Download presentation
Presentation is loading. Please wait.

1
重组人白介素-18诱导表达,纯化与免疫印迹鉴定
2012级生物科学专业课实验 重组人白介素-18诱导表达,纯化与免疫印迹鉴定 高朝辉 2015年3月13日

2
目 录 一、实验目的 二、实验原理 三、试剂配制及关键仪器使用 四、实验操作步骤及注意事项 五、相关理论知识(自学)
")
3
一、实验目的 熟悉工程菌发酵及源蛋白表达的特点和方法; 掌握镍柱亲和层析的基本原理和操作方法;
掌握蛋白质印迹技术(Western Blot)的基本原理和操作方法。
的基本原理和操作方法。")
4
熟悉工程菌发酵及源蛋白表达的特点和方法;
掌握镍柱亲和层析的基本原理和操作方法; 掌握蛋白质印迹技术(Western Blot)的基本原理和操作方法。
的基本原理和操作方法。")
5
熟悉工程菌发酵及源蛋白表达的特点和方法;
掌握镍柱亲和层析的基本原理和操作方法; 掌握蛋白质印迹技术(Western Blot)的基本原理和操作方法。
的基本原理和操作方法。")
6
了解工程菌发酵及外源蛋白表达的特点和方法;
掌握镍柱亲和层析的基本原理和操作方法; 掌握蛋白质印迹技术(Western Blot)的基本原理和操作方法。
的基本原理和操作方法。")
7
二、实验原理 1、菌株构建与目的蛋白 目的蛋白:IL-18含有157 个氨基酸残基,分子量为18.3kDa,pI=4.9。
菌株构建:分子生物学实验《重组人白介素-18(rhIL-18)基因工程菌的构建》
基因工程菌的构建》")
8
2、技术路线 pET32a hIL18 pET-hIL18 BL21中表达 Tax-hIL18 生物活性检测 双酶切 基因合成
纯化 hIL-18基因片段 线性pET32a Tax-hIL18 连接 酶切、PCR及测序鉴定 生物活性检测 SDS-PAGE检测分子量 Western Blot定性鉴别

9
pET-hIL18

10
3、pET表达系统的原理

11
内容介绍 1 乳糖操纵子的正负调控 2 T7 RNA 聚合酶 3 pET表达系统

12
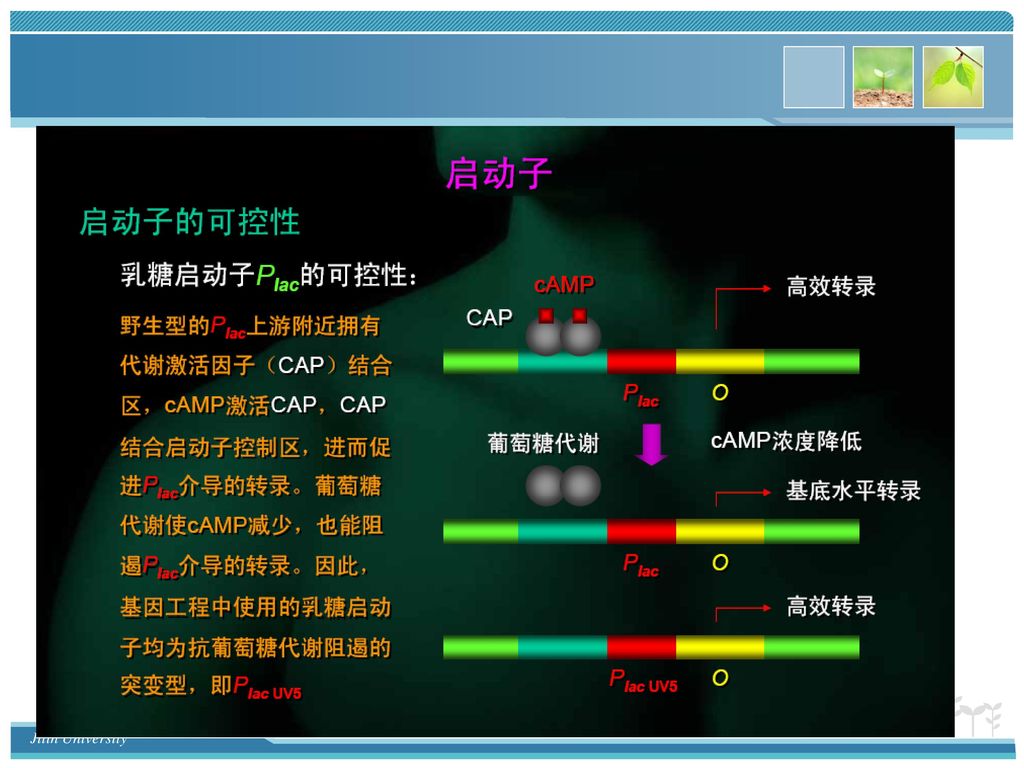
乳糖操纵子的结构 operator lac operon I P O Z Y A Lac I 阻遏蛋白 β-半乳糖苷酶 Lac 通透酶

13
乳糖操纵子的负调控 诱导物不存在时 I P O Z Y A 诱导物存在时 I P O Z Y A I mRNA
I mRNA翻译出的阻遏蛋白结合到启动子部位,阻止lac 操纵子基因的转录 诱导物存在时 I P O Z Y A lac mRNA I mRNA 诱导物 诱导物与阻遏蛋白结合使阻遏蛋白无法与启动子结合,转录顺利进行,表达相应的酶 诱导物

15
T7 RNA聚合酶 来源 专一性 转录效率 λ(DE3)噬菌体,单链蛋白。 只识别λ噬菌体启动子,不识别大肠杆菌启动子。
RNA的合成速度是大肠杆菌RNA聚合酶的5倍

16
pET系统表达原理 未诱导的情况下 E.Coli 基因组 目标基因 pET lacI 阻遏蛋白 T7 启动子 lacUV5
T7 RNA Polymerase gene 目标基因 Content Layouts pET lacI 阻遏蛋白 T7 启动子

17
pET系统表达原理 诱导的情况下 E.Coli 基因组 目标基因 pET T7 启动子 lacUV5
T7 RNA Polymerase gene 目标基因 lacI 阻遏蛋白 pET 乳糖或IPTG T7 启动子 IPTG:异丙基-β-D-硫代吡喃半乳糖苷,乳糖结构类似物,是一 种作用极强的诱导剂,不被细菌代谢而十分稳定,因此被实验室广泛应用。价格昂贵,工业生产不用。

18
4、原核表达的包涵体问题 包涵体是指细菌表达的蛋白在细胞内凝集,形成无活性的固体颗粒。
包涵体的成分由50%以上的重组蛋白,糖体元件、RNA聚合酶、内毒素、外膜蛋白ompC、ompF和ompA等,环状或缺口的质粒DNA,以及脂体、脂多糖等组成,大小为0.5-1um,密度高(约1.3mg/ml),无定形,呈非水溶性,只溶于变性剂如尿素、盐酸胍等。
,无定形,呈非水溶性,只溶于变性剂如尿素、盐酸胍等。")
19
包涵体形成主要原因 1、基因工程菌的表达产率过高,超过了细菌正常的代谢水平,由于细菌的δ因子的蛋白水解能力达到饱和,使之表达产物积累起来。
2、重组蛋白的氨基酸组成:一般说含硫氨基酸越多越易形成包涵体,而脯氨酸的含量明显与包涵体的形成呈正相关。 3、重组蛋白所处的环境:发酵温度高或胞内pH接近蛋白的等电点时容易形成包涵体。 4、重组蛋白是大肠杆菌的异源蛋白,由于缺乏真核生物中翻译后修饰所需酶类和辅助因子,如折叠酶和分子伴侣等,致使中间体大量积累,容易形成包涵体沉淀。 5、蛋白质在合成之后,于中性pH或接近中性pH的环境下,其本身固有的溶解度对于包涵体的形成比较关键,即是说,有的表达产率很高,如Aspartase和Cyanase,表达产率达菌体蛋白的30%,也不形成包涵体,而以可溶形式出现。 6、在细菌分泌的某个阶段,蛋白质分子间的离子键、疏水键或共价键等化学作用导致了包涵体的形成。

20
5、亲和层析原理

21
理想的亲和层析检测结果 M:protein marker 1:未诱导 2:诱导表达 3:挂柱穿透组分 4:洗涤组分 5:洗脱组分

22
金属螯合层析法 金属螯合亲合层析,又称固定化金属离子亲合层析(Immobilized metal ion affinity chromatography, IMAC) 是近30年发展起来的一种新型分离技术。最早由Paroth等人提出 该方法利用蛋白质表面的一些氨基酸,如组氨酸、色氨酸、半胱氨酸等能和金属离子发生特殊的相互作用的原理,从而对蛋白质加以分离 这些作用包括配价键结合、静电吸附、共价键结合,其中以配价键结合为主,而且这其中又以6组氨酸标签(His-Tag)应用最为广泛
应用最为广泛.")
23
Ni Protein His imidazole 固相载体 C-COOH -O-C-C-C-O-C-C-C-N elution OH
Juang RH (2004) ECX 圖 3.10 一種金屬螯合層析法:
 ECX. 圖 3.10 一種金屬螯合層析法:")
24
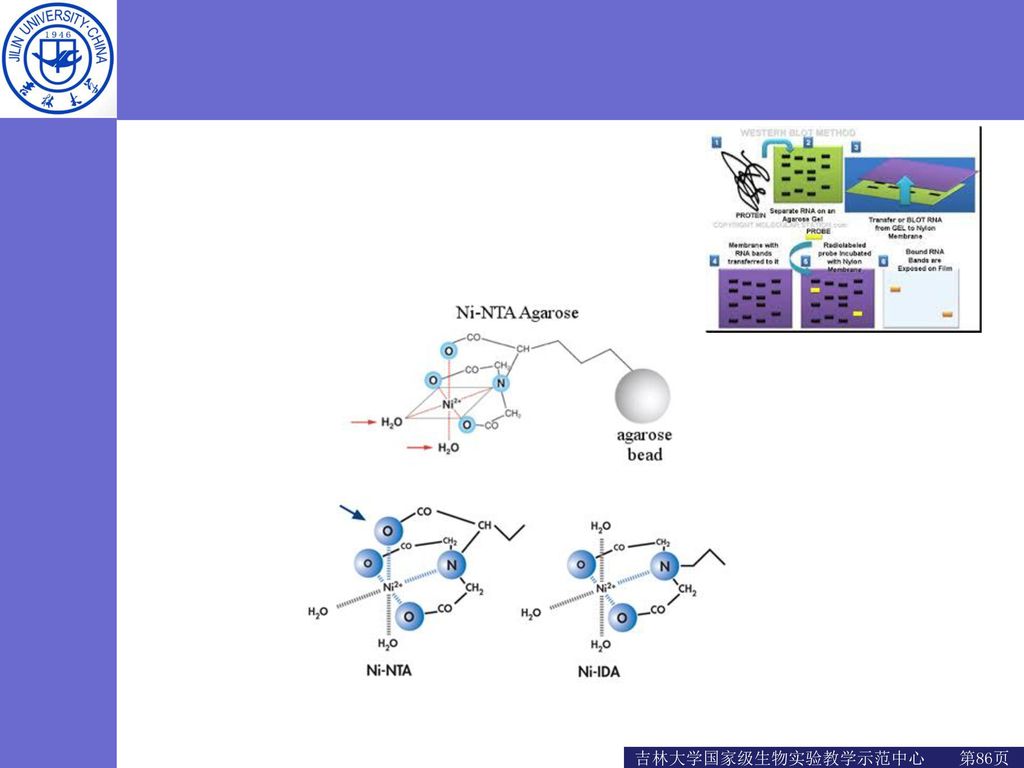
His-tag——历久弥新的6组氨酸标签 His-tag可与多种金属离子发生特殊的相互作用,包括Ca2+、Mg2+、Ni2+、Co2+等,其中以镍离子使用最为广泛。

25
常用镍柱树脂 Ni-IDA层析纯化介质是把琼脂糖介质(4%交联)通过3个结合位点共价耦联到可以结合Ni2+的三齿螯合剂(亚氨基二乙酸,IDA)制备而成。 Ni-NTA层析纯化介质是把琼脂糖介质(4%交联)通过4个结合位点共价耦联到可以结合Ni2+的四齿螯合剂(次氮基三乙酸,IDA)制备而成。
通过3个结合位点共价耦联到可以结合Ni2+的三齿螯合剂(亚氨基二乙酸,IDA)制备而成。 Ni-NTA层析纯化介质是把琼脂糖介质(4%交联)通过4个结合位点共价耦联到可以结合Ni2+的四齿螯合剂(次氮基三乙酸,IDA)制备而成。")
26
Ni-NTA Ni-IDA

27
镍琼脂糖凝胶FF的配基是最经典的IDA,镍离子有六个螯合价位,Ni-IDA螯合了三价,剩余三价
而Ni-NTA螯合了四价的,剩余是两价 因此,Ni-IDA琼脂糖凝胶作用力要比Ni-NTA琼脂糖凝胶的强 IDA的载量要比NTA高,而在同样条件下Ni-IDA洗杂质和目标蛋白的要比Ni-NTA的咪唑浓度高 NTA的填料更稳定,能耐受更强的还原剂,更不容易脱落。

28
6、SDS-PAGE原理 PAGE具有电泳和分子筛的双重作用。
蛋白质在凝胶中受电场的作用而发生迁移,迁移的速率取决于蛋白质所带电荷的多少和蛋白质的大小和形状。 使各种蛋白质—SDS复合物都带上相同密度的负电荷,其数量远远超过了蛋白质分子原有的电荷量,从而掩盖了不同种类蛋白质间原有的电荷差别。

29
7、Western Blot原理 Western免疫印迹(Western Blot)是将蛋白质转移到膜上后利用抗体进行检测的技术。
是将蛋白质转移到膜上后利用抗体进行检测的技术。")
30
Western Blot流程

31
三、试剂配制及关键仪器使用 1、诱导表达所需试剂 2、SDS-PAGE所需试剂 3、Western Blot所需试剂

32
1、诱导表达所需试剂 1. LB培养基 2. 氨苄青霉素 3. IPTG 4. 悬浮缓冲液 100mM Tris-HCl, pH8.0
2. 氨苄青霉素 3. IPTG 4. 悬浮缓冲液 100mM Tris-HCl, pH8.0 5. 2×SDS上样缓冲液 50mmol/LTris-Cl pH6.8 100mmol/L DTT(二硫苏糖醇) 4%SDS 0.2%溴酚蓝 20%甘油.
 4%SDS. 0.2%溴酚蓝. 20%甘油.")
33
2、SDS-PAGE所需试剂 10%SDS (w/v) (100ml) 凝胶储备液 (30% Acr-0.8% Bis)
电极缓冲液 (5×) 染色液 脱色液
 染色液. 脱色液.")
34
3、Western Blot所需试剂 TBS 缓冲液:20mml/L Tris-HCl,500mmol/LNaCl,PH:7.5。
TBST缓冲液:20mmol/L Tris-HCl,500mmol/L NaCl,0.05% Tween-20,pH:7.5。 抗体溶液:Blocking溶液加相应量抗体。 Blocking 溶液(封闭液):5%脱脂牛奶的TBS溶液 转移缓冲液:25mmol/L Tris,192 mmol/L 20%MethnolL,pH8.3。 一抗:His-tag小鼠单克隆抗体 二抗:辣根过氧化物酶标记山羊抗小鼠IgG抗体 DAB显色试剂盒
:5%脱脂牛奶的TBS溶液. 转移缓冲液:25mmol/L Tris,192 mmol/L 20%MethnolL,pH8.3。 一抗:His-tag小鼠单克隆抗体. 二抗:辣根过氧化物酶标记山羊抗小鼠IgG抗体. DAB显色试剂盒.")
35
四、实验操作步骤及注意事项 培养基配制与种子培养 诱导表达 细胞破碎 Western Blot 结果分析 配置培养基 种子液培养 表达量检测
收集菌体 超声破碎 离心分离 Western Blot SDS-PAGE 电转膜 酶免疫定位 结果分析 不同时间表达量 亲和层析 免疫印迹结果分析

36
1、培养基配制与种子培养 配置LB液体培养基100mL 种子液培养
LB 培养基:胰蛋白胨1%、酵母提取物0.5%、NaCl 1%、调pH值至7.0,121℃灭菌20min。 种子液培养 取一个LB液体培养基,加入Amp (100mg/mL)至终浓度50ug/mL。 接种保存的、含有目的基因的工程菌。 37 ℃培养过夜。
至终浓度50ug/mL。 接种保存的、含有目的基因的工程菌。 37 ℃培养过夜。")
37
2、诱导表达 取1mL种子液接入100mL培养液,加入Amp (100mg/mL)至终浓度50ug/mL,在250ml摇瓶中于30℃剧烈震荡培养2h,OD600为0.6。 吸出1mL未经诱导的培养物(0h)放在一个Ep管中,按下面步骤5所述进行处理。 在剩余培养物中加入IPTG(100mmol/L)至终浓度1mmol/L,30℃继续通气培养。 在诱导的不同时间(0.5h、1h、1.5h、2h、3h)取1mL培养液置于Ep管中,室温高速(12000rpm)离心2min。 沉淀悬于100uL的1×SDS凝胶加样缓冲液中,沸水浴加热5min,点动离心,冰上放置保存。 待全部样品处理完后进行菌体总蛋白SDS-PAGE,检测目的蛋白表达量。
至终浓度1mmol/L,30℃继续通气培养。 在诱导的不同时间(0.5h、1h、1.5h、2h、3h)取1mL培养液置于Ep管中,室温高速(12000rpm)离心2min。 沉淀悬于100uL的1×SDS凝胶加样缓冲液中,沸水浴加热5min,点动离心,冰上放置保存。 待全部样品处理完后进行菌体总蛋白SDS-PAGE,检测目的蛋白表达量。")
38
3、不同表达时间表达量检测 SDS-PAGE凝胶制备 先制分离胶,将分离胶注入玻板后,用去离子水封口,约30-40min后凝聚。
将胶面的水吸干后灌注浓缩胶,并插入梳子,胶凝固后取下备用。 单位/ml 去离子水 丙烯 酰胺 Tris 溶液 10% SDS AP TEMED 分离胶 2.6 1.7 pH8.8 0.6 0.05 0.003 5% 浓缩胶 0.5 pH6.8 0.76 0.02

39
电泳结束后染色1h,脱色至脱色液基本无色,背景透明无色,样品条带清晰,照相。
凝胶电泳 取30 ml 电极缓冲液分别注入阴极电泳槽和阳极电泳槽中。 在加样孔中加入15ul已处理样品,久置样品需煮沸 10 mA电泳至溴酚蓝前沿进入分离胶,调整电泳电流为20mA,电泳50min。 点样顺序 电泳结束后染色1h,脱色至脱色液基本无色,背景透明无色,样品条带清晰,照相。 第一道 第二道 第三道 第四道 第五道 第六道 第七道 第八道 第九道 第十道 样品6 样品1 Marker 0h 0.5h 1h 2h 3h

40
预期实验结果 诱导不同时间目的蛋白表达量检测结果

41
4、细胞破碎 诱导表达3h取样后,剩余培养物4℃、4000rpm离心10min,收集菌体,倒置离心管尽量去掉上清培养液。
用10ml去离子水重悬菌体。 超声破碎细胞,仪器设置:输出功率50%、超声时间5s,间隔时间5s,工作时间30min,将变幅杆深入液面下1-1.5cm,开始超声。超声全程冰浴试管,以保证在超声破碎的过程中菌体始终保持低温。 4℃、4000rpm离心10min,上清转移到新的离心管,10mL去离子水重悬沉淀,取50ul留样1(菌体)。 4℃、10000rpm离心20min,上清用0.45um的滤膜过滤以去除细胞碎片,过滤液转移到新的离心管中,取50uL留样2(可溶蛋白),储存于4℃冰箱中备用。用1ml去离子水重悬沉淀,取50uL留样3(包涵体)。
。 4℃、10000rpm离心20min,上清用0.45um的滤膜过滤以去除细胞碎片,过滤液转移到新的离心管中,取50uL留样2(可溶蛋白),储存于4℃冰箱中备用。用1ml去离子水重悬沉淀,取50uL留样3(包涵体)。")
42
5、Ni柱亲和层析 取一支10ml注射器,加去离子水检测底部筛板是否通透、橡皮管有无滴漏、紧固夹是否好使。
吸取1-2ml Ni-IDA柱料,加入注射器中。 用10倍柱床体积结合buffer平衡柱料,流速30ml/h。 连接紫外检测器,无气泡后,T档调100%旋钮至透光度95,然后调至0.1灵敏度,调0%旋钮至吸光度5。 将保存10mL上清液上样,流速15ml/h,另取容器承接穿透液,可反复上样以增加挂住蛋白量,取50ul最终穿透液留样4(挂柱效果)。
。")
43
结合buffer或50mM咪唑洗脱buffer洗涤5个柱床体积,流速30ml/h。取50ul洗涤液留样5。
500mM/L的咪唑洗脱buffer洗脱5-10个柱床体积,流速15mL/min,用紫外检测器同步检测,收集洗脱液,取洗脱峰50ul留样6。 用去离子水洗10个柱床体积,流速30ml/h。 再用20%乙醇洗5个柱床体积,流速30ml/h ,将柱子封存于4℃冰箱,留待下组实验使用。

44
6、破碎及层析效果检测 SDS-PAGE凝胶制备 先制分离胶,将分离胶注入玻板后,用去离子水封口,约30-40min后凝聚。
将胶面的水吸干后灌注浓缩胶,并插入梳子,胶凝固后取下备用。 单位/ml 去离子水 丙烯 酰胺 Tris 溶液 10% SDS AP TEMED 分离胶 2.6 1.7 pH8.8 0.6 0.05 0.003 5% 浓缩胶 0.5 pH6.8 0.76 0.02

45
电泳结束后染色1h,脱色至脱色液基本无色,背景透明无色,样品条带清晰,照相。
凝胶电泳 取30 ml 电极缓冲液注入中间电泳槽,外部电泳槽使用回收电极缓冲液。 在加样孔中加入15ul已处理样品,久置样品需煮沸 10 mA电泳至溴酚蓝前沿进入分离胶,调整电泳电流为20mA,电泳50min。 点样顺序 电泳结束后染色1h,脱色至脱色液基本无色,背景透明无色,样品条带清晰,照相。 第一道 第二道 第三道 第四道 第五道 第六道 第七道 第八道 第九道 第十道 Marker 样品1 样品2 样品3 样品4 样品5 样品6

46
理想的亲和层析检测结果 M:protein marker 1:未诱导 2:诱导表达 3:挂柱穿透组分 4:洗涤组分 5:洗脱组分

47
7、Western Blot SDS-PAGE 电转膜 酶免疫定位

48
7.1 SDS-PAGE 凝胶电泳 停止电泳 取30 ml 电极缓冲液注入中间电泳槽,外部电泳槽使用回收电极缓冲液。
在加样孔中加入15ul已处理样品,久置样品需煮沸 10 mA电泳至溴酚蓝前沿进入分离胶,调整电泳电流为20mA,电泳50min。 停止电泳 先将玻璃板撬掉才可以剥胶,将浓缩胶轻轻刮去,最后将溴酚蓝的前沿切下。 再将两侧的空白泳道切下不要,沿中间空白用刀将样品和Marker分开。 Marker泳道正常染色脱色,样品泳道进行转膜。

49
Marker 样品一 样品二 浓缩胶 分离胶 溴酚兰前沿

50
Marker 样品一 样品二 浓缩胶 分离胶 溴酚兰前沿

51
Marker 样品一 样品二 浓缩胶 分离胶 溴酚兰前沿

52
Marker 样品一 样品二 浓缩胶 分离胶 溴酚兰前沿

53
Marker 样品一 样品二 浓缩胶 分离胶 溴酚兰前沿

54
Marker 样品一 样品二 浓缩胶 分离胶 溴酚兰前沿

55
Marker 样品一 样品二 浓缩胶 进入转膜、免疫、显色操作 正常染色、脱色 分离胶 溴酚兰前沿

56
7.2 电 转 膜 切下的凝胶放入转移buffer浸泡15min。
7.2 电 转 膜 切下的凝胶放入转移buffer浸泡15min。 照凝胶大小剪取等大的硝酸纤维素膜,也浸泡在转移缓冲液中15min。将凝胶与硝酸纤维素膜重合,在左上角剪个小口做标记。 再剪取同样大小的两张厚滤纸,使用之前要在转移缓冲液中浸泡一下,滤纸不要比凝胶和膜大,以免短路。 将转移电泳槽中的夹子打开,浸泡在转移缓冲液中。白色一面在底下,在上面垫一张海绵垫,用玻璃棒来回擀几遍以擀走里面的气泡,在垫子上垫滤纸,一手固定滤纸,一手用玻璃棒擀去其中的气泡。

57
接着放上硝酸纤维素膜,再放上凝胶,凝胶之上再放滤纸。滤纸,膜,凝胶和滤纸之间要对整齐。最后盖上另一个海绵垫,擀几下后合起夹子。整个操作在转移缓冲液中进行。
将夹子放入转移槽中,要使夹子的黑面对应阴极,夹子的白面对应阳极。电转移时会产热,将转移电泳槽下端接上自来水,冷凝循环。打开电源,200mA,1.5h。转膜结束后,切断电源,取出硝酸纤维素膜。 为检测转膜是否成功,可用丽春红染色。 将硝酸纤维素膜取出放在平皿中,加入适量丽春红染色液,在摇床上染色5分钟,大量的水洗膜直至水变清,无色蛋白条带清晰,观察现象。

58
7.3 酶免疫定位 将膜用TBST浸湿后,移至含有封闭液的表面皿中,室温下,用封闭液在摇床上摇动封闭1.5h。封闭液现用现配,为1%
从封闭液中取出膜,在含有TBST的表面皿中洗涤5min,反复3次。 将膜(正面朝上)放入含有一抗的自封袋中,放入37℃摇床温育1h。 从封闭袋中取出膜,在含有TBST的表面皿中洗涤5min,反复3次。 将膜(正面朝上)放入含有二抗的自封袋中,放入37℃摇床温育1h
放入含有一抗的自封袋中,放入37℃摇床温育1h。 从封闭袋中取出膜,在含有TBST的表面皿中洗涤5min,反复3次。 将膜(正面朝上)放入含有二抗的自封袋中,放入37℃摇床温育1h.")
59
从DAB试剂盒中三种溶液分别吸取300uL稀释于6mL去离子水中,做显色液。
将膜放入显色液中,于暗处反应15min。观察照相。

60
Western Blot预期结果 1:正常sf9的表达产物 2:Bacmid转染sf9的表达产物 3:重组Bacmid转染sf9的表达产物
4:蛋白质marker.

61
实验时间安排 周三 周六 上午11:00,每组1人,接种 下午13:00,诱导表达,取样 看SDS-PAGE视频,练习配胶
每组配2板胶,取1块胶进行表达量检测,跑完电泳后切胶,一半胶用于转膜、封闭、一抗孵育过夜,另一半胶染色过夜 同时进行超声破碎、亲和层析、破碎层析效果检测,跑完电泳后染色过夜

62
周日 两块胶脱色,照相 TBST洗涤、二抗孵育、显色观察

63
The end!

64
五、相关理论知识(自学) 影响目的基因表达的因素 Ni柱纯化His标签融合蛋白注意事项 SDS-PAGE电泳常见问题的分析
Western Blot常见问题分析

65
影响目的基因表达的因素 外源基因表达产量与细胞浓度和细胞表达产量呈正相关。单个细胞产量取决于: 外源基因的拷贝数 外源基因的表达效率
启动子的强弱 核糖体接合位点的有效性 SD序列和起始密码ATG的间距 密码子组成 表达产物的稳定性 细胞代谢负荷 工程菌的培养条件

66
Ni柱纯化His标签融合蛋白注意事项 避免缓冲液中有高浓度的电子供体基团如NH4、甘氨酸、精氨酸、Tris等等;
各种缓冲液中不能有强螯合剂EDTA、EGTA等; 各种缓冲液里不能有高浓度的强还原剂如DTT,防止二价Ni被还原; 不能含离子型的去垢剂如SDS,防止Ni流失; 在破碎细胞的时候建议加入蛋白酶抑制剂如0.1-1mM的PMSF,防止目的蛋白被降解;

67
缓冲液里可以加入甘油,防止蛋白之间由于疏水相互作用而发生聚集沉淀,甘油浓度最高可达50%(v/v);
应避免含碳酸氢钠、柠檬酸等物质; 缓冲液里NaCl的浓度应在300mM到2M之间; 可加入变性剂促溶,盐酸胍(最高可6M),尿素(最高可8M); 可加入非离子型去垢剂,如Triton,Tween,NP40等,最高2%,可以减少背景蛋白污染和去除核酸污染;
,尿素(最高可8M); 可加入非离子型去垢剂,如Triton,Tween,NP40等,最高2%,可以减少背景蛋白污染和去除核酸污染;")
68
SDS-PAGE电泳常见问题的分析 配胶缓冲液系统对电泳的影响?
在SDS-PAGE不连续电泳中,制胶缓冲液使用的是Tris-HCL缓冲系统,浓缩胶是pH6.7,分离胶pH8.9;而电泳缓冲液使用的Tris-甘氨酸缓冲系统。 在浓缩胶中,其pH环境呈弱酸性,因此甘氨酸解离很少,其在电场的作用下,泳动效率低;而CL离子却很高,两者之间形成导电性较低的区带,蛋白分子就介于二者之间泳动。由于导电性与电场强度成反比,这一区带便形成了较高的电压剃度,压着蛋白质分子聚集到一起,浓缩为一狭窄的区带。 当样品进入分离胶后,由于胶中pH的增加,呈碱性,甘氨酸大量解离,泳动速率增加,直接紧随氯离子之后,同时由于分离胶孔径的缩小,在电场的作用下,蛋白分子根据其固有的带电性和分子大小进行分离。 所以,pH对整个反应体系的影响是至关重要的,实验中在排除其他因素之后仍不能很好解决问题的情况,应首要考虑该因素。

69
样品如何处理? 根据样品分离目的不同,主要有三种处理方法:还原SDS处理、非还原SDS处理、带有烷基化作用的还原SDS处理。
1)还原SDS处理:在上样buffer中加入SDS和DTT(或Beta巯基乙醇)后,蛋白质构象被解离,电荷被中和,形成SDS与蛋白相结合的分子,在电泳中,只根据分子量来分离。一般电泳均按这种方式处理,样品稀释适当浓度,加入上样Buffer,离心,沸水煮5min,再离心加样。 2)带有烷基化作用的还原SDS处理:碘乙酸胺的烷基化作用可以很好的并经久牢固的保护SH基团,得到较窄的谱带;另碘乙酸胺可捕集过量的DTT,而防止银染时的纹理现象。100ul样品缓冲液中10ul 20%的碘乙酸胺,并在室温保温30min。 3)非还原SDS处理:生理体液、血清、尿素等样品,一般只用1%SDS沸水中煮3min,未加还原剂,因而蛋白折叠未被破坏,不可作为测定分子量来使用。
还原SDS处理:在上样buffer中加入SDS和DTT(或Beta巯基乙醇)后,蛋白质构象被解离,电荷被中和,形成SDS与蛋白相结合的分子,在电泳中,只根据分子量来分离。一般电泳均按这种方式处理,样品稀释适当浓度,加入上样Buffer,离心,沸水煮5min,再离心加样。 2)带有烷基化作用的还原SDS处理:碘乙酸胺的烷基化作用可以很好的并经久牢固的保护SH基团,得到较窄的谱带;另碘乙酸胺可捕集过量的DTT,而防止银染时的纹理现象。100ul样品缓冲液中10ul 20%的碘乙酸胺,并在室温保温30min。 3)非还原SDS处理:生理体液、血清、尿素等样品,一般只用1%SDS沸水中煮3min,未加还原剂,因而蛋白折叠未被破坏,不可作为测定分子量来使用。")
70
SDS-PAGE电泳凝胶中各主要成分的作用?
聚丙烯酰胺的作用:丙烯酰胺与为蛋白质电泳提供载体,其凝固的好坏直接关系到电泳成功与否,与促凝剂及环境密切相关; 制胶缓冲液:浓缩胶选择pH6.7,分离胶选择pH8.9,选择tris-HCL系统, TEMED与AP:AP提供自由基,TEMED是催化剂,催化自由基引起的聚合反应进行; 十二烷基硫酸钠(SDS):阳离子去污剂,作用有四:去蛋白质电荷、解离蛋白质之间的氢键、取消蛋白分子内的疏水作用、去多肽折叠。
:阳离子去污剂,作用有四:去蛋白质电荷、解离蛋白质之间的氢键、取消蛋白分子内的疏水作用、去多肽折叠。")
71
提高SDS-PAGE电泳分辨率的途径? 聚丙烯酰胺的充分聚合,可提高凝胶的分辨率。
建议做法:待凝胶在室温凝固后,可在室温下放置一段时间使用。忌即配即用或4度冰箱放置,前者易导致凝固不充分,后者可导致SDS结晶。一般凝胶可在室温下保存4天,SDS可水解聚丙烯酰胺。 一般常用的有氨基黑、考马斯亮蓝、银染色三种染料,不同染料又各自不同的染色方法,具体可参照郭尧君编著的《蛋白质电泳技术手册》P82-103。

72
“ 微笑”(两边翘起中间凹下)形带原因? “皱眉”(两边向下中间鼓起)形带原因? 为什么带出现拖尾现象?
主要是由于凝胶的中间部分凝固不均匀所致,多出现于较厚的凝胶中。 处理办法:待其充分凝固再作后续实验。 “皱眉”(两边向下中间鼓起)形带原因? 主要出现在蛋白质垂直电泳槽中,一般是两板之间的底部间隙气泡未排除干净。 处理办法:可在两板间加入适量缓冲液,以排除气泡。 为什么带出现拖尾现象? 主要是样品融解效果不佳或分离胶浓度过大引起的。 处理办法:加样前离心;选择适当的样品缓冲液,加适量样品促溶剂;电泳缓冲液时间过长,重新配制;降低凝胶浓度。
形带原因? 主要出现在蛋白质垂直电泳槽中,一般是两板之间的底部间隙气泡未排除干净。 处理办法:可在两板间加入适量缓冲液,以排除气泡。 为什么带出现拖尾现象? 主要是样品融解效果不佳或分离胶浓度过大引起的。 处理办法:加样前离心;选择适当的样品缓冲液,加适量样品促溶剂;电泳缓冲液时间过长,重新配制;降低凝胶浓度。")
73
为什么带出现纹理现象? 为什么溴酚蓝不能起到指示作用? 为什么电泳的条带很粗?
主要是样品不溶性颗粒引起的。 处理办法:加样前离心;加适量样品促溶剂。 为什么溴酚蓝不能起到指示作用? 我们在实验中常会遇到溴酚蓝已跑出板底,但蛋白质却还未跑下来的现象。主要与缓冲液和分离胶的浓度有关。 处理办法:更换正确pH值的Buffer;降低分离胶的浓度。 为什么电泳的条带很粗? 电泳中条带很粗是常见的事,主要是未浓缩好的原因。 处理办法:适当增加浓缩胶的长度;保证浓缩胶贮液的pH正确(6.7);适当降低电压。
;适当降低电压。")
74
什么是“鬼带”,如何处理? “鬼带”就是在跑大分子构象复杂的蛋白质分子时,常会出现在泳道顶端(有时在浓缩胶中)的一些大分子未知条带或加样孔底部有沉淀,主要由于还原剂在加热的过程中被氧化而失去活性,致使原来被解离的蛋白质分子重新折叠结合和亚基重新缔合,聚合成大分子,其分子量要比目标条带大,有时不能进入分离胶。但它却于目标条带有相同的免疫学活性,在WB反应中可见其能与目标条带对应的抗体作用。 处理办法:在加热煮沸后,再添加适量的DTT或Beta巯基乙醇,以补充不足的还原剂;或可加适量EDTA来阻止还原剂的氧化。
的一些大分子未知条带或加样孔底部有沉淀,主要由于还原剂在加热的过程中被氧化而失去活性,致使原来被解离的蛋白质分子重新折叠结合和亚基重新缔合,聚合成大分子,其分子量要比目标条带大,有时不能进入分离胶。但它却于目标条带有相同的免疫学活性,在WB反应中可见其能与目标条带对应的抗体作用。 处理办法:在加热煮沸后,再添加适量的DTT或Beta巯基乙醇,以补充不足的还原剂;或可加适量EDTA来阻止还原剂的氧化。")
75
浓缩胶与分离胶断裂、板间有气泡对电泳有影响吗?
为什么电泳电压很高而电流却很低呢? 这种现象一般初学者易出现。比如电压50v以上,可电流却在5mA以下。主要是由于电泳槽没有正确装配,电流未形成通路。包括:a.内外槽装反;b.外槽液过少;c.电泳槽底部的绝缘体未去掉(比如倒胶用的橡胶皮)。 处理办法:电泳槽正确装配即可。 浓缩胶与分离胶断裂、板间有气泡对电泳有影响吗? 这主要出现在初学者中,一般对电泳不会有太大的影响。前者主要原因是拔梳子用力不均匀或过猛所致;后者是由于在解除制胶的夹子后,板未压紧而致空气进入引起的。
。 处理办法:电泳槽正确装配即可。 浓缩胶与分离胶断裂、板间有气泡对电泳有影响吗? 这主要出现在初学者中,一般对电泳不会有太大的影响。前者主要原因是拔梳子用力不均匀或过猛所致;后者是由于在解除制胶的夹子后,板未压紧而致空气进入引起的。")
76
凝胶时间不对,或慢或快,怎么回事? 电泳时间比正常要长?
通常胶在30MIN-1H内凝。如果凝的太慢,可能是TEMED,APS剂量不够或者失效。APS应该现配现用,TEMED不稳定,易被氧化成黄色。 如果凝的太快,可能是APS和TEMED用量过多,此时胶太硬易裂,电泳时易烧胶。 电泳时间比正常要长? 可能由于凝胶缓冲系统和电级缓冲系统地pH选择错误,即缓冲系统地pH和被分离物质的等电点差别太小,或缓冲系统的离子强度太高。

77
Western Blot常见问题分析 在western实验中,通常有人采用TBS,有人采用PBS,在使用中有什么区别
选择合适的缓冲液对于维持一定pH值下蛋白质的稳定及保证实验的重复性是很重要的。PBS的缓冲能力强于TBS,TBS在PH7.0以下缓冲能力较弱(不过PBS易污染)。但我们常常要根据我们实验目的选用合适的缓冲液,如在蛋白纯化中进行阴离子交换层析,阳离子缓冲液首选TBS;阳离子交换层析时,阴离子缓冲液则首选PBS。western blot中使用TBS和PBS均可,只是要根据需要选择合适的浓度。 没有注明可以用来做western blot的一抗,可以用来做western blot吗 不一定,因为有的抗体是识别线性表位的,有的是识别构象型表位的,识别构象型表位的抗体不能用于western blotting,因为在western blotting中抗体识别的是完全变性的蛋白质抗原,而蛋白质抗原的构象型表位变性时被破坏。
。但我们常常要根据我们实验目的选用合适的缓冲液,如在蛋白纯化中进行阴离子交换层析,阳离子缓冲液首选TBS;阳离子交换层析时,阴离子缓冲液则首选PBS。western blot中使用TBS和PBS均可,只是要根据需要选择合适的浓度。 没有注明可以用来做western blot的一抗,可以用来做western blot吗. 不一定,因为有的抗体是识别线性表位的,有的是识别构象型表位的,识别构象型表位的抗体不能用于western blotting,因为在western blotting中抗体识别的是完全变性的蛋白质抗原,而蛋白质抗原的构象型表位变性时被破坏。")
78
做western每次只加一种一抗,可以同时加两种或者多种一抗的吗?
最好还是不要同时加两种一抗。因为如果结果里面有非特异信号的话,就说不清楚了。做Western在同一张膜上检测目的蛋白和内对照,也是先上目的蛋白的一抗、二抗,ECL显色、压片、洗片;漂洗以后,再上内对照的一抗、二抗,ECL显色、压片、洗片。 western blot 使用的膜哪种最好啊,PVDF好还是NC膜,如何选择? PVDF膜价格较贵,可重复使用,特别适合蛋白印迹,结合能力较强,但价格比较昂贵。 NC膜价格比较便宜,应用较广,结合牢固性差一些,韧性也不如PVDF,不能重复使用,但蛋白吸附容量高,亲水性较好。 都可以用丽春红染色。具体使用还要参考相关文献。

79
为什么出现了多条带,每个加样槽中都出现了多条带,而且好象都很有规律,为什么?是封闭时间不够吗?做WESTERN必须要内参吗?
一抗、或二抗抗体浓度太高,多克隆抗体本身的非特异性反应,抗体的特异性不强,目的蛋白经过处理后发生变化,而内参的条带基本均匀一致.这样才有说服力,表明的确是处理因素造成目的蛋白的变化而不是加样误差或认为的造成目的条带浓度的变化.所以严格意义上说,内参是必须做的。

80
western用的一抗,单抗好还是用多抗好?用于western的抗体和用于ELISA的抗体有什么不同的要求?
作为western来讲,理论上单抗比多抗的特异性要好,但单抗种类较少一般价格偏高,所以一般来说多抗足够了。用于western的抗体和用于ELISA的抗体,一般来说用于western的抗体识别的是氨基酸序列特异性;而可用于ELISA的抗体则要看抗原种类,有些是识别氨基酸序列特异性,有些是识别构像特异性。所以一般根据抗体说明书来确定。 做免疫印迹时选择抗体主要应考虑两个问题,一是所选抗体是否能识别凝胶电泳后转印至膜上的变性蛋白,另一个是所选抗体是否会引起交叉反应条带。

81
半干转是否要求膜,滤纸,胶同样大小?因为胶大小不一定规则,膜和滤纸会大一点,那样会不会短路?什么是短路?如何控制和发现短路呢?
可以相等,但是如果纸、膜比凝胶大就比较容易形成短路了,上下两层虑纸也不能因过大而相互接触,这样同样会短路,电流不会通过胶和虑纸。转移前和转移过程中看看电压就行,正常的半干是慢慢变高的, 最后结束时一般是开始的1.5-3倍都是正常的。一般Buffer和滤纸选的对就不会短路。

82
Western Blot哪种染色好? 阴离子染料是较常用的,特别是氨基黑,脱色快,背景低检测极限可达到1.5μg,考马斯亮蓝虽然与氨基黑有相同的灵敏度,但脱色慢,背景高。丽春红S和快绿在检测后容易从蛋白质中除去 ,以便进行随后的氨基酸分析。缺点是:溶剂系统的甲醇会引起硝化纤维素膜的 皱缩或破坏。不能用语正电贺的膜。灵敏度低。 胶体金,灵敏度高,检测范围可到pg级,但染色比稳定。 生物素化灵敏度位于1、2之间,可用于任何一种膜。

83
不能很好地将大分子量蛋白转移到膜上, 转移效率低怎么样解决?
可以考虑:转移缓冲液中加入20%甲醇(是指终浓度)(优化的转移缓冲液,可以参考《蛋白质技术手册》),因为甲醇可降低蛋白质洗脱效率,但可增加蛋白质和NC膜的结合能力,甲醇可以防止凝胶变形,甲醇对高分子量蛋白质可延长转移时间;转移缓冲液加入终浓度0.1%SDS,也是为了增加转移效率;用优质的转移膜,或使用小孔径的NC膜(0.2微米);使用戊二醛交联;低浓度胶,如低至6%。太大时还可以考虑用琼脂糖胶;提高转移电压/电流;增加转移时间。
(优化的转移缓冲液,可以参考《蛋白质技术手册》),因为甲醇可降低蛋白质洗脱效率,但可增加蛋白质和NC膜的结合能力,甲醇可以防止凝胶变形,甲醇对高分子量蛋白质可延长转移时间;转移缓冲液加入终浓度0.1%SDS,也是为了增加转移效率;用优质的转移膜,或使用小孔径的NC膜(0.2微米);使用戊二醛交联;低浓度胶,如低至6%。太大时还可以考虑用琼脂糖胶;提高转移电压/电流;增加转移时间。")
84
DAB显色、碱性磷酸酶-抗碱性磷酸酶显色原理是什么?
DAB显色在辣根过氧化酶的作用能形成一种灰褐色的终末产物,该产物难溶于醇和其他有机溶剂,DAB的氧化还能引起聚合作用,导致与四氧化锇反应而增加其染色强度和电子密度。碱性磷酸酶-抗碱性磷酸酶显色中,酶将奈酚磷酸(底物)水解成酚类和磷酸。酚和无色的重氮盐(显色原)结合而产生有色的、不溶性偶氮染料。
水解成酚类和磷酸。酚和无色的重氮盐(显色原)结合而产生有色的、不溶性偶氮染料。")
85
酶显色与荧光显色各自的优缺点是什么? 免疫酶技术就是用酶(如辣根过氧化物酶)标记已知抗体(或抗原),然后与组织标本在一定条件下反应,如果组织中含有相应抗原(或抗体),抗原抗体相互结合形成的复合物中所带酶分子遇到底物时,能催化底物水解、氧化或还原,产生显色反应,这样就可以识别出标本抗原(抗体)分布的位置和性质,通过图像分析并可达到定量的目的。 免疫荧光技术虽已广泛应用于免疫学的研究与诊断,但是荧光抗体染色标本不能长期保存,对组织细胞的细微结构分辨不清,免疫酶技术则能克服上述不足,标记免疫酶技术的敏感性更优于免疫荧光法,对石蜡切片标本尤为适用,为免疫病理研究开辟了一条新途径,酶显色产物具有较高的电子密度,经过适当处理还可以进行免疫电镜观察,免疫酶组化技术分为酶标记法与非标记抗体技术,前者是将酶通过交联剂结合在抗体分子上,形成酶标记抗体。后者是将酶作为抗原与相应的特异性抗体连接进行的免疫反应,称为非标记抗体酶技术。
标记已知抗体(或抗原),然后与组织标本在一定条件下反应,如果组织中含有相应抗原(或抗体),抗原抗体相互结合形成的复合物中所带酶分子遇到底物时,能催化底物水解、氧化或还原,产生显色反应,这样就可以识别出标本抗原(抗体)分布的位置和性质,通过图像分析并可达到定量的目的。 免疫荧光技术虽已广泛应用于免疫学的研究与诊断,但是荧光抗体染色标本不能长期保存,对组织细胞的细微结构分辨不清,免疫酶技术则能克服上述不足,标记免疫酶技术的敏感性更优于免疫荧光法,对石蜡切片标本尤为适用,为免疫病理研究开辟了一条新途径,酶显色产物具有较高的电子密度,经过适当处理还可以进行免疫电镜观察,免疫酶组化技术分为酶标记法与非标记抗体技术,前者是将酶通过交联剂结合在抗体分子上,形成酶标记抗体。后者是将酶作为抗原与相应的特异性抗体连接进行的免疫反应,称为非标记抗体酶技术。")
Similar presentations

。>")
 是用标记的特异性抗体(或抗原)对 组织内的抗原(或抗体)的分布进行定位、 定性、定量检测,经过组织化学呈色反应.>")
















