Download presentation
Presentation is loading. Please wait.

1
園藝學原理 Chapter 9 - 植物賀爾蒙 PLANT HORMONES
張育森 教授 國立臺灣大學 園藝暨景觀學系 【本著作除另有註明外,採取創用CC「姓名標示-非商業性-相同方式分享」臺灣3.0版授權釋出】

2
前言 植物的外觀及大小主要以表現型( phenotypes )來表示。表現型的形態差異主要受到基因型(genotype )及環境的交互作用所致。 植物基因透過轉錄(transcription )和轉譯(translation )來形成蛋白質的代碼,其coding涉及一些植物荷爾蒙的生物合成( biosynthesis )。 植物荷爾蒙可以刺激細胞、組織及器官的生長及分化。 植物荷爾蒙的生合成也會受到環境刺激(包括溫度、水、光、營養、害蟲、逆境和人為操作如修剪)的影響。 因此,植物表現型除了受到自然環境的變化及人為的操作外,其內在荷爾蒙的平衡也相當的重要。 植物生長調節劑(plant growth regulators)可促進植物產量、品質及增加採收後貯藏的壽命,為園藝商業生長上重要的調節方法。
和轉譯(translation )來形成蛋白質的代碼,其coding涉及一些植物荷爾蒙的生物合成( biosynthesis )。 植物荷爾蒙可以刺激細胞、組織及器官的生長及分化。 植物荷爾蒙的生合成也會受到環境刺激(包括溫度、水、光、營養、害蟲、逆境和人為操作如修剪)的影響。 因此,植物表現型除了受到自然環境的變化及人為的操作外,其內在荷爾蒙的平衡也相當的重要。 植物生長調節劑(plant growth regulators)可促進植物產量、品質及增加採收後貯藏的壽命,為園藝商業生長上重要的調節方法。")
3
TERMINOLOGY 專有名詞 植物荷爾蒙( Plant hormone) 1. 需在植物體內合成 2. 為有機化合物
3. 僅需低濃度即可調節植物生長與發育 4. 其荷爾蒙須由生合成部位移至作用位置使得以發揮作用,此為一種訊號傳遞 5.養分(如醣類)及維他命並不包括在內 植物生長物質(Plant growth substance) 為自然或人工合成的物質,僅需低濃度即可調節植物的生長。養分(如醣類)及維他命並不包括在植物生長物質內。其同義詞為植物生長調節劑( plant growth regulator,PGR )
及維他命並不包括在內. 植物生長物質(Plant growth substance) 為自然或人工合成的物質,僅需低濃度即可調節植物的生長。養分(如醣類)及維他命並不包括在植物生長物質內。其同義詞為植物生長調節劑( plant growth regulator,PGR )")
4
TERMINOLOGY 專有名詞 向頂運輸(Acropetal)
植物體內有機或無機物質由基部運移到頂端的現象,如細胞分裂素在根部合成,由根部向地上部運輸 向基運輸(Basipetal) 植物體內有機或無機物質由頂端運移到基部的現象,如生長素在莖頂合成,而往枝條基部和根部運輸 檢定法(Assay) 確定一種或多種的化合物存在、不存在或者定量的分析方法 生物檢定法(Bioassay) 利用活體材料(living material )進行檢測的方法,特定植物或部位對某些化合物特別敏感,所測得為-like compound
 植物體內有機或無機物質由頂端運移到基部的現象,如生長素在莖頂合成,而往枝條基部和根部運輸. 檢定法(Assay) 確定一種或多種的化合物存在、不存在或者定量的分析方法. 生物檢定法(Bioassay) 利用活體材料(living material )進行檢測的方法,特定植物或部位對某些化合物特別敏感,所測得為-like compound.")
5
TERMINOLOGY 專有名詞 內生性( Endogenous ) 源自植物內部發生的一些事件;例如一些化合物在植物內部被合成
外生性(Exogenous ) 與內生性相反,源自植物外部所引發的一些事件;例如藉由人工外施ㄧ些化合物而引發植物的反應 In vitro(生體外) 拉丁話意為「試管內」,即在人為環境控制下,將植物的培植體置於試管或燒杯中並供給冨含營養分的培養基,以促進植物的生長。 In vivo (生體內) 即在活的有機體(living organism )發生一些反應。如植物行光合作用(photosynthesis occurs in vivo)
 與內生性相反,源自植物外部所引發的一些事件;例如藉由人工外施ㄧ些化合物而引發植物的反應. In vitro(生體外) 拉丁話意為「試管內」,即在人為環境控制下,將植物的培植體置於試管或燒杯中並供給冨含營養分的培養基,以促進植物的生長。 In vivo (生體內) 即在活的有機體(living organism )發生一些反應。如植物行光合作用(photosynthesis occurs in vivo)")
6
TERMINOLOGY 專有名詞 低濃度(Low concentration) 指濃度低於10-6 M或1 ppm
1 mg = l0-3g = milligram, or thousandth of a gram 1μg = l0-6g = microgram, or millionth of a gram 1 ng = l0-9g = nanogram, or billionth of a gram 1 pg = l0-12g = picogram, or trillionth of a gram 1 fg = l0-15g = femtogram, or quadrillionth of a gram 1 ppm = 10-6 = 1 mg‧liter-1 (milligrams per liter of water) = 1μg‧ml-1; 1% = 10,000 ppm.)
 = 1μg‧ml-1; 1% = 10,000 ppm.)")
7
TERMINOLOGY 專有名詞 主要植物荷爾蒙(Major plant hormone)
存在於所有高等植物中的一種或一類植物荷爾蒙,包括auxins, gibberellins, cytokinins, brassinosteroids, ethylene, abscisic acid, and jasmonates. 藥理學反應( Pharmacological response ) 非在自然環境下植物發生的反應,例如施用藥物及殺草劑過量對植物所產生的反應 生理學反應( Physiological response) 在自然環境下植物發生的反應 向性( Tropism ) 如向光性((phototropism )、向地性(geotropism)及背地性(gravitropism)
 非在自然環境下植物發生的反應,例如施用藥物及殺草劑過量對植物所產生的反應. 生理學反應( Physiological response) 在自然環境下植物發生的反應. 向性( Tropism ) 如向光性((phototropism )、向地性(geotropism)及背地性(gravitropism)")
8
主要的植物荷爾蒙 ( MAJOR PLANT HORMONES )
促進生長的荷爾蒙( Growth-promoting hormones ) 1. Auxins:促進細胞伸長、不定根形成 2. Gibberellins:促進細胞伸長及種子發芽 3. Cytokinins:促進細胞分裂及枝條分枝 4. Brassinosteroids :促進莖及其他器官的細胞伸長 創傷和逆境的荷爾蒙(Wounding and stress hormones) 1. Ethylene:為氣態,可促進莖及根的捲曲(swelling)、為老化荷爾蒙(aging hormone),可促進果實後熟及老化 2. Abscisic acid:為逆境荷爾蒙(stress hormone),在逆境下(如乾旱)其內生ABA會大量生成,此為植物抵抗逆境的機制 3. Jasmonates:在植物遭受創傷或病害侵襲時,茉莉酸會大量生成
 1. Auxins:促進細胞伸長、不定根形成. 2. Gibberellins:促進細胞伸長及種子發芽. 3. Cytokinins:促進細胞分裂及枝條分枝. 4. Brassinosteroids :促進莖及其他器官的細胞伸長. 創傷和逆境的荷爾蒙(Wounding and stress hormones) 1. Ethylene:為氣態,可促進莖及根的捲曲(swelling)、為老化荷爾蒙(aging hormone),可促進果實後熟及老化. 2. Abscisic acid:為逆境荷爾蒙(stress hormone),在逆境下(如乾旱)其內生ABA會大量生成,此為植物抵抗逆境的機制. 3. Jasmonates:在植物遭受創傷或病害侵襲時,茉莉酸會大量生成.")
9
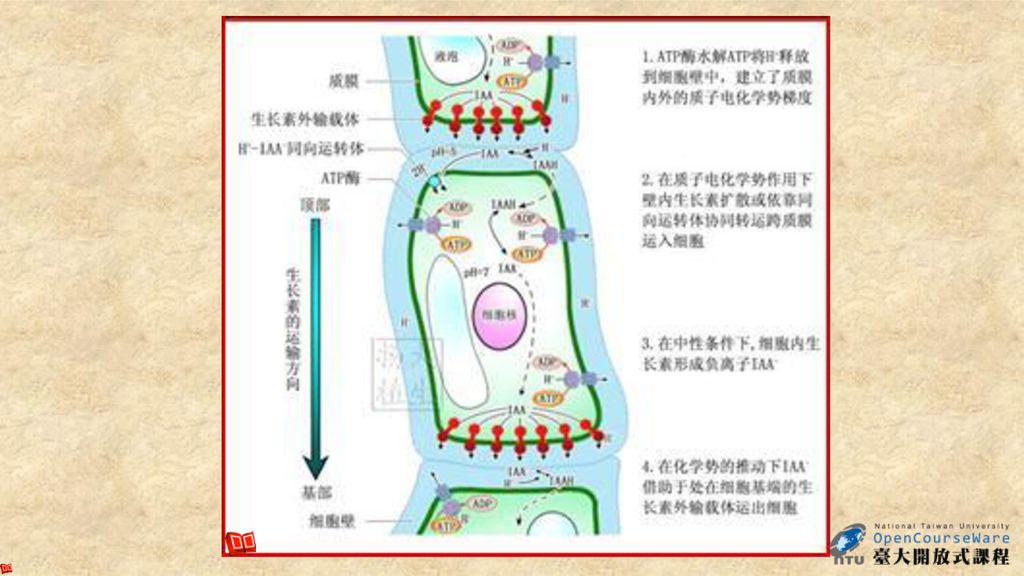
促進生長的荷爾蒙: 1.生長素( Auxin ) 生長素是最早被發現的植物激素,1880年達爾文(Darwin)父子利用胚芽鞘進行向光性實驗,發現在光線照射下,胚芽鞘向光彎曲。之後被證實在胚芽鞘頂端(coleoptile tip )有吲哚乙酸(indole-3-acetic acid, IAA)的存在。 IAA可由胚芽鞘頂端往基部運移,促進莖的伸長。生長素在植物體內的運輸具有極性,即生長素只能從植物的形態學上端向下端運輸,而不能向相反的方向運輸,這稱為生長素的極性運輸(polar transport)。其它植物激素則無此特點。
父子利用胚芽鞘進行向光性實驗,發現在光線照射下,胚芽鞘向光彎曲。之後被證實在胚芽鞘頂端(coleoptile tip )有吲哚乙酸(indole-3-acetic acid, IAA)的存在。 IAA可由胚芽鞘頂端往基部運移,促進莖的伸長。生長素在植物體內的運輸具有極性,即生長素只能從植物的形態學上端向下端運輸,而不能向相反的方向運輸,這稱為生長素的極性運輸(polar transport)。其它植物激素則無此特點。")
10
植物的向光性與IAA的分佈有關 室內植物往光源的方向彎曲

11
19.1 Summary of early experiments in auxin research (Part 1)
pp4e-fig jpg Plant physiology, Lincoln Taiz, Eduardo Zeiger, Sunderland, MA : Sinauer Associates, c2010, 5th edition, p. 547

12
19.1 Summary of early experiments in auxin research (Part 2)
Plant physiology, Lincoln Taiz, Eduardo Zeiger, Sunderland, MA : Sinauer Associates, c2010, 5th edition, p. 547

14
Examples of Auxins -IAA
IAA為天然的auxin類最具代表性的一種,於1934年被鑑定出來。除IAA外,在1982發現另一種天然的auxin—苯乙酸(phenylactic acid, PAA) ,且其在大麥、番茄、煙草、向日葵及玉米等植物中之含量比IAA還高,但其活性低於IAA。
 ,且其在大麥、番茄、煙草、向日葵及玉米等植物中之含量比IAA還高,但其活性低於IAA。")
15
(過量的auxin會有殺草劑效應) IAA若經過藍光或UV-B照射,極易降解,故不適於商業生產。若直接噴施於植株上則在兩小時即會有光降解現象(photodestruction )。 此外IAA oxidase和 peroxidases 也會使IAA崩解。 因此需人工合成一些穩定的auxin類藥劑,如indole-3-butyric acid (IBA); naphthalene-acetic acid (NAA); 和一些殺草劑2,4,-dichlorophenoxy acetic acid (2,4-D)和 4-chloro-2-methylphenoxyacetic acid (MCPA)
; naphthalene-acetic acid (NAA); 和一些殺草劑2,4,-dichlorophenoxy acetic acid (2,4-D)和 4-chloro-2-methylphenoxyacetic acid (MCPA)")
16
Auxin Biosynthesis IAA及PAA生物合成的前趨物質分別為:色氨酸(tryptophan)及苯丙氨酸(phenylalanine )。 Auxins主要合成部位:植物的莖端分生組織,特別是正在擴展的幼嫩葉片(<1/5 fully expanded )、幼嫩芽鞘頂端、根冠、發育中的種子及果實
、幼嫩芽鞘頂端、根冠、發育中的種子及果實.")
17
合成前体 直接前体 吲哚乙醛 色氨酸 色氨酸转氨E 吲哚丙酮酸 吲哚丙酮酸脱羧E 吲哚乙醛脱氢E 吲哚乙酸 吲哚丙酮酸途径 色氨酸脱羧E
色胺 胺氧化E 色胺途径 吲哚乙醇 吲哚乙醇氧化E 吲哚乙醇途径

18
IAA biosynthetic pathways in plants and bacteria
pp4e-fig jpg

19
蘋果或葡萄若缺乏種子,就缺乏 auxin等荷爾蒙的生成,其果實發育異常。外施auxin可促使子房及其周遭組織膨大而獲得無籽果實。
較大果實含 較多種子

20
Auxin Activity(1) 1. Auxin可促進莖的伸長,其細胞的伸長也與趨光性有相關。當植物置於缺乏光線的環境下,其內生IAA並不會photodestruction 。顯示出在缺乏光線的情況下,其內生auxin含量高,促使白化苗(etiolation ) 節間伸長。 2. Auxin並參與細胞的分裂,且auxin與cytokinin的交互作用也很重要。 3. Auxin參與細胞及組織的分化(differentiation )與再分化(dedifferentiation ,即returning to the meristematic state),此可證實在組織培養中(如大豆或樺樹),auxin可促進callus的生長並影響維管束形成層及木質部的再分化。
與再分化(dedifferentiation ,即returning to the meristematic state),此可證實在組織培養中(如大豆或樺樹),auxin可促進callus的生長並影響維管束形成層及木質部的再分化。")
21
Auxin Activity(2) 4. Auxin促進頂芽優勢( apical dominance )
(即頂芽對側芽生長的抑制) (Figure 11-6). Auxin在頂端產生,完全抑制側芽的生長,造成單一主幹
 (Figure 11-6). Auxin在頂端產生,完全抑制側芽的生長,造成單一主幹.")
22
Auxin Activity(3) Auxin合成主要在頂端及幼嫩的葉片,若摘去頂芽,其頂端內生auxin含量降低,促進側枝數增加 (Figure 11-7) 修剪去頂,造成多分枝的灌叢苗木
 Auxin合成主要在頂端及幼嫩的葉片,若摘去頂芽,其頂端內生auxin含量降低,促進側枝數增加 (Figure 11-7) 修剪去頂,造成多分枝的灌叢苗木")
23
Auxin Activity(4) 5.Auxin可藉由處理的時間、濃度、植物 生長的stage來促進或抑制花朵、葉片 和果實的離層。
雖然果樹會自然疏除過多的果實,如 蘋果和梨在夏天時會“June drop” , 但仍不足經濟生產,因此仍需人工疏 果處理。在盛花後3-30天施用較高 NAA可促使落果。 但施用NAA亦可減緩採收前的落果。 故利用auxin促進或抑制果實的脫落的 特性,對於增進果實產量、品質有很 大的影響。 太多果實會使果實變小,可使用 NAA或配合加保利進行疏果

24
Auxin Activity(5) 6. Auxin促進胚及器官(尤其是根)的形成,例如在胡蘿蔔培養基中添加2,4-D,可誘導體胚的形成
施用IBA粉劑,使聖誕紅根系發達 6. Auxin促進胚及器官(尤其是根)的形成,例如在胡蘿蔔培養基中添加2,4-D,可誘導體胚的形成 此外聖誕紅的插穗,施用IBA粉劑,可使根系更發達(Figure 11-9)
的形成,例如在胡蘿蔔培養基中添加2,4-D,可誘導體胚的形成. 此外聖誕紅的插穗,施用IBA粉劑,可使根系更發達(Figure 11-9)")
25
Auxin Activity(6) 7. 促進插穗不定根的形成普遍應用在植物繁殖上
生長素可以有效促進插穗不定根的形成,這主要是剌激了插穗基部切口處細胞的分裂與分化,誘導了根原基的形成。

26
Auxin Sensitivity Auxin效應與濃度有很大關係,而顯現出促進或抑制的效果。
美國白臘樹在試管內加入過量 Auxin造成短粗的根(右) Auxin效應與濃度有很大關係,而顯現出促進或抑制的效果。 植物根部對auxin敏感,低濃度auxin可促進根部伸長,較高一點濃度則顯現出抑制的效果而呈現較短粗的根(Figure 11-11) 高濃度的2,4-D為良好的殺草劑,但過量會對生長造成傷害。 芽及莖對auxin敏感低於根部,高濃度auxin可促進芽及莖的生長,但濃度太高則會呈現抑制生長。
 Auxin效應與濃度有很大關係,而顯現出促進或抑制的效果。 植物根部對auxin敏感,低濃度auxin可促進根部伸長,較高一點濃度則顯現出抑制的效果而呈現較短粗的根(Figure 11-11) 高濃度的2,4-D為良好的殺草劑,但過量會對生長造成傷害。 芽及莖對auxin敏感低於根部,高濃度auxin可促進芽及莖的生長,但濃度太高則會呈現抑制生長。")
27
10-4 根 莖 芽 10-10 10-8 抑制 促進 不同營養器官對不同濃度IAA的反應
生長素濃度(mol/L) 不同營養器官對不同濃度IAA的反應 抑制 促進 10-4 根 莖 芽 10-10 10-8
 不同營養器官對不同濃度IAA的反應. 抑制 促進 根. 莖. 芽")
28
2.激勃素( Gibberellins ,GA) 激勃素為日本人研究水稻馬鹿兒苗病(silly seedling, Bakanae Disease)時發現的,患病的水稻植株會呈現徒長並減少40%的產量。 此疾病主要由一種真菌所引起,無性階段的稱為Fusarium moniliforme (asexual form) 、有性階段的稱為 Gibberella fujikuori (sexual form) 。 1935年日本科學家藪田從誘發水稻馬鹿兒苗病的真菌中分離得到了能促進生長的非結晶固體,並稱之為激勃素。 目前已發現136種激勃素,有些激勃素只在真菌中才有,有些則只存在高等植物中,有些激勃素則是真菌及植物中皆有。
 、有性階段的稱為 Gibberella fujikuori (sexual form) 。 1935年日本科學家藪田從誘發水稻馬鹿兒苗病的真菌中分離得到了能促進生長的非結晶固體,並稱之為激勃素。 目前已發現136種激勃素,有些激勃素只在真菌中才有,有些則只存在高等植物中,有些激勃素則是真菌及植物中皆有。")
29
Gibberellin Biosynthesis and Commercial Availability
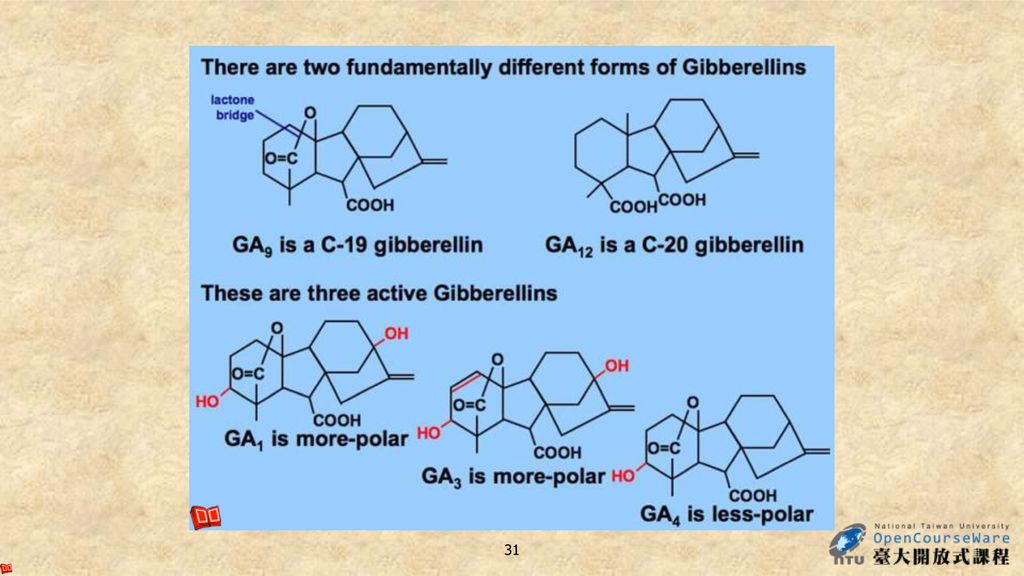
激勃素的種類雖然很多,但都是以激勃烷(gibberellane)為骨架的衍生物。激勃素是一種雙帖,由四個異戊二烯單位組成,有四個環。 A、B、C、D四個環對激勃素的活性都是必要的,環上各基團的種種變化就形成了各種不同的激勃素,但所有有活性的激勃素的第7個碳均為羧基。 A B C D
為骨架的衍生物。激勃素是一種雙帖,由四個異戊二烯單位組成,有四個環。 A、B、C、D四個環對激勃素的活性都是必要的,環上各基團的種種變化就形成了各種不同的激勃素,但所有有活性的激勃素的第7個碳均為羧基。 A. B. C. D.")
30
Gibberellin Biosynthesis and Commercial Availability
根據激勃素分子中碳原子的不同,可分為20-C和19-C。 20-C含有激勃烷中所有的20個碳原子(如GA15、GA24、GA19、GA25、GA17等), 而19-C只含有19個碳原子,第20位的碳原子已丟失(如GA1、GA3、GA4、GA9、GA20等)。19-C激勃素在數量上多於20-C激勃素,且活性也較高。
, 而19-C只含有19個碳原子,第20位的碳原子已丟失(如GA1、GA3、GA4、GA9、GA20等)。19-C激勃素在數量上多於20-C激勃素,且活性也較高。")
32
甲瓦龍酸(甲羥戊酸, mevalonic acid )
生物合成前體︰ 甲瓦龍酸(甲羥戊酸, mevalonic acid ) 植物體內合成部位︰ 頂端幼嫩部分,如根尖 和莖尖, 生長中的種子和果實, 其中正在發育的種 子是GA的豐富來源。 目前商業上販售的人工合成的GA有GA3、GA4+7或GA4+7結合BA的藥劑 GA在植物體內形成過程與矮化劑阻礙位置(太田 1985) CPD: copalyl pyrophosphate, MVA: mevalonic acid. GGPP: geranylgeranyl pyrophosphate
 植物體內合成部位︰ 頂端幼嫩部分,如根尖 和莖尖, 生長中的種子和果實, 其中正在發育的種 子是GA的豐富來源。 目前商業上販售的人工合成的GA有GA3、GA4+7或GA4+7結合BA的藥劑. GA在植物體內形成過程與矮化劑阻礙位置(太田 1985) CPD: copalyl pyrophosphate, MVA: mevalonic acid. GGPP: geranylgeranyl pyrophosphate.")
33
甲瓦龍酸(MVA) 異戊烯基焦磷酸(IPP) 法尼基焦磷酸(FPP) 蟒牛兒蟒牛兒焦磷酸(GGPP) 内-貝殼杉烯 貝殼杉烯酸 GA12-醛
GA GAS

34
Gibberellin Activities(1)
1. GA可促進細胞的延長 (1)GA可促進細胞的延長,如叢生的大豆及碗豆(bush beans or bush peas )施用GA後會形成攀爬型的大豆及碗豆( climbing pole beans or pole peas) (2)甘藍菜施用GA其節間伸長可使植株長至2公尺以上 (3)甘蔗施用GA可促進其節間伸長並提升產量 (4) 葡萄施用GA可促進其花梗伸長並減少其叢生 矮生豌豆缺乏GA(左) 施用GA促節間伸長(右)
GA可促進細胞的延長,如叢生的大豆及碗豆(bush beans or bush peas )施用GA後會形成攀爬型的大豆及碗豆( climbing pole beans or pole peas) (2)甘藍菜施用GA其節間伸長可使植株長至2公尺以上. (3)甘蔗施用GA可促進其節間伸長並提升產量. (4) 葡萄施用GA可促進其花梗伸長並減少其叢生. 矮生豌豆缺乏GA(左) 施用GA促節間伸長(右)")
35
Gibberellin Activities(2)
2. GA可促進細胞分裂 (1)白楊樹施用GA3可促進維管束形成層的細胞分裂,誘使新的韌皮部分化篩管。若同時施用GA3及IAA則可促進維管束形成層的細胞分裂並誘使韌皮部及木質部的分化。 (2)Auxin及GA的相互作用可促進植物的生長與發育 3. GA促進種子發芽 GA強化IAA的作用
白楊樹施用GA3可促進維管束形成層的細胞分裂,誘使新的韌皮部分化篩管。若同時施用GA3及IAA則可促進維管束形成層的細胞分裂並誘使韌皮部及木質部的分化。 (2)Auxin及GA的相互作用可促進植物的生長與發育. 3. GA促進種子發芽. GA強化IAA的作用.")
36
20.34 IAA promotes and is required for GA1 biosynthesis in subtending internodes of pea
IAA強化GA的作用

37
內生GA促進大麥種子發芽過程 乾燥種子浸水後,啟動內在生化機制 GA在胚處形成,此可證明大麥種子富含大量的GA
GA移至糊粉層(aleurone layer),此糊粉層有3-4層細胞厚,圍繞著胚乳 在糊粉層中,GA會誘導α-amylase和其他水解酵素的產生 α-amylase和其他水解酵素會移至胚乳,把澱粉水解成醣以進行呼吸作用 之後這些醣會移至胚提供能量給種子做為生長之用。 當種子發芽過程到此高潮時,會從種皮突出胚根,種子即可進行之後的生長及發育
,此糊粉層有3-4層細胞厚,圍繞著胚乳. 在糊粉層中,GA會誘導α-amylase和其他水解酵素的產生. α-amylase和其他水解酵素會移至胚乳,把澱粉水解成醣以進行呼吸作用. 之後這些醣會移至胚提供能量給種子做為生長之用。 當種子發芽過程到此高潮時,會從種皮突出胚根,種子即可進行之後的生長及發育.")
38
Gibberellin Activities(3)
4. GA可打破種子休眠 (1)對於需光和需低溫才能萌發的種子,如萵苣、煙草、紫蘇、李和蘋果等的種子,GA可代替光照和低溫打破休眠,這是因為GA可誘導α-澱粉酶、蛋白酶和其它水解酶的合成,催化種子內貯藏物質的降解,以供胚的生長發育所需。 (2)在啤酒製造業中,用GA處理萌動而未發芽的大麥種子,可誘導α-澱粉酶的產生,加速釀造時的糖化過程,並降低萌芽的呼吸消耗,從而降低成本。
對於需光和需低溫才能萌發的種子,如萵苣、煙草、紫蘇、李和蘋果等的種子,GA可代替光照和低溫打破休眠,這是因為GA可誘導α-澱粉酶、蛋白酶和其它水解酶的合成,催化種子內貯藏物質的降解,以供胚的生長發育所需。 (2)在啤酒製造業中,用GA處理萌動而未發芽的大麥種子,可誘導α-澱粉酶的產生,加速釀造時的糖化過程,並降低萌芽的呼吸消耗,從而降低成本。")
39
Gibberellin Activities(4)
5. 促進雄花分化 對於雌雄異花同株的植物,用GA處理后,雄花的比例增加;對於雌雄異株植物的雌株,如用GA處理,也會開出雄花。GA在這方面的效應與生長素和乙烯相反 6. 其它生理效應 (1)GA還可加強IAA對養分的動員效應,促進某些植物著果和單性結實、延緩葉片衰老等。 (2)此外,GA也可促進細胞的分裂和分化,GA促進細胞分裂是由於縮短了G1期和S期。但GA對不定根的形成卻起抑制作用,這與生長素又有所不同。
GA還可加強IAA對養分的動員效應,促進某些植物著果和單性結實、延緩葉片衰老等。 (2)此外,GA也可促進細胞的分裂和分化,GA促進細胞分裂是由於縮短了G1期和S期。但GA對不定根的形成卻起抑制作用,這與生長素又有所不同。")
40
Gibberellin Sensitivity
不存在超最適濃度的抑制作用,即使GA濃度很高,仍可表現出最大的促進效應,這與生長素促進植物生長具有最適濃度的情況顯著不同。

41
3.細胞分裂素( Cytokinins ) 斯庫格(F. Skoog)和崔 (1948)等在尋找促進組織培養中細胞分裂的物質時,發現生長素存在時腺嘌呤具有促進細胞分裂的活性。 1954年,雅布隆斯基(J. R. Jablonski)和斯庫格發現煙草髓組織在只含有生長素的培養基中細胞不分裂而只長大,如將髓組織與維管束接觸,則細胞分裂。 1955年米勒(C. O. Miller)和斯庫格等偶然將存放了4年的鯡魚精細胞DNA加入到煙草髓組織的培養基中,發現也能誘導細胞的分裂,且其效果優於腺嘌呤,但用新提取的DNA卻無促進細胞分裂的活性,如將其在pH〈4的條件下進行高壓滅菌處理,則又可表現出促進細胞分裂的活性。他們分離出了這種活性物質,並命名為激動素(kinetin, KT)。 1956年,米勒等從高壓滅菌處理的鯡魚精細胞DNA分解產物中純化出了激動素結晶,並鑑定出其化學架構為6- 喃氨基嘌呤(N6-furfurylaminopurine),分子式為C10H9N5O,分子量為215.2,接著又人工合成了這種物質。
和斯庫格發現煙草髓組織在只含有生長素的培養基中細胞不分裂而只長大,如將髓組織與維管束接觸,則細胞分裂。 1955年米勒(C. O. Miller)和斯庫格等偶然將存放了4年的鯡魚精細胞DNA加入到煙草髓組織的培養基中,發現也能誘導細胞的分裂,且其效果優於腺嘌呤,但用新提取的DNA卻無促進細胞分裂的活性,如將其在pH〈4的條件下進行高壓滅菌處理,則又可表現出促進細胞分裂的活性。他們分離出了這種活性物質,並命名為激動素(kinetin, KT)。 1956年,米勒等從高壓滅菌處理的鯡魚精細胞DNA分解產物中純化出了激動素結晶,並鑑定出其化學架構為6- 喃氨基嘌呤(N6-furfurylaminopurine),分子式為C10H9N5O,分子量為215.2,接著又人工合成了這種物質。")
42
1964年確定其化學架構為6-(4-羥基-3-甲基 -反式-2-丁烯基氨 基)嘌呤,分子式為C10H13N5O,分子量為129.7。
1963年,萊撒姆(D.S.Letham)從未成熟的玉米籽粒中分離出 了一種類似於激動素的細胞分裂促進物質,命名為玉米素 (zeatin, Z,ZT), 1964年確定其化學架構為6-(4-羥基-3-甲基 -反式-2-丁烯基氨 基)嘌呤,分子式為C10H13N5O,分子量為129.7。 1965年斯庫格等提議 將來源於植物的、其 生理活性類似於激動 素的化合物統稱為細 胞分裂素(cytokinin, CTK,CK) 目前在高等植物中已 至少鑑定出了30多種 細胞分裂素。 常見的天然細胞分裂素和人工合成的細胞分裂素的結構式
從未成熟的玉米籽粒中分離出 了一種類似於激動素的細胞分裂促進物質,命名為玉米素 (zeatin, Z,ZT), 1964年確定其化學架構為6-(4-羥基-3-甲基 -反式-2-丁烯基氨 基)嘌呤,分子式為C10H13N5O,分子量為129.7。 1965年斯庫格等提議 將來源於植物的、其 生理活性類似於激動 素的化合物統稱為細 胞分裂素(cytokinin, CTK,CK) 目前在高等植物中已 至少鑑定出了30多種 細胞分裂素。 常見的天然細胞分裂素和人工合成的細胞分裂素的結構式.")
43
Cytokinin Biosynthesis and Commercial Availability
主要分佈位置為可進行細胞分裂的部位,如莖尖、根尖、未成熟的種子、萌發的種子和生長中的果實等。 一般細胞分裂素在根尖合成經木質部運到地上部。

44
Cytokinin生物合成 合成部位:根尖、生長中的種子和果實,在細胞内的合成部位是微粒體。 CTK有兩類:游離的和結合在tRNA上的
游離的CTKS来源: tRNA降解 重新合成:前體:甲瓦龍酸 CTK有兩類:游離的和結合在tRNA上的

45
甲瓦龍酸 異戊烯基焦磷酸 5’-AMP 異戊烯基腺苷-5’-磷酸鹽 異戊烯基腺嘌呤 玉米素 (MVA) (IPP) (〔9R-5’P〕iP)
(Zeatin)
")
46
Cytokinin Activities(1)
1. 促進細胞分裂 (1)細胞分裂素的主要生理功能就是促進細胞的分裂。生長素、激勃素和細胞分裂素都有促進細胞分裂的效應,但它們各自所起的作用不同。 (2)生長素只促進核的分裂(因促進了DNA的合成),而與細胞質的分裂無關。 (3)而細胞分裂素主要是對細胞質的分裂起作用,所以,細胞分裂素促進細胞分裂的效應只有在生長素存在的前提下才能表現出來。
細胞分裂素的主要生理功能就是促進細胞的分裂。生長素、激勃素和細胞分裂素都有促進細胞分裂的效應,但它們各自所起的作用不同。 (2)生長素只促進核的分裂(因促進了DNA的合成),而與細胞質的分裂無關。 (3)而細胞分裂素主要是對細胞質的分裂起作用,所以,細胞分裂素促進細胞分裂的效應只有在生長素存在的前提下才能表現出來。")
47
Cytokinin Activities(2)
2. 促進細胞擴大(cell enlargement)︰ 此指細胞全方位增大,而非Auxin和GA所促進的細胞伸長(cell elongation) ,且cytokinin常抑制細胞伸長。 3.因此 cytokinin可促進葉片擴展︰ 【佐證】彩葉草及印度橡膠的葉片可誘使其生成不定根, 但無法形成不定芽。 然其葉片擴展速度會比正常植株的葉片快,此可能與cytokinin的作用有關。(圖11-16)
︰ 此指細胞全方位增大,而非Auxin和GA所促進的細胞伸長(cell elongation) ,且cytokinin常抑制細胞伸長。 3.因此 cytokinin可促進葉片擴展︰ 【佐證】彩葉草及印度橡膠的葉片可誘使其生成不定根, 但無法形成不定芽。 然其葉片擴展速度會比正常植株的葉片快,此可能與cytokinin的作用有關。(圖11-16)")
48
Cytokinin Activities(3)
4.促進芽的分化 1957年斯庫格和米勒在進行煙草的組織培養時發現,細胞分裂素(激動素)和生長素的相互作用控制著癒傷組織根、芽的形成。 (1)當培養基中CTK/IAA的比值高時,癒傷組織形成芽; (2)當CTK/IAA的比值低時,癒傷組織形成根; (3)如二者的濃度相等,則癒傷組織保持生長而不分化;所以,透過調整二者的比值,可誘導癒傷組織形成完整的植株。 5. 打破種子及芽的休眠 6. 促進氣孔打開
和生長素的相互作用控制著癒傷組織根、芽的形成。 (1)當培養基中CTK/IAA的比值高時,癒傷組織形成芽; (2)當CTK/IAA的比值低時,癒傷組織形成根; (3)如二者的濃度相等,則癒傷組織保持生長而不分化;所以,透過調整二者的比值,可誘導癒傷組織形成完整的植株。 5. 打破種子及芽的休眠. 6. 促進氣孔打開.")
49
Cytokinin Activities(4)
(左)培養基中〔CTK〕/〔IAA〕的比值高時,癒傷組織形成芽; (右)當〔CTK〕/〔IAA〕的比值低時,癒傷組織形成根;
培養基中〔CTK〕/〔IAA〕的比值高時,癒傷組織形成芽; (右)當〔CTK〕/〔IAA〕的比值低時,癒傷組織形成根;")
50
Cytokinin Activities(5)
7. 延緩老化 (1)如在離體葉片上局部突以細胞分裂素則在葉片其餘部位變黃衰老時,塗抹細胞分裂素的部位仍保持鮮綠。 這不僅說明了細胞分裂素有延緩葉片衰老的作用,而且說明了細胞分裂素在一般組織中是不易移動的。 (2)當把細胞分裂素基因clone出來ipt,將此基因轉入煙草中,其含有高量的細胞分裂素故可比對照組(未轉入ipt gene)具較深綠的葉片,產量也會增加。
如在離體葉片上局部突以細胞分裂素則在葉片其餘部位變黃衰老時,塗抹細胞分裂素的部位仍保持鮮綠。 這不僅說明了細胞分裂素有延緩葉片衰老的作用,而且說明了細胞分裂素在一般組織中是不易移動的。 (2)當把細胞分裂素基因clone出來ipt,將此基因轉入煙草中,其含有高量的細胞分裂素故可比對照組(未轉入ipt gene)具較深綠的葉片,產量也會增加。")
51
8. 植物受到感染時其細胞分裂也會增加 (1)豆科植物被根瘤菌(Rhizobium or Bradyrhizobium )感染形成根瘤,其細胞分裂增加進行固氮作用。 (2)被農桿菌(Agrobacterium tumefaciens)感染後,其基部會形成虫癭,此疾病稱為crown gall (Figure 11-18) ,若將ipt gene轉植入寄主植物,過量表現細胞分裂素,則會使腫瘤產生不定芽 (3)藉由昆蟲、螨類、寄生植物或基因突變所造成的疾病稱為簇葉病( Witches’ brooms),此並會造成許多分枝且節間很短,與細胞分裂素有關(Figure ) 。
藉由昆蟲、螨類、寄生植物或基因突變所造成的疾病稱為簇葉病( Witches’ brooms),此並會造成許多分枝且節間很短,與細胞分裂素有關(Figure ) 。")
52
Tumor induction by Agrobacterium tumefaciens
Plant physiology, Lincoln Taiz, Eduardo Zeiger, Sunderland, MA : Sinauer Associates, c2010, 5th edition, p. 626

53
Cytokinin Activities(6)
9. 促進側芽發育,消除頂端優勢 CTK能解除由生長素所引起的頂端優勢,促進側芽生長發育。 pp4e-fig jpg

54
補充:促進側芽發育,消除頂端優勢 CTK能解除由生長素所引起的頂端優勢,促進側芽生長發育。 香脂冷杉上的眾生枝 轉ipt基因的菸草
41*2

55
4.油菜素類固醇( Brassinosteroids )
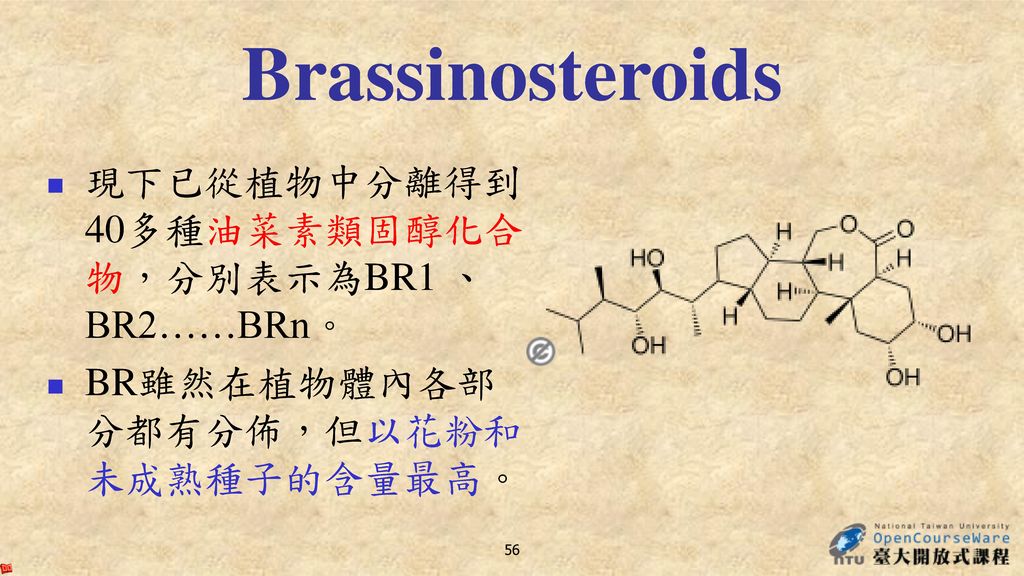
1970年,美國的米契爾(Mitchell)等報導在油菜的花粉中發現了一種新的生長物質,它能引起菜豆幼苗節間伸長、彎曲、裂開等異常生長回應,並將其命名為油菜素(brassin)。 1979年,格羅夫(Grove)等從227 kg油菜花粉中提取得到10 mg的高活性結晶物,因為它是甾醇內酯化合物,故將其命名為油菜素內酯(brassinolide, BR1)。 此後油菜素內酯及多種架構相似的化合物紛紛從多種植物中被分離鑑定,這些以甾醇為基本架構的具有生物活性的天然產物統稱為油菜素類固醇(brassinosteroids, BR,BRs),BR在植物體內含量極少,但生理活性很強。 目前,BR以及多種類似化合物已被人工合成,用於生理生化及田間試驗
等報導在油菜的花粉中發現了一種新的生長物質,它能引起菜豆幼苗節間伸長、彎曲、裂開等異常生長回應,並將其命名為油菜素(brassin)。 1979年,格羅夫(Grove)等從227 kg油菜花粉中提取得到10 mg的高活性結晶物,因為它是甾醇內酯化合物,故將其命名為油菜素內酯(brassinolide, BR1)。 此後油菜素內酯及多種架構相似的化合物紛紛從多種植物中被分離鑑定,這些以甾醇為基本架構的具有生物活性的天然產物統稱為油菜素類固醇(brassinosteroids, BR,BRs),BR在植物體內含量極少,但生理活性很強。 目前,BR以及多種類似化合物已被人工合成,用於生理生化及田間試驗.")
56
Brassinosteroids 現下已從植物中分離得到40多種油菜素類固醇化合物,分別表示為BR1 、BR2……BRn。
57
BR Biosynthesis BR生合成亦從mevalonic acid 而來,與GA同,其與CTK及ABA皆有五個碳(isoprene)具有相似的生合成。 Campesterol (菜油甾醇)為mevalonate 的衍生物,亦為castasterone和brassinolide的前驅物質。
具有相似的生合成。 Campesterol (菜油甾醇)為mevalonate 的衍生物,亦為castasterone和brassinolide的前驅物質。")
58
Isopentenyl-pyrophosphate
Acetyl-CoA * 3 乙醯輔酶A Mevalonic acid 甲瓦龍酸 Isopentenyl-pyrophosphate 異戊烯基-焦磷酸 Dimethylalleyl pyrophosphate 二甲基烯丙基焦磷酸 呼吸作用中斷的中間產物 很有效率的萜前驅物 同分異構物 (Modified from 徐和廖, 2006) Fig. 9.甲瓦龍酸路徑。 IPP DMAPP MVA
 Fig. 9.甲瓦龍酸路徑。 IPP. DMAPP. MVA.")
59
Geranyl pyrophosphate(C10)
IPP DMAPP 異戊烯基-焦磷酸 二甲基烯丙基焦磷酸 Cytokinins Geranyl pyrophosphate(C10) 牻牛兒基焦磷酸 2 Farnesyl pyrophosphate(C15) 法尼基焦磷酸 Spualene(C30) 鯊烯 Cycloartenol 環木鳳梨烯醇 Gibberellins Campesterol 菜油甾醇 Geranylgeranyl pyrophosphate(C20) 牻牛兒基牻牛兒基焦磷酸 2 Phytoene(C40) 八氫番茄紅素 BR Abscisic acid Fig. 10.萜的生合成。
 牻牛兒基焦磷酸. 2. Farnesyl pyrophosphate(C15) 法尼基焦磷酸. Spualene(C30) 鯊烯. Cycloartenol. 環木鳳梨烯醇. Gibberellins. Campesterol. 菜油甾醇. Geranylgeranyl pyrophosphate(C20) 牻牛兒基牻牛兒基焦磷酸. 2. Phytoene(C40) 八氫番茄紅素. BR. Abscisic acid. Fig. 10.萜的生合成。")
60
BR Activities 1. 促進莖的伸長 BR可促進莖的伸長,若是mutant plant(無BR)則植株呈現低矮,此時即使施用GA或auxin也無法恢復生長。若施用brassinolide 或 castasterone可恢復生長,尤其在光照環境下 2. 促進根的生長 促進intact plant根的生長,但抑制excised root的生長 3. 抵抗環境逆境 植物在淹水、鹽害及施用殺草劑的環境下,施用brassinosteroids 可有效抵抗環境逆境。 4. 提升種子產量:可提升 45%種子產量

61
創傷和逆境的荷爾蒙: 1.乙烯(Ethylene)
在19世紀,當瓦斯用作街道照明時,發現到此街道樹木落葉的程度廣泛地大於其他街道,最後證實瓦斯中的乙烯是導致樹木落葉的成分。 在1901年,Dimitry Neljubov,發現生長在乙烯中的豌豆具有三相反應︰減緩莖的縱向伸長,增加橫向膨大和植株的不正常生長。 在1910年,H. H. Cousins指出乙烯是植物組織中自然的產物,他觀察到將柑桔儲藏在chamber中會釋放出一種氣體,造成香蕉後熟。而柑桔果實上的霉菌亦會產生大量的乙烯。 在1934年R. Gene利用化學方法證實乙烯在植物中的代謝反應,對植物有顯著的影響,被列為可能的植物荷爾蒙之一。 多年來乙烯被認為不是重要的植物荷爾蒙,因為許多生理學家相信乙烯的形成是由於auxin的關係(第一個被發現的植物荷爾蒙)。Auxin被認為是主要的荷爾蒙,而乙烯被認為是間接的生理過程。到了1959年,氣相層析儀的發明,才開始對乙烯的研究,證實乙烯確實是一種老化荷爾蒙(aging hormone)
。Auxin被認為是主要的荷爾蒙,而乙烯被認為是間接的生理過程。到了1959年,氣相層析儀的發明,才開始對乙烯的研究,證實乙烯確實是一種老化荷爾蒙(aging hormone)")
62
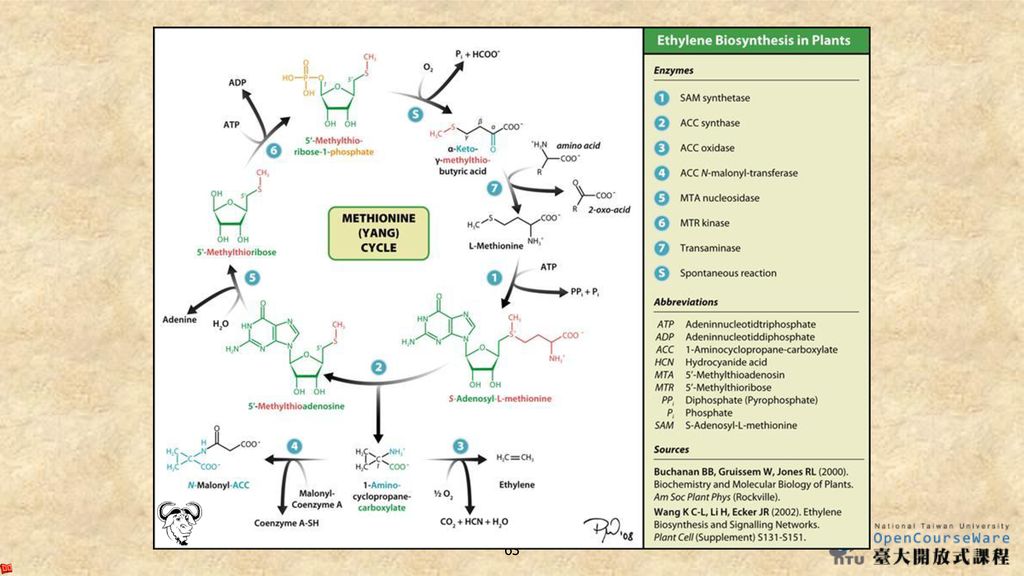
Ethylene Biosynthesis
蛋胺酸(甲硫胺酸)是乙烯的前驅物質,而S-腺苷蛋胺酸(SAM)經氨基環丙烷-1-羧酸合成酶(ACC synthase)催化下轉變成氨基環丙烷-1-羧酸(ACC, 1-aminocyclopropane-1-carboxylic acid),再經由ACC氧化酶,最終產生乙烯。
是乙烯的前驅物質,而S-腺苷蛋胺酸(SAM)經氨基環丙烷-1-羧酸合成酶(ACC synthase)催化下轉變成氨基環丙烷-1-羧酸(ACC, 1-aminocyclopropane-1-carboxylic acid),再經由ACC氧化酶,最終產生乙烯。")
64
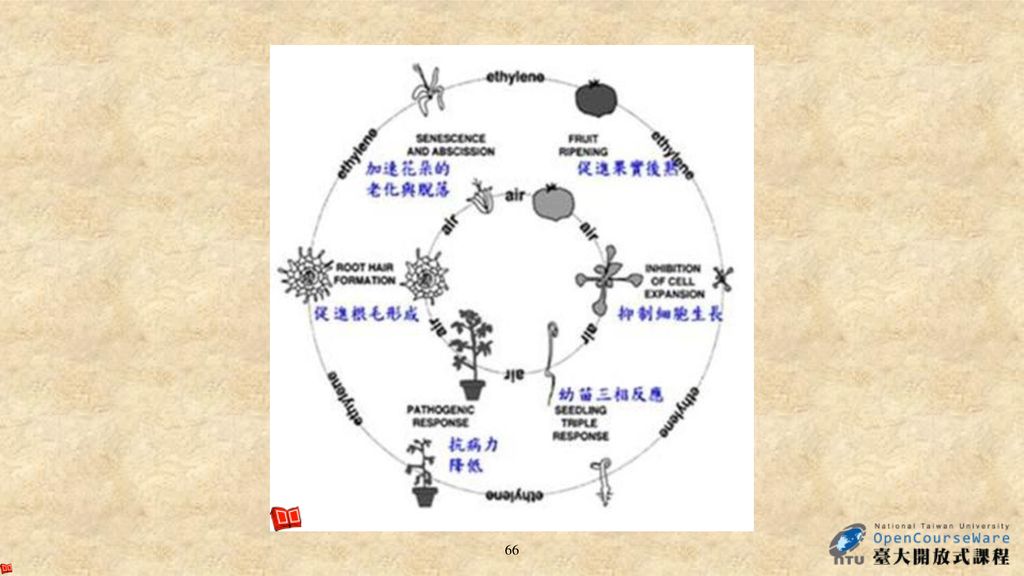
Ethylene Activity(1) 1. 乙烯具有三相反應(triple response)︰減緩莖的縱向伸長,增加橫向膨大和植株的不正常生長(橫向生長)。
︰減緩莖的縱向伸長,增加橫向膨大和植株的不正常生長(橫向生長)。")
65
2. 乙烯導致葉片的上偏生長性(epinasty)即葉柄上半部細胞生長較快,使葉片下垂︰ 乙烯或一些殺草劑(如2,4-D)均會導致葉片的上偏生長。 3. 乙烯參與誘導不定根形成,而且與auxin有關。乙烯與auxin在不定根形成上可能為互補的角色。在內生auxin被移除(如去除頂梢)或乙烯生合成被抑制時均使發根受到抑制。 番茄的葉片上偏生長

67
4.乙烯影響一些乳液的分泌︰ 如巴西橡膠樹(Hevea brasiliensis)受傷後即有乳液(latex)由乳汁管(laticifers)流出 ,當用ethephon處理之後,其乳液產量會增加一倍。用2,4-D、2,4,5-T及NAA也會促進乳液增加,此可能為施用auxin藥劑後促進乙烯的生合成所致。
受傷後即有乳液(latex)由乳汁管(laticifers)流出 ,當用ethephon處理之後,其乳液產量會增加一倍。用2,4-D、2,4,5-T及NAA也會促進乳液增加,此可能為施用auxin藥劑後促進乙烯的生合成所致。")
68
Ethylene Activity(2) 5. 乙烯可使植物適應淹水(缺氧)逆境
(1)植株淹水時,ACC在根部合成,但是它在淹水的無氧狀態下轉變成乙烯非常緩慢。ACC會經由木質部運移到枝條和葉片,再轉變成乙烯。導致葉片下垂、莖部膨大、新根形成。 (2)新形成的根和莖,由於乙烯可活化一些酵素(如cellulase)因而分解一些細胞,形成通氣組織(aerenchyma),所以較為適應淹水(缺氧)逆境。 (3)蓮花、睡蓮 和水稻等水生植物在淹水時會促進節間和葉柄的伸長,而使得葉片或上部枝條維持在水面上,而乙烯的處理也可獲得和淹水處理相似的結果。 3. 因為乙烯積聚在組織中而刺激水生植物的生長。當O2缺乏時,通氣組織(aerenchyma)提供水面下部分生長所需的氧氣。
植株淹水時,ACC在根部合成,但是它在淹水的無氧狀態下轉變成乙烯非常緩慢。ACC會經由木質部運移到枝條和葉片,再轉變成乙烯。導致葉片下垂、莖部膨大、新根形成。 (2)新形成的根和莖,由於乙烯可活化一些酵素(如cellulase)因而分解一些細胞,形成通氣組織(aerenchyma),所以較為適應淹水(缺氧)逆境。 (3)蓮花、睡蓮 和水稻等水生植物在淹水時會促進節間和葉柄的伸長,而使得葉片或上部枝條維持在水面上,而乙烯的處理也可獲得和淹水處理相似的結果。 3. 因為乙烯積聚在組織中而刺激水生植物的生長。當O2缺乏時,通氣組織(aerenchyma)提供水面下部分生長所需的氧氣。")
69
番茄植株淹水時,ACC在根部合成,但是它在淹水的無氧狀態下轉變成乙烯非常緩慢。ACC會經由木質部運移到枝條和葉片,然後再轉變成乙烯。氣體態的乙烯並不能運移,所以通常大多僅影響乙烯生成附近的組織。乙烯前趨物ACC則可運移且可以從遠離ACC合成的位置來產生乙烯(Bradford and Yang 1980)。 Plant physiology, Lincoln Taiz, Eduardo Zeiger, Sunderland, MA : Sinauer Associates, c2010, 5th edition, p. 661

70
Ethylene Activity(3) 6.乙烯促進葉片及果實的離層和老化
(1)外生乙烯或ACC(乙烯前趨物)的施用會加速葉片的離層和老化,而外生細胞分裂素的處理可以延緩葉片老化 。 (2)乙烯生成的增加與葉綠素的減少及褪色有關,這可作為將來葉片和花朵老化的特徵;另外亦發現葉片中細胞分裂素的含量和老化開始的時間具有負相關。因此,生理學研究指出老化可藉由乙烯和細胞分裂素間的平衡來調節。 (3)乙烯合成抑制劑(例如AVG或Co2+)和作用抑制劑(例如Ag+或CO2) 以及auxin會延緩葉片的離層和老化。
外生乙烯或ACC(乙烯前趨物)的施用會加速葉片的離層和老化,而外生細胞分裂素的處理可以延緩葉片老化 。 (2)乙烯生成的增加與葉綠素的減少及褪色有關,這可作為將來葉片和花朵老化的特徵;另外亦發現葉片中細胞分裂素的含量和老化開始的時間具有負相關。因此,生理學研究指出老化可藉由乙烯和細胞分裂素間的平衡來調節。 (3)乙烯合成抑制劑(例如AVG或Co2+)和作用抑制劑(例如Ag+或CO2) 以及auxin會延緩葉片的離層和老化。")
71
7. 乙烯促進果實後熟(Fruit Ripening)
圖. 果實成熟時乙烯、ACC含量和ACC氧化酶活性的改變。 黃元帥蘋果(Golden Delicious)ACC氧化酶活性、乙烯及ACC含量改變之情形,此數據為採收後天數的變化圖。乙烯、ACC濃度、ACC氧化酶活性的增加與後熟有密切的關係(Yang 1987)。
ACC氧化酶活性、乙烯及ACC含量改變之情形,此數據為採收後天數的變化圖。乙烯、ACC濃度、ACC氧化酶活性的增加與後熟有密切的關係(Yang 1987)。 sa=t&rct=j&q=&esrc=s&source=web&cd=3&ved=0CEIQFjAC&url=http%3A%2F%2Fwww.chnfood.cn%2Findex.php%3Fm%3DDownload%26a%3Ddown%26id%3D9757&ei=KcmZUfbfCIeAlQWgqoGQBw&usg=AFQjCNEUDdkTNYqxeYTZ7BqzPGsP5CUgpw&sig2=Y2qZ1BS6iaeP6HPsWpxKZw.")
72
Ethylene Activity(4) 8. 乙烯的其他作用 例如乙烯可促進葉綠素分解、 打破芽體和種子休眠、 促進種子發芽、
促進鳳梨科等植物的開花、 促進雌花的花性表現、 促進側芽生長

73
益收生長素( Ethephon ) Ethephon 為一乙烯釋放物質,化學名稱為2-chloroethylphosphonic acid,國外商品名「Ethrel、Florel、CEPA」,國內商品名「益收生長素」, 是一濃度39.5%之溶液。於pH值小於3.5的酸性環境下可穩定存在,處理後進入植體內(pH值多半高於4.0)則水解釋放出乙烯與磷酸,便能造成與內生乙烯相同的生理作用。
則水解釋放出乙烯與磷酸,便能造成與內生乙烯相同的生理作用。")
74
2.Abscisic acid(ABA) 離層酸(脫落酸)
它是人們在研究植物體內與休眠、脫落和種子萌發等生理過程有關的生長抑制物質時發現的。 1961年劉(W. C. Liu)等在研究棉花幼鈴的脫落時,從成熟的乾棉殼中分離純化出了促進脫落的物質,並命名這種物質為離層素,后來阿迪柯特將其稱為離層素Ⅰ(Abscisin I)。 1963年大熊和彥和阿迪柯特(K. Ohkuma and F. T. Addicott)等從225 kg 4~7 d齡的鮮棉鈴(Gossypium hirsutum)中分離純化出了9 mg具有高度活性的促進脫落的物質,命名為離層素Ⅱ(abscisinⅡ)。
等在研究棉花幼鈴的脫落時,從成熟的乾棉殼中分離純化出了促進脫落的物質,並命名這種物質為離層素,后來阿迪柯特將其稱為離層素Ⅰ(Abscisin I)。 1963年大熊和彥和阿迪柯特(K. Ohkuma and F. T. Addicott)等從225 kg 4~7 d齡的鮮棉鈴(Gossypium hirsutum)中分離純化出了9 mg具有高度活性的促進脫落的物質,命名為離層素Ⅱ(abscisinⅡ)。")
75
Abscisic acid(ABA) 離層酸(脫落酸)
在阿迪柯特領導的小組研究棉鈴脫落的同時,英國的韋爾林和康福思(P. F. Wareing and J. W. Cornforth)領導的小組正在進行著木本植物休眠的研究。 幾乎就在離層素Ⅱ發現的同時,伊芳芳格爾斯(C. F. Eagles)和韋爾林從楓樹葉(Acer pseudoplatanus)中提取出了一種能抑制生長並誘導旺盛生長的枝條進入休眠的物質,他們將其命名為休眠素(dormin)。 1965年康福思等從28kg秋天的乾楓樹葉中得到了260μg的休眠素純結晶,透過與脫落素Ⅱ的分子量、紅外光譜和熔點等的比較鑑定,確定休眠素和脫落素Ⅱ是同一物質。 1967年在渥太華召開的第六屆國際植物生長物質會議上,這種生長調節物質正式被定名為離層酸(脫落酸)
領導的小組正在進行著木本植物休眠的研究。 幾乎就在離層素Ⅱ發現的同時,伊芳芳格爾斯(C. F. Eagles)和韋爾林從楓樹葉(Acer pseudoplatanus)中提取出了一種能抑制生長並誘導旺盛生長的枝條進入休眠的物質,他們將其命名為休眠素(dormin)。 1965年康福思等從28kg秋天的乾楓樹葉中得到了260μg的休眠素純結晶,透過與脫落素Ⅱ的分子量、紅外光譜和熔點等的比較鑑定,確定休眠素和脫落素Ⅱ是同一物質。 1967年在渥太華召開的第六屆國際植物生長物質會議上,這種生長調節物質正式被定名為離層酸(脫落酸)")
76
ABA 生物合成 合成部位︰主要在根尖和葉片細胞的質體內(葉中是葉綠體,根中是澱粉體)。此外,在莖、種子、花和果等器官中也能合成ABA
前體︰甲瓦龍酸(MVA) 合成途徑︰ 直接途徑─由MVA合成而來 間接途徑─由類胡蘿蔔素(紫黃質、葉黃素等)裂解而來
 合成途徑︰ 直接途徑─由MVA合成而來. 間接途徑─由類胡蘿蔔素(紫黃質、葉黃素等)裂解而來.")
77
甲瓦龍酸 C5 異戊烯基焦磷酸 古巴焦磷酸 C10 法尼焦磷酸 C15 ABA 紫黄質 黄質醛 C15 間接途徑 直接途徑 (MVA)
(IPP) (Violaxanthin) (FPP) (Xantholal) 直接途徑
 (Violaxanthin) (FPP) (Xantholal) 直接途徑.")
78
甲瓦龍酸 細胞分裂素 異戊烯基焦磷酸 胡蘿蔔素 離層酸 激勃素

79
(Taiz and Zeiger,2010)
")
80
ABA Activities 1. 抑制生長︰ABA能抑制整株植物或離體器官的生長,也能抑制種子的萌發。
2. 於低濃度( and l0-8 M )下可促進生長,如在乾旱逆境下促根生長 3. 促進植物器官脫落,如葉、花、果實脫落 4. 促進種子及芽的休眠︰外施ABA時,可使旺盛生長的枝條停止生長而進入休眠,這是它最初也被稱為“休眠素”的原因。 5. 促進氣孔關閉︰外施ABA,氣孔可於2-3分鐘關閉。於逆境環境下(如水分及溫度逆境、鹽害),ABA上升,氣孔關閉,增加抗逆性,故ABA又稱為逆境荷爾蒙。 6. 維持頂芽優勢,抑制分枝形成
下可促進生長,如在乾旱逆境下促根生長. 3. 促進植物器官脫落,如葉、花、果實脫落. 4. 促進種子及芽的休眠︰外施ABA時,可使旺盛生長的枝條停止生長而進入休眠,這是它最初也被稱為 休眠素 的原因。 5. 促進氣孔關閉︰外施ABA,氣孔可於2-3分鐘關閉。於逆境環境下(如水分及溫度逆境、鹽害),ABA上升,氣孔關閉,增加抗逆性,故ABA又稱為逆境荷爾蒙。 6. 維持頂芽優勢,抑制分枝形成.")
81
3.茉莉酸( Jasmonates ) 茉莉酸(jasmonic acid,JA) 、茉莉酸甲酯(methyl jasmonate,JA-ME)和其他JA的衍生物,是一種特殊之環戊酮化合物。這一類的化合物總稱為Jasmonates (Jose Leon et al., 1999) 。 JA-ME的發現比JA稍早,約在1962年 從大花茉莉的精油分離出來,是精油裡香氣的成分之ㄧ(Aldrige et al., 1971)。 JA在1971年最早由座腔雙胞菌(Lasiodiplodia theobromae)的培養液中分離出來,後來發現為座腔雙胞菌的代謝產物。 JA 是植物體內最豐富的形式, JA 羧基中的氫被甲基取代後即形成JA-Me (Sembdner and Partjhoer ,1993) 。由於JA-ME 較易揮發且不被離子化,易透過細胞膜,因而JA-ME 形式在外源施用時活性較強,是首選的形式。
。 JA在1971年最早由座腔雙胞菌(Lasiodiplodia theobromae)的培養液中分離出來,後來發現為座腔雙胞菌的代謝產物。 JA 是植物體內最豐富的形式, JA 羧基中的氫被甲基取代後即形成JA-Me (Sembdner and Partjhoer ,1993) 。由於JA-ME 較易揮發且不被離子化,易透過細胞膜,因而JA-ME 形式在外源施用時活性較強,是首選的形式。")
82
Jasmonates JA分佈量以在植物 sink 部位,如花、幼芽和幼葉等位置的較高,植株成熟部位的含量較少
亞麻酸經脂氧合酶(lipoxygenase)催化加氧作用產生脂肪酸過氧化氫物,再經過氧化氫物環化酶(hydroperoxide cyclase)的作用轉變為18碳的環脂肪酸(cyclic fatty acid),最後經還原及多次β-氧化而形成JA。
催化加氧作用產生脂肪酸過氧化氫物,再經過氧化氫物環化酶(hydroperoxide cyclase)的作用轉變為18碳的環脂肪酸(cyclic fatty acid),最後經還原及多次β-氧化而形成JA。")
83
茉莉酸之生物合成 亞麻酸 LOX 氫過氧化物脫氫酶 13-氫過氧化亞麻酸 12 ,13-環氧亞麻酸 AOC 12-氧代植二烯酸
衍生出各種茉莉酸鹽(jasmonates) 氫過氧化物脫氫酶 12 ,13-環氧亞麻酸 (Sembdner, G. and B. Parthier, 1993)
 氫過氧化物脫氫酶. 12 ,13-環氧亞麻酸. (Sembdner, G. and B. Parthier, 1993)")
84
Jasmonate Activity(1) 1. 參與植物防禦機制
植物遭受創傷時,其內生JA含量增加,促使一些蛋白質(如proteinase inhibitors和chitinases)的生成以防禦微生物的病原體和昆蟲的侵襲。另外也會增加volatile aldehydes及oxoacids的含量,以增進植物的防禦能力。 2. 促進植物卷鬚纏繞 3. 抑制莖及根的生長、種子發芽及光合作用 JA與abscisic acid(ABA)的架構相似,具有相似分子量與溶解度以及促進葉片老化的作用,因此JA所造成的生理作用常被拿來與ABA比較,總之JA和ABA皆在逆境環境下抑制植物生長以抵御環境逆境。
的生成以防禦微生物的病原體和昆蟲的侵襲。另外也會增加volatile aldehydes及oxoacids的含量,以增進植物的防禦能力。 2. 促進植物卷鬚纏繞. 3. 抑制莖及根的生長、種子發芽及光合作用. JA與abscisic acid(ABA)的架構相似,具有相似分子量與溶解度以及促進葉片老化的作用,因此JA所造成的生理作用常被拿來與ABA比較,總之JA和ABA皆在逆境環境下抑制植物生長以抵御環境逆境。")
85
Jasmonate Activity(2) 4. jasmonates可誘導一些特殊蛋白質的合成
塊莖、球莖及種子其營養器官具有一些貯藏性蛋白質,對於植物抵御環境逆境有重要的影響,而這些貯藏性蛋白質藉由genes coding 證實為jasmonates誘導生成。 5. 促進塊莖形成 6.促進乙烯生合成、具有與乙烯類似之作用,例如促進果實成熟、促進葉片、花朵老化及脫落

86
Other plant growth substances
SA(水楊酸) PA(多胺)
 PA(多胺)")
87
一、水楊酸salicylic acid,SA
1. 水楊酸的發現 1763年英國的斯通(E.Stone)首先發現柳樹皮有很強的收斂作用,可以治療瘧疾和發燒。
首先發現柳樹皮有很強的收斂作用,可以治療瘧疾和發燒。")
88
salicylic acid,SA 後來發現這是柳樹皮中所含的水楊酸鹽-水楊醇配醣體(salicyl alcohol glucoside, SAG) 在起作用,於是經過許多藥物學家和化學家的努力,醫學上便有了阿斯匹林(aspirin)藥物的問世。阿斯匹林即乙醯水楊酸(acetylsalicylic acid),在生物體內可很快轉化為水楊酸(salicylic acid, SA)。
 在起作用,於是經過許多藥物學家和化學家的努力,醫學上便有了阿斯匹林(aspirin)藥物的問世。阿斯匹林即乙醯水楊酸(acetylsalicylic acid),在生物體內可很快轉化為水楊酸(salicylic acid, SA)。")
89
2. 水楊酸的分布 SA在植物體中的分佈一般以產熱植物的花序較多,如天南星科海芋屬植物的花序,該類植物開花時溫度上升,比環境溫度高得多。(產熱作用) 海芋

90
3. 水楊酸的生物合成 植物體內SA的合成來自反式肉桂酸(trans-cinnamic acid),即由莽草酸(shikimic acid)經苯丙氨酸(phenylalanine)形成的反式肉桂酸可經鄰香豆酸(o-coumaric acid)或苯甲酸轉化成SA。 (Hayat et al., 2007)
")
91
4. 水楊酸的生理效應 增強抗性 某些抗病植物在受到病原侵染后,其體內SA含量立即升高,SA能誘導抗病基因的活化而使植株產生抗性。
誘導開花 用5.6μmol‧L-1的SA處理可使長日植物浮萍gibbaG3在非誘導光週期下開花。 其它 SA還抑制蒸散作用、抑制乙烯生成,被用於切花保鮮、水稻抗寒等方面。

92
二、多胺─ 是一類脂肪族含氮鹼 生理功能: 1、促進生長:加快 DNA轉錄,增强RNA聚合酶活性和加快氨基酸掺入蛋白质的速度,促進核酸和蛋白質合成。 多胺有五种:精胺(spermine)、亞精胺(spermidine)、腐胺(putrescine)、鲱精胺(agmatine)、屍胺(cadaverine); 它們都是由精氨酸和賴氨酸生物合成而來。
、亞精胺(spermidine)、腐胺(putrescine)、鲱精胺(agmatine)、屍胺(cadaverine); 它們都是由精氨酸和賴氨酸生物合成而來。")
93
2、延緩衰老:保持光合膜完整性,減緩蛋白質喪失和RNase活性,阻止葉綠素破壞。另外多胺與乙烯爭奪SAM,故抑制乙烯生成 。
3、適應逆境條件:多胺帶正電荷,可代替K+、Mg2+等陽離子維持細胞pH。另外,滲透脅迫下,多胺顯著增多,維持滲透平衡,保護膜穩定和原生質完整。

94
植物荷爾蒙代謝的相互關係 GA3 抑制 結合態IAA CTKS ABA 生物合成 GA 結合態GA 促進 生物合成 IAA IAA氧化E
生物合成 ETH 低濃度促進 CTKS ABA 生物合成 GA 結合態GA 植物荷爾蒙代謝的相互關係 高濃度抑制

95
下課了 下次見
96
版權聲明 頁碼 作品 版權標示 作者 / 來源 10 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.298 依據著作權法第46、52、65條合理使用。 植物的向光性與IAA的分佈有關 作品來自 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.298。 由國立臺灣大學 園藝暨景觀學系 張育森 教授 整理翻譯。 依據著作權法第46、52、65條合理使用。 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.299 室內植物往光源的方向彎曲 作品來自 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.299。 由國立臺灣大學 園藝暨景觀學系 張育森 教授 整理翻譯。 依據著作權法第46、52、65條合理使用。 11 Plant physiology, Lincoln Taiz, Eduardo Zeiger, Sunderland, MA : Sinauer Associates, c2010, 5th edition, p. 547

97
頁碼 作品 版權標示 作者 / 來源 12 Plant physiology, Lincoln Taiz, Eduardo Zeiger, Sunderland, MA : Sinauer Associates, c2010, 5th edition, p. 547 依據著作權法第46、52、65條合理使用。 13 揚州大學生物科學與技術學院植物生理教研室 ( 瀏覽日期2013/5/23,依據著作權法第46、52、65條合理使用。 14 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.299 15 過量的auxin會有殺草劑效應 作品來自 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.299。 由國立臺灣大學 園藝暨景觀學系 張育森 教授 整理翻譯。 依據著作權法第46、52、65條合理使用。

98
17 18 19 頁碼 作品 版權標示 作者 / 來源 國立臺灣大學 園藝暨景觀學系 張育森 教授。
Plant physiology, Lincoln Taiz, Eduardo Zeiger, Sunderland, MA : Sinauer Associates, c2006, 4th edition, 依據著作權法第46、52、65條合理使用。 19 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.300 蘋果或葡萄若缺乏種子,……其果實發育異常。 作品來自 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.300。 由國立臺灣大學 園藝暨景觀學系 張育森 教授 整理翻譯。 依據著作權法第46、52、65條合理使用。 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.332

99
頁碼 作品 版權標示 作者 / 來源 19 較大果實含較多種子 作品來自 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.332。 由國立臺灣大學 園藝暨景觀學系 張育森 教授 整理翻譯。 依據著作權法第46、52、65條合理使用。 21 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.301 依據著作權法第46、52、65條合理使用。 Auxin促進頂芽優勢……造成單一主幹 作品來自 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.301。 由國立臺灣大學 園藝暨景觀學系 張育森 教授 整理翻譯。 依據著作權法第46、52、65條合理使用。 22 修剪去頂,造成多分枝的灌叢苗木

100
頁碼 作品 版權標示 作者 / 來源 23 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.301 依據著作權法第46、52、65條合理使用。 但施用NAA亦可減緩採收前的落果……品質有很大的影響。太多果實會使果實變小,……或配合加保利進行疏果 作品來自 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.301。 由國立臺灣大學 園藝暨景觀學系 張育森 教授 整理翻譯。 依據著作權法第46、52、65條合理使用。 24 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.302 此外聖誕紅的插穗…… (Figure 11-9) 施用IBA粉劑,使聖誕紅根系發達 作品來自 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.302。 由國立臺灣大學 園藝暨景觀學系 張育森 教授 整理翻譯。 依據著作權法第46、52、65條合理使用。 25 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.303
 施用IBA粉劑,使聖誕紅根系發達. 作品來自 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.302。 由國立臺灣大學 園藝暨景觀學系 張育森 教授 整理翻譯。 依據著作權法第46、52、65條合理使用。 25. The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.303.")
101
頁碼 作品 版權標示 作者 / 來源 25 滑石粉稀釋的auxin,可促進插穗發根 作品來自 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.303。 由國立臺灣大學 園藝暨景觀學系 張育森 教授 整理翻譯。 依據著作權法第46、52、65條合理使用。 26 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.303 依據著作權法第46、52、65條合理使用。 植物根部對auxin敏感……(Figure 11-11) 美國白臘樹在試管……造成短粗的根(右) 27 國立臺灣大學 園藝暨景觀學系 張育森 教授。 29 Wikipedia / Calvero. ( 瀏覽日期2013/5/23,依據著作權法第46、52、65條合理使用。
 美國白臘樹在試管……造成短粗的根(右) 27. 國立臺灣大學 園藝暨景觀學系 張育森 教授。 29. Wikipedia / Calvero. ( 瀏覽日期2013/5/23,依據著作權法第46、52、65條合理使用。")
102
頁碼 作品 版權標示 作者 / 來源 29 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.304 依據著作權法第46、52、65條合理使用。 31 Koning, Ross E Gibberellins. Plant Physiology Information Website. ( )., ,依據著作權法第46、52、65條合理使用。 32 Constance N. White2, William M. Proebsting, Peter Hedden, and Carol J. Rivin Gibberellins and Seed Development in Maize. I. Evidence That Gibberellin/Abscisic Acid Balance Governs Germination versus Maturation Pathways. Plant Physiology,Vol. 122,pp.1081–1088. 33 國立臺灣大學 園藝暨景觀學系 張育森 教授。 34 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.306
., ,依據著作權法第46、52、65條合理使用。 32. Constance N. White2, William M. Proebsting, Peter Hedden, and Carol J. Rivin Gibberellins and Seed Development in Maize. I. Evidence That Gibberellin/Abscisic Acid Balance Governs Germination versus Maturation Pathways. Plant Physiology,Vol. 122,pp.1081– 國立臺灣大學 園藝暨景觀學系 張育森 教授。 34. The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.306.")
103
頁碼 作品 版權標示 作者 / 來源 34 矮生豌豆缺乏GA(左) 施用GA促節間伸長(右) 作品來自 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.306。 由國立臺灣大學 園藝暨景觀學系 張育森 教授 整理翻譯。 依據著作權法第46、52、65條合理使用。 35 揚州大學生物科學與技術學院植物生理教研室 ( 瀏覽日期2013/5/23,依據著作權法第46、52、65條合理使用。 36 Plant physiology, Lincoln Taiz, Eduardo Zeiger, Sunderland, MA : Sinauer Associates, c2006, 4th edition。 依據著作權法第46、52、65條合理使用。 IAA強化GA的作用 作品來自 Plant physiology, Lincoln Taiz, Eduardo Zeiger, Sunderland, MA : Sinauer Associates, c2006, 4th edition。 由國立臺灣大學 園藝暨景觀學系 張育森 教授 整理翻譯。 依據著作權法第46、52、65條合理使用。 37 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.306

104
頁碼 作品 版權標示 作者 / 來源 37 內生GA促進大麥種子發芽過程 作品來自 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.306。 由國立臺灣大學 園藝暨景觀學系 張育森 教授 整理翻譯。 依據著作權法第46、52、65條合理使用。 42 揚州大學生物科學與技術學院植物生理教研室 ( 瀏覽日期2013/5/23,依據著作權法第46、52、65條合理使用。 45 國立臺灣大學 園藝暨景觀學系 張育森 教授。 Flickr / J€RRY ( ), 瀏覽日期:2013/01/03。 47 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.309 依據著作權法第46、52、65條合理使用。
, 瀏覽日期:2013/01/03。 47. The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.309. 依據著作權法第46、52、65條合理使用。")
105
頁碼 作品 版權標示 作者 / 來源 47 彩葉草及印度橡膠的葉片可誘使其生成不定根, ……此可能與cytokinin的作用有關。 作品來自 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.309。 由國立臺灣大學 園藝暨景觀學系 張育森 教授 整理翻譯。 依據著作權法第46、52、65條合理使用。 49 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.309 依據著作權法第46、52、65條合理使用。 (左)培養基中〔CTK〕/〔IAA〕的比值高時,……癒傷組織形成根; 50 Proceedings of the Royal Society Biological Sciences ( From:Proc Biol Sci. 2010 August 7; 277(1692): 2311–2319. Published online 2010 March 31. doi: /rspb ,瀏覽日期 2013/06/19,依據著作權法第 46、52、65 條合理使用。 51 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.310
培養基中〔CTK〕/〔IAA〕的比值高時,……癒傷組織形成根; 50. Proceedings of the Royal Society Biological Sciences. ( From:Proc Biol Sci August 7; 277(1692): 2311–2319. Published online 2010 March 31. doi: /rspb ,瀏覽日期 2013/06/19,依據著作權法第 46、52、65 條合理使用。 51. The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.310.")
106
頁碼 作品 版權標示 作者 / 來源 51 被農桿菌(Agrobacterium tumefaciens)感染後,…… (Figure 11-18) 作品來自 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.310。 由國立臺灣大學 園藝暨景觀學系 張育森 教授 整理翻譯。 依據著作權法第46、52、65條合理使用。 牙醫保健與生態網 / 雲林縣西螺鎮公所 版權所有 ( ,依據著作權法第 46、52、65 條合理使用。 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.310 依據著作權法第46、52、65條合理使用。 藉由昆蟲、螨類、寄生植物或基因突變所造成…… (Figure ) 。 52 Plant physiology, Lincoln Taiz, Eduardo Zeiger, Sunderland, MA : Sinauer Associates, c2010, 5th edition, p. 626
,瀏覽日期 ,依據著作權法第 46、52、65 條合理使用。 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.310. 依據著作權法第46、52、65條合理使用。 藉由昆蟲、螨類、寄生植物或基因突變所造成…… (Figure ) 。 52. Plant physiology, Lincoln Taiz, Eduardo Zeiger, Sunderland, MA : Sinauer Associates, c2010, 5th edition, p")
107
54 56 57 58 頁碼 作品 版權標示 作者 / 來源 豆丁網 / 植物生長生理p.41
( 瀏覽日期2013/5/23,依據著作權法第46、52、65條合理使用。 56 Wikipedia / Wostr ( 57 Uday K. Divi and Priti Krishna.2009.Brassinosteroid: a biotechnological target for enhancing crop yield and stress tolerance.P 依據著作權法第46、52、65條合理使用。 58 徐善德、廖玉琬 編譯.2006.植物生理學.偉明圖書有限公司.pp602. 依據著作權法第46、52、65條合理使用。

108
59 62 63 64 頁碼 作品 版權標示 作者 / 來源 徐善德、廖玉琬 編譯.2006.植物生理學.偉明圖書有限公司.pp602.
依據著作權法第46、52、65條合理使用。 62 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.312 蛋胺酸(甲硫胺酸)是乙烯的前驅物質,……最終產生乙烯。 作品來自 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.312。 由國立臺灣大學 園藝暨景觀學系 張育森 教授 整理翻譯。 依據著作權法第46、52、65條合理使用。 63 WIKIPEDIA / Crenim ( 瀏覽日期 2013/06/19。 64 Proceedings of the National Academy of Sciences / Copyright © 2013 National Academy of Sciences ( 瀏覽日期 2013/06/19,依據著作權法第 46、52、65 條合理使用。
是乙烯的前驅物質,……最終產生乙烯。 作品來自 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.312。 由國立臺灣大學 園藝暨景觀學系 張育森 教授 整理翻譯。 依據著作權法第46、52、65條合理使用。 63. WIKIPEDIA / Crenim. ( 瀏覽日期 2013/06/19。 64. Proceedings of the National Academy of Sciences / Copyright © 2013 National Academy of Sciences. ( 瀏覽日期 2013/06/19,依據著作權法第 46、52、65 條合理使用。")
109
頁碼 作品 版權標示 作者 / 來源 65 The biology of horticulture: an introductory textbook, John E. Preece, Paul E. Read, Hoboken, NJ : J. Wiley, c2005, 2nd edition, p.313 依據著作權法第46、52、65條合理使用。 66 花博士網路科學教室 / 著作權所有人:臺北市政府 ( 瀏覽日期 2013/06/19,依據著作權法第 46、52、65 條合理使用。 67 Wikipedia / Irvin calicut ( 瀏覽日期2013/05/30。 Wikipedia / Solitude ( 瀏覽日期:2013/06/19。 68 IOP Publishing (

110
頁碼 作品 版權標示 作者 / 來源 69 Plant physiology, Lincoln Taiz, Eduardo Zeiger, Sunderland, MA : Sinauer Associates, c2010, 5th edition, p. 661 依據著作權法第46、52、65條合理使用。 71 桃果实成熟软化过程中α -L- 阿拉伯呋喃糖苷酶活性和乙烯合成相关因子的变化,金昌海,阚 娟,索 标,汪志君,陆兆新,郁志芳, 食品科学 2006, Vol. 27, No. 1 63頁 ( 2013/06/19,依據著作權法第 46、52、65 條合理使用。 73 WIKIMEDIA COMMONS / Edgar181 ( 瀏覽日期 2013/06/19,依據著作權法第 46、52、65 條合理使用。 77 國立臺灣大學 園藝暨景觀學系 張育森 教授。 78
,瀏覽日期 2013/06/19,依據著作權法第 46、52、65 條合理使用。 73. WIKIMEDIA COMMONS / Edgar181. ( 瀏覽日期 2013/06/19,依據著作權法第 46、52、65 條合理使用。 77. 國立臺灣大學 園藝暨景觀學系 張育森 教授。 78.")
111
頁碼 作品 版權標示 作者 / 來源 79 Plant physiology, Lincoln Taiz, Eduardo Zeiger, Sunderland, MA : Sinauer Associates, c2010, 5th edition,pA3-10. 依據著作權法第46、52、65條合理使用。 83 The Biochemistry and the Physiological and Molecular Actions of Jasmonates / Gunther Sembdner and Benno Parthier ( 瀏覽日期2013/5/23,依據著作權法第46、52、65條合理使用。 87 科學月刊雜誌社.金台灣資訊事業有限公司 ( 89 Wikimedia commons / Fanghong ( 瀏覽日期:2012/12/31。 Flickr / Sinchen.Lin ( 瀏覽日期2013/5/23。

112
頁碼 作品 版權標示 作者 / 來源 90 Salicylic Acid: Biosynthesis, Metabolism and Physiological Role in Plants / S. Hayat, B. Ali and A. Ahmad ( 瀏覽日期2013/5/23,依據著作權法第46、52、65條合理使用。 94 國立臺灣大學 園藝暨景觀學系 張育森 教授。 95 Scholars Junction ( Open Clip Art Library / palomaironique ( 2013/06/19。 1-112 轉載自 Microsoft Office 2010 PowerPoint 設計主題範本, 依據 Microsoft 服務合約及著作權法第 46、52、65 條合理使用。

Similar presentations

 皆為異營真核生物大多為陸生大多具菌絲 ( 營養菌絲, 繁殖菌絲 )>")
>")



>")
>")
>")




細胞衰老(senescence) (2)基因體的不穩定(genomic instability)>")





