Download presentation
1
染色质与表观遗传调控 Chromatin and Epigenetic Regulation
三峡大学医学院 盛 德 乔

2
一、表观遗传学的概念

3
Epi-genetics Epi—“在…上面”,“在…之上”,“在…之外,另加”之义 不遵守经典的遗传学规律!!

4
经典遗传学 Law of segregation 分离规律 Law of Independent Assortment自由组合规律
遗传学之父,发现了遗传学基本规律。 ( Published in 1865 and 1866 and “re-discovered” in 1900 ) Gregor Mendel ( )
 Gregor Mendel ( )")
5
基因连锁互换定律 (Published in 1915) Thomas Hunt Morgan ( )
 Thomas Hunt Morgan ( )")
6
经典的遗传学规律!! 基因型决定表现型 ? 基因型 (碱基排列顺序) 中心法则 基因决定论 表现型 (生物学性状)
 中心法则 基因决定论 表现型 (生物学性状)")
7
表现型改变了,但其碱基序列并未发生改变??
异常现象: 基因型相同,但表现型不同? 表现型改变了,但其碱基序列并未发生改变??

8
From Phenomenon to Field
Epigenetics: From Phenomenon to Field

9
经典的遗传学无法解释 —同卵双胞胎的差异 同卵双胞胎——相貌、气质、体质及一些其他特征非常相似(如皮纹、头发的颜色、头发的形式和密度、眼虹膜的颜色和结构、前后发际、耳廓、鼻形、脸形、唇形、眼睑、皮肤的颜色和结构、雀斑等)
")
11
研究者发现,一些孪生子的情况并不符合预期的理论。往往在长大成人后出现性格、健康方面的很大差异。这种反常现象长期困扰着遗传学家。

12
自然界不符合经典遗传学规律现象 动物正交/反交差异
骡子是马和驴种间交配产生的种间杂种,有雌雄之分,但是没有生育的能力。具有劲头大、耐力强、健壮、不易病而且耐粗饲等优于双亲的特点,是“杂种优势”的典型例子。 骡子的母亲决定后代更像马或驴,有马骡和驴骡之分。由公驴和母马所生又称为马骡,反过来则称为驴骡。

13
马骡力大无比,是马和驴远远不可相及的;而驴骡则善于奔跑,也是驴所无法比拟的。
驴骡像驴,马骡像马,说的当然不只是外表方面,内在品质上它们也是各有特色 。 Why?

14
虎狮兽

15
狮虎兽 Liger

16
虎狮兽:体形比虎或狮都小,雄性不育 狮虎兽:体形比虎和狮都大,雄性不育 虎、狮正反交的后代差别较大;同卵双生的两人具有完全相同的基因组,在同样的环境中长大后,他们在性格、健康等方面会有较大的差异,并不符合经典遗传学理论预期的情况。

17
老虎控制体型生长的基因是靠公虎遗传的,狮子控制体型生长的基因却是靠母狮遗传的(基因印记
老虎控制体型生长的基因是靠公虎遗传的,狮子控制体型生长的基因却是靠母狮遗传的(基因印记!),所以公虎母狮所生的虎狮兽受到双重的基因控制变得明显比狮子或老虎都娇小许多反而比較像猫或豹。 问题? 基因印记?
,所以公虎母狮所生的虎狮兽受到双重的基因控制变得明显比狮子或老虎都娇小许多反而比較像猫或豹。 问题? 基因印记")
18
在相应的基因碱基序列没有发生变化的情况下,一些生物体的表型却发生了改变!?
基因型—表现型?

19
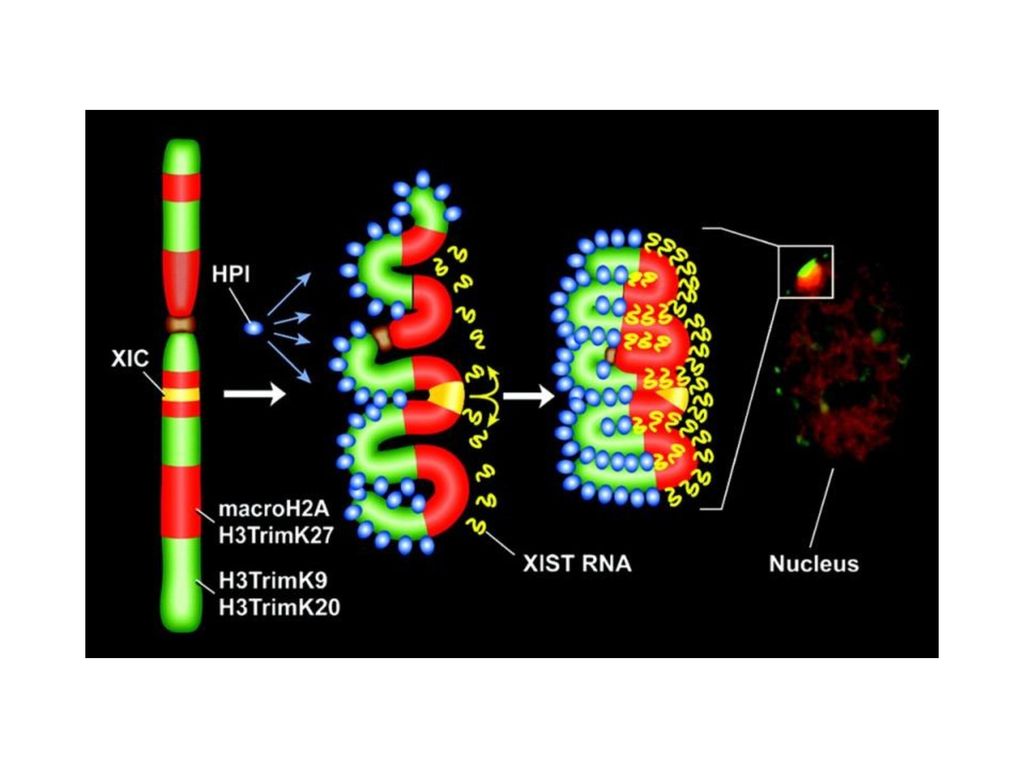
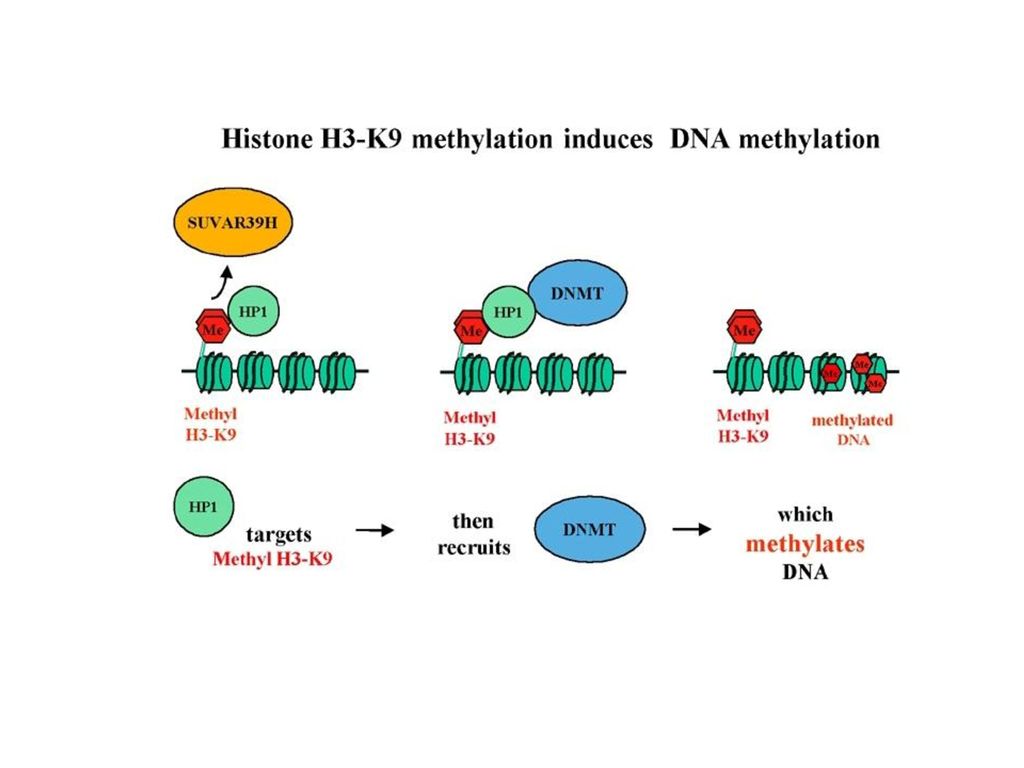
X-染色体失活 一般女性虽然都有两条X染色体,但事实上有一条会惰性化,变成巴尔体(Barr body),这个作用称为X去活性(X inactivation)或里昂化作用(lyonization,以发现者Mary F. Lyon命名)。 机制? DNA 甲基化 组蛋白修饰 RNAi

20
实验证据 以基因型为a/a的母鼠及其孕育的基因型为AVY/a的仔鼠作实验对象。
孕鼠分为两组,试验组孕鼠除喂以标准饲料外,从受孕前两周起还增加富含甲基的叶酸、乙酰胆碱等补充饲料,而对照组孕鼠只喂饲标准饲料。

21
结果实验组孕鼠产下的仔鼠大多数在身体的不同部位出现了大小不等的棕色斑块,甚至出现了以棕褐色为主要毛色的小鼠。而对照组孕鼠的仔鼠大多数为黄色。
From Dr. Randy Jirtle's laboratory at Duke University
22
分析表明喂以富甲基饲料的孕鼠所产仔鼠的IAP所含CpG岛的甲基化平均水平远高于对照组,转录调控区的高甲基化使原该呈异位表达的基因趋于沉默,毛色也趋于棕褐色。 (DNA 甲基化影响基因表达水平!)
")
23
科学家们发现:可以在不影响DNA序列的情况下改变基因组(DNA/组蛋白)的修饰.这种改变不仅可以影响个体的发育(?),而且还可以遗传(?)下去。
表观遗传学 Epigenetics DNA修饰:甲基化 组蛋白修饰:甲基化、乙酰化、磷酸化…… 表观遗传修饰!

24
表观遗传学 表观遗传学——是研究没有DNA序列变化,可遗传的基因表达(活性)的改变。
主要涉及DNA甲基化作用的改变、染色质组蛋白的修饰作用、基因印记、RNAi,等

25
表观遗传学的特点: 可遗传的,即这类改变通过有丝分裂或减数 分裂,能在细胞或个体世代间遗传;
可遗传的,即这类改变通过有丝分裂或减数 分裂,能在细胞或个体世代间遗传; 可逆性的基因表达调节,也有较少的学者描 述为基因活性或功能的改变; 没有DNA序列的改变或不能用DNA序列变化 来解释。

26
表观遗传学的研究内容 表观遗传学的概念; 表观遗传调控的分子机制; 表观遗传学的应用。

27
发展简史 1942年,Conrad Hal Waddington提出现代Epigenetics的概念,认为基因型通过一些“偶然的、不确定的机制”决定了不同的表型 1941年,Hermann J. Muller发现Position-effect variegation (PEV)——第一种表观遗传学现象 1958年,R.A.Brink发现Paramutation现象 1961年,Mary Lyon发现X染色体失活现象 1983年,DNA甲基化的发现
——第一种表观遗传学现象. 1958年,R.A.Brink发现Paramutation现象. 1961年,Mary Lyon发现X染色体失活现象. 1983年,DNA甲基化的发现.")
28
1993年, miRNA & siRNA 1998年,Non-coding RNA 2000年,组蛋白共价修饰(Histone modification) &组蛋白密码( Histon code ) 2004年,提出表观遗传学现象 2006年, Long non-coding RNA(lncRNA) 2008年,“Epigenetics” 形成共识 2010年,RNA 甲基化(?)
 2008年, Epigenetics 形成共识. 2010年,RNA 甲基化(?)")
29
Waddington's epigenetics
基因型 表型

30
基因型 表型

31
In the summer of 2004, the 69th Cold Spring Harbor Symposium on Quantitative Biology
David Haig define an epigenetic phenomenon as a change in phenotype that is heritable but does not involve DNA mutation.

32
Berger S, Kouzarides T, Shiekhattar R, Shilatifard A (2009)
Berger S, Kouzarides T, Shiekhattar R, Shilatifard A (2009). "An operational definition of epigenetics". Genes Dev. 23 (7): 781–3
. An operational definition of epigenetics . Genes Dev. 23 (7): 781–3.")
33
二、染色质结构与表观 遗传调控

34
Multiple levels of chromatin folding

35
染色质结构影响基因表达? 常染色质 常染色质 ? 异染色质 异染色质(核内深染部分) 常染色质(核内浅染部分)
 常染色质(核内浅染部分)")
36
核心组蛋白的结构 核小体是真核细胞染色质的基本单位。核小体的核心结构是由146个碱基对的DNA盘绕四对核心组蛋白的八聚体外面近两周所构成。
核心组蛋白组成 H2A/H2B H3 H4

37
核心组蛋白—N-端尾巴和保守的组蛋白折叠结构域组成,是一类较小而带有丰富正电荷的核蛋白。
非常保守! 近年成为研究的热点之一。 核小体组装是一个干扰DNA复制、基因表达和细胞周期进展的过程,因此在细胞生命过程中极为重要!

38
染色质结构或状态的改变: 组蛋白的共价修饰 DNA的甲基化 ……

39
N-末端共价修饰 H3和H4组蛋白富含赖氨酸(K)和精氨酸(R),在H3/H4的N端含有多个可以受乙酰化调节的赖氨酸残基,以及受甲基化调节的赖氨酸和精氨酸残基;丝氨酸残基的磷酸化也在调节组蛋白修饰进程中起作用。(共价修饰!) 表观遗传修饰的基础!

40
组蛋白N-端尾巴的共价修饰

41
DNA甲基化 所谓DNA甲基化是指在DNA甲基化转移酶的作用下,在基因组CpG二核苷酸的胞嘧啶5 碳位共价键结合一个甲基基团。
5-甲基化胞嘧啶(5mC) DNA甲基化可抑制基因的表达。
 DNA甲基化可抑制基因的表达。")
42
组蛋白修饰 DNA甲基化 表观遗传机制

43
表观遗传学的研究内容 表观遗传现象 表观遗传修饰 基因组印记 X染色体失活 副突变 位置效应异质性 … … DNA 甲基化 组蛋白修饰
染色质重塑 RNA编辑 非编码RNA(non-coding RNA):siRNA, miRNA,lncRNA… … …
:siRNA, miRNA,lncRNA… … …")
44
三、表观遗传现象

45
X 染色体失活 在哺乳动物中,雌、雄性个体X 染色体的数目不同,这类动物需要以一种方式来解决X 染色体剂量的差异。
哺乳动物雌性个体的X染色体失活遵循n-1法则,不论有多少条X染色体,最终只能随机保留一条的活性。

46
在雌性哺乳动物中,两条X 染色体有一个是失活的,称为X染色体的剂量补偿。X染色体失活的选择和起始发生在胚胎发育的早期,这个过程被X 失活中心(X - inactivation center , Xic) 所控制,是一种反义转录调控模式。 X 染色体的失活状态需要表观遗传修饰如DNA 甲基化来维持。这种失活可以通过有丝或减数分裂遗传给后代。

47
Xic长约1Mb,包括4个已知基因:Xist、Xce、Tsix和DXPas34。
Exist:X染色体失活特异性转录因子,是X染色体上启动转录最早的基因。 Xce:在基因组中的组成与X-染色体随机失活中的选择有关,当Xce处于纯合状态时,体细胞中的X-失活是完全随机的,而杂合态时,失活不是完全随机的。 Tsix:位于Xist下游的瞬时调控元件, 与CTCF可能协同起着Xist的外源开关功能。 DXPas34:富含CpG,包括一个15Kb的微卫星重复序列,对X-失活有一定调控作用。

49
X染色体失活 X染色体失活过程模式图

50
X-Inactivation in Mammalian Females
玳瑁猫

51
基因组印记 (genomic imprinting)
高等动物的基因组为两倍体,受精卵从双亲中各继承了一套基因组。每一个常染色体均为双拷贝,父源和母源常染色体基因都能有均等的机会表达或受到抑制,这是孟德尔遗传定律的基本要素。

52
上世纪90年代开始,发现了不符合这个定律的遗传现象。某些基因呈单等位基因表达,即父源与母源的基因拷贝不能同时表达,并且,父源或母源等位基因通过某种特异的基因修饰机制,特异性地抑制另一母源或父源染色体等位基因表达。

53
来自双亲的某些等位基因,在子代的表达不同,有些只有父源的基因有转录活性,而母源的同一基因则始终处于沉默状态,另一些基因的情况则相反。
胰岛素样生长因子2基因(insulin-like growth factor 2, IGF2)只表达源自父亲的等位基因,母源等位基因被抑制。 胰岛素样生长因子2受体基因(insulin-like growth factor 2 receptor, IGF2R)为源自父亲的等位基因不表达,只表达母源等位基因。
只表达源自父亲的等位基因,母源等位基因被抑制。 胰岛素样生长因子2受体基因(insulin-like growth factor 2 receptor, IGF2R)为源自父亲的等位基因不表达,只表达母源等位基因。")
54
♀ ♂ 正 交 反 交 Igf-2 Igf-2m Igf-2m Igf-2 正常小鼠 矮小型小鼠 矮小型小鼠 正常小鼠 Igf-2m
正 交 反 交 Igf-2 Igf-2m ♀ ♂ Igf-2m Igf-2 正常小鼠 矮小型小鼠 矮小型小鼠 正常小鼠 Igf-2m Igf-2 Igf-2 Igf-2m 正常小鼠 矮小型小鼠 54 54

55
由正反交实验可以看出: 印迹基因的正反交结果不一致、不符合孟德尔定律。
小鼠 Igf-2 基因总是母本来源的等位基因被印迹,父本来源的等位基因表达,因此是母本印迹。 基因印迹使基因的表达受到抑制,导致被印迹的基因的生物功能的丧失。 55 55

56
这种现象可能是在父母受精卵形成过程中,特异性地对源自父亲或源自母亲的等位基因做一印记使其只表达父源或母源等位基因。这种现象被称作基因印记(gene imprinting)或基因组印记(genomic imprinting)。
或基因组印记(genomic imprinting)。")
57
四、表观遗传调控的 分子机制

58
表观遗传调控 DNA修饰-甲基化 核心组蛋白修饰-组蛋白密码(Histone code)
染色质重塑(Chromatin Remodeling)ATP-依赖 Non-coding RNA (Regulatory RNA): siRNA , miRNA, Long non-coding RNA RNA编辑(RNA editing) ……
ATP-依赖. Non-coding RNA (Regulatory RNA): siRNA , miRNA, Long non-coding RNA. RNA编辑(RNA editing) ……")
59
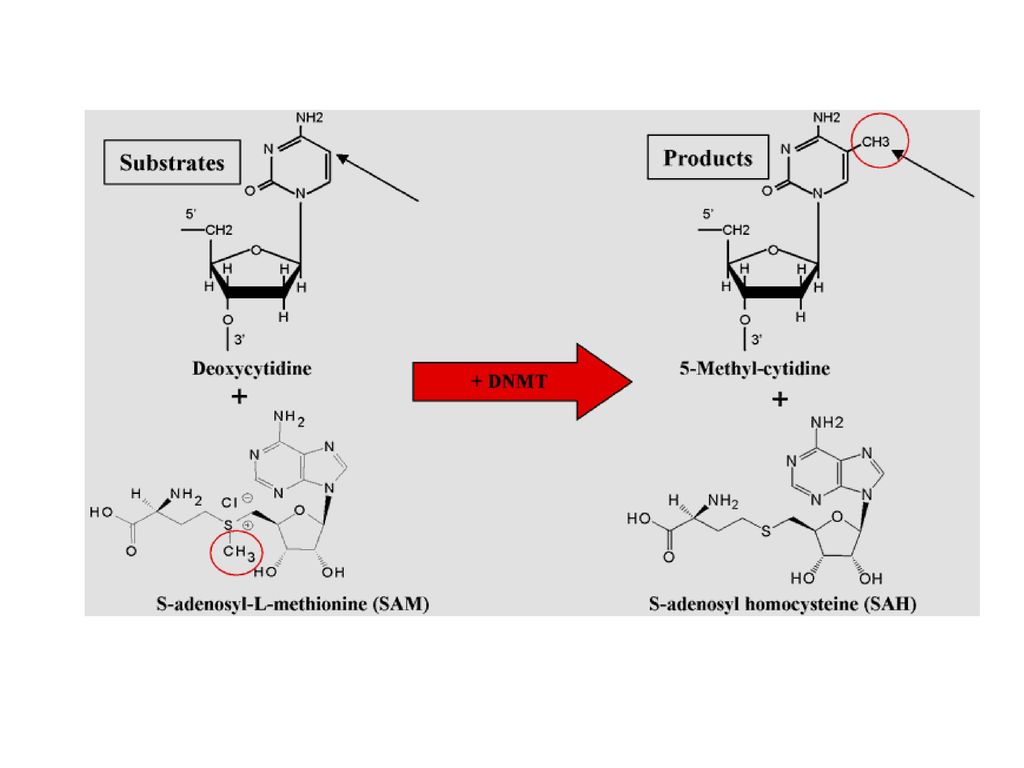
DNA甲基化 所谓DNA甲基化是指在DNA甲基化转移酶的作用下,在基因组CpG二核苷酸的胞嘧啶5 碳位共价键结合一个甲基基团。
5-甲基化胞嘧啶(5mC) 这种DNA修饰方式并没有改变基因序列,但它由此调控了基因的表达。 甲基化是基因组DNA 的一种主要表观遗传修饰形式,是调节基因组功能的重要手段。
 这种DNA修饰方式并没有改变基因序列,但它由此调控了基因的表达。 甲基化是基因组DNA 的一种主要表观遗传修饰形式,是调节基因组功能的重要手段。")
60
哺乳动物中,CpG序列在基因组中出现的频率仅有1%,远低于基因组中的其它双核苷酸序列。
在基因组的某些区域中,CpG序列密度很高,可以达均值的5倍以上,成为鸟嘌呤和胞嘧啶的富集区,形成所谓的CpG岛(CpG island) ,通常长度在1kb~2kb 左右。 CpG岛通常位于基因的启动子区或是第一个外显子区 。
 ,通常长度在1kb~2kb 左右。 CpG岛通常位于基因的启动子区或是第一个外显子区 。")
61
DNA甲基化沉默基因表达

62
DNA 甲基转移酶 DNA 甲基化主要是通过DNA 甲基转移酶家族(DNA methyltransferase, Dnmt ) 来催化。
在DNA 甲基化过程中,胞嘧啶突出于DNA 双螺旋并进入与胞嘧啶甲基转移酶结合部位的裂隙中,该酶将S - 腺苷甲硫氨酸(SAM) 的甲基转移到胞嘧啶的5′位,形成5 - 甲基胞嘧啶(5 - methylcytosine , 5mC) 。
 的甲基转移到胞嘧啶的5′位,形成5 - 甲基胞嘧啶(5 - methylcytosine , 5mC) 。")
64
甲基化CpG结合蛋白(MBD) 甲基化的CpG位点由特异的蛋白质识别,这类蛋白称为DNA甲基化结合蛋白(MBD家族)
MeCP2(methyl-CpG-binding protein 2) 单mCpG结合蛋白 MBD1 MBD2 MBD3 MBD4 MeCP1是甲基化结合蛋白,但是后来发现它不是一个单一的蛋白质,而一是个蛋白的复合体,即主要由MBD3、MBD2、HDAC1 /2 和RbAp46 /48 等所组成。
 单mCpG结合蛋白. MBD1. MBD2. MBD3. MBD4. MeCP1是甲基化结合蛋白,但是后来发现它不是一个单一的蛋白质,而一是个蛋白的复合体,即主要由MBD3、MBD2、HDAC1 /2 和RbAp46 /48 等所组成。")
65
MBD介导的转录抑制— 募集转录抑制相关蛋白

66
DNA甲基化的调控机理 DNA甲基化直接干扰特异性转录因子与各基因启动子中识别位置的结合。
序列特异性的甲基化DNA结合蛋白与启动子区甲基化CpG岛结合,募集一些蛋白,形成转录抑制复合物,阻止转录因子与启动子区靶序列的结合,从而影响基因的转录。 DNA甲基化通过改变染色质结构,抑制基因表达。染色质构型的变化伴随着组蛋白的乙酰化和去乙酰化。

67
DNA甲基化的生物学作用 DNA甲基化与遗传印记、胚胎发育 DNA甲基化与肿瘤
甲基化状态的改变是引起肿瘤的一个重要因素,这种变化包括基因组整体甲基化水平降低和CpG岛局部甲基化水平的异常升高,从而导致基因组的不稳定(如染色体的不稳定、可移动遗传因子的激活、原癌基因的表达)和抑癌基因的不表达。如果抑癌基因中有活性的等位基因失活,则发生癌症的机率提高。
和抑癌基因的不表达。如果抑癌基因中有活性的等位基因失活,则发生癌症的机率提高。")
68
DNA甲基化与肿瘤 DNA甲基化是肿瘤中最常见的分子改变之一,包括基因组总体甲基化水平降低和某些基因启动区域发生高甲基化(hypermethylation)。 DNA甲基化和组蛋白去乙酰化都可以引起基因的转录抑制,这种调控机制是可逆的,因而对肿瘤临床治疗来说是非常令人振奋的靶点。目前,DNA甲基化酶抑制剂和组蛋白去乙酰化酶抑制剂都已经应用于临床。 甲基化转移酶抑制剂应用于肿瘤治疗 5 -氮胞苷(5 -aza-cytidine, 5 -aza-C) 5 -氮-2-脱氧胞苷
 5 -氮-2-脱氧胞苷.")
69
因甲基化而在肿瘤中沉默的基因 Cancer epigenetics

70
DNA 甲基化抑制剂

71
表观遗传调控 DNA修饰-甲基化 核心组蛋白修饰-组蛋白密码(Histone code)
染色质重塑(Chromatin Remodeling)ATP-依赖 Non-coding RNA (Regulatory RNA): siRNA , miRNA, Long non-coding RNA RNA编辑(RNA editing)
ATP-依赖. Non-coding RNA (Regulatory RNA): siRNA , miRNA, Long non-coding. RNA RNA编辑(RNA editing)")
72
核心组蛋白修饰 核心组蛋白N-末端氨基酸残基的共价修饰 乙酰化 Lys 甲基化 Lys , Arg 磷酸化 Ser , Thr 泛素化
SUMO化 ADP-核糖基化

73
组蛋白是染色体的基本单位 — 核小体的蛋白质组成部分在进化上十分保守与遗传信息DNA 的包装有关。
每个核心组蛋白都含有一个球 状体和一个氨基酸末端-即 “ 尾巴” (富含碱性氨基酸)。
。")
74
组蛋白是高度保守的碱性小分子; 碱性氨基酸残基易于共价修饰; 共价修饰是可逆的。 H1 H2A H2B H3 H4
Linker histone H2A H2B helix Core histones variable H3 H4 conserved N

75
组蛋白的修饰可通过影响组蛋白与DNA双链的亲和性,从而改变染色质的疏松或凝集状态,或通过影响其它转录因子与结构基因启动子的亲和性来发挥基因调控作用。组蛋白修饰对基因表达的调控有类似DNA遗传密码的调控作用。

76
一般来说,组蛋白乙酰化能选择性的使某些染色质区域的结构从紧密变得松散,开放某些基因的转录,增强其表达水平。(活性转录区的标志)
组蛋白甲基化既可抑制也可增强基因表达。乙酰化修饰和甲基化修饰往往是相互排斥的。 在细胞有丝分裂和凋亡过程中,磷酸化修饰能调控蛋白质复合体向染色质集结。

77
组蛋白乙酰化 HAT HDAC Acetyl-CoA

78
组蛋白甲基化 精氨酸 赖氨酸

79
组蛋白共价修饰 组蛋白修饰的书写 H3-K9Me H3-K9Me H3K9Ac

80
组蛋白不仅包装DNA成核小体,其末端的各种共价修饰也构成了独特的“组蛋白密码(histone code)”,可以被一系列特定蛋白质或蛋白质复合物所识别,从而将这种密码翻译成特定的染色质状态,调节基因的表达。
 ,可以被一系列特定蛋白质或蛋白质复合物所识别,从而将这种密码翻译成特定的染色质状态,调节基因的表达。")
81
组蛋白密码? 书写(Writer) 擦写(Eraser) 阅读(Reader)
 擦写(Eraser) 阅读(Reader)")
82
核心组蛋白的共价修饰 与组蛋白密码 组蛋白的共价修饰可直接影响染色质或核小体的结构;
组蛋白的共价修饰可募集(Recruit)其他转录因子参与调控基因表达。 “组蛋白密码”学说 Laboratory of Chromatin Biology and epigenetics , Rockefeller University Jenuwein, T. and Allis, C.D. (2001) Translating the histone code. Science 293,
其他转录因子参与调控基因表达。 组蛋白密码 学说. Laboratory of Chromatin Biology and epigenetics , Rockefeller University. Jenuwein, T. and Allis, C.D. (2001) Translating the histone code. Science 293,")
83
2015年生命科学突破奖 C. David Allis, The Rockefeller University, for the discovery of covalent modifications of histone proteins and their critical roles in the regulation of gene expression and chromatin organization, advancing the understanding of diseases ranging from birth defects to cancer.

84
核心组蛋白的共价修饰 乙酰化 Lys 甲基化 Lys , Arg 磷酸化 Ser , Thr 泛素化 SUMO化 ADP-核糖基化

85
组蛋白乙酰化 组蛋白乙酰化修饰 乙酰化干扰染色质的结构,有利于维持转录核小体的开放状态
组蛋白乙酰基转移酶 Histone acetyl transferase,HAT Acetyl-CoA 组蛋白去乙酰基酶 Histone deacetylase,HDAC 乙酰化干扰染色质的结构,有利于维持转录核小体的开放状态 活性转录区的标志!!!

86
组蛋白乙酰化与基因活化以及DNA复制相关,组蛋白的去乙酰化和基因的失活相关。
乙酰化转移酶(HATs)主要是在组蛋白H3、H4的N端尾上的赖氨酸加上乙酰基,去乙酰化酶(HDACs)则相反,不同位置的修饰均需要特定的酶来完成。
主要是在组蛋白H3、H4的N端尾上的赖氨酸加上乙酰基,去乙酰化酶(HDACs)则相反,不同位置的修饰均需要特定的酶来完成。")
87
乙酰化 去乙酰化

88
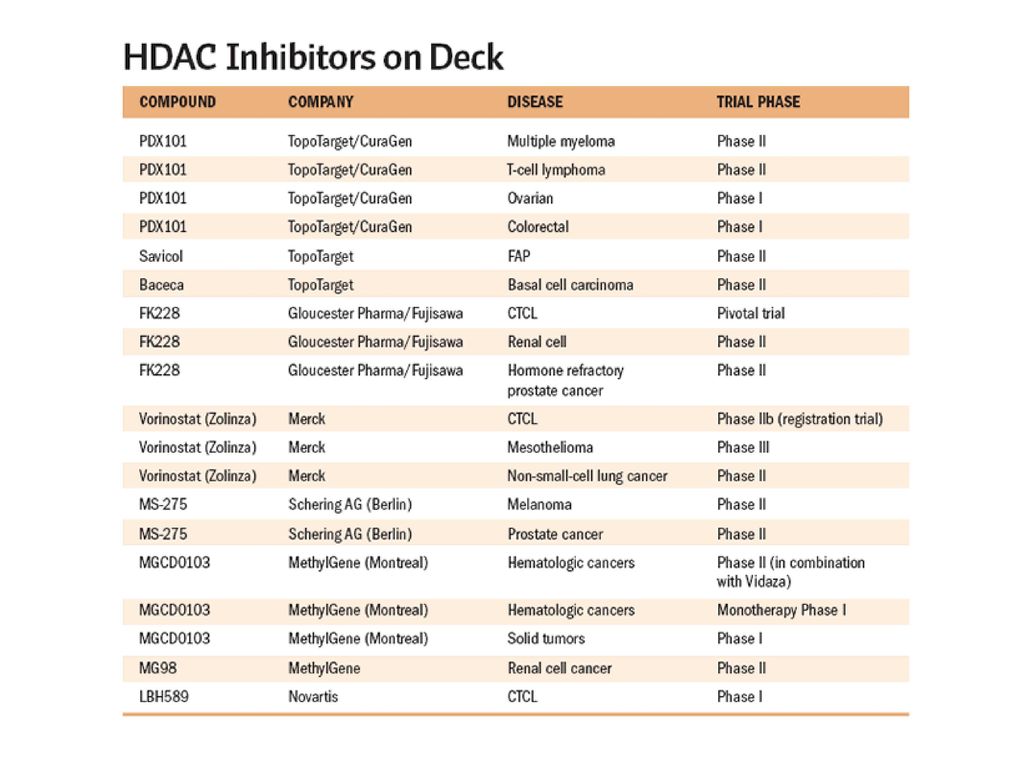
HDAC Inhibitors

90
核心组蛋白的共价修饰 乙酰化 Lys 甲基化 Lys , Arg 磷酸化 Ser , Thr 泛素化 SUMO化 ADP-核糖基化

91
组蛋白甲基化 Histone methylation
组蛋白甲基化修饰对基因转录的调控 赖氨酸残基(K)或精氨酸残基(R) 组蛋白甲基转移酶(histone methyltransferases, HMTs) 组蛋白去甲基化酶(histone demethylases, HDMs) 组蛋白赖氨酸甲基转移酶介导的 阻遏 (H3-K9, H3-K27, H4-K20) 组蛋白赖氨酸甲基转移酶介导的 激活 (H3-K4, H3-K36, H3-K79)
或精氨酸残基(R) 组蛋白甲基转移酶(histone methyltransferases, HMTs) 组蛋白去甲基化酶(histone demethylases, HDMs) 组蛋白赖氨酸甲基转移酶介导的 阻遏 (H3-K9, H3-K27, H4-K20) 组蛋白赖氨酸甲基转移酶介导的 激活 (H3-K4, H3-K36, H3-K79)")
92
精氨酸 赖氨酸

93
组蛋白赖氨酸甲基化酶及其功能 HMTs

94
The Methylation of Histone Lysines Is Reversible and Regulates Gene Expression

95
组蛋白甲基化和DNA甲基化系统可联合作用以建立一种长期的沉默,并且可通过DNA复制而传递下去。
组蛋白赖氨酸甲基化在表观遗传调控中,不仅不同位点的甲基化有不同的效应,而且同一位点不同的甲基数量其效应也不相同!!

96
不同位点精氨酸甲基化具有不同效应

97
核心组蛋白的共价修饰 乙酰化 Lys 甲基化 Lys , Arg 磷酸化 Ser , Thr 泛素化 SUMO化 ADP-核糖基化

98
组蛋白磷酸化 组蛋白磷酸化对于有丝分裂和减数分裂过程中染色质的凝聚及细胞周期的演进至关重要,除此之外还与真核基因的转录激活、细胞凋亡以及DNA的损伤和修复有关。 磷酸化的位点:Ser,Thr

99
核心组蛋白的共价修饰 乙酰化 Lys 甲基化 Lys , Arg 磷酸化 Ser , Thr 泛素化 SUMO化 ADP-核糖基化

100
组蛋白不同共价修饰间相互作用

103
表观遗传调控 DNA修饰-甲基化 核心组蛋白修饰-组蛋白密码(Histone code)
染色质重塑(Chromatin Remodeling)ATP-依赖 Non-coding RNA (Regulatory RNA): siRNA , miRNA, Long non-coding RNA RNA编辑(RNA editing)
ATP-依赖. Non-coding RNA (Regulatory RNA): siRNA , miRNA, Long non-coding RNA. RNA编辑(RNA editing)")
104
染色质重塑 chromatin remodeling
染色质重塑( remodeling) 指染色质位置和结构的变化。主要涉及密集的染色质丝在核小体连接处发生松解造成染色质解压缩,从而暴露基因转录启动子区中的顺式作用元件,为反式作用蛋白(转录因子) 与之结合提供了一种称为可接近性(accesibility) 的状态。
 指染色质位置和结构的变化。主要涉及密集的染色质丝在核小体连接处发生松解造成染色质解压缩,从而暴露基因转录启动子区中的顺式作用元件,为反式作用蛋白(转录因子) 与之结合提供了一种称为可接近性(accesibility) 的状态。")
105
ATP依赖的染色质重塑复合物(ATP-dependent chromatin remodeling complex)与组蛋白共价修饰、DNA分子甲基化等机制共同控制染色质的结构。
与组蛋白共价修饰、DNA分子甲基化等机制共同控制染色质的结构。")
106
chromatin remodeling complex

107
染色体重塑过程由两类结构介导: ATP 依赖型核小体重塑复合体 组蛋白修饰复合体 前者通过水解作用改变核小体构型;后者对核心组蛋白N 端尾部的共价修饰进行催化。修饰直接影响核小体的结构,并为其他蛋白提供了和DNA 作用的结合位点。

108
染色质重塑(chromatin remodeling)
广义-是指染色质结构的动态调整或重新塑造染色质结构。 狭义-专指ATP提供能量的、通过依赖ATP的染色质重塑复合物改变组蛋白与DNA的结合状态,在靠近核心组蛋白的DNA表面建立特殊的构象,使转录因子易于接近DNA的过程。

109
染色质结构的动态改变对基因的表达调控非常重要
染色质结构的改变是快速的、可逆的。

110
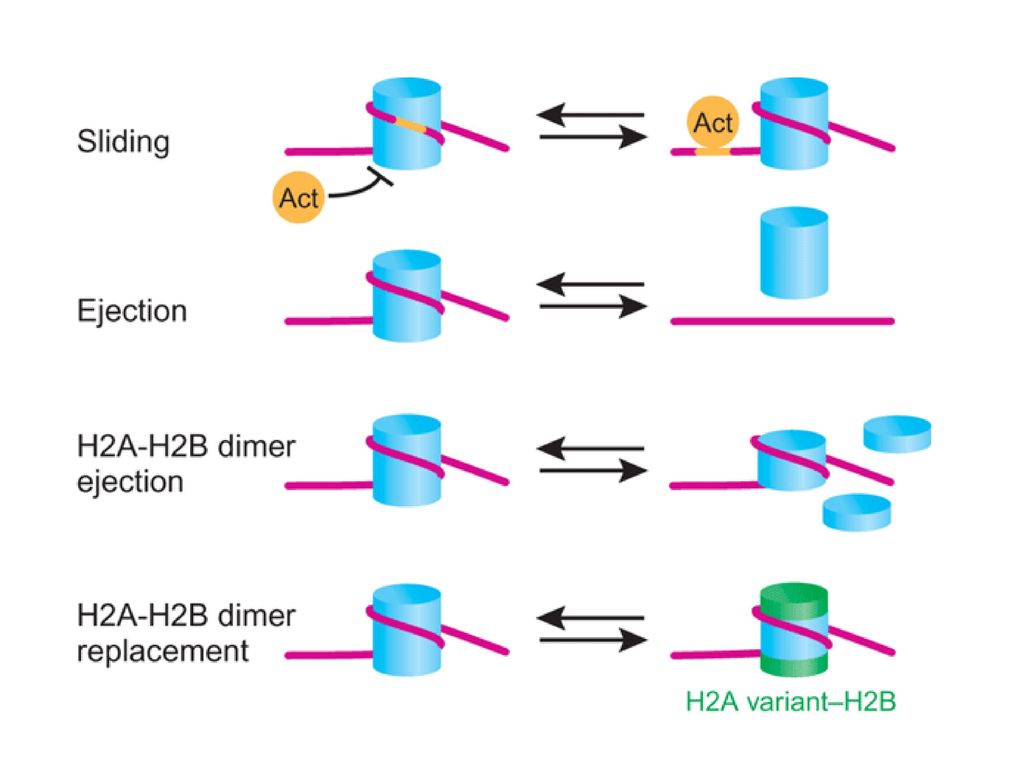
染色质重塑的模式 核小体滑动:核心组蛋白位置变化 重塑核小体: 核小体置换:核心组蛋白完全被取代 组蛋白替换:核心组蛋白成分改变

112
染色质重塑

113
染色质重塑复合物 SWI/SNF family (SWI2/SNF2) ISWI family Mi-2 family Ino80/SWR1
SWI (switching ) SNF (sucrose non-fermenting)
 SNF (sucrose non-fermenting)")
114
ATP-dependent Remodeling Complexes
CHD1 dMi-2 Cell 2002,108:

115
表观遗传调控 DNA修饰-甲基化 核心组蛋白修饰-组蛋白密码(Histone code)
染色质重塑(Chromatin Remodeling)ATP-依赖 非编码RNA (Non-coding RNA ,Regulatory RNA): Antisence RNA, siRNA , miRNA, Long non-coding RNA RNA编辑(RNA editing)
ATP-依赖. 非编码RNA (Non-coding RNA ,Regulatory RNA): Antisence RNA, siRNA , miRNA, Long non-coding RNA. RNA编辑(RNA editing)")
116
非编码RNA(Non-coding RNA)是指不编码蛋白质的RNA。其中包括rRNA,tRNA,snRNA,snoRNA 和microRNA 等多种已知功能的 RNA,还包括未知功能的RNA。
Anti-sence RNA miRNA siRNA Long non-coding RNA

117
Mol.Cell-special issue- Regulatory RNA

118
MicroRNAs(miRNAs)是一种小的内源性非编码RNA分子,大约由21-25个核苷酸组成。这些小的miRNA通常靶向一个或者多个mRNA,通过翻译水平的抑制或断裂靶标mRNAs而调节基因的表达。 最近的研究表miRNA参与各种各样的调节途径,包括发育、病毒防御、造血过程、器官形成、细胞增殖和凋亡、脂肪代谢等等。

119
成熟的miRNAs是由较长的初级转录物经过一系列核酸酶的剪切加工而产生的,随后组装进RNA诱导的沉默复合体,通过碱基互补配对的方式识别靶mRNA,并根据互补程度的不同指导沉默复合体降解靶mRNA或者阻遏靶mRNA的翻译。

122
siRNA(Small interfering RNA ):是一种小RNA分子(~21-25核苷酸),由Dicer(RNAase Ⅲ家族中对双链RNA具有特异性的酶)加工而成。SiRNA是siRISC的主要成员,激发与之互补的目标mRNA的沉默。 实验室常用技术!

123
2006年的诺贝尔生理学奖获得者

124
miRNA与 siRNA

125
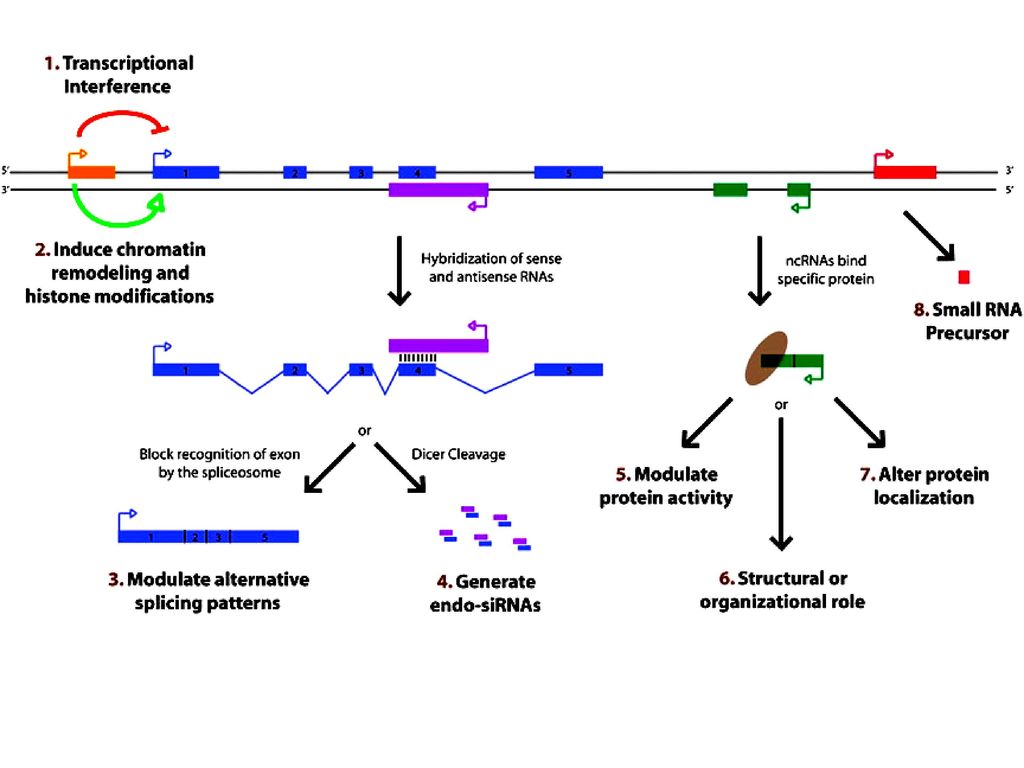
Long non-coding RNA 长链非编码RNA(Long non-coding RNA, lncRNA)是一类本身不编码蛋白、转录本长度超过200nt的长链非编码RNA分子,它可在多层面上(表观遗传调控、转录调控以及转录后调控等)调控基因的表达。 lncRNA最初被认为是RNA聚合酶II转录的副产物,是一种“噪音”,不具有生物学功能。然而,今年来的研究表面,lncRNA参与了X染色体沉默、染色体修饰和基因组修饰、转录激活、转录干扰、核内运输等过程,其调控作用正在被越来越多的人研究。

126
lncRNA的作用机制 编码蛋白的基因上游启动子区(橙色)转录,干扰下游基因(蓝色)的表达;
抑制RNA聚合酶II或者介导染色质重构以及组蛋白修饰,影响下游基因(蓝色)的表达; 与编码蛋白基因的转录本形成互补双链(紫色),干扰mRNA的剪切,形成不同的剪切形式; 与编码蛋白基因的转录本形成互补双链(紫色),在Dicer酶的作用下产生内源性siRNA;
的表达; 与编码蛋白基因的转录本形成互补双链(紫色),干扰mRNA的剪切,形成不同的剪切形式; 与编码蛋白基因的转录本形成互补双链(紫色),在Dicer酶的作用下产生内源性siRNA;")
127
与特定蛋白质结合,lncRNA转录本(绿色)可调节相应蛋白的活性;
作为结构组分与蛋白质形成核酸蛋白质复合体; 结合到特定蛋白质上,改变该蛋白质的细胞定位; 作为小分子RNA(如miRNA、piRNA)的前体分子。
的前体分子。")
129
Heart, Lung and Circulation. 2012;21:131–142
A MicroRNA Guide for Clinicians and Basic Scientists: Background and Experimental Techniques

130
ChR, chromatin remodelers; DNMTs, DNA methyltransferases;
HATs, histone acetyltransferases; HDACs, histone deacetylases; HMTs, histone methyltransferases; HDMs, histone demethylases; DDMs,DNA demethylases [unidentified in mammals to date]; and TFs, transcription factors [reflecting the genetic component of the epigenetic process] Epigenetics: A Landscape Takes Shape Aaron D. Goldberg, C. David Allis, and Emily Bernstein Cell 128, February 23, 2007

131
Human Epigenome Project
研究基因组水平上表观遗传修饰的科学称为表观基因组学(epigenomics) 。1999 年在欧洲成立了一个研究表观基因组的机构,即人类表观基因组协会( Human Epigenome Consortium , HEC ,http :/ / www. epigenome. org) 。该协会在2003 年10月正式宣布开始实施人类表观基因组计划( HumanEpigenome Proiect, HEP) 。
 。1999 年在欧洲成立了一个研究表观基因组的机构,即人类表观基因组协会( Human Epigenome Consortium , HEC ,http :/ / www. epigenome. org) 。该协会在2003 年10月正式宣布开始实施人类表观基因组计划( HumanEpigenome Proiect, HEP) 。")
132
Human Epigenome Project
The Human Epigenome Project aims to identify, catalogue and interpret genome-wide DNA methylation patterns of all human genes in all major tissues.

133
人类表观基因组计划是要绘制出不同组织类型和疾病状态下的人类基因组甲基化可变位点(methylation variable position ,MVP) 图谱。
这项计划可以进一步加深研究者对于人类基因组的认识,为探寻与人类发育和疾病相关的表观遗传变异提供蓝图。

134
Cell 155, September 26, 2013

135
Epigenetic drugs Nat Rev Drug Discov. 2013 Aug 30;12(9):661-2.
ALL, acute lymphoblastic leukaemia; AML, acute myeloid leukaemia; CRC, colorectal cancer; HCC, hepatocellular carcinoma; HDAC, histone deacetylase; MDS, myelodysplastic syndromes; NSCLC, non-small-cell lung cancer; RCC, renal cell carcinoma.

136
Epigenetic drugs by phase of clinical development and mechanism of action
Nat Rev Drug Discov Aug 30;12(9):661-2.
:")
137
转录因子结合到启动子区! 真核生物转录复合物

138
染色质水平上的调控—表观遗传调控

139
? 中心法则 基因表达调控: 转录水平 翻译水平 染色质重塑因子 组蛋白共价修饰 DNA甲基化 DNA 调节性RNA 组蛋白
Chromatin 染色质 DNA 组蛋白 改变染色质 状态 表观遗传调控 ? 中心法则 基因表达调控: 转录水平 翻译水平



染色体结构不同; ( 2 )原核生物具有正调控和负调控并重的特点,真核 生物目前已知的主要是正调控; ( 3 )原核生物的转录和翻译是相偶联的,真核生物的.>")
:核蛋白体组成成分 转移 RNA ( tRNA ):转运氨基酸 信使 RNA ( mRNA ):蛋白质合成模板 不均一核 RNA ( hnRNA ):成熟 mRNA 的前体 小核 RNA ( snRNA ):>")

>")
>")











 A.染色体 B.质体 C.线粒体 D.核糖体>")


