Download presentation
Presentation is loading. Please wait.

1
寒武纪物种大爆发 刘洋 张飞 冯俊 北京师范大学生命科学院2003级

2
摘要 寒武纪物种大爆发对达尔文提出的进化论的补充,以及地质环境对寒武纪物种大爆发形成的影响,及我国云南澄江动物群充当寒武纪物种大爆发的有利证据。

3
前言 寒武纪是地质年代古生代的第一个纪,开始于5.7亿年左右,结束于5.1亿年左右。寒武纪之前的地层的动物化石较少,而在寒武纪的地层中,发现了种类繁多的动物化石,所以古生物学家认为动物各门的祖先在这个时期都已出现,称其为“寒武纪物种大爆发。

4
成因 寒武纪物种大爆发的可能解释有两种:一种观点认为,寒武爆发是一种假象,这是达尔文主义者所持的观点。由于进化是渐进的,所谓的“爆发”只是表明首次在生物化石记录中发现了早在前寒武纪就已经广泛存在并发展的生物,其它的生物化石群则可能由于地质记录的不完全而“缺档”,造成这种“缺档”的原因是前寒武纪地层经历着热与压力,其中的化石被销毁了。由于发现前寒武纪化石沉积层中存在大量像细菌和蓝藻这样简单的原核生物,因而这一解释不再有说服力。另一种观点认为,寒武爆发代表了生物进化过程中的真实事件,科学家从物理环境和生态环境的变化两个方面来解释这一现象。

5
中国科学家构想寒武纪大致图

6
六十年代两位美国物理学家提出了寒武爆发是由于地球大气的氧水平这个物理因素造成的。他们认为,在早期地球的大气中含有很少或根本就没有自由氧,氧是前寒武纪藻类植物光合作用的产物并逐渐积累形成的。后生动物需要大量的氧,一方面用于呼吸作用,另一方面氧还以臭氧的形式在大气中吸收大量有害的紫外线,使后生动物免于有害辐射的损伤。 而生物学家则希望从生物本身的生态关系来探讨这一问题,因为地质学的证据否定了这种氧理论的观点。大约在距今10亿年至20亿年之间广泛沉积层中含有大量严重氧化的岩石,这说明在这一时期内已经存在足够生命爆发的氧条件。因而生物学家从两个重要事件的出现来探索造成寒武爆发的原因,即有性生殖的产生和生物收割者的出现。

7
从化石资料来看,真核藻类大约在9亿年前出现了有性生殖,实际上,有性生殖出现得更早。有性生殖的发生在整个生物界的进化过程中有着极其重大的作用,由于有性生殖提供了遗传变异性,从而有可能进一步增加了生物的多样性,这是造成寒武爆发的原因之一。

8
生物收割者假说是美国生态学家斯坦利提出的,是一种解释寒武爆发的生态学理论,即收割原则。斯坦利认为,在前寒武纪的25亿年的多数时间里,海洋是一个以原核蓝藻这样简单的初级生产者所组成的生态系统。这一系统内的群落在生态学上属于单一不变的群落,营养级也是简单唯一的。由于物理空间被这种种类少但数量大的生物群落顽强地占据着,所以这种群落的进化非常缓慢,从未有过丰富的多样性。寒武爆发的关键是草食收割者的出现和进化,即食用原核细胞(蓝藻)的原生动物的出现和进化。收割者为生产者有更大的多样性制造了空间,而这种生产者多样性的增加又导致了更特异的收割者的进化。营养级金字塔按两个方向迅速发展:较底层次的生产者增加了许多新物种,丰富了物种多样性,在顶端又增加了新的“收割者”,丰富了营养级的多样性。从而使得整个生态系统的生物多样性不断丰富,最终导致了寒武纪生命大爆发的产生。
的原生动物的出现和进化。收割者为生产者有更大的多样性制造了空间,而这种生产者多样性的增加又导致了更特异的收割者的进化。营养级金字塔按两个方向迅速发展:较底层次的生产者增加了许多新物种,丰富了物种多样性,在顶端又增加了新的 收割者 ,丰富了营养级的多样性。从而使得整个生态系统的生物多样性不断丰富,最终导致了寒武纪生命大爆发的产生。")
9
对于“收割理论”科学家们目前还没有找到直接的证据来证明其正确性,然而,一些间接的证据支持了这一理论。间接证据之一来自于前寒武纪叠层石,这些由藻类组成的叠层石中保存了前寒武纪最丰富的生产者群落。今天,叠层石仅盛产于缺少后生动物收割者的贫瘠环境中,如超盐量的咸水湖中。藻类在前寒武纪地层中的大量存在,大概反映了当时收割者的贫乏。另外,生态学野外研究也提供了一些间接的证据,研究表明,在一个人工池塘中,放进捕食性鱼,会增加浮游生物的多样性;从多样的藻类群落中去掉海胆,会使某一藻类在该群落中占统治地位而多样性下降。

10
对比 寒武纪物种大爆发已直接对达尔文的进化论产生了挑战,达尔文提出的进化论主要包括四方面:1一般进化论 2 共同祖先学说 3 自选选择学说 4渐变论 而“渐变论”——生物进化的步调是渐变式的,是一个在自然选择作用下累积微小的优势变异的逐渐改进的过程,而不是跃变式的。这是达尔文进化论中较有争议的部分。在达尔文在世时以及死后相当长一段时间,大部分生物学家,特别是古生物学家,都相信生物进化是能够出现跃变的,认为新的形态和器官是源自大的跃变,而不是微小的变异在自然选择的作用下缓慢而逐渐地累积下来的。包括赫胥黎在内的一些古生物学家由于强调生物化石的不连续性,而持这种观点。在遗传学诞生之后的一段时间内,早期遗传学家们由于强调遗传性状的不连续性,也普遍接受跃变论。在20世纪40年代,“现代综合”学说将遗传学和自然选择学说成功地结合起来,渐变论逐渐占了优势。

11
但是近二、三十年来,古生物学和进化发育生物学的研究表明,生物进化过程很可能是渐变和跃变两种模式都存在的,跃变论又有抬头的趋势。不过,进化论所说的跃变,除了某些非常特殊的情形(例如植物经杂交出现新种),并非是指在一代或数代之间发生的进化,而可能经历了数千年、数万年乃至数百万年,只不过以地质年代来衡量显得很短暂而已——又与寒武纪物种大爆发产生严重差异,但它即使成立,也不过表明进化有时候是能够以跃变的方式进行的,并不能否认进化在其他时候是以渐变的方式进行的。几乎所有动物的“门”都在寒武纪早期出现,绝不意味着这些动物祖先不是进化而来的,更不意味着它们之后没有发生进化。第一,在寒武纪之前,动物已经过了漫长的进化过程。自五十年代以来,古生物学家已在世界各地三十个地方发现了大量的寒武纪之前的多细胞生物乃至动物,数量最多、最为闻名的在四个地方:澳大利亚的埃迪亚加拉山(Ediacara Hill)、加拿大纽芬兰的错误点(Mistake Point)、俄罗斯的白海海岸和纳米比亚。此外还有中国瓮安动物群,据称是迄今发现的最古老的实体化石动物群。这些寒武纪之前的多细胞生物包括软珊瑚、海蜇、蠕虫和其他稀奇古怪的生物。对这些多细胞生物是否是寒武纪动物的直接祖先,以前有争议,因为在1995年之前从这些多细胞生物到寒武纪动物还存在一段地质空白,所以有专家主张这些早期多细胞生物全部灭绝,在寒武纪又再来一次从单细胞到多细胞的进化。
,并非是指在一代或数代之间发生的进化,而可能经历了数千年、数万年乃至数百万年,只不过以地质年代来衡量显得很短暂而已——又与寒武纪物种大爆发产生严重差异,但它即使成立,也不过表明进化有时候是能够以跃变的方式进行的,并不能否认进化在其他时候是以渐变的方式进行的。几乎所有动物的 门 都在寒武纪早期出现,绝不意味着这些动物祖先不是进化而来的,更不意味着它们之后没有发生进化。第一,在寒武纪之前,动物已经过了漫长的进化过程。自五十年代以来,古生物学家已在世界各地三十个地方发现了大量的寒武纪之前的多细胞生物乃至动物,数量最多、最为闻名的在四个地方:澳大利亚的埃迪亚加拉山(Ediacara Hill)、加拿大纽芬兰的错误点(Mistake Point)、俄罗斯的白海海岸和纳米比亚。此外还有中国瓮安动物群,据称是迄今发现的最古老的实体化石动物群。这些寒武纪之前的多细胞生物包括软珊瑚、海蜇、蠕虫和其他稀奇古怪的生物。对这些多细胞生物是否是寒武纪动物的直接祖先,以前有争议,因为在1995年之前从这些多细胞生物到寒武纪动物还存在一段地质空白,所以有专家主张这些早期多细胞生物全部灭绝,在寒武纪又再来一次从单细胞到多细胞的进化。")
12
在1995年,在纳米比亚火山灰层中出现了大量的寒武纪之前的多细胞生物,恰好补上了这段空白,所以,现在已很少有专家怀疑前寒武纪的多细胞生物和寒武纪的动物没有相承关系。第二,寒武纪的动物并不是“同时”出现的,而是持续了几百万年,这在进化史上当然是短时间,但对神创论来说,却是长得不可思议。第三,“几乎所有动物的门”在寒武纪地层出现并不等于“几乎所有动物的种”在那时候都已出现。事实上,寒武纪的动物一般地只是那个门的原始物种,以后几乎全都灭绝了,后来的物种是进化来的。比如,寒武纪只存在少数几种原始的脊索动物,而丰富多彩的脊椎动物各类群,包括鱼类、两栖类、爬行类、哺乳类和鸟类,都是在寒武纪之后从原始脊索动物逐渐进化来的。现代脊椎动物各物种更都有了几亿年的进化史。

13
几乎所有动物的门在这较短的时间内进化出来,生物学家提出了不少的假设,目前被较为合理的是Hox基因调控理论。Hox基因是一种“同源异形”基因,是动物形态蓝图的设计师,在发育过程中控制身体各部分形成的位置。如果同源异形基因发生突变,会使动物某一部位的器官变成其他部位的器官,叫做同源异形。比如,让某个同源异形基因发生突变,能使果蝇的身体到处长眼睛,在该长眼睛的地方长出翅膀,或者在该长触角的地方长出了脚。Hox基因在所有的脊椎动物和绝大部分无脊椎动物中都存在,调控的机理也相似,这表明它可能是最古老的基因之一,在最早的动物祖先中就已存在。Hox的突变一开始时在胚胎早期引起的变化不大,但随着组织、器官的分化定型,突变的影响逐步被放大,导致身体结构发生重大的改变。这可以解释寒武纪物种大爆发。那时候基因结构、发育过程都较简单,Hox的基因突变容易被保留,结果导致了身体结构的多姿多彩。

14
证据 寒武爆发的典型代表是被称为20世纪最惊人的科学发现之一的云南澄江动物群,它是世界上目前所发现的最古老、保存最为完整的带壳后生生物群。 该动物群是青年古生物学家侯先光教授于1984年在云南澄江县帽天山首先发现的。这是一个内容十分丰富、保存非常完美,距今约5.7亿年的化石群,其成员包括水母状生物、三叶虫、具附肢的非三叶的节肢动物、金臂虫、蠕形动物、海绵动物、内肛动物、环节动物、无绞纲腕足动物、软舌螺类、开腔骨类,以及藻类等,甚至还有属于低等脊索动物或半索动物等。由于许多动物的软组织保存完好,为研究早期无脊椎动物的形态结构、生活方式、生态环境等提供了极好的材料。

16
大量新的化石类型的发现,特别是箭虫、尾索动物、甲壳动物和螯肢类等现生动物高级分类单元化石代表的发现,为寒武纪大爆发是现生高级分类系统主要历史起点提供了新证据,也对寒武纪大爆发有了更趋完整的认识, 在脊椎动物起源的研究方面取得重要进展。大量海口鱼化石的新发现和详细研究证明海口鱼已进入有头类的进化轨道;对海口虫解剖学和分支生物学的深入研究,证实海口虫处在在无脊椎动物向脊椎动物演化的中间阶段,提出了神经脊动物的概念,并对脊椎动物脑、脊椎骨、头部感觉器等的起源进行了探讨;将古虫重新 解释为后口类,并据此提出无脊椎动物向脊椎动物演化划分为五个阶段的新假说,通过生物食性和粪化石的研究,对寒武纪早期生态链的特征增进了解,为寒武纪大爆发是生命复杂生态体系的历史起点提供了新的依据。

17
环境 寒武纪以前,由于地质环境尚未演化到适合的阶段,生物的出现也仅仅是低级的原始的,个体数量少,分布面积不广,密度也不大,而且更重要的是还没有条件形成硬壳保护其遗体。所以,保存化石的机率就很少。但是,寒武纪以前长达26亿年(自32亿年前最古老的微化石到6 亿年前寒武纪开始)的漫长时代,为生物大爆发准备了质变的飞跃和大量繁殖出现的条件。寒武纪早期,地球三圈—岩石圈、水圈、气圈发生剧烈变化,游离氧增至现代氧压的10% ,足以支持生物在液体介质中借助本身器官进行气体循环。此时的海水中富磷、镁、硅并含钙,为生物造壳提供了物质基础。生物一旦有壳,可保护半定型的软体,适应多变环境和防御外敌能力大大加强。于是,发生了生物界具有质变飞跃意义的带壳演化,促使了梅树村动物群和澄江动物群两次生命大爆发。
的漫长时代,为生物大爆发准备了质变的飞跃和大量繁殖出现的条件。寒武纪早期,地球三圈—岩石圈、水圈、气圈发生剧烈变化,游离氧增至现代氧压的10% ,足以支持生物在液体介质中借助本身器官进行气体循环。此时的海水中富磷、镁、硅并含钙,为生物造壳提供了物质基础。生物一旦有壳,可保护半定型的软体,适应多变环境和防御外敌能力大大加强。于是,发生了生物界具有质变飞跃意义的带壳演化,促使了梅树村动物群和澄江动物群两次生命大爆发。")
18
思考 寒武纪,除个别门类外,现在生活在地球上的生物几乎全部出现了,揭开了生物演化史上的宏伟帷幕。当时的大陆可能是荒凉的,还没有找到任何真实的陆生生物遗迹。低级的苔藓和地衣类的植物,可能生活在潮湿的低地,还缺少根与纤维组织,难以蔓生到干燥地区;当时的无脊椎动物也还没有在空气中生活的机能。

20
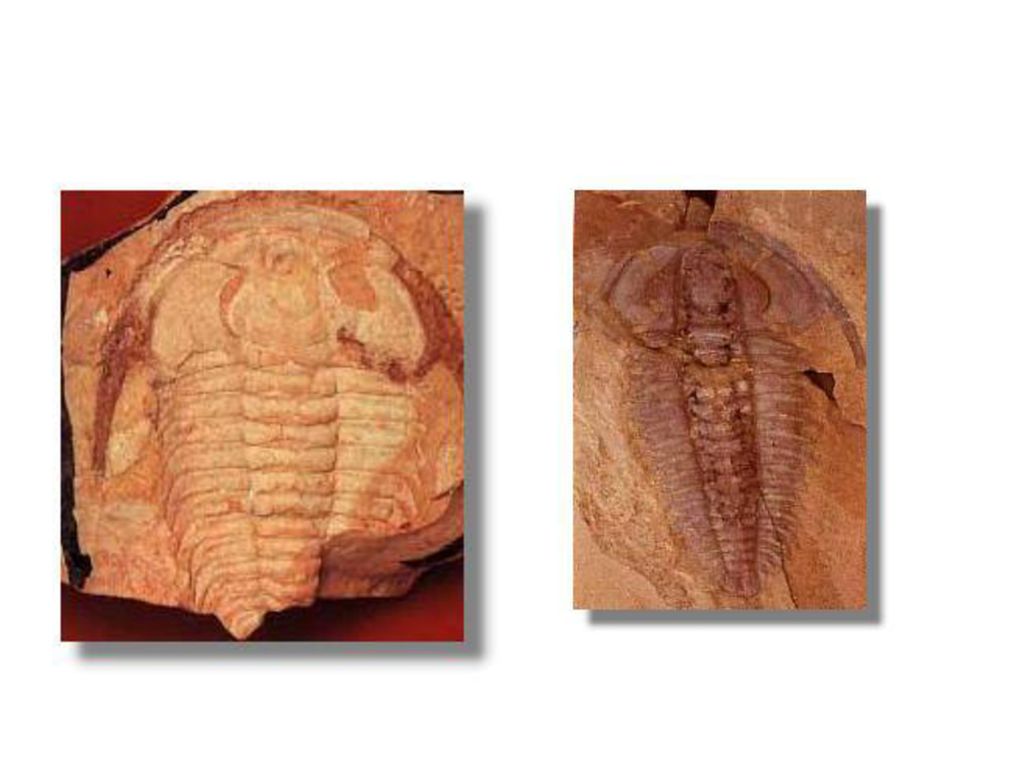
(1)无脊椎动物 节肢动物的三叶虫是寒武纪最繁盛的生物,约占当时全部生物的60%,是古生代早期一类比较高级的无脊椎动物。根据形态概略的讲,早寒武纪的三叶虫,一般是头部巨大,尾部短小,如小遇仙寺虫);中寒武纪的,头尾大小近相等,尾部经常生长着不同型式的棘刺,如德氏虫、叉尾虫;晚寒武世的头尾多半是光滑的圆浑的。
;中寒武纪的,头尾大小近相等,尾部经常生长着不同型式的棘刺,如德氏虫、叉尾虫;晚寒武世的头尾多半是光滑的圆浑的。")
21
(2)腕足动物 约占寒武纪全部生物的30%。早、中寒武纪的腕足动物,以贝体小、几丁质壳的无铰类为多,如小舌形贝、滇东贝等。原始的、具钙质壳的有铰类已出现在早寒武纪。
腕足动物 约占寒武纪全部生物的30%。早、中寒武纪的腕足动物,以贝体小、几丁质壳的无铰类为多,如小舌形贝、滇东贝等。原始的、具钙质壳的有铰类已出现在早寒武纪。")
22
寒武纪物种大爆发为物种开启了另一扇进化之门,然而单凭现在的证据以及在此之上建立起的假设还不足以说明物种的进化,并且寒武纪是否真正存在大爆发还不能确定。

Similar presentations


 、 ( 資料蒐集 ) 組員: 9466402 林富娟 ( 書面整理 ) 、 ( 資料蒐集 ) 9466415 陳佳琪 (Power Point) 、 ( 資料蒐集 ) 9466419.>")



 優質通識學生學習檔案 授課教授:○○○教授 學 生:○○○ 學 號:○○○○ 範本 個人照片一張 中國科技大學.>")
 教師手冊. 影片欣賞 -- 愛的晚霞 單純的阿霞人生第一次的愛情,卻是帶來身心嚴重 的傷害,阿霞要如何面對感染愛滋後的生活 …>")
 貳、教學檔案平台介紹(主菜) 參、檔案目錄範例及小叮嚀(甜點)>")


 地址:閃電鎮同人區10.8號 電話:(08)>")



>")

: 掌握谋职类、条据类文书的作用、写作要求。>")


