Download presentation
Presentation is loading. Please wait.

1
第九章 植物的成花生理 植物营养生长到一定阶段,在外界条件的诱导下,就会进入生殖生长。一般植物必须长到一定大小,达到一定生理状态后,才能感受所要求的外界条件而开花。植物具有的能感受环境条件而诱导开花的生理状态称花熟状态。花熟状态是植物营养生长转入生殖生长的标志。通常将植物达到花熟状态之前的营养生长时期称幼年期。幼年期各类植物长短不一。 植物的开花大致可分为3个阶段: (1)成花诱导:指能感受环境条件,启动开花基因,使植物营养生长转化为生殖生长。 (2)花的发端:分生组织开始分化,形成花原基。 (3)花的发育:花原基进一步发育,形成各种花器官。
成花诱导:指能感受环境条件,启动开花基因,使植物营养生长转化为生殖生长。 (2)花的发端:分生组织开始分化,形成花原基。 (3)花的发育:花原基进一步发育,形成各种花器官。")
2
第一节 春化作用 一、春化作用现象 1918年Gassner等用冬黑麦进行试验,发现冬黑麦在萌发期或苗期必须经历一个低温阶段才能开花,而春小麦则不需要。冬小麦必须冬播,次年才正常开花,若改为春播,整个夏季便进行营养生长不开花。Gassner将小麦和黑麦分为冬性和春性两种。 1928年Lysenko(李森科)将吸水萌动的冬小麦种子经低温处理后春播,发现其可在当年夏季抽穗开花。他将低温促进植物开花的作用称春化作用(Vernalization)。
将吸水萌动的冬小麦种子经低温处理后春播,发现其可在当年夏季抽穗开花。他将低温促进植物开花的作用称春化作用(Vernalization)。")
3
2、植物对低温的要求是相对的:这类植物若不经过低温春化,只延长营养生长期,时间一长照样开花。如冬小麦春播,时间一长照样开花。
二、植物对低温反应的类型: 现在研究表明,除了冬小麦、冬黑麦以外,某些二年生植物如白菜、萝卜、芹菜、甜菜、甘蓝、天仙子等以及一些多年生植物如牧草的开花也需经春化作用。但各种植物对低温的要求不同:有三类: 1、植物对低温的要求是绝对的:这类植物若不经过一定天数的低温处理,可保持无限的营养生长,不开花。主要是些二年生和多年生草本植物。如油菜、胡萝卜、白菜等。 2、植物对低温的要求是相对的:这类植物若不经过低温春化,只延长营养生长期,时间一长照样开花。如冬小麦春播,时间一长照样开花。

4
3、多数喜温植物和许多木本植物不需要春化。
如水稻、玉米、黄瓜等。 根据小麦和黑麦开花对低温的要求和通过春化的时间,可将小麦和黑麦分为冬性、半冬性、春性三个类型,一般冬性越强,要求春化的温度越低,时间越长。冬小麦:0-3 ℃, 40-45天; 半冬性小麦:3-6 ℃,10-15天;春小麦:8-15℃,5-8天。 三、植物通过春化的条件 1、低温:对于大多数植物,1-2℃是最有效的春化温度。但只要有足够的时间,在-3-10℃范围内对春化都有效。各类植物通过春化所要求的低温和时间有所不同,一般冬性越强,要求春化的温度越低,时间越长。

5
在植物春化过程结束前,如将植物放到较高的生长温度下,低温的效果会被减弱或消除,这种现象称去春化作用。一般解除春化的温度为25-40℃。去春化作用呈现下列特点:
(1)去春化作用只能发生在低温处理的时期,当春化过程结束, 就不会被高温去春化。 (2)植物经低温处理时间愈长,解除春化愈困难。 (3)去春化的植物可用低温重新春化,而且低温的效应可以累加。这种解除春化之后,再进行的春化作用称再春化作用。 (4)缺氧也可发生去春化作用。 推测低温诱导植物开花可能与在低温下某种物质的合成有关。
去春化作用只能发生在低温处理的时期,当春化过程结束, 就不会被高温去春化。 (2)植物经低温处理时间愈长,解除春化愈困难。 (3)去春化的植物可用低温重新春化,而且低温的效应可以累加。这种解除春化之后,再进行的春化作用称再春化作用。 (4)缺氧也可发生去春化作用。 推测低温诱导植物开花可能与在低温下某种物质的合成有关。")
6
2、水分:将已萌动的小麦种子失水干燥,当其含水量低于40%时,就不能通过低温春化。
3、氧气:在缺氧条件下,即使满足了水分、低温要求,仍不能通过春化。据测定,春化期间,某些酶的活性会提高,氧化还原作用会加强,呼吸作用增加。缺氧也会发生去春化作用。 4、呼吸底物营养:通过春化作用还需要足够的营养物质。若将小麦种子的胚培养在富含蔗糖的培养基中,在低温下通过春化,但若培养基中缺乏蔗糖,则不能通过春化。 此外,许多植物在感受低温后还需经过长日照诱导才能开花。如天仙子,经低温处理后放在短日照下,就不开花。

7
四、春化作用机理 (一)春化刺激部位和传递 1、春化作用的时期和部位 (1)春化作用的时期 一般植物在种子萌发到植物营养生长的苗期都可感受低温而通过春化。如冬小麦、黑小麦可在萌动期,也可在苗期,甚至可在母体正在发育的幼胚中进行。 但胡萝卜、甘蓝和芹菜等植物只有在幼苗长到一定大小以后,才能通过春化。例甘蓝幼苗的茎的直径达0.6cm,叶宽达5cm以上才能通过春化。月见草至少要有6-7个叶片,才接受低温处理。

8
(2)感受春化的部位 分生组织和某些进行细胞分裂的部位。 1)茎尖分生组织:将芹菜的茎尖给予0℃左右的低温,其它部分放在25℃的高温中,植物可通过春化,反之,将茎尖置于高温中,其它部分置于低温中,则不能通过春化。 2)幼胚:Gregory和Purvis将冬黑麦的胚分开,用1-2℃低温处理离体胚后,转移到培养基上培养,长出的小植株表现出春化效果。 3)叶柄基部:椴花的叶柄基部在适当低温处理后,经组织培养可培养出花茎,但如果将叶柄基部切除0.5cm,再生的植株就不能形成花茎。
幼胚:Gregory和Purvis将冬黑麦的胚分开,用1-2℃低温处理离体胚后,转移到培养基上培养,长出的小植株表现出春化效果。 3)叶柄基部:椴花的叶柄基部在适当低温处理后,经组织培养可培养出花茎,但如果将叶柄基部切除0.5cm,再生的植株就不能形成花茎。")
9
2、春化作用的传递 有两类结果相反的实验: (1)低温处理的植株可能产生了某种可以传递的物质,并可经过嫁接传递给未经春化的植株。 天仙子嫁接实验:Melchers和Lang将已春化的二年生植物天仙子枝条或一片叶片嫁接到未春化天仙子枝条上,能诱导未春化植株开花。不但如此,将已春化的天仙子枝条嫁接到烟草或矮牵牛植株上,也能使这两种植物开了花。 (2)低温处理不产生传递物质,低温感应效应经细胞分裂由一个细胞传递到另一个细胞。
低温处理不产生传递物质,低温感应效应经细胞分裂由一个细胞传递到另一个细胞。")
10
1)菊花嫁接实验:将菊花已春化植株和未春化植株嫁接,未春化植株不能开花。如将春化后的芽移植到未春化植株上,则仅这个芽长出的梢开花。
2)萝卜嫁接实验:将未春化的萝卜植株的顶芽嫁接到已春化的萝卜植株上,该顶芽长出的梢不能开花。 (二)春化作用的生理生化变化: 1、呼吸作用增加:试验表明: 1)春化处理过的冬小麦呼吸速率要比对照高;2)春化作用需氧气和呼吸底物;3)用氧化磷酸化解偶联剂DNP处理,可抑制春化的过程;4)在春化过程中,呼吸氧化酶系统会发生了变化,前期以细胞色素氧化酶为主,后期细胞色素氧化酶活性逐渐降低,抗坏血酸氧化酶活性增加。
萝卜嫁接实验:将未春化的萝卜植株的顶芽嫁接到已春化的萝卜植株上,该顶芽长出的梢不能开花。 (二)春化作用的生理生化变化: 1、呼吸作用增加:试验表明: 1)春化处理过的冬小麦呼吸速率要比对照高;2)春化作用需氧气和呼吸底物;3)用氧化磷酸化解偶联剂DNP处理,可抑制春化的过程;4)在春化过程中,呼吸氧化酶系统会发生了变化,前期以细胞色素氧化酶为主,后期细胞色素氧化酶活性逐渐降低,抗坏血酸氧化酶活性增加。")
11
2、增加了RNA的合成和性质的改变:试验证明:
低温处理可提高冬小麦幼苗的可溶性RNA和rRNA含量。从经过60d春化处理的冬小麦幼苗中提取出来的染色体,主要合成大于20S的mRNA,而从常温下萌发的冬小麦幼苗中提取出来的染色体,主要合成9-20S的mRNA,说明,低温处理下,植物有新的mRNA的合成。 3、增加可溶性蛋白质和游离Aa的含量:实验表明: 低温处理可提高冬小麦种子增加可溶性蛋白质和游离Aa的含量。电泳分析显示,春化处理的冬小麦有新的蛋白质谱带出现。在去春化作用的冬小麦中,原来经低温诱导产生的新蛋白质消失了。在增加的游离Aa中,以Proline增加最多。 说明,春化作用是一个基因启动、表达与调节的复杂过程,涉及到某些特定基因的活化,特定mRNA和蛋白质的合成。

12
(三)春化作用机理 1、春化基因去甲基化假说 认为:低温导致DNA去甲基化,而改变基因表达。 依据:DNA去甲基化试剂5-氮胞苷处理拟南芥晚花突变体和冬小麦,总DNA甲基化水平降低,开花提早。而拟南芥早花突变体和春小麦对5-氮胞苷不敏感。因此,认为拟南芥晚花突变体之所以迟开花,是由于基因被甲基化,而阻碍了开花相关基因表达, 2、与开花抑制物基因(FLC,flowering locus c)的表达有关: 在非春化植株的顶端分生组织中,FLC强烈表达,但低温处理后FLC表达就减弱。低温时间越长,FLC表达越弱。低温抑制FLC基因的表达,从而减弱了其对开花的抑制作用。
的表达有关: 在非春化植株的顶端分生组织中,FLC强烈表达,但低温处理后FLC表达就减弱。低温时间越长,FLC表达越弱。低温抑制FLC基因的表达,从而减弱了其对开花的抑制作用。")
13
3、与开花基因Ver203的表达有关 种康等(1994)得到与春化相关的三个基因Ver17、 Ver69、 Ver203的cDNA,将Ver203反义基因转入植物,发现开花显著被抑制。 4、春化作用与GA 的关系 (1)春化作用与GA有关: 1)二年生植物天仙子、白菜、甜菜等若不经过低温处理,植物呈莲座状,不能抽苔开花。但使用GA可使这些植物不经低温处理就能开花。 2)许多植物如菊、小麦、油菜等经春化处理后,体内GA含量会增加。3)用GA生物合成抑制剂如福斯方D、AMO-1618等处理,可抑制春化作用。
春化作用与GA有关: 1)二年生植物天仙子、白菜、甜菜等若不经过低温处理,植物呈莲座状,不能抽苔开花。但使用GA可使这些植物不经低温处理就能开花。 2)许多植物如菊、小麦、油菜等经春化处理后,体内GA含量会增加。3)用GA生物合成抑制剂如福斯方D、AMO-1618等处理,可抑制春化作用。")
15
(2)GA不是春化开花刺激物 1)GA并不能诱导所有需春化的植物开花。GA对低温的替代作用只对长日植物有效。 2)植物对GA的反应不同于低温:被低温诱导的植物抽苔时就出现花芽,而GA虽可引起植物茎的伸长或抽苔,但不一定开花。 近年的研究发现植物体的的赤霉素烯酮与植物的春化作用有关。 五、农业生产应用: 1、人工春化处理 冬小麦萌动后,对其进行低温处理可进行春天补种。在育种中可利用春化处理,在一年中培育3-4代冬性作物,加速育种过程。

16
2、调种引种 在南北方地区之间引种时,必须注意品种对温度的要求。北种南引,植物因得不到合适的低温,致使植物只进行营养生长,不开花结果。 3、控制花期 1)低温处理可使秋播的一、二年生草本花卉改为春播,当年开花。 2)利用解除春化控制某些植物开花:例洋葱鳞茎在春季种植前,用高温处理,解除春化作用,可防止其在生长期开花而增加鳞茎的产量。
利用解除春化控制某些植物开花:例洋葱鳞茎在春季种植前,用高温处理,解除春化作用,可防止其在生长期开花而增加鳞茎的产量。")
17
第二节 光周期现象 1920年美国的贝尔兹维尔农业研究小组的W.W.Garner和H.A.Alard注意到烟草的一个变种,在夏季进行旺盛的营养生长,株高可达3-5 M,但不开花.但在冬季转入温室后,株高不足1 M就可开花。 他们试验了温度、光质、营养等各种条件,发现昼夜长度是影响烟草开花的关键因素。如果在夏天用黑布遮光,人为缩短日照长度,烟草就能开花,冬季在温室内用人工光照延长日照长度,则烟草保持营养状态而不开花。 由此他们得出结论:植物的开花与昼夜的相对长度有关。

18
现在已证明,不仅是植物的开花,植物的休眠和落叶,鳞茎、块茎、球茎等地下贮藏器官的形成都受光周期的影响。如菊芋块茎的形成需短日照,马铃薯块茎、洋葱鳞茎的形成需长日照。人们将一昼夜间光暗交替(昼夜光暗的相对长度)称光周期。而植物对昼夜长度所发生的反应现象称光周期现象。研究最多的是植物的成花光周期诱导。 冬季 夏季

19
一、植物光周期的类型 依据自然界植物对日照长短反应的不同,可将植物分为: 1、长日植物(LDP):指在24小时昼夜周期中,日照长度长于一定时间才能成花的植物。延长光照可促进或提早这类植物开花。一般是上半年开花的植物。如小麦、大麦、油菜、菠菜、萝卜、白菜、芹菜,山茶、天仙子等。 2、短日植物(SDP):指在24小时昼夜周期中,日照长度短于一定时间才能成花的植物。对于这类植物,适当延长暗期或缩短光期可促进或提早开花。一般是下半年开花的植物。如大豆、高梁、水稻、玉米、苍耳、烟草、菊花等。 3、日中性植物(NDP):这类植物成花对日照长度无特别要求,只要其它条件能满足,可在任何日照长度下开花。如黄瓜、茄子、番茄、辣椒、菜豆、月季、君子兰、向日葵等。
:指在24小时昼夜周期中,日照长度短于一定时间才能成花的植物。对于这类植物,适当延长暗期或缩短光期可促进或提早开花。一般是下半年开花的植物。如大豆、高梁、水稻、玉米、苍耳、烟草、菊花等。 3、日中性植物(NDP):这类植物成花对日照长度无特别要求,只要其它条件能满足,可在任何日照长度下开花。如黄瓜、茄子、番茄、辣椒、菜豆、月季、君子兰、向日葵等。")
20
4、长短日植物和短长日植物: 这些植物花的诱导和形成两个过程明显分开,需双重的日照条件。大叶落地生根、芦荟、夜香树等植物长日照有花的诱导作用,但花器官的形成需短日照是长短日植物。风铃草、瓦松、白三叶草等植物花的诱导在短日下进行,而花器官的形成需长日照是短长日植物。此外,还有: 中日照植物:指只能在一定的中等日照长度下才能开花,延长或缩短日照都抑制开花的植物。常见有甘蔗,其要求 h的日照。 两极光周期植物:在中等日照条件下植物保持营养生长,而在较长或较短日照条件下才开花。如狗尾草。

21
我们将诱导长日植物开花所需最短的日照时数或短日植物开花最长的日照时数,称临界日长是(CDL:critical daylength)
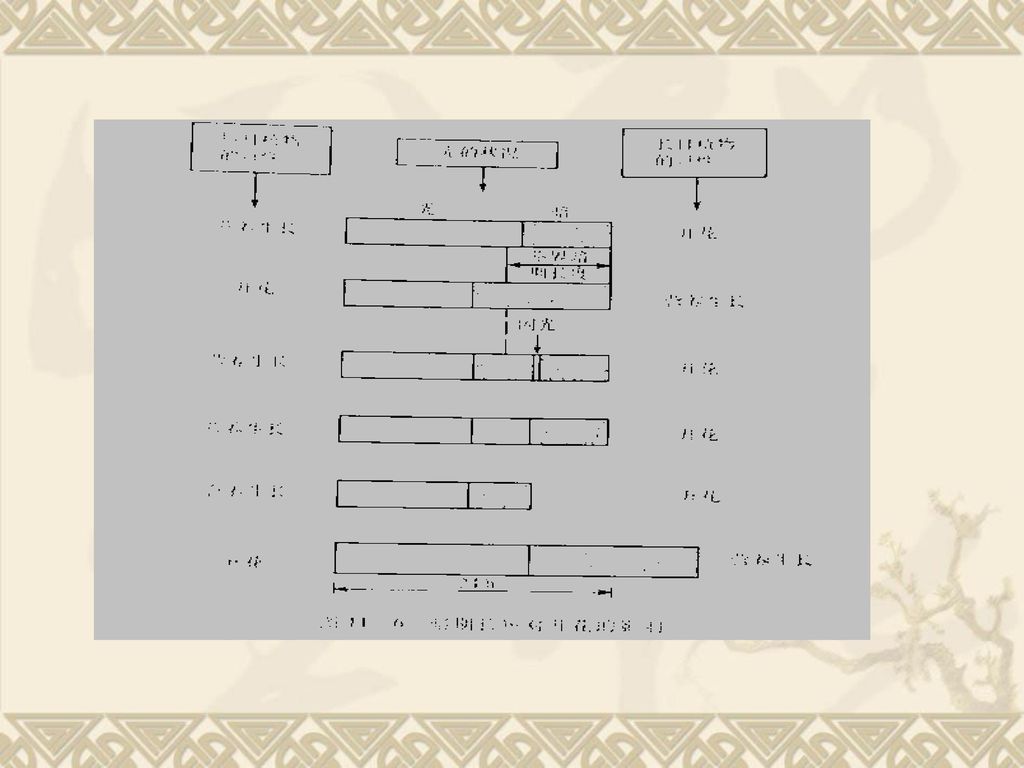
我们将诱导长日植物开花所需最短的日照时数或短日植物开花最长的日照时数,称临界日长是(CDL:critical daylength).长日植物的光照时间只要大于CDL就开花,而短日植物光照时间只要小于CDL就可开花。例SDP大豆的CDL为14h,LD P冬小麦的CDL为12h,现光照为13h,两种植物都开花。 二、光周期诱导 (一)光周期诱导 1、光周期诱导天数:植物开花必须感受适当的光周期刺激,但在适宜的光周期下要放多长时间呢?研究表明:为了引起开花,植物并不需要长时间地都放在适当的日照条件下,只要得到足够天数的适宜的光周期,以后即使放在不适宜的光周期下,仍然可以长期保持刺激效果而开花,这种诱导效应称光周期诱导。
.长日植物的光照时间只要大于CDL就开花,而短日植物光照时间只要小于CDL就可开花。例SDP大豆的CDL为14h,LD P冬小麦的CDL为12h,现光照为13h,两种植物都开花。 二、光周期诱导. (一)光周期诱导. 1、光周期诱导天数:植物开花必须感受适当的光周期刺激,但在适宜的光周期下要放多长时间呢?研究表明:为了引起开花,植物并不需要长时间地都放在适当的日照条件下,只要得到足够天数的适宜的光周期,以后即使放在不适宜的光周期下,仍然可以长期保持刺激效果而开花,这种诱导效应称光周期诱导。")
22
各种植物通过光周期诱导的天数不同。 短日照植物:苍耳、日本牵牛、水稻、浮萍只需1d,大豆要2-3d,大麻要4d,菊花要12d。
2、光周诱导光强 用于光周期诱导的光强只要50-100lx,低于光合作用所需光强。

23
(二)光周期诱导中光期和暗期的作用 对于LDP植物,只要日照长度大于CDL就开花,而对于SDP植物而言:日照长度小于CDL就开花。 与临界日长相对应的概念为临界夜长:短日植物开花的最短的暗期长度或长日植物开花的最长暗期长度称临界夜长( critical night length ,CNL)。 对于SDP,暗期只要大于CNL,就开开花,对于LDP,暗期小于CNL才开花。 光期和暗期在光周期诱导中的作用?

24
1、暗期比光期更重要 (1) SDP大豆:CNL为10h, 发现暗期小于10h无花芽分化,暗期长于10h形成花芽,暗期13-14h 花芽最多。 (2) LDP天仙子:CDL为11h。发现12h的光期+12H的暗期下不开花,但6h光期+6h暗期植物开了花。 因此,SDP只要满足一个长的(大于CNL )连续的暗期就开花,SDP称长夜植物。而LDP只要满足一个短暗期就能开花,LDP称短夜植物。 2、暗期起决定性作用,光期也必不可少 短日植物的成花诱导要求暗期,但光期太短也不能成花,也就是说,在一定范围内,暗期愈长对开花越有利,但并不可以无限制地延长暗期。在大于临界夜长固定的长暗期中,增加光照时间,SDP的开花数会增加,因为花的发育需光合作用提供足够的营养物质。
连续的暗期就开花,SDP称长夜植物。而LDP只要满足一个短暗期就能开花,LDP称短夜植物。 2、暗期起决定性作用,光期也必不可少. 短日植物的成花诱导要求暗期,但光期太短也不能成花,也就是说,在一定范围内,暗期愈长对开花越有利,但并不可以无限制地延长暗期。在大于临界夜长固定的长暗期中,增加光照时间,SDP的开花数会增加,因为花的发育需光合作用提供足够的营养物质。")
26
对于长日植物,日照只要大于临界日长就开花,日照愈长开花愈早,连续光照开花最早。
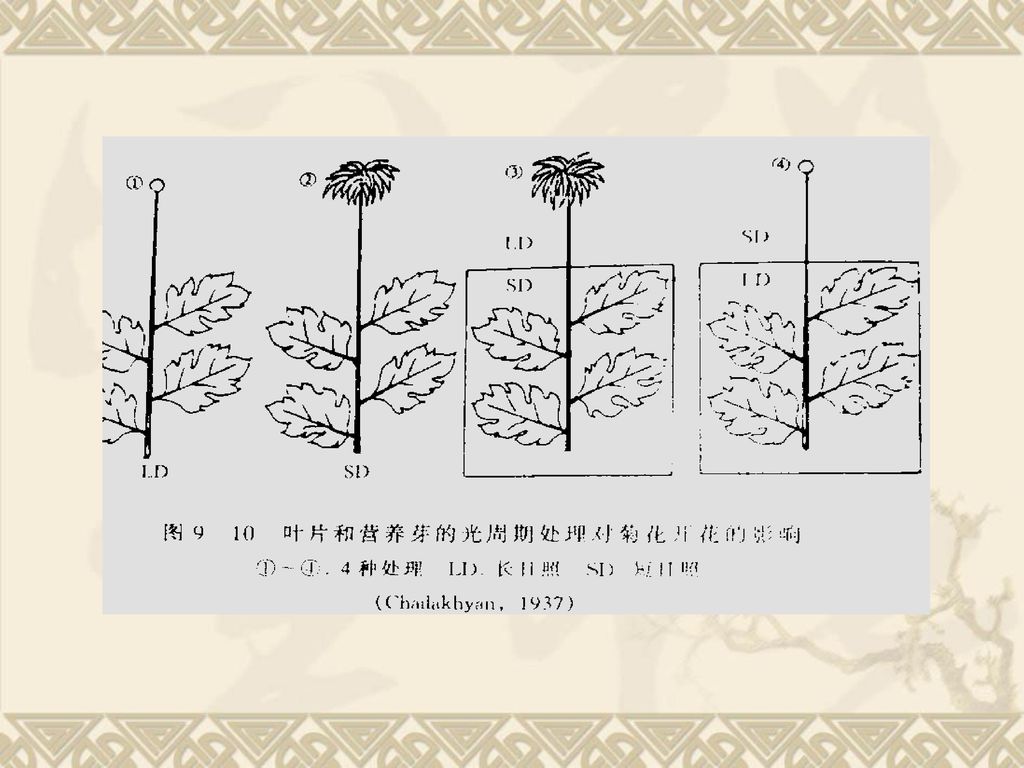
(三)光周期刺激的感受和传递 1、感受光刺激的部位:叶片,菊花实验 A、将SDP植物菊花整株放在长日条件下,植物保持营养生长而不开花。 B、将SDP植物菊花整株放在短日条件下,植物开花。C、将菊花的叶片处于短日条件下而茎尖给予长日照,植株开花。 D、将菊花的叶片处于长日条件下而茎尖给予短日照,植株不开花。 表明:只要叶片处于适宜的光周期条件下就能诱导成花,而于茎尖所处的光周期条件无关, 叶片是感受光刺激的主要部位。
光周期刺激的感受和传递. 1、感受光刺激的部位:叶片,菊花实验. A、将SDP植物菊花整株放在长日条件下,植物保持营养生长而不开花。 B、将SDP植物菊花整株放在短日条件下,植物开花。C、将菊花的叶片处于短日条件下而茎尖给予长日照,植株开花。 D、将菊花的叶片处于长日条件下而茎尖给予短日照,植株不开花。 表明:只要叶片处于适宜的光周期条件下就能诱导成花,而于茎尖所处的光周期条件无关, 叶片是感受光刺激的主要部位。")
28
2、感受光周期刺激的时期 通常植物生长到一定大小后,才能感受光周期刺激。不同植物对光周期表现的敏感开始年龄不同,如大豆在子叶伸展期,水稻在7 叶期前后,红麻在6 叶期。幼小的和衰老的叶片敏感性差,刚成熟的叶片敏感性最强。 3、光周期刺激的传递 接受光周期的部位是叶片,诱导开花的部位是茎尖生长点。在最初的感受部位和发生成花反应的部位之间存在着叶柄和一段茎的距离。 因此推测:叶片接受光周期刺激后,可能产生了开花刺激物,被运输到了茎尖,从而产生了成花反应。

29
(1)接受光周期刺激后叶片有开花刺激物的形成
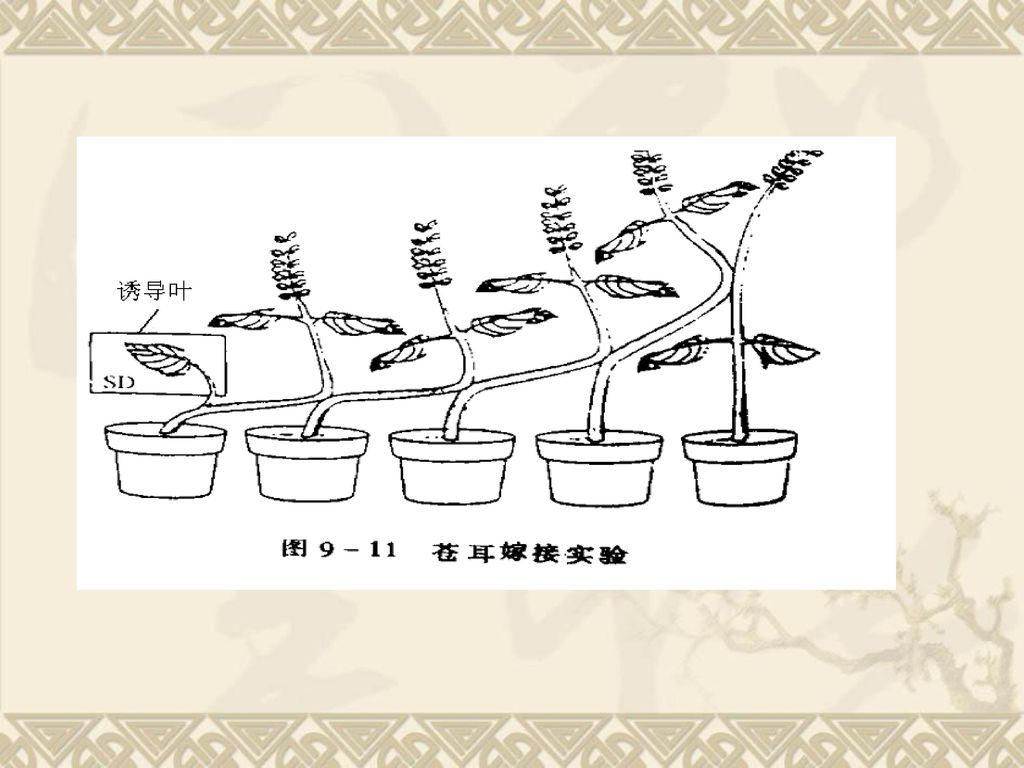
柴拉轩(Chailakhyan)5株苍耳嫁接实验: 将5株苍耳嫁接串连在一起,只要其中一株的一片叶接受了适宜的光周期诱导,即使其他植株都在长日照条件下,最后所有植株都能开花。 证实存在开花刺激物质,可以通过嫁接在株间进行传递。 (2)不同光周期类型的开花刺激物的性质相似:嫁接实验 LDP与SDP嫁接实验: A.LDP天仙子和SDP烟草嫁接,发现在LD或SD下两种植物都能开花。 B.将经适宜光周期诱导的LDP大叶落地生根的花序切去,嫁接上未经诱导的SDP高凉菜植物,置于长日照下,高凉菜可大量开花。而用未经光周期诱的大叶落地生根嫁接上高凉菜,高凉菜仍保持营养生长。
5株苍耳嫁接实验: 将5株苍耳嫁接串连在一起,只要其中一株的一片叶接受了适宜的光周期诱导,即使其他植株都在长日照条件下,最后所有植株都能开花。 证实存在开花刺激物质,可以通过嫁接在株间进行传递。 (2)不同光周期类型的开花刺激物的性质相似:嫁接实验. LDP与SDP嫁接实验: A.LDP天仙子和SDP烟草嫁接,发现在LD或SD下两种植物都能开花。 B.将经适宜光周期诱导的LDP大叶落地生根的花序切去,嫁接上未经诱导的SDP高凉菜植物,置于长日照下,高凉菜可大量开花。而用未经光周期诱的大叶落地生根嫁接上高凉菜,高凉菜仍保持营养生长。")
31
(3)合成开花刺激物需要相当长的时间:2-4h 去叶实验:
(4)开花刺激物是经韧皮部运输 利用环割或蒸汽处理植物的叶柄或茎,干扰或阻止韧皮部的运输,可延迟或抑制开花。说明开花刺激物是经韧皮部运输的。 (5)开花刺激物运输速度一般小于或接近同化物 柴拉轩1957年用14CO2进行试验,发现开花的紫苏由诱导叶片运到花芽的同位素很高,而不开花的紫苏芽的同位素含量较低。毒麦的蔗糖的运输速度为100cm/h,而开花刺激物为10-24mm/h,牵牛花的同化物为33-37 cm/h,开花刺激物运输速度为24-37 cm/h。
开花刺激物是经韧皮部运输. 利用环割或蒸汽处理植物的叶柄或茎,干扰或阻止韧皮部的运输,可延迟或抑制开花。说明开花刺激物是经韧皮部运输的。 (5)开花刺激物运输速度一般小于或接近同化物. 柴拉轩1957年用14CO2进行试验,发现开花的紫苏由诱导叶片运到花芽的同位素很高,而不开花的紫苏芽的同位素含量较低。毒麦的蔗糖的运输速度为100cm/h,而开花刺激物为10-24mm/h,牵牛花的同化物为33-37 cm/h,开花刺激物运输速度为24-37 cm/h。")
32
4、开花刺激物提取 1961年(Lincoln)等人以甲醇从开花的苍耳植株提取了成花刺激物,并将它们的粗提物加入羊毛脂,施于在长日照下生长的苍耳植株上,可诱发50%的植株开花。而从营养生长的苍耳植株内提取的物质不具有诱导开花的活性,但是在进一步提纯这种物质时活性消失。 1967年Carr重复了Lincoln的工作,并发现在开花活性提取物中加入GA3后,诱导开花效果特别明显,被处理苍耳100%开花。 1970年Hadson和Hammer也重复这个实验,但用丙酮提取,也证明在加入GA3的粗提物可诱导浮萍、苍耳开花,还进一步证明这种粗提物对长日植物无效。

33
开花刺激物可能与GA有关。依据:1)LDP植物菠菜在SD下,GA含量低,植株不开花,当把植株转移到LD下时,进行13-OH途径合成GA,GA1水平增加5倍。2)GA可代替某些LDP植物如天仙子、金光菊等的LD条件,使它们在非诱导条件SD下开花。3)GA可代替某些冬性LDP植物的低温条件,使它们不经低温而开花。 还有证据表明,水杨酸、ETH、CTK、多胺可能也是开花刺激物组成成分。 5、 成花抑制物 由于明确的成花刺激物没找到,推测植物在非诱导条件下不能开花是由于叶片中存在某种开花抑制物。依据: LDP天仙子、SDP草莓和藜在非诱导光周期条件下不能开花,但是去掉全部叶片时,在任何日长下都能开花。但成花抑制物至今也没找到。

34
三、光敏素在植物成花中的作用 1、夜间断现象 在一个足够使SDP开花,LDP不开花的长暗期中,给予一个短时间足够强度的闪光处理,发现可抑制SDP开花,促进LDP开花,此现象称夜间断现象或短夜反应。短夜反应的效果: (1)时相分布:暗期中间给于闪光处理最有效。 (2)光强和照光时间有关:短时间闪光处理需高强度光,长时间光照,光强可弱点,例对于苍耳,10-100lx照光1分钟便有效。 (3)光质有关:闪光处理抑制SDP开花,促进LDP成花,红光最有效,红光的夜间断反应可被远红光所转逆,远红光逆转红光的效果又可被红光逆转。推测光敏素参于光周期成花反应。
时相分布:暗期中间给于闪光处理最有效。 (2)光强和照光时间有关:短时间闪光处理需高强度光,长时间光照,光强可弱点,例对于苍耳,10-100lx照光1分钟便有效。 (3)光质有关:闪光处理抑制SDP开花,促进LDP成花,红光最有效,红光的夜间断反应可被远红光所转逆,远红光逆转红光的效果又可被红光逆转。推测光敏素参于光周期成花反应。")
35
2、光敏素与植物的开花 暗期间断实验表明:光敏素对成花的作用与Pr和Pfr的可逆转化有关。 进一步的研究表明:

36
(1)成花作用不是决定于Pr和Pfr绝对含量,而是受Pfr/Pr比值的影响:
1)Pfr/Pr比值高,促进LDP成花 长光期,Pfr合成增加,短间期,Pfr逆转为Pr减少,结果Pfr/Pr比值高,LDP成花。 2)Pfr/Pr比值低,有利于SDP成花: 短光期Pfr合成少,长暗期Pfr又逆转为Pr,Pfr/Pr比值降低,SDP成花。 (2)诱导SDP或LDP开花还需Pfr的变化 1)SDP在暗期的前期要求高的Pfr含量,后期才要求低的Pfr水平: SDP牵牛或红藜若在暗期就以远红光照射,前期Pfr水平低,反而抑制成花,但若暗诱导的后期,降低Pfr水平,就促进SDP成花。
Pfr/Pr比值高,促进LDP成花. 长光期,Pfr合成增加,短间期,Pfr逆转为Pr减少,结果Pfr/Pr比值高,LDP成花。 2)Pfr/Pr比值低,有利于SDP成花: 短光期Pfr合成少,长暗期Pfr又逆转为Pr,Pfr/Pr比值降低,SDP成花。 (2)诱导SDP或LDP开花还需Pfr的变化. 1)SDP在暗期的前期要求高的Pfr含量,后期才要求低的Pfr水平: SDP牵牛或红藜若在暗期就以远红光照射,前期Pfr水平低,反而抑制成花,但若暗诱导的后期,降低Pfr水平,就促进SDP成花。")
37
2)LDP:暗期的前期要求由高到低的Pfr含量变化,后期才要求由低到高的Pfr水平:
LDP毒麦在短日状态时,以红光延长光照8h,增加前期Pfr含量,植株成花不显著,但如在暗期前期红光中加入几小时的远红光,则明显促进成花。 四、光周期限理论在农业生产上的应用 1、 植物的地理起源和分布: (1) 低纬度地区:只分布短日植物; (2)高纬度地区: 多分布长日植物; (3)中等纬度地区,长短日植物并存。 但需指出的是:人工栽培的品种,同一品种可在不同纬度地区都有分布,如大豆是SDP植物,东北和海南岛都有当地的育成品种,水稻也是如此。已形成对当地光周期的适应。
 低纬度地区:只分布短日植物; (2)高纬度地区: 多分布长日植物; (3)中等纬度地区,长短日植物并存。 但需指出的是:人工栽培的品种,同一品种可在不同纬度地区都有分布,如大豆是SDP植物,东北和海南岛都有当地的育成品种,水稻也是如此。已形成对当地光周期的适应。")
38
2、北半球一年四季光周期的变化:

39
3、在农业生产上的应用: (1)引种和育种 1)引种:同纬度引种容易成功,不同纬度引种应考虑引种植物的光周期性质和原产地与被引种地日照的长短。还可用品种的早晚性加以调节: SDP植物:南种北引,延迟开花,应引早熟种;而北种南引,提前开花,应引晚熟种。LDP植物:南种北引,提前开花,应引晚熟种;而北种南引,延迟开花,应引早熟种。 2)育种:通过人工光周期调节,加速良种繁育,缩短育种年限。方法: 培育新品种:通过人工光周期调节,使生长在不同光周期地区的植物,同时开花进行有性杂交,培育新品种。缩短育种年限:SDP水稻、玉米可在海南岛加快繁育种子。长日植物小麦夏季在黑龙江、冬季在云南种植。一年内可繁育2-3代。
育种:通过人工光周期调节,加速良种繁育,缩短育种年限。方法: 培育新品种:通过人工光周期调节,使生长在不同光周期地区的植物,同时开花进行有性杂交,培育新品种。缩短育种年限:SDP水稻、玉米可在海南岛加快繁育种子。长日植物小麦夏季在黑龙江、冬季在云南种植。一年内可繁育2-3代。")
40
(2)控制花期 可用人工控制光周期来提前或推迟花卉植物开花。SDP植物采用遮光法可使其在上半年开花,LDP植物进行人工延长光照处理,则可提早开花。 (3)调节营养生长和生殖生长 以收获营养体为主的植物可采用:SDP植物南种北引,延迟开花,促进营养生长如SDP麻类植物;LDP植物采用北种南引,延迟开花,促进营养生长。

41
第三节 花器官的形成和性别表现 一、花器官的形成 植物经过一定时期的营养生长后,就能感受低温和光周期诱导,产生开花刺激物。开花刺激物运输到茎端分生组织,由茎端分生组织分化形成花器官。 花的发育可分为3个阶段: 1)成花诱导:植物感受外界刺激如光周期、低温等以及自身产生的开花信号。 2)形成花原基:感受成花刺激后,茎端分生组织转变为花分生组织。 3)形成花器官:花分生组织分化形成不同的花器官。
成花诱导:植物感受外界刺激如光周期、低温等以及自身产生的开花信号。 2)形成花原基:感受成花刺激后,茎端分生组织转变为花分生组织。 3)形成花器官:花分生组织分化形成不同的花器官。")
42
(一)成花诱导途径 以拟南芥为材料,采用现代分子生物学研究手段,发现拟南芥有4条成花诱导发育途径。 1、光周期途径 红光促进LDP成花,抑制SDP成花,而远红光促进SDP成花,抑制LDP成花。光是植物成花的诱导因子之一。现已知,光敏素(Phytochrome,PHY)和隐花素(Cryptochrome,CRY)参与了光诱导成花的过程。 拟南芥至少有5个基因编码光敏素蛋白,它们是PHYA、PHYB、PHYC、PHYD和PHYE。这些编码基因有各自不同的时间、空间分布和生理功能。其中: (1) PHYB接受红光后抑制CONSTANS(CO)基因的表达,而抑制开花;
和隐花素(Cryptochrome,CRY)参与了光诱导成花的过程。 拟南芥至少有5个基因编码光敏素蛋白,它们是PHYA、PHYB、PHYC、PHYD和PHYE。这些编码基因有各自不同的时间、空间分布和生理功能。其中: (1) PHYB接受红光后抑制CONSTANS(CO)基因的表达,而抑制开花;")
43
(2) PHYA与隐花素基因CRY1和CRY2互作,通过生理钟基因,促进开花素的形成,从而促进CO的表达, CO编码一个转录因子,通过作用于分生组织决定基因AGL20(AGAMOUS-LIKE20),促进开花。 2、春化/自主途径 植物要达到一定的年龄才能开花,称开花自主途径。植物达到花熟状态后或经低温处理后,会抑制开花阻抑物基因FLC(FLOWERING LOCUS C)的表达,而解除了FLC对分生组织决定基因AGL20的抑制作用,而促进下游基因的表达。 3、赤霉素GA途径 GA与受体结合后,通过自身的信号转导途径,促进分生组织决定基因AGL20的表达,促进开花和在非诱导条件下开花。
的表达,而解除了FLC对分生组织决定基因AGL20的抑制作用,而促进下游基因的表达。 3、赤霉素GA途径. GA与受体结合后,通过自身的信号转导途径,促进分生组织决定基因AGL20的表达,促进开花和在非诱导条件下开花。")
44
4、蔗糖(糖类)途径 蔗糖可通过促进分生组织决定基因AGL20的表达,而促进拟南芥开花。 (二)花器官发育的基因控制 花器官的形成依赖于器官特征基因在时间和空间位置上的表达。近几年,对花发育特异性基因的表达研究有了突破性进展。最突出的是对花发育同源异型基因的研究。 器官特征基因的鉴定是通过花器官改变突变体的研究进行的。由于基因发生突变,会发生如花瓣部分被雄蕊代替、萼片被心皮代替等花器官改变。这种遗传变异现象被称为花发育同源异型突变。

45
促进 抑制 促进 解除抑制

46
花形态建成控制的ABC理论( Coen提出)
在拟南芥中发现有5种花器官特征基因,AP1、AP2、AP3、PI和AG基因,可分为A、B、C 3类: A类: AP2基因,控制萼片的形成。 B类: AP3和PI基因,和A类基因一起控制花瓣形成。 C类: AG基因,和B类基因一起,形成雄蕊;仅C类基因表 达,则形成心皮。 (三)花器官形成所需条件 1、营养状态 营养是花芽分化和器官形成的物质基础。其中:1).碳水化合物是合成其它物质的碳源和能源。2)蛋白质是花芽分化和器官形成所必需的,氮素营养不足,花芽分化少且慢,但若氮素过多,C/N比失调,会造成植物贪青晚熟。3)磷的化合物和核酸也参与了花芽的分化。
花器官形成所需条件. 1、营养状态. 营养是花芽分化和器官形成的物质基础。其中:1).碳水化合物是合成其它物质的碳源和能源。2)蛋白质是花芽分化和器官形成所必需的,氮素营养不足,花芽分化少且慢,但若氮素过多,C/N比失调,会造成植物贪青晚熟。3)磷的化合物和核酸也参与了花芽的分化。")
47
ABC三类基因突变后花器官表型的变化

48
2、内源激素 体内营养状况与内源激素的平衡相互调节着花芽的分化。已知:CTK、ABA和乙烯含量高、GA、IAA含量低促进花芽的分化;高浓度GA、 IAA有抑制作用。高GA由于可诱导a-淀粉酶的活性,促进淀粉的水解,对花芽形成不利。 3、外部因素 (1)光:光照有利于花器官的形成,自然光照时间越长,光强越大,形成的有机物质越多,对花形成越有利。不同植物开花和器官形成所要求的最低光强有所不同:一般阳地植物高于阴生植物;雄蕊高于雌蕊。例小麦花药发育处于花粉母细胞形成前夕时遮光72h,花粉全部败育。
光:光照有利于花器官的形成,自然光照时间越长,光强越大,形成的有机物质越多,对花形成越有利。不同植物开花和器官形成所要求的最低光强有所不同:一般阳地植物高于阴生植物;雄蕊高于雌蕊。例小麦花药发育处于花粉母细胞形成前夕时遮光72h,花粉全部败育。")
49
(2)温度 温度由于影响光合作用、呼吸作用和物质转化和运输等过程而间接影响花芽的分化。一般在一定温度范围内,温度越高,花芽分化越快。低温下花粉母细胞往往分裂受阻;绒毡层细胞异常,不能向花粉粒供应能量,花粉败育。 例:水稻减数分裂时如遇17 ℃以下低温。花粉母细胞不能正常分裂,绒毡层细胞肥大,不能向花粉粒供应充足的能量,花粉败育。 (3) 水 小麦、水稻的孕穗期往往是植物对水分最敏感的时期,若此时缺水,就会影响雌雄蕊的分化。但夏季适度干旱可提高果树的C/N比,对形成花芽有利。
 水 小麦、水稻的孕穗期往往是植物对水分最敏感的时期,若此时缺水,就会影响雌雄蕊的分化。但夏季适度干旱可提高果树的C/N比,对形成花芽有利。")
50
(4)肥:氮素营养不足,花芽分化少且慢,但若氮素过多,C/N比失调,会造成植物贪青晚熟。增施磷肥可增加花数。在适宜的氮肥条件下,配合使用磷钾肥,并注意补Mn、Mo等微量元素,对开花有益。
二、性别分化与表达 (一) 植物性别表现类型:常见: 1、雌雄同花植物:大多数植物属雌雄同花植物,一朵花中既有雄蕊又有雌蕊,为两性花。如小麦、蕃茄、茶花等。 2、雌雄异株植物:只具雌花或雄花(单性花)。例杨、柳、桑 、银杏等。 3、雌雄同株异花植物:一株植株上既有雌花,又有雄花(单性花)。如瓜类:西瓜、甜瓜、玉米、丝瓜等。 4、雄性系或雌性系:指雌雄同株异花植物仅开雄花称雄性系;仅开雌花的称雌性系。常为人工栽培的特别品系。
 植物性别表现类型:常见: 1、雌雄同花植物:大多数植物属雌雄同花植物,一朵花中既有雄蕊又有雌蕊,为两性花。如小麦、蕃茄、茶花等。 2、雌雄异株植物:只具雌花或雄花(单性花)。例杨、柳、桑 、银杏等。 3、雌雄同株异花植物:一株植株上既有雌花,又有雄花(单性花)。如瓜类:西瓜、甜瓜、玉米、丝瓜等。 4、雄性系或雌性系:指雌雄同株异花植物仅开雄花称雄性系;仅开雌花的称雌性系。常为人工栽培的特别品系。")
51
(二)雌雄个体生理差异:雌雄株间: 1、呼吸代谢的差异:一般雄株的呼吸速率高于雌株。例大麻、桑、番木瓜等植物雄株的CAT酶活性比雌株高约50-70%,而银杏、菠菜雄株幼叶POD同工酶谱带少于雌株。由于呼吸代谢的差异,氧化还原能力上也有差异:一般雄株的氧化能力强,雌株的还原能力强。 2、光合作用的差异:一般雌株的RNA、叶绿素、胡萝卜素和碳水化合物含量高于雄株。 3、内源激素差异:如大麻雌株叶片内IAA含量高,而雄株叶片中的GA含量较高;玉米雌穗IAA水平高,而雄穗中GA含量高;雌雄异株的野生葡萄中,雌株的CTK含量高于雄株。

52
(三)外界条件对性别分化的调控: 植物性别的表达受基因的控制。多数雌雄异花或雌雄异株植物,其器官发育的早期均为两性的,但在发育的不同时期,一种性器官败育而另一种性器官得到完全发育至成熟。 1、光:光不仅能诱导开花,而且能调节花性别的分化:一般是适宜的光周期促进雌花的分化,而在非诱导光周期下,多开雄花。即SD促进SDP植物多开雌花,LDP多开雄花;LD促进LDP植物多开雌花,SDP多开雄花。 例:LDP菠菜,在光周期诱导后给予短日照,在其雌株上也能形成雄花。

53
水稻农垦58S:在短日条件下是可育的,但在长日条件下,其花粉完全败育,称光周期敏感核不育水稻。其与对日照长度敏感的—隐性雄性不育基因的表达有关,光敏素是这种水稻接受光周期的光敏受体,也称光敏素雄性不育水稻。 2、温度:昼夜温差大对雌花发育有利。例:夜间温度低利于菠菜、大麻、葫芦雌花的发育,对于番木瓜,低温下雌花占优势,高温下以雄花为主。但黄瓜夜间温度低时雌花分化少。 3、水肥:水肥充足多开雌花,土壤干旱、氮肥少,雄花分化多。

54
4、植物激素 (1)IAA、ETH、CTK:利于雌株的分化: GA:利于雄株分化。 (2)三碘苯甲酸(TIBA)、马来酰肼抑制雌花产生,诱导出 雄花;CCC:对抗GA长雌花。 5、机械损伤或烟熏 烟中含CO和乙烯,可抑制IAA氧化酶的活性,提高体内IAA水平,而多开雌花,植物损伤,增加乙烯产生,多开雌花。

55
第四节 受精生理 一、花粉的结构 1、花粉粒的发育
花粉母细胞减数分裂形成4个单核细胞称小孢子。小孢子继续分裂为二,形成较大的一个称营养细胞,较小的一个称生殖细胞(生殖核外有一团细胞质,整个游离于营养细胞的细胞质中),形成有二细胞花粉,一些原始的科如木兰科、百合科等的花粉为二细胞花粉。 在一些较进化的科如十字花科、菊科中,生殖细胞再分裂一次形成2个精细胞,加上一个营养细胞,构成三细胞花粉。两个互相连接的精细胞与营养核作为一个功能团被称作雄性生殖单位。成熟花粉粒也称雄配子体。
,形成有二细胞花粉,一些原始的科如木兰科、百合科等的花粉为二细胞花粉。 在一些较进化的科如十字花科、菊科中,生殖细胞再分裂一次形成2个精细胞,加上一个营养细胞,构成三细胞花粉。两个互相连接的精细胞与营养核作为一个功能团被称作雄性生殖单位。成熟花粉粒也称雄配子体。")
56
2、花粉粒的结构 有两层: (1) 外壁:较坚固,主要由纤维素、类黄酮素、孢粉素和外壁蛋白等物质组成。 孢粉素:是类胡萝卜素的氧化聚合物,其化学性能稳定,耐温、耐压、抗酸能力强,增加花粉粒的生存力。 外壁蛋白:其由绒毡层细胞合成,参于与柱头的相互识别。外壁上还有许多孔和沟,称萌发孔和萌发沟。 (2) 内壁:主要由纤维素、半纤维素、果胶质和活性内壁蛋白组成。活性内壁蛋白主要是些花粉萌发以及穿入柱头的有关酶类。萌发孔处的内壁蛋白十分丰富,内壁对水分子有很大的亲和力。
 内壁:主要由纤维素、半纤维素、果胶质和活性内壁蛋白组成。活性内壁蛋白主要是些花粉萌发以及穿入柱头的有关酶类。萌发孔处的内壁蛋白十分丰富,内壁对水分子有很大的亲和力。")
57
(3)营养细胞:含有丰富的细胞器和贮藏物质,营养细胞核膜上有较多的核孔。营养细胞的主要功能是积累与提供营养物质。
(4)生殖细胞:无细胞壁,游离于营养细胞的细胞质中,生殖细胞的细胞质稀少,几乎不含贮藏物质,核孔也少。 二、避免自花授粉: 花粉落到柱头上能否萌发,花粉管能否生长并通过花柱进入胚囊受精,取决于花粉与雌蕊的亲和性和识别反应,以或排斥远缘或异种花粉。 1、自交不亲和性:指植物花粉粒落在同花雌蕊的柱头上不能受精的现象。 研究表明,自交不亲和性受一系列复等位S基因的控制。当花粉粒和雌蕊组织具有相同的S等位基因时,就表现出不亲和性,反之亲和,以避免自花受精。
生殖细胞:无细胞壁,游离于营养细胞的细胞质中,生殖细胞的细胞质稀少,几乎不含贮藏物质,核孔也少。 二、避免自花授粉: 花粉落到柱头上能否萌发,花粉管能否生长并通过花柱进入胚囊受精,取决于花粉与雌蕊的亲和性和识别反应,以或排斥远缘或异种花粉。 1、自交不亲和性:指植物花粉粒落在同花雌蕊的柱头上不能受精的现象。 研究表明,自交不亲和性受一系列复等位S基因的控制。当花粉粒和雌蕊组织具有相同的S等位基因时,就表现出不亲和性,反之亲和,以避免自花受精。")
58
被子植物的自交不亲和性有2种表型: 1)配子体不亲和型(GSI): 它的不亲和性发生在花粉中,花粉管进入花柱后,中途生长停止、破裂。 如二核花粉的茄科、百合科植物,三核花粉的禾本科植物为GSI型。配子体不亲和型与雌蕊花柱S基因编码的S-核酸酶有关,S-核酸酶能进入花粉管,把花粉管内的RNA水解,花粉管就停止生长。 2)孢子体不亲和型(SSI型): 表现为花粉和柱头相互作用,阻碍花粉的水合作用,而不能形成花粉管。三核花粉的十字花科、菊科植物属SSI型。
孢子体不亲和型(SSI型): 表现为花粉和柱头相互作用,阻碍花粉的水合作用,而不能形成花粉管。三核花粉的十字花科、菊科植物属SSI型。")
59
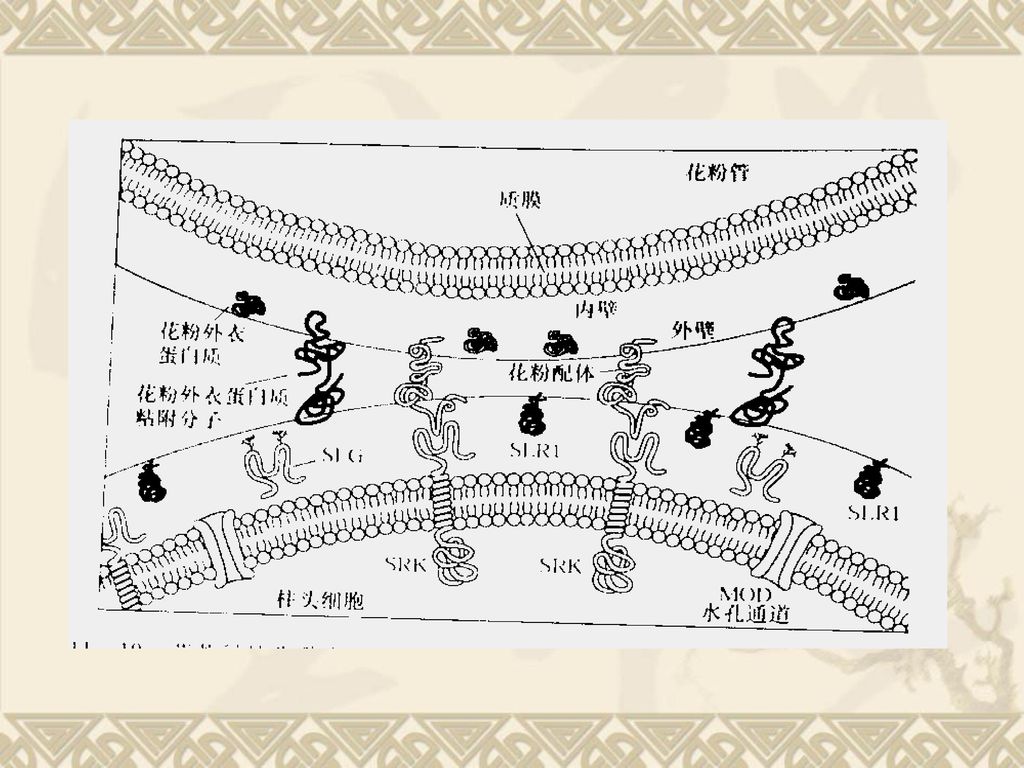
孢子体不亲和型与雌蕊的类受体蛋白激酶有关。研究比较清楚的有两个基因:
(1)S基因座糖蛋白(S locus glycoprotein, SLG)基因,编码糖蛋白并分泌到柱头乳突细胞的细胞壁和胞间区。SLG表达时间与植物自交不亲和性的反应时间相关。 (2)S基因座受体激酶(S locus receptor kinase,SRK)基因,编码一种跨膜的类受体蛋白激酶,它主要由两个区域组成,一是与质膜蛋白质相连的胞外区,另一个是与细胞质丝氨酸/苏氨酸激酶相连的胞内区。生化研究表明,SRK参与了柱头上的磷酸化过程,介导了自交不亲和性。
S基因座糖蛋白(S locus glycoprotein, SLG)基因,编码糖蛋白并分泌到柱头乳突细胞的细胞壁和胞间区。SLG表达时间与植物自交不亲和性的反应时间相关。 (2)S基因座受体激酶(S locus receptor kinase,SRK)基因,编码一种跨膜的类受体蛋白激酶,它主要由两个区域组成,一是与质膜蛋白质相连的胞外区,另一个是与细胞质丝氨酸/苏氨酸激酶相连的胞内区。生化研究表明,SRK参与了柱头上的磷酸化过程,介导了自交不亲和性。")
60
2、经生理或形态的改变达到 1)单性花(unisexual flower):具有单性花的植物必须进行异花传粉。如雌雄同株的玉米、瓜类等,雌雄异株的桑、大麻、杨、柳、菠菜等。 2)雌雄蕊异熟(dichogamy):指一株植物或一朵花上的雌雄蕊成熟时间不一致。如: 玉米的雄花序比雌花序先熟。梨、苹果的两性花雄蕊先熟,而油菜的两性花则雌蕊先熟。 3)雌雄蕊异长(heterogony): I)二型花柱植物:如荞麦、报春花的花柱有两种类型:一种为雄蕊高于花柱,另一种为雌蕊花柱高于雄蕊。只有高雄蕊花药掉到高柱头上或低雄蕊的花粉粒传到低柱头上才能受精。
雌雄蕊异熟(dichogamy):指一株植物或一朵花上的雌雄蕊成熟时间不一致。如: 玉米的雄花序比雌花序先熟。梨、苹果的两性花雄蕊先熟,而油菜的两性花则雌蕊先熟。 3)雌雄蕊异长(heterogony): I)二型花柱植物:如荞麦、报春花的花柱有两种类型:一种为雄蕊高于花柱,另一种为雌蕊花柱高于雄蕊。只有高雄蕊花药掉到高柱头上或低雄蕊的花粉粒传到低柱头上才能受精。")
61
II)三型花柱植物: 如千屈菜属植物,花柱和雄蕊分别有长、中、短3种,只须相同高度的雄蕊和雌蕊间才能传粉,否则传粉不孕。 4)雌雄蕊异位(herkogamy) 指雌、雄蕊通过在空间的排列位置不同,避免自花受粉。 如石竹科植物: 两性花的雌蕊高于雄蕊,花粉不易传到柱头上。再如百合科的嘉兰属植物,雌蕊的花柱基部在近子房处呈直角状向外伸出,远离雄蕊,避免自花传粉。

63
三、花粉的萌发和花粉管的生长 1、花粉粒的萌发 成熟花粉从花粉囊中散出,落到柱头上的过程称授粉。栽培上,将异株异花间的授粉称异花授粉,而同株异花、同株同花间的授粉称自花授粉。 花粉落到柱头上,相互识别以后,吸收柱头表皮细胞分泌物和水分,由于营养细胞的吸胀作用使花粉内壁及营养细胞的质膜在萌发孔处向外突出,形成花粉管的乳状顶,此过程称花粉萌发。

64
植物能否成功授粉依赖于柱头表面与花粉的粘附能力和水合能力。
1)柱头若是干性的,授粉时柱头就不产生分泌物,由于花粉的胞被含脂质,可与柱头细胞反应引起水合作用而形成花粉管,如芸苔属植物的柱头。 2)若是湿性柱头,柱头能分泌脂肪富集的分泌物,刺激水合作用。 2、花粉管的生长 花粉粒一方面消耗自身的贮藏物质,另一方面从花柱中吸收营养供花粉管的生长和新壁的合成。花粉管的生长一般局限于花粉管顶端区。
柱头若是干性的,授粉时柱头就不产生分泌物,由于花粉的胞被含脂质,可与柱头细胞反应引起水合作用而形成花粉管,如芸苔属植物的柱头。 2)若是湿性柱头,柱头能分泌脂肪富集的分泌物,刺激水合作用。 2、花粉管的生长. 花粉粒一方面消耗自身的贮藏物质,另一方面从花柱中吸收营养供花粉管的生长和新壁的合成。花粉管的生长一般局限于花粉管顶端区。")
66
研究指出,花粉管顶端区代谢旺盛,内含有新壁合成的相关细胞器如线粒体、内质网、高尔基体等,用于花粉管的伸长;另外花粉管从顶端到基部还存在由高到低的Ca2+浓度梯度,以保持顶端的极性生长。为减少生长阻力,花粉管还向柱头分泌许多水解酶,如角质酶、淀粉酶、转化酶等。 花粉管穿过柱头,通过花柱向子房生长。 1)在空心的花柱中,花柱中央有1到数条纵行的沟称花柱道,花粉管沿着花柱道表面上的分泌物生长,如百合。 2)在实心的花柱中,常在花柱中央引导组织细胞间隙的分泌物中生长,若无引导组织还可经过中央薄壁细胞中穿过。
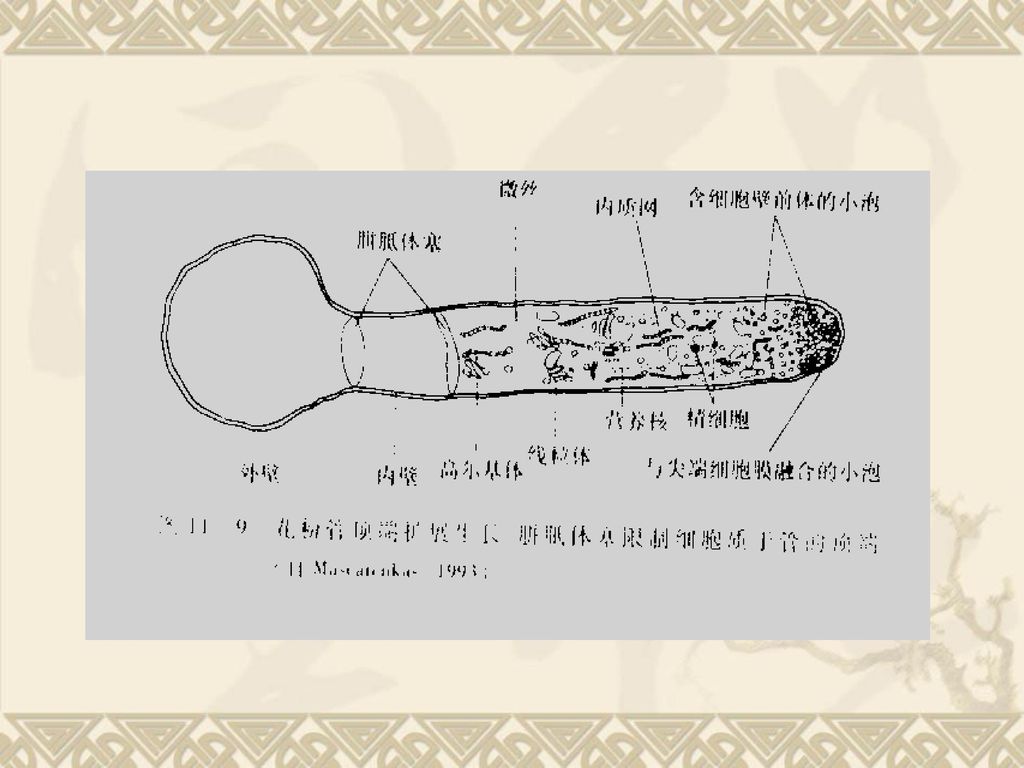
在空心的花柱中,花柱中央有1到数条纵行的沟称花柱道,花粉管沿着花柱道表面上的分泌物生长,如百合。 2)在实心的花柱中,常在花柱中央引导组织细胞间隙的分泌物中生长,若无引导组织还可经过中央薄壁细胞中穿过。")
67
3 、花粉管定向伸长机理 花粉管总是朝着胚囊组织方向生长。现在研究表明:这种定向生长与助细胞释放的Ca2+有关。花粉管会释放出一些信号物质如GA,可引起助细胞首先解体,助细胞的解体会释放大量的Ca2+,造成花柱与珠孔间的Ca2+浓度梯度,而Ca2+是一种向化性物质,花粉管会朝着Ca2+浓度高的方向生长,最后穿过珠孔,进入胚囊。Machlis(US)也发现金鱼草从柱头到子房Ca2+浓度也依次递增。 认为花粉管的定向生长是与雌蕊组织中的向化性物质Ca2+有关,此物在雌蕊组织中的浓度不同,花粉管总是朝着雌蕊组织中Ca2+浓度递增的方向伸长。 在花柱上的花粉密度大,对花粉的萌发和花粉管的伸长有明显的刺激作用,称花粉的集体(群体)效应。
也发现金鱼草从柱头到子房Ca2+浓度也依次递增。 认为花粉管的定向生长是与雌蕊组织中的向化性物质Ca2+有关,此物在雌蕊组织中的浓度不同,花粉管总是朝着雌蕊组织中Ca2+浓度递增的方向伸长。 在花柱上的花粉密度大,对花粉的萌发和花粉管的伸长有明显的刺激作用,称花粉的集体(群体)效应。")
68
四、受精生理 (一)受精过程 随着花粉管的伸长,三细胞花粉的2个精细胞、1个营养核和细胞质全部移入花粉管,通常营养核在前,2个精细胞在后,将内含物全部释放给胚囊。二核花粉生殖细胞在花粉管中再分裂一次,产生的2个精子与营养核也一并进入花粉管。花粉管到达子房后,一般沿着子房的内壁生长,直达胚珠,经珠孔、合点或中部受精。 (二)受精前后雌蕊的代谢变化 花粉落到雌蕊柱头上,在摄取柱头和花柱物质的同时,还主动分泌一些物质到雌蕊,其中主要是酶,例磷酸化酶、淀粉酶、转化酶等。以取得营养保证花粉管的生长。等到双受精结束,花粉将全部营养交给了雌蕊。雌蕊组织的代谢发生明显的变化。
受精前后雌蕊的代谢变化. 花粉落到雌蕊柱头上,在摄取柱头和花柱物质的同时,还主动分泌一些物质到雌蕊,其中主要是酶,例磷酸化酶、淀粉酶、转化酶等。以取得营养保证花粉管的生长。等到双受精结束,花粉将全部营养交给了雌蕊。雌蕊组织的代谢发生明显的变化。")
69
1、增加水分和无机盐的吸收 例:兰科植物授粉以后,合蕊柱吸水增加1/3,N、P含量显著增加。再例:玉米授粉后大量32P由植物的其它部分流入雌蕊,使雌蕊磷含量增加0.7倍. 2、雌蕊组织呼吸加强 受精后的雌蕊的呼吸速率一般要比受精前或未受精的高。例:棉花受精时雌蕊的呼吸速率增高2倍;百合花的呼吸速度在授粉后出现2次高峰,一次在精卵细胞发生融合时,另一次在胚乳游离核分裂旺盛期。

70
3、雌蕊中的IAA、CTK含量显著增加 例:烟草授粉20h,花柱中的生长素含量增加3倍多。且合成部位由花柱顶端向子房转移,即由花柱顶部—花柱基部—子房。另外发现CTK也明显增加。 雌蕊中的IAA一部分由花粉带入,绝大部分由花柱和子房合成。花粉将由Try合成IAA的酶系统分泌到了雌蕊,就促进了雌蕊IAA的合。 IAA、CTK含量的增加,会促进合子与初生胚乳的分裂与生长,促使营养物质向子房的运输。 在生产上,用2,4-D、NAA、GA 和CTK处理未受精的子房,可使营养向子房输送,产生无籽果实。不经受精作用而形成不含种子的果实称单性结实。

71
(三)影响受精的因素 1、花粉的活力 成熟花粉在离开花药以后可维持一定时间的生命力。我们将离体花粉维持具有受精能力的时间称花粉的寿命。各种植物花粉寿命不同,例水稻花粉为5-10min,小麦15-30min,玉米1d,梨、苹果70-210d。一般二核花粉寿命比三核花粉长,因三核花粉代谢活跃。果树花粉寿命长。刚从花约中散失的成熟花粉活力最强。 外界环境条件影响花粉的寿命,在高温、高氧、高湿条件下花粉易失活。因为此时花粉的代谢旺盛,营养物质消耗多;酶活性衰退;易受病菌感染,积累有毒物质会加速花粉的衰老与死亡。

72
反之,低湿、低温、低氧有利于花粉的贮藏。通常采用的温度:1-5度;相对湿度:10-40%。例苹果花粉:在3度、10-25%相对湿度的环境中可保存350d,仍有60%以上的萌发率。此外,光线对花粉的贮存也有影响,一般以遮荫或黑暗中贮存较好。 2、柱头活力 柱头活力关系到花粉落到柱头上能否萌发,花粉管能否伸长,所以柱头生活力直接关系到受精的成败。柱头活力持续时间的长短因植物种类而异: 例:水稻能持续6-7d,以开花当天活力最强,以后逐渐下降。小麦柱头可维持9d,但在麦穗抽出第3天结实率最高,到第6d下降。

73
3、环境条件 1)温度:温度影响花药的开裂,也影响花粉的萌发和花粉管的伸长。如水稻,15℃时花药不能开裂,40-50℃时花药易干燥,柱头易失活,25-30℃是受精的最适温度。 2)湿度:花粉萌发需要一定的湿度,湿度过小,会影响花药的生活力和花丝的生长,并使雌蕊的花柱和柱头干枯;湿度过大或雨天开花,花粉会过度吸水而破裂。一般湿度在70-80%较合适。 3)矿质营养:B能促进花粉的萌发和花粉管的伸长。一方面硼促进糖的吸收和代谢,另一方面硼参与果胶物质的形成,有利于花粉管的形成。 4)田间管理:株间通风、透光利于雌雄蕊的发育胺精。
湿度:花粉萌发需要一定的湿度,湿度过小,会影响花药的生活力和花丝的生长,并使雌蕊的花柱和柱头干枯;湿度过大或雨天开花,花粉会过度吸水而破裂。一般湿度在70-80%较合适。 3)矿质营养:B能促进花粉的萌发和花粉管的伸长。一方面硼促进糖的吸收和代谢,另一方面硼参与果胶物质的形成,有利于花粉管的形成。 4)田间管理:株间通风、透光利于雌雄蕊的发育胺精。")
Similar presentations







 安阳市实验中学 宋军芬 1 、 观察桃花的结构,由外到内依次生有 ____ 、 ____ 、 ____ 、 ____ 、 ____ 、 ____ 。 2 、雄蕊包括 ____ 、 ____ 。 雌蕊包括 ____ 、 __ _ 、 。 课堂预习: 观察课本第 116 页桃花结构图,完成学案。>")
:核蛋白体组成成分 转移 RNA ( tRNA ):转运氨基酸 信使 RNA ( mRNA ):蛋白质合成模板 不均一核 RNA ( hnRNA ):成熟 mRNA 的前体 小核 RNA ( snRNA ):>")











