Download presentation
1
第十一章 表观遗传学 (Epigenetics)
")
2
遗传学:是研究遗传信息的组织、表达和传递的科学,
也是揭示生命本质和遗传规律的科学 基因(gene):含有遗传信息的可遗传单位,是编码一 条多肽链的特定DNA片段。 基因型(Genotype):个体的遗传结构; 表现型(Phenotype):环境条件与基因型相互作 用而使个体呈现的性状。 性状遗传符合一定的规律
:含有遗传信息的可遗传单位,是编码一. 条多肽链的特定DNA片段。 基因型(Genotype):个体的遗传结构; 表现型(Phenotype):环境条件与基因型相互作. 用而使个体呈现的性状。 性状遗传符合一定的规律.")
3
机体是由多种不同类型的细胞组成; 机体不同的细胞具有相同的基因型; 机体不同的细胞:形态存在差异 执行不同的生理功能 基础:基因表达模式(expression pattern) 不同。 组织细胞特异性的表达模式通过细胞分裂来传递并稳定维持。
 不同。 组织细胞特异性的表达模式通过细胞分裂来传递并稳定维持。 .")
4
生物中的非孟德尔遗传现象 果蝇中的杂色(眼)位置效应(position-effect variegation): 野生红眼基因W+(显性)
基因定位于X染色体长臂末端

5
“W+/W+”和“W+/w”均表现正常红眼

6
“W+/w”杂合体表现为花斑眼(杂色),即:部分细胞正常红色,部分少量红色,部分白色。
7
表观遗传存在的现象

8
遗传印迹现象 “马骡”和“驴骡”有较大差异 人胚胎三倍体的自然流产现象:多出的染色体父、母源性不同,则产生不同的病理特征。

9
遗传印迹现象 马骡 驴骡 差异较大

10
The history of epigenetics is linked with the study of evolution and development.
The meaning of the term “epigenetics” has itself undergone an evolution. Present working definition: the study of mitotically and/or meiotically heritable changes in gene function that cannot be explained by changes in DNA sequence. (Riggs et al.1996)
")
11
表观遗传学(epigenetics):是研究没有DNA序列变化的、可以遗传的表达改变(Wolffe,1999)
表观遗传(epigenetic inheritance):通过有丝分裂和减数分裂来传递非DNA序列信息的现象。
:通过有丝分裂和减数分裂来传递非DNA序列信息的现象。")
12
人类基因组含有两类信息 传统意义上的遗传学信息,它提供生命所必需的所有蛋白质合成的蓝图;
表遗传(epigenetic)信息,提供何时、何地、如何应用遗传学信息的指令,保证基因适时启闭 One genome multiple epigenome
信息,提供何时、何地、如何应用遗传学信息的指令,保证基因适时启闭. One genome multiple epigenome.")
13
一、表观遗传修饰 表达模式的信息标记: DNA特定碱基的修饰:胞嘧啶的甲基化; 染色质构型重塑:如,组蛋白的乙酰化、甲基化

14
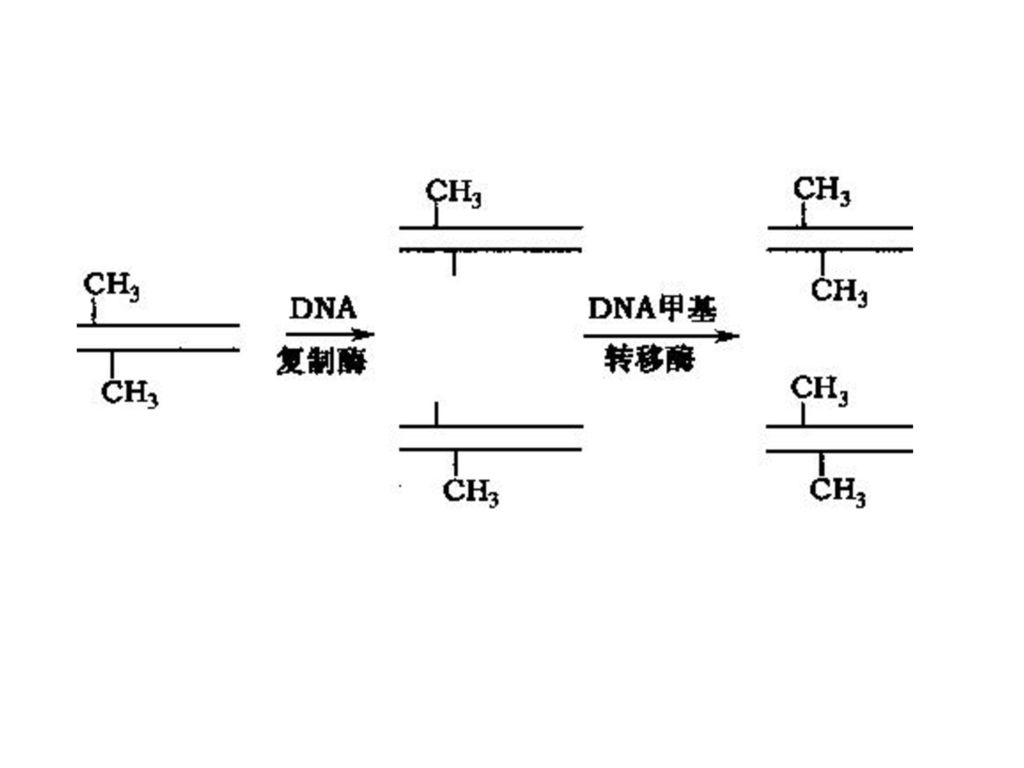
表观遗传修饰机制1:DNA甲基化 DNA甲基化:基因组DNA上的胞嘧啶第5位碳原子和甲基的共价结合—5-甲基胞嘧啶( 5-methylcytosine, 5-mC)。 DNA甲基化通过DNA甲基转移酶(DMTs)来完成,将S-腺苷甲硫氨酸上的甲基→胞嘧啶的第5位碳原子。 5-mC主要发生在CpG二联核苷。 CpG二联核苷常常以成簇串连的形式存在于结构基因的调控区段。 CpG岛(CpG islands):结构基因5‘端富含CpG二联核苷的区域。 DNA甲基化→基因沉寂; DNA非甲基化→基因活化。
:结构基因5‘端富含CpG二联核苷的区域。 DNA甲基化→基因沉寂; DNA非甲基化→基因活化。")
15
DNMT (de novo methylase):从头甲基化酶
SAM (S-adenosylmethiomine): S-腺苷甲硫氨酸
: S-腺苷甲硫氨酸.")
18
表观遗传修饰机制2:染色质重塑 组蛋白的化学修饰:乙酰化、甲基化 (1)组蛋白中不同氨基酸残基的乙酰化一般与活化的
染色质构型和有表达活性的基因相关联; (2)组蛋白中氨基酸残基的甲基化与浓缩的异染色质 核基因表达受抑有关。 也有例外: 组蛋白甲基化抑制或激活基因表达取决于被修饰的赖氨酸的位置, 如, H3 Lys9甲基化→基因沉寂; H3 Lys4甲基化→基因激活.
组蛋白中氨基酸残基的甲基化与浓缩的异染色质. 核基因表达受抑有关。 也有例外: 组蛋白甲基化抑制或激活基因表达取决于被修饰的赖氨酸的位置, 如, H3 Lys9甲基化→基因沉寂; H3 Lys4甲基化→基因激活.")
19
表观遗传修饰调控的复杂性 机体表观遗传模式的变化在整个发育过程中高度有序/严格受控: RNA干涉(影响组蛋白修饰) 组蛋白结构修饰 调控网络
组蛋白结构修饰 调控网络 DNA甲基化 动态而有序地调控组织细胞特异性的基因表达模式

20
RNA 干涉? 2006年10月2日,瑞典皇家卡罗林医学院宣布,本年度诺贝尔生理学和医学奖的桂冠授予两位美国人:
A.Z. Fire (安德鲁·法尔) C.C. Mello (克雷格·梅洛) 他们获奖,是因为发现了RNA的干扰机制。 诺贝尔奖评委会的公报说, Fire 和 Mello 获奖是因为他们“发现了控制遗传信息流动的基本机制。”
 C.C. Mello (克雷格·梅洛) 他们获奖,是因为发现了RNA的干扰机制。 诺贝尔奖评委会的公报说, Fire 和 Mello 获奖是因为他们 发现了控制遗传信息流动的基本机制。")
21
Fire 和 Mello (1998) 提出双链RNA(dsRNA )可以作为调控元件而触发基因的沉寂
当时普遍认为反义RNA诱导的相关基因沉寂是通过与互补mRNA结合,从而阻碍蛋白质的合成。 Fire 和 Mello 的工作推翻了这一定论。

22
实验性细胞内引入RNA 以干扰内源性基因的功能。
当时一般认为这种 RNA 干扰依赖于引入的RNA与mRNA杂交,以反义链封闭mRNA的翻译。 设计实验拟解决:“RNA 干扰”是否与转入的RNA结构有关。

23
意外发现:导入双链RNA的产生功能干扰的有效性远高于导入单链RNA, sense or antisense RNA导入均如此。
仅需少数分子即可产生干扰效应,提示酶促反应或分子扩增的存在。

24
上述现象提示: 1. 存在超越简单反义RNA作用的机理。 2. RNA靶向的作用也不能排除。 3. 同时可能存在RNA与染色质的直接作用,影响RNA的转录。

26
miRNAs与siRNAs区别 分子起源: miRNAs来源于特异性前体,在基因组内具有明确的编码序列;
siRNAs 可能更随机的来源于较长的dsRNAs ,它亦可由外源导入或内源基因的双向转录来源。 作用机制: 多数情况下,miRNAs 与靶mRNA 3‘-UTRs 区多位点非精确的互补结合; siRNAs常与靶mRNA单个位点精确配对,导致靶mRNAs 裂解。

27
Lin-4的分子特征

28
miRNA和siRNA 的生物发生和调节特征比较

29
siRNAs和miRNAs 在分子特征,生物发生和发挥功能作用的机制均相似。
目前将两者割裂为两类分子,这更多体现了人为因素的存在,可能与它们的发现过程不同有关。 事实上,今天有关miRNAs 认知,其中许多来源于对siRNAs和 RNAi的认识途径。

30
Distinction between Euchromatic and Heterochromatic Domains
RITS: RNA-induced transcriptional silencing effector Distinction between Euchromatic and Heterochromatic Domains

31
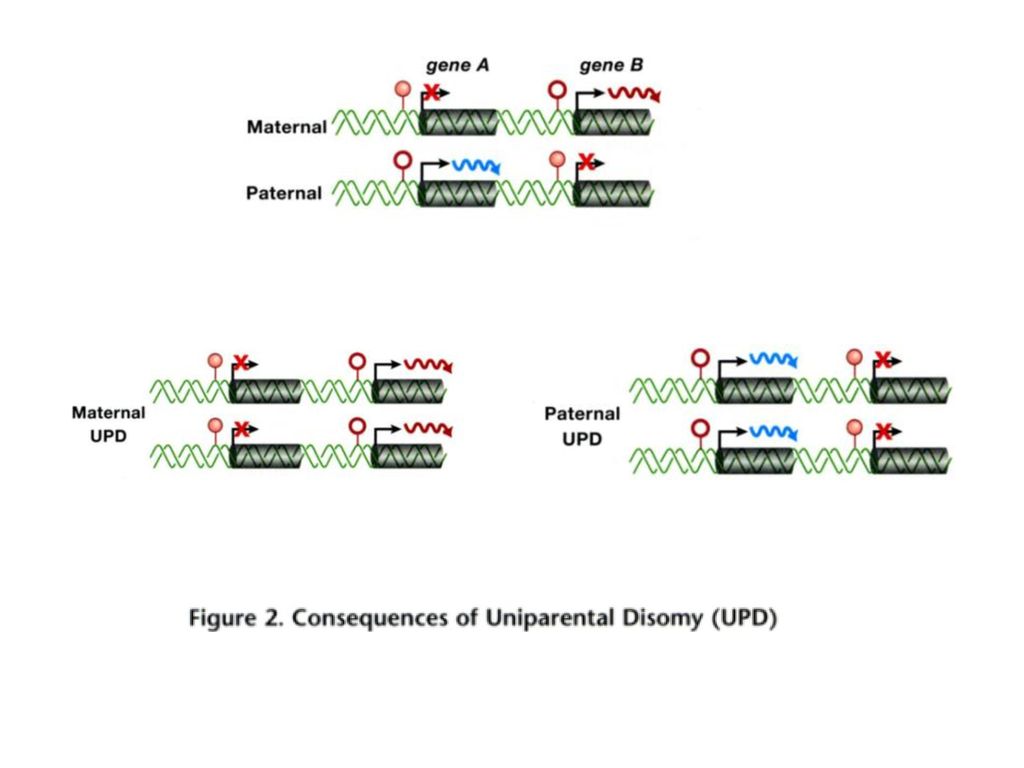
二、基因组印迹(genomic imprinting)
概念:依赖于父、母源性的等位基因的差异性表达,即父亲和母亲的基因组在个体发育中有着不同的影响,这种现象称基因组印迹。 两个亲本的等位基因差异性甲基化是基因组印迹现象的基础。

32
疾病的基础:15q11-13 微缺失 Angelman syndrome, AS(母源):共济失调, 严重智力低下
Prader-Willi syndrome, PWS(父源):肥胖、矮小, 中度智力低下
33
Both syndromes can be caused by genetic or epigenetic defects
Uni-parental disomy Both syndromes can be caused by genetic or epigenetic defects

35
基因组改变: 微缺失的关键区域有成簇排列的,富含CpG岛的基因表达调控元件—— 印迹中心(imprinting centers, ICs)
父源非甲基化 父源 母源 染色体上的ICs呈现差异甲基化 母源甲基化 人类基因组已发现ICs 50多个,分布于基因组的12 个区 域,大多成簇排列,与疾病发生相关。 Beckwith-Wiedemann syndrome, BWS:11p15.5

36
印迹调控区 IGF2:胰岛素样生长因子2 P:启动之 H19:胎肝文库第19 cDNA克隆 E:增强子 DMR1:差异甲基化区域
CTCF:锌指蛋白 P:启动之 E:增强子 :非甲基化CpG :甲基化CpG 父源等位基因表达 母源等位基因表达

37
CTCF(CCCTC-binding factor)——Transcriptional Repressor
Transcriptional insulators are DNA elements that set boundaries on the actions of enhancer and silencer elements All vertebrate insulators appear to use the versatile CTCF protein. CTCF uses various combinations of its 11 zinc fingers to recognize a variety of unrelated DNA sequences. Once bound to DNA, CTCF can function as a transcriptional insulator, repressor, or activator, depending on the context of the binding site

38
印迹基因的DNA甲基化型都是在生殖细胞成熟过程中建立的。
基因组印迹是性细胞系的一种表观遗传修饰,是一整套分布于染色体不同部位的印迹中心来协调。 印迹中心直接介导了印迹标记的建立及其在发育过程中的维持和传递,导致亲本来源特异性的优先表达两亲本等位基因中的一个。

39
印迹基因的特点 印迹基因的群集性:目前已知的印迹基因在基因组中不是单一基因的分散分布,而是多基因簇集,其中部分父源表达、部分母源表达。
印迹基因DNA复制的不同步性。

40
雄性生殖系 雌性生殖系 父系染色体 母系染色体 合子 亲代基因组印迹在生殖系的重新编程 父系配子 母系配子

41
Key features of genomic imprinting in mammals
cis-Acting mechanism A consequence of inheritance Imprints are epigenetic modification acquired by one parental gamete Imprinted genes are mostly clustered together with a noncoding RNA Imprints can modify long-range regulatory elements that act on multiple genes Imprinted genes play a role in mammalian development

42
个体发育过程中 表观基因组的重编程 所有结构和功能各不相同的细胞虽具有完全一样的基因组,却有着很不一样的表达模式。
建立和维持细胞特异性表达模式的信息,必须可以通过细胞分裂而遗传,同时应具备被删除和重建的可能性。

43
早期原始生殖细胞(primordial germ cells, PGC)携带体细胞样的表观遗传型。在PGC进入性腺前后,这一表观基因组开始被删除。
之后,性别特异性和序列特异性的表观遗传型在两性生殖细胞中被建立。 受精后,合子基因组在卵细胞质的生理环境中,启动与胚胎发育相关、有严格时空特异性的基因表达程序。

44
删除生殖细胞成熟过程中建立的、除印迹基因以外的全部表观遗传修饰标记,重新建立胚胎发育特有的表观基因组。
表观遗传修饰的重新编程对环境变化非常敏感:辅助生育干预了配子形成和胚胎发育早期(表观遗传编程和维持的关键时期),注意条件的控制十分重要。
,注意条件的控制十分重要。")
45
三、X染色体失活—表观遗传修饰 1.表观遗传证据: 雌性哺乳动物细胞的一条X染色体失活,且呈异染色质状态;
2.可能机制:Xq13.3存在X染色体失活中心(X-inaction center, Xic), X染色体失活启动区段。
, X染色体失活启动区段。")
46
Xic区段含4个基因,约1Mb大小 Xist( X-inactive specific transcript):X染色体启动转录最早的基因,没有“开读框架”(ORF),约17Kb 大小。转录后只有一条染色体的Xist RNA将这条染色体包裹,并启动异染色质化。 Xce(X chromosome controlling element):与X染色体“随机”失活的选择有关,Xce纯合时,随机失活;杂合时,非随机。 Xsix(瞬时调控元件):CTCF结合位点,表达仅在X染色体失活发生前,可能起到Xist的外源开关功能。 DXPas34:富含CpG.
:与X染色体 随机 失活的选择有关,Xce纯合时,随机失活;杂合时,非随机。 Xsix(瞬时调控元件):CTCF结合位点,表达仅在X染色体失活发生前,可能起到Xist的外源开关功能。 DXPas34:富含CpG.")
47
两条X染色体表达不稳定的Xist基因RNA:

48
Polycomb

49
四、表观遗传与疾病 表观遗传修饰异常引起的疾病可分为两类:
(1)表观突变(epi-mutation):发育的重新编程过程中造成的特定基因表观遗传修饰的异常。 (2)表观遗传修饰的分子构件与功能相关蛋白质编码基因的突变。 DNA甲基转移酶基因突变或表观突变 差异甲基化CpG岛结合蛋白CTCF的突变或表观突变
表观突变(epi-mutation):发育的重新编程过程中造成的特定基因表观遗传修饰的异常。 (2)表观遗传修饰的分子构件与功能相关蛋白质编码基因的突变。 DNA甲基转移酶基因突变或表观突变. 差异甲基化CpG岛结合蛋白CTCF的突变或表观突变.")
50
表观遗传病: 由表观遗传修饰异常而导致的疾病。
染色体病 单基因遗传病 多基因遗传病 线粒体遗传病 表观遗传病: 由表观遗传修饰异常而导致的疾病。

51
B. Hagberg(1983)报道Rett 综合征 表现为一种进行性神经系统病变,脑功能的迅速恶化和严重痴呆。
病因:X染色体编码MeCP2蛋白质的基因发生突变。 机制:MeCP2是一种甲基结合蛋白,能专一性地识别甲基化CpG岛并与之结合,作为一种分子“榫头”将染色质修饰复合物和DNA甲基化域连接在一起,发挥阻遏基因转录的功能。

52
A Rett syndrome patient illustrates the unusual stereotyped hand movements, teeth grinding, and abnormal posture.

53
ICF(immunodeficiency,centromeric region instability and facial anomalies syndrome )综合征
表现:不同程度的免疫缺陷,伴有面部畸形和智力低下。(常染色体隐性遗传) 病因:催化DNA从头甲基化的DNMT3B编码基因的突变,造成随体序列的低甲基化,以及环着丝粒序列的不稳定。 机制: DNMT3B基因的突变通过降低转甲基化活性而减少了对基因转录的阻遏作用,从而间接影响了淋巴细胞部分基因的表达模式。
 病因:催化DNA从头甲基化的DNMT3B编码基因的突变,造成随体序列的低甲基化,以及环着丝粒序列的不稳定。 机制: DNMT3B基因的突变通过降低转甲基化活性而减少了对基因转录的阻遏作用,从而间接影响了淋巴细胞部分基因的表达模式。")
54
脆性X综合征(Fragile X syndrome)
病因:X染色体智障基因(fragile X mental retardation-1, FMR1)5‘非翻译区CCG重复序列的扩展。 机制: (CGG)n重复序列的扩展会引起CGG中二联核苷的甲基化,使FMR1基因沉默,并使染色质构型发生改变。 表观遗传病(epigenetic diseases):包括复杂的遗传综合征、印迹综合征、免疫性疾病、神经发育性疾病、衰老和癌症等。
5‘非翻译区CCG重复序列的扩展。 机制: (CGG)n重复序列的扩展会引起CGG中二联核苷的甲基化,使FMR1基因沉默,并使染色质构型发生改变。 表观遗传病(epigenetic diseases):包括复杂的遗传综合征、印迹综合征、免疫性疾病、神经发育性疾病、衰老和癌症等。")
55
Fragile X Syndrome(FRAXA)
")
56
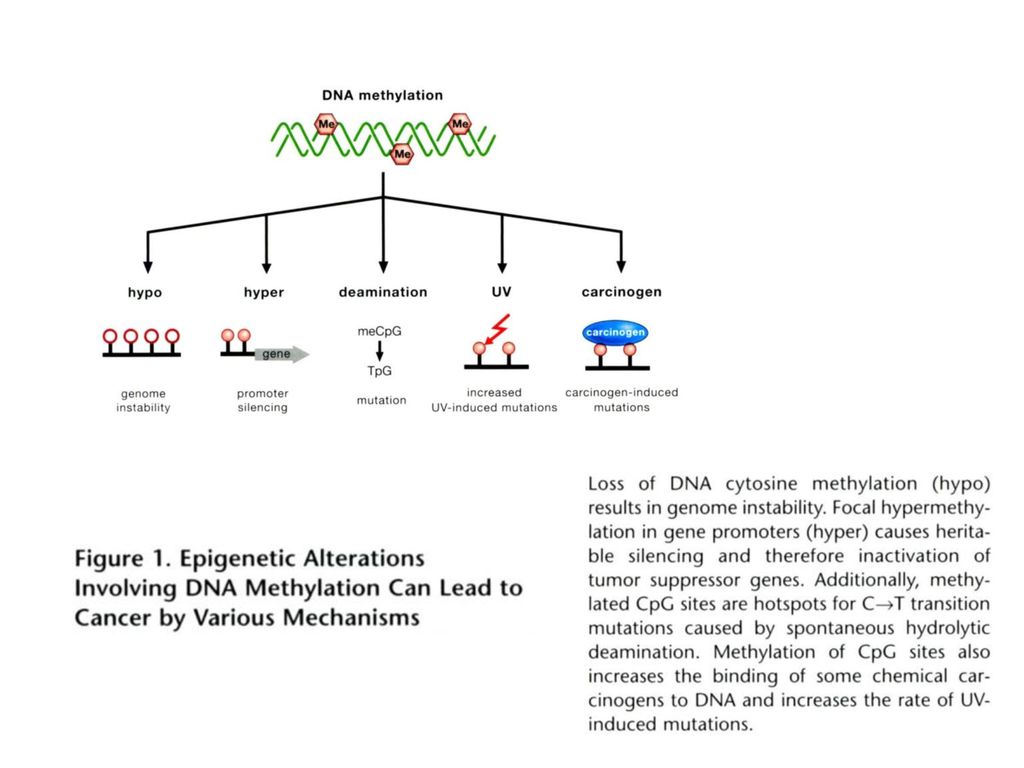
五、表观遗传与肿瘤 1979年Holliday提出DNA甲基化可能在癌症过程中起重要作用。
(2)人类基因组的内源性转座子等可移动因子的激活可与细胞的遗传不稳定相关联:肺癌细胞DNA总体甲基化水平的降低与K-ras等基因的激活。
人类基因组的内源性转座子等可移动因子的激活可与细胞的遗传不稳定相关联:肺癌细胞DNA总体甲基化水平的降低与K-ras等基因的激活。")
57
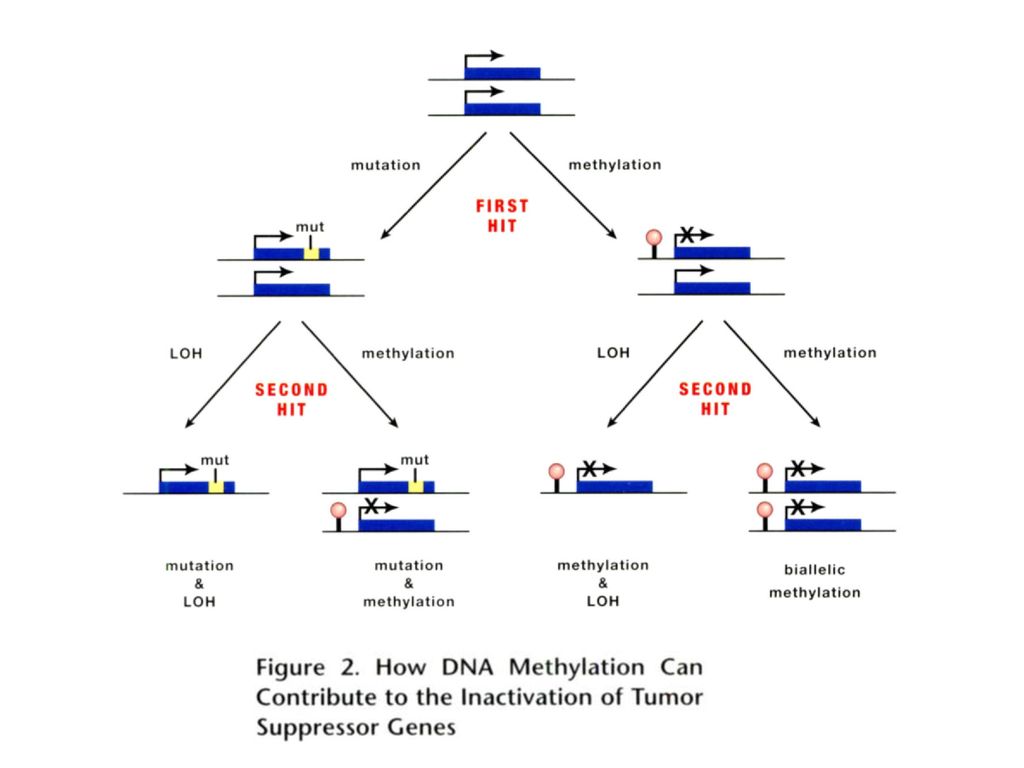
DNA甲基化与癌症

58
癌细胞局部序列的高甲基化 抑癌基因启动子区的高甲基化:如APC基因、VHL基因等
细胞周期相关基因、DNA损伤修复基因可呈现肿瘤特异性甲基化(基因启动子等调控元件的CpG岛)——肿瘤的极早期事件。 表观遗传病与肿瘤相关:BWS患者的Wilm发病率是常人的1000倍。 IGF2等基因的印迹丢失可提高个体肺癌、神经胶质瘤、乳腺癌的发病风险。
——肿瘤的极早期事件。 表观遗传病与肿瘤相关:BWS患者的Wilm发病率是常人的1000倍。 IGF2等基因的印迹丢失可提高个体肺癌、神经胶质瘤、乳腺癌的发病风险。")
59
癌细胞局部序列的高甲基化的原因 抑癌基因的甲基化可能渐进发展:特定CpG岛从头甲基化 更多CpG岛甲基化基因沉寂。
实验提示:最初的甲基化可能源自在DNA自发损伤中产生的5-甲基脱氧单磷酸胞嘧啶(5mdCMP)错误掺入。 5mdCMP由专项酶脱氨后转变为脱氧单磷酸胸腺嘧啶(dTMP)。 分析表明:癌细胞中5mdCMP脱氨酶活性降低,导致5mdCMP衍生为5mdCDP、5mdCTP形式掺入。
错误掺入。 5mdCMP由专项酶脱氨后转变为脱氧单磷酸胸腺嘧啶(dTMP)。 分析表明:癌细胞中5mdCMP脱氨酶活性降低,导致5mdCMP衍生为5mdCDP、5mdCTP形式掺入。")
62
The early role for abnormal DNA methylation in tumor progression

63
六、表观遗传与衰老 年龄相关(衰老)性疾病多表现为局灶性病变。
衰老过程中某些细胞会发生年龄相关性变化,如某个基因CpG岛的从头甲基化,从而关闭相关基因,丧失该基因的生理功能。 另一方面,个别基因甲基化的丢失也会激活正常情况下的沉寂基因,造成相关基因不恰当的“异位表达”。

64
例:动脉粥样硬化 是一种局灶性增生性疾病,存在表遗传因素,平滑肌细胞增值可使血管变窄。 曾观察到年龄相关的基因(ERα)启动子区甲基化。
异常甲基化的细胞可使组织或器官呈现表观遗传上的异质性和镶嵌性。 这种年龄相关的表遗传镶嵌性可能是血管上皮细胞、平滑肌细胞局灶性病变的重要病因。 胰岛素抵抗性糖尿病:胰岛素受体信号传导相关系列基因的表观遗传异常。

65
表观遗传分析在医学中的潜在应用 表观基因组在发育、生长和衰老过程中存在一个动态变化过程,体细胞的表观基因组存在重新编程的可能性,这有助于将表观遗传分析应用于诊断、风险评估和治疗。 需要解决的问题: (1)确定表观遗传修饰与特定生理和病理指标的关系(抑癌基因启动子区甲基化); (2)技术分析的可行性; (3)确定表观遗传修饰——病理指标——个体临床表现之间的关系 亚硫酸氢盐选择性地使胞嘧啶脱氨,但不作用于5-mC
确定表观遗传修饰与特定生理和病理指标的关系(抑癌基因启动子区甲基化); (2)技术分析的可行性; (3)确定表观遗传修饰——病理指标——个体临床表现之间的关系. 亚硫酸氢盐选择性地使胞嘧啶脱氨,但不作用于5-mC.")
66
甲基化的胞嘧啶不变,未甲基化的胞嘧啶转变为尿嘧啶
DNA 经亚硫酸氢盐处理后, 甲基化的胞嘧啶不变,未甲基化的胞嘧啶转变为尿嘧啶

67
甲基特异性的PCR 扩增(MS-PCR):
亚硫酸氢盐处理后DNA作为模板,加入甲基化特异性的引物(primerⅠ)或非甲基化的引物(primerⅡ),进行特异性的扩增
或非甲基化的引物(primerⅡ),进行特异性的扩增.")
68
七、表观遗传的生物学意义 Jirtle和Waterland的实验(2003):Agouti小鼠实验
Agouti(A)基因编码一种信号分子,合成黄色素。在鼠发育过程中一般短暂表达,鼠毛一段呈现黄色,使野生型Agouti鼠呈棕褐色。 在A基因上游插入一个IAP(intracisternal A particle,源自逆转座子LTR序列),使A基因受IAP中的启动子调控而持续表达,毛色呈黄色。插入了IAP的“A”基因称“AVY”(Agouti viable yellow gene allele)
基因编码一种信号分子,合成黄色素。在鼠发育过程中一般短暂表达,鼠毛一段呈现黄色,使野生型Agouti鼠呈棕褐色。 在A基因上游插入一个IAP(intracisternal A particle,源自逆转座子LTR序列),使A基因受IAP中的启动子调控而持续表达,毛色呈黄色。插入了IAP的 A 基因称 AVY (Agouti viable yellow gene allele)")
69
IAP启动子CpG岛的甲基化又会使有些细胞中的AVY 表达受抑。

70
Agouti小鼠实验 隐性纯合型 a/a 母鼠孕 AVY/a 杂合型仔鼠。 孕鼠分为两组: (1)仅喂食标准饲料
(2)标准饲料 + 富含甲基的食物 结果: 第一组鼠孕产的仔鼠大多数为黄色,并对肥胖、糖尿病和肿瘤易感 第二组鼠孕产的仔鼠在身体的不同部位出现变化的棕色斑块,甚出现以棕褐色为主要毛色的仔鼠。
标准饲料 + 富含甲基的食物. 结果: 第一组鼠孕产的仔鼠大多数为黄色,并对肥胖、糖尿病和肿瘤易感. 第二组鼠孕产的仔鼠在身体的不同部位出现变化的棕色斑块,甚出现以棕褐色为主要毛色的仔鼠。")
71
With no more than a change in diet.
Laboratory agouti mice (left) were prompted to give birth to young ( right) that differed markedly in appearance and disease susceptibility.
72
喂食富含甲基的饲料对孕鼠产仔表型的影响

73
分析显示: 富含甲基饲料的孕鼠所产仔鼠IAP启动子区CpG岛的甲基化水平高于对照组。
转录调控区高甲基化使原呈异位表达的基因趋于沉默,毛色也趋于棕褐色。 结论: 营养等不致使DNA序列发生改变的环境因素变化,可以通过甲基化型而改变表遗传型,造成明显的表型效应。

74
理论意义 表观遗传修饰的环境因子敏感性可以解释基因结构一致的个体在不同环境中可以产生明显的表型差异。
细胞基因组可转座因子插入位点的异常甲基化会引起小鼠在细胞水平上的表观遗传镶嵌性,扩大了表型变化的范围。 每一个哺乳动物个体都能成为表观遗传镶嵌性,因此更容易在保持基因组稳定的前提下,提高机体对环境的适应能力。

75
表观遗传学发展而产生的一些概念 表观遗传:通过有丝分裂和减数分裂来传递非DNA序列信息的现象。
表观遗传突变:发育的重新编程过程中造成的特定基因表观遗传修饰的异常。 表观遗传病:由表观遗传修饰异常而导致的疾病。

76
表观等位基因:一个基因座(位点)上的特定的表遗传修饰。不同的表遗传修饰构成不同的表观等位基因。
表观基因组:基因组的全部表观遗传修饰标记。 表观基因组学:在基因组的背景下研究表遗传修饰与表型的关系。 表观基因治疗:以表观遗传修饰为靶点的基因治疗。

77
第十一章 掌握 熟悉 了解: 表观遗传概念,表观遗传修饰机制,人类疾病中的表观遗传现象,肿瘤发生中的表遗传修饰。
人类疾病中的基因印迹现象及其机制。 了解: 表观遗传的生物学意义。

79
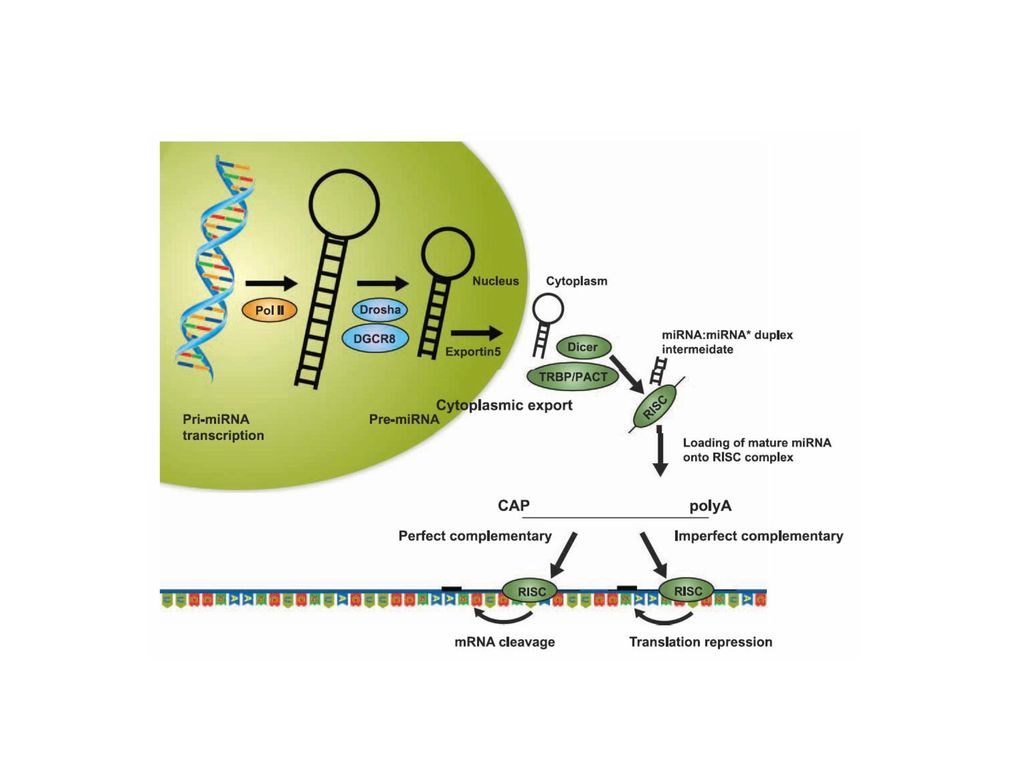
Small Interfering RNAs, siRNAs
几乎与第二种miRNA( let-7) 发现的同时, 通过不同的体系鉴定了small RNAs 和RNA干扰( RNA interference, RNAi)现象的存在。 RNAi 是一种进化上保守的序列特异性基因沉寂机制,通过导入双链RNA可以始动RNAi 的过程。 其细胞内的最后过程仍是将导入的dsRNA转化为21–25 bp大小的干涉RNA(small interfering RNAs, siRNAs) siRNA的一条链选择性的整合入发挥功能作用的复合体 (the RNA-induced silencing complex, RISC)
 发现的同时, 通过不同的体系鉴定了small RNAs 和RNA干扰( RNA interference, RNAi)现象的存在。 RNAi 是一种进化上保守的序列特异性基因沉寂机制,通过导入双链RNA可以始动RNAi 的过程。 其细胞内的最后过程仍是将导入的dsRNA转化为21–25 bp大小的干涉RNA(small interfering RNAs, siRNAs) siRNA的一条链选择性的整合入发挥功能作用的复合体 (the RNA-induced silencing complex, RISC)")


染色体结构不同; ( 2 )原核生物具有正调控和负调控并重的特点,真核 生物目前已知的主要是正调控; ( 3 )原核生物的转录和翻译是相偶联的,真核生物的.>")
:核蛋白体组成成分 转移 RNA ( tRNA ):转运氨基酸 信使 RNA ( mRNA ):蛋白质合成模板 不均一核 RNA ( hnRNA ):成熟 mRNA 的前体 小核 RNA ( snRNA ):>")

>")












 A.染色体 B.质体 C.线粒体 D.核糖体>")


