Download presentation
Presentation is loading. Please wait.

1
水生生物学——养殖水域生态学 第 十章 初级生产力

2
第一节 初级生产力及其测定方法 一、生物生产力及有关概念 二、水体中的初级生产过程 三、初级生产力的测定方法

3
一、生物生产力及有关概念 生物生产力是生态系统提供生物产品高低的一种性能,它既是生态系中能量流动和物质循环这两大功能的综合表征,又是生物种群通过同化作用生产或积累有机质的能力。水体生物生产力是与土壤肥力相类似的概念,不仅取决于水体的特性,而且与种群的特性密切联系。

4
现存量或生物量 现存量(standing crop)或生物量(biomass):指水体单位面积或单位体积内生物有机质的重量。例如底栖生物用g/m2或kg/m2来表示;浮游生物量通常用g/m3或mg/L来表示;鱼类现存量通常用kg/hm2来表示。水体单位面积内所能维持的最高的鱼重量称为水体鱼载力。
或生物量(biomass):指水体单位面积或单位体积内生物有机质的重量。例如底栖生物用g/m2或kg/m2来表示;浮游生物量通常用g/m3或mg/L来表示;鱼类现存量通常用kg/hm2来表示。水体单位面积内所能维持的最高的鱼重量称为水体鱼载力。")
5
生产量 生产量(production):指一定时间内单位面积(m2,hm2)或单位水体积(m3,L)内所产生的生物有机质的重量,现存量和生产量也常用能量单位(J,kJ)表示。
:指一定时间内单位面积(m2,hm2)或单位水体积(m3,L)内所产生的生物有机质的重量,现存量和生产量也常用能量单位(J,kJ)表示。")
6
收获量 收获量(yield):一定时间内捕捞出的那一部分产量。池塘和其他小水体可以一次把鱼全部捕出,收获量和鱼产量较接近(加上死亡的鱼就是生产量),大水面的渔获量占生产量或多或小的一部分,它与捕捞技术和需要有关。
:一定时间内捕捞出的那一部分产量。池塘和其他小水体可以一次把鱼全部捕出,收获量和鱼产量较接近(加上死亡的鱼就是生产量),大水面的渔获量占生产量或多或小的一部分,它与捕捞技术和需要有关。")
7
周转率和周转时间 周转率(turnover rate):一定时间内新增加的生物量(P)与这段时间内平均生物量(B)的比率(通称P/B系数)。周转率的倒数(B/P)就是周转时间(turnover time),它表示生物量周转一次所需时间。
:一定时间内新增加的生物量(P)与这段时间内平均生物量(B)的比率(通称P/B系数)。周转率的倒数(B/P)就是周转时间(turnover time),它表示生物量周转一次所需时间。")
8
初级产量、次级产量 根据生物的营养特点,生产量可分为初级产量(primary production)和次级产量(secondary production)。自养生物通过光合作用或化合作用在单位时间、 单位面积或容积内所合成的有机质的量称为初级产量,异养生物在单位时间内同化、生长和繁殖而增加的生物量或所贮存的能量,称为次级产量。生产量是生产力的体现,一般说来,初级产量和初级生产力是同义词,但次级产量不一定代表次级生产力。
和次级产量(secondary production)。自养生物通过光合作用或化合作用在单位时间、 单位面积或容积内所合成的有机质的量称为初级产量,异养生物在单位时间内同化、生长和繁殖而增加的生物量或所贮存的能量,称为次级产量。生产量是生产力的体现,一般说来,初级产量和初级生产力是同义词,但次级产量不一定代表次级生产力。")
9
初级产量的进一步划分 对于初级产量要区分初级毛产量(gross primary production)、初级净产量(net primary production)和群落净产量(net community production)三个概念。初级毛产量指自养生物所固定的总能量或所合成的全部有机质量(包括已被本身消耗的);初级净产量指自养生物本身呼吸消耗以外剩余的能量或有机质量;群落净产量,也称生态系净产量,指整个生态系中自养生物所固定的能量除去全部生物呼吸消耗以外的剩余部分,即: 群落净产量=初级净产量-异养生物呼吸量
、初级净产量(net primary production)和群落净产量(net community production)三个概念。初级毛产量指自养生物所固定的总能量或所合成的全部有机质量(包括已被本身消耗的);初级净产量指自养生物本身呼吸消耗以外剩余的能量或有机质量;群落净产量,也称生态系净产量,指整个生态系中自养生物所固定的能量除去全部生物呼吸消耗以外的剩余部分,即: 群落净产量=初级净产量-异养生物呼吸量.")
10
胞外产物 植物在生活过程中经常向水中释放溶解有机质(DOM),这一部分有机质可占光合产物的相当比重,称为胞外产物。
从表8-1可见,胞外产物占光合合成碳的百分比变化很大,从7%到83%,Fogg指出最高值可达95%。但在培养条件下则低得多,一般仅0.5%~4.5%。

11
水柱呼吸量 水柱呼吸量代表水层中有机质的分解速率,前已指出,浮游植物毛产量减去水柱群落的呼吸量就是群落或生态系的净产量。
水柱呼吸量也就是黑瓶中氧的消耗量,主要包括浮游植物本身以及细菌和浮游动物呼吸的耗氧量。据Винберг(1976)对原苏联10个湖泊和2个水库的统计,浮游植物呼吸平均占群落呼吸量的19.1%(4%~32%),细菌占57.4%(44%~73%),浮游动物占23.5%(5%~34%),但在富营养程度很高的武汉东湖,藻类群落的呼吸速率(66 mgC/m3·h)高于异养细菌(51 mgC/m3·h)。
对原苏联10个湖泊和2个水库的统计,浮游植物呼吸平均占群落呼吸量的19.1%(4%~32%),细菌占57.4%(44%~73%),浮游动物占23.5%(5%~34%),但在富营养程度很高的武汉东湖,藻类群落的呼吸速率(66 mgC/m3·h)高于异养细菌(51 mgC/m3·h)。")
12
水柱毛产量(P)和呼吸量(R)的比值是生态系统生产力特点的一个重要指标。在贫营养型湖P/R通常小于1,群落或生态系统净产量(Pe)常为负值;在富营养型湖P/R值近于1。
养鱼池P/R值与施肥种类有关,国外多施无机肥料,P/R大于1,有时达到6~7(Винбрг 等,1965)。P/R值过高,表明初级产量的利用率低,物质循环速率不高,是肥效低的标志。中国高产鱼池以施有机肥料为主,P/R值常低于1或近于1,有些兼施化肥的鱼池大于1,如南汇渔场成鱼池以化肥养鱼,P/R值高达3.41。
。P/R值过高,表明初级产量的利用率低,物质循环速率不高,是肥效低的标志。中国高产鱼池以施有机肥料为主,P/R值常低于1或近于1,有些兼施化肥的鱼池大于1,如南汇渔场成鱼池以化肥养鱼,P/R值高达3.41。")
13
二、水体中的初级生产过程 (一) 光合作用 (二)化合作用
 光合作用 (二)化合作用")
14
(二氧化碳) (水) (光能) (葡萄糖) (氧)
(一)光合作用 绿色植物的光合作用是水体内自养生产过程的主要部分。自养生物是借助太阳能合成有机质的,光合作用的全过程并不是都需要光,而是需光的光反应和不需光的暗反应两个不可分割过程的综合。 光反应的第一步是叶绿素吸收光能使水分解: H2O → (H) + (OH) 两个OH-再形成H2O并放出O2 2(OH) → H2O2 → H2O + 1/2 O2 光分解时分离出来的氢原子(H)经过一段复杂的化学反应,和二氧化碳形成碳水化合物。这段过程不需要光,为暗反应。 整个光合作用的方程式表示如下: 6CO2 + 6H2O kJ=C6H12O6 + 6O2 (二氧化碳) (水) (光能) (葡萄糖) (氧)
光合作用. 绿色植物的光合作用是水体内自养生产过程的主要部分。自养生物是借助太阳能合成有机质的,光合作用的全过程并不是都需要光,而是需光的光反应和不需光的暗反应两个不可分割过程的综合。 光反应的第一步是叶绿素吸收光能使水分解: H2O → (H) + (OH) 两个OH-再形成H2O并放出O2 2(OH) → H2O2 → H2O + 1/2 O2 光分解时分离出来的氢原子(H)经过一段复杂的化学反应,和二氧化碳形成碳水化合物。这段过程不需要光,为暗反应。 整个光合作用的方程式表示如下: 6CO2 + 6H2O kJ=C6H12O6 + 6O2. (二氧化碳) (水) (光能) (葡萄糖) (氧)")
15
6CO2 + 12H2S+(光能)= C6H12O6 + 12S +6H2O
藻类的其他色素也能吸收光能,但吸收的能量必须传递给叶绿素才起作用,因为只有叶绿素能进行水的光分解作用。 在光合过程中光反应和暗反应交替进行,光反应阶段约经10-5s,暗反应经历时间就慢得多,约为光反应的10 000倍。 光合作用中所形成的葡萄糖进一步转化为难溶性的高分子糖类、脂肪和其他贮藏物质。如果只考虑到有机质成分中碳、氢、氧、氮、磷五种最主要的元素,就可以用下列近似方程式表示: 214CH2O + 16NH3 + HPO O2 →C106H263O110N16P + 108CO2+107H2O 在静水水体中一些具叶绿素的细菌(光合细菌)能够利用光能使二氧化碳和硫化氢合成葡萄糖,这种光合作用的方程式如下: 6CO2 + 12H2S+(光能)= C6H12O6 + 12S +6H2O 例如某些红色硫细菌在湖泊和池塘中都很常见,当水体中缺氧但有光线透入时,它们可以利用硫化氢而大量繁殖起来,使水面呈鲜红色。
能够利用光能使二氧化碳和硫化氢合成葡萄糖,这种光合作用的方程式如下: 6CO2 + 12H2S+(光能)= C6H12O6 + 12S +6H2O 例如某些红色硫细菌在湖泊和池塘中都很常见,当水体中缺氧但有光线透入时,它们可以利用硫化氢而大量繁殖起来,使水面呈鲜红色。")
16
(二)化合作用 化能营养性的自养过程,仅在特殊情况下才有显著作用。进行这一过程的主要是硝化细菌、硫化细菌、铁细菌、氢细菌、沼气细菌等。这类细菌最常集中于好气条件和嫌气条件的交界处,因为在它们的生命活动中既需要氧又需要从有机质的嫌气性分解中形成的还原性化合物。在水体中具备这种条件的主要是水底土壤和底层水中。因此化合细菌的数量也是在水底土壤中最多,底层水中次之,表层水最少。因而,化能营养的强度在水层中通常只有水底土壤中的几十分之几甚至几百分之几。如在雷滨水库的水层中化合作用的最高值为0.061 mgC/L,约等于水底土壤中这个数值的1/25,而在表层水中这一数值(0.057 mgC/L)又只及底层水中的1/11左右。 化合作用的强度随温度而增高。用雷滨水库水底土壤所作试验表明:温度从0.5℃增到20℃,化合强度从每天0.95~1.8 mgC/L提高到3.09~67 mgC/L。该水库7月份水层中的化合作用值较9月份高几倍。
又只及底层水中的1/11左右。 化合作用的强度随温度而增高。用雷滨水库水底土壤所作试验表明:温度从0.5℃增到20℃,化合强度从每天0.95~1.8 mgC/L提高到3.09~67 mgC/L。该水库7月份水层中的化合作用值较9月份高几倍。")
17
三、初级生产力的测定方法 (一)收获量法 (二)黑白瓶测氧法 (三)放射性14C示踪法 (四)叶绿素法
收获量法 (二)黑白瓶测氧法 (三)放射性14C示踪法 (四)叶绿素法")
18
(一)收获量法 主要用于水生维管束植物和大型藻类生产量的测定。在一定面积内将所有植株连根取出,洗净风干到重量不变时秤重即得出单位面积的生物量,前后两次生物量之差即为其生产量。此法所得为净产量。由于采样间隔期间可能有一部分生物量被动物摄食和微生物分解,测定值通常偏低。
收获量法 主要用于水生维管束植物和大型藻类生产量的测定。在一定面积内将所有植株连根取出,洗净风干到重量不变时秤重即得出单位面积的生物量,前后两次生物量之差即为其生产量。此法所得为净产量。由于采样间隔期间可能有一部分生物量被动物摄食和微生物分解,测定值通常偏低。")
19
(二)黑白瓶测氧法 原理:藻类光合作用的途经和同化产物虽然因种类组成而有差异,但一般都可以用下列简单公式表示:
6CO2 + 6H2O C6H12O6 + 6O2 由于氧的生成量和有机质的合成量之间存在着一定的当量关系,即放出1 g氧相当于合成0.37g碳、0.937 g葡萄糖或14.70 J,因而通过测定水中溶氧的变化可间接计算有机质的生成量。

20
测定方法 将所采的水样分装于透明的白瓶和不透明的黑瓶,再放于原采样水层曝光一定时间(一般为24 h)后计算前后溶氧的变化。
白瓶中藻类进行光合作用和呼吸作用,同时细菌、浮游动物等异养生物也进行呼吸。黑瓶中仅进行群落的呼吸,因此: 白瓶中溶氧-原初溶氧=群落净产量 白瓶溶氧-黑瓶溶氧=藻类毛产量 藻类的净产量只能以假定其呼吸量占毛产量的一定比例来估算(一般估计占0.2~0.3)。
。")
21
方法的优缺点 此法简单易行,也有一定准确度,已广泛用于湖库池沼浮游植物生产力的测定,但缺点也不少:(1)不能测定藻类净产量;(2)灵敏度低,对贫营养型水体不适用;(3)瓶内外条件不尽相同,瓶内藻类易死亡,有时细菌附着瓶壁加速养分的周转,这些都能影响测定的准确度; (4)有光和黑暗中呼吸强度不完全相同;(5)玻瓶容积大小和曝光时间长短都会影响结果,容器越大,产氧量越高。武汉东湖浮游植物毛产量在连续曝光24 h的测定结果显著低于每次曝光2 h的全天累计结果。 此法也可在室内模拟自然条件进行,即灌满水样后黑白瓶挂于水族箱中,模拟同样的光照和温度条件下进行曝光。
不能测定藻类净产量;(2)灵敏度低,对贫营养型水体不适用;(3)瓶内外条件不尽相同,瓶内藻类易死亡,有时细菌附着瓶壁加速养分的周转,这些都能影响测定的准确度; (4)有光和黑暗中呼吸强度不完全相同;(5)玻瓶容积大小和曝光时间长短都会影响结果,容器越大,产氧量越高。武汉东湖浮游植物毛产量在连续曝光24 h的测定结果显著低于每次曝光2 h的全天累计结果。 此法也可在室内模拟自然条件进行,即灌满水样后黑白瓶挂于水族箱中,模拟同样的光照和温度条件下进行曝光。")
22
(三)放射性14C示踪法 将一定数量的放射性碳酸氢盐(H14CO3-)或碳酸盐(14CO32-)加入到已知二氧化碳总量的水样瓶中,曝光一定时间后将藻类滤出,干燥后测定藻细胞内14C数量,即可计算被同化的总碳量。
放射性14C示踪法 将一定数量的放射性碳酸氢盐(H14CO3-)或碳酸盐(14CO32-)加入到已知二氧化碳总量的水样瓶中,曝光一定时间后将藻类滤出,干燥后测定藻细胞内14C数量,即可计算被同化的总碳量。")
23
14C法优缺点 此法的采样、曝光等过程与黑白瓶法基本相似,但灵敏度高得多,可用于贫营养型水体和大洋中初级生产力的测定,也可采用模拟法在室内进行工作。 此法的缺点是设备和技术较难掌握,此外藻类分泌出的溶解有机质(胞外产物)流入滤液中,可能产生巨大的误差。因此,必须同时测定滤液中的放射性。如不需要区分细胞和胞外产物的产量时,可将曝光后的水样不经过滤直接测定其放射性。 一般认为14C法所得数值为净产量或接近于净产量,但也有作者认为仍属于毛产量,可能是介于两者之间的一种数值。
流入滤液中,可能产生巨大的误差。因此,必须同时测定滤液中的放射性。如不需要区分细胞和胞外产物的产量时,可将曝光后的水样不经过滤直接测定其放射性。 一般认为14C法所得数值为净产量或接近于净产量,但也有作者认为仍属于毛产量,可能是介于两者之间的一种数值。")
24
(四)叶绿素法 在一定条件下光合作用强度与细胞内叶绿素含量直接相关,因此根据叶绿素量和藻类的同化指数可计算其生产量。
测定叶绿素量目前已广泛作为浮游植物的定量方法,与此同时测定现场的同化系数进而计算初级生产力,是简便又易掌握的方法。

25
第二节 决定初级生产力的因素 初级生产力取决于自养生物的现存量及其组成、养分、光、温度、水的运动以及动物的摄食等生态因子。

26
一. 现存量 自养生物生物量是初级生产力的物质基础,又是生产力的产物。
当养分充足和光照条件良好时,净产量和生物量都迅速增大,单位面积光合速率随生物量的增加而增高,水柱中藻类种群密度也越来越高。与此同时由于自荫作用使透明度降低,下层水中光照不足使光合速率急降,光合层的厚度缩减。当生物量极高时,初级生产只在表层几厘米到十几厘米薄层进行。水柱的毛产量急剧下降,随着呼吸量的增大净产量降到零或负值。

27
在海洋、湖库等天然水体浮游植物量不会达到严重自荫程度,生产量一般随现存量而增高, Brylinsky(1980)根据IBP大量资料的系统分析表明,湖库初级生产力与现存量有极显著的相关,又如武汉东湖水中叶绿素a浓度增加1 μg/L,水柱初级产量相应增加0.28 gO2/m2(王骥等,1981),但在施肥鱼池中过高的浮游植物量常使生产力下降。据Куэъмичева (1970)材料养鱼池(约1 m深)浮游植物毛产量(以叶绿素a计)在20 μg/L(约相当于生物量40~60 mg/L)时达到最高值。雷衍之(1986)研究越冬池冰下生物增氧的氧平衡数学模式时,得出最佳浮游植物量为25~50 mg/L,与上述数值接近。但生产力与生物量的关系比较复杂,最适生物量还与本身的种类组成有关。
根据IBP大量资料的系统分析表明,湖库初级生产力与现存量有极显著的相关,又如武汉东湖水中叶绿素a浓度增加1 μg/L,水柱初级产量相应增加0.28 gO2/m2(王骥等,1981),但在施肥鱼池中过高的浮游植物量常使生产力下降。据Куэъмичева (1970)材料养鱼池(约1 m深)浮游植物毛产量(以叶绿素a计)在20 μg/L(约相当于生物量40~60 mg/L)时达到最高值。雷衍之(1986)研究越冬池冰下生物增氧的氧平衡数学模式时,得出最佳浮游植物量为25~50 mg/L,与上述数值接近。但生产力与生物量的关系比较复杂,最适生物量还与本身的种类组成有关。")
28
同化指数 通常用单位生物量或单位叶绿素量在单位时间内合成的有机质量(或产氧量)作为植物的同化指数.
在饱和光照条件下,每毫克叶绿素a在1 h中的产氧量(mg)代表其光合能力,据Константинов(1979),浮游植物光合能力平均为3.7 mg C/mg·h,Talling(1975)认为此值多在1~10 mg C/mg·h之间。在具体条件下浮游植物的同化指数与种类组成、年龄和生理状态有关。藻类细胞越小,光合能力越强,处于指数增长期的藻类其光合能力远高于平衡期的老化细胞。
代表其光合能力,据Константинов(1979),浮游植物光合能力平均为3.7 mg C/mg·h,Talling(1975)认为此值多在1~10 mg C/mg·h之间。在具体条件下浮游植物的同化指数与种类组成、年龄和生理状态有关。藻类细胞越小,光合能力越强,处于指数增长期的藻类其光合能力远高于平衡期的老化细胞。")
29
二. 养分 自养生物在光合过程中,除了需要碳源外,尚需要各种矿物质,其中具有最大生态意义的应该是那些构成生物体质的主要成分,而生境中含量又常常不足的化学元素。水生植物最易缺乏的是磷和氮,其次是碳、硅、钾等。

30
二氧化碳-1 二氧化碳是光合作用的碳源,天然水中二氧化碳的浓度随温度和盐度而变化,一般在0.2~0.5 ml/L之间。在浮游植物丰富的水中,白天光合作用每小时可消耗二氧化碳0.2~0.3 ml/L以上,因此需要不断地输入二氧化碳,才能补偿其迅速的消耗。在水中二氧化碳的来源除大气溶解和水生生物呼吸放出以外,还能从二氧化碳平衡系统中得到,一般是不会缺乏的。但在浮游植物极为繁盛或低碱度和低硬度的浅水水体中在高温强光的情形下,也会出现二氧化碳的缺乏。在我国肥水鱼池和日本的养鳗池都常出现因二氧化碳不足而抑制浮游植物光合作用的情况。这时水失去鲜绿色而发白(隐约的乳白色),我国渔农称为水质老化,松井魁等(1971)称为“白化”。有时甚至因二氧化碳不足使浮游植物大量死亡而引起泛塘现象。在实验条件下,增强光照强度的同时,增加二氧化碳可使植物的光合作用速率增强,反之单纯增加照度,有时甚至使光合速率减弱。
,我国渔农称为水质老化,松井魁等(1971)称为 白化 。有时甚至因二氧化碳不足使浮游植物大量死亡而引起泛塘现象。在实验条件下,增强光照强度的同时,增加二氧化碳可使植物的光合作用速率增强,反之单纯增加照度,有时甚至使光合速率减弱。")
31
二氧化碳-2 各种植物最适的二氧化碳浓度是不同的,而且还随照度及其他因子的变化而变化,如粉核小球藻和四刺栅藻最大光合率时的二氧化碳浓度为0.1%,水藓(Fontinalis)则为0.35%。柱孢鱼腥藻(Anobaena cylindrica)在5500 lx的照度下,水温15℃时为0.10%,25℃时为0.25%。 二氧化碳含量过多也能起抑制作用,如二氧化碳浓度超过30%时对依乐藻(Elodea)和水藓的光合作用就有不良的影响,超过0.5%时对柱孢鱼腥藻有抑制作用。
则为0.35%。柱孢鱼腥藻(Anobaena cylindrica)在5500 lx的照度下,水温15℃时为0.10%,25℃时为0.25%。 二氧化碳含量过多也能起抑制作用,如二氧化碳浓度超过30%时对依乐藻(Elodea)和水藓的光合作用就有不良的影响,超过0.5%时对柱孢鱼腥藻有抑制作用。")
32
二氧化碳-3 很多藻类(特别是红藻)的光合强度仅依赖于二氧化碳浓度,但是生活在高pH水中的一些植物,除了吸收游离二氧化碳以外,还能直接利用碳酸氢根中的碳,如铜绿微囊藻、多变鱼腥藻(Anabaena variabilis)和其他一些蓝藻都能利用碳酸氢根为碳源,普通小球藻和水藓等也能利用这种碳。四尾栅藻(Scenedesmns quadricauda)和钝顶螺旋藻(Spirulina plateusis)甚至对碳酸氢根的利用率超过对二氧化碳,但后一种情况是罕见的。实际上碳酸氢根中的碳大多数是在转化为碳酸根沉淀和放出二氧化碳时被利用的。在这种情形下形成的难溶性碳酸盐粘附在植物体上,在体外形成一层白色皮膜从而阻挡光线,并使植物体下沉。两种情况都能恶化光合条件,所以保证光合过程进行的碳源,还是以溶解二氧化碳为最好。
的光合强度仅依赖于二氧化碳浓度,但是生活在高pH水中的一些植物,除了吸收游离二氧化碳以外,还能直接利用碳酸氢根中的碳,如铜绿微囊藻、多变鱼腥藻(Anabaena variabilis)和其他一些蓝藻都能利用碳酸氢根为碳源,普通小球藻和水藓等也能利用这种碳。四尾栅藻(Scenedesmns quadricauda)和钝顶螺旋藻(Spirulina plateusis)甚至对碳酸氢根的利用率超过对二氧化碳,但后一种情况是罕见的。实际上碳酸氢根中的碳大多数是在转化为碳酸根沉淀和放出二氧化碳时被利用的。在这种情形下形成的难溶性碳酸盐粘附在植物体上,在体外形成一层白色皮膜从而阻挡光线,并使植物体下沉。两种情况都能恶化光合条件,所以保证光合过程进行的碳源,还是以溶解二氧化碳为最好。")
33
无机氮 水中无机氮主要以铵、亚硝酸盐、硝酸盐和溶解气体氮(分子氮)形式存在。除某些固氮蓝藻外,水生植物只能利用前三种形式氮;而亚硝酸盐一般仅在低浓度时可为某些藻类所利用,因此作为光合作用的主要氮源是氨和硝酸盐。当这两种氮同时存在时,藻类首先利用铵氮,一般是铵氮被吸收尽以后,才利用硝酸盐氮。这是因为:藻类必须把硝酸盐氮还原为铵氮以后才能同化利用。这个“准备”过程需要硝酸盐还原酶参加并且多耗费能量。但是,处于氮饥饿状态的藻类,可以同时吸收铵氮和硝酸盐氮。由于铵氮进入藻类细胞的速度远较硝酸盐为快,通常较后者低得多的含量就能满足藻类的需要。但是也有一些藻类更善于利用硝酸盐氮。
形式存在。除某些固氮蓝藻外,水生植物只能利用前三种形式氮;而亚硝酸盐一般仅在低浓度时可为某些藻类所利用,因此作为光合作用的主要氮源是氨和硝酸盐。当这两种氮同时存在时,藻类首先利用铵氮,一般是铵氮被吸收尽以后,才利用硝酸盐氮。这是因为:藻类必须把硝酸盐氮还原为铵氮以后才能同化利用。这个 准备 过程需要硝酸盐还原酶参加并且多耗费能量。但是,处于氮饥饿状态的藻类,可以同时吸收铵氮和硝酸盐氮。由于铵氮进入藻类细胞的速度远较硝酸盐为快,通常较后者低得多的含量就能满足藻类的需要。但是也有一些藻类更善于利用硝酸盐氮。")
34
各种藻类对氮浓度的要求 各种藻类在同化作用中要求的氮浓度是不一样的,通常绿藻和蓝藻较高,硅藻较低,如据古谢娃的培养试验,硅藻在硝酸盐氮0.01~0.8 mg/L时生长最好,而绿球藻类则必须5 mg/L,团藻类必须2~5 mg/L时生长最好。

35
磷酸盐 藻类同化作用的磷源主要是磷酸盐。各种藻类要求的含磷量也不一样,如美星杆藻可以在磷低到0.1 μg/L浓度下吸收磷。这种硅藻的最适磷浓度为0.002~0.01 mg/L,磷浓度增高到0.2 mg/L时即可能起抑制作用;大约1 μg/L能产生2~5 mm3/L的生物量。其他藻类最大密度时的磷浓度和单位容积生物量所需要的最低磷量一般较此为大。平板藻和桅杆藻在磷为45 μg/L以内达到最大密度,栅藻则必需在磷500 μg/L时、红色颤藻(Oscillatoria rubesceus)必须达到3000 μg/L磷时才达到最大密度。
必须达到3000 μg/L磷时才达到最大密度。")
36
现已证实,藻类能够利用有机磷源供代谢和生长的需要,如甘油磷酸脂、植酸盐(phytin)等有机磷都可满足藻类的需要。藻类利用有机磷的能力明显地依赖于磷酸酶的活动能力。这是一种在环境磷酸盐浓度很低时诱发合成的酶。但是并不是任何藻类在磷酸盐不足时都能诱发合成磷酸酶,所以有机磷源的作用显然不及磷酸盐。 长期以来在水化学和水生生物学研究中,只注意磷酸盐的作用。以后在研究藻类对养分的需要时发现,培养液中最适生长的磷浓度总较其生长的天然水为高,起初以为是由于湖水中有某种未知的有机生长因子存在所致,近年用示踪原子研究表明:水中有机磷可以不断地转化为磷酸盐。因此溶解无机磷量和浮游植物生长情况的关系不是主要的,重要的是磷的循环速度和颗粒磷同溶解磷之间的交换。因此用总磷量代替现在通用的磷酸盐量来表示水体的肥度是较为适宜的,因为在任何时候水中大部分磷是以结合粒状磷或有机磷状态存在的。
等有机磷都可满足藻类的需要。藻类利用有机磷的能力明显地依赖于磷酸酶的活动能力。这是一种在环境磷酸盐浓度很低时诱发合成的酶。但是并不是任何藻类在磷酸盐不足时都能诱发合成磷酸酶,所以有机磷源的作用显然不及磷酸盐。 长期以来在水化学和水生生物学研究中,只注意磷酸盐的作用。以后在研究藻类对养分的需要时发现,培养液中最适生长的磷浓度总较其生长的天然水为高,起初以为是由于湖水中有某种未知的有机生长因子存在所致,近年用示踪原子研究表明:水中有机磷可以不断地转化为磷酸盐。因此溶解无机磷量和浮游植物生长情况的关系不是主要的,重要的是磷的循环速度和颗粒磷同溶解磷之间的交换。因此用总磷量代替现在通用的磷酸盐量来表示水体的肥度是较为适宜的,因为在任何时候水中大部分磷是以结合粒状磷或有机磷状态存在的。")
37
硅酸盐 硅在水中以溶解性硅酸盐、胶体及悬浮物存在,溶解性硅酸盐和胶体硅大多能为硅藻直接吸收,是有效硅化合物。海水浮游植物以硅藻为主,而海水中有效硅含量又较低,常成为初级产量的限制因子。内陆水域在春末形成硅藻水华时,也可能缺硅。 其他如铁、锰等微量元素以及某些有机微养分的缺乏,也可能影响初级生产。

38
藻类对养分的吸收率 各种藻类对营养盐类浓度要求的不同亦即对养分吸收能力的差别取决于细胞中酶对养分的亲和力大小。一般可用米氏方程表示藻类对养分的吸收率(V): V=Vmax·S/( Ks + S) 式中Vmax为最大吸收率,S为养分浓度,KS为半饱和常数或称米氏常数。藻类的KS越小,表明其酶对养分的亲和力越大,达到最大吸收率(Vmax)所需养分浓度(S)越低,种群越易在低养分的水中增长。因此KS值可作为某种藻类正常生长所需的有效形式的营养盐类的临界浓度,可作为对养分吸收能力的指标。实验表明,保证正常初级生产所需的养分浓度约为临界浓度的3倍,即S=3 Ks。
所需养分浓度(S)越低,种群越易在低养分的水中增长。因此KS值可作为某种藻类正常生长所需的有效形式的营养盐类的临界浓度,可作为对养分吸收能力的指标。实验表明,保证正常初级生产所需的养分浓度约为临界浓度的3倍,即S=3 Ks。")
39
藻类的KS值因种类、对环境的适应性和细胞大小而不同。海洋浮游植物对NO3-—N的KS值,近岸种1. 0 μg/L,大洋种仅0
藻类的KS值因种类、对环境的适应性和细胞大小而不同。海洋浮游植物对NO3-—N的KS值,近岸种1.0 μg/L,大洋种仅0.2 μg/L。从表8-2可见,在培养条件下海洋浮游植物大洋种和贫营养海区种类的Ks值均较低于近海种和富营养型海区种类。淡水藻类的KS值通常高于海洋种类。淡水浮游植物对磷的KS值多在0.2~0.8 μmol/L之间,正常生活所需浓度约为0.6~2.4 μmol/L。硅藻中星杆藻对PO4—P的KS值为2.0 μg/L,而绿藻的纤维藻和栅藻则高达10.0和100~1000 μg/L。小型种类因相对面积大,吸收能力也强,KS值较大型种类为低。实验表明当藻类细胞大小在0.1~10 μm之间变化时,对NO2-和NH4+—N的KS值均随体积增大而增高。硅藻对SiO2—Si的KS值多在0.2~3.4 μmol/L之间,以2 μmol/L最常见。

40
N:P 各种养分的相对含量对生产力也有重要影响。水生植物(藻类、水草)细胞和组织中碳、氮、磷的原子量比值为106∶16∶1,按重量比约为40∶7∶1。这3种元素中某一种的相对量低于此比值,其他两种就成为生理需要上的多余,而不足的这一种即成为限制因子,因此,一般认为氮和磷之比大于7时为缺磷,小于7时为缺氮。 然而,浮游植物细胞中氮和磷的含量不仅因种类而有差异,并且同一种类还因生活条件而有很大变化,氮和磷的重量比在缺氮培养液中可能低于1.5∶1,在缺磷培养液中则可能达到15∶1以上。有些藻类在缺磷时还会诱发合成磷酸酶使有机磷分解出磷酸供本身利用。此外,磷的循环速率较氮为快,在热带的George湖,氮的周转时间为0.66 d,磷仅0.5 d,因而磷在光合作用中的利用效率较氮为高。因此,在实际应用时一些作者用较宽的N/P指标:如Ryding(1980)以溶解性氮和溶解性磷的比值5~12为指标,低于5时缺氮,大于12时缺磷,同时用总氮和总磷比值10~17为指标,低于10时缺氮,高于17时缺磷。
细胞和组织中碳、氮、磷的原子量比值为106∶16∶1,按重量比约为40∶7∶1。这3种元素中某一种的相对量低于此比值,其他两种就成为生理需要上的多余,而不足的这一种即成为限制因子,因此,一般认为氮和磷之比大于7时为缺磷,小于7时为缺氮。 然而,浮游植物细胞中氮和磷的含量不仅因种类而有差异,并且同一种类还因生活条件而有很大变化,氮和磷的重量比在缺氮培养液中可能低于1.5∶1,在缺磷培养液中则可能达到15∶1以上。有些藻类在缺磷时还会诱发合成磷酸酶使有机磷分解出磷酸供本身利用。此外,磷的循环速率较氮为快,在热带的George湖,氮的周转时间为0.66 d,磷仅0.5 d,因而磷在光合作用中的利用效率较氮为高。因此,在实际应用时一些作者用较宽的N/P指标:如Ryding(1980)以溶解性氮和溶解性磷的比值5~12为指标,低于5时缺氮,大于12时缺磷,同时用总氮和总磷比值10~17为指标,低于10时缺氮,高于17时缺磷。")
41
P-N-C 通常贫营养型湖水中磷的来源较少,浮游植物的发展受到磷的限制,而多余的氮和碳不能被充分利用。在富营养化过程中首先是磷的增加。由于磷极快被利用并且循环速度很快,所以总储量虽然增大但瞬时浓度不一定增大甚至常常降低;同时变化幅度增大。磷的增加促进了浮游植物的产量。当浮游植物大量发展时,氮的消耗量可能超过补充量,因此引起上层水中氮的不足。这时固氮蓝藻的出现扩大了氮的来源。 在磷和氮都不缺乏的罕见情况下,碳也可能成为限制因素。在软水湖中较易出现碳的缺乏,施肥池塘当形成强烈水华时,可能因二氧化碳不足而限制生产力。

42
特例 一般说来花岗岩地区湖库水中富含磷和硅,而缺氮;变质岩地区则富含氮。岩石和土壤受淋溶时进入水中的磷较多,因而地下水带入的磷较多。大气降水带入的氮较多,因而集水区面积小的水体易缺氮。火山活动地区湖水氮、磷的比例较特殊,如内蒙东部达里湖磷酸盐的含量甚至高于无机氮。

43
三. 光 光是光合作用的能源,光照强度、光谱组成和光照时间均影响光合速率。在补偿点以上,光合速率随光强而加速,在达到饱和点以后光合强度即不随光强而增高,并且光强继续增高反而抑制光合过程。

44
光抑制现象 一般水的深度呈算术级数增加时,光照强度则呈指数递减的趋势。通常晴天水面的光强已达到对藻类起抑制作用,浮游植物初级生产力的最高水层不在水面,而在水面以下某一深度 (常位于相当于透明度一半的水层),但阴天则位于水表面。由于光强随深度而递减,到了某一水层光强低到仅相当浮游植物的补偿点,净产量等于零,这一水层称为补偿水层,所在深度称为补偿深度。通常补偿深度为透明度的2~3倍。
,但阴天则位于水表面。由于光强随深度而递减,到了某一水层光强低到仅相当浮游植物的补偿点,净产量等于零,这一水层称为补偿水层,所在深度称为补偿深度。通常补偿深度为透明度的2~3倍。")
45
四. 温度 温度影响光合作用中暗反应的速率。温度升高暗反应加快,如1℃时暗反应过程要0.4s,25℃时仅0.04s。在0~20℃间,温度每增高1℃海洋浮游植物的光合作用增强2.2倍 (Ведерников ,1976),武汉东湖水温每升高1℃,初级生产量大致增长0.17~0.20gO2/(m2·d)。一般情况只有光强达到饱和点时,光合速率和生产力才随温度而升高,反之单方面提高温度则因藻类呼吸强度的增加而降低生产力。
,武汉东湖水温每升高1℃,初级生产量大致增长0.17~0.20gO2/(m2·d)。一般情况只有光强达到饱和点时,光合速率和生产力才随温度而升高,反之单方面提高温度则因藻类呼吸强度的增加而降低生产力。")
46
实例 在深水湖库真光层下部,初级生产力通常受光强的限制,而在真光层上部温度常成为限制因子。在光线可透射水底的浅水湖沼,温度和CO2成为控制光合速率的决定性因子。 在天然水体温度对初次生产力的影响是一个比较复杂的问题,它除了直接影响光合速率外,还通过对水体中养分的再循环和其他生物学及理化过程的影响而间接影响生产力。例如,有时温度降低使光合速率减弱,但由于增高了藻类光合色素和酶的含量从而更有效地利用光能。有时温度的升高不仅增强光合速率,还可促进对养分的吸取率。

47
五. 垂直水流 各种原因引起的水的垂直涡动,对初级生产力均有重要影响。垂直水流一方面将深层养分带到真光层供藻类在光合过程中利用,另一方面又可能将藻类带到无光层而停止光合作用,因此当混合层的厚度≤真光层时,垂直水流将促进生产力的提高;反之当混合层深度超过补偿深度时,浮游植物有相当时间处于黑暗水层,生产力将下降。

48
六 动物滤食 当浮游植物现存量不高时,浮游动物和鱼类的滤食常使藻类生物量和生产量下降,但当浮游植物密度很大时,则动物的滤食将起到调节密度改善光照条件和提高生产力的作用。

49
关于鲢、鳙等滤食性鱼类对浮游植物初级生产力的影响,研究结果尚有分岐。Kajak(1979)研究白鲢对浮游植物种群的影响,发现在水温分层的水体中放养白鲢后,浮游植物的产量和生物量都降低,因为大量的浮游植物形成鱼粪沉积水底。反之在不分层的浅水水体或者当有底层鱼存在时,放养白鲢后不仅浮游植物的产量增高,并且其生物量也常增高。这是因为白鲢在滤食浮游植物同时也食进大量浮游动物,由于浮游动物量的降低,浮游植物的消耗量不但未增高,甚至反而减少。此外白鲢的滤食活动促进了水层中物质循环速度,使养分的周转率加大有利于浮游植物的增长。滤食动物的影响,随其密度而有变化。据东湖的试验,当鲢的放养密度为105~780 kg/hm2,同时进行施肥和投饵的条件下,浮游植物生物量和生产量均显著增长,当鲢放养密度为450~3000 kg/hm2时,两者都显著下降(阮景荣等 1995)。
研究白鲢对浮游植物种群的影响,发现在水温分层的水体中放养白鲢后,浮游植物的产量和生物量都降低,因为大量的浮游植物形成鱼粪沉积水底。反之在不分层的浅水水体或者当有底层鱼存在时,放养白鲢后不仅浮游植物的产量增高,并且其生物量也常增高。这是因为白鲢在滤食浮游植物同时也食进大量浮游动物,由于浮游动物量的降低,浮游植物的消耗量不但未增高,甚至反而减少。此外白鲢的滤食活动促进了水层中物质循环速度,使养分的周转率加大有利于浮游植物的增长。滤食动物的影响,随其密度而有变化。据东湖的试验,当鲢的放养密度为105~780 kg/hm2,同时进行施肥和投饵的条件下,浮游植物生物量和生产量均显著增长,当鲢放养密度为450~3000 kg/hm2时,两者都显著下降(阮景荣等 1995)。")
50
贝类的下行效应 在近海以贝类为主的浮筏养殖区内,贝类通过生物过滤作用对水体中浮游生物和颗粒有机质有着巨大的影响。80年代初,庙岛海峡浮游植物量年平均值高达3.0×106个细胞/m3,而开展贝类养殖后1988、1989年两年的平均值为0.54×106个细胞/m3,仅为前者的1/6。大量研究结果表明,在某些海区滤食性贝类可以控制浮游植物的生物量,而且在以下几种条件下,这种控制会得到加强:一是水体较浅;二是滞留时间长;三是滤食性贝类的生物量很高。我国浅海筏式养殖海区具备上述条件,尤其是在养殖的中后期(8月份以后),养殖贝类的生物量以接近养殖容量,因此贝类摄食对海区浮游植物的控制尤为明显(杨红生,1998)。
,养殖贝类的生物量以接近养殖容量,因此贝类摄食对海区浮游植物的控制尤为明显(杨红生,1998)。")
51
第三节 浮游植物初级生产力 一、内陆水体 (一)分布概况 (二)现存量及P/B系数 (三)生物量和生产力的时空分布 二、 海 洋
(二)全世界海洋的估算 (三)新生产力及有关概念 (一)现存量
全世界海洋的估算. (三)新生产力及有关概念. (一)现存量.")
52
概述 浮游植物是水圈的主要生产者,浮游植物产量占世界海洋初级生产力的90%以上,内陆水体除浅水湖泊和大湖的沿岸带水草和附生藻类在年产量中可能占主要地位外,一般也是以浮游植物为主要生产者。水越深、越肥,浮游植物的作用越显著。有些内陆盐沼因盐度高,水草难以丛生,初级产量主要由浮游植物和底生藻类组成,在水深、流缓、河床宽阔的大河下游,浮游植物产量也起主要作用。

53
一、内陆水体 在测定浮游植物初级生产力时,有几个主要指标: 1.水柱的日产量 Pd(mg/m2·d),每平方米水面每日的毛产量;
2.水柱年产量或生长期产量,Pa(mg/m2·a)一般用平均日产量乘生长期日数; 3.生长期中最大日产量Pmax(mg/m2·d)。
一般用平均日产量乘生长期日数; 3.生长期中最大日产量Pmax(mg/m2·d)。")
54
分布概况 内陆水域水柱日产量差别极大,毛产量可以从零点几克到10 gC以上。印度一个池塘当发生蓝藻水华时达到13~24 gC/m2·d,水柱日产量(Pd)与最大日产量Pmax和透明度(T)之间有一定关系(БуΛьон ,1979): Pd=Pmax(0.90±0.30)T 或简化为: Pd=Pmax·T 生长期或年毛产量变化更大,据IBP 43个湖和12个水库资料,从极地到热带变动于125.6~ kJ/m2之间,极值相差近400倍,并且同一水体不同年份也有较大变化,最大值出现于印度北纬10~12°N水体。此外加拿大有些湖泊年产量仅41.9~83.7kJ/m2。
T 或简化为: Pd=Pmax·T 生长期或年毛产量变化更大,据IBP 43个湖和12个水库资料,从极地到热带变动于125.6~ kJ/m2之间,极值相差近400倍,并且同一水体不同年份也有较大变化,最大值出现于印度北纬10~12°N水体。此外加拿大有些湖泊年产量仅41.9~83.7kJ/m2。")
55
初级产量与纬度极显著负相关 日照强度和照射时间及水温均随纬度而变化,同时低纬度水温高,养分周转快。所以全球初级产量有从热带向极地降低的趋势。Brylinsky(1980)根据IBP资料分析,表明纬度和初级产量有极显著的负相关。然而,同一纬度内都存在不同营养类型的水体,其生产力也是不同的。
根据IBP资料分析,表明纬度和初级产量有极显著的负相关。然而,同一纬度内都存在不同营养类型的水体,其生产力也是不同的。")
56
不同营养类型的日净产量-1 据Liken的统计,全世界富营养型湖平均日净产量为600~8000 mgC/m2,中营养型湖为250~1000 mgC/m2,贫型湖为50~300 mgC/m2,超贫营养型湖低于50 mgC/m2。 在河流和水库由于水的透明度较低,初级产量通常低于富营养型和中营养型湖。

57
Винберг(1960) 据Винберг(1960)的材料,湖泊按浮游植物产量可以分为四类:
1.最高产的高养分多营养型湖,如苏联黑湖和科星诺湖,日产量为7.5~10 gO2/m2,年产量在10467~14653kJ/m2之间; 2.富营养湖,日产量为2.5~7.5 gO2/m2,年产量4186~10467 kJ/m2; 3.中营养型湖,按水的深度和透明度的不同,产量变化很大,日产量1~7.5 gO2/m2,年产量1256~10467 kJ/m2; 4.低产的沼泽型或贫营养型湖,前者日产量不超过0.1~0.2 mgO2/L或0.5~1.0 gO2/m2,年产量低于1256kJ/m2;后者日产量一般低于0.1 mgO2/L,当水深、透明度高时,达到0.5~1.0 gO2/m2,年产量628~1256 kJ/m2。

58
何志辉,1987 根据80年代对四大水系102个湖库的调查材料(何志辉,1987),中国湖泊水库浮游植物生产力按日毛产量(gO2/m2)可分5种类型: 1.低型,≤1, 占总数26.7% 2.中型,1.0~3.0,占总数35.6% 3.较高型,3.0~5.0,占总数27.7% 4.高型,5.0~10.0,占总数8.9% 5.极高型,>10.0,占总数1%

59
中国湖库浮游植物日毛产量特点 可见中国湖库浮游植物日毛产量以中型和较高型为主,亦即63.4%水体在1.0~5.0 gO2/m2之间。从地域分布看,珠江水系以低型为主,占本水系湖库数70%;长江水系以中型和低型为主,两者占本水系总数的72.7%,黄河水系全为中型和较高型,黑龙江(包括辽河)水系以较高型和高型为主,两者占本水系总数的62.2%。总的趋势是从南向北递增。 按年毛产量(kJ/m2)可分为3种类型: 1.低型,<1256,占总数8%; 2.中型,1256~4186,占总数32%; 3.高型,>4186,占总数60%。 可见年毛产量92%属高型和中型,虽然生长期日数珠江和长江水系远高于黑龙江和黄河水系,但年产量和日产量同样地呈从南向北增高的趋势。
水系以较高型和高型为主,两者占本水系总数的62.2%。总的趋势是从南向北递增。 按年毛产量(kJ/m2)可分为3种类型: 1.低型,<1256,占总数8%; 2.中型,1256~4186,占总数32%; 3.高型,>4186,占总数60%。 可见年毛产量92%属高型和中型,虽然生长期日数珠江和长江水系远高于黑龙江和黄河水系,但年产量和日产量同样地呈从南向北增高的趋势。")
60
中国浮游植物日产量和年产量分布特点 全球湖库初级产量有从低纬度向高纬度降低的趋势,而中国浮游植物日产量和年产量则呈相反的趋势。这种情况决定于我国土壤和气候分布的特点。东北和华北地区土壤肥力较高且气候干燥,水中营养盐类和有机质较易积累。此外北方生长期中日照时间较长,晴天日数多,有利于初级生产的进行。反之珠江和长江流域土壤肥力较低且常受雨水的稀释,生长期云量大,晴天日数少,日照时间短。珠江水系pH较低(有时近酸性)也是不利因素之一。这些原因造成我国浮游生物初级生产力地理分布和世界其他地域显然不同的这种特点。
也是不利因素之一。这些原因造成我国浮游生物初级生产力地理分布和世界其他地域显然不同的这种特点。")
61
(二)现存量及P/B系数 生物量是浮游植物丰盛度的重要标志,当前浮游植物量主要用叶绿素含量和以细胞体积换算的生物重量来表示,由于不同藻类细胞中叶绿素含量有很大差别,同一种类所含叶绿素量也因外界条件和本身生理状况而有变化,此外还难免有测定方法上的一些问题,两者很难准确换算,当需要对两种方法所得数据相互比较时,只能粗略估算。大致说来,叶绿素a量多在生物量的0.2%到0.6%之间,富营养型水体高于贫营养型水体,一般可采用1 mgB≈3μgchla估算。

62
Pn =Lc + Lw + Ls + Lz + Lm + B
Lz——浮游动物、贝类和某些鱼类的滤水率相当可观,如浮游甲壳类1 mg湿重每天可过滤0.25~0.5L水,在生物量达到2~6 mg/L的富营养型水体,每天可过滤全水量的10%~90%,鲢、鳙和淡水贝类每克湿重每天的滤水率可达2L左右。可见在养殖水体浮游植物每天几乎要被过滤0.5~1次以上,生物量不断被消耗; Lw——在河流和河川型水库有相当一部分浮游植物随水流流出,水的交换率越大,流出量越多。 Ls——原生质比重大于1,浮游植物虽有各种浮游适应,但当水体静止时也要下沉。 Lm——包括病毒和真菌寄生,代谢产物的抑制和非生物环境的急变等造成的死亡以及藻类的自然死亡,这部分损失率目前还不清楚。

63
水质按浮游植物量的分级 从渔业利用出发,内陆水体水质可按浮游植物量分为10个等级(何志辉,1985)(表8—5),其中0级为贫营养型水,1~2级为中营养型水,3~4级为富到超富营养型水,5~6级为最佳的鱼池肥水,7级为肥水和老水的过渡区,亦即肥水的上限。
(表8—5),其中0级为贫营养型水,1~2级为中营养型水,3~4级为富到超富营养型水,5~6级为最佳的鱼池肥水,7级为肥水和老水的过渡区,亦即肥水的上限。")
65
P/B值及其影响因素 浮游植物量的周转率多用毛产量(Pg)和生物量(B)的比值表示。P/B值在不同水体和不同时期均有明显变化,主要与种类组成、水体深浅和生物量本身的高低有关: 1.种类组成 浮游植物群落中藻类细胞越小,光合能力和生产力就越高,生物量则越低,P/B值越高。淡水浮游植物日P/B值变动于0.1~5.0之间,大型种类(>30 μm)平均0.7,小型种类(<30 μm=达2.0(Walters等,1980)。 2.水域深度 浅水水体光合层在整个水层中所占比例较大,生产量较高。如果深度不超过透明度的2~3倍,从水面到水底全水层都可进行光合作用,生产量和P/B值达到最高值。 3.浮游植物生物量 生物量过低,生产量受限制,生物量过高又因自荫作用降低生产量。 中国湖泊水库浮游植物的日P/B系数平均值多在0.3~0.8之间,高产鱼池常在0.5~1.5之间。
平均0.7,小型种类(<30 μm=达2.0(Walters等,1980)。 2.水域深度 浅水水体光合层在整个水层中所占比例较大,生产量较高。如果深度不超过透明度的2~3倍,从水面到水底全水层都可进行光合作用,生产量和P/B值达到最高值。 3.浮游植物生物量 生物量过低,生产量受限制,生物量过高又因自荫作用降低生产量。 中国湖泊水库浮游植物的日P/B系数平均值多在0.3~0.8之间,高产鱼池常在0.5~1.5之间。")
66
(三)生物量和生产力的时空分布 1. 水平分布 水体的不同区段的环境条件有所不同,浮游植物生物量和生产力也有差异。湖越大,生境越多样化,这种差异也越明显。
生物量和生产力的时空分布 1. 水平分布 水体的不同区段的环境条件有所不同,浮游植物生物量和生产力也有差异。湖越大,生境越多样化,这种差异也越明显。")
67
1. 水平分布-湖泊 水体的不同区段的环境条件有所不同,浮游植物生物量和生产力也有差异。 。湖越大,生境越多样化,这种差异也越明显。例如太湖7个湖区浮游植物量变动于1.73~9.28 mg/L,极值相差5倍多,生产力也是以生物量最高的五里湖和三山湖为首,其他湖泊生物量和生产力的水平分布基本上也是一致的。

68
水平分布-水库 但在河川型水库中,生物量和生产力的水平分布常呈相反的趋势。如辽宁清河水库浮游植物量上游或中游为下游的1.6~18.6倍,生产力则下游为中游的1.5~2.0倍。这是因为这类水库浮游植物以硅藻为主,随着从上游到下游水流的递减,硅藻因细胞壁较重首先随泥砂等悬浮物而下沉,流程越长,下沉越多。因此以硅藻为主的河川型水库浮游植物量从上游向下游呈递降趋势,有时因上游水的含砂量较大,生物量的高峰移到中游。但生产力的分布主要受光照条件的控制,随着水流速度的减弱和悬浮物的下沉,下游透明度增大,许多静水型藻类迅速增长,生产力逐渐达到高峰。其他河川型水库也有类似情况,如珠江水系泗维河水库初级产量中游为上游1.9倍,下游为上游的2.8倍。

69
水平分布-池塘 养鱼池面积虽小,浮游植物量和生产力也有明显的水平差异。通常迎风岸由于对岸风流带来的浮游植物积累起来,生物量较高;背风岸由于底层缺浮游植物的水上升,生物量较低。此外某些鞭毛藻类有趋光运动,常集中在光照较强水区,使池水呈现渔农所谓的“半塘红、半塘绿”现象。由于自荫作用,生物量密集区的生产力一般不高。

70
2. 垂直分布-1 深水湖泊和水库下游深水区浮游植物种量常有显著的垂直差异,特别是在夏季水温分层期,上层远高于下层。如镜泊湖夏季0.5 m水层生物量12.0 mg/L,以蓝藻为主,10 m深层降到3.0 mg/L,以硅藻为主。在春秋循环期,随着水团的混合,种、量的垂直分布趋于均匀,但在以硅藻为主的河川型水库中,由于藻类的下沉常出现中层和底层高于表层的情况。

71
垂直分布-2 由于水层光照强度随深度而递减,浮游植物生产力更是集中于上层,中国湖泊水库0~1 m水层的初级产量通常占水柱产量的30%~50%以上,如辽宁二龙山水库达到70%~75%,武汉东湖达到44.4%~63.3%。多数湖库最高生产力水层位于相当一半透明度深处的水层,但在纬度低、透明度又很大的深水水库,如珠江水系的锦江水库,0~1 m水层的产量仅占水柱产量的13.4%,最高生产层的深度移到4~6 m,接近于补偿深度。

72
3. 昼夜分布 浮游植物光合作用仅在白天进行,并且光合强度上午大于下午。如武汉东湖毛产量从6∶00开始增高,10∶00~12∶00达到最高值,12∶00~14∶00开始下降,18∶00以后降到最低点(戎克文等,1995)。在相近条件下,下午低于上午的原因有各种解释,如下午养分枯竭、藻类本身代谢产物积累、呼吸作用的增强、细胞中光合色素淡化和浮游动物滤食增加等等,但Westlake(1980)认为也可能是一种类似生物钟的内在现象。
。在相近条件下,下午低于上午的原因有各种解释,如下午养分枯竭、藻类本身代谢产物积累、呼吸作用的增强、细胞中光合色素淡化和浮游动物滤食增加等等,但Westlake(1980)认为也可能是一种类似生物钟的内在现象。")
73
生物量分布的昼夜节律 生物量的分布也常呈现昼夜节律。黑海浮游植物量以21∶00为最高,此后下降,到翌日1∶00达最低值,5∶00以后又开始回升。夜间下降的原因是浮游动物从深层上升强烈摄食,白天回升是因为藻类本身的繁殖。昼夜藻类数量差别可达2~3倍以上。 但是在其他情况下,浮游植物的昼夜变化特点可能两样。例如在无锡郊区河埒口公社养鱼池中,扁平膝口藻数量从清晨5∶00以后就开始减少,17∶00降到最低点,以后又逐渐回升,到半夜1∶00达到最高值,这种情况可能和鲢、鳙等鱼类滤食节律有关。膝口藻是白鲢的优质食物,白鲢主要在白天摄食,特别在4∶00~8∶00之间,这种情况和膝口藻数量从5∶00到9∶00间急剧降低完全一致。

74
4.季节分布 由于太阳辐射能的周期性变化和随之而来的其他环境条件的变化,导致浮游植物生产力和生物量的季节变化,变化状况与水体所处的纬度、深度和营养类型等有密切关系。

75
冬季初级生产力 在温带中或富营养型湖泊,冬季在低光照、短日照和低温下,浮游植物生产力和生物量一般较低。当水面封冰时,如果冰层不厚且无积雪复盖,冰下的照度通常远高于藻类的补偿点,光合作用仍可不同程度地进行着。如黑龙江省一些越冬池明冰时,甚至冰下1~2 m深的水层,照度仍有3000~ lx,接近于某些藻类的最适光照,一昼夜产氧量常在1 mg/L以上,最高达2.71 mg/L,浮游植物量可达10 mg/L以上(李永函等,1979)。又如印度东北部Sylvar湖,冰下3个月浮游植物生产量可占全年的四分之一(Wetzel,1975)。当冰层由厚的乌冰组成或冰上长期覆雪时,净产量转为负值,由于冰下无湍流藻类易下沉,生物量降到最低点。冰下浮游植物主要由隐藻、甲藻、金藻等鞭毛藻类组成。
。又如印度东北部Sylvar湖,冰下3个月浮游植物生产量可占全年的四分之一(Wetzel,1975)。当冰层由厚的乌冰组成或冰上长期覆雪时,净产量转为负值,由于冰下无湍流藻类易下沉,生物量降到最低点。冰下浮游植物主要由隐藻、甲藻、金藻等鞭毛藻类组成。")
76
春季 春季随着冰层融化,在对流和风力混合下产生水层的垂直流转,养分从底层上升,加上光照和温度的升高,为浮游植物的发展创造良好的条件。首先是硅藻种群的大量增长。硅藻高峰期一般不超过3个月,此后由于硅酸盐枯竭(<0.5 mg/L=或其他原因(动物滤食、菌类寄生等),种群开始消退并为绿球藻类或某些甲藻所取代。这段时期如果生产层的养分能及时得到补充,生产力仍然很高。但由于浮游动物的强烈滤食,生物量难以增长。
,种群开始消退并为绿球藻类或某些甲藻所取代。这段时期如果生产层的养分能及时得到补充,生产力仍然很高。但由于浮游动物的强烈滤食,生物量难以增长。")
77
夏、秋季节 随着绿藻的发展,水中含氮量降到极低点,因而中夏以后固氮蓝藻(鱼腥藻、束丝藻等)取代绿藻而急剧增长。蓝藻此时占优势的原因还与高的温度(25℃以上)、强光照、高pH以及较少被食等有关。蓝藻水华期生物量很高,但生产力通常下降。 秋后光照的减弱和温度下降等原因,引起蓝藻种群突然性地消退。此后随着秋季水层的垂直混合,环境条件又和春季类似,因而出现了硅藻的第二次高峰,在湖泊秋季高峰一般不及春季。

78
评述 上述浮游植物季节分布是温带中等深度湖泊的模式,其中所有环节不一定都能出现,有时硅藻高峰之前有一个金藻的优势期,有时硅藻水华以后直接出现蓝藻水华而没有绿藻和甲藻的优势期。在贫营养型湖硅藻全年占优势通常在晚春或初夏有一个弱的高峰。在水浅的富营养型湖,从春到秋都保持高的生产力和生物量,随着优势种的更替,生产力和生物量都呈不规则的波动,蓝藻的作用增强。超富营养型湖和肥水池塘中鞭毛藻类、蓝藻、绿藻占极大优势,高的生物量常达到自荫程度,生产力和生物量形成相互消长的颤动状态。热带湖泊浮游植物季节变化较不显著,极地湖泊生产力和生物量的高峰常在夏季。

79
生产力和生物量变化 在季节变化中,生产力和生物量的极值未必同时出现,生产力极值的变幅要大于生物量。在一个高山湖Vorderer Finstertaler See中,生物量最高值为最低值的5倍,而日毛产量变幅达270倍,这是因为沉淀、流出、动物滤食的作用随生产力的提高而加强,对生物量的变化起了调控作用。生物量年变幅随水体的富营养化而增大,最高值与最低值之比在贫营养型湖<30,富营养型湖>30(Westlake,1980)。
。")
80
水库浮游植物的季节分布 中国位于东亚季风气候区,每年由海洋暖气流带来的春讯和夏讯,使集雨区大量养分流进湖泊,浮游植物生产力和生物量的高峰多在洪水期过后的夏秋季。如贵州省草海7~9月的生产量占全年产量的47%。 水库浮游植物的季节分布,除自然因素外,还受到管理和使用方式等人为因素的影响。一般从春到夏由于农业用水增加,库水大量排放,浮游植物量来不及积累即随水流排出,中夏排水量虽减少但由于雨水的稀释和透明度降低,生产力和生物量未见显著提高。秋季水位增高,消落区被淹没的大量旱生和沼生植物腐烂分解起着施肥的作用;此外此时水位稳定,随着悬浮物的沉淀,透明度开始升高。两方面结合起来有力地促进了浮游植物的秋季高峰。如东北的龙凤山水库、清河水库、云峰水库、珠江水系的澄碧河水库、泗维河水库、锦江水库等都是这种情况。辽宁二龙山水库秋季含砂量仍然很高,直到冬季封冰以后泥砂才充分下沉,春汛融雪水带入大量养分,导致其生产力和生物量高峰出现在春末夏初。

81
5. 年间变化 同一水体在正常情况下各年份浮游植物的时空分布特点相对说来是稳定的,但当气候特殊变化或在人类的影响下,可能出现较大的变化。例如,清河水库70年代浮游植物以硅藻为主,生物量在上游和中游远高于下游,高峰期在秋季,但1978年夏季奇旱,水位降到最低点,库水停止排放,由于水变浅未出现温跃层。浮游植物量在富养分、高水温和强光照下提前积累并形成特大的夏季高峰。同时优势种类也从硅藻转为静水性的甲藻(飞燕角藻)和蓝藻(铜绿微囊藻),生物量也集中于水质最肥的中游区,中游生物量达到上游的4倍和下游的10倍多。
和蓝藻(铜绿微囊藻),生物量也集中于水质最肥的中游区,中游生物量达到上游的4倍和下游的10倍多。")
82
二 海 洋 海洋浮游植物初级生产力的地区差异极为显著,日产量每平方米从几毫克到100毫克以上。年产量从不到1克到几百克之间。其分布的总趋势是: 1. 高纬度高,低纬度低; 2. 从近岸向外海降低; 3. 三大洋中印度洋高于大西洋和太平洋; 4. 同纬度和同海区中,上升流处生产力最高。

83
(一)分布概况 热带海洋光照充足,水温高,但常年存在着温跃层,阻碍了水层的垂直混合和养分从底层向上层输送,从而限制了浮游植物的发展,其生产力和生物量全年都较低,并且变化不大。 温带海洋夏季出现温跃层,因此生产力和生物量的峰值在春秋两季,夏冬较低。季节变化极为显著。 极地海洋养分不受限制,但除夏季短暂的一段时间外,都受冰雪覆盖,光照不足限制了浮游植物的发展,仅在夏季有一个生产力和生物量的高峰。南极有些海区由于长的日照时间和上升流提供丰富的养分,日产量曾达到1.56gC/m2·d。 近岸和沿岸海区初级生产力高是因为可以从陆地输入大量营养盐,同时水较浅,其深度通常小于补偿深度,也很少出现持久的温跃层。

84
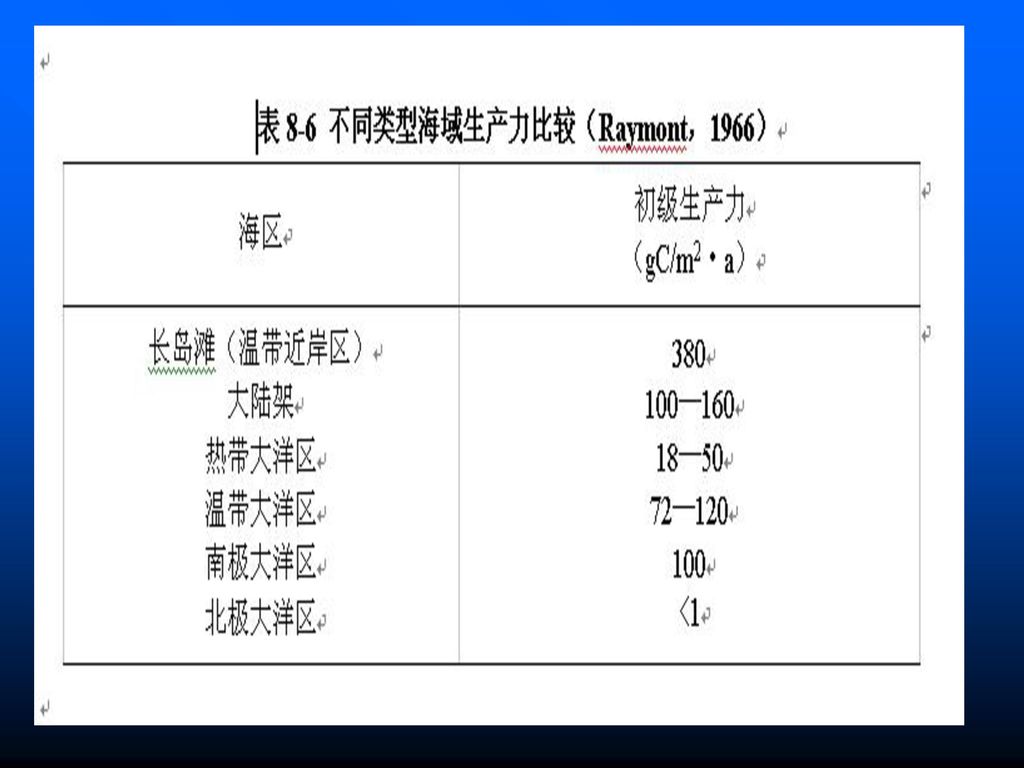
水平分布 根据Raymont(1966)从宏观上的推算,近岸海区生产力超过大陆架2倍以上,温带和南极大洋区远超过热带大洋区,北极大洋区最低(表8-6) 在三大洋中,印度洋年产量81.0 gC/m2·a,年均日产量0.89 gC/m2·d,大西洋为69.4 gC/m2·d和0.52 gC/m2·d,太平洋为46.4 gC/m2·d和0.710 gC/m2·d。 全球的大量资料表明,不论是寒、温带和热带、近海或大洋,上升流海区的生产力总是特别高,如非洲南岸上升流区浮游植物生产力达3.8 gC/m2·d ,秘鲁沿岸上升流区甚至高达10 gC/m2·d。在同一个上升流区的生产力,一般也是在近海较高,而在外海较低。例如,美国俄勒冈上升流区的初级生产力,近海为162 gC/m2·a, 外海仅61 gC/m2·a。据Ryther(1969)估算,全世界海洋浮游植物年产量,大洋:沿岸:上升流之比约为1:2:6。
估算,全世界海洋浮游植物年产量,大洋:沿岸:上升流之比约为1:2:6。")
86
(一)全世界海洋的估算 海洋占地球面积的四分之三,全世界海洋究竟能提供多少初级产量不仅决定着海洋渔业资源的潜力,对生物圈所能容纳人类的总数量也有重要影响。因此,一些学者对全世界海洋的初级生产力作过估算,虽然对各海区的初级生产力进行了很多调查,但是要把这些结果进行比较却不容易。因为各海区调查的深入程度不同,而且调查方法与分析统计方法不一样也使结果有很大的差别。

87
实 例-1 Riley(1944)最早用黑白瓶测氧法的数据估计全世界海洋初级生产量为1.55×109t C/a。
实 例-1 Riley(1944)最早用黑白瓶测氧法的数据估计全世界海洋初级生产量为1.55×109t C/a。 Sverdrup等(1955)用14C法的数据估算,为12-15×109t C/a。两种方法的结果相差10倍。 Ryther(1969)把世界海洋分大洋区、沿岸区和上升流区三种类型,估计它们的平均产量分别为50、100和300(gC/m2·a),他认为全世界海洋最可能的初级产量约为20×109tC/a( Koblentz-Mishke等(1970)根据7000个以上测站所得数据,把世界海洋水域分成五种类型。其中,亚热带中部的贫营养水的初级生产水平仅70 mgC/(m2·d),而在沿岸水域可达1000 mgC/(m2·a)。他们估计全世界浮游植物的总初级产量为23×109tC/a。
最早用黑白瓶测氧法的数据估计全世界海洋初级生产量为1.55×109t C/a。 Sverdrup等(1955)用14C法的数据估算,为12-15×109t C/a。两种方法的结果相差10倍。 Ryther(1969)把世界海洋分大洋区、沿岸区和上升流区三种类型,估计它们的平均产量分别为50、100和300(gC/m2·a),他认为全世界海洋最可能的初级产量约为20×109tC/a( Koblentz-Mishke等(1970)根据7000个以上测站所得数据,把世界海洋水域分成五种类型。其中,亚热带中部的贫营养水的初级生产水平仅70 mgC/(m2·d),而在沿岸水域可达1000 mgC/(m2·a)。他们估计全世界浮游植物的总初级产量为23×109tC/a。")
88
实 例-2 70年代以后,一些学者对海洋初级生产力的估计都超过上述的估计值。例如,Platt(1975)和Berger(1987)估计为30×109tC/a,而Shushkina(1985)根据1968—1982年间从130个站位获得的资料估计为56×109tC/a。这些差别的原因之一是各自对特定的海区(直接测定)代表类型的范围估计不同,并用这些估计作为推算整个海洋初级生产力的依据。
和Berger(1987)估计为30×109tC/a,而Shushkina(1985)根据1968—1982年间从130个站位获得的资料估计为56×109tC/a。这些差别的原因之一是各自对特定的海区(直接测定)代表类型的范围估计不同,并用这些估计作为推算整个海洋初级生产力的依据。")
89
海洋超微型浮游生物 据近几年研究,海洋超微型浮游生物(原核细菌和真核细菌)在海洋初级生产中占有极其重要的地位,它们的贡献率高达60%。目前,对蓝细菌(Cyanobacteria)的研究表明,它们在光补偿层占叶绿素的25%—90%。这是初级生产力研究中的一个重要进展。过去把热带反气旋区看作“沙漠”不毛之地,认为那里浮游植物生产量昼夜不超过0.18g/m2。但目前这种观点有了改变,特别是这类海区也有数量众多的金枪鱼、箭鱼、鲐鱼、竹刀鱼、竹荚鱼及大量食微生物的种类,这种现象只有在浮游植物产量较高的情况下才能出现。
在海洋初级生产中占有极其重要的地位,它们的贡献率高达60%。目前,对蓝细菌(Cyanobacteria)的研究表明,它们在光补偿层占叶绿素的25%—90%。这是初级生产力研究中的一个重要进展。过去把热带反气旋区看作 沙漠 不毛之地,认为那里浮游植物生产量昼夜不超过0.18g/m2。但目前这种观点有了改变,特别是这类海区也有数量众多的金枪鱼、箭鱼、鲐鱼、竹刀鱼、竹荚鱼及大量食微生物的种类,这种现象只有在浮游植物产量较高的情况下才能出现。")
90
(一) 新生产力及有关概念 “新生产力”一词最早是由Dugdale & Goering(1967)提出来的。他们认为,进入初级生产者细胞内的任何一种元素都可以划分为新结合的和再循环的两类。其中,N是构成细胞的主要元素,而且其N/C和N/P的比值也相对较为稳定,因此,用N描述初级生产者的生长比用其它元素(如C、P)更为精确。此外,N常常是海洋环境中的限制性营养元素,因而建立N源基础上的生产力研究更具有实际意义。根据以上观点,他们提出:在真光层中再循环的N为再生N(主要是NH+4—N),由真光层之外提供的N为新生N(主要是N-3—N),由再生N源支持的那部分初级生产力称为再生生产力(regeneration production),由新生N源支持的那部分初级生产力称为新生产力(new production)。显然新生产力和再生生产力之和就是总初级生产力。表达新生产力的单位与初级生产力相同。
提出来的。他们认为,进入初级生产者细胞内的任何一种元素都可以划分为新结合的和再循环的两类。其中,N是构成细胞的主要元素,而且其N/C和N/P的比值也相对较为稳定,因此,用N描述初级生产者的生长比用其它元素(如C、P)更为精确。此外,N常常是海洋环境中的限制性营养元素,因而建立N源基础上的生产力研究更具有实际意义。根据以上观点,他们提出:在真光层中再循环的N为再生N(主要是NH+4—N),由真光层之外提供的N为新生N(主要是N-3—N),由再生N源支持的那部分初级生产力称为再生生产力(regeneration production),由新生N源支持的那部分初级生产力称为新生产力(new production)。显然新生产力和再生生产力之和就是总初级生产力。表达新生产力的单位与初级生产力相同。")
91
Eppley和Peterson(1979)明确指出,新生N来自:①上升流或梯度扩散,②陆源供应(如径流),③大气沉降或降水,④N2固定。再生N来自真光层中生物的代谢产物(如氨态N、尿素N和氨基酸N等)。同时他们利用15N的实验数据(新生N与再生N)和以往14C测得的初级生产力数据联系起来,提出“f”比的概念( 即新生产力与总生产力的比率,f = Pn / PG×100%)就可能依据这种定量关系对全球新生产力水平做出大致的估计。他们还将新生产力与向底部的碳输出,既输出生产力(export production)联系起来,赋予新生产力以生物地球化学方面的含义。此外,提出颗粒态营养元素下沉出真光层之前的循环次数(r)与f比的关系,即r=(1—f)/f。
明确指出,新生N来自:①上升流或梯度扩散,②陆源供应(如径流),③大气沉降或降水,④N2固定。再生N来自真光层中生物的代谢产物(如氨态N、尿素N和氨基酸N等)。同时他们利用15N的实验数据(新生N与再生N)和以往14C测得的初级生产力数据联系起来,提出 f 比的概念( 即新生产力与总生产力的比率,f = Pn / PG×100%)就可能依据这种定量关系对全球新生产力水平做出大致的估计。他们还将新生产力与向底部的碳输出,既输出生产力(export production)联系起来,赋予新生产力以生物地球化学方面的含义。此外,提出颗粒态营养元素下沉出真光层之前的循环次数(r)与f比的关系,即r=(1—f)/f。")
92
意义-1 (1)新生产力研究有助于从更深层次阐明海洋生态系统的结构、功能 新生产力的概念将海洋初级生产力划分为再生和新生的两部分,前者主要反映真光层营养物质循环的效率,后者反映从真光层之外的营养物质补充的比例。而不同海区的f比的差异往往与理化水文环境、初级生产者、消费者的组成(特别是粒径组成)、食物网的结构等特征以及能量的转移效率等特点有关,也就是说,新生产力与再生生产力的比例不同是各个生态系统的结构及其运转过程特征的表现。因此,新生产力的研究促使对海洋水层生态系统(pelagic ecosystem)物质转移、能量传递、营养元素再循环等的理论研究进入一个更深的层次。
新生产力研究有助于从更深层次阐明海洋生态系统的结构、功能 新生产力的概念将海洋初级生产力划分为再生和新生的两部分,前者主要反映真光层营养物质循环的效率,后者反映从真光层之外的营养物质补充的比例。而不同海区的f比的差异往往与理化水文环境、初级生产者、消费者的组成(特别是粒径组成)、食物网的结构等特征以及能量的转移效率等特点有关,也就是说,新生产力与再生生产力的比例不同是各个生态系统的结构及其运转过程特征的表现。因此,新生产力的研究促使对海洋水层生态系统(pelagic ecosystem)物质转移、能量传递、营养元素再循环等的理论研究进入一个更深的层次。")
93
意义-2 (2)新生产力的研究对阐明全球碳循环过程有重要意义 人类活动(主要是矿物燃料的燃烧造成大量CO2进入大气,所谓“温室效应”就是大气中CO2等温室气体含量增大导致全球气温上升的现象(大气中的水气量、云量以及颗粒物的作用与CO2相反)。海洋是地球最大的碳库之一,海洋中碳的生物地球化学过程在全球碳循环起重要作用。新生产力的水平是反映海洋真光层吸收CO2的能力,气—海界面碳的净通量在很大程度上就是由新生产力决定的。因此,研究新生产力的规模、时空变化和制约机智,就可能对全球变化中海洋的调节能力做出估计和预测。
新生产力的研究对阐明全球碳循环过程有重要意义 人类活动(主要是矿物燃料的燃烧造成大量CO2进入大气,所谓 温室效应 就是大气中CO2等温室气体含量增大导致全球气温上升的现象(大气中的水气量、云量以及颗粒物的作用与CO2相反)。海洋是地球最大的碳库之一,海洋中碳的生物地球化学过程在全球碳循环起重要作用。新生产力的水平是反映海洋真光层吸收CO2的能力,气—海界面碳的净通量在很大程度上就是由新生产力决定的。因此,研究新生产力的规模、时空变化和制约机智,就可能对全球变化中海洋的调节能力做出估计和预测。")
94
评述 目前有关海洋新生产力的现场调查还很少,现有的一些估计值主要时根据原有的初级生产力资料结合f比值间接计算出来的。根据Eppley和Peterson(1979)估计,全球海洋年初级生产力为1.90~24.0Gt,平均f比为0.18~0.20,因而年新生产力约为3.4~4.7Gt。Chavez和Barger(1987)、Berger等(1989)估计,海洋年总初级生产力约为30.0Gt,其中大约20%(6Gt)时新生产力。由此可见,他们估计值的差别主要是对总初级生产力的估计不同,而二者提出f比却相差不大。另据Martin(1987)的估计,海洋总初级生产力可达51.0Gt,f比约为0.145,因而年新生产力达7.40Gt(表8-8)。
估计,全球海洋年初级生产力为1.90~24.0Gt,平均f比为0.18~0.20,因而年新生产力约为3.4~4.7Gt。Chavez和Barger(1987)、Berger等(1989)估计,海洋年总初级生产力约为30.0Gt,其中大约20%(6Gt)时新生产力。由此可见,他们估计值的差别主要是对总初级生产力的估计不同,而二者提出f比却相差不大。另据Martin(1987)的估计,海洋总初级生产力可达51.0Gt,f比约为0.145,因而年新生产力达7.40Gt(表8-8)。")
95
(四)现存量 现有的海洋浮游植物现存量资料以细胞数表示的多。在富营养型海域浮游植物现存量一般大于106/L。而贫营养型海域每升水仅几百个甚至几十个细胞。在大多数情形下海洋浮游植物密度在每升几千到几万个之间。 生物量资料较少,大致说来贫营养型海域低于1mg/m3,富营养型海域高于1mg/m3,最高可达10g/m3以上。 贫营养型海域浮游植物密度和生物量的年变幅较小,最大密度和生物量和最小密度或生物量一般为2到十几倍,而在富营养型海域这个比值达到几百到几千倍以上。

96
第四节 其他生物的初级生产力 内陆水体的生产者除浮游植物外,还包括水生维管束植物、底生藻类、光合细菌、化合细菌等,后两类的生产力目前还没有足够的了解,

97
一、水 草 水草生产量的测定首先要测定生物量,然后以一定时间内两次生物量之差或以平均生物量乘以P/B系数(一般1.2~1.3)来计算生产量。也可采用测氧法求同化指数,再以同化指数乘生物量即为生产量。 水草生物量可用湿重(风干后)、干重、无灰分干重、碳值、叶绿素(chl)或叶绿素a(chla)来表示,各单位关系可按下式粗略换算(IBP,1980): 1 g干重≈0.85 g(无灰分干重或有机质干重)≈0.4 gC ≈5~15 mg(chl)≈3.75~11.25 mg(chla) 1 g水草C≈ mg(chla),简化≈10~25 mg(chla) 1 g藻类C≈2~43 mg(chla) 1 g无灰分干重C≈19 kJ
、干重、无灰分干重、碳值、叶绿素(chl)或叶绿素a(chla)来表示,各单位关系可按下式粗略换算(IBP,1980): 1 g干重≈0.85 g(无灰分干重或有机质干重)≈0.4 gC. ≈5~15 mg(chl)≈3.75~11.25 mg(chla) 1 g水草C≈ mg(chla),简化≈10~25 mg(chla) 1 g藻类C≈2~43 mg(chla) 1 g无灰分干重C≈19 kJ.")
98
(一)现存量 据IBP(1980)材料,湖泊水草生物量按有机质干重计为2.4~130 g/m2,水草区为27~360(880) g/m2,按叶绿素量计约为200~5000 mg/m2。湖库水草叶绿素a量的实测值为100~3000 mg/m2,远高于浮游植物的现存量(以叶绿素a计)34~1800 mg/m2。
材料,湖泊水草生物量按有机质干重计为2.4~130 g/m2,水草区为27~360(880) g/m2,按叶绿素量计约为200~5000 mg/m2。湖库水草叶绿素a量的实测值为100~3000 mg/m2,远高于浮游植物的现存量(以叶绿素a计)34~1800 mg/m2。")
99
中国湖库水草现存量 中国湖库水草现存量全湖均值为38.4~13630 g/ m2湿重,若水分按8.85%计相当于3.4~1206 g/ m2干重或2.89~1025 g/ m2有机质干重,高于IBP资料。长江以北平均4871 g/ m2湿重,长江以南仅3166 g/ m2湿重,这点和南方水草经常被采捞利用有关。 按单位面积计算,挺水植物生物量远较沉水植物为高。按地上植株有机质干重计算,挺水植物生物量在温带湖泊多在1.5~3.5 kg/ m2之,热带湖通常超过2.0 kg/ m2。最大值可达3~9 kg/ m2。沉水植物温带肥水区多在0.5 kg/ m2有机质干重左右,最大值约0.6~0.7 g/ m2有机质干重。

100
水草生物量的空间差异 水草生物量的水平差异极为显著,同一水体极值可达0~1500 g/ m2有机质干重。最高值一般出现于有厚的淤泥沉淀和避风的浅水处,但不在易受波浪冲击和水位变化影响的最浅水区。如鄱阳湖水草生物量(湿重)湖均1542 g/ m2,水草区1939 g/ m2,有8处高于2000 g/ m2的均位于淤泥很厚的湖区接近水面浅处。 水草垂直分布下限约为10 m深处,但轮藻和水藓可分布到30 m深处,通常从0.3 m往下生物量渐减。
湖均1542 g/ m2,水草区1939 g/ m2,有8处高于2000 g/ m2的均位于淤泥很厚的湖区接近水面浅处。 水草垂直分布下限约为10 m深处,但轮藻和水藓可分布到30 m深处,通常从0.3 m往下生物量渐减。")
101
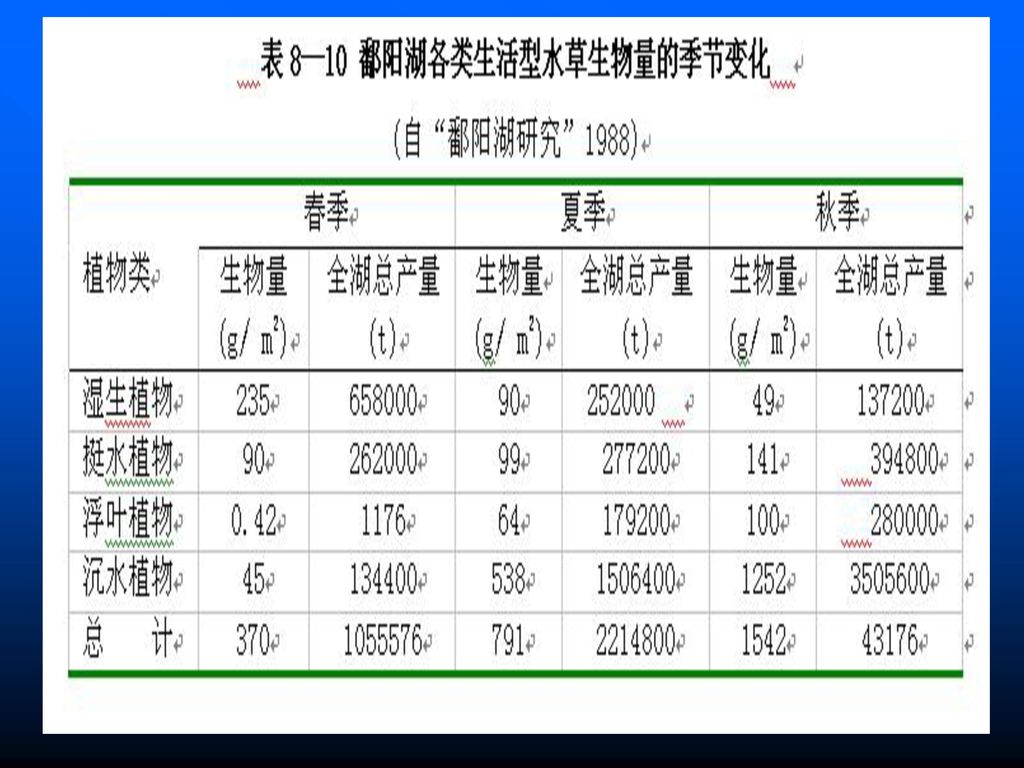
水草生物量的季节分布 从季节分布看,生物量一般都是从春到秋增高,冬季最低,如鄱阳湖除湿生植物受水位影响以春季最高,夏季次之,秋季最低外,挺水植物、浮叶植物和沉水植物都是从春到秋递增(表8—10)。有些湖泊水草因被草鱼摄食,秋季低于春夏。
。有些湖泊水草因被草鱼摄食,秋季低于春夏。")
103
(二)光合能力 水草在最适光照下的净同化指数通常为6~40 mgO2/g干重,平均为10 mgO2/g,粗略换算尚不到1 mgC/mgchla·h,最大值挺水植物可达1~2.5,沉水植物达0.9 mgC/mgchla·h,与浮游植物平均值(4~5 mgC/mgchla·h)相比较低得多,这是因为水草主要由叶部分进行光合作用,而藻类整个细胞都可进行。 水草光的饱和值较低,一般为40~80 J/m2·s,约相当于10 000~ lx,因种类和适应情况而不同,如果逐渐增加光照,饱和点可达到160 J/m2·s。喜阴种的饱和点很低,如日本萍蓬草(Nuphar japonicum)沉水叶在5000 lx(约为20 J/m2·s)时即受抑制,但浮水叶未受影响。喜阴种的叶宽而薄,叶绿素a含量多,呼吸率和光合能力都较低,喜光种则相反。
相比较低得多,这是因为水草主要由叶部分进行光合作用,而藻类整个细胞都可进行。 水草光的饱和值较低,一般为40~80 J/m2·s,约相当于10 000~ lx,因种类和适应情况而不同,如果逐渐增加光照,饱和点可达到160 J/m2·s。喜阴种的饱和点很低,如日本萍蓬草(Nuphar japonicum)沉水叶在5000 lx(约为20 J/m2·s)时即受抑制,但浮水叶未受影响。喜阴种的叶宽而薄,叶绿素a含量多,呼吸率和光合能力都较低,喜光种则相反。")
104
(三)生产量和呼吸量 毛产量的日变化主要由太阳辐射的变化引起的,但在不同地点或在一段较长时期内,生物量、透明度和光合能力的不同也可能起重要作用。温带水草的光合能力通常冬季低,春季增高,夏末或秋季达到最高点或有夏秋2个最高点。 在肥水中当水草生长迅速期,草丛区的净产量(有机质干重)通常在2~10 g/m2·d之间,温带湖年产量多是按生物量最高季节计算的,常在20~1000 g/ m2 (有机质干重)。在热带湖泊生产中也有季节变化,生产量(有机质干重)可能超过2000 g/ m2。
通常在2~10 g/m2·d之间,温带湖年产量多是按生物量最高季节计算的,常在20~1000 g/ m2 (有机质干重)。在热带湖泊生产中也有季节变化,生产量(有机质干重)可能超过2000 g/ m2。")
105
在大型植物中挺水植物提供的产量最高,在温带和热带的水体中,其净产量一般可达7~11 t/hm2·a(有机质干重后同)。马来西亚的一种蒲草(dw)毛产量达25 g/ m2·d,如呼吸所消耗的按25%计算,全年生长期按300 d计,则年净产量达到60 t/hm2。 沉水植物产量较低的原因可能是二氧化碳供应不足,因为二氧化碳在水中的扩散率只有大气中的1%。 水草(dw)的呼吸强度平均约为1 mgO2/g·h,在低温、低氧量和无湍流情况下较低,呼吸量和光合能力之比常为0.1~0.15,但极值达0.001~0.4。 在晴天白天水草呼吸量(R)约为毛产量(P)的0.3~0.5,24 h的R/P值晴天为0.35,雨天达0.9,月均值在0.5~0.7之间(Lkusima,1965),但Westlake(1966)指出水草24 hR/P值变幅很大,晴天可能不到0.1,阴天在浑浊度很大的水中高达5以上,正常情况呼吸量约为7 gO2/ m2·d。
的呼吸强度平均约为1 mgO2/g·h,在低温、低氧量和无湍流情况下较低,呼吸量和光合能力之比常为0.1~0.15,但极值达0.001~0.4。 在晴天白天水草呼吸量(R)约为毛产量(P)的0.3~0.5,24 h的R/P值晴天为0.35,雨天达0.9,月均值在0.5~0.7之间(Lkusima,1965),但Westlake(1966)指出水草24 hR/P值变幅很大,晴天可能不到0.1,阴天在浑浊度很大的水中高达5以上,正常情况呼吸量约为7 gO2/ m2·d。")
106
二、底生或周生藻类 淡水底生藻类群落有3种类型(IBP,1980):
1. 坚实地附着在各种基质上,形成稠密的毛毡状物,其中混杂很多死细胞和老细胞,生物量(chla)达到189~1960 mg/m2。 2. 丝状藻类集合成簇状体或束状体,有时还有胶状物使连结成团,生物量(chla)为12~2350 mg/m2。 3. 在底质上可活动的藻类,因易被水流冲走,生物量不高,仅为2.6~124.8 mg/m2 (chla)。 1类和2类的生物量主要受光的限制,通常集中分布于表层一薄层中,密度极大,即使生物量(chla)达到200 mg/m2,厚度也不到5mm。3类的生物量常随养分多寡而变化。
达到189~1960 mg/m2。 2. 丝状藻类集合成簇状体或束状体,有时还有胶状物使连结成团,生物量(chla)为12~2350 mg/m2。 3. 在底质上可活动的藻类,因易被水流冲走,生物量不高,仅为2.6~124.8 mg/m2 (chla)。 1类和2类的生物量主要受光的限制,通常集中分布于表层一薄层中,密度极大,即使生物量(chla)达到200 mg/m2,厚度也不到5mm。3类的生物量常随养分多寡而变化。")
107
底生藻类生物量 周生藻类常和其他附着生物一起,在河流水流缓慢处或湖泊的沿岸,有时几乎完全掩盖基质。在Ontario湖岸,一个350 m宽地带,66%为刚毛藻所复盖,生物量达15700 kg/km2,其中一个277 m宽带,79%被复盖,生物量达26000 kg/km2。 在温带湖泊生物量高峰出现于春季或夏初,中夏以后因受雨水冲刷,浮游植物吸收养分和遮光以及动物的摄食等影响,生物量下降。底生藻类生物量的年变幅较浮游植物和水草小,年最大值和最小值的比值浮游植物为1.5~2.5到83倍,水草为1.5到190倍,底生藻类仅4.5~18倍。

108
周生藻类的作用 周生藻类产量在淡水水体的初级产量中有时占很大比重,通常在流水水体中所起作用大于静水水体。在有些河流中周生藻类可能利用外来有机质进行异养生活。 在沿岸带面积很小的深水湖泊,周生藻类生产力比重所占很小。光线可透射到水底的浅水区越大,水草越丰富,周生藻类分布区越广,在初级生产力中起的作用也越大。据Wetzel(1975)统计,周生藻类在静水水体的初级生产力所占比重可以从1%(无水草的贫营养型湖)到62%(水交换量很大的浅水湖,浮游植物被大量排出)。
统计,周生藻类在静水水体的初级生产力所占比重可以从1%(无水草的贫营养型湖)到62%(水交换量很大的浅水湖,浮游植物被大量排出)。")
109
在水流湍急的河流,底生藻类几乎是唯一的生产者,但在水流缓慢的深水河流,浮游植物产量仍高于周生藻类。当河水十分浑浊时,浮游植物也受限制,某些周生藻类可能通过异养性营养而生活。在一个河流上游盛产鲑鱼的急流区,周生藻类生产力曾达到1050 mgC/m2·d,远超过富营养型湖的水平(McConnell等,1959)。 不同水体和同一水体的不同时期,底生藻类的毛产量都差别很大。由于很难排除沉淀中有机质分解的影响,底生藻类群落的呼吸量尚很少研究。Hargrave(1969)用加抗菌素的方法,发现群落年呼吸量中细菌占30%~45%,藻类呼吸在6月份仅占总量20%~25%。底生藻的生物量和生产力均与水流速度有密切关系,流速大,藻类代谢率升高,生产量和生物量周转率都加大,但流速过大藻类易被冲走,生物量降低,流速约50 cm/s时,冲走量就大于生物量的积累(Horner,1978)。试验表明流速还与种类组成有关,38 cm/s时硅藻占优势,9 cm/s时鞘藻、毛枝藻、黄藻等占优势。
用加抗菌素的方法,发现群落年呼吸量中细菌占30%~45%,藻类呼吸在6月份仅占总量20%~25%。底生藻的生物量和生产力均与水流速度有密切关系,流速大,藻类代谢率升高,生产量和生物量周转率都加大,但流速过大藻类易被冲走,生物量降低,流速约50 cm/s时,冲走量就大于生物量的积累(Horner,1978)。试验表明流速还与种类组成有关,38 cm/s时硅藻占优势,9 cm/s时鞘藻、毛枝藻、黄藻等占优势。")
110
三、各类生产力间的关系 浮游植物和水草通过光和养分的竞争以及释放化学物质的相生相克关系,对彼此的生产力都有明显的影响。浮游植物水华降低透明度抑制水草的生长,而水草丛生处光线受阻,浮游植物也难增长。水草主要从底层中吸收养分,因此与浮游植物在养分上的直接矛盾并不尖锐。对水葫芦(Eichhornia crassipes)的试验也表明,在短时间内营养的竞争不是藻类受水草抑制的原因。大木规 晃(1984)、俞子文等(1992)的试验都表明,漂浮植物对藻类有不同程度的抑制效应。俞子文等指出分泌物的相生相克关系有两种含意: 1)不同植物种产生的分泌物对同一受体植物产生不同的影响,如水葫芦、水花生(Alternanthera philoxeroides)、水浮莲(Pissia stratiotes)、满江红(Azolla imbricata)、紫萍 (Spirodela polyrhiza)、浮萍(Lemna minor)的分泌物对衣藻产生抑制作用,而西洋菜(Nasturfium officinale)对衣藻没有抑制作用,甚至稍有促进作用。2)浓度效应,水葫芦、水花生、水浮莲等分泌物在低浓度下表现促进作用,而当浓度达到一定阈值后产生克制作用。
的试验也表明,在短时间内营养的竞争不是藻类受水草抑制的原因。大木规 晃(1984)、俞子文等(1992)的试验都表明,漂浮植物对藻类有不同程度的抑制效应。俞子文等指出分泌物的相生相克关系有两种含意: 1)不同植物种产生的分泌物对同一受体植物产生不同的影响,如水葫芦、水花生(Alternanthera philoxeroides)、水浮莲(Pissia stratiotes)、满江红(Azolla imbricata)、紫萍 (Spirodela polyrhiza)、浮萍(Lemna minor)的分泌物对衣藻产生抑制作用,而西洋菜(Nasturfium officinale)对衣藻没有抑制作用,甚至稍有促进作用。2)浓度效应,水葫芦、水花生、水浮莲等分泌物在低浓度下表现促进作用,而当浓度达到一定阈值后产生克制作用。")
111
在水草丛生区浮游植物量和生理活性都降低,生产力也激降。
野外观测也发现,在水草丛生区浮游植物量和生理活性都降低,生产力也激降。据捷克一个浅水湖的试验,把湖心开旷区的浮游植物移到沿岸沉水植物带,初级产量降低27%~56%,从沿岸移到开旷区则生产力增高230%~500%(Brandl等,1970);反之,在浮游植物丰富水区,水草很难丛生。戎克文等(1995)研究武汉东湖三种不同类型湖区浮游植物初级生产力,发现在藻型湖区年产量(dw)最高,达 g/m2·a,草型湖区最低,仅228.6 g/ m2·a(dw),草一藻过渡型湖区居中,为572.1 g/ m2·a(dw),显示水草对浮游植物生产力的巨大影响。根据1963~1990年草型湖区浮游植物日毛产量和水草生物量的关系,得出两者呈显著负相关。水草生物量(湿重)每增加1 g/m2,浮游植物日产量将降低 gO2/m2·d。
;反之,在浮游植物丰富水区,水草很难丛生。戎克文等(1995)研究武汉东湖三种不同类型湖区浮游植物初级生产力,发现在藻型湖区年产量(dw)最高,达 g/m2·a,草型湖区最低,仅228.6 g/ m2·a(dw),草一藻过渡型湖区居中,为572.1 g/ m2·a(dw),显示水草对浮游植物生产力的巨大影响。根据1963~1990年草型湖区浮游植物日毛产量和水草生物量的关系,得出两者呈显著负相关。水草生物量(湿重)每增加1 g/m2,浮游植物日产量将降低 gO2/m2·d。")
112
特殊情况 水草丛生也可能促进浮游植物生产力的提高,如太湖3个湖区的浮游植物毛产量和年P/B值均随水草的增多而增高(表8—11)。这是因为,太湖属高度富营养化的浅水湖,营养盐类有剩余,但风大水浅,在大风的搅动下泥砂含量高,低的透明度限制了浮游植物的光合强度。在沉水植物丛生区风浪和水流受阻,水的透明度升高,因而提高了初级产量。在内蒙乌梁素海和辽宁团结水库也发现类似情况。
。这是因为,太湖属高度富营养化的浅水湖,营养盐类有剩余,但风大水浅,在大风的搅动下泥砂含量高,低的透明度限制了浮游植物的光合强度。在沉水植物丛生区风浪和水流受阻,水的透明度升高,因而提高了初级产量。在内蒙乌梁素海和辽宁团结水库也发现类似情况。")
113
底生藻类和浮游植物 底生藻类和浮游植物在光和养分方面的竞争也极为明显,养鱼池早春清塘注水施肥后,如果水绵之类底生藻类先繁殖起来,浮游植物由于养分被吸收而增长极慢,反之当浮游植物已经大量出现,导致透明度降低,底生藻类也难于孳生。 一向认为附生藻类和水草之间是偏利关系:藻类附着在水草茎叶上生活,使本身处在光照和温度条件较好的环境中,并且还能从水草腐朽的组织的淋滤中得到养分,而水草并未受到不良的影响,然而现在已觉察到,情况要复杂些。水草的分泌物可能对附生藻类产生不良影响,藻类的分泌物也可能克制水草的生长。藻类大量附着在沉水植物体和浮叶植物根系上,可能对水草生长不利。但总的现象是:不同水体和同一水体中,底生藻类的生产力通常随沉水植物的生物量而升高。

114
四、生物圈的初级生产力和光能利用效率 养生物通过光合和化合作用合成的有机质,是人类赖以生存的食物和其他原料的基础,生物圈初级生产力的大小规定了地球能养活多少人口的限度,水圈初级生产力的大小则规定了海洋捕鱼业发展的限度。 据Whittaker和Likens估算,全地球包括海陆在内所有生态系统,每年初级净产量约1640×108t有机质干重,其中1/3在海洋,2/3在陆地(包括内陆水体),从海洋面积接近陆地2.5倍来看,应当说海洋初级生产力是比较低的。 从表8—12可见,年均初级净产量陆地几乎为海洋的5倍,虽然海藻床和珊瑚礁、河口湾的年均净产量(dw)达到1500~2500 g/m2,和陆地的森林、泡沼相近,上升流和大陆架也可与湖泊和河流相比,但占海洋总面积90%以上的大洋生产力极低,只相当于陆地的荒原、高山、荒漠等的水平。若按生物量计,海洋更低得多,不及陆地的千分之一,这是因为海洋的生产者几乎全是微型藻类,而陆地则以大型植物为主。
,从海洋面积接近陆地2.5倍来看,应当说海洋初级生产力是比较低的。 从表8—12可见,年均初级净产量陆地几乎为海洋的5倍,虽然海藻床和珊瑚礁、河口湾的年均净产量(dw)达到1500~2500 g/m2,和陆地的森林、泡沼相近,上升流和大陆架也可与湖泊和河流相比,但占海洋总面积90%以上的大洋生产力极低,只相当于陆地的荒原、高山、荒漠等的水平。若按生物量计,海洋更低得多,不及陆地的千分之一,这是因为海洋的生产者几乎全是微型藻类,而陆地则以大型植物为主。")
115
光合利用率 通常以单位地面(或水面)植物光合作用所积累的能量(初级产量)和同一时间所接受的有效辐射能(约相当于可见光部分,大致为太阳总辐射的50%)的百分比来表示光能利用率。 从理论上说,8~10个量子可以使1个水分子的水分解并和一个分子的二氧化碳合成碳水化合物。按此计算,以毛产量估计的光能利用率可达到10%以上。但实际上远低于此值。

116
地球和森林的光能利用率 计算(Stern,1975)全球地表每年进入的有效辐射能约为100×1022 J,自养生物年总产量约为100×109 t碳,相当于170×1019 J,因此全球的光能利用率平均仅0.2%,在生长最快季节可达3%~4%。 在陆生态系中,温带森林净产量(dw)(不计地下根系部分)约5~10 t/hm2·a到20 t/hm2·a,热带森林净产量(dw)可达30 t/hm2·a以上,毛产量(dw)达40 t/hm2·a。按此计算净产量的光能效率<1%~1.5%,毛产量(dw)也不过2.0%~3.5%。草本植物净产量(dw)约4~13 t/hm2·a,光能效率约0.5%~1%,但在最适时期和最适条件下短期计算可达8%~10%。
(不计地下根系部分)约5~10 t/hm2·a到20 t/hm2·a,热带森林净产量(dw)可达30 t/hm2·a以上,毛产量(dw)达40 t/hm2·a。按此计算净产量的光能效率<1%~1.5%,毛产量(dw)也不过2.0%~3.5%。草本植物净产量(dw)约4~13 t/hm2·a,光能效率约0.5%~1%,但在最适时期和最适条件下短期计算可达8%~10%。")
117
海藻、水草和浮游植物的效率 海带、巨藻等大型海藻净产量(dw)达25~50 t/hm2·a,光能效率达4%~6%。
淡水挺水植物净产量(dw)约7~11 t/hm2·a,沉水植物仅0.8~2.0 t/hm2·a,马来西亚一种蒲草的生物量(dw)达370~520 t/hm2·a,毛产量(dw)达25 g/hm2·a以上,如呼吸消耗按25%,生长期按300 d计算,则净产量(dw)为60 t/hm2·a,光能效率达4%~6% 。 海洋浮游植物光能效率不过0.16%~0.20%,淡水浮游植物最高产量达10 gC/m2·d(Talling,1975),但在印度一个蓝藻水华池曾报导13~24 gC/m2·d的高产量,约相当于40~70 t/hm2·a(dw),光能效率达到3.7%~7%。 淡水微藻在大量培养条件下,据捷克报导栅藻的最高产量(dw)达44 g/m2·d,5~7月间平均日产量(dw)为22.8 g/m2,光能效率平均4.8%,最高值达10%~11%。
约7~11 t/hm2·a,沉水植物仅0.8~2.0 t/hm2·a,马来西亚一种蒲草的生物量(dw)达370~520 t/hm2·a,毛产量(dw)达25 g/hm2·a以上,如呼吸消耗按25%,生长期按300 d计算,则净产量(dw)为60 t/hm2·a,光能效率达4%~6% 。 海洋浮游植物光能效率不过0.16%~0.20%,淡水浮游植物最高产量达10 gC/m2·d(Talling,1975),但在印度一个蓝藻水华池曾报导13~24 gC/m2·d的高产量,约相当于40~70 t/hm2·a(dw),光能效率达到3.7%~7%。 淡水微藻在大量培养条件下,据捷克报导栅藻的最高产量(dw)达44 g/m2·d,5~7月间平均日产量(dw)为22.8 g/m2,光能效率平均4.8%,最高值达10%~11%。")
118
光能低于理论值的原因 由此可见,除人工培养的特殊条件下,各类生态系统光能利用效率都远低于理论值,主要原因包括:(1)射到植物体上的光能一部分被反射,一部分透过植物体,仅部分被吸收;(2)有时光照过强,起了抑制作用;(3)二氧化碳供应不足,特别是在水生态系中;(4)养分的限制;(5)环境压力;(6)植物体老化。
射到植物体上的光能一部分被反射,一部分透过植物体,仅部分被吸收;(2)有时光照过强,起了抑制作用;(3)二氧化碳供应不足,特别是在水生态系中;(4)养分的限制;(5)环境压力;(6)植物体老化。")
119
复习思考题 1. 弄清现存量或生物量、生产量、收获量、周转率、周转时间、生物生产力、初级生产力、次级生产力、初级净产量、初级毛产量、群落或生态系净产量等概念的含意和彼此间的关系。 2. 初级生产力的测定方法有哪些?各有何优缺点?适宜在什么条件下采用? 3. 决定初级生产力的因素有那些?作用如何? 4. 湖泊和水库浮游植物初级生产力的分布规律如何?中国有何特点? 5. 养鱼池初级生产力主要受那些因素的影响? 6. 何谓胞外产物?一般占多大比例? 7. P/R值指什么?有何生态意义? 8. 生物量和生产量之间的关系如何? P/B值与那些因素有关? 9. 淡水浮游植物量可分为那些等级?各级水的渔业意义如何? 10. 浮游植物的生物量和生产力的时空分布有何趋势? 11. 水草和底生藻类的现存量和生产力分布有何特点? 12.浮游植物、水草和底生藻类在生产力上相互关系如何? 13. 生物圈中各类生物群落对光能的利用效率达到什么程度?为什么远低于理论值?

Similar presentations






的過濾作用 腎小管的再吸收作用 腎小管的分泌作用 尿液的組成>")








>")




