处于相同的构象状态。
只有一个结合位点。 3.每种亚基有两种构象状态,一种为有利于结合底物或调节物的松弛型构象(R型)另一种为不利于底物或调节物结合的紧张型构象(T型) 各亚基在同一时间内均处于相同的构象状态。")
122
2.序变模型(KNF模型)(1966) 1.当配体不存在时,别构酶只有一种构象状态存在(T)而不是处于R T的平衡状态,只有当配体与之结合后才诱导T态向R态转变 2.构象是以序变方式进行的,而不是齐变。(有各种TR杂合型) 3.亚基间的相互作用可能是正协同效应,也可能是负协同效应,前者导致下一亚基对配体有更大的亲和力,后者则降低亲和力。 T态 TR态 R态

123
LDH5(M4)、 LDH4(M3H)、 LDH3(M2H2)、 LDH2(MH3)、 LDH1(H4)
二、同工酶 1.概念 : 同工酶:是指能催化相同的化学反应,但酶蛋白的分子结构、理化性质和免疫性能等方面都存在明显差异的一组酶。一般为寡聚蛋白。 2.同工酶举例: 1959年,用电泳分离法发现动物的乳酸脱氢酶具有多种分子形式。 LDH5(M4)、 LDH4(M3H)、 LDH3(M2H2)、 LDH2(MH3)、 LDH1(H4) 心肌型(H)和肌肉型(M)
、 LDH4(M3H)、 LDH3(M2H2)、 LDH2(MH3)、 LDH1(H4) 心肌型(H)和肌肉型(M)")
124
心、肝病变时引起的血清LDH同工酶的变化规律: 心脏疾病 LDH2和LDH4上升,LDH3和LDH5下降。
(H3M) LDH3 (H2M2) LDH4 (H1M3) LDH5 (M4) H亚基 M亚基 心、肝病变时引起的血清LDH同工酶的变化规律: 心脏疾病 LDH2和LDH4上升,LDH3和LDH5下降。 急性肝炎 LDH5明显上升,随病情好转而恢复正常。
 LDH3. (H2M2) LDH4. (H1M3) LDH5. (M4) H亚基. M亚基. 心、肝病变时引起的血清LDH同工酶的变化规律: 心脏疾病 LDH2和LDH4上升,LDH3和LDH5下降。 急性肝炎 LDH5明显上升,随病情好转而恢复正常。")
125
三、诱导酶 根据酶的形成与代谢的关系,人们把酶相对地分为结构酶和诱导酶 结构酶—是指细胞中天然存在的酶,其含量较为稳定,受外界的影响小。
诱导酶—指细胞中加入特定诱导物后诱导产生的酶,其含量在诱导物存在下显著增高,这种诱导物往往是该酶底物的类似物或底物本身.

126
四、抗体酶 抗体酶—指专一于抗原分子的, 具有催化活性的免疫球蛋白,即在其高可变区赋予了酶的属性。是抗体的高度选择性与酶的高效催化性相结合的产物。

127
(五)核酶 1.概念 核酶:具有催化活性的RNA. 2.种类 按作用底物分: (1)催化分子内反应(如自我剪接和自我剪切)的核酶。
(2)催化分子间反应(如原核生物RNaseP中的RNA)的核酶。
催化分子间反应(如原核生物RNaseP中的RNA)的核酶。")
128
第七节、酶活力测定与分离提纯 一、酶活力测定与酶活力单位 (一)酶活力与酶反应速度
1. 酶活力:也称酶活性,指酶催化一定化学反应的能力。其大小可用在一定条件下,它所催化的某一化学反应的反应速度来表示,两者呈线性关系。所以测定酶的活力就是测定酶的反应速率。 2. 酶反应速度:用单位时间内、单位体积中底物的减少量或产物的增加量来表示。单位:浓度/单位时间

129
引起酶反应速度降低的主要原因: 1、底物浓度的降低; 2、酶的部分失活; 3、产物对酶的抑制; 斜率=浓度/时间=
4、产物增加引起的逆反应速度的增加 斜率=浓度/时间= 产物浓度 可以用初速度来测定制剂中酶的含量。 时间 酶反应速度曲线

130
3、酶的活力单位 (1).酶的活力单位: 1961年,提出用“国际单位”(IU)表示酶活力, 即:1个酶活力单位:是指在最适条件下,1分钟内能转化1微摩尔底物的酶量,或转化底物中1微摩尔有关基团的酶量。(25C,最适底物浓度和最适pH) 1IU=mol/min 1972年,提出新的酶活力国际单位:Kat单位:最适条件下,每秒钟能催化1mol底物转化为产物所需的酶量,定为1Kat=1mol/s 所以:1Kat=60×106 IU 习惯用法:每小时催化1克底物所需的酶量。

131
比活力=活力IU/mg蛋白=总活力IU/总蛋白mg (3)转换数
(2).酶的比活力: 代表酶的纯度,比活力用每mg蛋白质所含的酶活力单位数表示,对同一酶来说,比活力愈大,表示酶的纯度愈高。用IU/mg蛋白、 Kat/mg蛋白表示。 比活力大小可用来比较每单位质量蛋白质的催化能力。 比活力=活力IU/mg蛋白=总活力IU/总蛋白mg (3)转换数 每秒钟每个酶分子能催化底物发生变化的微摩尔数,用kcat表示( mol/S )。
.酶的比活力: 代表酶的纯度,比活力用每mg蛋白质所含的酶活力单位数表示,对同一酶来说,比活力愈大,表示酶的纯度愈高。用IU/mg蛋白、 Kat/mg蛋白表示。 比活力大小可用来比较每单位质量蛋白质的催化能力。 比活力=活力IU/mg蛋白=总活力IU/总蛋白mg. (3)转换数. 每秒钟每个酶分子能催化底物发生变化的微摩尔数,用kcat表示( mol/S )。")
132
(二)酶活力的测定方法 1.分光光度法 利用底物和产物在紫外或可见光部分的光吸收的不同,选择一适当的波长,测定反应过程中反应进行的情况。
优点:简便、节省时间和样品,可检测到nmol/L水平的变化。 2.荧光法 主要是根据底物或产物的荧光性质的差别来进行测定。 3.同位素测定方法 4.电化学方法

133
二、酶分离提纯的一般原则 1、选材:目前常用微生物为材料制备各种酶制剂 2、破碎细胞:超声波、细菌磨、冻融等处碎壁制成组织匀浆。
3、抽提:在低温下,以水或低盐缓冲液,从组织匀浆中抽提酶,得到酶的粗提液 4、分离与提纯:一般在0-5℃间进行防止酶变性失活 5、结晶:酶的结晶过程进行得很慢,如果要得到好的晶体也许需要数天或数星期。 6、保存:通常将纯化后的酶溶液经透析除盐后冰冻干燥得到酶粉,低温下可较长时期保存。

134
酶的保存:1.低温(0-4℃,-20℃);2.高浓度较稳定;3.加入稳定剂;4.固定化。
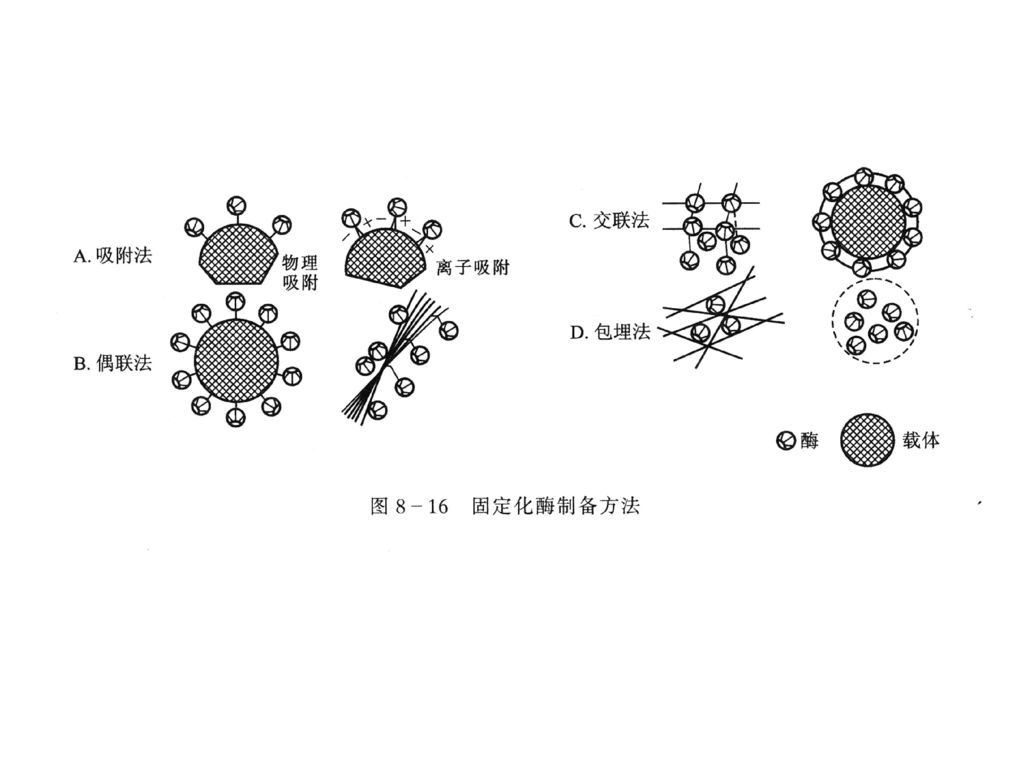
酶的固定化就是把水溶性酶经物理(吸附法与包埋法)或化学方法(共价偶联法与交联法)处理后,使酶与惰性载体结合或将酶包埋起来成为一种不溶于水的状态。 1、吸附法:使酶被吸附于惰性固体的表面,或吸附于离子交换剂上。 2、包埋法:使酶包埋在凝胶的格子中或聚合物半透膜小胶囊中。 3、偶联法:使酶通过共价键连接于适当的不溶于水的载体上。 4、交联法:使酶分子依靠双功能基团试剂交联聚合成“网状”结构。
或化学方法(共价偶联法与交联法)处理后,使酶与惰性载体结合或将酶包埋起来成为一种不溶于水的状态。 1、吸附法:使酶被吸附于惰性固体的表面,或吸附于离子交换剂上。 2、包埋法:使酶包埋在凝胶的格子中或聚合物半透膜小胶囊中。 3、偶联法:使酶通过共价键连接于适当的不溶于水的载体上。 4、交联法:使酶分子依靠双功能基团试剂交联聚合成 网状 结构。")
136
第八节、酶工程简介 一、酶工程的概念: 酶工程:是指酶制剂在工业上的大规模生产及应用。
1971年第一届国际酶工程会议上得到命名。主要研究:酶的生产、纯化、固定化技术、酶分子结构的修饰和改造及其在工、农、医药等领域的应用。 天然酶在开发和应用方面受到限制: 1.酶的不稳定性 2.酶的分离、纯化较难,成本高,价格贵

137
目前在酶的应用方面所采取的一般方法: 1. 化学方法:通过对酶的化学修饰或固定化处理,改善酶的性质以提高酶的效率和降低成本,或通过化学合成法制造人工酶。 2. 利用基因重组技术生产酶以及对酶基因进行修饰或设计新基因,生产出性能稳定、具有新的生物活性以及催化效率更高的酶。

138
二、化学酶工程 指天然酶、化学修饰酶、固定化酶及人工模拟酶的研究和应用。 1.天然酶
化学酶工程亦称初级酶工程: 指天然酶、化学修饰酶、固定化酶及人工模拟酶的研究和应用。 1.天然酶 主要指工业用酶,从微生物发酵得到的酶。如:洗涤剂、皮革生产中用的蛋白酶;纸张制造用的淀粉酶;乳制品用的凝乳酶等。

139
2.化学修饰酶:用于医药及研究工作,通过 (1)化学修饰酶的功能基 ; (2)通过交联反应; (3)大分子修饰作用 3.固定化酶:将水溶性酶用物理或化学方法,使之成为不溶于水的,但仍具有酶活性的状态。 4.人工模拟酶—化学法合成酶(已知酶的活性中心和作用机理)。
。")
140
三、生物酶工程 生物酶工程亦称高级酶工程 是酶学和以DNA重组技术为主的现代分子生物学技术相结合的产物。 生物酶工程的主要内容:
克隆酶—用基因工程技术大量生产酶( -淀粉酶,青霉素酰胺酶、亮氨酸合成酶) 突变酶—对酶基因进行修饰,产生遗传修饰酶 制造新酶—设计新酶基因,合成自然界不曾有过的酶
 突变酶—对酶基因进行修饰,产生遗传修饰酶. 制造新酶—设计新酶基因,合成自然界不曾有过的酶.")
141
本章总结 第一节 酶的概念 第二节 酶的命名与分类 一、酶是生物催化剂 二、酶的作用特点 三、酶的分布 四、酶的专一性 一、酶的分类
第一节 酶的概念 一、酶是生物催化剂 二、酶的作用特点 三、酶的分布 四、酶的专一性 第二节 酶的命名与分类 一、酶的分类 二、酶的命名

142
第三节 酶的组成与结构 一、酶的组成 二、几个重要的辅助因子 (一)辅酶、辅基 (二)金属离子 三、酶的结构
第三节 酶的组成与结构 一、酶的组成 二、几个重要的辅助因子 (一)辅酶、辅基 (二)金属离子 三、酶的结构 (一)酶蛋白与蛋白质相同,都有一定的空间结构 (二)根据酶蛋白结构上的特点,酶分为: (三)酶的活性中心
辅酶、辅基. (二)金属离子. 三、酶的结构. (一)酶蛋白与蛋白质相同,都有一定的空间结构. (二)根据酶蛋白结构上的特点,酶分为: (三)酶的活性中心.")
143
第四节、酶的催化作用机理 第五节、影响酶促反应速度的因素 一. 酶作用专一性机理 二. 酶作用高效性机理 三. 影响酶高效性的因素
四. 酶原与酶原的激活 第五节、影响酶促反应速度的因素 一、底物浓度对反应速度的影响 二、pH对反应速度的影响 三、温度对反应速度的影响 四、酶浓度对反应速度的影响 五、激活剂对反应速度的影响 六、抑制剂对反应速度的影响

144
第六节、别构酶 同工酶 诱导酶 抗体酶 核酶 第七节、酶活力测定与分离提纯 第八节、酶工程简介 一、酶活力测定与酶活力单位
(一)酶活力与酶反应速度 (二)酶活力的测定方法 二、酶分离提纯的一般原则 第八节、酶工程简介
酶活力与酶反应速度. (二)酶活力的测定方法. 二、酶分离提纯的一般原则. 第八节、酶工程简介.")
Similar presentations

生物生长发育和代谢所必需 的一类微量有机物质;( 2 )需量少,每日仅需 mg 或 g 级;( 3 )在体内不能合成或合成不足, 必需由食物供给( 4 )机体缺乏会导致物质谢障 碍,引起缺乏症。 分类:维生素一般习惯分为脂溶性和水溶性两大 类。其中脂溶性维生素在体内可直接参与代谢的.>")
 一、维生素的一般概念和类别. 维生素是机体维持正常生命活动所必不可 少的一类有机物质。 在机体内含量很少,在生命活动中,维生 素既不是构成组织的基础物质,也不是能 量物质,但它是一类重要的生命物质,在 代谢中起调节作用,如果缺乏会导致一定.>")



;>")
:核蛋白体组成成分 转移 RNA ( tRNA ):转运氨基酸 信使 RNA ( mRNA ):蛋白质合成模板 不均一核 RNA ( hnRNA ):成熟 mRNA 的前体 小核 RNA ( snRNA ):>")







 2004.1 酶.>")


.>")
