Download presentation
Presentation is loading. Please wait.

1
8 基因的表达与调控 ——真核基因表达调控 真核生物(除酵母、藻类和原生动物等单细胞类之外)主要由多细胞组成,每个细胞基因组中蕴藏的遗传信息量及基因数量都大大高于原核生物。人类细胞单倍体基因组有3×109bp,为大肠杆菌总DNA的800倍,噬菌体的10万倍左右!
主要由多细胞组成,每个细胞基因组中蕴藏的遗传信息量及基因数量都大大高于原核生物。人类细胞单倍体基因组有3×109bp,为大肠杆菌总DNA的800倍,噬菌体的10万倍左右! .")
2
真核生物染色质被包裹在细胞核内,基因的转录(核内)和翻译(细胞质内)被核膜所隔开,核内RNA的合成与转运,细胞质中RNA的剪接和加工等都属于真核生物基因调控的范围。
和翻译(细胞质内)被核膜所隔开,核内RNA的合成与转运,细胞质中RNA的剪接和加工等都属于真核生物基因调控的范围。")
3
真核生物基因调控主要包括: (1)瞬时调控或称可逆性调控:相当于原核细胞对环境条件变化作出的反应,包括激素水平的改变、酶活性的调节。
(2)发育调控或称不可逆调控:发育调控是真核基因调控的精髓部分,它决定了真核细胞生长、分化、发育的全部进程。
发育调控或称不可逆调控:发育调控是真核基因调控的精髓部分,它决定了真核细胞生长、分化、发育的全部进程。")
4
根据基因调控在同一事件中发生的先后次序,又可将其分为
转录水平调控、转录后水平调控 转录后水平调控主要包括: RNA加工成熟过程的调控; 翻译水平调控; 蛋白质加工水平的调控。

5
图8-1 真核基因表达调控的主要步骤示意图

6
研究基因调控主要应回答3个问题 1.什么是诱发基因转录的信号? 2.基因调控的关键在什么水平上? 3.不同水平基因调控的分子机是什么?

7
8.1 真核生物的基因结构与转录活性 1.在真核细胞中,一条成熟的mRNA链只能翻译出一条多肽链,很少原核生物中常见的多基因操纵子形式。
2.真核细胞DNA与组蛋白和大量非组蛋白相结合,只有一小部分DNA是裸露的。 3.高等真核细胞DNA中很大部分是不转录的,由几个或几十个碱基组成的DNA序列,在整个基因组中重复几百次甚至上万次。

8
4.真核生物能够有序地根据生长发育阶段的需要进行DNA片段重排,还能增加细胞内某些基因的拷贝数。
5.基因转录的调节区很大,可能远离转录起始位点达几百个甚至上千个碱基对,主要通过改变整个所控制基因5'上游区DNA构型来影响它与RNA聚合酶的结合能力。

9
6.真核生物的RNA在细胞核中合成,只有经转运穿过核膜,到达细胞质后,才能被翻译成蛋白质。原核生物中不存在这样严格的空间间隔。
7.许多真核生物的基因只有经过复杂的成熟和剪接过程,才能被顺利地翻译成蛋白质。

10
8.1.1 基因家族(gene family) 真核细胞中许多相关的基因常按功能成套组合,被称为基因家族。同一家族中的成员有时紧密地排列在一起,成为一个基因簇,更多的时候,分散在同一染色体的不同部位,甚至位于不同的染色体上,具有各自不同的表达调控模式。
 真核细胞中许多相关的基因常按功能成套组合,被称为基因家族。同一家族中的成员有时紧密地排列在一起,成为一个基因簇,更多的时候,分散在同一染色体的不同部位,甚至位于不同的染色体上,具有各自不同的表达调控模式。")
11
1、简单多基因家族 简单多基因家族中的基因一般以串联方式前后相连。细菌中所有rRNA和部分tRNA都来自这个分子量为30S(约6500个核苷酸)的前rRNA。 在真核生物中,前rRNA转录产物的分子量为45S,(约有14 000个核苷酸),包括18S,28S和5.8S三个主要rRNA分子。5S rRNA作为一个独立的转录单位,由RNA聚合酶III(而不是聚合酶I)完成转录。
,包括18S,28S和5.8S三个主要rRNA分子。5S rRNA作为一个独立的转录单位,由RNA聚合酶III(而不是聚合酶I)完成转录。")
12
图8-2 细菌中rRNA基因家族各成员的分布与成熟过程分析
图8-2 细菌中rRNA基因家族各成员的分布与成熟过程分析

13
图8-3 脊椎动物中rRNA基因家族主要成员的分布与成熟过程分析。

14
2、复杂多基因家族 复杂多基因家族一般由几个相关基因家族构成,基因家族之间由间隔序列隔开,并作为独立的转录单位。
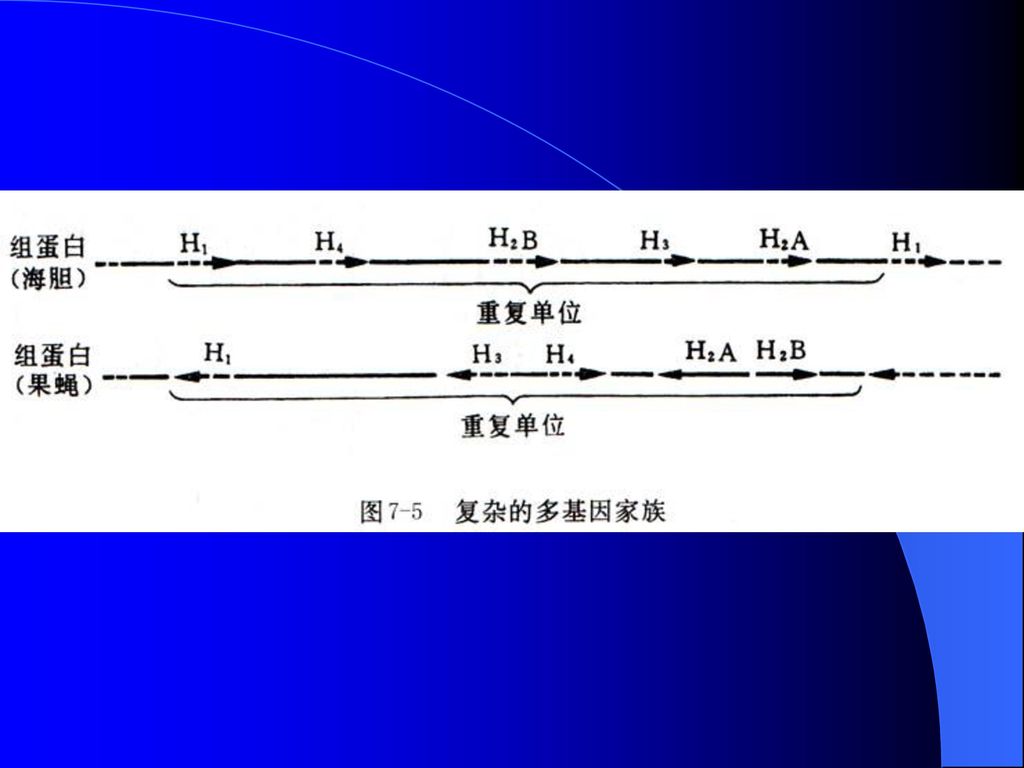
海胆组蛋白基因家族(图8-4):编码不同组蛋白的基因处于一个约为6 000bp的片段中,分别被间隔序列所隔开。这5个基因组成的串联单位在整个海胆基因组中可能重复多达1 000次。
:编码不同组蛋白的基因处于一个约为6 000bp的片段中,分别被间隔序列所隔开。这5个基因组成的串联单位在整个海胆基因组中可能重复多达1 000次。")
16
海胆串联单位中每个基因都在同一条DNA链上按同一方向单独转录,都没有内含子,基因转录与翻译速度都受到调节。H2A、H2B、H3和H4摩尔数相等,H1恰好是前者的一半。

17
3、发育调控的复杂多基因家族 血红蛋白是所有动物体内输送分子氧的主要载体,由2α2β组成的四聚体加上一个血红素辅基(结合铁原子)后形成功能性血红蛋白。在生物个体发育的不同阶段出现几种不同形式的α和β亚基。
后形成功能性血红蛋白。在生物个体发育的不同阶段出现几种不同形式的α和β亚基。")
18
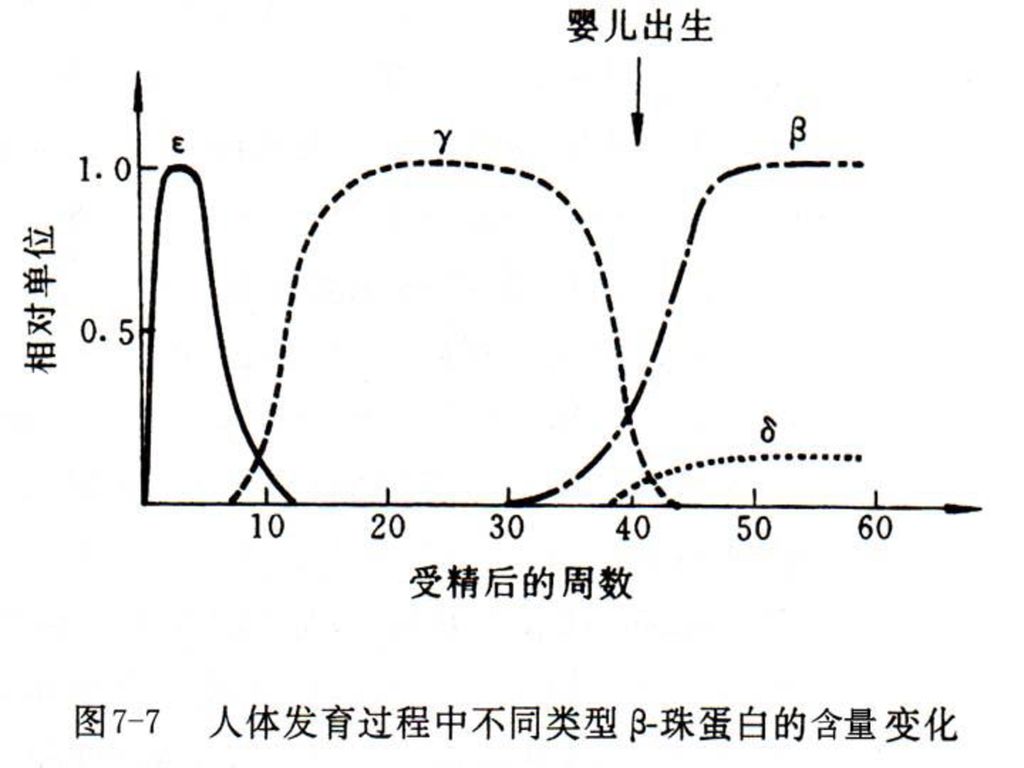
(1)人α珠蛋白基因簇位于16号染色体短臂上,约占30kb左右,其中ζ为胚胎期基因(表8-1)。
(2)β珠蛋白基因簇位于11号染色体短臂上,约占50-60kb,其中ε为胚胎期基因,Gγ和Aγ为胎儿型基因,δ和β为成人期基因。
β珠蛋白基因簇位于11号染色体短臂上,约占50-60kb,其中ε为胚胎期基因,Gγ和Aγ为胎儿型基因,δ和β为成人期基因。")
19
表8-1 不同发育阶段血红蛋白亚型 发 育 阶 段 组 成 胚胎期(8周以前) ζ2ε2、ζ2γ2和α2ε2 胎儿期(8~41周) α2γ2
表8-1 不同发育阶段血红蛋白亚型 发 育 阶 段 组 成 胚胎期(8周以前) ζ2ε2、ζ2γ2和α2ε2 胎儿期(8~41周) α2γ2 成人期 α2δ2和α2β2
 ζ2ε2、ζ2γ2和α2ε2. 胎儿期(8~41周) α2γ2. 成人期. α2δ2和α2β2.")
20
图8-5 人β-珠蛋白基因的基本结构。

22
图8-7 人细胞中α和β-珠蛋白基因簇结构示意图

23
8.1.2 真核基因的断裂结构 1.外显子与内含子 “intron” 是指存在于原始转录物或基因组DNA中,但不存在于成熟mRNA、rRNA或tRNA中的那部分核苷酸序列。大多数真核基因都是由蛋白质编码序列和非蛋白质编码序列两部分组成的。

24
基因中的内含子数量和大小都不同。胶原蛋白基因长约40kb,至少有40个内含子,其中短的只有50bp,长的可达到2000bp。

25
表8-2 不同真核生物基因的平均长度及 单个基因平均含有外显子数量比较 物 种 外显子数/基因 核基因平均长度(kb) mRNA平均长度
物 种 外显子数/基因 核基因平均长度(kb) mRNA平均长度 酵 母 1 1.6 真 菌 3 1.5 线 虫 4 4.0 3.0 果 蝇 11.3 2.7 鸡 9 13.9 2.4 哺乳类 7 16.6 2.2
 mRNA平均长度. 酵 母 真 菌 线 虫 果 蝇 鸡 哺乳类")
26
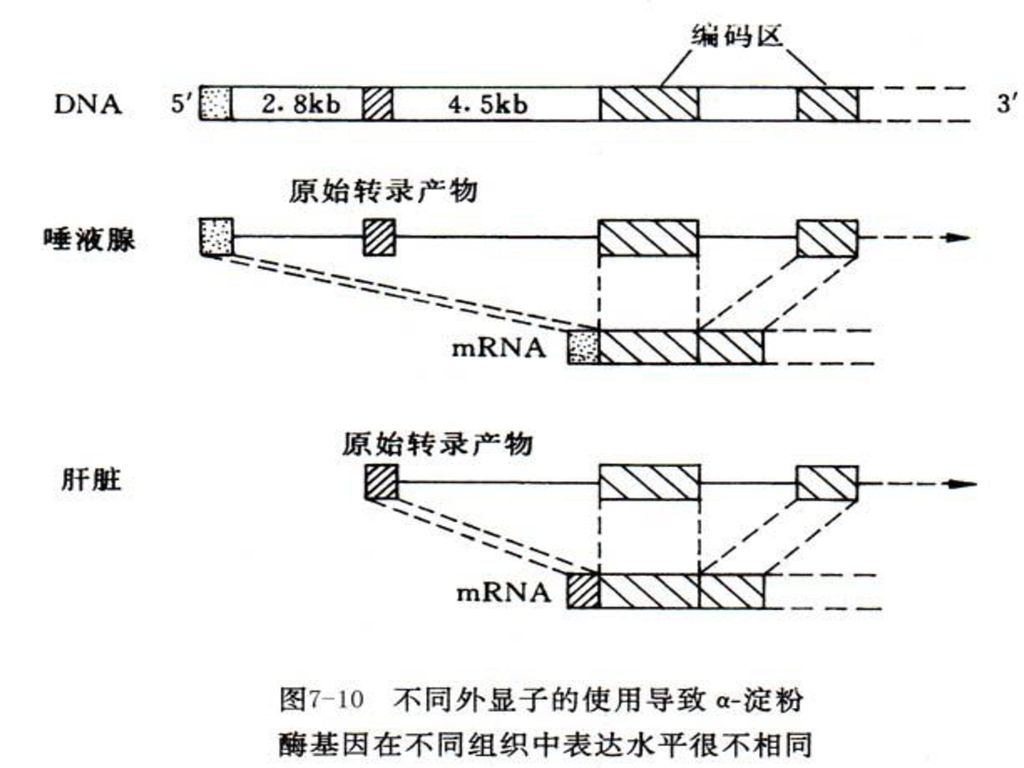
图8-8是哺乳动物二氢叶酸还原酶基因,全长25-31kb左右,但其6个外显子总长只有2kb。
少数基因,如组蛋白及α型、β型干扰素基因,根本不带内含子。

27
图8-8 哺乳动物二氢叶酸还原酶的基因结构示意图

28
2、外显子与内含子的连接区 断裂结构的一个重要特点是外显子-内含子连接区(exon—intron junction)的高度保守性和特异性碱基序列。连接区特征: (1)内含子的两端序列之间没有广泛的同源性,因此内含子两端序列不能互补,说明在剪接加工之前,内含子上游序列和下游序列不可能通过碱基配对形成发卡式二级结构。 (2)短,高度保守。
内含子的两端序列之间没有广泛的同源性,因此内含子两端序列不能互补,说明在剪接加工之前,内含子上游序列和下游序列不可能通过碱基配对形成发卡式二级结构。 (2)短,高度保守。")
29
序列分析表明,几乎每个内含子5‘ 端起始的两个碱基都是GT,而3’ 端最后两个碱基总是AG,由于这两个碱基的高度保守性和广泛性,有人把它称为GT-AG法则,即:5‘GT……AG 3’。
——P95内含子剪切部分内容。

30
3、外显子与内含子的可变调控 (1)一个基因的转录产物通过剪接只能产生一种成熟的mRNA,编码一个多肽——组成型剪接。
(2)有些基因选择了不同的启动子,或者在转录产物上选择了不同的多聚(A)位点而使原始转录物具有不同的二级结构,产生不同的mRNA分子。同一基因的转录产物由于不同的剪接方式形成不同mRNA的过程称为选择性剪接。
有些基因选择了不同的启动子,或者在转录产物上选择了不同的多聚(A)位点而使原始转录物具有不同的二级结构,产生不同的mRNA分子。同一基因的转录产物由于不同的剪接方式形成不同mRNA的过程称为选择性剪接。")
32
8.1.3 真核生物DNA水平上的基因表达调控 在个体发育过程中,模板DNA会发生规律性变化,从而控制基因表达和生物体发育。 DNA水平的调控是真核生物发育调控的一种形式,包括基因丢失、扩增、重排和移位等,通过这些方式可以消除或变换某些基因并改变它们的活性。

33
1、“开放”型活性染色质(active chromatin)结构对转录的影响
真核基因的活跃转录发生在常染色质上。转录发生前,特定的染色质区域解旋,DNA局部结构发生变化,核小体消除或呈松散状,从右旋型变为左旋型等,导致结构基因暴露,促进转录因子与启动区DNA的结合。

34
有证据表明,存在于“灯刷型”染色体(lamp brush)上的环形结构可能与基因的活性转录有关。“灯刷型” 是染色体充分伸展时的一种形态,此时,两对姐妹染色体常常通过“交叉点”(chiasmata)连成一体。 灯刷型染色体上存在许多突起的“泡”或环状结构,有时看到RNP沿突起结构移动,表明这部分DNA正被DNA聚合转录。

35
图8-10 “灯刷型”染色体(A)及存在于“活性”染色质区的DNA环状结构(B)。图A中箭头所指为不同姐妹染色体对之间的交叉点。
图8-10 “灯刷型”染色体(A)及存在于“活性”染色质区的DNA环状结构(B)。图A中箭头所指为不同姐妹染色体对之间的交叉点。
及存在于 活性 染色质区的DNA环状结构(B)。图A中箭头所指为不同姐妹染色体对之间的交叉点。")
36
2、基因扩增 是指某些基因的拷贝数专一性大量增加的现象,它使细胞在短期内产生大量的基因产物以满足生长发育的需要,是基因活性调控的一种方式。
非洲爪蟾的卵母细胞中原有rRNA基因(rDNA)约500个拷贝,在减数分裂粗线期,基因开始迅速复制,到双线期拷贝数约为200万个,扩增近4000倍,可用于合成1012个核糖体。
约500个拷贝,在减数分裂粗线期,基因开始迅速复制,到双线期拷贝数约为200万个,扩增近4000倍,可用于合成1012个核糖体。")
37
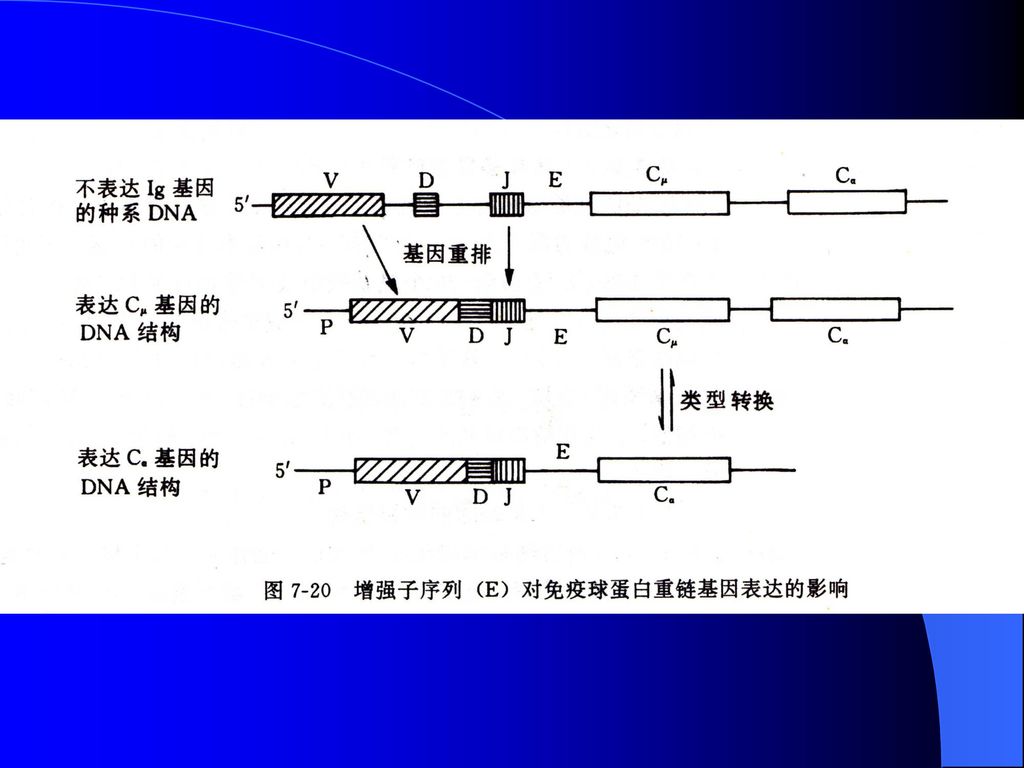
3、基因重排与变换 将一个基因从远离启动子的地方移到较近的位点从而启动转录,被称为基因重排。
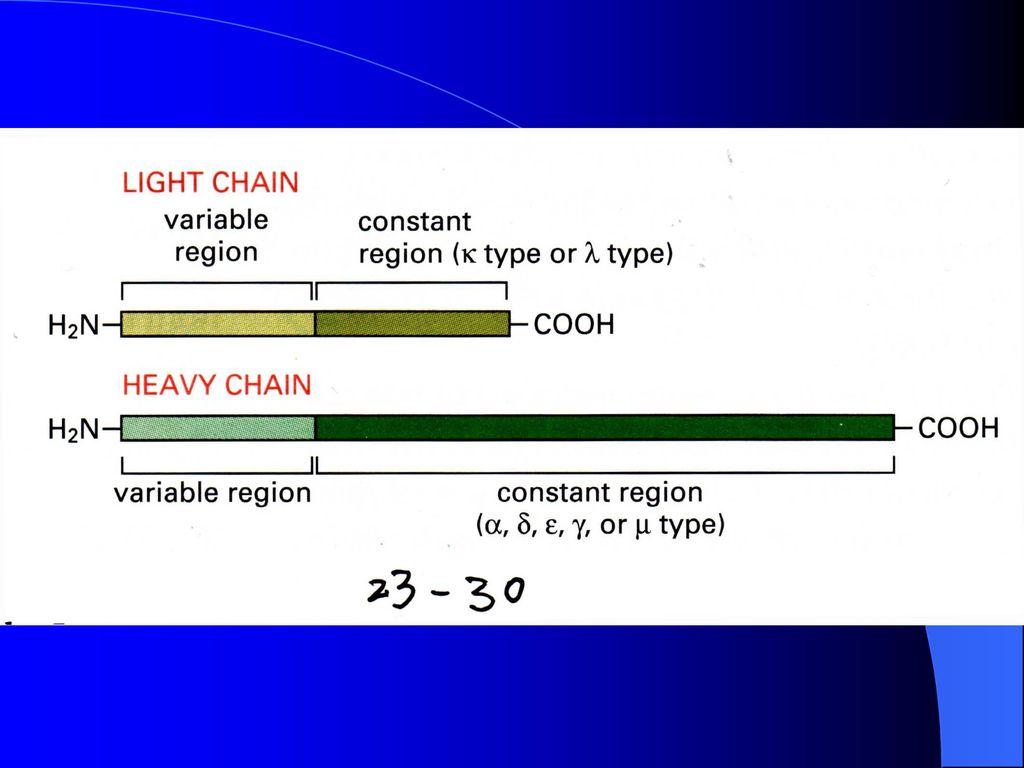
免疫球蛋白的肽链主要由可变区(V区)、恒定区(C区)以及两者之间的连接区(J区)组成,V、C和J基因片段在胚胎细胞中相隔较远。编码产生免疫球蛋白的细胞发育分化时,通过染色体内DNA重组把4个相隔较远的基因片段连接在一起,产生具有表达活性的免疫球蛋白基因。
、恒定区(C区)以及两者之间的连接区(J区)组成,V、C和J基因片段在胚胎细胞中相隔较远。编码产生免疫球蛋白的细胞发育分化时,通过染色体内DNA重组把4个相隔较远的基因片段连接在一起,产生具有表达活性的免疫球蛋白基因。")
38
表8-3 人类基因组中免疫球蛋白基因 主要片段的数量比较
成份 基因位点 染色体 基因片段数量 V D J C 重链 IGH 14 86 30 9 11 轻链(K链) IGK 2 76 5 1 轻链(λ链) IGL 22 52 7
 IGK 轻链(λ链) IGL")
39
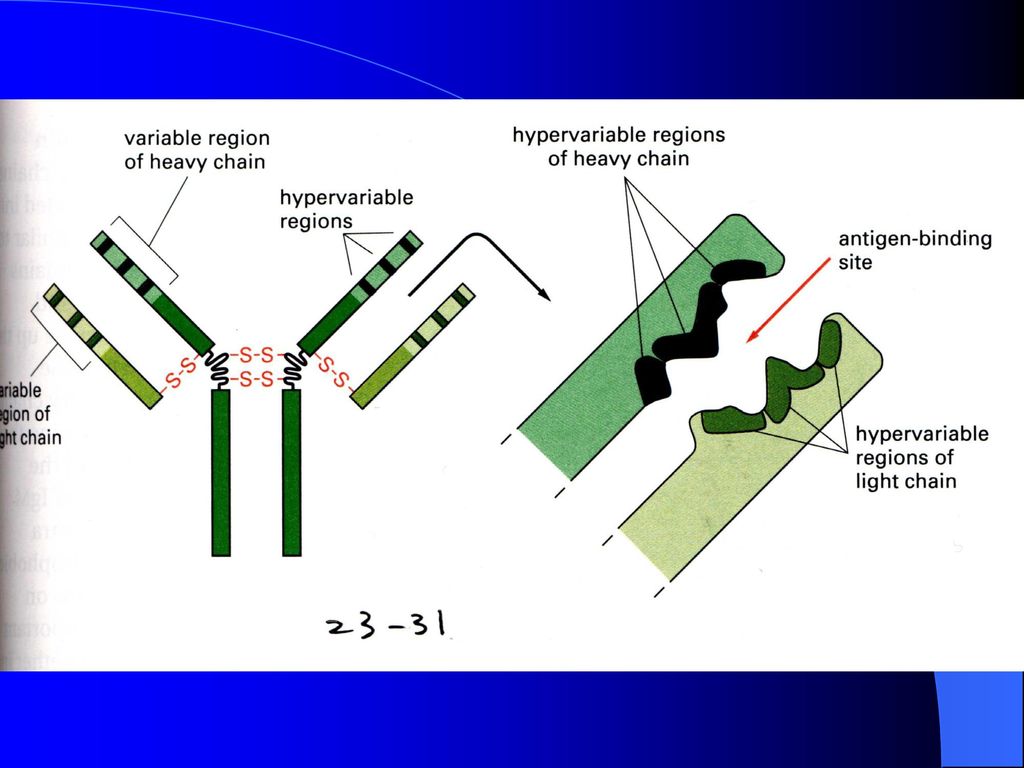
图7-11 免疫球蛋白重链基因片段重排与组织特异性表达

43
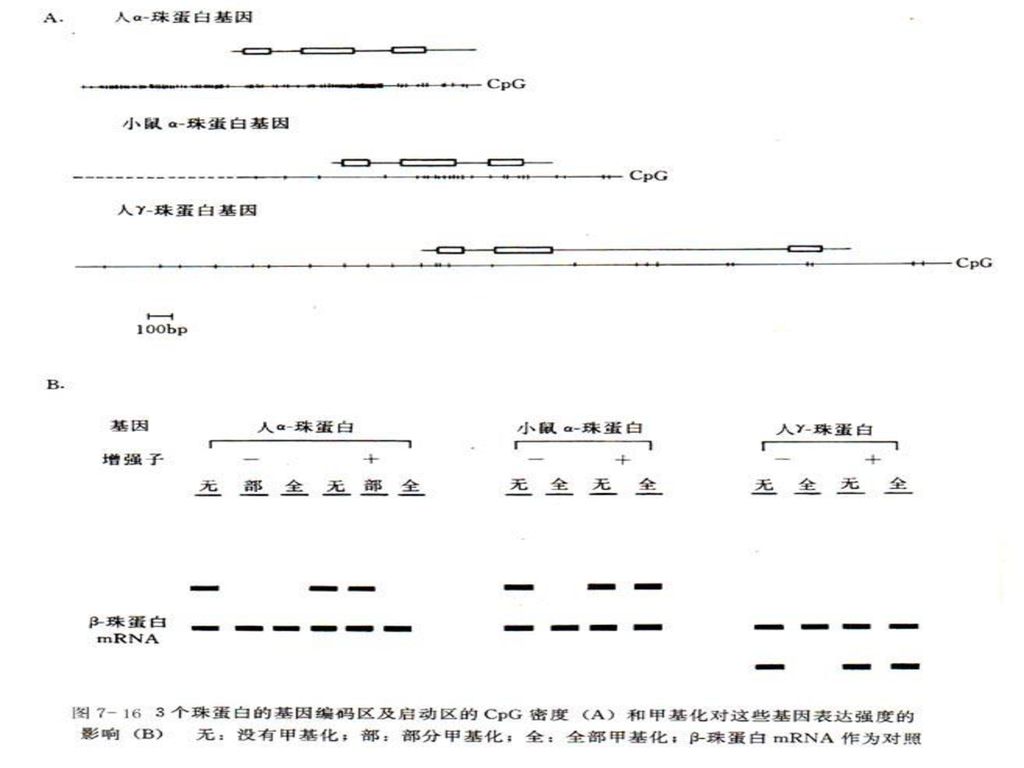
DNA甲基化与基因调控 DNA甲基化能引起染色质结构、DNA构象、DNA稳定性及DNA与蛋白质相互作用方式的改变,从而控制基因表达。研究证实,CpG二核苷酸中胞嘧啶的甲基化导致了人体1/3以上由于碱基转换而引起的遗传病。

44
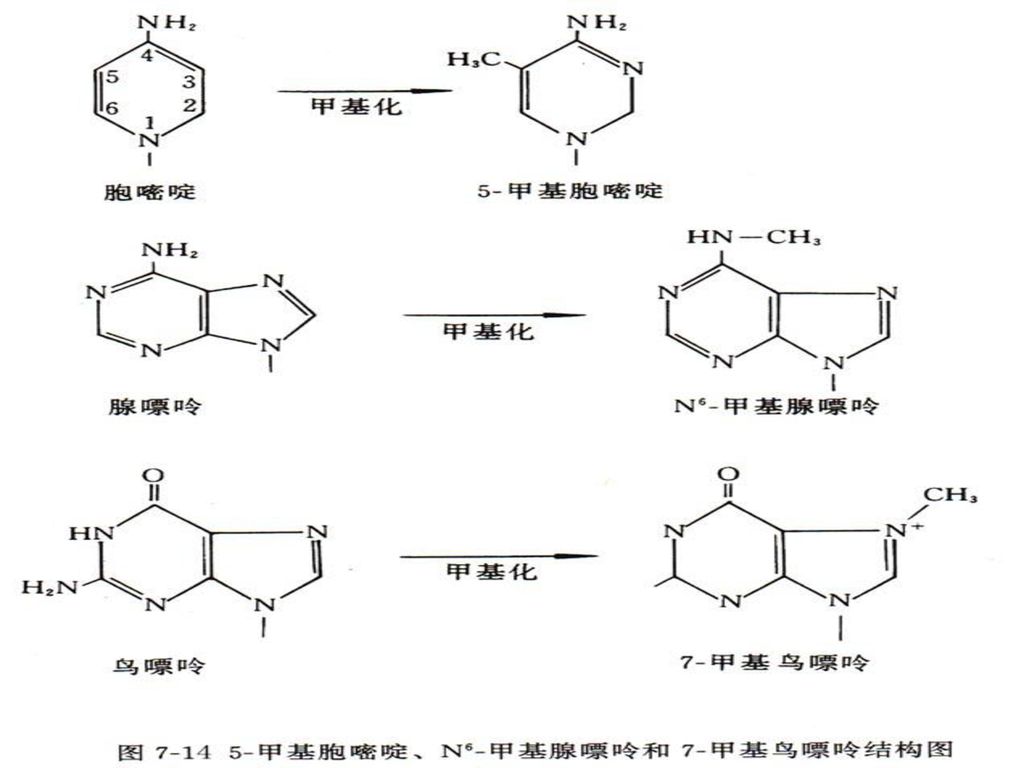
1、DNA甲基化 DNA甲基化主要形成5-甲基胞嘧啶(5-mC)和少量的N6-甲基腺嘌呤(N6-mA)及7-甲基鸟嘌呤(7-mG)。真核生物中,5-甲基胞嘧啶主要出现在CpG、CpXpG、CCA/TGG和GATC中。
和少量的N6-甲基腺嘌呤(N6-mA)及7-甲基鸟嘌呤(7-mG)。真核生物中,5-甲基胞嘧啶主要出现在CpG、CpXpG、CCA/TGG和GATC中。")
46
高等生物CpG二核苷酸中的C通常被甲基化,极易自发脱氨,生成胸腺嘧啶,所以CpG二核苷酸序列出现的频率远远低于按核苷酸组成计算出的频率。

47
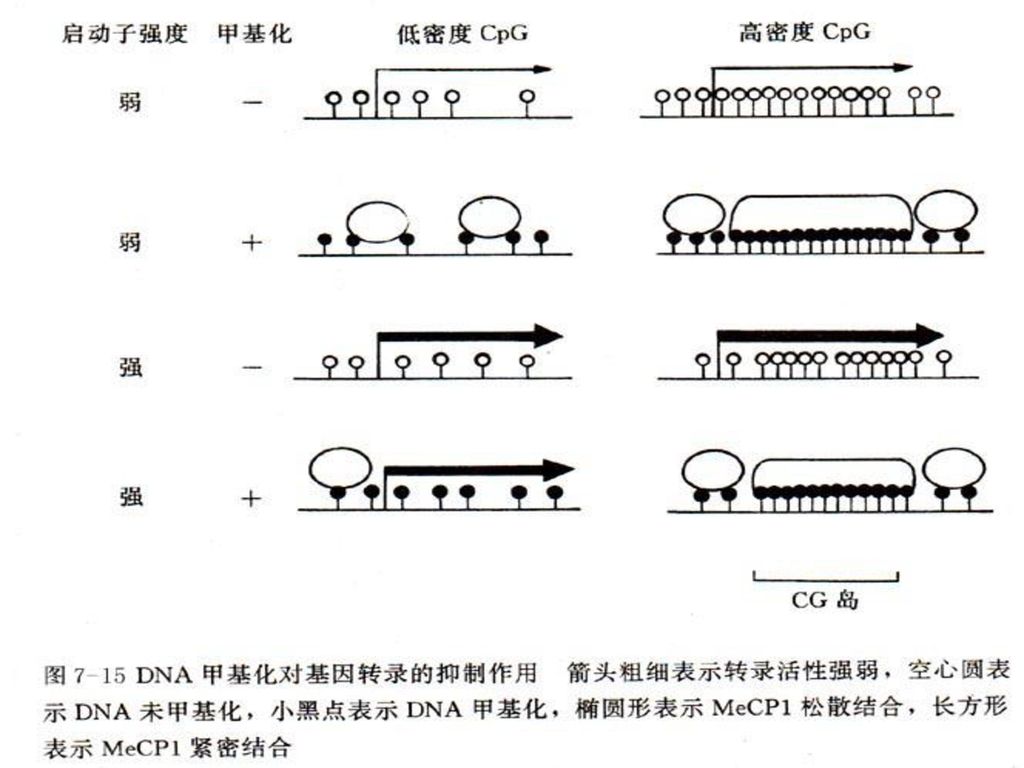
2、DNA甲基化抑制基因转录的机制 DNA甲基化导致某些区域DNA构象变化,从而影响DNA与蛋白质相互作用,抑制转录因子与启动区DNA的结合效率。 对于弱启动子来说,稀少的甲基化就能使其完全失去转录活性。当这一类启动子被增强时(带有增强子),即使不去甲基化也可以恢复其转录活性。若进一步提高甲基化密度,即使增强后的启动子仍无转录活性。
,即使不去甲基化也可以恢复其转录活性。若进一步提高甲基化密度,即使增强后的启动子仍无转录活性。")
50
3、DNA甲基化与X染色体失活 Xist基因的表达是决定X染色体失活的关键因素。Xist表达,则X染色体失活;不表达, X染色体有活性。

51
7. 2 真核基因转录机器的主要组成 真核基因调控主要也是在转录水平上进行,受大量特定的顺式作用元件(cis-acting element)和反式作用因子(transacting factor,又称跨域作用因子)调控。
和反式作用因子(transacting factor,又称跨域作用因子)调控。 .")
52
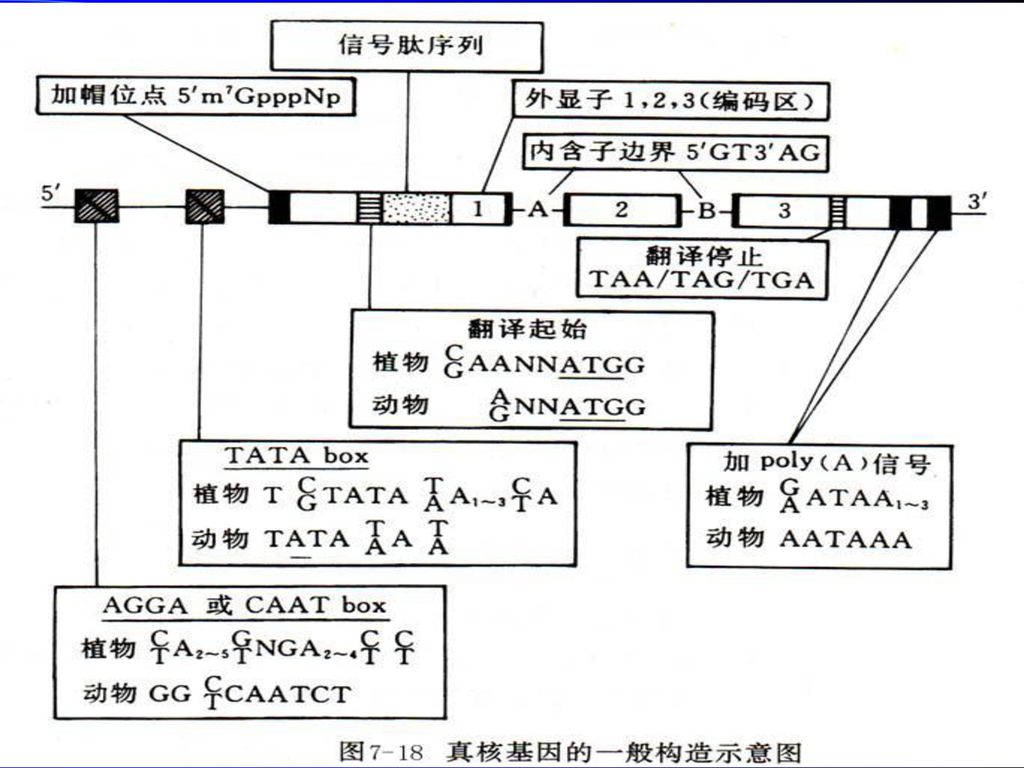
8.2.1 真核基因的转录 一个完整的基因,不但包括编码区(coding region),还包括5’和3’端长度不等的特异性序列,它们虽然不编码氨基酸,却在基因表达的过程中起着重要作用。所以,“基因”的分子生物学定义是:产生一条多肽链或功能RNA所必需的全部核苷酸序列。
,还包括5’和3’端长度不等的特异性序列,它们虽然不编码氨基酸,却在基因表达的过程中起着重要作用。所以, 基因 的分子生物学定义是:产生一条多肽链或功能RNA所必需的全部核苷酸序列。")
54
1、启动子(promoter) 真核基因启动子由核心启动子和上游启动子两个部分组成,是在基因转录起始位点(+1)及其5’上游大约100~200bp以内的一组具有独立功能的DNA序列,每个元件长度约为7~20bp,是决定RNA聚合酶II转录起始点和转录频率的关键元件。
 真核基因启动子由核心启动子和上游启动子两个部分组成,是在基因转录起始位点(+1)及其5’上游大约100~200bp以内的一组具有独立功能的DNA序列,每个元件长度约为7~20bp,是决定RNA聚合酶II转录起始点和转录频率的关键元件。")
55
(1)核心启动子(core promoter):是指保证RNA聚合酶II转录正常起始所必需的、最少的DNA序列,包括转录起始位点及转录起始位点上游-25-30bp处的TATA盒。核心启动子确定转录起始位点并产生基础水平的转录。
核心启动子(core promoter):是指保证RNA聚合酶II转录正常起始所必需的、最少的DNA序列,包括转录起始位点及转录起始位点上游-25-30bp处的TATA盒。核心启动子确定转录起始位点并产生基础水平的转录。")
56
(2)上游启动子元件(upstream promoter element,UPE)包括通常位于-70bp附近的CAAT盒(CCAAT)和GC盒(GGGCGG)等,能通过TFIID复合物调节转录起始的频率,提高转录效率。
上游启动子元件(upstream promoter element,UPE)包括通常位于-70bp附近的CAAT盒(CCAAT)和GC盒(GGGCGG)等,能通过TFIID复合物调节转录起始的频率,提高转录效率。")
57
2、转录模板 3、RNA聚合酶II 4、RNA聚合酶II基础转录所需蛋白因子

58
8.2.2 增强子及其对转录的影响 增强子是指能使与它连锁的基因转录频率明显增加的DNA序列,最早发现于SV40早期基因的上游,有两个长72bp的正向重复序列。

59
增强子通常具有下列特性: A、增强效应十分明显。 B、增强效应与其位置和取向无关。 C、大多为重复序列(50bp)。 D、其增强效应有严密的组织和细胞特异性。 E、没有基因专一性,可以在不同的基因组合上表现 增强效应; F、许多增强子还受外部信号的调控。

60
8.2.3 反式作用因子 根据各个蛋白质成分在转录中的作用,能将整个复合物分为3部分:
反式作用因子 根据各个蛋白质成分在转录中的作用,能将整个复合物分为3部分: (1)参与所有或某些转录阶段的RNA聚合酶亚基,不具有基因特异性。 (2)与转录的起始或终止有关的辅助因子,不具有基因特异性。 (3)与特异调控序列结合的转录因子。
参与所有或某些转录阶段的RNA聚合酶亚基,不具有基因特异性。 (2)与转录的起始或终止有关的辅助因子,不具有基因特异性。 (3)与特异调控序列结合的转录因子。")
61
1、反式作用因子中的DNA识别或结合域 反式作用因子是能直接或间接地识别或结合在各类顺式作用元件核心序列上(表8-5),参与调控靶基因转录效率的蛋白质。
,参与调控靶基因转录效率的蛋白质。")
62
表8-5 部分球蛋白基因5’调控区序列的保守性比较。 斜体表示高度保守的序列
表8-5 部分球蛋白基因5’调控区序列的保守性比较。 斜体表示高度保守的序列 基 因 -80区 -30区 小鼠βmin GGCCAATCTGCTC GGTATATAAA 小鼠βmaj AGCATATAAG 兔β GGCCAATCTAC-- GGCATAAAAG 山羊βA AGCCAATCTGCTC 山羊βC 人β GGCCAATCTACTC 人δ AACCAATCTGCTC -GCATAAAAG 人Aγ GACCAATAGCCTT 人Gγ 人α GACCAATGACTTT

63
(1)螺旋-转折-螺旋(helix-turn-helix, H-T-H)结构
这一类蛋白质分子中有至少两个α螺旋,中间由短侧连氨基酸残基形成“转折”,近羧基端的α螺旋中氨基酸残基的替换会影响该蛋白质在DNA双螺旋大沟中的结合。

65
(2)锌指(Zinc finger)结构 包括锌指、锌钮(twist)和锌簇(cluster)结构,有锌参与时才具备转录调控活性。与DNA的结合较为牢固,特异性也很高。 类固醇激素受体家族含有连续的两个锌指结构,以同源或异源性二聚体的方式将两个α螺旋结合在相邻的两个大沟中。
锌指(Zinc finger)结构 包括锌指、锌钮(twist)和锌簇(cluster)结构,有锌参与时才具备转录调控活性。与DNA的结合较为牢固,特异性也很高。 类固醇激素受体家族含有连续的两个锌指结构,以同源或异源性二聚体的方式将两个α螺旋结合在相邻的两个大沟中。")
66
图8-19 典型的类固醇激素受体结构示意图

67
(3)碱性-亮氨酸拉链(basic-leucine zipper),即bZIP结构
蛋白中每隔6个氨基酸就有一个亮氨酸残基,导致第7个亮氨酸残基都在螺旋的同一方向出现。 以二聚体形式与DNA结合,亮氨酸拉链区并不直接结合DNA,肽链氨基端20~30个富含碱性氨基酸结构域与DNA结合,但以碱性区和亮氨酸拉链结构域整体作为基础。

68
图7-20 碱性亮氨酸拉链结构域转录激活因子。

69
图8-21 碱性亮氨酸拉链转录激活蛋白与调控区DNA相结合的示意图

70
(4)碱性-螺旋-环-螺旋(basic-helix/loop/helix,即bHLH结构)
在免疫球蛋白κ轻链基因的增强子结合蛋白E12与E47中,羧基端100~200个氨基酸残基可形成两个双性α螺旋,被非螺旋的环状结构所隔开,蛋白质的氨基端则是碱性区,其DNA结合特性与亮氨酸拉链类蛋白相似。

71
bHLH类蛋白只有形成同源或异源二聚体时,才具有足够的DNA结合能力。当这类异源二聚体中的一方不含有碱性区(如Id或E12蛋白)时,该二聚体明显缺乏对靶DNA的亲和力。
时,该二聚体明显缺乏对靶DNA的亲和力。")
72
图8-22 bHLH蛋白的DNA结合能力分析

73
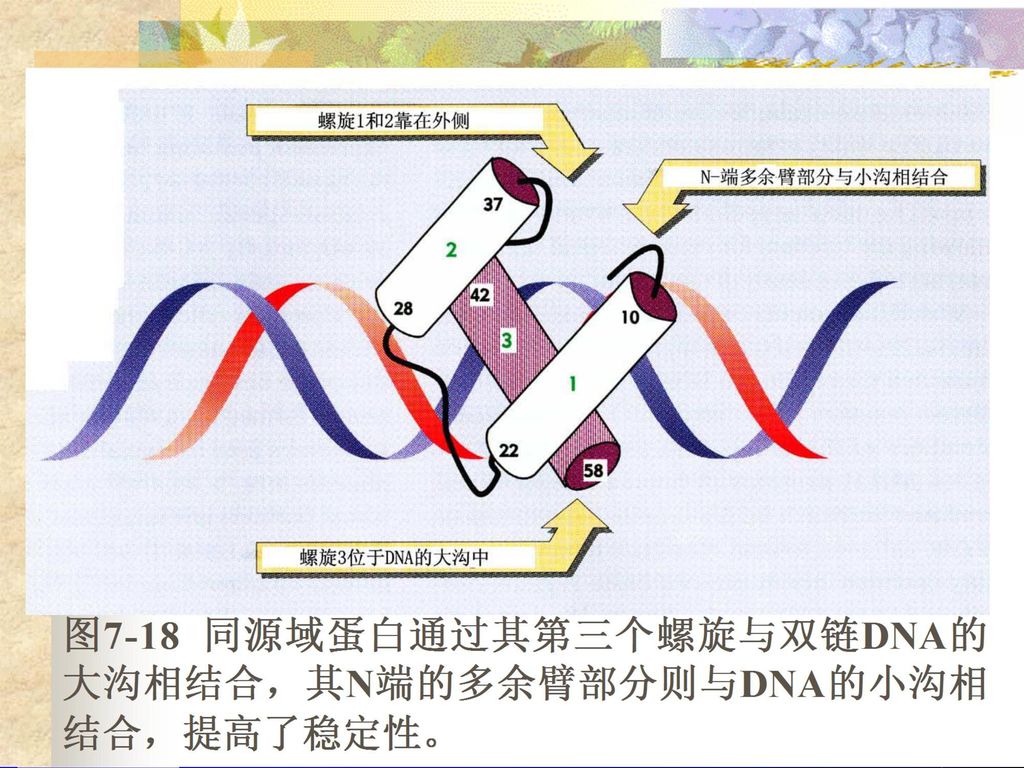
(5)同源域(homeo domains) 又称发育同源结构域,是指存在于某些蛋白质中的与DNA结合的模体(基序),这些蛋白质对发育过程中基因的表达有调节作用。发育同源结构域在结构上及与DNA相互作用的方式上均极为保守,60氨基酸残基长,由三个α螺旋及一个氨基末断臂组成。
同源域(homeo domains) 又称发育同源结构域,是指存在于某些蛋白质中的与DNA结合的模体(基序),这些蛋白质对发育过程中基因的表达有调节作用。发育同源结构域在结构上及与DNA相互作用的方式上均极为保守,60氨基酸残基长,由三个α螺旋及一个氨基末断臂组成。")
74
第三个α螺旋的残基与大沟的特异碱基接触,氨基末断臂的残基与邻近的DNA小沟接触。它识别特定的DNA共有序列。它广泛存在于真核生物基因组内,由于最早从果蝇homeotic loci(该遗传位点的基因产物决定了躯体发育)中克隆得到而命名。
中克隆得到而命名。")
75
表8-8 含有同源转换区的转录因子及其DNA靶序列
蛋白因子 靶 DNA 序 列 Oct-1、Oct-2 ATTTGCAT Pit-1/GHF-1 TATGCATAA en、ftz、ubx TCAATTAAAT Eve TCAATTAAAT TCAGCACCG Bicoid TCAATCCC

76
图8-23 各种同源域DNA结合蛋白中保守序列分析

77
2、 转录活化结构域 在真核生物中,反式作用因子的功能由于受蛋白质-蛋白质之间相互作用的调节变得精密、复杂,完整的转录调控功能通常以复合物的方式来完成,因此,是否具有转录活化域就成为反式作用因子中唯一的结构基础。

78
P 酵母单杂、双杂体系 图8-24 几种常见的转录活化结构域

79
不同的转录活化域有下列特征性结构: 1、带负电荷的螺旋结构。哺乳动物细胞中糖皮质激素受体的两个转录活化域,AP1家族的Jun及GAL4都有酸性的螺旋结构,它们可能与TFIID复合物中某个通用因子或RNA聚合酶II本身结合,具有稳定转录起始复合物的作用。

80
2、富含谷氨酰胺的结构。 SP1是启动子GC盒的结合蛋白,共有4个参与转录活化的区域,其中最强的转录活化域含25%左右的谷氨酰胺。 Oct1/2、Jun、AP2、血清应答因子(SRF)等都有相同的富含谷氨酰胺的结构域。
等都有相同的富含谷氨酰胺的结构域。")
81
3、富含脯氨酸的结构。 CTF-NF1因子的羧基端富含脯氨酸(达20%~30%),很难形成α螺旋。在Oct2、Jun、AP2、SRF等哺乳动物因子中也有富含脯氨酸的结构域。
,很难形成α螺旋。在Oct2、Jun、AP2、SRF等哺乳动物因子中也有富含脯氨酸的结构域。")
82
真核基因转录调控的主要模式: (1)蛋白质磷酸化对基因转录的调控。 (2)蛋白质乙酰化对基因转录的调控。
(3)激素与热激蛋白对基因转录的调控。
激素与热激蛋白对基因转录的调控。")
83
8.3 蛋白质磷酸化对基因转录的调控 细胞是生命活动的基本单位。细胞通过DNA的复制和细胞分裂将本身所固有的遗传信息由亲代传至子代,实现增殖繁衍。 它们还不断地“感知”环境变化,并对其作出特定的应答。

84
细胞对外界信息的应答可以分为3个阶段: 1.外界信息的“感知”,即由细胞膜到细胞核内的信号传递; 2.染色质水平上的基因活性调控;
3.特定基因的表达,即从DNA→RNA→蛋白质的遗传信息传递过程。

85
细胞信号类型 (1)近分泌信号(paracrine) (2)内分泌信号(endocrine) (3)神经信号(synaptic)
近分泌信号(paracrine) (2)内分泌信号(endocrine) (3)神经信号(synaptic)")
86
细胞信号的一般概念 信号分子和信号受体 信号分子:指细胞外信号分子,有几百种,包括蛋白质、小肽、氨基酸、核酸、固醇类和脂肪酸类似物
信号受体: 1.细胞表面受体 2.细胞内受体

87
细胞表面受体和细胞内受体 细胞表面受体 (1)离子通道关联受体 (2)G蛋白关联受体 (3)酶关联受体 细胞内受体
离子通道关联受体 (2)G蛋白关联受体 (3)酶关联受体 细胞内受体")
88
图8-27 细胞表面的三类受体示意图。

89
跨膜信息传递的三个环节 受体识别 信号传导 细胞内效应
细胞表面受体与配体分子的高亲和力特异性结合,能诱导受体蛋白构象变化,使胞外信号通过各种方式被传至细胞内。

90
蛋白质磷酸化、信号转导及基因表达 近年来,数条跨膜信号转导途径已被逐步阐明,在信号转导的上游途径中,以酪氨酸蛋白激酶(PTK)途径及受体偶联的G蛋白途径较为引人注目——主要通过磷酸化/去磷酸化来进行调控。
途径及受体偶联的G蛋白途径较为引人注目——主要通过磷酸化/去磷酸化来进行调控。")
91
表8-9 真核细胞主要跨膜信号转导途径 体 系 作用方式 蛋白激酶受体 配体-受体结合激活受体细胞质内蛋白酪氨酸激酶或丝氨酸/苏氨酸激酶
体 系 作用方式 蛋白激酶受体 配体-受体结合激活受体细胞质内蛋白酪氨酸激酶或丝氨酸/苏氨酸激酶 受体耦联的G蛋白 配体结合后激活特定的G蛋白,活化其α亚单位,并调节特定靶分子,如腺苷酸环化酶、磷脂酶、cGMP磷酸二酯酶和离子通道的活性 离子通道 配体结合特定可关闭或开启的离子通道 心钠素(ANF)受体 配体结合活化鸟苷酸环化酶 细胞因子受体 细胞因子与受体结合,抑制或激活核基因表达 类固醇激素基因家族的核受体 与配体结合诱导或抑制靶基因表达
受体. 配体结合活化鸟苷酸环化酶. 细胞因子受体. 细胞因子与受体结合,抑制或激活核基因表达. 类固醇激素基因家族的核受体. 与配体结合诱导或抑制靶基因表达.")
92
蛋白质的磷酸化与去磷酸化过程是生物体内普遍存在的信息传导调节方式,几乎涉及所有的生理及病理过程,如糖代谢、光合作用、细胞的生长发育、神经递质的合成与释放甚至癌变等等。

93
图8-28 蛋白质磷酸化和GTP结合蛋白 参与的信号转导过程

94
蛋白质的磷酸化及去磷酸化

95
已发现蛋白激酶(protein kinase,PK)基因2000余个和1000个左右蛋白质去磷酸化酶基因。根据底物蛋白质被磷酸化的氨基酸残基可分为:
①丝氨酸/苏氨酸型; ②酪氨酸型; ③组氨酸型。

96
根据是否有调节物参与蛋白激酶活性可分为两大类:信使依赖型(又分胞内信使、调节因子依赖型和激素或生长因子依赖型,表8-10)和非信使依赖型(表8-11)。
细胞受刺激后,通过蛋白质磷酸化及一系列级联放大过程将胞外信号转化为胞内信号,从而引起广泛的生理反应。

97
表8-10 信使依赖型蛋白激酶 胞内信使或调节因子依赖型PK 激素或生长因子依赖型PK 名称 调节物 动物 植物
cAMP依赖型PK (PKA) cAMP + ? 表皮生长因子(EGF) 依赖型PK EGF - cAMP依赖型PK(PKG) cGMP 胰岛素依赖型PK (胰岛素样生长因子) 胰岛素 Ca2+、CaM 依赖型PK Ca2+、 CaM 血小板衍生生长因子(PDGF)依赖型PK PDGF Ca2+、磷脂依赖型PK(PKC) 磷脂 类胰岛素生长因子-1 (IGF-1)依赖型PK IGF-1 只依赖于Ca2+的PK Ca2+ 集落刺激因子-1 (CSF-1)依赖型PK CSF-1
 cAMP. + ? 表皮生长因子(EGF) 依赖型PK. EGF. - cAMP依赖型PK(PKG) cGMP. 胰岛素依赖型PK (胰岛素样生长因子) 胰岛素. Ca2+、CaM. 依赖型PK. Ca2+、 CaM. 血小板衍生生长因子(PDGF)依赖型PK. PDGF. Ca2+、磷脂依赖型PK(PKC) 磷脂. 类胰岛素生长因子-1 (IGF-1)依赖型PK. IGF-1. 只依赖于Ca2+的PK. Ca2+ 集落刺激因子-1 (CSF-1)依赖型PK. CSF-1.")
98
表8-11 非信使依赖型蛋白激酶 名 称 动物 植物 酪蛋白激酶(I,II型) + 糖原合成酶激酶 ? 组蛋白激酶II 视紫红质蛋白激酶 -
名 称 动物 植物 酪蛋白激酶(I,II型) + 糖原合成酶激酶 ? 组蛋白激酶II 视紫红质蛋白激酶 - β-肾上腺素受体蛋白激酶
 + 糖原合成酶激酶. ? 组蛋白激酶II. 视紫红质蛋白激酶. - β-肾上腺素受体蛋白激酶.")
99
受体分子活化细胞功能的途径主要有两条: 一是受体本身或受体结合蛋白具有内源酪氨酸激酶活性,胞内信号通过酪氨酸激酶途径得到传递; 二是配体与细胞表面受体结合,通过G蛋白介异的效应系统产生介质,活化丝氨酸/苏氨酸或酪氨酸激酶,从而传递信号。

100
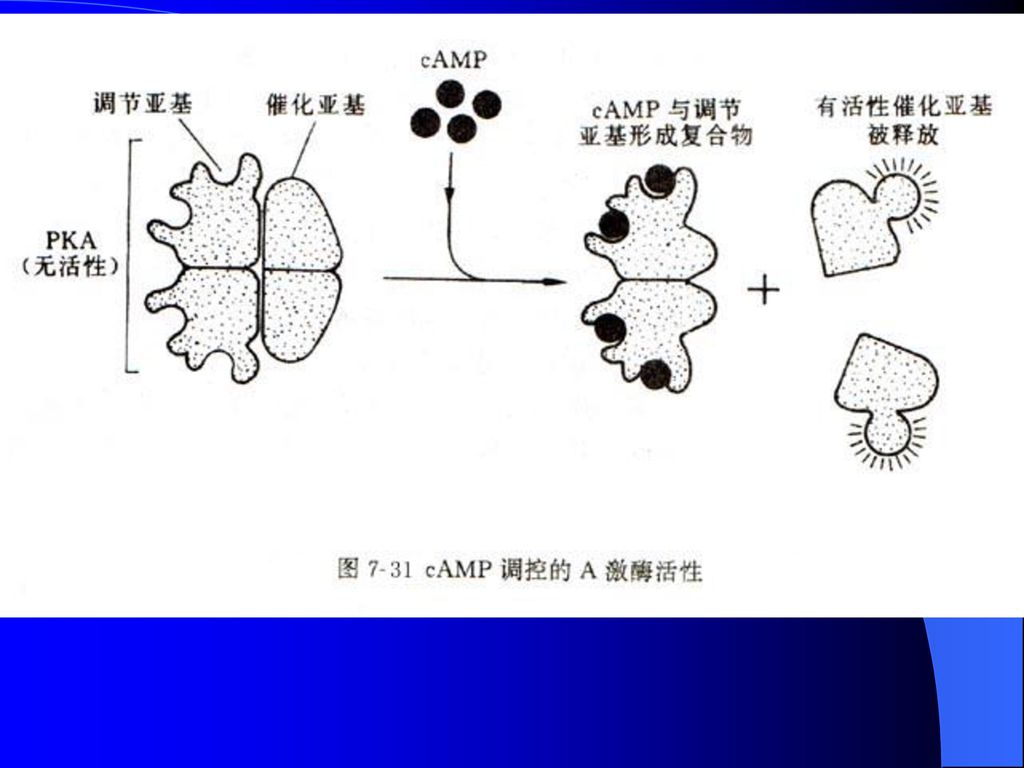
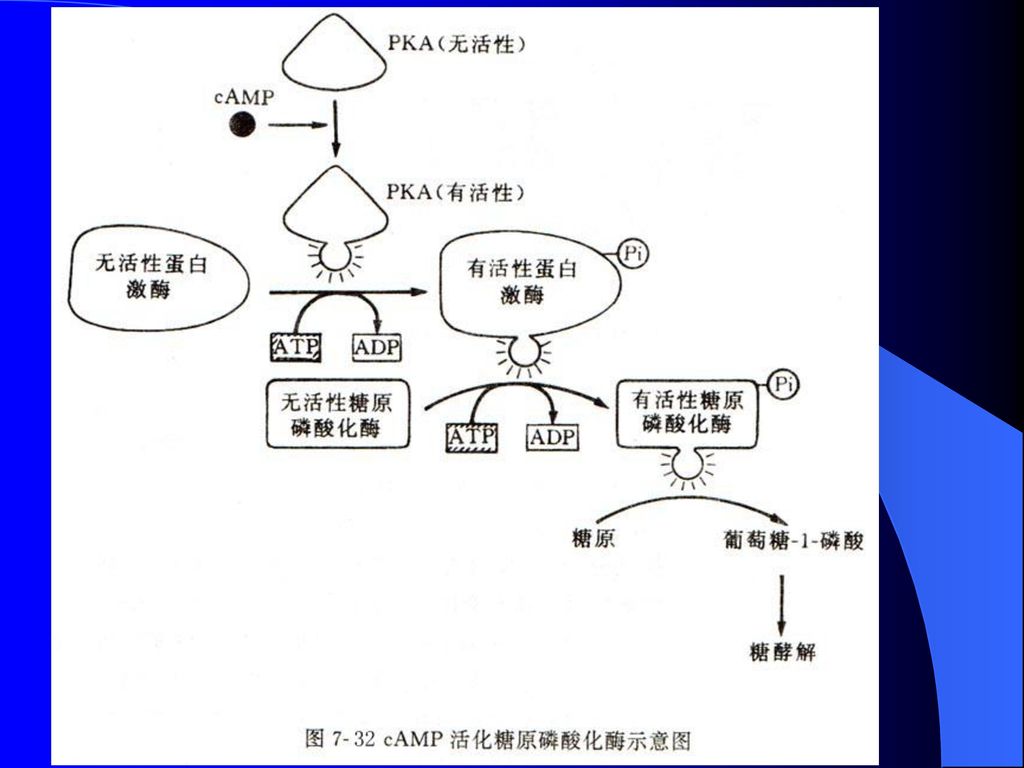
受cAMP水平调控的A激酶 依赖于cAMP的蛋白激酶称为A激酶(PKA),非活性状态的PKA全酶由4个亚基R2C2所组成,分子量约为 ,调节亚基与cAMP相结合,引起构象变化并释放催化亚基,后者随即成为有催化活性的单体(图8-30)。 激活后的PKA能把ATP分子上的末端磷酸基团加到某个特定蛋白质的丝氨酸或苏氨酸残基上。

102
糖原代谢: 激素与其受体在肌肉细胞外表面相结合,诱发细胞质cAMP的合成并活化A激酶,后者再将活化磷酸基团传递给无活性的磷酸化酶激酶,活化糖原磷酸化酶,最终将糖原磷酸化,进入糖酵解过程并提供ATP(图8-30)。
。")
104
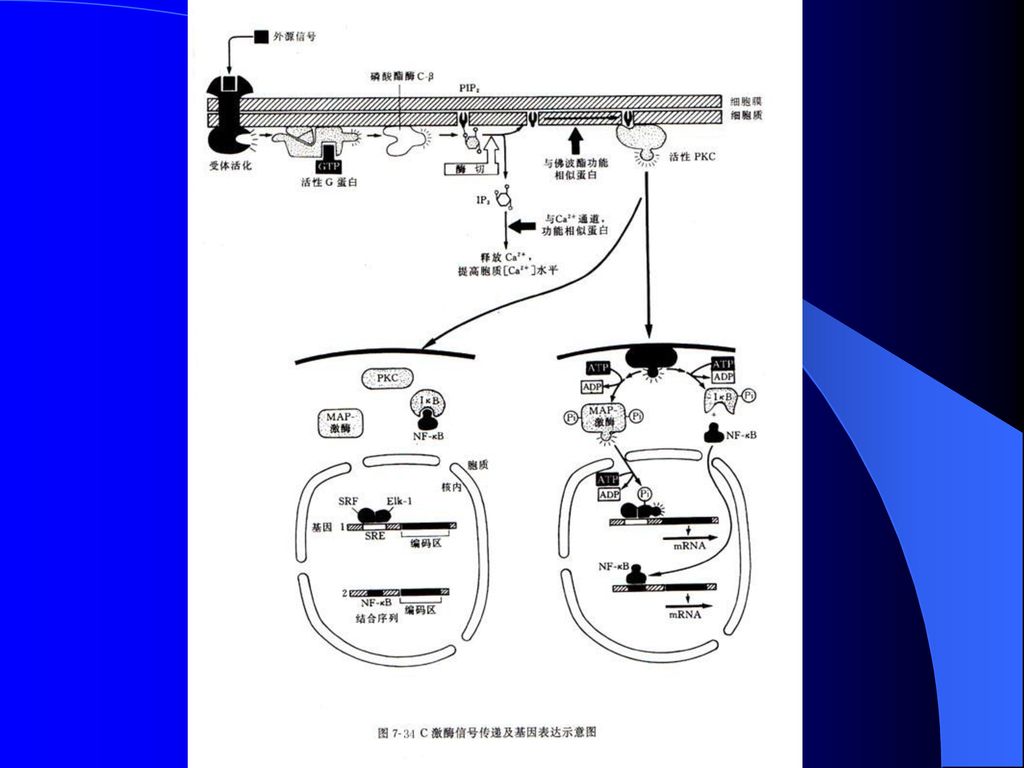
8.3.2 C激酶与PIP2、IP3和DAG 该蛋白激酶活性是依赖于Ca2+的,所以称为C激酶(PKC)。
IP3引起细胞质Ca2+浓度升高,导致C激酶从胞质转运到靠近原生质膜内侧处,并被DAG和Ca2+的双重影响所激活。DAG提高了C激酶对于Ca2+的亲和力。

105
C激酶是一个7.7×104的蛋白质,主要实施对丝氨酸、苏氨酸的磷酸化,它具有一个催化结构域和一个调节结构域。

107
CaM激酶及MAP激酶 Ca2+的细胞学功能主要通过钙调蛋白激酶(CaM-kinase)来实现的,它们也是一类丝氨酸/苏氨酸激酶,但仅应答于细胞内Ca2+水平。
来实现的,它们也是一类丝氨酸/苏氨酸激酶,但仅应答于细胞内Ca2+水平。")
108
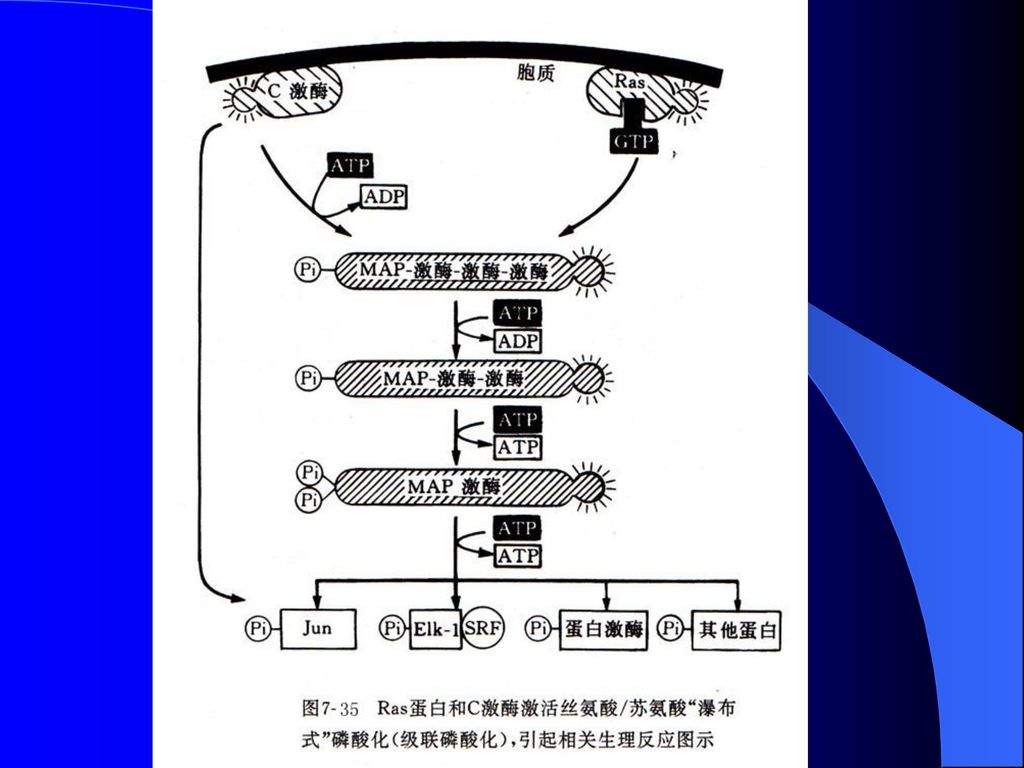
MAP激酶(mitogen-activated proteinkinase,MAP-kinase,又称为extracellular-signal-regulated kinase,ERKS)活性受许多外源细胞生长、分化因子的诱导。MAP-激酶的活性取决于该蛋白中仅有一个氨基酸之隔的酪氨酸、丝氨酸残基是否都被磷酸化。
活性受许多外源细胞生长、分化因子的诱导。MAP-激酶的活性取决于该蛋白中仅有一个氨基酸之隔的酪氨酸、丝氨酸残基是否都被磷酸化。")
109
能同时催化这两个氨基酸残基磷酸化的酶被称为MAP-激酶-激酶,它的反应底物是MAP激酶。MAP-激酶-激酶本身能被MAP-激酶-激酶-激酶所磷酸化激活,后者能同时被C激酶或酪氨酸激酶家族的Ras蛋白等激活,从而在信息传导中发挥功能。

111
8.3.4 酪氨酸激酶(PTK)途径 包括跨膜受体家族与胞质非受体家族两大类。
途径 包括跨膜受体家族与胞质非受体家族两大类。")
112
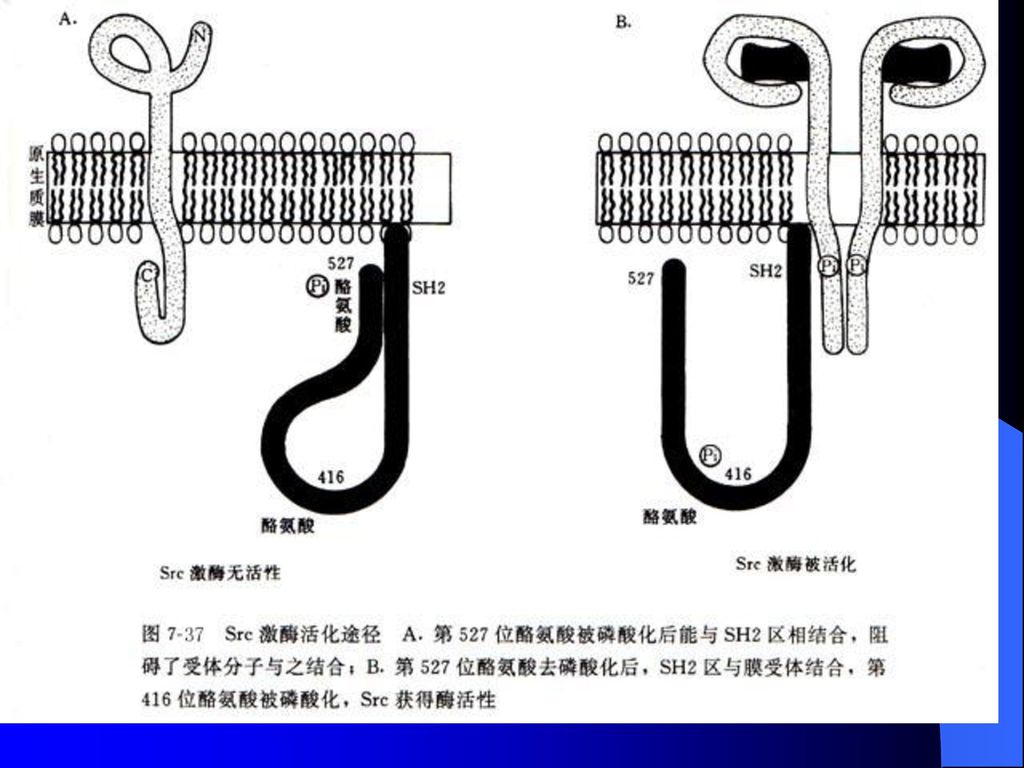
因为Src亚家族非受体酪氨酸激酶以及许多PTK作用底物分子都存在被称为Src同源结构域(Src homology domain,SH)。
举例:Src亚家族成员pp60c-Src

113
在正常细胞中,pp60c-Src上的Tyr527被磷酸化并以头尾相连的缔合方式折入同一分子的SH2结构域中,从而抑制了PTK活性。pp60c-Src与靶分子PI3激酶,rasGAP等的缔合作用均涉及pp60c-SrcSH2结构域与PI3激酶及rasGAP分子上酪氨酸磷酸化区的结合。

117
8.5 激素与热激蛋白对基因表达的影响 许多类固醇激素(如雌激素、孕激素、醛固酮、糖皮质激素和雄激素)以及一般代谢性激素(如胰岛素)的调控作用都是通过起始基因转录而实现的。
以及一般代谢性激素(如胰岛素)的调控作用都是通过起始基因转录而实现的。")
118
图8-43 几种常见的疏水性小分子激素的结构式。
类皮质激素(肾上腺类固醇) 图8-43 几种常见的疏水性小分子激素的结构式。
 图8-43 几种常见的疏水性小分子激素的结构式。")
119
表7-12 三种激素应答序列成分分析 激 素 元 件 序 列 糖皮质激素 GRE TGGTACANNNTGTTCT 雌激素 ERE GGTCANNNTGTCC 甲状腺素 TRE CAGGGACGTGACCGCA

120
体内存在的许多糖皮质类激素应答基因都有一段大约20bp的顺式作用元件(激素应答元件,简称HRE),该序列具有类似增强子的作用,其活性受激素制约。靶细胞中含有大量激素受体蛋白,而非靶细胞中没有或很少有这类受体。
,该序列具有类似增强子的作用,其活性受激素制约。靶细胞中含有大量激素受体蛋白,而非靶细胞中没有或很少有这类受体。")
121
固醇类激素的受体蛋白分子有相同的结构框架,包括保守性极高并位于分子中央的DNA结合区,位于C端的激素结合区和保守性较低的N端。如果糖皮质激素受体蛋白激素结合区的某个部分丢失,就变成一种永久型的活性分子。

122
图8-44 激素及其胞内受体介导的基因表达调控模式。(A)小分子激素激活其胞内受体分子的机制;(B)胞内受体的基本结构分析。
小分子激素激活其胞内受体分子的机制;(B)胞内受体的基本结构分析。")
123
多肽激素胰高血糖素接触靶细胞时,首先与受体结合,并激活膜上的腺苷酸环化酶,使之以Mg2+-ATP为底物生成环腺苷酸和焦磷酸。细胞内cAMP浓度升高,导致蛋白激酶活性增强,特定酶系的磷酸化水平及酶活性都得到改善,促进糖原最终分解为葡萄糖-1-磷酸(图8-45)。
。")
124
图7-39

125
热激蛋白诱导的基因表达 能与某个(类)专一蛋白因子结合,从而控制基因特异表达的DNA上游序列称为应答元件(response element)。
专一蛋白因子结合,从而控制基因特异表达的DNA上游序列称为应答元件(response element)。")
126
主要有: 热激应答元件(heat shock response element,HSE) 糖皮质应答元件(glucocorticoid response element,GRE) 金属应答元件(metal response element,MRE)。 应答元件与细胞内专一的转录因子相互作用,协调相关基因的转录 。

127
表8-14特异转录因子与各类应答元件的相互作用
调控因子 应答元件 DNA序列 结合蛋白 相对分子质量 热激 HSE CNNGAANNTCCNNG HSF 9.3×104 镉 MRE CGNCCCGGNCNC ? ? 佛波酯 TRE TGACTCA AP1 3.9×104 血清 SRE CCATATTAGG SRF 5.2×104

128
许多生物在最适温度范围以上,能受热诱导合成一系列热休克蛋白(heat shock protein,HSP)。
如受热后,果蝇细胞内Hsp70 mRNA水平提高1 000倍,就是因为热激因子(heat shock factor,HSF)与hsp70基因TATA区上游60bp处的应答元件HSE相结合,诱发转录起始。
与hsp70基因TATA区上游60bp处的应答元件HSE相结合,诱发转录起始。")
129
表8-15 主要HSP家族及其生理功能 家 族 主 要 成 员 主 要 生 理 功 能 免疫应答中的可能作用 HSP90
家 族 主 要 成 员 主 要 生 理 功 能 免疫应答中的可能作用 HSP90 HSP90, HSP83 促进甾醇激素受体与激素的结合及与DNA结合,调节激酶磷酸化活性。 抗肿瘤,提高自身免疫性能 HSP70 HSP70, BiP, Dnat, Hsc70, Grp78 参与蛋白质折叠和去折叠、蛋白转位及多聚复合物的组装。 免疫球蛋白装配,类抗原加工,病原体抗原及自身免疫性诱导。 小分子HSP Hsp27, Gro23, HSP16 参与蛋白质折叠和去折叠及多聚复合物的装配。 病原体抗原及自身免疫性诱导。 泛 素 蛋白质降解。 类抗原加工,淋巴细胞回巢,自身免疫性诱导。

130
不受热或其他环境胁迫时,热激因子HSF主要以单体的形式存在于细胞质和核内。单体HSF没有DNA结合能力,Hsp70可能参与了维持HSF的单体形式。
受热激或其他环境胁迫时,细胞内变性蛋白增多,它们与HSF竞争结合Hsp70,从而释放HSF,使之形成三体并输入核内。HSF的三体能与HSE特异结合,促进基因转录。

131
图7-40

132
热激后,HSF不但形成三体,还会迅速被磷酸化。故磷酸化水平也影响HSF对基因表达的调控能力。
HSF与HSE的特异性结合,引起包括Hsp70在内的许多热激应答基因表达,大量产生Hsp70蛋白。

133
随着热激温度消失,细胞内出现大量游离的Hsp70蛋白,它们与HSF相结合,形成没有DNA结合能力的单体并脱离DNA。

134
图7-41

135
HSF具有多个可形成拉链的疏水DNA区域,其中3个位于N端,靠近DNA结合区,参与促进三体的形成。第4个拉链位于C末端,与位于第452~488位氨基酸残基之间的保守区一道参与维持HSF单体构象。

136
序列删除实验表明,无论去除第4个拉链区,还是更换该区甲硫氨酸391→赖氨酸,亮氨酸395→脯氨酸,都会导致HSF突变体对HSE在常温下的高亲和力。删除第452~488位氨基酸,也可部分替代热激效应 。

137
在常温下,Hsp蛋白第1和4号拉链发生相互作用,从而阻断了三体的形成;热激或其他环境胁迫导致内源拉链之间的解离,蛋白质伸展成长链,不同链上的拉链发生作用,形成具有DNA结合能力的三体。

138
图7-42

139
8.6 其他水平上的基因调控 8.6.1 RNA的加工成熟 1、rRNA和tRNA的加工成熟 rRNA:分子内切割和化学修饰。 tRNA:前体tRNA→成熟tRNA。

140
2、mRNA加工成熟 hnRNA→mRNA。

141
3、真核生物基因转录后加工的多样性 简单转录单位。这类基因只编码产生一个多肽,其原始转录产物有时需要加工,有时则不需要加工。这类基因转录后加工有3种不同形式。

142
图7-44

143
复杂转录单位。含有复杂转录单位的主要是一些编码组织和发育特异性蛋白质的基因,它们除了含有数量不等的内含子以外,其原始转录产物能通过多种不同方式加工成两个或两个以上的mRNA。

144
利用多个5’端转录起始位点或剪接位点产生不同的蛋白质。
肌球蛋白碱性轻链基因选用了不同的5’转录起始位点及剪接不同外显子产生蛋白质异构体LC1和LC3的过程。

146
大鼠降钙素基因编码与血钙代谢有关的蛋白质。它有一个转录起始位点和6个外显子,两个加多聚(A)位点。
初级转录产物以0—1—2—3 —4号外显子连接在一起,并利用外显子4后面的多聚(A)位点,产生降钙素mRNA。
位点,产生降钙素mRNA。")
147
若初级转录产物以0—1—2—3—5—6号外显子连接在一起,并利用外显子6以后的加多聚(A)位点时,就产生了编码与降钙素基因表达有关的多肽CGRP的mRNA 。
位点时,就产生了编码与降钙素基因表达有关的多肽CGRP的mRNA 。")
148
图8-51 前mRNA不同的剪接方式造成了不同组织中不同的降钙素样蛋白

149
4、mRNA 有效性的调控 mRNA的稳定性与蛋白质表达水平密切相关。

150
8.6.2 翻译水平的调控 1、 真核生物mRNA的“扫描模式”与蛋白质合成的起始 2、 mRNA 5′末端帽子结构的识别与蛋白质合成

151
3、 mRNA的稳定性与基因表达调控 高等生物中转铁蛋白受体(TfR)和铁蛋白负责铁吸收和铁解毒,这两个mRNA上存在相似的顺式作用元件,称为铁应答元件(iron response element,IRE)。
和铁蛋白负责铁吸收和铁解毒,这两个mRNA上存在相似的顺式作用元件,称为铁应答元件(iron response element,IRE)。")
152
研究表明,位于5’非翻译区的IRE控制了铁蛋白mRNA的翻译效率,去掉这个非翻译区IRE,可造成铁蛋白的永久性高水平翻译。
IRE与IRE结合蛋白(IREBP)相互作用控制了这两个mRNA的翻译效率。细胞中能产生两个数量级的蛋白水平差异,却没有在mRNA水平上发现显著差异。
相互作用控制了这两个mRNA的翻译效率。细胞中能产生两个数量级的蛋白水平差异,却没有在mRNA水平上发现显著差异。")
153
当细胞缺铁时,IREBP与IRE具有高亲和力,两者的结合有效地阻止了铁蛋白mRNA的翻译。此时,TfR mRNA上3’非翻译区中的IRE也与IREBP特异结合,有效地阻止TfR mRNA的降解,促进TfR蛋白的合成。

155
另外,催乳素能明显延缓酪蛋白(casein)mRNA的降解。
不加入催乳素时,体系中的酪蛋白mRNA在1小时内降解50%,而加入催乳素后40小时,酪蛋白mRNA才降解50%。

156
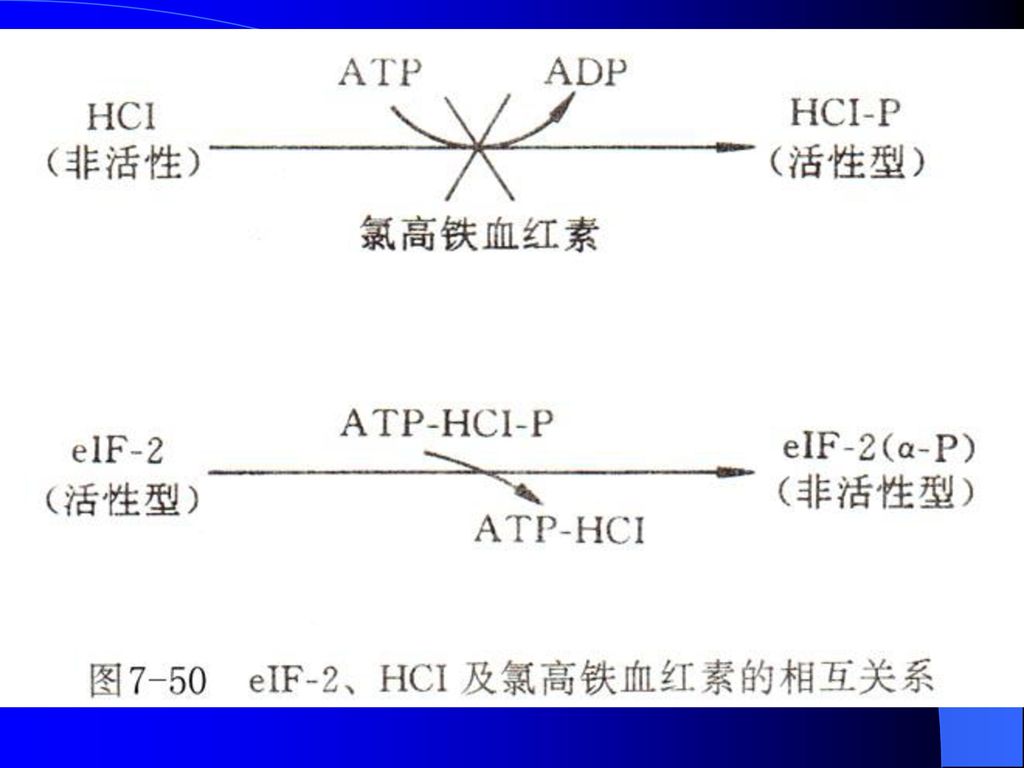
4、 可溶性蛋白因子的修饰与翻译起始调控 用兔网织红细胞粗抽提液研究蛋白质合成时发现,如果不向这一体系中添加氯高铁血红素,几分钟之内蛋白质合成活性急剧下降,直到完全消失。 研究表明,当没有氯高铁血红素存在时,网织红细胞粗抽提液中的蛋白质合成抑制剂就被活化,从而抑制蛋白质合成。

158
该抑制剂HCI受氯高铁血红素调节,是eIF-2的激酶,使eIF-2的α亚基磷酸化,并由活性型变成非活性型。没有生物活性的HCI也可以通过自身的磷酸化变成活性型。氯高铁血红素阻断了HCI的活化过程。

Similar presentations
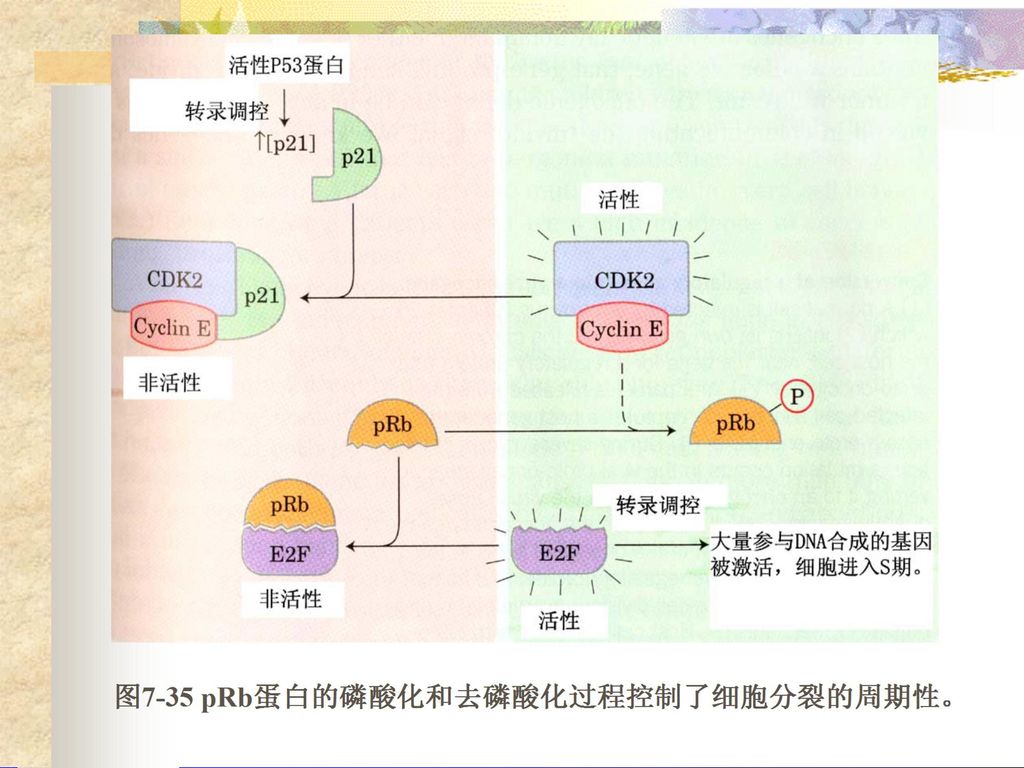
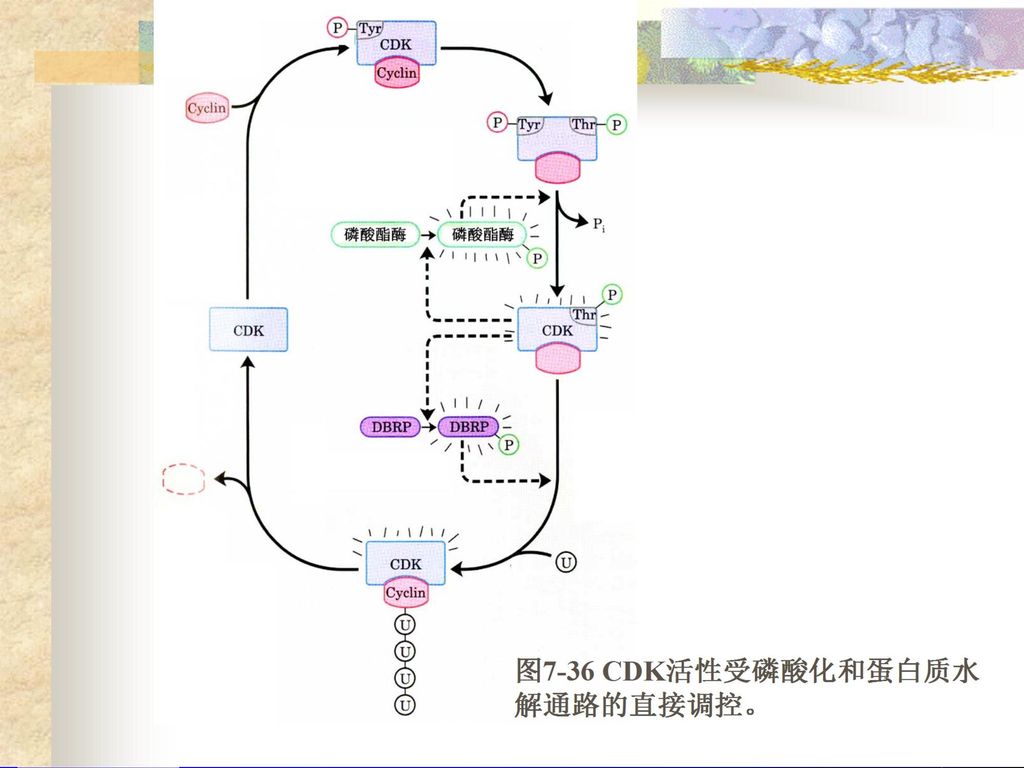
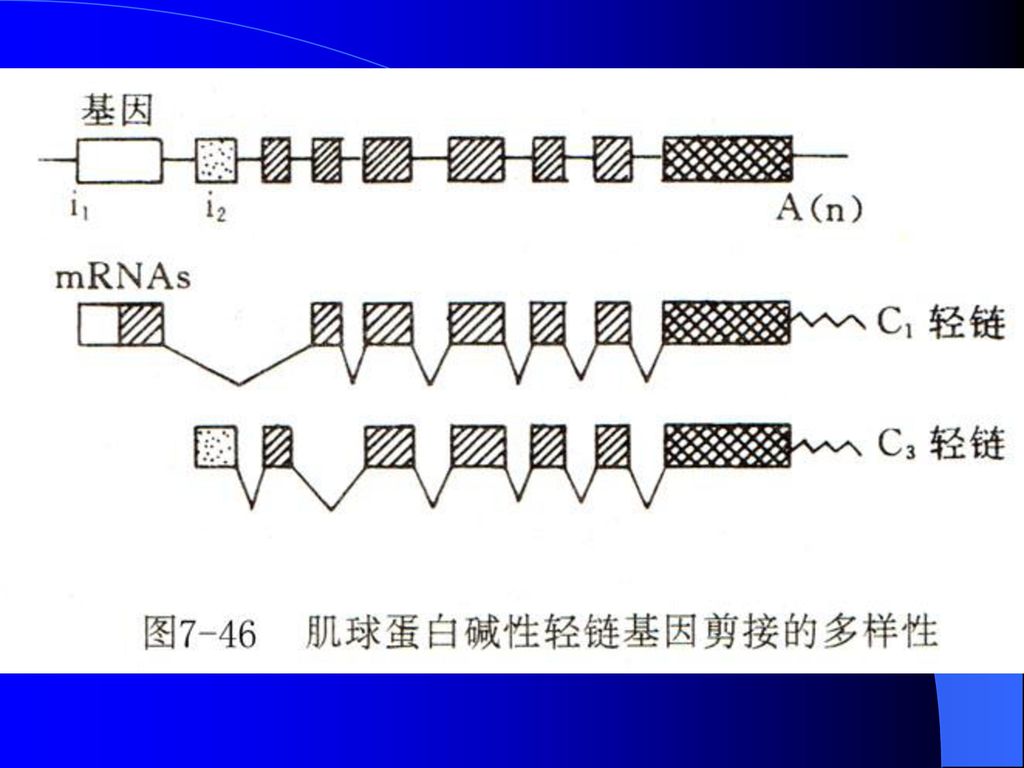
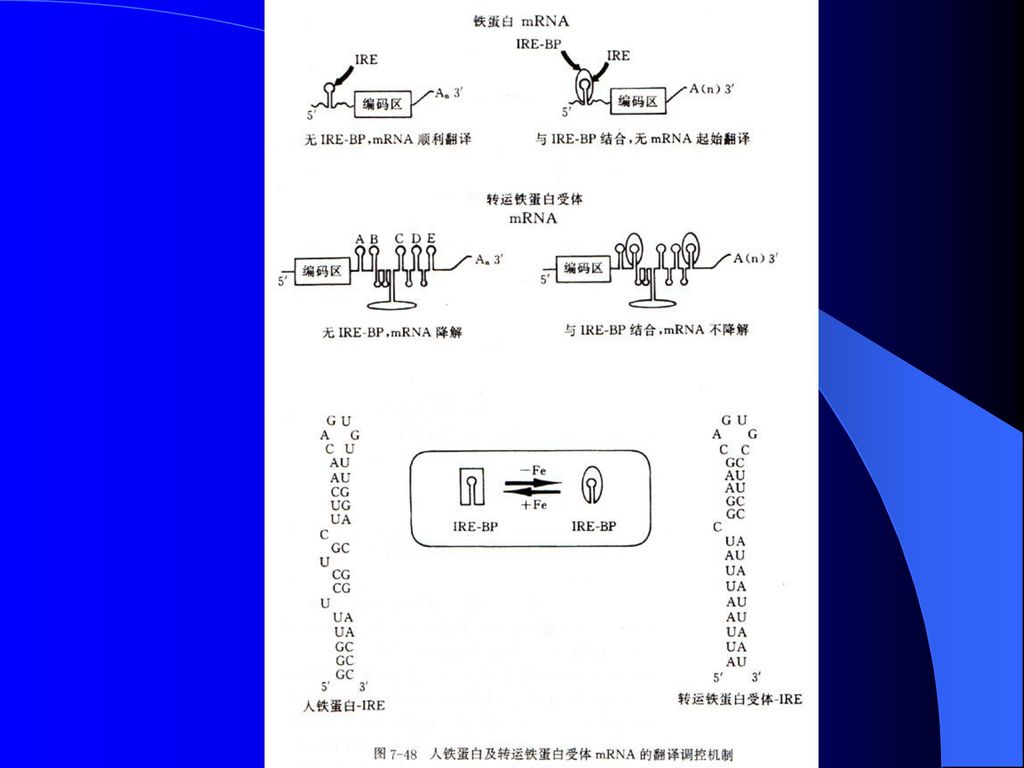
染色体结构不同; ( 2 )原核生物具有正调控和负调控并重的特点,真核 生物目前已知的主要是正调控; ( 3 )原核生物的转录和翻译是相偶联的,真核生物的.>")



.>")





是一群不受生长调控而繁殖的细胞,也称恶性肿瘤。良性肿瘤则是一群仅局限在自己的正常位置,且不侵染周围其它组织和器官的细胞。>")


>")


 Chapter 12 Protein Biosynthesis,Translation>")


