Download presentation
Presentation is loading. Please wait.

1
脂質代謝

2
脂肪的分解和合成是在細胞的不同部位進行。
脂肪合成: 細胞質中,由NADPH提供原動力 脂肪分解: 粒腺體中,以FAD和NAD+為質子與電子的 受體。 脂肪比醣類釋出高達一倍多的能量,因為脂肪分子中的 和氧的比值比醣類高: 棕櫚酸 C:H:O = 8:16:1, 葡萄糖 C:H:O = 1:2:1

3
14.1 脂質的消化與吸收 一、脂質的消化 存於消化道中以備水解的脂肪幾乎全為三酸甘油酯,只有一小 部份的食物脂肪屬於膽固醇酯及磷脂類。 1.舌脂肪酶(Lingal lipase)胃中的脂肪酶活性大部分是來自舌背之漿液性舌腺所分泌之脂肪酶,這種舌脂肪酶(Lingal lipase)是針對中鏈及短鏈三酸甘油酯,舌脂肪酶對於幼小動物及新生兒(早產兒)之脂肪消化是特別重要的。
胃中的脂肪酶活性大部分是來自舌背之漿液性舌腺所分泌之脂肪酶,這種舌脂肪酶(Lingal lipase)是針對中鏈及短鏈三酸甘油酯,舌脂肪酶對於幼小動物及新生兒(早產兒)之脂肪消化是特別重要的。")
4
2.胃解脂酶: 食物中10%的脂肪是在胃部分解的,但是缺乏膽鹽作用,成為不帶電且難容於水,聚集在指解酶周圍

5
3.胰解脂酶:在小腸腔中有胰臟分泌的胰解脂酶 (pancreatic lipase),分為-解脂酶和-解脂 酶。
三醯甘油可被-解脂酶分解為二醯甘油以及單醯甘油. 單醯甘油再被-解脂酶水解去除-碳原子上的脂肪酸

6
4. 膽固醇酯酶:膽固醇酯酶(cholesterol. esterase),可將膽固醇酯(cholesterol ester)
4.膽固醇酯酶:膽固醇酯酶(cholesterol esterase),可將膽固醇酯(cholesterol ester) 水解成膽固醇(cholesterol)和脂肪酸。 5.磷酸解脂酶:磷酸解脂酶(phospholipase)可將磷脂質(phospholipid)水解產生甘油、脂肪酸、磷酸以及膽鹼、乙醇胺或serine.
,可將膽固醇酯(cholesterol ester) 水解成膽固醇(cholesterol)和脂肪酸。 5.磷酸解脂酶:磷酸解脂酶(phospholipase)可將磷脂質(phospholipid)水解產生甘油、脂肪酸、磷酸以及膽鹼、乙醇胺或serine.")
7
二、脂質的吸收 在腸黏膜細胞中,游離脂肪酸被轉化成醯基CoA (acyl coenzyme A),首先合成二醯甘油,再合成三醯甘油,並形成直徑為0.5~1.0m的乳糜微粒(chylomicron),被釋放到黏膜細胞外。
,首先合成二醯甘油,再合成三醯甘油,並形成直徑為0.5~1.0m的乳糜微粒(chylomicron),被釋放到黏膜細胞外。")
8
三、脂質的運輸 脂肪、磷脂質和膽固醇及其酯衍生物分別以乳糜微粒(CM)、極低密度脂蛋白(VLDL) 、LDL、HDL、VHDL等形式,由血液運送(transportation)。 脂肪酸的運輸:游離脂肪酸除以極高密度脂蛋白(VHDL)運送外,也有一部分由血清蛋白運送。 在血液中各種脂質成分即構成血脂質(blood lipid)。 在血管中有C6~C10低分子量的游離脂肪酸與血漿蛋白結合,由微血管經肝門靜脈進入肝臟氧化 C10以上的長鏈脂肪酸與血漿蛋白結合運送。 C6~C10短鏈游離脂肪酸比C12~C18長鏈游離脂肪酸容易吸收,不飽和脂肪酸比飽和脂肪酸易吸收
。 在血管中有C6~C10低分子量的游離脂肪酸與血漿蛋白結合,由微血管經肝門靜脈進入肝臟氧化. C10以上的長鏈脂肪酸與血漿蛋白結合運送。 C6~C10短鏈游離脂肪酸比C12~C18長鏈游離脂肪酸容易吸收,不飽和脂肪酸比飽和脂肪酸易吸收.")
9
2.膽固醇的運輸:膽固醇的吸收和運送必須依賴 脂蛋白。被吸收的膽固醇可透過膽汁(bile)再 排入腸腔,稱為再循環(re-cycle)。
被腸黏膜細胞吸收的膽固醇會與脂肪酸形成膽固醇酯,可通過淋巴系統進入血液循環。

11
14.2 脂肪的分解代謝: 在解脂酶(lipase)水解下生成甘油和脂肪酸
一、甘油代謝 甘油在ATP參與下,由甘油激酶(glycerol kinase)催化,首先轉變為-磷酸甘油(α-phosphoglycerol)。 -磷酸甘油在磷酸甘油脫氫酶(phosphoglycerol dehydrogenase)作用下,轉變成磷酸二羥丙酮。磷酸二羥丙酮可進入三羧酸循環被徹底氧化;也可經由EMP逆向反應用以合成葡萄糖或肝醣
催化,首先轉變為-磷酸甘油(α-phosphoglycerol)。 -磷酸甘油在磷酸甘油脫氫酶(phosphoglycerol dehydrogenase)作用下,轉變成磷酸二羥丙酮。磷酸二羥丙酮可進入三羧酸循環被徹底氧化;也可經由EMP逆向反應用以合成葡萄糖或肝醣.")
12
14.2 脂肪的分解代謝: 在解脂酶(lipase)水解下生成甘油和脂肪酸
一、甘油代謝 甘油在ATP參與下,由甘油激酶(glycerol kinase)催化,首先轉變為-磷酸甘油(α-phosphoglycerol)。 -磷酸甘油在磷酸甘油脫氫酶(phosphoglycerol dehydrogenase)作用下,轉變成磷酸二羥丙酮。磷酸二羥丙酮可進入三羧酸循環被徹底氧化;也可經由EMP逆向反應用以合成葡萄糖或肝醣
催化,首先轉變為-磷酸甘油(α-phosphoglycerol)。 -磷酸甘油在磷酸甘油脫氫酶(phosphoglycerol dehydrogenase)作用下,轉變成磷酸二羥丙酮。磷酸二羥丙酮可進入三羧酸循環被徹底氧化;也可經由EMP逆向反應用以合成葡萄糖或肝醣.")
13
圖14-1 甘油代謝途徑

14
從a和b碳原子切斷,b-碳原子被氧化成為COO-,
二、脂肪酸的-氧化 1904年德國生物化學家Franz Knoop做動物實驗時,發現: 偶數碳原子的苯脂酸均變為苯乙尿酸(phenyl-ethyl-glycine)(苯乙酸與甘胺酸的縮合產物) 奇數碳原子的苯脂酸均變為馬尿酸(hippuric acid)(苯甲酸與甘胺酸的縮合產物)。 脂肪酸的分解每次是切下一個二碳單位, 從a和b碳原子切斷,b-碳原子被氧化成為COO-, 所以提出脂肪酸b-氧化作用
(苯乙酸與甘胺酸的縮合產物) 奇數碳原子的苯脂酸均變為馬尿酸(hippuric acid)(苯甲酸與甘胺酸的縮合產物)。 脂肪酸的分解每次是切下一個二碳單位, 從a和b碳原子切斷,b-碳原子被氧化成為COO-, 所以提出脂肪酸b-氧化作用.")
15
脂肪酸的β-氧化作用(β-oxidation):
脂肪酸的-氧化作用是脂肪酸氧化分解的主要途徑,最後產生乙醯CoA (acetyl CoA),進入TCA循環,被徹底氧化。 步驟: 1.脂肪酸的活化 2.脂醯CoA轉入粒腺體 3.脫氫作用 4.水合作用 5.再脫氫作用 6.硫解作用
,進入TCA循環,被徹底氧化。 步驟: 1.脂肪酸的活化. 2.脂醯CoA轉入粒腺體. 3.脫氫作用. 4.水合作用. 5.再脫氫作用. 6.硫解作用.")
16
O 1.脂肪酸的活化 ∥ ATP水解為AMP提供能量 脂肪酸與CoA生成 高能硫酯鍵”脂醯CoA (fatty acyl CoA)”
催化酶:脂醯CoA合成酶 (fatty acyl CoA synthetase) 為在粒線體外膜和內質網。 O ∥ 脂醯CoA合成酶 Fatty acyl CoA
 為在粒線體外膜和內質網。 O. ∥ 脂醯CoA合成酶. Fatty acyl CoA.")
17
1.脂肪酸的活化 特例: 在微生物及動物肌肉細胞中,脂肪酸的活化主要由CoA轉移酶(CoA transferase)催化, 琥珀酸CoA提供CoASH,生成脂醯CoA。
催化, 琥珀酸CoA提供CoASH,生成脂醯CoA。")
18
2.脂醯CoA轉入粒線體 脂醯CoA無法通過粒線體內膜,需藉由 肉酸素(canitine)運輸。 反應: 肉酸素是由Lys轉化而成的,脂肪酸的連接位置是第3位置的羥基與脂肪酸羧基縮合成酯鍵。
運輸。 反應: 肉酸素是由Lys轉化而成的,脂肪酸的連接位置是第3位置的羥基與脂肪酸羧基縮合成酯鍵。")
19
肉酸素醯基轉移酶(carnitine acyl transferase)
位在粒線體的內膜上,可分為酶I和II 酶I:在粒線體內膜的外側,催化脂醯CoA變成 脂醯肉酸素, 並使脂醯肉酸素轉入膜內 酶II:在粒線體內膜的內側,催化上述逆反應, 使脂醯CoA進入粒腺體的基質(matrix) 進行氧化分解
 進行氧化分解.")
20
圖14-2 脂醯CoA轉入粒線體示意圖

21
3.脫氫作用

22
4.水合作用 H

23
-羥脂醯CoA脫氫酶(-hydroxyacyl-CoA dehydrogenase)具有立體異構專一性,只能催化L-羥脂醯CoA氧化。
5.再脫氫作用: -羥脂醯CoA脫氫酶(-hydroxyacyl-CoA dehydrogenase)具有立體異構專一性,只能催化L-羥脂醯CoA氧化。 _
具有立體異構專一性,只能催化L-羥脂醯CoA氧化。 _.")
24
6.硫解作用 再進行脫氫, 水合作用, 再脫氫和硫解作用 即脫下一個乙醯CoA--- TCA cycle和生糖

25
圖14-3 脂肪酸-氧化途徑

26
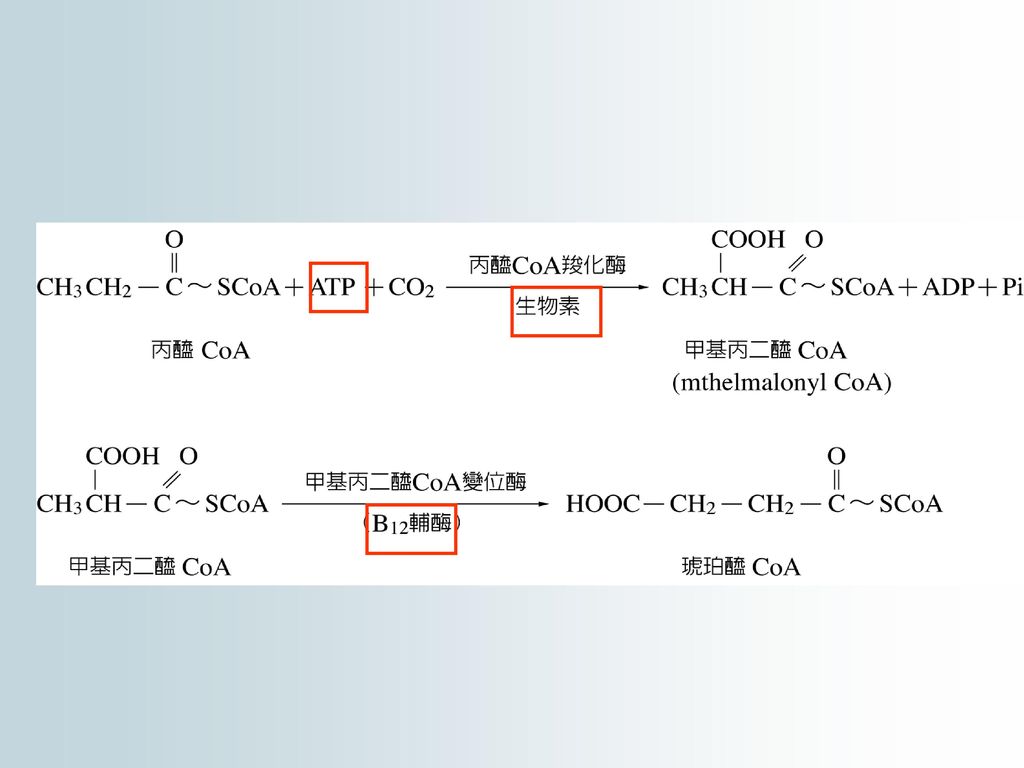
自然界存在的大多數脂肪酸都是含偶數碳原子,經過-氧化全部轉變成乙醯CoA,而後進入TCA循環被完全氧化。
動物和人體內丙醯基CoA可轉化為琥珀醯基CoA (succinyl CoA),而後併入TCA循環 。 在植物體內的丙醯基CoA可轉化為丙二醯基CoA (malonyl CoA),再脫去COO-變成乙醯CoA而後併入TCA循環 。
,而後併入TCA循環 。 在植物體內的丙醯基CoA可轉化為丙二醯基CoA (malonyl CoA),再脫去COO-變成乙醯CoA而後併入TCA循環 。")
28
不飽和脂肪酸的氧化分解,除按正常的-氧化過程外,還需兩個酶的參與:順-反-烯脂醯CoA異構酶(cis-trans-enoyl-CoA isomerase) 和-羥醯基CoA異構酶(-hydroxyacyl-CoA epimerase) 。
 和-羥醯基CoA異構酶(-hydroxyacyl-CoA epimerase) 。")
29
-氧化作用(-oxidation)是P. K. Stumpf(1965)首先在植物種子和葉組織中發現,而後也在腦及肝細胞中被發現。
三、-氧化作用和-氧化作用 -氧化作用(-oxidation)是P. K. Stumpf(1965)首先在植物種子和葉組織中發現,而後也在腦及肝細胞中被發現。 1. 加單氧 2. 脫氫 3. 脫去COO- 奇數碳原子脂肪酸
是P. K. Stumpf(1965)首先在植物種子和葉組織中發現,而後也在腦及肝細胞中被發現。 1. 加單氧. 2. 脫氫. 3. 脫去COO- 奇數碳原子脂肪酸.")
30
-氧化作用(-oxidation)存在於動物的肝及某些消化石油的微生物中。
1.加單氧 2.脫氫: 氧化成醛與酸 3.轉入粒線體進行b-氧化成琥珀醯CoA TCA

31
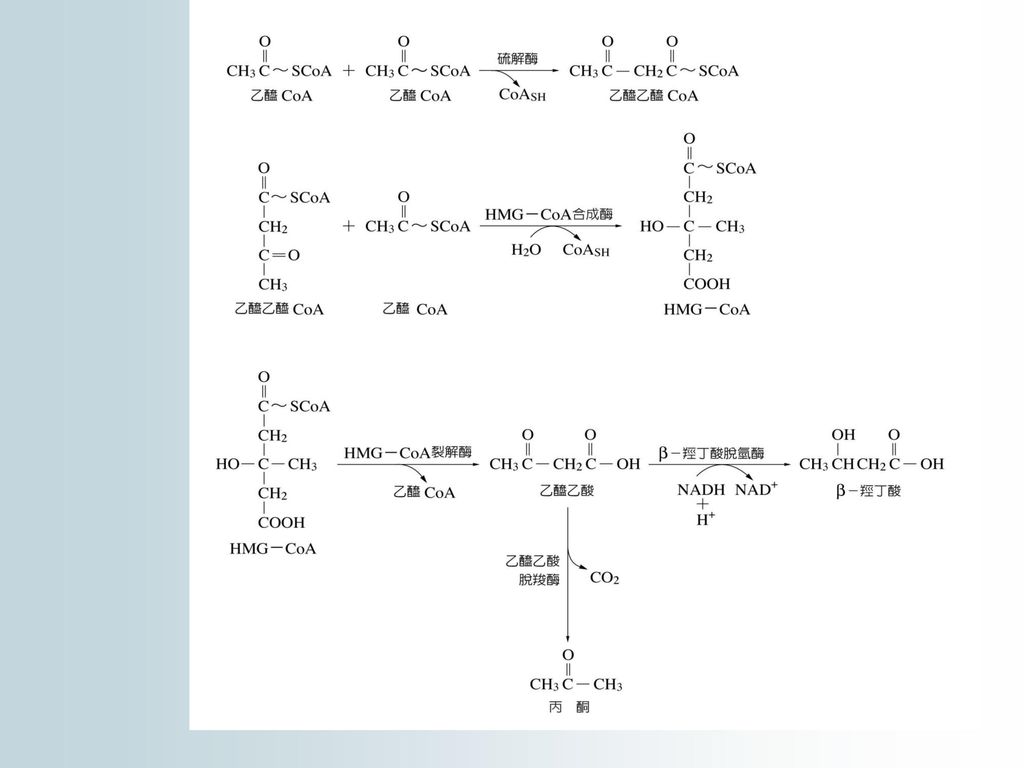
14.3 酮體代謝 脂肪酸進行b-氧化產生過量的乙醯CoA,若不能及時氧化分解,可轉變成丙酮(acetone)、乙醯乙酸(acetoacetic acid)、-羥基丁酸等,統稱為酮體(ketone body)。 一、酮體的生成 生成酮體的原料是乙醯CoA。

34
二、酮體的氧化 肝中缺乏解酮體酵素, 所以無法在肝中氧化, 酮體可容於水, 所以可由肝細胞滲入血液中循環運送到肝以外的組織進行氧化。 1.-羥基丁酸及乙醯乙酸的路徑

36
TCA cycle

37
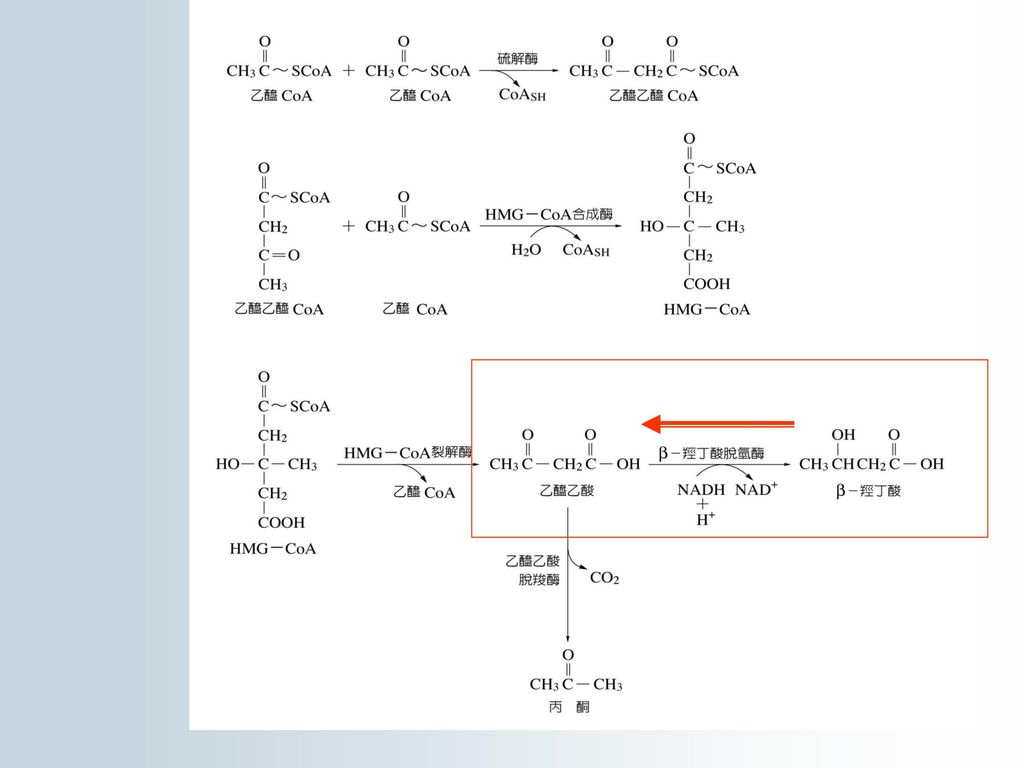
2.丙酮的去路:丙酮可以加水生成1,2-丙二醇 (malonol),繼而氧化成丙酮酸,也可氧化 成甲酸和乙酸。
丙酮酸和乙酸(活化成乙醯CoA)進入TCA cycle 酮體代謝是人和動物的一種 正常脂代謝途徑,它是脂代謝中聯繫肝臟及肝外組織代謝的一種方式。
進入TCA cycle. 酮體代謝是人和動物的一種 正常脂代謝途徑,它是脂代謝中聯繫肝臟及肝外組織代謝的一種方式。")
38
14.4 脂肪分解的能量轉換 脂肪用來儲存能源,在分解時能為有機體提供大量能量,其中脂肪酸完全氧化所提供的能量比醣類要大得多。 以硬脂酸(stearic acid)為例, 共有18個碳原子, 可經由8次-氧化產生9個乙醯CoA進入TCA cycle, 每次一個乙醯CoA進入TCA cycle即會產生12ATP。 9X12=108 ATP

39
NADH ATP FADH2

40
14.4 脂肪分解的能量轉換 -氧化作用一次產生一個FADH2(2ATP)和NADH(3ATP),共生成5個ATP。八次生成40ATP。 因此,每一分子硬脂酸完全分解,共生成148個ATP。 但是在-氧化時會 消耗2ATP (ATP AMP) 共生成148個ATP 所以共生成146個ATP
 共生成148個ATP. 所以共生成146個ATP.")
41
對於任何飽和或不飽和脂肪酸完全分解產生ATP數可用下式計算:
N: 脂肪酸碳原子數 D:不飽和脂肪酸的雙鍵數 -2: 活化時需要消耗2個ATP

42
N: 脂肪酸碳原子數 D:不飽和脂肪酸的雙鍵數 -2: 活化時需要消耗2個ATP 式中:n為脂肪酸碳原子數;d為不飽和脂肪酸的雙鍵數。5×( -1)為-氧化作用產生ATP數;12×( )為乙醯CoA產生的ATP數;因為有一個雙鍵就少生成一個FADH2,故少2個ATP;最後減2表示活化時需消耗的ATP數。
為-氧化作用產生ATP數;12×( )為乙醯CoA產生的ATP數;因為有一個雙鍵就少生成一個FADH2,故少2個ATP;最後減2表示活化時需消耗的ATP數。")
43
14.5 脂肪的合成作用 一、脂肪酸的合成作用 脂肪酸(fatty acid)的主要合成途徑(anabolism)是在粒線體外的細胞質中。 1963年,P. R. Vegelos等人以大腸桿菌(E. coli)為材料,分離出單獨起作用的七種與脂肪酸合成有關的酶,其中心是由77個胺基酸組成的脂醯基載體蛋白(acyl carrier protein, ACP)。 於是,基本上確立了脂肪酸的合成途徑。
為材料,分離出單獨起作用的七種與脂肪酸合成有關的酶,其中心是由77個胺基酸組成的脂醯基載體蛋白(acyl carrier protein, ACP)。 於是,基本上確立了脂肪酸的合成途徑。")
44
脂醯基運送蛋白與CoASH的結構是部分相同的,都具有磷酸泛酸硫基乙胺(phosphantheine),其功能基也是-SH基。所以脂醯基運送蛋白也可簡寫為ACPSH。其部分結構如下:
,其功能基也是-SH基。所以脂醯基運送蛋白也可簡寫為ACPSH。其部分結構如下:")
45
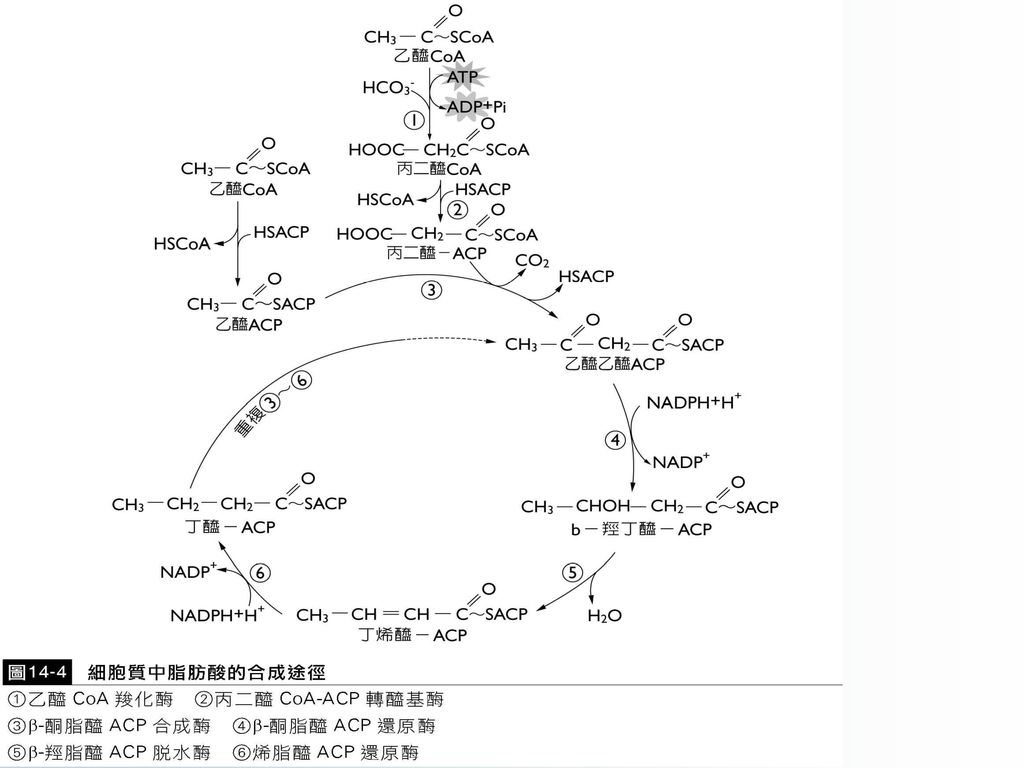
細胞質中脂肪酸的合成途徑包括下列六個步驟:
1.乙醯CoA羧化:作為脂肪酸合成的直接原料是 丙二醯CoA,丙二醯CoA由乙醯CoA羧化產 生,由乙醯CoA羧化酶(acetyl-CoA carboxylase)催化,生物素(biotin)為其輔因 子,ATP水解提供能量。
催化,生物素(biotin)為其輔因 子,ATP水解提供能量。")
47
乙醯CoA羧化酶是脂肪酸合成的調節,控制著整個脂肪酸合成的速度。
2.轉醯基反應

48
3.縮合反應

49
直接由兩分子乙醯ACP縮合,反應平衡不利於乙醯乙醯ACP的生成;而以丙二醯ACP作為反應物易使脫羧反應的自由能降低許多,而有利於此反應的平衡。

50
4.還原作用

51
5.脫水作用

52
6.再還原作用

53
經由上述途徑生成的脂醯基ACP經轉醯基酶(acyl-transferase)作用,將脂肪酸與CoASH結合,生成脂醯CoA,用於脂肪的合成;或者在硫酯酶(thioesterase)催化下,脫掉ACP而成游離脂肪酸。
作用,將脂肪酸與CoASH結合,生成脂醯CoA,用於脂肪的合成;或者在硫酯酶(thioesterase)催化下,脫掉ACP而成游離脂肪酸。")
54
細胞質中脂肪酸合成途徑總結於(圖14-4)。
。")
56
參與脂肪酸合成的酵素實際上是一個多酶系統(multienzyme system),ACP為其核心,各個酵素在ACP周圍依次排列(圖14-5)。
,ACP為其核心,各個酵素在ACP周圍依次排列(圖14-5)。")
57
圖14-5 脂肪酸合成多酵素系統(李,1990)
")
58
二、脂肪酸合成中乙醯CoA的來源 脂肪酸合成所需的乙醯CoA (acetyl-CoA),主要來自粒線體內基質中脂肪酸-氧化作用和丙酮酸的氧化作用的產物。乙醯CoA必須由肉酸素運輸、-酮戊二酸或檸檬酸自粒線體運輸到細胞質。
,主要來自粒線體內基質中脂肪酸-氧化作用和丙酮酸的氧化作用的產物。乙醯CoA必須由肉酸素運輸、-酮戊二酸或檸檬酸自粒線體運輸到細胞質。")
59
圖14-6 丙酮酸—檸檬酸循環

60
三、脂肪酸的碳鏈延長 飽和脂肪酸碳鏈的延長基本上是-氧化的逆轉反應,將乙醯CoA連續加到其羧基末端。 內質網(微粒體)也延長飽和或不飽和脂醯CoA的碳鏈。但所使用的原料是丙二醯基CoA而不是乙醯CoA,還原過程需NADPH供氫。
也延長飽和或不飽和脂醯CoA的碳鏈。但所使用的原料是丙二醯基CoA而不是乙醯CoA,還原過程需NADPH供氫。")
61
四、不飽和脂肪酸的合成 棕櫚酸和硬脂酸去飽和後即形成相對應的棕櫚油酸(palmitoleic acid,9-十六烯酸)和油酸(oleic acid,9-十八烯酸),這兩種烯脂酸在碳9位有一順式雙鍵。 1.細菌的合成方法:許多細菌的單烯脂肪酸的合 成則不需要氧分子,而是由特定的、中等碳鏈 長度的-脂醯基ACP脫水、延伸而成。

62
2.高等動植物合成方法: 哺乳動物(mammal)的多烯脂肪酸根據其雙鍵的數目常分為四大類,即棕櫚油酸(palmitoleic acid)、油酸(oleic acid)、亞麻油酸(linoleic acid)和亞麻仁油酸(linolenic acid)。其他多烯脂肪酸都由這四種脂肪酸作為前驅物再經通過碳鏈的延長和再次去飽和而衍生成。
的多烯脂肪酸根據其雙鍵的數目常分為四大類,即棕櫚油酸(palmitoleic acid)、油酸(oleic acid)、亞麻油酸(linoleic acid)和亞麻仁油酸(linolenic acid)。其他多烯脂肪酸都由這四種脂肪酸作為前驅物再經通過碳鏈的延長和再次去飽和而衍生成。")
63
哺乳動物只有從植物獲得亞麻油酸和亞麻仁油酸,因此,這兩種脂肪酸稱為必需脂肪酸(essential fatty acid)。
五、三醯甘油酯的合成 合成三醯甘油酯(油脂)除需有脂醯CoA外,還需磷酸化的甘油,即L-α-磷酸甘油(L-α-phosphoglycerol)
除需有脂醯CoA外,還需磷酸化的甘油,即L-α-磷酸甘油(L-α-phosphoglycerol)")
65
14.6 磷脂的分解代謝 一、磷脂的構造 磷脂是構成生物細胞膜的主要成分 ,在小腸中及微生物、植物中均存在多種磷脂酶(phospholipase),不同磷脂酶作用於磷脂的不同酯鍵上。 1.磷脂酶A1 :A1酶作用於C1位。 2.磷脂酶A2:作用在C2位,此處常連接不飽和脂 肪酸。 A1和A2作用於磷脂切下一個脂肪酸後 的產物具有溶血作用,故稱為溶血磷脂 (lysophospholipid)。
。")
66
3.磷脂酶C:分解鍵C3之鏈結。 4.磷脂酶D:作用於鍵C4, D酶主要存在於高等 植物中。 二、磷脂的分解代謝

68
1. 在動物組織中的分解途徑:卵磷脂通過磷脂酶. A1或A2的作用,切下一個脂肪酸,再被A1或. A2酶作用,生成甘油磷酸膽鹼,然後再被二酯
1.在動物組織中的分解途徑:卵磷脂通過磷脂酶 A1或A2的作用,切下一個脂肪酸,再被A1或 A2酶作用,生成甘油磷酸膽鹼,然後再被二酯 酶和磷酸單酯酶(phosphomonoesterase)作 用,水解生成膽鹼、甘油和脂肪酸。 2. 在細菌中的分解途徑:卵磷脂被磷脂酶C作 用, 最終生成膽鹼和磷酸。
作 用,水解生成膽鹼、甘油和脂肪酸。 2. 在細菌中的分解途徑:卵磷脂被磷脂酶C作 用, 最終生成膽鹼和磷酸。")
69
14.7 磷脂的合成代謝 在人體及動物體中,差不多在所有組織中都能合成磷脂質(phospholipids),但主要合成部位是肝臟(liver)。合成磷脂的材料脂肪酸、甘油和磷酸膽鹼再由不同的代謝途徑生成。
,但主要合成部位是肝臟(liver)。合成磷脂的材料脂肪酸、甘油和磷酸膽鹼再由不同的代謝途徑生成。")
70
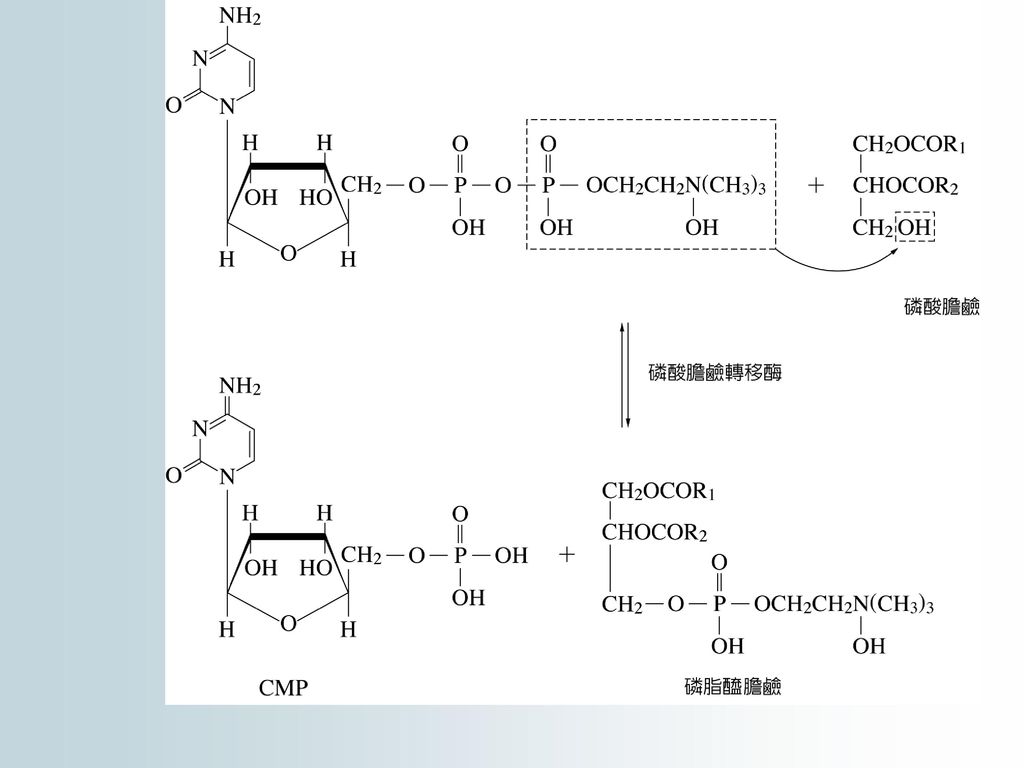
二醯基甘油酯和磷酸膽鹼可作為合成卵磷脂的直接原料;合成分兩步驟進行,並由兩個轉移催化,亦需要CTP參與:
1.生成膽鹼二磷酸胞苷

72
2.生成卵磷脂(CMP)
")
74
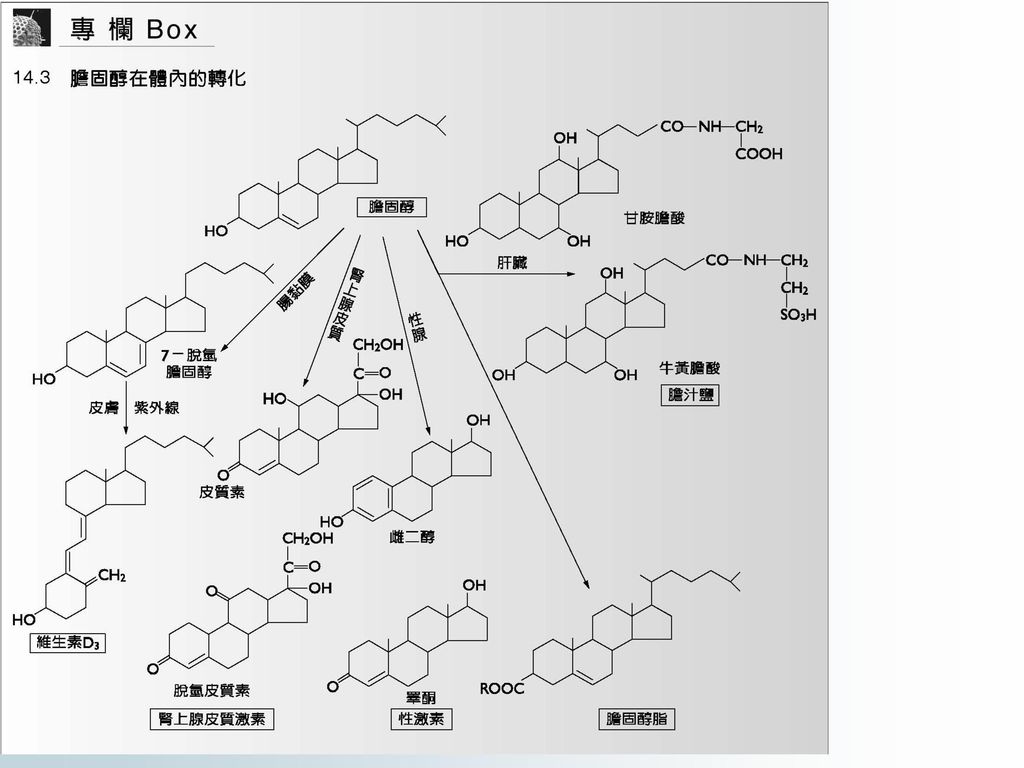
14.8 膽固醇的生物合成 膽固醇(cholesterol)的生物合成主要在肝中進行,其次是小腸(small intestine)、腎上腺(adrenal)和腦(brain)。 生物體內不少的次生物質如胡蘿蔔素、橡膠、膽固醇等都是以異戊二烯(isoprenoid)為碳經過加成作用而形成的。整個合成過程包括三個階段。第一階段是由二碳的乙醯CoA縮合成五碳的甲羥戊酸(mevalonic acid, MVA);第二階段由MVA轉變為開鏈的三十碳鯊烯(squalene)。這兩個階段的反應都在細胞質中進行;第三階段在內質網中,形成膽固醇。膽固醇完整的合成途徑見 (圖14-8)。
為碳經過加成作用而形成的。整個合成過程包括三個階段。第一階段是由二碳的乙醯CoA縮合成五碳的甲羥戊酸(mevalonic acid, MVA);第二階段由MVA轉變為開鏈的三十碳鯊烯(squalene)。這兩個階段的反應都在細胞質中進行;第三階段在內質網中,形成膽固醇。膽固醇完整的合成途徑見. (圖14-8)。")
75
圖14-8 膽固醇的生物合成途徑

Similar presentations







─ 廢食用油脂資源化 循環圖 什麼叫做生化柴油? 生化柴油係由可再生的油脂原料,諸如植物或 動物油脂,經合成 ( 交酯化 ) 所得的長鏈脂肪酸 甲脂,可代替柴油的一種 環保燃油 。經實驗 證明生化柴油可直接作為現有柴油引擎而不需.>")
 ( 一 ) 皆源自於動植物內的脂肪組織,是一種羧酸 酯,由碳 (C) 、氫 (H) 、氧 (O) 三種元素所組 成。 ( 二 ) 脂肪( Fat ) : 在室溫下呈固態者,如 豬油、 牛脂、羊油、奶油、乳酪等。 ( 三 ) 油質 (Oil): 在室溫下呈液態者,如黃豆油、 玉米油.>")

 不饱和脂肪酸的氧化 酮体 磷脂的代谢 脂肪酸代谢的调控.>")
 信号传导:激素、胞内信使.>")





.>")

物质在生物体的分解及合成代谢。要求学生重点掌握脂肪酸在生物体内的氧化分解途径—β-氧化和从头合成途径,了解脂类物质的功能和其他的氧化分解途径。 思考 脂类代谢 返回.>")
是参与生物生长发育和代谢所必需的一类微量有机物质。这类物质由于体内不能合成或者合成量不足,所以必需由食物供给。已知绝大多数维生素作为酶的辅酶或辅基的组成成分,在物质代谢中起重要作用。机体缺乏维生素时,物质代谢发生障碍,引起维生素缺乏症。 脂溶性维生素和水溶性维生素.>")
>")




