Download presentation
1
胺基酸的代謝 : 生合成 (biosynthesis of amino acid) 固氮作用 : 大氣中的氮在酵素催化下還原成氨 (NH 3 ) 的 作用,只有少數的原核微生物 (prokaryotic microorganism) 具有固氮作用的能力。
 固氮作用 : 大氣中的氮在酵素催化下還原成氨 (NH 3 ) 的 作用,只有少數的原核微生物 (prokaryotic microorganism) 具有固氮作用的能力。")
2
支鏈胺基酸 (BCAA:branched chain amino acids): 包含 leucine 、 isoleucine 與 valine 。在肝臟中大量的 NAA 與 EAA 被分解, 但 EAA 中的 BCAA 卻是例外。 BCAA 是肝臟 攜帶 amino group 至各組織中進行合成胺基 酸的物質。
: 包含 leucine 、 isoleucine 與 valine 。在肝臟中大量的 NAA 與 EAA 被分解, 但 EAA 中的 BCAA 卻是例外。 BCAA 是肝臟 攜帶 amino group 至各組織中進行合成胺基 酸的物質。")
3
轉胺反應 (transamination reaction): 將 -amino acid 的 -amino group 轉給 - keto acid 。 transamination 是一個可逆反應,是胺基酸 合成與分解時所必需的反應。
: 將 -amino acid 的 -amino group 轉給 - keto acid 。 transamination 是一個可逆反應,是胺基酸 合成與分解時所必需的反應。")
4
轉胺作用 (transamination)
")
5
胺基酸的生合成

6
氮的利用 : 1 植物無法進行固氮作用,需由土壤吸取 NH 4 + 然後用來合成胺基酸與核酸。 2 動物利用胺基酸上的胺基合成胺基酸與核 酸。 氮的循環 (nitrogen cycle): 各種生物體之間利用氮原子的循環。
: 各種生物體之間利用氮原子的循環。")
7
固氮作用 (nitrogen fixation) N N 鍵能為 225 kcal/mole 是很不活潑的氣 體。 工業固氮法是 Fritz Haber 在 1907-1909 年所 發明的,目前用在肥料的製造。反應式為 N 2 +3H 2 = 2NH 3 在鐵的催化、 500 0 C 高溫和 300atm 高壓力 下作用。
 N N 鍵能為 225 kcal/mole 是很不活潑的氣 體。 工業固氮法是 Fritz Haber 在 年所 發明的,目前用在肥料的製造。反應式為 N 2 +3H 2 = 2NH 3 在鐵的催化、 C 高溫和 300atm 高壓力 下作用。")
8
自然界只有少數的原核生物具有固氮能力, 如一些共生的根瘤菌 (Rhizobium) 。在豆科 植物的根部有一些寄生在根部的根瘤菌能 合成 NH 4 + 供植物利用,但豆科植物必需附 出相當代價 : (1) 在根瘤內需維持非常低的 O 2 (2) 耗用植物約 1/5 的 ATP 。
 。在豆科 植物的根部有一些寄生在根部的根瘤菌能 合成 NH 4 + 供植物利用,但豆科植物必需附 出相當代價 : (1) 在根瘤內需維持非常低的 O 2 (2) 耗用植物約 1/5 的 ATP 。")
9
圖 14.1 在酵素性氮固定作用中雙氮酶複 合物的模式圖說明電子流動和能量

10
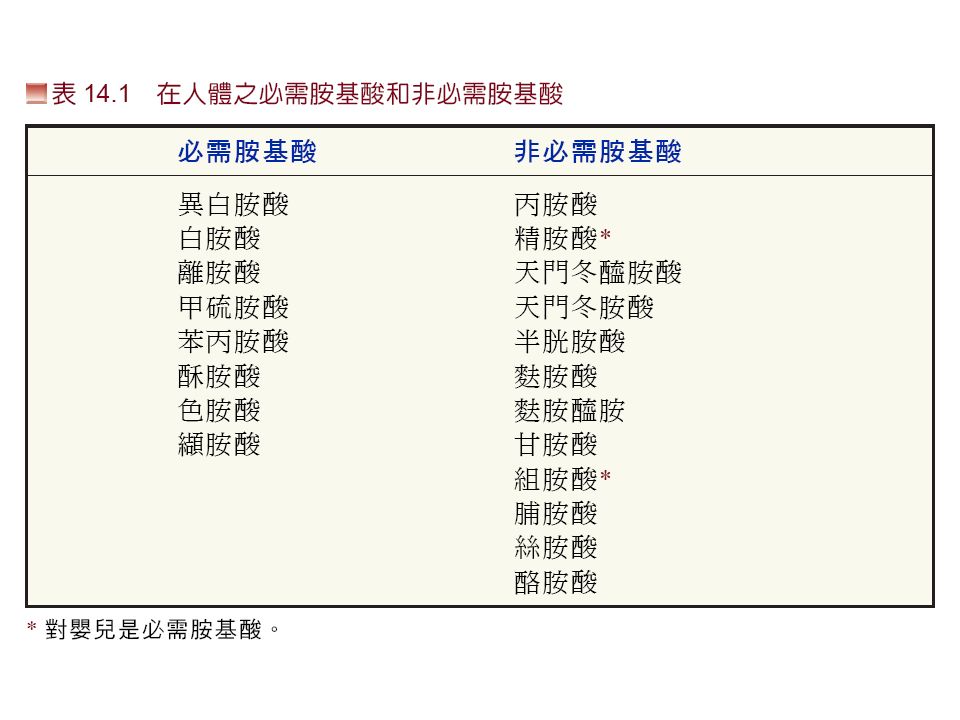
胺基酸生合成 (amino acid biosynthesis): 植物與微生物可以自行合成全部的胺基酸。 哺乳動物 : 僅能合成一些少數的非必需胺基 酸 (NAA) , EAA 必需由食物供給。
: 植物與微生物可以自行合成全部的胺基酸。 哺乳動物 : 僅能合成一些少數的非必需胺基 酸 (NAA) , EAA 必需由食物供給。")
12
氮平衡 (nitrogen balance): 當吃進的氮源 ( 主要是胺基酸 ) 與排出的氮源相等時稱氮平 衡。這是健康成人的情況。 Positive nitrogen balance: 吃入的氮源較 排出的氮源多。這是生長中的小孩、孕婦 與康復中的病人的狀況。 Negative nitrogen balance: 食物中匱乏蛋 白質無法補充排出的氮源。
: 當吃進的氮源 ( 主要是胺基酸 ) 與排出的氮源相等時稱氮平 衡。這是健康成人的情況。 Positive nitrogen balance: 吃入的氮源較 排出的氮源多。這是生長中的小孩、孕婦 與康復中的病人的狀況。 Negative nitrogen balance: 食物中匱乏蛋 白質無法補充排出的氮源。")
13
Kwashiorkor: 是一種惡性的營養缺乏症,肇因於長期蛋 白的攝取不足。症狀包括 : 生長停滯,肝腫 大、潰瘍、下痢、心臟、腎臟功能受損等。 流行於非洲。治療方法 : 大量餵食高蛋白物 質,如牛奶、雞蛋與肉類。

14
Transport: 輸送胺基酸進入細胞是由膜上特異的輸送 蛋白負責,至少分成兩類 : 1 需鈉離子的輸送 (Na+-dependent amino acid transport) 是一種 second active transport system 。 2 不需鈉離子的輸送 : 如 -glutamyl cycle 。
 是一種 second active transport system 。 2 不需鈉離子的輸送 : 如 -glutamyl cycle 。")
15
胺基的反應 : 1 轉胺作用 (transamination): 反應由 aminotransferase 催化 胺基酸的合成大部分以 glutamate 充當 amino group donor 。 有三個轉胺作用在代謝上具有重要的功能。 1 -ketoglutarate/glumate pair 在胺基酸合成與分 解反應 2 oxaloacetate/asparate pair 在 urea cycle 3 pyruvate/alanine pair 在 alanine cycle
: 反應由 aminotransferase 催化 胺基酸的合成大部分以 glutamate 充當 amino group donor 。 有三個轉胺作用在代謝上具有重要的功能。 1 -ketoglutarate/glumate pair 在胺基酸合成與分 解反應 2 oxaloacetate/asparate pair 在 urea cycle 3 pyruvate/alanine pair 在 alanine cycle")
16
胺基酸的生合成

17
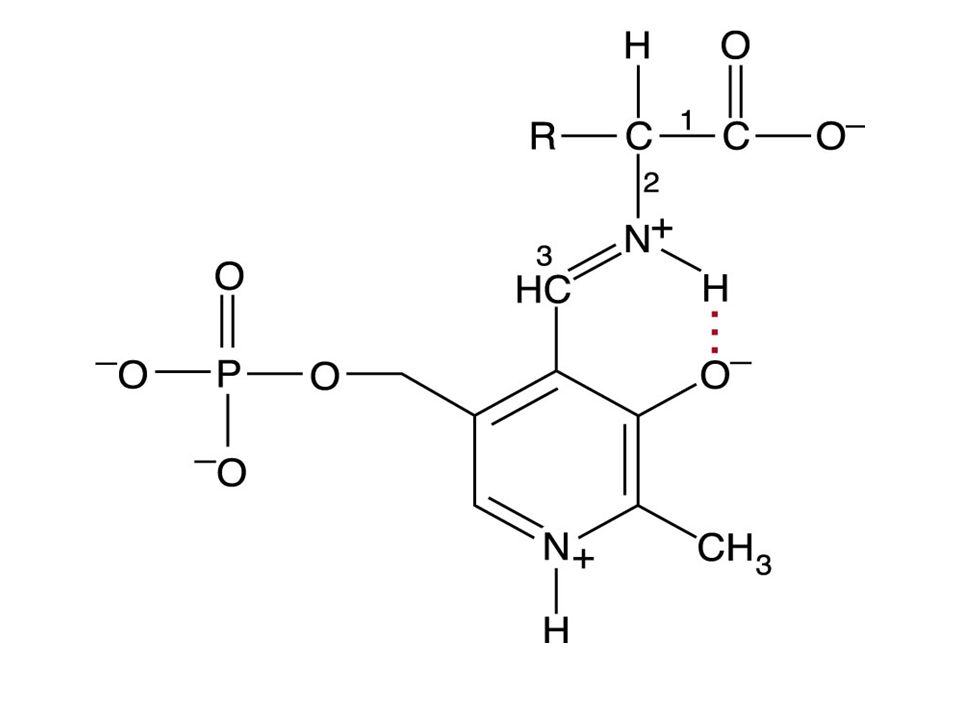
圖 14.2 維生素 B 6

18
Fig. 14.2

20
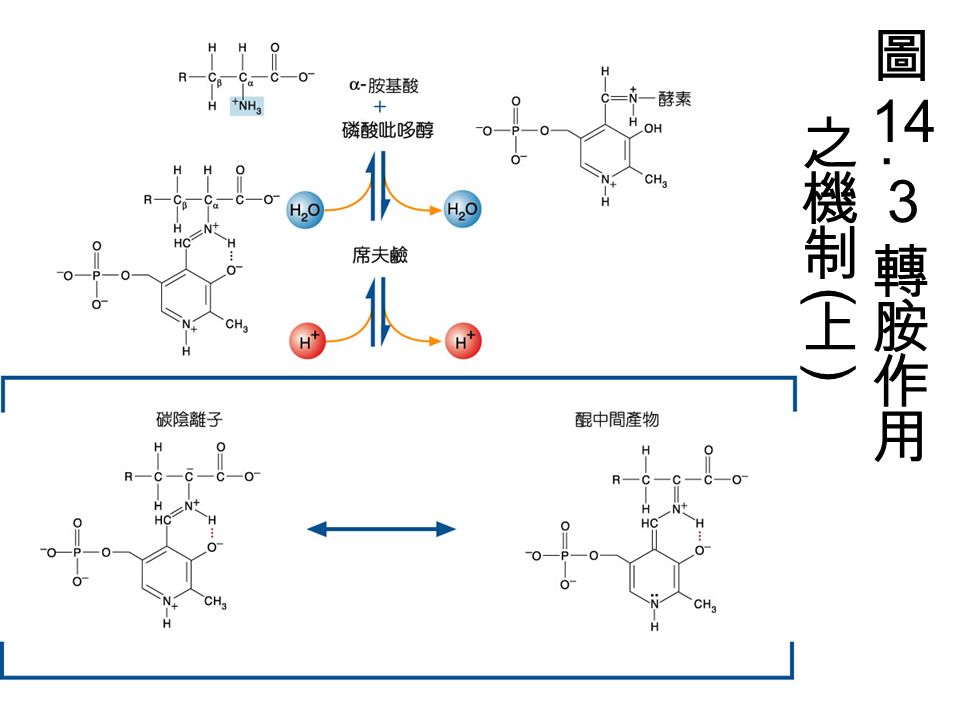
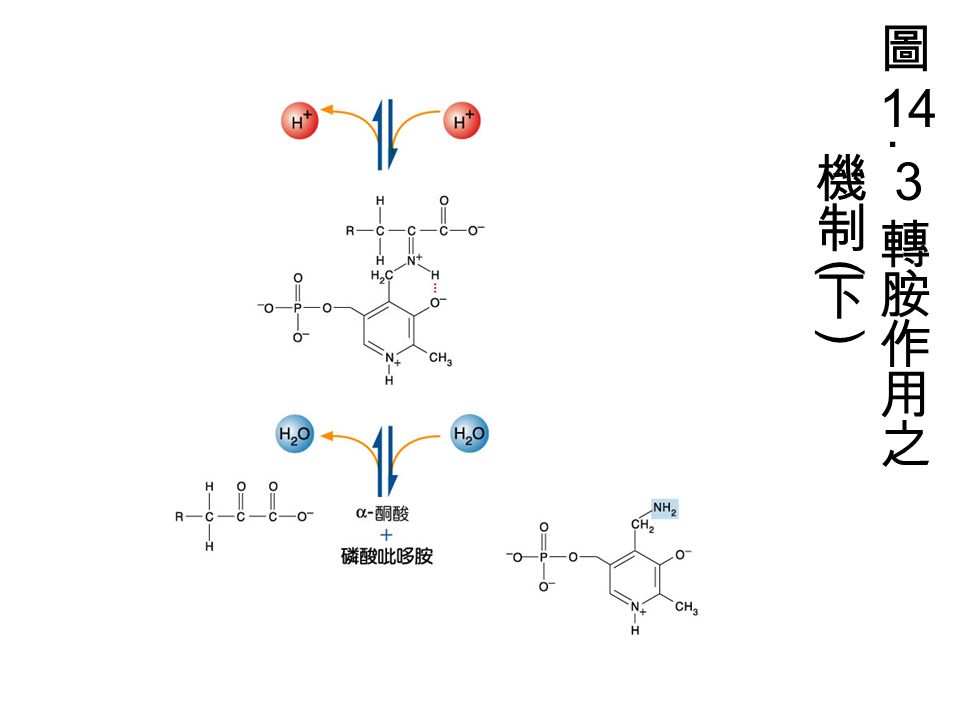
pyridoxal-5-phosphate(PLP): 由 pyridoxine(Vitamin B 6 ) 組成是轉胺作用的 coenzyme 。需 PLP 的酵素中, PLP 是接在 Lysine residue 上。 PLP 與胺基酸上的 amino group 結合形成中間物 (intermediate)( 如圖 ) ,形成 Schiff base
: 由 pyridoxine(Vitamin B 6 ) 組成是轉胺作用的 coenzyme 。需 PLP 的酵素中, PLP 是接在 Lysine residue 上。 PLP 與胺基酸上的 amino group 結合形成中間物 (intermediate)( 如圖 ) ,形成 Schiff base")
23
2 胺離子 (NH 4 + ) 併入有機分子中 : 分兩類 (1) 對 -keto acid 還原加胺作用 :Glutamate dehydrogenase 催化 -ketoglutarate 加胺作 用。此方應具可逆性,但真核生物中似乎 傾向於 NH 4 + 的釋出。 (2) 使 Asparate 與 glutamate 轉成 asparagine 與 glutamine 在腦中存有豐富的 glutamine synthetase ,因腦對 NH 4 + ) 的毒性特別敏感。
 併入有機分子中 : 分兩類 (1) 對 -keto acid 還原加胺作用 :Glutamate dehydrogenase 催化 -ketoglutarate 加胺作 用。此方應具可逆性,但真核生物中似乎 傾向於 NH 4 + 的釋出。 (2) 使 Asparate 與 glutamate 轉成 asparagine 與 glutamine 在腦中存有豐富的 glutamine synthetase ,因腦對 NH 4 + ) 的毒性特別敏感。")
24
The biosynthetic pathways to glutamate and glutamine 1.Glutamine synthetase catalyzes the reaction of glutamine and NH 4 + to yield glutamine. Glutamate + ATP → -glutamyl phosphate + ADP -glutamyl phosphate + NH 4 + → glutamine + Pi + H + Sum: Glutamate + NH 4 + +ATP → Glutamine + ADP + Pi + H + Glutamine synthetase is found in all organisms.

26
3. L-glutamate dehydrogenase -Ketoglutarate + NH 4 + +NADPH → L-Glutamate + NADP + + H 2 O In eukaryotic cells, L-glutamate dehydrogenase is located in the mitochondrial matrix. The reaction equilibrium favors reactants, and the Km for NH 4 + (about 1 mM) is so high that the reaction probably makes only a modest contribution to NH 4 + assimilation into amino acids and other metabolites
 is so high that the reaction probably makes only a modest contribution to NH 4 + assimilation into amino acids and other metabolites.")
27
Glutamine synthetase is a primary regulatory point in nitrogen metabolism In E. coli, glutamine synthetase has 12 identical subunits of Mr 50,000 and is regulated both allosterically and by covalent modification. Alanine, glycine,and at least six end products of glutamine metabolism are allosteric inhibitors of the enzyme. Each inhibitor alone produces only partial inhibition, but the effects of multiple inhibitors are move than additive, and all eight together virtually shut down the enzyme.

28
Allosteric regulation of glutamine synthetase

29
X-ray crystal structure of glutamine synthetase

30
Regulation of glutamine synthetase by covalent modification Adenylylation of Tyr 397 of glutamine synthetase increases sensitivity to the allosteric inhibitors, and activity decreases as more subunits are adenylylated. Both adenylylation and deadenylylation are promoted by adenylyltransferase (AT). The activity of AT is modulated by binding to a regulatory protein called P II, and the activity of P II, in turn, is regulated by covalent modification (uridylylation), again at a Tyr residue. The AT complex with P II -UMP stimulates deadenylylation, where the same complex with P II stimulated adenylylation of glutamine synthetase. Both uridylylation and deuridylylation of P II are brought about by a single enzyme, uridylyltransferase (UT). Uridylylation is inhibited by binding of glutamine and Pi to UT and is stimulated by binding of -ketoglutarate and ATP to P II.
. The activity of AT is modulated by binding to a regulatory protein called P II, and the activity of P II, in turn, is regulated by covalent modification (uridylylation), again at a Tyr residue. The AT complex with P II -UMP stimulates deadenylylation, where the same complex with P II stimulated adenylylation of glutamine synthetase. Both uridylylation and deuridylylation of P II are brought about by a single enzyme, uridylyltransferase (UT). Uridylylation is inhibited by binding of glutamine and Pi to UT and is stimulated by binding of -ketoglutarate and ATP to P II..")
31
The uridylylated P II also mediates the activation of transcription of the gene encoding glutamine synthetase, thus increasing the cellular concentration of the enzyme; the deuridylylated P II brings about a decrease in transcription of the same gene.

32
An adenylylated Tyr residue of glutamine synthetase

33
Cascade leading to adenylylation of glutamine synthetase

34
(2) 使 Asparate 與 glutamate 轉成 asparagine 與 glutamine 在腦中存有豐富的 glutamine synthetase ,因腦對 NH 4 + ) 的毒性特別敏感 glutamine synthetase 催化之反應 : glutamate + ATP + NH 4 + glutamine + ADP + P i Asparagine 的合成,需 Asparagine synthase
 使 Asparate 與 glutamate 轉成 asparagine 與 glutamine 在腦中存有豐富的 glutamine synthetase ,因腦對 NH 4 + ) 的毒性特別敏感 glutamine synthetase 催化之反應 : glutamate + ATP + NH 4 + glutamine + ADP + P i Asparagine 的合成,需 Asparagine synthase")
36
胺基酸的合成 (synthesis of the amino acid) 在動物中所有的 NAA 是由 glycerate-3- phosphate 、 pyruvate 、 -ketoglutarate 或 oxaloacetate 合成而來。另外 tyrosine 是 由 phenylalanine 合成而來。
 在動物中所有的 NAA 是由 glycerate-3- phosphate 、 pyruvate 、 -ketoglutarate 或 oxaloacetate 合成而來。另外 tyrosine 是 由 phenylalanine 合成而來。")
37
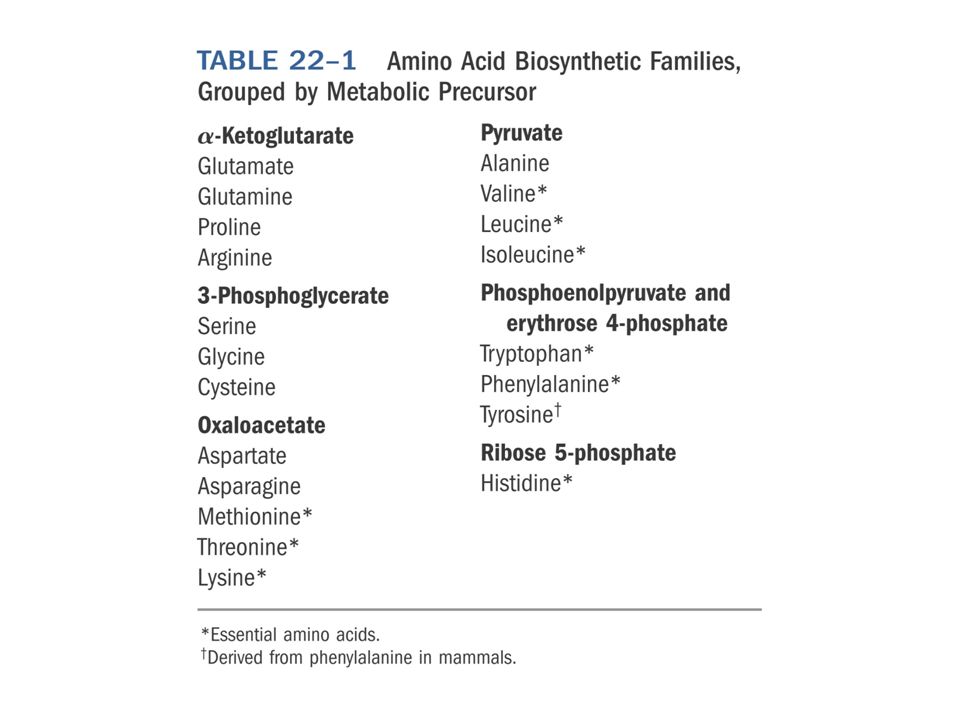
Biosynthesis of amino acids All amino acids are derived from intermediates in glycolysis, the citric acid cycle, or the pentose phosphate pathway. Nitrogen enters these pathways by way of glutamate and glutamine. Whereas most bacteria and plants can synthesize all 20 amino acids, mammals can synthesize only about half of them. These are the nonessential amino acids, not needed in the diet. The remainder, the essential amino acids, must be obtained from food.

38
Overview of amino acid biosynthesis The carbon skeleton precursors derived from three sources: glycolysis (pink), the citric acid cycle (blue), and the pentose phosphate pathway (purple).
, the citric acid cycle (blue), and the pentose phosphate pathway (purple).")
41
Serine, glycine, and cysteine are derived from phosphoglycerate

42
Biosynthesis of serine from 3- phosphoglycerate and of glycine from serine in all organisms In the liver of vertebrates, glycine can be made by another route: the reaction catalyzed by Glycine synthase (also called glycine cleavage enzyme): CO 2 + NH4 + + N 5,N 10 -methylene tetrahydrofolate + NADH + H + → Glycine + tetrahydrofolate + NAD +
: CO 2 + NH4 + + N 5,N 10 -methylene tetrahydrofolate + NADH + H + → Glycine + tetrahydrofolate + NAD +")
43
Biosynthesis of cysteine from homocysteine and serine in mammals

44
Three nonessential and six essential amino acids are synthesized from oxaloacetate and pyruvate

45
Chorismate is a key intermediate in the synthesis of tryptophan, phenylalanine, and tyrosine

46
Concerted Inhibition Six products derived from glutamine serve as negative feedback modulators of the enzyme, and the overall effects of these and other modulators are more than additive. Such regulation is called Concerted inhibition.

47
Fig. 14.4

48
胺基酸的生合成途徑有一個共同特性 : 它們 的碳骨架是由 glycolysis 、 pentose phosphate pathway 與 citric acid cycle 的中 間產物而來,可分為 6 個族群。 1 Glutamate family: 以 -ketoglutarate 為先 驅物,合成 glutamate 、 glutamine 、 proline 與 arginine 。

49
Fig. 14.5

50
2 Serine family: 以 glycerate-3-phosphate 為先驅物,合成 serine 、 glycine 與 cysteine 這一族的成員在生合成 (anabolism) 中扮演 重要的角色。
 中扮演 重要的角色。")
51
Fig. 14.6

52
Glycine 是合成嘌呤 (purine) 、 porphyrin( 紫 質 ) 、 glutathione 的先驅物 (precursor) 。 Serine 是合成 ethanolamine 與 sphingosine 的先驅物。 cysteine 在硫代謝中具重要的角色。
 、 porphyrin( 紫 質 ) 、 glutathione 的先驅物 (precursor) 。 Serine 是合成 ethanolamine 與 sphingosine 的先驅物。 cysteine 在硫代謝中具重要的角色。")
53
Fig. 14.7

54
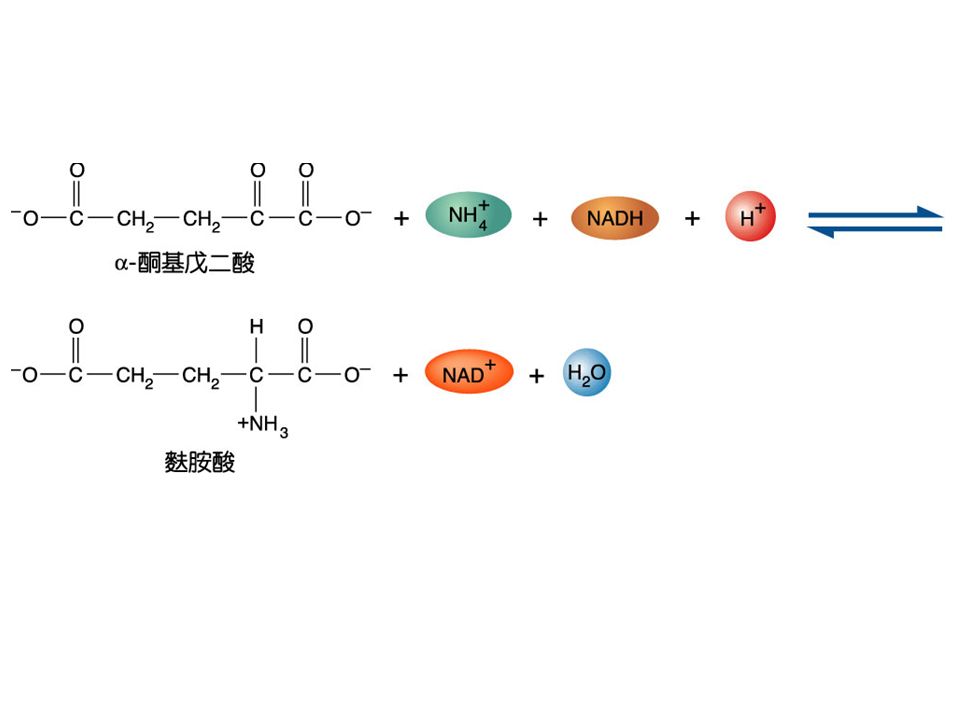
( 三 )Asparate 家族 : 以 oxaloacetate 為先驅物, 合成 aspartate 、 asparagine 、 lysine 、 methionine 與 threonine 。 Asparate 的合成 : glutamate + oxaloacetate aspartate + -ketoglutarate 此反應是由 asparate transaminase 催化又稱 GOT(glutamic oxaloacetic transaminase
Asparate 家族 : 以 oxaloacetate 為先驅物, 合成 aspartate 、 asparagine 、 lysine 、 methionine 與 threonine 。 Asparate 的合成 : glutamate + oxaloacetate aspartate + -ketoglutarate 此反應是由 asparate transaminase 催化又稱 GOT(glutamic oxaloacetic transaminase")
55
Fig. 14.8

56
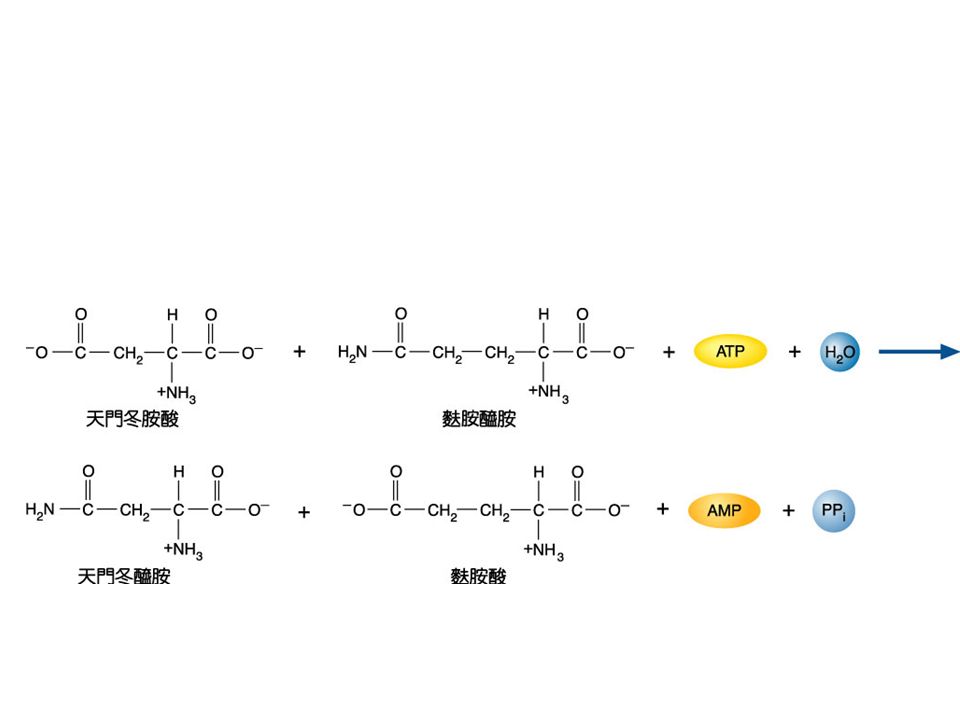
Asparagine 的合成 : asparagine + glutamine +ATP +H2O Asparagine + glutamate +AMP + PPi Asparagine 並不是直接由 aspartate 與 NH 4 + 合成,而經由 glutamine 轉胺作用形成, 這個反應耗掉約 2ATP 。

57
( 四 )pyruvate 家族 : 以 pyruvate 為先驅物,包 含 alanine 、 valine 、 leucine 與 isoleucine alanine 是由 pyruvate 進行轉胺作用合成的 glutamate + pyruvate alanine + - ketoglutarate 催化此反應的酵素為 alanine transaminase( 又稱 glutamic pyruvic transaminase:GPT)
pyruvate 家族 : 以 pyruvate 為先驅物,包 含 alanine 、 valine 、 leucine 與 isoleucine alanine 是由 pyruvate 進行轉胺作用合成的 glutamate + pyruvate alanine + - ketoglutarate 催化此反應的酵素為 alanine transaminase( 又稱 glutamic pyruvic transaminase:GPT)")
58
Fig. 14.9

59
( 五 )Aromatic 家族 : 是由 phosphoenolpyruvate 與 erythrose-4- phosphate 起始合成,包含 phenylalanine 、 tyrosine 與 tryptophan 。而 tyrosine 是 phenylalanine 合成而得。 chorismate: 可合成 phenylalanine 、 tyrosine 與 tryptophan 。若加入 terpenoids 則可合成 tocopherol 或 ubiquinone 。
Aromatic 家族 : 是由 phosphoenolpyruvate 與 erythrose-4- phosphate 起始合成,包含 phenylalanine 、 tyrosine 與 tryptophan 。而 tyrosine 是 phenylalanine 合成而得。 chorismate: 可合成 phenylalanine 、 tyrosine 與 tryptophan 。若加入 terpenoids 則可合成 tocopherol 或 ubiquinone 。")
60
Fig. 14.10

61
Fig. 14.11

62
( 六 )Histidine: 在健康的成人是屬於 NAA , 是由 phosphoribosylpyrophosphate(PRPP) , ATP 與 glutamine 所合成。
Histidine: 在健康的成人是屬於 NAA , 是由 phosphoribosylpyrophosphate(PRPP) , ATP 與 glutamine 所合成。")
63
Fig. 14.12

64
單碳的代謝 (one-carbon metabolism): 在生合成途徑 (biosynthetic pathway) 中,最 重要單碳的攜帶者 (one carbon carrier) 包含 (1) 葉酸 (folic acid) (2)S-Adenosylmethionine 。 另外 Vit B 12 也有此功能。
: 在生合成途徑 (biosynthetic pathway) 中,最 重要單碳的攜帶者 (one carbon carrier) 包含 (1) 葉酸 (folic acid) (2)S-Adenosylmethionine 。 另外 Vit B 12 也有此功能。")
65
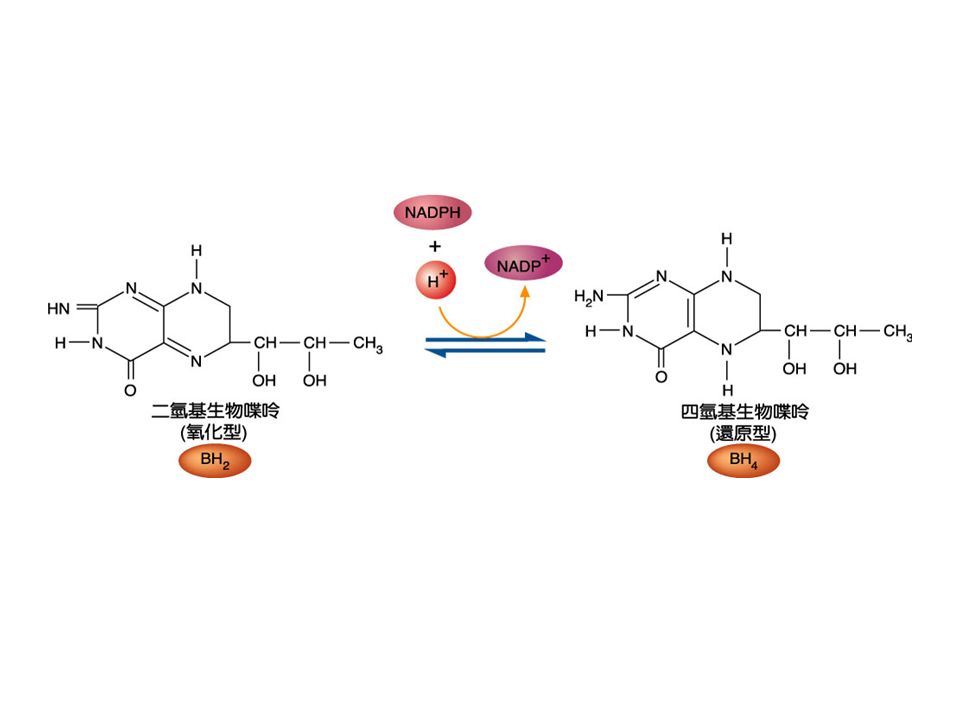
Folic acid: 分子包含一個 pteridine nucleus 與 para-aminobenzoic acid ,然後接在 glutamic acid 上。 四氫葉酸 (tetrahydrofolic acid:THF) 是生物 活性的形式。能攜帶 methyl 、 methylene 、 methenyl 與 formyl group ,結合位置在 pteridine ring 的 N 5 與 N 10 的位置。 Folic acid 可經由 dihydrofolate reductase 的 催化還原成 THF , NADPH 是輔因子。
 是生物 活性的形式。能攜帶 methyl 、 methylene 、 methenyl 與 formyl group ,結合位置在 pteridine ring 的 N 5 與 N 10 的位置。 Folic acid 可經由 dihydrofolate reductase 的 催化還原成 THF , NADPH 是輔因子。")
66
Fig. 14.13

67
Table 14.2

68
Fig. 14.14

69
Table 14.3

70
Fig. 14.15

71
S-adenosylmethionine(SAM): 單碳的代謝 中, SAM 是甲烷基的供應者 (methyl group donor) 。由 ATP 與 methionine 在 SAM synthase 催化下形成。 SAM 失去一個 -CH 3 後會形成 S- Adenosylhomocysteine(SAH) ,可經由 N 5 - methyl THF 補充。若 methionine 缺乏時也 可利用 choline 的甲烷基加入 homocysteine 成 methionine 。
: 單碳的代謝 中, SAM 是甲烷基的供應者 (methyl group donor) 。由 ATP 與 methionine 在 SAM synthase 催化下形成。 SAM 失去一個 -CH 3 後會形成 S- Adenosylhomocysteine(SAH) ,可經由 N 5 - methyl THF 補充。若 methionine 缺乏時也 可利用 choline 的甲烷基加入 homocysteine 成 methionine 。")
72
Fig. 14.16

73
Vitamine B 12 (cobalamin): 是一個含鈷的分 子,鈷離子上接一個氰胺 (cyanide) ,僅少 數的微生物能合成。動物可從小腸的菌叢 中獲得或從肝臟、雞蛋、蝦與肉類等食物 中獲得。
: 是一個含鈷的分 子,鈷離子上接一個氰胺 (cyanide) ,僅少 數的微生物能合成。動物可從小腸的菌叢 中獲得或從肝臟、雞蛋、蝦與肉類等食物 中獲得。")
74
Fig. 14.17

75
惡性貧血 (pernicious anemia):Vitamin B 12 缺乏會導致嚴重的病症,症狀除了低紅血 球數量,還包括虛弱與中樞神經受損。 1926 年三位美國醫生發現服用肝臟可冶療 惡性貧血 ( 只發生在人類 ) ,才揭開 Vit B12 的 神秘面紗, 1956 年 Hohgkin 女士解出其三度 空間結構。
:Vitamin B 12 缺乏會導致嚴重的病症,症狀除了低紅血 球數量,還包括虛弱與中樞神經受損。 1926 年三位美國醫生發現服用肝臟可冶療 惡性貧血 ( 只發生在人類 ) ,才揭開 Vit B12 的 神秘面紗, 1956 年 Hohgkin 女士解出其三度 空間結構。")
76
造成惡性貧血的原因如下 : 1 大部分是因為一個糖蛋白「具 Vit B 12 吸收 因子 (intrinsic factor) 」缺乏造成 2 胃腸病造成 Vit B 12 吸收被抑制。 3 大量使用抗生素造成腸內益菌減少, Vit B 12 吸收減少。
 」缺乏造成 2 胃腸病造成 Vit B 12 吸收被抑制。 3 大量使用抗生素造成腸內益菌減少, Vit B 12 吸收減少。")
77
Glutathione(GSH): -Glutamylcysteinylglycine:GSH 的功能 如下 : 1 幫助合成生物分子,如 eicosanoids 、 DNA 與 RNA 等。 ( 把催化反應的酵素變成還原態 ) 。 2 抗氧化劑 (Antioxidant): 抗 oxidative stress 的分子,阻止細胞膜被破壞。 3 在細胞間輸送胺基酸特別是 cysteine 與 methionine 。 4 形成 GSH conjugation ,以利一些物質排 出。
: -Glutamylcysteinylglycine:GSH 的功能 如下 : 1 幫助合成生物分子,如 eicosanoids 、 DNA 與 RNA 等。 ( 把催化反應的酵素變成還原態 ) 。 2 抗氧化劑 (Antioxidant): 抗 oxidative stress 的分子,阻止細胞膜被破壞。 3 在細胞間輸送胺基酸特別是 cysteine 與 methionine 。 4 形成 GSH conjugation ,以利一些物質排 出。")
78
Fig. 14.18

79
Fig. 14.19

80
-Glutamyl cycle: 存在一些組織中,具主 動運輸 (active transport) 數個胺基的特性 ( 特 別是 cysteine 與 methionine) 。
 數個胺基的特性 ( 特 別是 cysteine 與 methionine) 。")
81
神經傳導物質 (neurotransmitters): 具有激發性 (excitatory) 與抑制性 (inhibitory) 效應 : Excitatory neurotransmitters: 如乙醯膽鹼 (acetylcholine) 或 glutamate 等,能促進被傳 導的細胞膜去極化 (depolarization) ,能增加 action potential 。 足量的去極化反應 ( 即達到 action potential 的門檻 ) ,能啟動神經傳導物質的釋出。
: 具有激發性 (excitatory) 與抑制性 (inhibitory) 效應 : Excitatory neurotransmitters: 如乙醯膽鹼 (acetylcholine) 或 glutamate 等,能促進被傳 導的細胞膜去極化 (depolarization) ,能增加 action potential 。 足量的去極化反應 ( 即達到 action potential 的門檻 ) ,能啟動神經傳導物質的釋出。")
82
Table 14.4

86
Inhibitory neurotransmitters: 如 glycine 與 GABA ,會造成 Repolarization 即抑制 action potential 的形成。 許多神經傳導物質胺基酸或其衍生物 (derivatives) 後者又稱 biogenic amines 。
 後者又稱 biogenic amines 。")
87
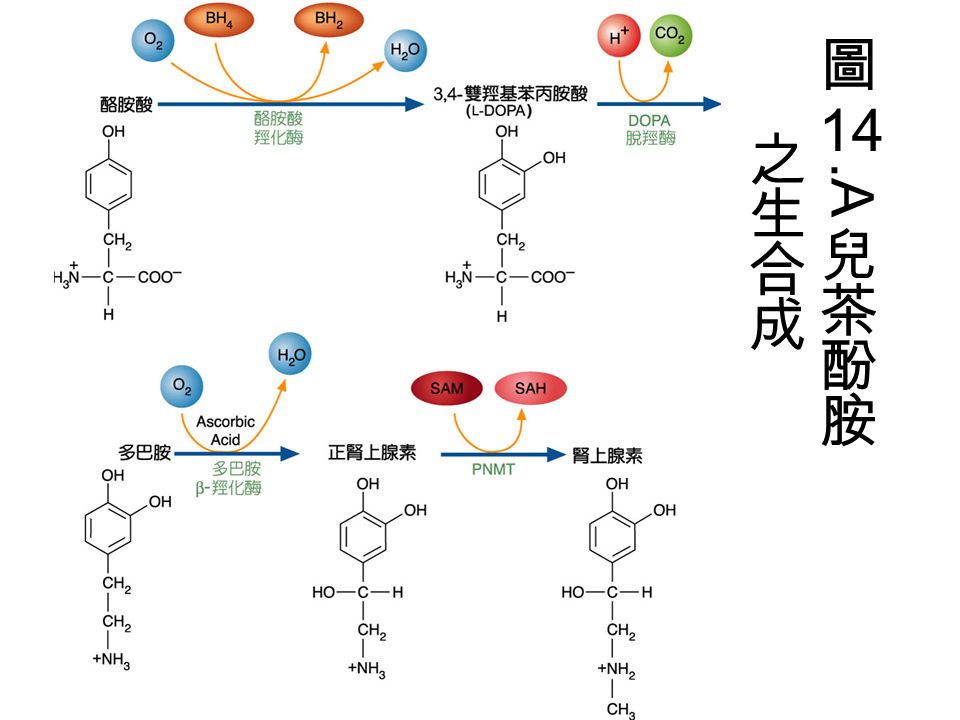
兒茶酚胺 (catecholamine): 包括多巴明 (dopamine) 、正腎上腺素 (nonepinephrine) 與腎上腺素 (epinephrine) ,是酪胺酸 (tyrosine) 的衍生物。在腦中是屬於激發性 的神經傳導物質。
: 包括多巴明 (dopamine) 、正腎上腺素 (nonepinephrine) 與腎上腺素 (epinephrine) ,是酪胺酸 (tyrosine) 的衍生物。在腦中是屬於激發性 的神經傳導物質。")
88
N (nonepinephrine) 與 NE (epinephrine) 皆具有調 控代謝作用的功能。 正腎上腺素 : 是腎上腺髓質的嗜鉻細胞 (chromaffin cells) 分泌,能促進三酸甘油脂與肝糖的分解,在 驚嚇、遇冷或運動與低血糖時分泌。 腎上腺素 : 是腎上腺髓質的嗜鉻細胞 (chromaffin cells) 分泌,在受壓力、創傷或激烈運動時,能促 進三酸甘油脂與肝糖的分解。兩者皆是激素 (hormone)
 與 NE (epinephrine) 皆具有調 控代謝作用的功能。 正腎上腺素 : 是腎上腺髓質的嗜鉻細胞 (chromaffin cells) 分泌,能促進三酸甘油脂與肝糖的分解,在 驚嚇、遇冷或運動與低血糖時分泌。 腎上腺素 : 是腎上腺髓質的嗜鉻細胞 (chromaffin cells) 分泌,在受壓力、創傷或激烈運動時,能促 進三酸甘油脂與肝糖的分解。兩者皆是激素 (hormone)")
89
L-DOPA: dopamine 是由 L-DOPA 合成而得, 因為 dopamine 無法通過 blood-brain barrier( 是一種障壁,用來阻止毒物進入腦 內 ) ,用來冶療巴金森氏症 (Parkinson’s disease) 。
 ,用來冶療巴金森氏症 (Parkinson’s disease) 。")
90
巴金森氏症 (Parkinson’s disease): 以震顫 麻痺 (paralysis agitans) 而聞名,是一種腦 部受損造成移動失調,常出現在 40 歲以上 的成人。症狀 : 為顫抖、肌肉僵硬、移動囷 難,目前原因並不清楚。只知在腦的黑質 (substantia nigra) 部位的一些神經細胞 (neuron) 無法產生多巴明 (dopamine) 可能是 造成此疾病的原因。
: 以震顫 麻痺 (paralysis agitans) 而聞名,是一種腦 部受損造成移動失調,常出現在 40 歲以上 的成人。症狀 : 為顫抖、肌肉僵硬、移動囷 難,目前原因並不清楚。只知在腦的黑質 (substantia nigra) 部位的一些神經細胞 (neuron) 無法產生多巴明 (dopamine) 可能是 造成此疾病的原因。")
91
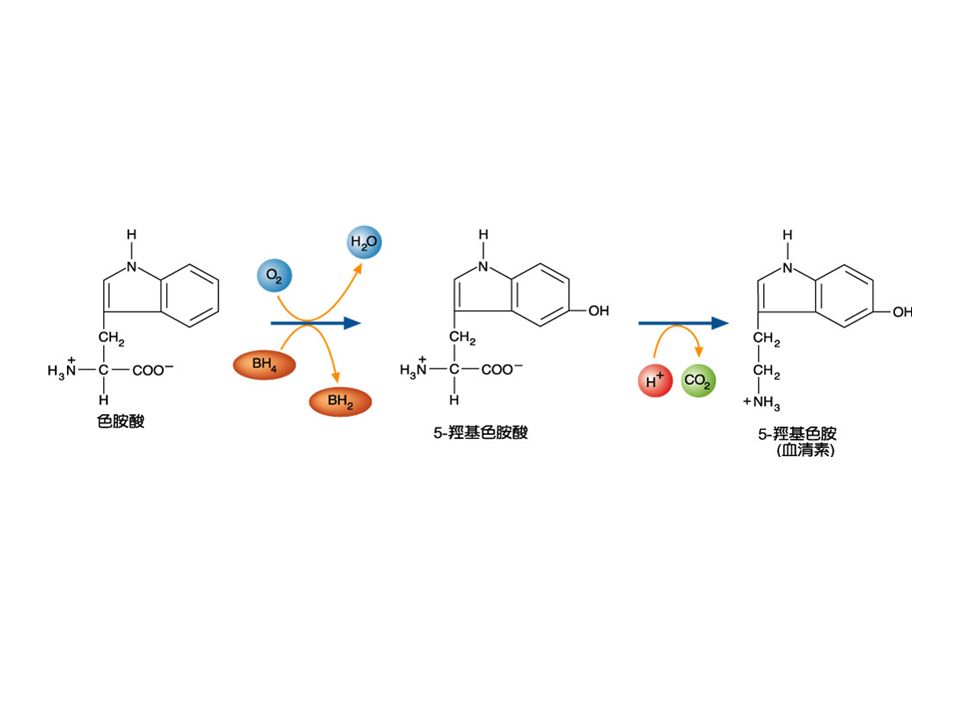
組織胺 (histamine): 是一個胺類,許多組織都可以產生,它是 過敏 (allergic) 與發炎 (inflammatory) 反應的 媒介物 (mediator) ,可剌激胃酸的產生等。 由 L-histidine 失去二氧化碳 (decarboxylation) 而得。此反應是由 histidine decarboxylase 催化, PLP 是其輔因子。抗組織胺的藥物常 用來冶療過敏性鼻炎、蕁麻疹等。但會有 嗜唾、疲勞副作用。
: 是一個胺類,許多組織都可以產生,它是 過敏 (allergic) 與發炎 (inflammatory) 反應的 媒介物 (mediator) ,可剌激胃酸的產生等。 由 L-histidine 失去二氧化碳 (decarboxylation) 而得。此反應是由 histidine decarboxylase 催化, PLP 是其輔因子。抗組織胺的藥物常 用來冶療過敏性鼻炎、蕁麻疹等。但會有 嗜唾、疲勞副作用。")
92
一氧化氮 (nitric oxide): 是一個自由基,非常活潑的氣體,生命期很短, 但扮演許多重要的生理角色。許多細胞都能分泌 NO ,生理功能包括促進血管的舒張、抑制血小板 的凝集與破壞外來細胞 ( 由巨噬細胞分泌 ) 。在神 經細胞 NO 是具有 neurotranmitter 的作用,可能與 學習與記憶形成有關。 NO 的合成 : 由 arginine 轉成 citrulline 並釋出 NO ,此 反應是由 nitric oxide synthase 所催化。 NOS 在神 經細胞是受鈣離子活化,若在一般細胞則由細胞 激素 (cytokine) 所活化。
: 是一個自由基,非常活潑的氣體,生命期很短, 但扮演許多重要的生理角色。許多細胞都能分泌 NO ,生理功能包括促進血管的舒張、抑制血小板 的凝集與破壞外來細胞 ( 由巨噬細胞分泌 ) 。在神 經細胞 NO 是具有 neurotranmitter 的作用,可能與 學習與記憶形成有關。 NO 的合成 : 由 arginine 轉成 citrulline 並釋出 NO ,此 反應是由 nitric oxide synthase 所催化。 NOS 在神 經細胞是受鈣離子活化,若在一般細胞則由細胞 激素 (cytokine) 所活化。")
93
核苷酸 (Nucleotide) Nucleotide: 含三個部分 : 一個氮鹼基、一個 五碳糖與一個、二個或三個磷酸。 氮鹼 : 含嘌呤 (purine) 或嘧啶 (pyrimidine) ,由 具有類苯環 (aromatic ring) 的結構,因此在 pH 7 時對 260 nm 的 UV 光具有吸收性。 Purine: 含 adenine 、 guanine 、 xanthine 與 hypoxanthine 。 Pyrimidine: 含 thymine 、 cytocine 與 uracil
 Nucleotide: 含三個部分 : 一個氮鹼基、一個 五碳糖與一個、二個或三個磷酸。 氮鹼 : 含嘌呤 (purine) 或嘧啶 (pyrimidine) ,由 具有類苯環 (aromatic ring) 的結構,因此在 pH 7 時對 260 nm 的 UV 光具有吸收性。 Purine: 含 adenine 、 guanine 、 xanthine 與 hypoxanthine 。 Pyrimidine: 含 thymine 、 cytocine 與 uracil")
94
Fig. 14.20

95
Fig. 14.21

96
Fig. 14.23a

97
Fig. 14.23b

98
五碳糖分成兩類 : 核糖 (ribose) 與去氧核糖 (deoxyribose) Nucleosine: 不接磷酸的核苷。 purine 或 pyrimidine 接在五碳糖的 C-1 的部位, 以 -N-Glycosidic 形式接合。 五碳糖也分成核糖 (ribose) 與去氧核糖 (deoxyribose) 。
 與去氧核糖 (deoxyribose) Nucleosine: 不接磷酸的核苷。 purine 或 pyrimidine 接在五碳糖的 C-1 的部位, 以 -N-Glycosidic 形式接合。 五碳糖也分成核糖 (ribose) 與去氧核糖 (deoxyribose) 。")
99
含 ribose 的 nucleiside:adenosine 、 guanosine 、 cytosine 與 uridine 。 含 deoxyribose 的 nucleoside: deoxyadenosine 、 deoxyguanosine 、 deoxycytosine 與 deoxythyminosine 。

101
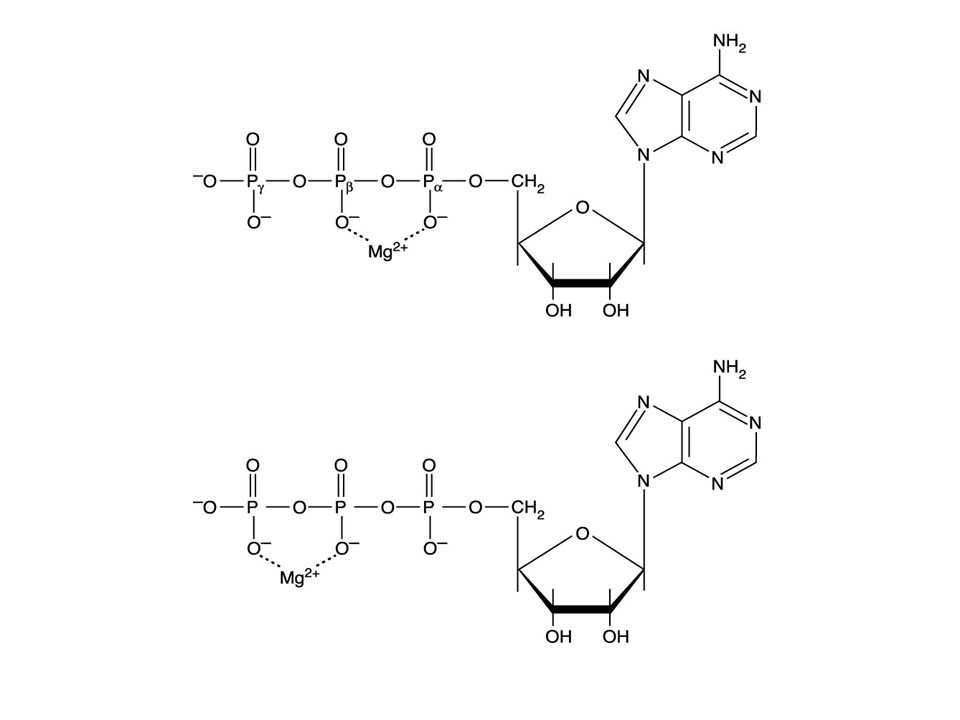
Nucleotide 就是 nucleoside 在五碳糖的 C-5 上接了 1 個或 2 、 3 個磷酸。 接一個磷酸稱 NMP 如 AMP 、 GMP 、 CMP 、 UMP 接二個磷酸稱 NDP 如 ADP 、 GDP 、 CDP 、 UDP 接三個磷酸稱 NTP 如 ATP 、 GTP 、 CTP 、 UTP 、 dATP 、 dGTP 、 dCTP 與 dTTP 。 ATP 如圖接在五 碳糖第一個磷酸稱 、第二個磷酸稱 ,第三個 磷酸稱 。

102
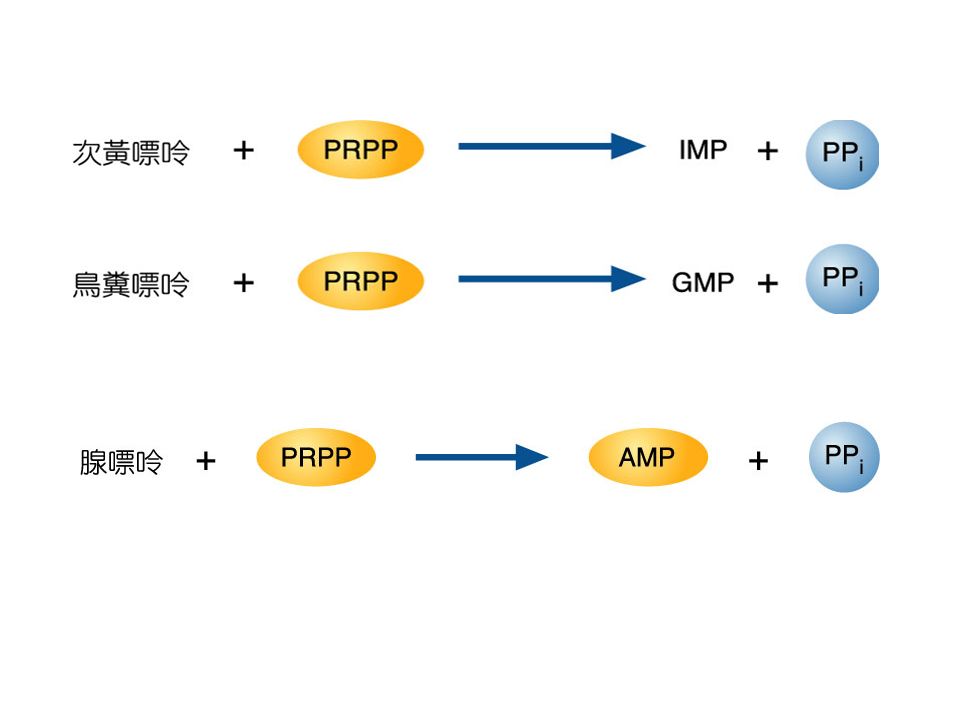
Purine nucleotide 合成 -D-ribose-5-phosphate + ATP PRPP + AMP -D-ribose-5-phosphate 是來自 Pentose phosphate pathway 。反應是由 PRPP synthetase , 再經過 9 個步驟後形成 IMP , IMP 再合成 AMP 或 GMP 。 AMP + ATP 2ADP NMP + ATP NDP + ADP 再由 Nucleotide diphosphate kinase 合成 NTP N 1 DP + N 2 TP N 1 TP + N 2 DP

103
– 嘌呤核苷酸

104
Fig. 14.24

105
Fig. 14.25

106
嘌呤回收途徑 (Purine salvage pathway): nucleotide 除了由 PRPP 生合成 (de novo 方 式 ) 外也可以直接與 purine 的鹼基合成 ( 因為 do novo 需耗掉 4 個 ATP)
: nucleotide 除了由 PRPP 生合成 (de novo 方 式 ) 外也可以直接與 purine 的鹼基合成 ( 因為 do novo 需耗掉 4 個 ATP)")
108
Hypoxanthine + PRPP IMP + PPi Guanine + PRPP GMP + ppi 催化此二反應的酵素為 HGPRT Lesch-Nyhan syndrome 是 HGPRT 缺乏會 造成過量的尿酸堆積與一些神經症狀 ( 導致 智能延緩,行動能力退化,自殘行為等 ) 。 從 HGPRT 的缺乏造成的症狀可以看出嘌呤 回收 (Purine salvage pathway) 途徑的重要 性。 Pyrimidine nucleotides 的合成
 。 從 HGPRT 的缺乏造成的症狀可以看出嘌呤 回收 (Purine salvage pathway) 途徑的重要 性。 Pyrimidine nucleotides 的合成")
109
–AMP 只有在ㄧ個方面不同於 IMP :ㄧ個胺基基 團來取代伊一個酮基氧 – 核苷三磷酸是最常使用於代謝之核苷酸

110
圖 14.26 嘌呤核苷酸生合成調節作用

111
Pyrimidine nucleotides 的合成 Carbamoyl phosphate synthetase II 位於細 胞質中催化生成 Carbamoyl phosphate 反應 如下 : 是合成 pyrimidine 途徑的第一反應,也是關 鍵酵素。 Carbamoyl phosphate synthetase I 在粒線 體中,是合成 Urea 第一個反應。

112
Fig. 14.27

113
pyrimidine 的合成與 purine 不一樣,先合成 出 orotate orotate + PRPP OMP UMP UMP 是合成 CTP 的先驅物 UMP UTP CTP

114
Fig. 14.28

115
Deoxyribonucleotides 的合成 : 以上所提及都是 ribonucleotide(A 、 G 、 U 與 C) 是組成 RNA 的建材分子 (building block) 。 而 DNA 的合成是由 2’- deoxyribonucleotide(A 、 T 、 C 、 G) 組成。 2’-deoxyribonucleotide 是由 NDP(nucleotide diphosphate) 還原合成而 來。
 是組成 RNA 的建材分子 (building block) 。 而 DNA 的合成是由 2’- deoxyribonucleotide(A 、 T 、 C 、 G) 組成。 2’-deoxyribonucleotide 是由 NDP(nucleotide diphosphate) 還原合成而 來。")
116
反應是由還原態的 Ribonucleotide reductase 催化, Thioredoxin reductase(13 kDa NADPH 是其輔因子 ) 再催化氧化態的 Ribonucleotide reductase 成還原態。
 再催化氧化態的 Ribonucleotide reductase 成還原態。")
117
Heme 的合成 : Heme 是哺乳動物細胞能合成的複雜分子之 一,具有一個結合鐵的 porphyrin ring 。是 血紅素 (hemoglobin) 、肌蛋白 (myoglobin) 與 cytochrome c 的輔因子。是由 glycine 與 succinyl-CoA 所合成。
 、肌蛋白 (myoglobin) 與 cytochrome c 的輔因子。是由 glycine 與 succinyl-CoA 所合成。")
118
Fig. 14.29

119
Nucleotide degradation 在人類與鳥類 Purine(AMP 、 GMP) 分解的終產物是 uric acid Pyrimidine 被分解成 -alanine 或 - aminoisobutyric acid cytidine 、 uridine -alanine thymine -aminoisobutyric acid
 分解的終產物是 uric acid Pyrimidine 被分解成 -alanine 或 - aminoisobutyric acid cytidine 、 uridine -alanine thymine -aminoisobutyric acid")
120
Purine 分解途徑缺陷會造成一些疾病 1 痛風 (gout) 是一種 purine catabolism 異常 的疾病,造成血液高濃度尿酸。因 uric acid 的溶解度不高,易造成尿酸鹽在關節上堆 積,引起關節發炎。
 是一種 purine catabolism 異常 的疾病,造成血液高濃度尿酸。因 uric acid 的溶解度不高,易造成尿酸鹽在關節上堆 積,引起關節發炎。")
121
2 Adenosine deaminase deficiency: 酵素缺 乏的先天性疾病,造成幼兒易受感染而死 亡。是一種先天體液免疫不全症。 3 Purine nucleoside phosphorylase deficiency: 高濃度的 dGTP 易造成 T cells 損 害,也是一種先天免疫不全症。








 Metabolism, the sum of all of the enzyme-catalyzed reactions in a living organisms, is highly coordinated and purposeful cell activity.>")


 電子傳遞鏈 傳導概論>")





 原料、来源 1、脂肪合成原料 脂肪酸和甘油。 生物体能利用糖类或简单碳原物质转化为脂肪酸。>")




