Download presentation
Presentation is loading. Please wait.

1
第二节 酶的结构与功能 一、酶的分子组成与分类 二、酶的辅助因子 三、酶的结构与功能 酶的活性中心和必需基团 酶的活性中心与酶作用的专一性
第二节 酶的结构与功能 一、酶的分子组成与分类 二、酶的辅助因子 三、酶的结构与功能 酶的活性中心和必需基团 酶的活性中心与酶作用的专一性 酶的活性中心与酶原的激活

2
三、酶的结构与功能 (一)、酶的活性中心和必需基团
酶分子上具有一定空间构象的部位,该部位化学基团集中,直接参与将底物转变为产物的反应过程,即与催化作用直接相关的部位,称为酶的活性中心(active center)。
。")
3
对于复合酶,辅因子常常是活性中心的组成部分。
(1)酶活性中心的组成: 由一些氨基酸残基的侧链基团组成。 用化学修饰法对多种酶的活性中心进行研究发现,活性中心处频率最高的氨基酸残基侧链基团主要有: Glu和Asp的-COOH,Lys的ε-NH2,His的咪唑基, Ser的-OH,Cys的-SH,Tyr的侧链基团。 对于复合酶,辅因子常常是活性中心的组成部分。
酶活性中心的组成: 由一些氨基酸残基的侧链基团组成。 用化学修饰法对多种酶的活性中心进行研究发现,活性中心处频率最高的氨基酸残基侧链基团主要有: Glu和Asp的-COOH,Lys的ε-NH2,His的咪唑基, Ser的-OH,Cys的-SH,Tyr的侧链基团。 对于复合酶,辅因子常常是活性中心的组成部分。")
4
His57 His57 His57

5
(2)酶活性中心的特点 a. 活性中心在酶分子总体积中只占相当小的部分 (约1%2%),相当于23个氨基酸残基。
b. 酶分子表面的一个凹穴,有一定的大小和形状, 但不是刚性的,而具有一定的柔性。 c. 活性中心为非极性的微环境,有利于与底物结合。

6
酶催化活性有关的基团 必需残基 酶蛋白 结合残基 接触残基 催化基团 活性中心 辅助残基 活性中心外 结构残基 非必需残基
--- 与酶的其它活性有关 如识别、定位、免疫等

7
必需基团 (1) 接触残基:结合基团,催化基团。 (2)辅助残基:不与底物接触,辅助酶与底物结合, 协助接触残基构成酶活性中心。
(3) 结构残基:维持酶分子三维构象,与酶活性相关, 但不在酶活性中心范围内,属于酶活 性中心以外的必需残基。
 结构残基:维持酶分子三维构象,与酶活性相关, 但不在酶活性中心范围内,属于酶活. 性中心以外的必需残基。")
8
酶分子中必需基团氨基酸残基的作用 接触残基: R1、R2、R6、R8、 R9、R163 辅助残基: R3、R4、R5、R164、 R165
结构残基: R10、R162、R169

9
底 物 活性中心以外的必需基团 催化基团 结合基团 活性中心

10
(二)、酶的活性中心与酶作用的专一性 1、酶的活性中心的一级结构研究
用同位素标记法:标记酶的活性中心,然后将酶水解并分离,对带标记片段进行一级结构测定。 分析结论:功能类似的酶在一级结构上有惊人的 相似性。

11
…Ser.Cys.Gly.Gly.Asp.Ser.Gly.Gly.Pro.Val…
活性丝氨酸 例:丝氨酸蛋白水解酶族的一级结构 酶 氨基酸顺序 牛胰蛋白酶 …Ser.Cys.Gly.Gly.Asp.Ser.Gly.Gly.Pro.Val… 牛胰凝乳蛋白酶 …Ser.Cys.Met.Gly.Asp.Ser.Gly.Gly.Pro.Leu… 猪弹性蛋白酶 …Gly.Cys.Gln.Gly.Asp.Ser.Gly.Gly.Pro.Leu… 猪凝血酶 ..Asp.Ala.Cys.Gln.Gly.Asp.Ser.Gly.Gly.Pro.… 且从微生物到哺乳动物,活性丝氨酸最邻近的5~6氨基酸顺序都一样 ——说明蛋白质活性中心在种系进化上有严格的保守性。

12
2、丝氨酸蛋白酶家族——空间构象具有一定相似性

13
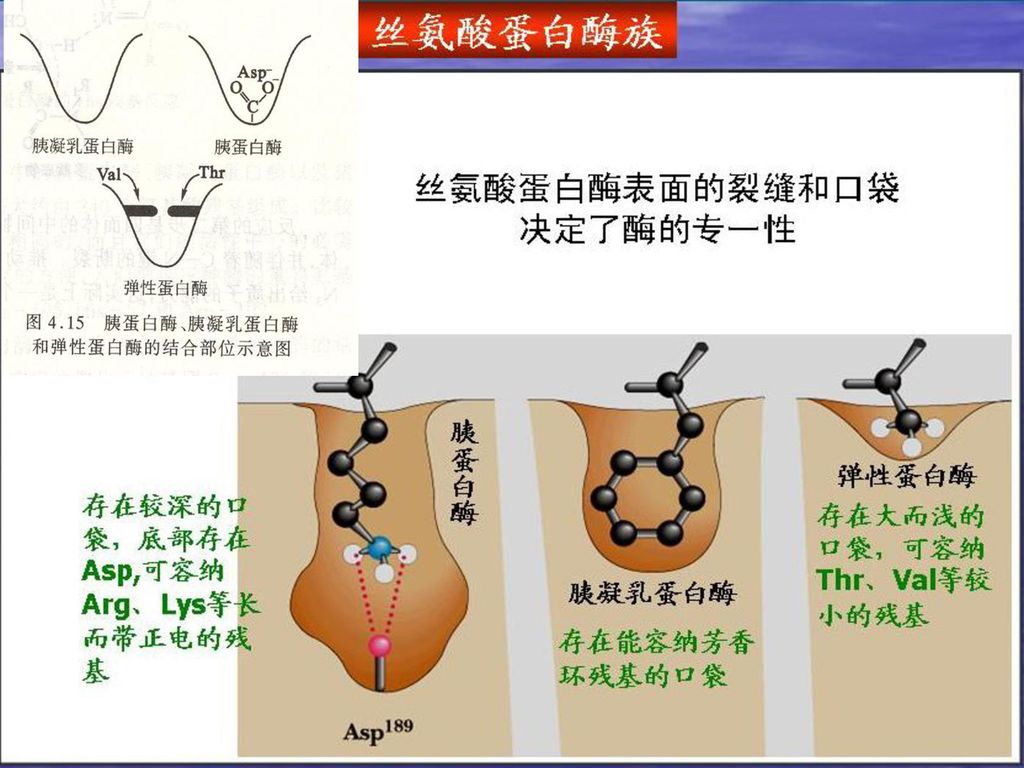
3、酶专一性与结合基团有关 胰凝乳蛋白酶:具有一个疏水性大口袋, 结合部位含有Ser残基 ,可与底物侧链为芳香基团氨基酸残基结合。
胰蛋白酶:与胰凝乳蛋白酶相似,具有口袋, 但结合部位由一个带负电荷的Asp代替了Ser,只能与带有正电荷氨基酸残基如 Lys 、 Arg 结合。 弹性蛋白酶: 结合部位入口处是Val和Thr(侧链长), 只能让侧链体积小Gly和Ala等进入。
, 只能让侧链体积小Gly和Ala等进入。")
15
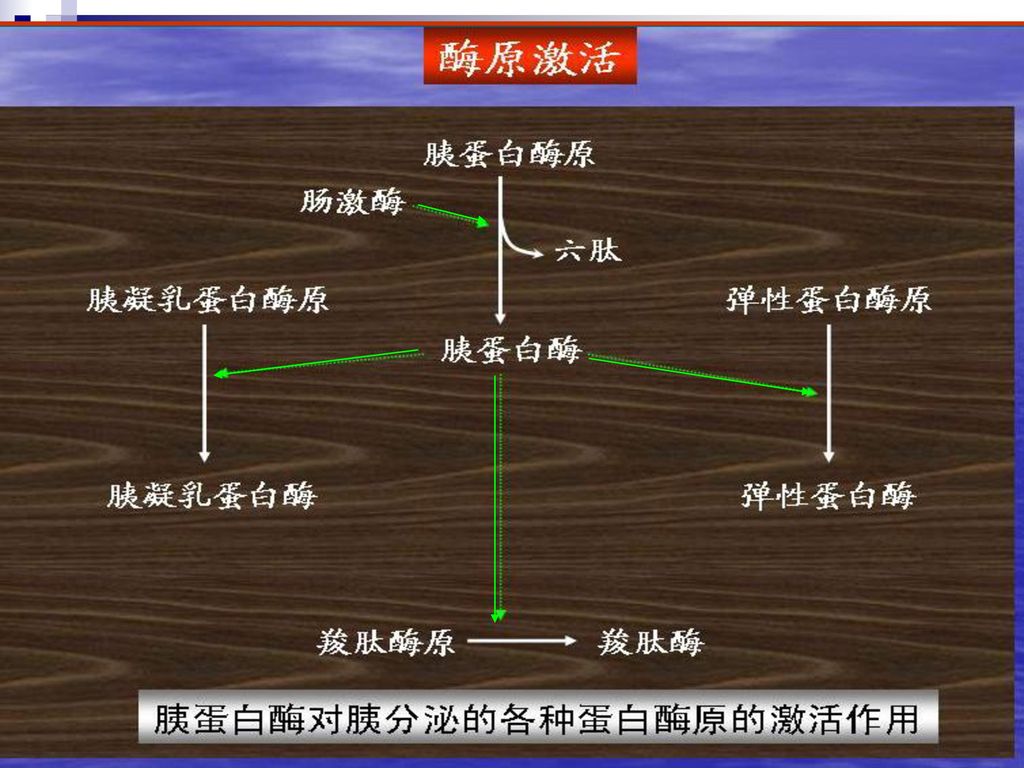
(三)酶的活性中心与酶原激活 胰蛋白酶原激活过程 胰蛋白酶活性中心 ?
酶的活性中心与酶原激活 胰蛋白酶原激活过程 胰蛋白酶活性中心")
16
酶 原 分子构象发生改变 形成或暴露出酶的活性中心 一个或几个特定的肽键断裂
酶原(zymogen) 无活性状态的酶的前体。 酶原的激活 酶原向活性的酶转化的过程。 酶原激活的机理 酶 原 分子构象发生改变 形成或暴露出酶的活性中心 一个或几个特定的肽键断裂 在特定条件下
 无活性状态的酶的前体。 酶原的激活 酶原向活性的酶转化的过程。 酶原激活的机理. 酶 原. 分子构象发生改变. 形成或暴露出酶的活性中心. 一个或几个特定的肽键断裂. 在特定条件下.")
18
胃蛋白酶原

19
胃蛋白酶原 胃蛋白酶

20
某些酶原的激活过程

22
酶原的激活 酶原激活的生理意义 ——酶活性的调节方式之一:酶结构的调节 避免对细胞进行自身消化 ——保护作用 酶在特定的部位和环境中发挥作用
——保证体内代谢正常进行 酶原是部分酶的储存形式

23
第三节 酶作用的机制 (一)酶-底物复合物的形成 酶底物复合物 E + S E + P ES
酶-底物复合物的形成 酶底物复合物 E + S E + P ES")
24
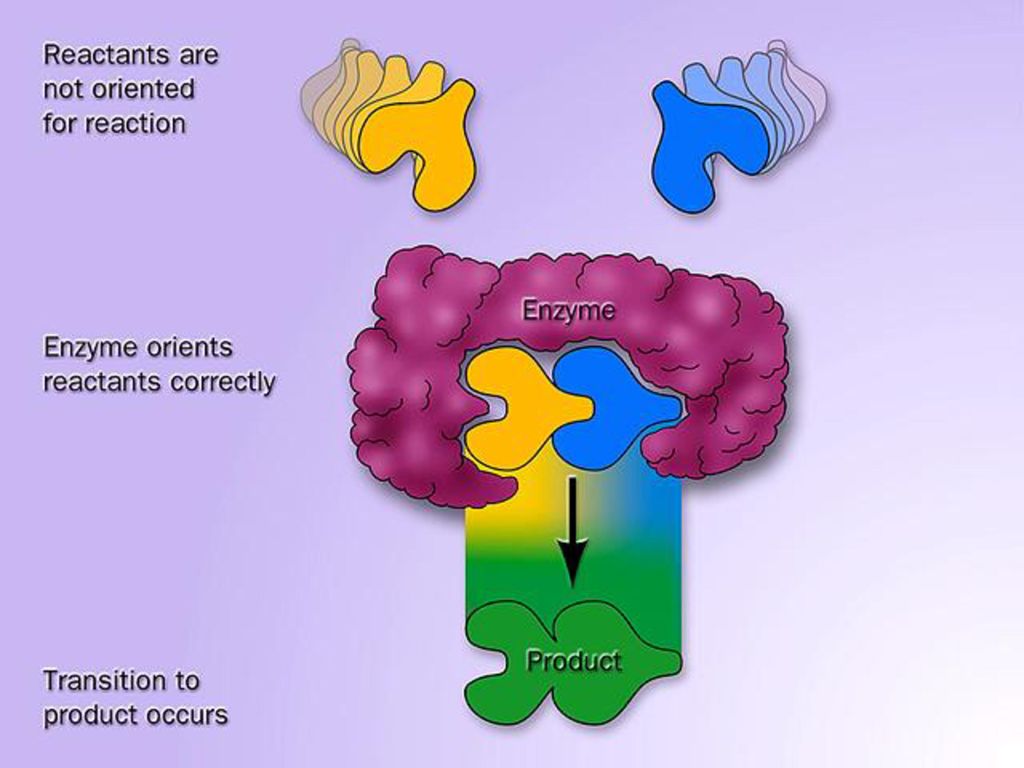
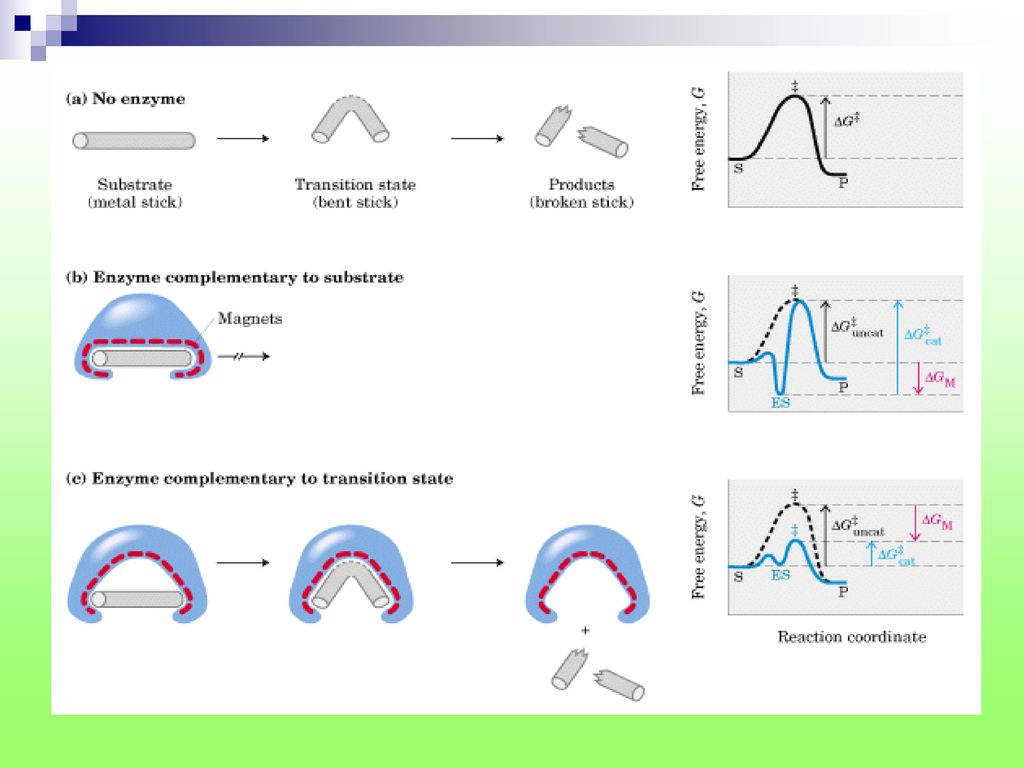
(二)、诱导契合学说(induced-fit hypothesis)
酶与底物接近→结构相互诱导→变形和相互适应→结合。

25
(三)、酶作用高效率的机制 趋近与定向效应 亲电催化 多元催化 亲核催化 酸、碱催化 离子催化 表面效应 “张力”和“形变” 酶
、酶作用高效率的机制 趋近与定向效应 亲电催化 多元催化 亲核催化 酸、碱催化 离子催化 表面效应 张力 和 形变 酶")
26
1、趋近与定向效应 邻近效应(proximity effect) 酶将诸底物结合到其活性中心,使它们的反应
基团相互靠近,降低进入过渡态所需的活化 能;底物在酶活性中心的有效浓度大大增加 定向排列(orientation arrange ): 酶活性中心的立体构型和相关基团的诱导、定向 作用,使诸底物参与反应的基团形成正确的几 何定向关系,相互接近,产生有效碰撞。
: 酶活性中心的立体构型和相关基团的诱导、定向. 作用,使诸底物参与反应的基团形成正确的几. 何定向关系,相互接近,产生有效碰撞。")
27
靠近

29
2、“张力”和“形变” 底物与酶结合,诱导酶的分子构象变化。
变化的酶分子又使底物分子的敏感键产生“张力”甚至“形变”,促使酶-底物中间产物进入过渡态。 酶与反应物过渡状态的亲和力远大于酶与底物或产物的亲和能力;

31
3、多元催化(multielement catalysis)
包括:酸-碱催化, 共价催化(亲核、亲电催化), 或多种催化形式的协同作用等。 同种基团在不同的微环境下解离度亦不同,这种多功能基团兼有酸、碱双重催化效能。
, 或多种催化形式的协同作用等。 同种基团在不同的微环境下解离度亦不同,这种多功能基团兼有酸、碱双重催化效能。")
32
酸碱催化: 酶分子中广义的酸碱基团: 氨基、羧基、巯基、酚羟基、咪唑基。 这类反应有:
酶活性部位上的某些基团成为质子(H+)供体或受体对底物酸碱催化。 酶分子中广义的酸碱基团: 氨基、羧基、巯基、酚羟基、咪唑基。 这类反应有: 羰基的加成作用,酮基和烯醇的互变异构, 肽和酯的水解及磷酸和焦磷酸参与的反应。
供体或受体对底物酸碱催化。 酶分子中广义的酸碱基团: 氨基、羧基、巯基、酚羟基、咪唑基。 这类反应有: 羰基的加成作用,酮基和烯醇的互变异构, 肽和酯的水解及磷酸和焦磷酸参与的反应。")
33
共价催化(又称亲核催化或亲电催化) 亲核催化剂或亲电催化剂在催化时放出电子或汲取电子并作用于底物的缺电子中心或负电中心,迅速形成不稳定共价复合物 酶中心最常见的亲核基团进攻底物的亲电中心, 底物中典型的亲电中心:磷酰基、酰基、糖基

34
酶分子中可作为亲核基团和酸硷催化功能基团

35
例:胰凝乳蛋白酶催化肽键水解的多种类催化机制
肽凝乳蛋白酶活性中心的结构特性 Ser195、His57和Asp102,通过氢键 “ 催化三元区 ” Asp102 —— 起稳定His57构象作用 His57 —— 质子受体,促进Ser195羟基氧对底物 肽酰基亲核进攻作用 “ 催化三元区 ” 特性:Ser-195 的羟基高度极化 , 具有很高的亲核进攻能力。 稳定作用

36
(2) 亲核共价催化作用: ① 四面体结构中间体的形成: Ser195羟基氧对底物肽酰基亲核进攻时, 形成一个共价四面体中间体
 亲核共价催化作用: ① 四面体结构中间体的形成: Ser195羟基氧对底物肽酰基亲核进攻时, 形成一个共价四面体中间体")
37
② His-57广义酸催化过程 酰基一酶中间体生成 :
受质子H同时 C-N肽键的断裂,形成产 物1R1NH2并从酶中离去; 环境中水 亲核进攻底物羰基碳,另一个过渡态四面 体产生; 产物1

38
③His-57作为广义碱-酸催化作用 His57作为碱进攻H2O并接受其质子,随后它作为酸又将质子转移给Ser195氧,——以此方式帮助酰基-酶中间体的解离。 酰基一酶中间体虽然 不稳定, 但 它的存在已经被X射线衍 射实验所证实。 产物2

39
4. 表面效应(surface effect):
酶活性中心的疏水环境排除了水分子对酶和底物功能基团的干扰、吸引或排斥,防止酶和底物之间形成水化膜,有利于酶和底物的密切接触。——疏水的为微环境极大利于酶的催化作用 + -

40
酶活性中心的低介电区(表面效应 surface effect)
化学基团的反应活性和反应速率在非极性介质 和水性介质有显著差别

41
第六节 重要的酶类及酶活性的调节 重要的酶类 : 寡聚酶 同工酶 诱导酶 调节酶 固定化酶

42
一、寡聚酶 由两个或两个以上,乃至多达数十个亚基组成的酶称为寡聚酶。其分子量从35000到几百万,可分为几种不同的类型:
1)含相同亚基的寡聚酶 2)含不同亚基的寡聚酶
含相同亚基的寡聚酶. 2)含不同亚基的寡聚酶.")
43
含不同亚基的寡聚酶 1. 双功能寡聚酶 2. 含有专一性的、非酶蛋白亚基的寡聚酶 3. 具有底物载体亚基的寡聚酶

44
1. 双功能寡聚酶 所含的亚基结构不同,每种亚基表现不同的功能,整个酶分子可以催化两个相关的反应。 举例:大肠杆菌的色氨酸合成酶

45
色氨酸合成酶 含A和B两种蛋白质 蛋白质A:含有一个亚基α, 蛋白质B:含有两个亚基ββ。 ——这两种蛋白质各自催化一个化学反应:
吲哚甘油磷酸 吲哚 + 3-磷酸甘油醛 吲哚 + 丝氨酸 色氨酸 蛋白质B

46
吲哚甘油磷酸 + 丝氨酸 色氨酸 + 3-磷酸甘油醛
色氨酸合成酶催化的化学反应 两个A蛋白 结合形成 一个B蛋白 它能将上述两个反应偶联而催化总反应: 色氨酸合成酶 (α2β2聚合) 吲哚甘油磷酸 + 丝氨酸 色氨酸 + 3-磷酸甘油醛
 吲哚甘油磷酸 + 丝氨酸 色氨酸 + 3-磷酸甘油醛.")
47
2.含有专一性、非酶蛋白亚基的寡聚酶 例如:乳糖合成酶催化乳糖的合成反应 这种寡聚酶含有两种亚基: 一种有催化作用, 另一种没有催化作用。
后者为非酶蛋白,它能决定酶促反应的专一性。 例如:乳糖合成酶催化乳糖的合成反应

48
乳糖合成酶催化乳糖的合成反应 UDP-半乳糖+葡萄糖 乳糖+UDP(1) UDP-半乳糖 + 葡萄糖 乳糖 + UDP(1)
此酶含有A及B两种蛋白质亚基:A 具有催化活性, B 为非酶蛋白。 A独存在 催化下列反应: UDP-半乳糖+N-乙酰葡萄糖胺 N-乙酰乳糖胺+UDP(2) 当B存在时,A才具有催化乳糖能力,A的专一性由B决定
 当B存在时,A才具有催化乳糖能力,A的专一性由B决定.")
49
乳糖合成酶催化乳糖的合成反应 B蛋白:是α-乳清蛋白,一个特殊的非酶蛋白。 研究表明: 反应(2)中加入B后,该反应受到抑制,
反应(1)则加强。 蛋白质A:广泛地存在于动物的各种组织中, 蛋白质B:仅存在于乳腺之中。 ——所以,只有在乳腺中才能合成乳糖
则加强。 蛋白质A:广泛地存在于动物的各种组织中, 蛋白质B:仅存在于乳腺之中。 ——所以,只有在乳腺中才能合成乳糖.")
50
3. 具有底物载体亚基的寡聚酶 由酶蛋白部分和底物载体蛋白部分组成。 例如:大肠杆菌的乙酰辅酶A羧化酶 三个蛋白质部分组成:
2个具有催化活性蛋白 生物素羧化酶 转酰基酶 1个羧基载体蛋白(BCCP) ——专一性的生物素羧基载体 ——这三个部分联结起来催化的反应分两步进行:
 ——专一性的生物素羧基载体. ——这三个部分联结起来催化的反应分两步进行:")
51
乙酰辅酶A羧化酶 三个部分联结起来催化的反应
BCCP + CO2 + ATP CO2~BCCP + ADP + Pi CO2~BCCP + RH BCCP + RCOOH 乙酰辅酶A 羧化酶 生物素羧基载体 蛋白亚基

52
二、变构酶与变构调节 变构酶(allosteric enzyme) 具有变构调节作用,多亚基构成的聚合体: 含有催化亚基(结合底物),
和调节亚基(结合变构剂)。 这催化和调节 可以在不同的亚基上, 或者在同一亚基的两个不同部位。
。 这催化和调节 可以在不同的亚基上, 或者在同一亚基的两个不同部位。")
53
变构剂(allosteric effector)
——凡能使酶分子构像改变并改变其催化活性的物质 变构激活剂allosteric effector ——引起酶活性增加的变构剂。 变构抑制剂allosteric effector ——引起酶活性降低的变构剂。

55
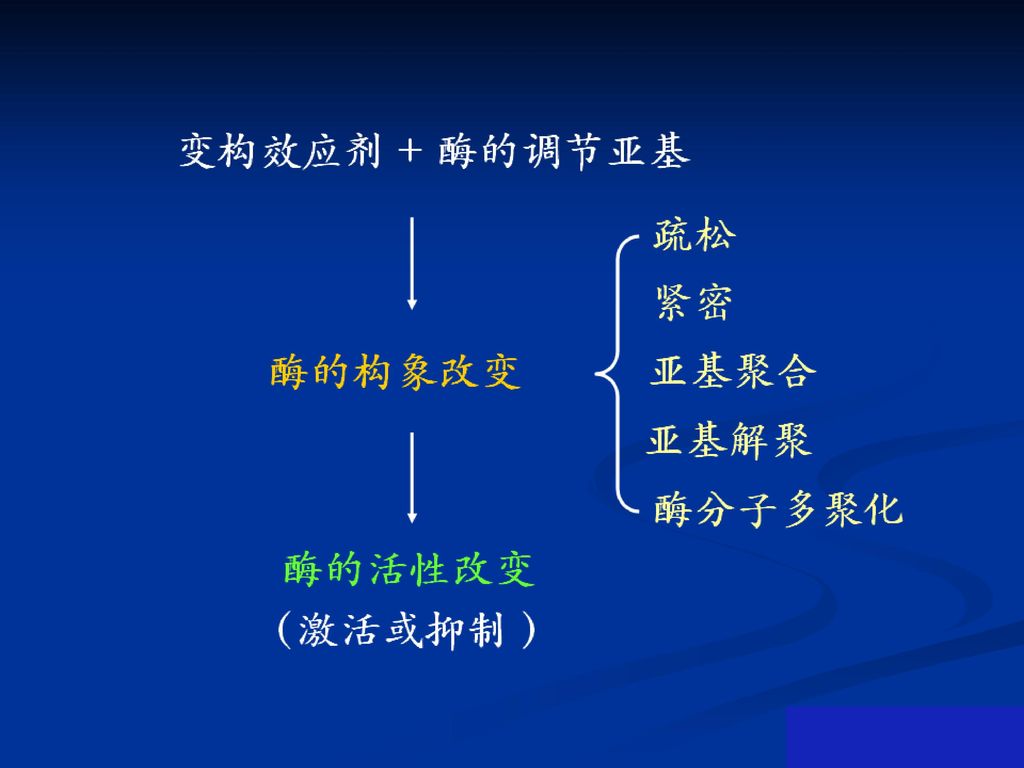
1、变构调节的机制:变构剂与酶调节亚基以非共价键特异结合,致其调节亚基的构象发生改变,进而改变其催化亚基的构象而使酶活性改变。

56
蛋白激酶A的变构调节

57
例磷酸果糖激酶: ATP是其变构抑制剂, 而ADP、AMP为其变构激活剂。

58
——变构调节 葡萄糖氧化分解提供能量,使AMP、ADP转变成ATP: 当ATP过多时:ATP抑制磷酸果糖激酶的活性, 限制葡萄糖的分解;
当ADP、AMP增多:AMP、ADP激活磷酸果糖激酶活性, 促进糖的分解。 所以机体可随时调节ATP/ADP的水平,以维持细胞内能量的正常供应。 ——变构调节

59
A (产物或中间产物) E D C B 变构酶 —常处于代谢通路的入口,通过反馈抑制,及早地调节整个代谢通路,减少不必要的底物消耗。 —
关键酶

60
变构调节的方式: 反馈抑制作用:是终产物抑制途径前面的一步关键 反应控制它自己合成的速度 前馈激活作用:当代谢途径前面步骤中产生的代谢
物激活下游某个反应的酶而发生前馈激 活作用。

61
糖和脂肪代谢酶系中某些变构酶及其变构效应剂
代谢途径 变构酶 变构激活剂 变构抑制剂 糖酵解 己糖激酶 AMP、ADP、FDP、Pi G-6-P 磷酸果糖激酶 FDP 柠檬酸 丙酮酸激酶 FDP ATP、乙酸CoA 三羧酸循环 柠檬酸合成酶 AMP ATP、长链脂酰CoA 异柠檬酸脱氢酶 AMP、 ADP ATP 糖异生 丙酮酸羧化酶 乙酰CoA、ATP AMP 果糖-1,6-二磷酸酶 5’-AMP AMP 糖原分解 磷酸化酶b AMP、G-1-P、Pi ATP、G-6-P 糖原合成 糖原合酶 G-6-P 脂肪酸合成 乙酰CoA羧化酶 柠檬酸、异柠檬酸 长链脂酰CoA 胆固醇合成 HMG-CoA还原酶 胆固醇 氨基酸代谢 谷氨酸脱氢酶 ADP、亮氨酸、甲硫氨酸 ATP、GTP、NADH 嘌呤合成 PRPP酰胺转移酶 PRPP AMP、ADP、GMP、 GDP 嘧啶合成 天冬氨酸氨基甲酰转 CTP 移酶 血红素合成 ALA合成酶 血红素

62
三、共价调节酶 ——通过其它酶对其多肽链上某些基团进行可逆的共价修饰,使其处于活性与非活性的互变状态,从而调节酶活性。
——这种作用简称为共价修饰调节 共价修饰调节也是体内快速调节代谢活动的一种重要的方式。

63
常见共价调节类型: — 磷酸化与脱磷酸化(最常见) — 乙酰化和脱乙酰化 — 甲基化和脱甲基化 — 腺苷化和脱腺苷化
— 尿苷酰化/脱尿苷酰化 — -SH与-S-S互变

64
共价修饰反应的例子 反应类型 共价修饰 被修饰的氨基酸残基 磷酸化 Tyr,Ser,Thr.His 腺苷酰化 Tyr 尿苷酰化 Tyr
反应类型 共价修饰 被修饰的氨基酸残基 磷酸化 Tyr,Ser,Thr.His 腺苷酰化 Tyr 共价修饰反应的例子 尿苷酰化 Tyr S-腺苷-Met S-腺苷-同型 Cys Glu 甲基化

65
酶的磷酸化修饰与去修饰 酶蛋白中丝、苏、酪氨酸的羟基是磷酸化的位点。 蛋白激酶 酶-OH 酶-P ATP ADP H2O Pi 磷蛋白磷酸酶

66
共价修饰的机制: 共价修饰通常在两种不同的酶的催化下发生共价修饰(或去修饰),从而引起酶分子在有活性形式与无活性形式之间进行相互转变。
目前已发现有几百种酶被翻译后都需要进行共价修饰,其中一部分处于代谢分支途径,是对其代谢流量起调节作用的关键酶。

67
共价修饰调节的生理意义广泛: 反应灵敏,节约能源,机制多样,在体内显得十分灵活,加之它们常受激素调节甚至神经的指令,导致为级联放大调节方式而日益引人注目。 共价修饰一般为耗能过程;

68
酶促化学修饰对酶活性的调节 糖原磷酸化酶 磷酸化/脱磷酸 激活/抑制 磷酸化酶b激酶 磷酸化/脱磷酸 激活/抑制
酶 化学修饰类型 酶活性改变 糖原磷酸化酶 磷酸化/脱磷酸 激活/抑制 磷酸化酶b激酶 磷酸化/脱磷酸 激活/抑制 糖原合成酶 磷酸化/脱磷酸 抑制/激活 丙酮酸脱羧酶 磷酸化/脱磷酸 抑制/激活 磷酸果糖激酶 磷酸化/脱磷酸 抑制/激活 丙酮酸脱氢酶 磷酸化/脱磷酸 抑制/激活 HMGCoA还原酶 磷酸化/脱磷酸 抑制/激活 HMGCoA还原酶激酶 磷酸化/脱磷酸 激活/抑制 乙酰CoA羧化酶 磷酸化/脱磷酸 抑制/激活 脂肪细胞脂肪酶 磷酸化/脱磷酸 激话/抑制 黄嘌呤氧化酶 SH/-S-S- 脱氢酶/氧化酶

69
例如:乳酸脱氢酶(lactate dehydrogenase,LDH)
四、同工酶(isoenzyme) 概念:在同一种属中,催化相同化学反应,但酶分子组成、结构、理化性质乃至免疫学性质和电泳行为都不同的一组酶。 例如:乳酸脱氢酶(lactate dehydrogenase,LDH)
 概念:在同一种属中,催化相同化学反应,但酶分子组成、结构、理化性质乃至免疫学性质和电泳行为都不同的一组酶。 例如:乳酸脱氢酶(lactate dehydrogenase,LDH)")
70
LDH1 LDH2 LDH3 LDH4 LDH5 H4 H3M H2M2 HM3 M4 乳酸脱氢酶( LDH):
四聚体,两种亚型 5种同工酶 (M型-骨骼肌型;H型-心肌型) LDH1 LDH2 LDH3 LDH4 LDH5 H4 H3M H2M2 HM3 M4 适应不同组织或不同细胞器在 代谢上的不同需要。
 LDH1. LDH2. LDH3. LDH4. LDH5. H4. H3M. H2M2. HM3. M4. 适应不同组织或不同细胞器在. 代谢上的不同需要。")
71
乳酸脱氢酶同工酶电泳图谱 LDH同工酶有组织特异性,LDH1在心肌含量高,而LDH5在肝、骨骼肌含量高。

72
LDH同工酶的亚单位组成及在人体各组织器官中的分布同工酶百分比
组织器官 LDH LDH LDH LDH LDH5 心肌 < <1 肾 <1 肝 骨骼肌 红细胞 肺 胰腺 — 脾 子宫

73
同工酶的生理意义:适应不同组织或细胞器在代谢上的不同需要——是代谢调节的一种重要方式。
血清检测指标: 相对含量的改变作为某脏器病变鉴别 诊断的依据 。 肝细胞受损,M4升高 血清含量高,心肌受损程 度高,心肌梗塞指标

74
同工酶在各学科中的应用 (1) 遗传学和分类学:提供了一种精良的判别遗传标志的工具。
(2) 发育学:有效地标志细胞类型及细胞在不同条件下的分化情 况,以及个体发育和系统发育的关系。 (3) 生物化学和生理学:根据不同器官组织中同工酶的动力学、 底物专一性、辅助因子专一性、酶的变构性等性质的差异, 从而解释它们代谢功能的差别。 (4)医学和临床诊断:体内同工酶的变化,可看作机体组织 损伤,或遗传缺陷,或肿瘤分化的的分子标志。
 发育学:有效地标志细胞类型及细胞在不同条件下的分化情. 况,以及个体发育和系统发育的关系。 (3) 生物化学和生理学:根据不同器官组织中同工酶的动力学、 底物专一性、辅助因子专一性、酶的变构性等性质的差异, 从而解释它们代谢功能的差别。 (4)医学和临床诊断:体内同工酶的变化,可看作机体组织. 损伤,或遗传缺陷,或肿瘤分化的的分子标志。")
75
五、固定化酶(immobilized enzyme)
将水溶性酶固定于高分子支持物上(或载体如琼脂糖、聚丙烯酰胺)使成为不溶于水,但仍有酶活性的一种酶制剂。模式图:
使成为不溶于水,但仍有酶活性的一种酶制剂。模式图:")
76
吸附法(Adsorption) 吸附法通常有物理吸附、离子吸附及螯合或金属结合法。 常用的载体:
淀粉、谷蛋白等有机类载体,活性炭、多孔玻璃、硅胶等无机类载体;大孔型的合成树脂,陶瓷以及纤维素衍生物类。阴、阳离子交换剂 吸附法(Adsorption)
")
77
共价键合法(Covalent bonds)
是酶蛋白的侧链基团和载休表面上的功能基团之间 形成共价键而固定的方法。 氨基:赖氨酸的氨基和多肽链的末端氨基; 羧基:天冬氨酸、谷氨酸的羧基和末端羧基; 酚基:酪氨酸的酚环; 巯基:半胱氨酸、蛋氨酸的巯基; 羟基:丝氨酸、苏氨酸和酪氨酸的羟基; 咪唑基:组氨酸的咪唑基; 吲哚基:色氨酸的吲哚基。

79
包埋法(Entrapment) 包埋法是将游离酶包埋于格子或微胶囊内,格子的结构可以防止酶渗出到周围的培养基中,而底物分子仍能渗入格子内与酶接触。 包埋类型可有:网格型、微囊型及脂质体液膜型。
 包埋法是将游离酶包埋于格子或微胶囊内,格子的结构可以防止酶渗出到周围的培养基中,而底物分子仍能渗入格子内与酶接触。 包埋类型可有:网格型、微囊型及脂质体液膜型。")
80
包埋法

81
催化侧链连接到环糊精上,可模拟胰凝乳蛋白酶
人工模拟酶-benzyme 环糊精分子结构 环糊精结构模型 催化侧链 环糊精 催化侧链连接到环糊精上,可模拟胰凝乳蛋白酶

82
固定化酶的优点 保持着酶的高度专一性及高催化效率还有类似于离子交换树脂的优点: 稳定性大为提高,可较长期使用或贮藏 有一定的机械强度
可以充分洗涤,不带进杂质 分离方便,可反复使用 稳定性大为提高,可较长期使用或贮藏

83
固定化酶的缺点 但由于固定化酶是通过反应而被结合在载体上,固定化过程中酶的活力难免有一定损失;
而底物则要求是水溶性的,这样才能够接触酶而发生反应; 不适宜于需要辅助因子的反应。

84
酶与医学的关系 酶与某些疾病发生的关系 酶在疾病诊断上的应用 酶在疾病治疗中的应用

85
酪氨酸酶缺陷——白化病 皮肤乳白色,毛发淡黄或银白色,瞳孔淡红,虹膜淡灰或淡红,半透明视网膜缺乏色素。

86
苯丙氨酸羟化酶缺陷——苯丙酮酸尿症 智力低下,60%患儿有脑电图异常,头发细黄,皮肤色淡和虹膜淡黄色,惊厥,尿有“发霉”臭味或鼠尿味。
87
胱硫醚合成酶缺陷——同型胱氨酸尿症 多发性血栓形成,晶体脱位,身体瘦长,蜘蛛样指(趾),轻中度智力低下
,轻中度智力低下")
88
酶活性测定与疾病诊断 ALT(GPT)——肝脏疾患 LDH同工酶谱——心肌梗塞、肝癌等 碱性磷酸酶——骨与肝脏疾患
酸性磷酸酶——前列腺转移癌 肌酸激酶——心肌梗塞、肌肉疾患

89
酶在疾病治疗中的应用 消化酶类 抗炎清创酶类 抗氧化酶类 抗栓酶类 抗肿瘤细胞生长的酶

90
1990年首次基因疗法 美国一位4岁小女孩Ashanti DiSilva,她患了一种严重联合免疫缺陷征(SCID)的疾病,机体对任何微生物都缺乏抵抗力,她只能呼吸过滤了的空气,饮用严格消毒的水和吃严格消毒的食物。 病因:腺苷酸脱氨酶(ADA)基因先天遗传缺陷。 Dr.W.French Anderson 和他的同事在 小女 孩的T细胞中插入一个正常的 ADA基因,将其注入她的血液系统。 正常的T细胞以每月增长25%的速度生 长,改善了她的免疫功能
基因先天遗传缺陷。 Dr.W.French Anderson 和他的同事在. 小女 孩的T细胞中插入一个正常的. ADA基因,将其注入她的血液系统。 正常的T细胞以每月增长25%的速度生. 长,改善了她的免疫功能.")
91
关于酶? 20世纪80年代发现某些RNA有催化活性,还有一些抗体也有催化活性,甚至有些DNA也有催化活性,使酶是蛋白质的传统概念受到很大冲击。 1.某些RNA有催化活性 1982年美国T. Cech等人发现四膜虫的rRNA前体能在完全没 有蛋白质的情况下进行自我加工, 发现RNA有催化活性 Thomas Cech University of Colorado at Boulder, USA

92
Cech和Altman各自独立地发现了RNA的催化活性,并命名这一类酶为ribozyme(核酶),2人共同获1989年诺贝尔化学奖。
1983年美国S.Altman等研究RNaseP (由20%蛋白质和80%的RNA组成),发现 RNaseP中的RNA可催化 E. coli tRNA的前体加工。 Cech和Altman各自独立地发现了RNA的催化活性,并命名这一类酶为ribozyme(核酶),2人共同获1989年诺贝尔化学奖。 Sidney Altman Yale University New Haven, CT, USA
,发现. RNaseP中的RNA可催化. E. coli tRNA的前体加工。 Cech和Altman各自独立地发现了RNA的催化活性,并命名这一类酶为ribozyme(核酶),2人共同获1989年诺贝尔化学奖。 Sidney Altman. Yale University New. Haven, CT, USA.")
95
2.抗体酶(abzyme) 抗体: 与抗原特异结合的免疫球蛋白。 抗体酶:指具有催化功能的抗体分子,在抗体分子
抗体: 与抗原特异结合的免疫球蛋白。 抗体酶:指具有催化功能的抗体分子,在抗体分子 的可变区(肽链N端)是识别抗原的活性 区域,这部分区域被赋予了酶的属性。 1986年美国Schultz和Lerner两个实验室同时在Science上发表论文,报道他们得到的具有催化活性的抗体。
是识别抗原的活性. 区域,这部分区域被赋予了酶的属性。 1986年美国Schultz和Lerner两个实验室同时在Science上发表论文,报道他们得到的具有催化活性的抗体。")
96
3.有些DNA也有催化活性 1995年Cuenoud等发现有些DNA分子亦具有催化活性。

97
核酸酶——指所有可以水解核酸的酶 DNA酶(DNase):专一降解DNA。 RNA酶 (RNase):专一降解RNA。
依据底物不同分类 DNA酶(DNase):专一降解DNA。 RNA酶 (RNase):专一降解RNA。 依据切割部位不同 核酸内切酶:分为限制性核酸内切酶和 非特异性限制性核酸内切酶。 核酸外切酶:5´→3´或3´→5´核酸外切酶。
:专一降解DNA。 RNA酶 (RNase):专一降解RNA。 依据切割部位不同. 核酸内切酶:分为限制性核酸内切酶和. 非特异性限制性核酸内切酶。 核酸外切酶:5´→3´或3´→5´核酸外切酶。")
98
核酸酶的功能 参与DNA的合成与修复、RNA合成后的剪接等基因复制和基因表达
负责清除多余的、结构和功能异常的核酸,还可以清除侵入细胞的外源性核酸 在消化液中降解食物中的核酸以利吸收 是体外重组DNA技术中的重要工具酶

Similar presentations

 反应部位 器官,细胞内定位 生理意义 代谢调节 主要调节点,主要变构抑制剂、变构激活剂 各代谢途径之间的联系和调控.>")
:核蛋白体组成成分 转移 RNA ( tRNA ):转运氨基酸 信使 RNA ( mRNA ):蛋白质合成模板 不均一核 RNA ( hnRNA ):成熟 mRNA 的前体 小核 RNA ( snRNA ):>")



 2004.1 酶.>")



蛋白质的降解: 外源蛋白的消化 内源性蛋白的选择性降解 (2)氨基酸的分解代谢:>")


.>")



受体拮抗剂 抑制性氨基酸受体受体拮抗剂 神经肽Y受体拮抗剂>")


