Download presentation
Presentation is loading. Please wait.

1
第十三、十四章 细胞增殖及其调控

2
细胞增殖(cell proliferation)的意义
第一节、细胞周期与细胞分裂 第二节、细胞周期调控

3
细胞增殖的意义 细胞增殖是细胞生命活动的重要特征之一,是生物繁育的基础。 单细胞生物细胞增殖导致生物个体数量的增加。
多细胞生物由一个单细胞(受精卵)分裂发育而来,细胞增殖是多细胞生物繁殖基础。 成体生物仍然需要细胞增殖,主要取代衰老死亡的细胞,维持个体细胞数量的相对平衡和机体的正常功能。 机体创伤愈合、组织再生、病理组织修复等,都要依赖细胞增殖。
分裂发育而来,细胞增殖是多细胞生物繁殖基础。 成体生物仍然需要细胞增殖,主要取代衰老死亡的细胞,维持个体细胞数量的相对平衡和机体的正常功能。 机体创伤愈合、组织再生、病理组织修复等,都要依赖细胞增殖。")
4
第一节 细胞周期与细胞分裂 一、细胞周期(cell cycle)概述 二、有丝分裂(mitosis) 三、胞质分裂(Cytokinesis)
第一节 细胞周期与细胞分裂 一、细胞周期(cell cycle)概述 二、有丝分裂(mitosis) 三、胞质分裂(Cytokinesis) 四、减数分裂(Meiosis)
概述. 二、有丝分裂(mitosis) 三、胞质分裂(Cytokinesis) 四、减数分裂(Meiosis)")
5
第二节 细胞周期的调控 (Cell-Cycle Control)
一、细胞周期调控系统的主要作用 二、细胞周期检验点(Cell Cycle Checkpoint) 三、MPF 四、Cyclin-Cdk复合物的多样性及细胞周期运转 五、细胞周期运转的阻遏(细胞周期运转的负调控)
 三、MPF. 四、Cyclin-Cdk复合物的多样性及细胞周期运转. 五、细胞周期运转的阻遏(细胞周期运转的负调控)")
6
一、细胞周期(cell cycle)概述 细胞周期 细胞周期中各个不同时相及其主要事件 细胞周期长短测定 细胞周期同步化 特异的细胞周期
概述 细胞周期 细胞周期中各个不同时相及其主要事件 细胞周期长短测定 细胞周期同步化 特异的细胞周期")
7
二、有丝分裂(mitosis) 一、有丝分裂过程 二、与有丝分裂直接相关的亚细胞器结构 三、有丝分裂过程中染色体运动的动力机制
前期(prophase) 前中期(prometaphase) 中期(metaphase) 后期(anaphase) 末期(telophase) 胞质分裂期(cytokinesis) 二、与有丝分裂直接相关的亚细胞器结构 中心体 动粒与着丝粒 纺缍体 三、有丝分裂过程中染色体运动的动力机制
 前中期(prometaphase) 中期(metaphase) 后期(anaphase) 末期(telophase) 胞质分裂期(cytokinesis) 二、与有丝分裂直接相关的亚细胞器结构. 中心体 动粒与着丝粒 纺缍体. 三、有丝分裂过程中染色体运动的动力机制.")
9
中心体是高等动物细胞内与微管装配和细胞分裂密切相关的细胞器。
中心体一般由一对位于中央的中心粒和其周围的无定型物质构成。 中心体在细胞周期过程中也要进行复制,并经历一系列的发育过程,称为中心体周期。

10
动粒称为着丝点,是附于着丝粒上的一种细胞器。着丝粒是指染色体主缢痕部位的染色质。
目前已分离了几种动粒蛋白成分。CENP-E是一种驱动蛋白,定位于动粒外层表面的冠上。CENP-E被认为在促使与染色体与来自两极的微管相联结过程中起重要作用。

11
纺缍体是细胞分裂过程中的一种与染色体分离直接相关的细胞器。高等细胞的纺缍体呈纺缍状,主要由微管和微管结合蛋白组成。纺缍体的两端为星体。
纺缍体的装配

14
三、胞质分裂(Cytokinesis) 动物细胞胞质分裂 植物细胞胞质分裂
 动物细胞胞质分裂 植物细胞胞质分裂")
15
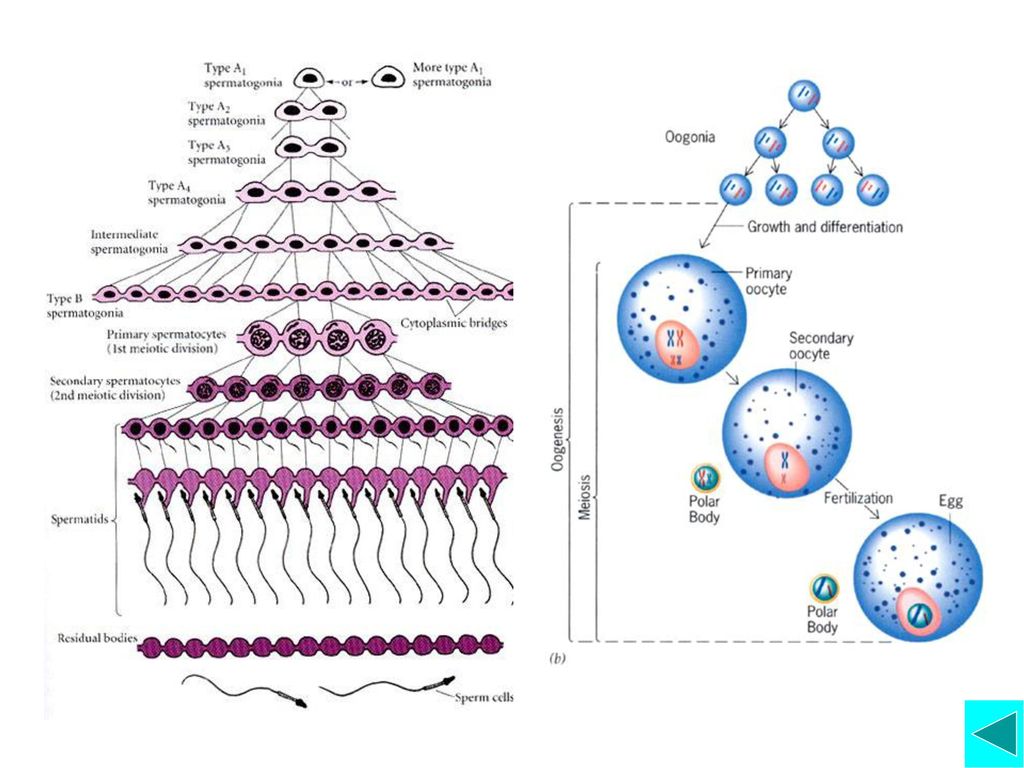
四、减数分裂(Meiosis) 减数分裂概念与过程 减数分裂的意义 减数分裂各时相及其主要事件 脊椎动物配子发生过程
 减数分裂概念与过程 减数分裂的意义 减数分裂各时相及其主要事件 脊椎动物配子发生过程")
16
细胞周期 细胞周期时相组成 细胞周期时间 根据增殖状况,细胞分类三类
连续分裂的细胞从一次有丝分裂结束到下一次有丝分裂完成所经历的一个有序过程。其间细胞遗传物质和其他内含物分配给子细胞。整个细胞周期又可分为G1期、S期、G2期和M期四个时期。 细胞周期时相组成 细胞周期时间 根据增殖状况,细胞分类三类

17
细胞周期时相组成 间期(interphase): G1期,S期,G2期
M期: 有丝分裂期(Mitosis), 胞质分裂期(Cytokinesis) 细胞沿着G1→S→G2→M→G1周期性运转,在间期细胞体积增大(生长),在 M 期细胞先是核分裂,接着胞质分裂,完成一个细胞周期。
, 胞质分裂期(Cytokinesis) 细胞沿着G1→S→G2→M→G1周期性运转,在间期细胞体积增大(生长),在 M 期细胞先是核分裂,接着胞质分裂,完成一个细胞周期。")
18
细胞周期时间 不同细胞的细胞周期时间差异很大 S+G2+M 的时间变化较小,细胞周期时间长 短主要差别在G1期
有些分裂增殖的细胞(早期胚胎细胞)缺乏G1、 G2期
缺乏G1、 G2期.")
19
根据增殖状况,细胞分类三类 连续分裂细胞(cycling cell) 休眠细胞(G0细胞) 终末分化细胞
体内的某些细胞由于分化程度很高,一经形成终生不再 分裂。 G0期细胞和终末分化细胞的界限有时难以划分,有的细胞过去认为属于终末分化细胞,目前可能被认为是G0期细胞。

20
细胞周期中不同时相及其主要事件 ◆ G1期 ◆ S 期 ◆ G2期 ◆ M 期

21
G1期 与DNA合成启动相关,开始合成细胞生长所需要的多种蛋 白质、RNA、碳水化合物、脂等,同时染色质去凝集。
G1期早期合成的物质主要与细胞生长相关;后期合成的物 质主要与细胞进入S期相关。 不同的物种G1期的长短不同。 G1期能否进入S期取决于细胞能否通过限制点(restriction point R点)。如能通过R点则细胞就可以进入S期,如不能 通过则细胞就会进入G0期或进一步分化。 影响细胞从G1期进入S期的因素
。如能通过R点则细胞就可以进入S期,如不能 通过则细胞就会进入G0期或进一步分化。 影响细胞从G1期进入S期的因素.")
22
G2期 DNA复制完成,在G2期合成一定数量的蛋白质和RNA分子,为细胞进入M期做准备。
细胞在从G2期向M期过渡的时候也存在R点。只有少数种类的细胞不能通过R点从而停在G2期。

23
M 期 M期即细胞分裂期,真核细胞的细胞分裂主要包括两种方式,即有丝分裂(mitosis)和减数分裂(meiosis)。
遗传物质和细胞内其他物质分配给子细胞。

24
S期 DNA的复制是以复制单位来进行双向复制的; DNA复制与组蛋白合成同步·,组成核小体串珠 结构,新合成的组蛋白结合于滞后链上;
复制单位的打开有先后顺序,常染色质先于异染 色质复制,富含GC的区域先于富含AT的区域, 因此造成的DNA合成不同步。

25
细胞周期长短测定 ◆脉冲标记DNA复制和细胞分裂指数观察测定法 ◆流式细胞仪测定法(Flow Cytometry) ◆缩时摄像技术
可以得到准确的细胞周期时间及分裂间期和分裂期的准确时间。

26
细胞周期 n S M 1 G2

27
细胞周期同步化 ◆自然同步化,如有一种粘菌的变形体plasmodia, 某些受精卵早期卵裂 ◆人工选择同步化 ◆药物诱导法
◆条件依赖性突变株在细胞周期同步化中的应用: 将与细胞周期调控有关的条件依赖性突变株转移到限定条件下培养,所有细胞便被同步化在细胞周期中某一特定时期。

28
人工选择同步化 有丝分裂选择法:用于单层贴壁生长细胞。优点是 细 胞未经任何药物处理,细胞同步化效率高。缺 点是分离的细胞数量少。
有丝分裂选择法:用于单层贴壁生长细胞。优点是 细 胞未经任何药物处理,细胞同步化效率高。缺 点是分离的细胞数量少。 密度梯度离心法:根据不同时期的细胞在体积和重 量上存在差别进行分离。优点是方法简单省时,效 率高,成本低。缺点是对大多数种类的细胞并不适 用。

29
药物诱导法 DNA合成阻断法 ─ G1/S-TdR双阻断法:最终将细胞群阻 断于G1/S交界处。优点是同步化效率高,几乎适合于所 有体外培养的细胞体系。缺点是诱导过程可造成细胞非均 衡生长。 分裂中期阻断法:通过抑制微管聚合来抑制细胞 分裂器 的形成,将细胞阻断在细胞分裂中期。优点是操作简便, 效率高。缺点是这些药物的毒性相对较大。

30
特异的细胞周期 ◆爪蟾早期胚胎细胞的细胞周期 特异的细胞周期是指那些特殊的细胞所具有的与标准的细胞周期相比有着鲜明特点的细胞周期。
◆酵母细胞的细胞周期 ◆植物细胞的细胞周期 ◆细菌的细胞周期

31
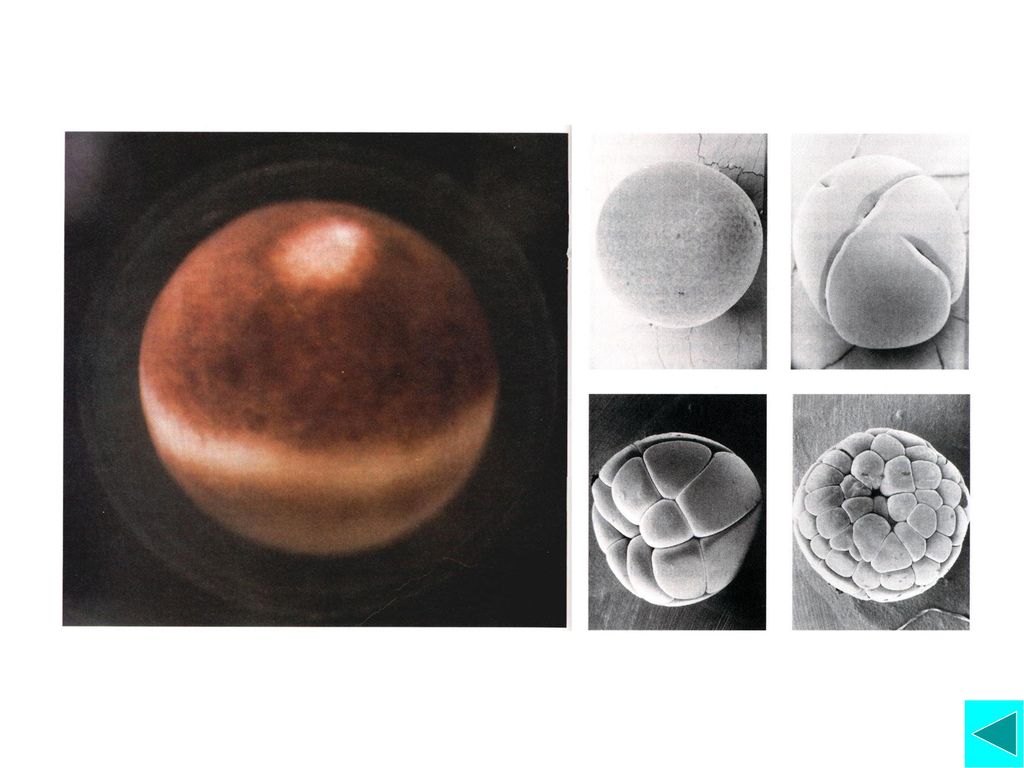
爪蟾早期胚胎细胞的细胞周期 细胞分裂快,无G1期, G2期非常短,S期也短(所有复 制子都激活), 以至认为仅含有S期和M期无需临时 合成其它物质。 子细胞在G1、G2期并不生长,越分裂体积越小。 细胞周期调控因子和调节机制与一般体细胞标准的 细胞周期基本是一致的。

32
酵母细胞的细胞周期 酵母细胞的细胞周期与标准的细胞周期在时相和调控方 面相似,常用种类有芽殖酵母和裂殖酵母 酵母细胞周期明显特点:
酵母细胞的细胞周期与标准的细胞周期在时相和调控方 面相似,常用种类有芽殖酵母和裂殖酵母 酵母细胞周期明显特点: 酵母细胞周期持续时间较短; 封闭式细胞分裂 ,即细胞分裂时核膜不解聚; 纺锤体位于细胞核内; 在一定环境下,也进行有性繁殖

35
植物细胞的细胞周期 植物细胞的细胞周期与动物细胞的标准细胞周 期非常相似,含有G1期、S期、G2期和M期四 个时期。
植物细胞不含中心体,但在细胞分裂时可以正 常组装纺锤体。 植物细胞以形成中板的形式进行胞质分裂

36
细菌的细胞周期 慢生长细菌细胞周期过程与真核细胞周期过 程有一定相似之处。其DNA复制之前的准备 时间与G1期类似。分裂之前的准备时间与G2 期类似。再加上S期和M期,细菌的细胞周期 也基本具备四个时期。 细菌在快速生长情况下,如何协调快速分裂和最基本的DNA复制速度之间的矛盾。

37
前期(prophase) 标志前期开始的第一个特征是染色质开始浓缩 (condensation)形成有丝分裂染色体(mitotic chromosome) 第二个特征细胞骨架解聚,有丝分裂纺锤体 (mitotic spindle)开始装配 Golgi体、ER等细胞器解体,形成小的膜泡

38
这种染色体由两条染色单体(chromatid)构成
在前期末,染色体主缢痕部位形成一种蛋白复 合物称为动粒(kinetochore)
")
39
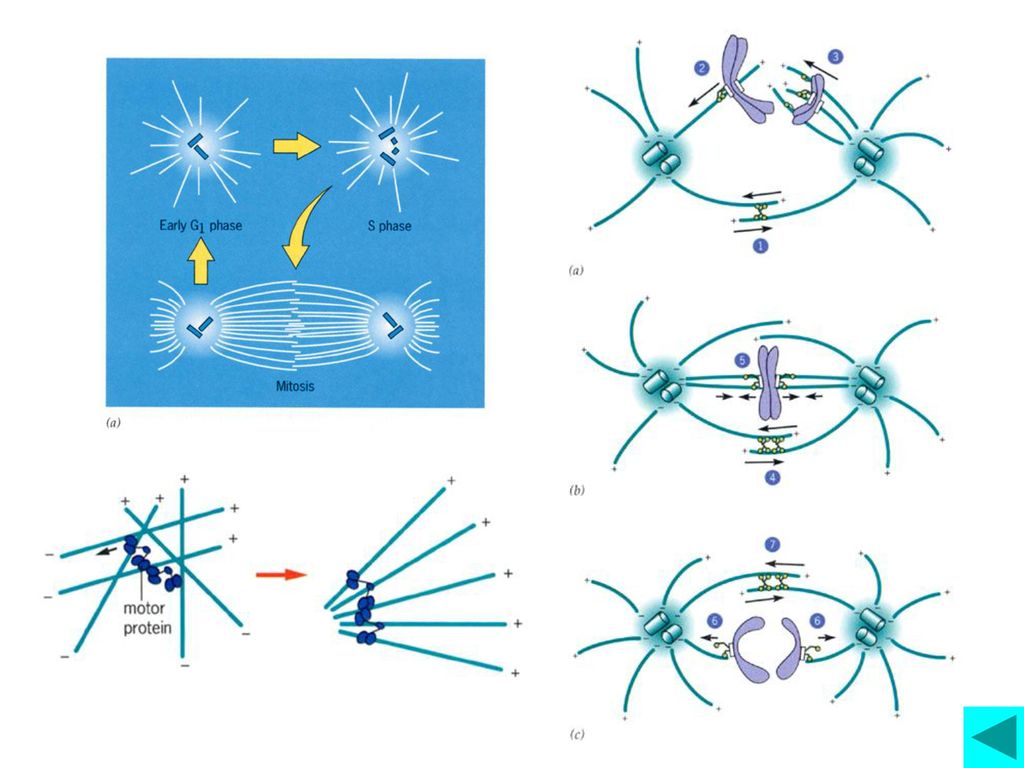
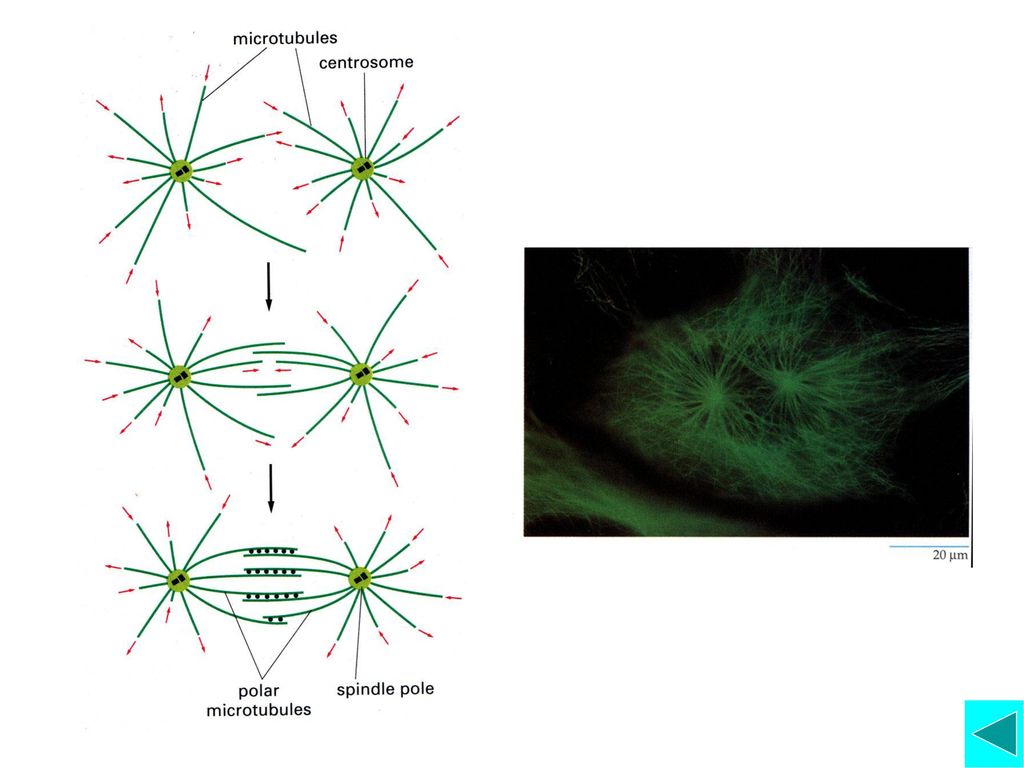
间期动物细胞含一个MTOC,即中心体,在S期末,两个中心粒在各自垂直的方向复制出一个中心粒,形成两个中心体。当前期开始时,2个中心体移向细胞两极,并同时组织微管生长,由两极形成的微管通过微管结合蛋白在正极末端相连,最后形成有丝分裂纺锤体。

40
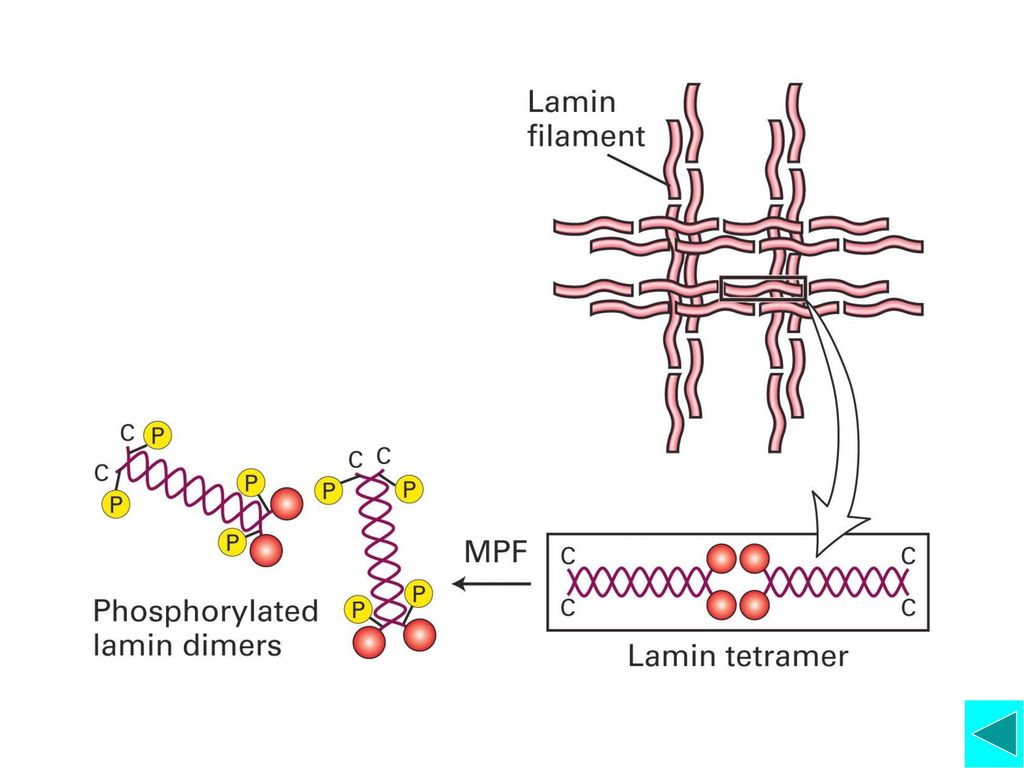
前中期(prometaphase) ◆核膜破裂成小的膜泡,这一过程是由核纤层蛋白中特 异的Ser残基磷酸化导致核纤层解体
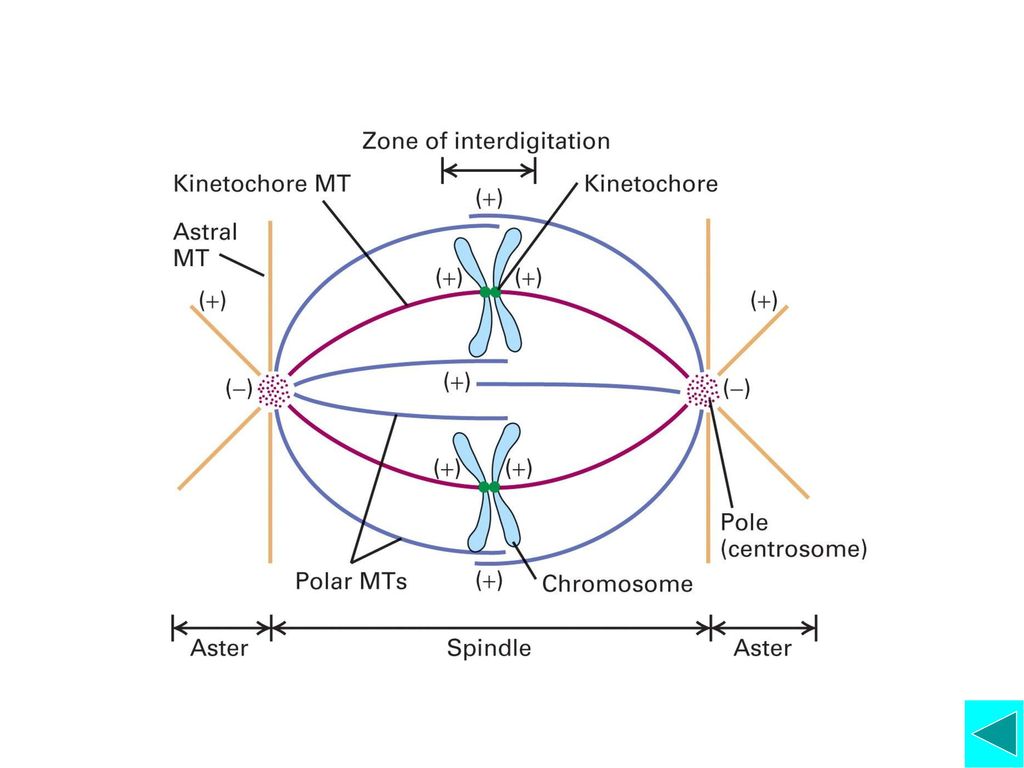
◆纺锤体微管与染色体的动粒结合,捕捉住染色体每个 已复制的染色体有两个动粒,朝相反方向,保证与两极 的微管结合;纺锤体微管捕捉住染色体后,形成三种类 型的微管 ◆不断运动的染色体开始移向赤道板。细胞周期也由前 中期逐渐向中期运转。

41
中期(metaphase) ◆所有染色体排列到赤道板(Metaphase Plate) 上,标志着细胞分裂已进入中期
◆是什么机制确保染色体正确排列在赤道板上? 着丝粒微管动态平衡形成的张力

42
后期(anaphase) 排列在赤道面上的染色体的姐妹染色单体分离产生 向极运动
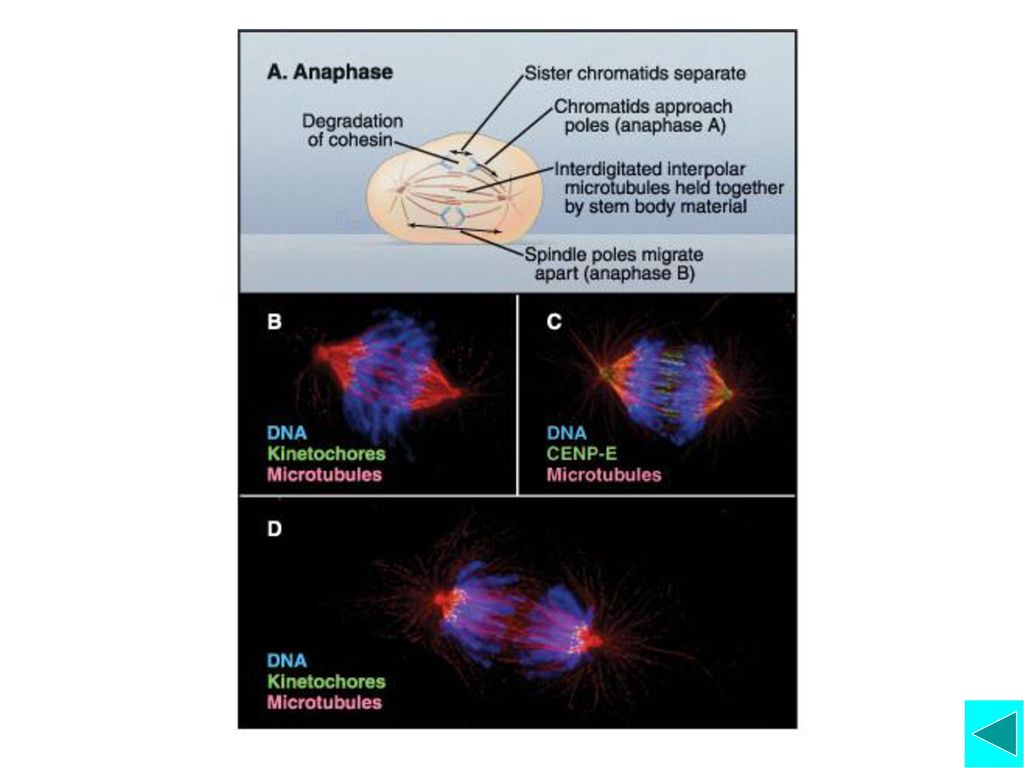
排列在赤道面上的染色体的姐妹染色单体分离产生 向极运动 后期(anaphase)大致可以划分为连续的两个阶段, 即后期A和后期B ·后期A,动粒微管去装配变短,染色体产生两极运动 ·后期B,极间微管长度增加,两极之间的距离逐渐拉长, 介导染色体向极运动
大致可以划分为连续的两个阶段, 即后期A和后期B. ·后期A,动粒微管去装配变短,染色体产生两极运动. ·后期B,极间微管长度增加,两极之间的距离逐渐拉长, 介导染色体向极运动.")
43
末期(telophase) 染色单体到达两极,即进入了末期(telophase), 到达两极的染色单体开始去浓缩 核膜开始重新组装
Golgi体和ER重新形成并生长 核仁也开始重新组装,RNA合成功能逐渐恢复,有丝 分裂结束

44
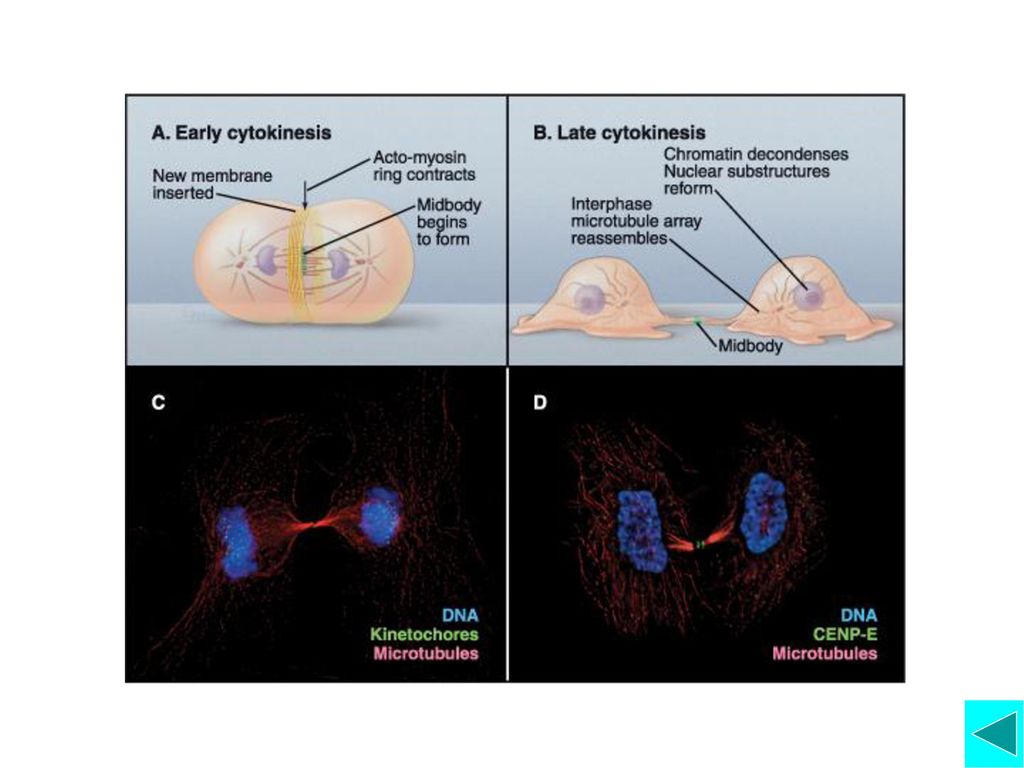
动物细胞胞质分裂 胞质分裂(cytokinesis)开始于细胞分裂后期,在赤 道板周围细胞表面下陷,形成环形缢缩,称为 分 裂沟(furrow)。分裂沟的位置与纺锤体极性微管和 钙离子浓度升高的变化有关。 胞质分裂开始时,大量肌动蛋白和肌球蛋白在中体处组装成微丝并相互组成微丝束,环绕细胞,称为收缩环(contractile ring)。收缩环收缩、收缩环处细胞膜融合并形成两个子细胞。
。收缩环收缩、收缩环处细胞膜融合并形成两个子细胞。")
45
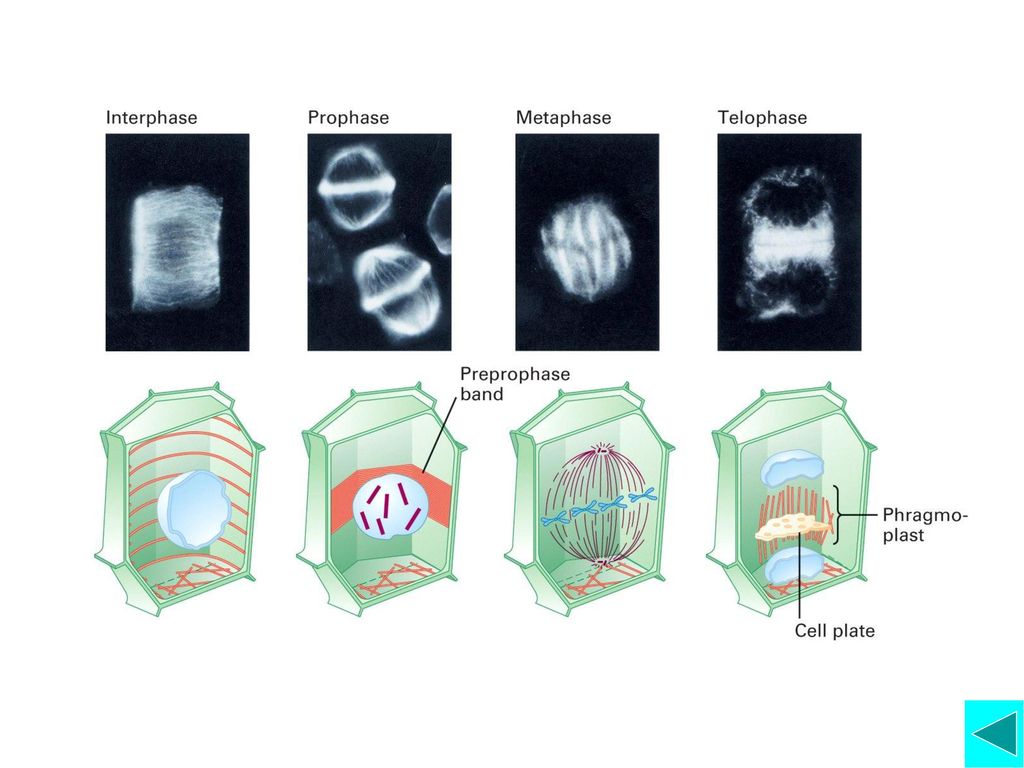
植物细胞胞质分裂 与动物细胞胞质分裂不同的是,植物细胞胞质分裂是因为在细胞内形成新的细胞膜和细胞壁而将细胞分开。

46
减数分裂概念与过程 概念:减数分裂是细胞仅进行一次DNA复制, 随后进行两次分裂,染色体数目减半的一种特 殊的有丝分裂 减数分裂过程

47
减数分裂的意义 确保世代间遗传的稳定性; 增加变异机会,确保生物的多样性,增强生物适应环境变化的能力。
减数分裂是生物有性生殖的基础,是生物遗传、生物进化和生物多样性的重要基础保证。

48
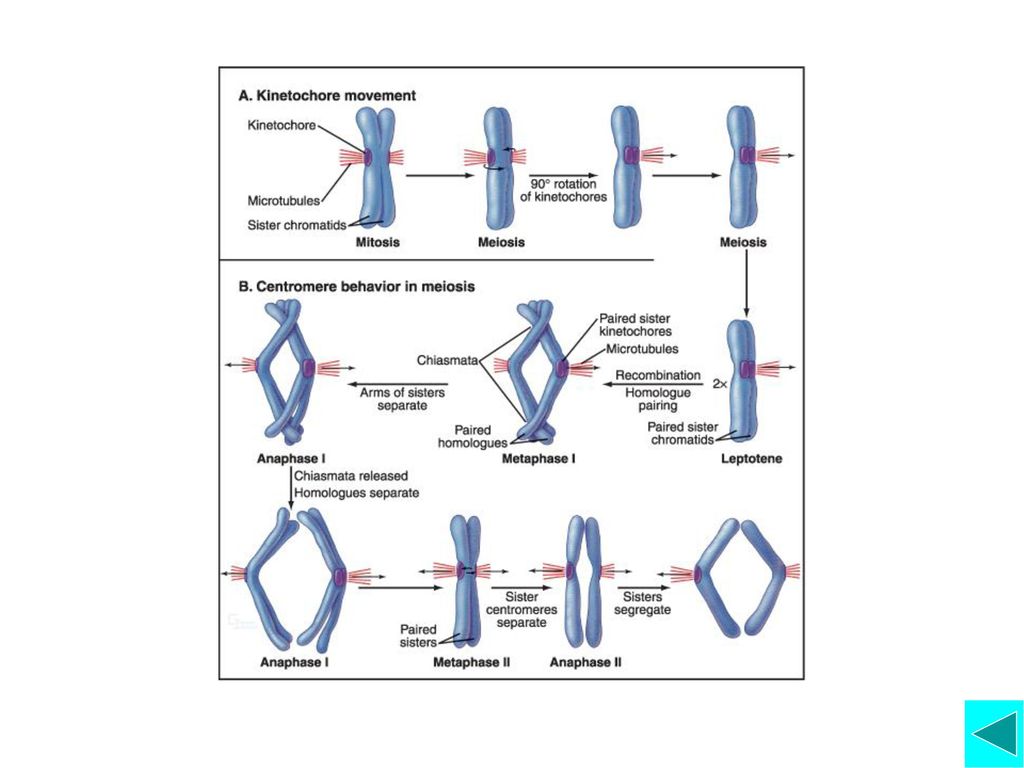
减数分裂各时相及其主要事件 间期中S期持续时间较长 第一次减数分裂 前期I分为细线期,偶线期,粗线期,双线期, 终变期等五个阶段
第二次减数分裂

50
细线期 染色质凝集,但是两条染色单体的臂并不分离; 在细纤维样的染色体上,出现颗粒状的染色粒 染色体端粒通过接触斑与核膜相连

51
偶线期 偶线期同源染色体配对形成联会复合体 (Synaptonemal Complex, SC)同源染色体配对 的过程称为联会;
合成在S期未合成的0.3%的DNA(偶线期DNA,即zDNA)
")
52
粗线期 发生等位基因之间部分DNA片段的交换和重组 合成一小部分DNA,称为P-DNA,这些DNA编码一些与DNA点切和修复相关的酶类。
合成减数分裂期专有的组蛋白,并将体细胞类型的组蛋白置换下来。

53
双线期 同源染色体相互分离,仅有几入相互联系同源染色体间遗传物质重组,产生新的基因组合; 同源染色体或多或少地发生去凝集,RNA转录活跃;

54
终变期 染色体凝集成棒状结构; 交叉向染色体臂的端部移行,这个过程称为端化。

55
减数分裂前S期与有丝分裂前S期长度比较 减数分裂前S期 有丝分裂前S期 蝾螈 10天 12小时 小鼠 14小时 5---6小时 小麦
3.8小时 酵母 1小时 0.5小时

56
一、细胞周期调控系统的主要作用 在适当时候激活细胞周期各个时相的相关酶和蛋白,确保细胞周期向下一个阶段进行,然后以某种方式失活(正调控)。
当细胞周期中出现异常时,细胞会启动一系列的负调控机制,使细胞停滞在某个特定的时期,这种调节机制可确保每一时相事件的全部完成。 外界环境因子(如多细胞生物对增殖信号的反应) 通过酪氨酸蛋白激酶受体偶联的信号通路完成信号向细胞内的传递。
 通过酪氨酸蛋白激酶受体偶联的信号通路完成信号向细胞内的传递。")
57
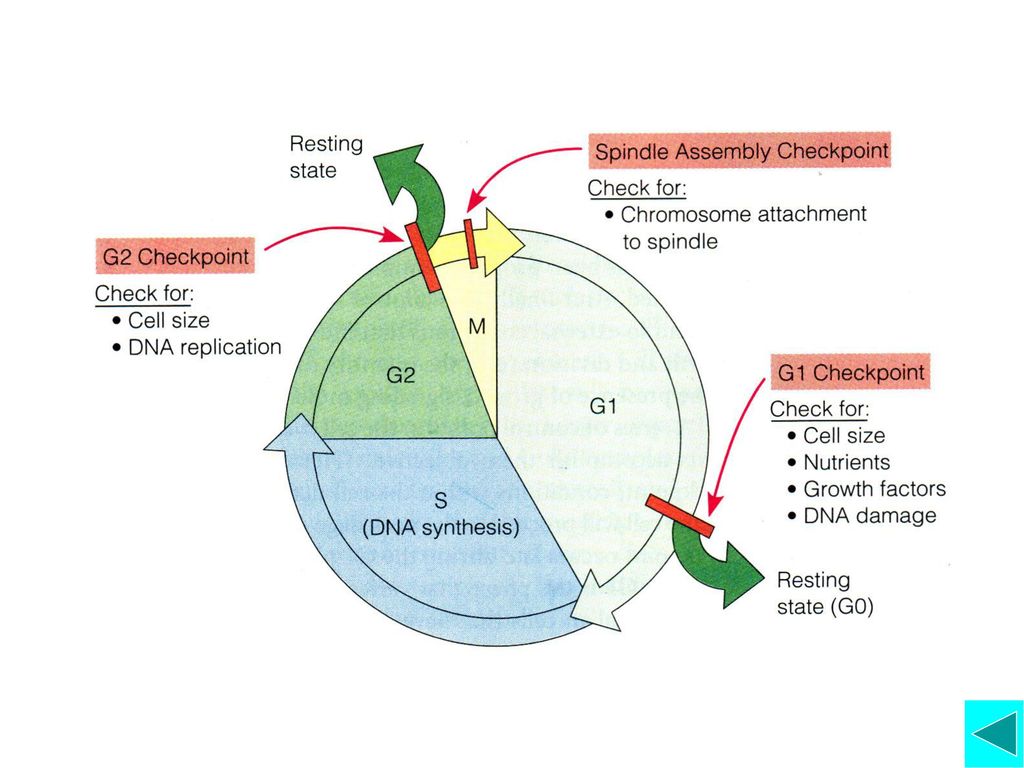
二、细胞周期检验点(checkpoint)
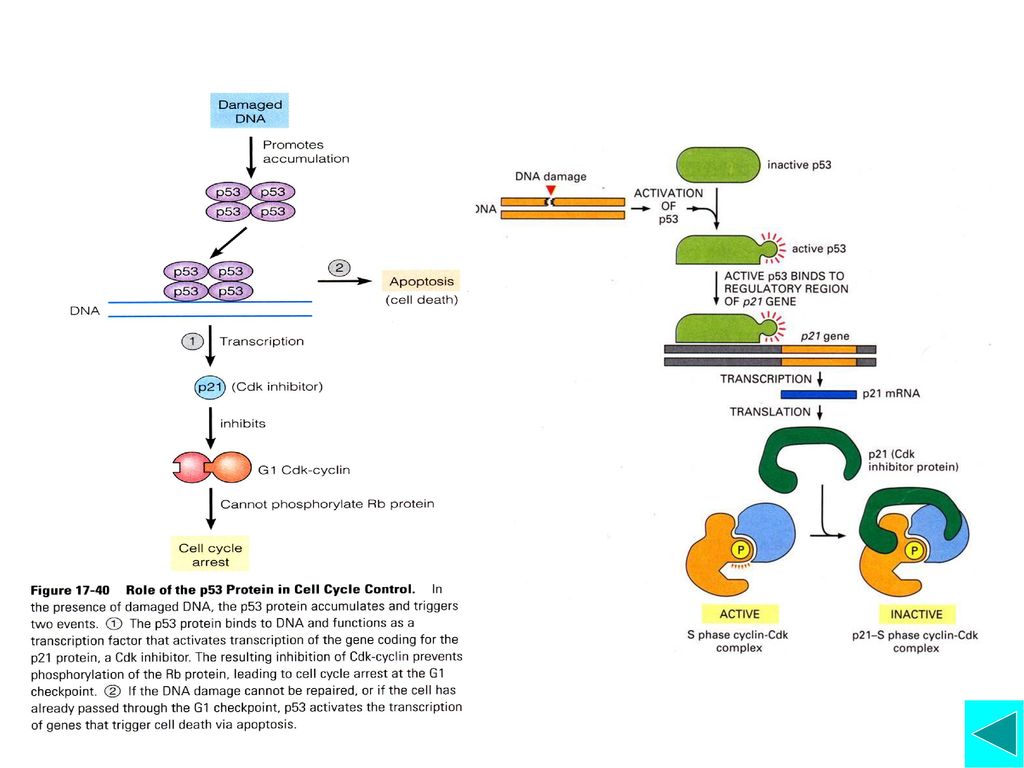
细胞周期检验点是细胞周期调控的一种机制,主要是确保周期每一时相事件的有序、全部完成并与外界环境因素相联系 细胞周期检验点及其作用 G1期检验点: 酵母——Start; 动物细胞——Restriction Point

58
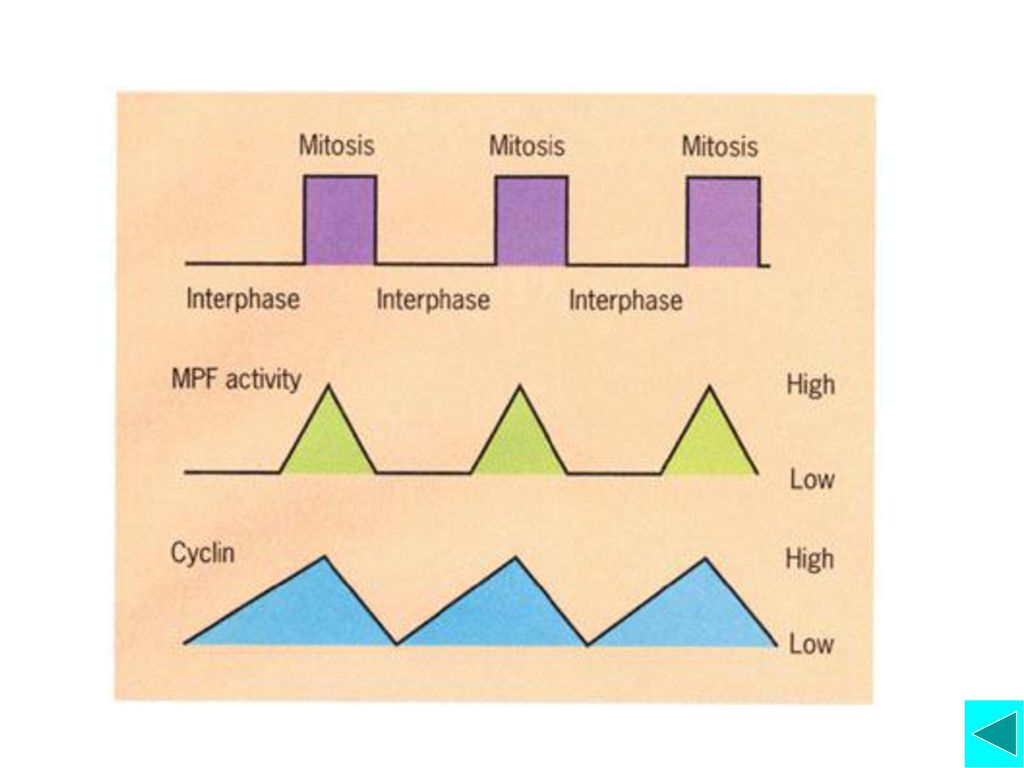
MPF( Maturation-promoting factor)
周期蛋白 CDK Mitotic Cyclin-Cdk(MPF)的活化与功能 2001年Nobel Prize M&P
的活化与功能. 2001年Nobel Prize M&P.")
59
2001年Nobel Prize M&P授予在细胞周期关键调控因子中作出突出贡献的三位科学家
L. Hartwell,发现芽殖酵母中控制细胞周期的cdc28基因,并于1988年提出检测点的概念 P. Nurse,发现裂殖酵母中控制细胞周期的cdc2基因,并提出了CDK的概念 Tim Hunt,发现了细胞周期蛋白cyclin

60
周期蛋白 人们从不同生物中分离克隆出了数十种周期蛋白,这些周期蛋白不细胞内表达的时期有所不同。
G1期表达并起作用的称为G1期周期蛋白,主要有E和D;M期表达并起作用的称为M期周期蛋白,主要有B和A。 各种周期蛋白之间的共性: 1、含有一段保守序列,称为周期蛋白框,这一序列是它和催化亚基结合的位点; 2、A和B在N端具有破坏框,其余分子在C端具有PEST序列,这些均与周期蛋白的降解相关。

62
CDK 人们从不同物种成功分离到了十多个cdc相关基因,它们编码的产物共同之处在于: 含有一段类似的AA序列
都可以与周期蛋白结合并将周期蛋白作为其调节亚单位,进而表现出蛋白激酶的活性。 这些蛋白被称为周期蛋白依赖性蛋白激酶(cyclin-dependent kinase),简称CDK。由于cdc2是第一个发现的cdc蛋白,因此被命名为CDK1。
,简称CDK。由于cdc2是第一个发现的cdc蛋白,因此被命名为CDK1。")
63
在MPF的发现过程中的重要事件 Johnson和Rao进行的实验及其发现 Mausi利用爪蟾卵母细胞进行的实验及MPF的提出
Hartwell等人利用芽殖酵母进行的实验及其发现 P. Nurse等人利用裂殖酵母进行的实验及其发现 Tim Hunt利用海胆卵细胞进行的实验及其发现 结论

64
Hartwell等利用芽殖酵母进行的实验
实验是利用温度敏感的芽殖酵母为材料来进行的。在正常温度下这些突变体可以正常分裂增殖,但是当温度升高时就不能进行正常的分裂,这是由于在这些温变突变体内缺少某个特定的基因而引起的。 这些基因是与细胞分裂和细胞周期调控有关的基因,称为cdc(cell diversion cycle)基因。 在芽殖酵母中分离得到的cdc基因是cdc 28,它的表达产物为p34cdc28,它是一种蛋白激酶。 cdc 28突变可使细胞停在G1/S交界处或G2/M交界处。
基因。 在芽殖酵母中分离得到的cdc基因是cdc 28,它的表达产物为p34cdc28,它是一种蛋白激酶。 cdc 28突变可使细胞停在G1/S交界处或G2/M交界处。")
68
Johnson和Rao进行的实验及其发现
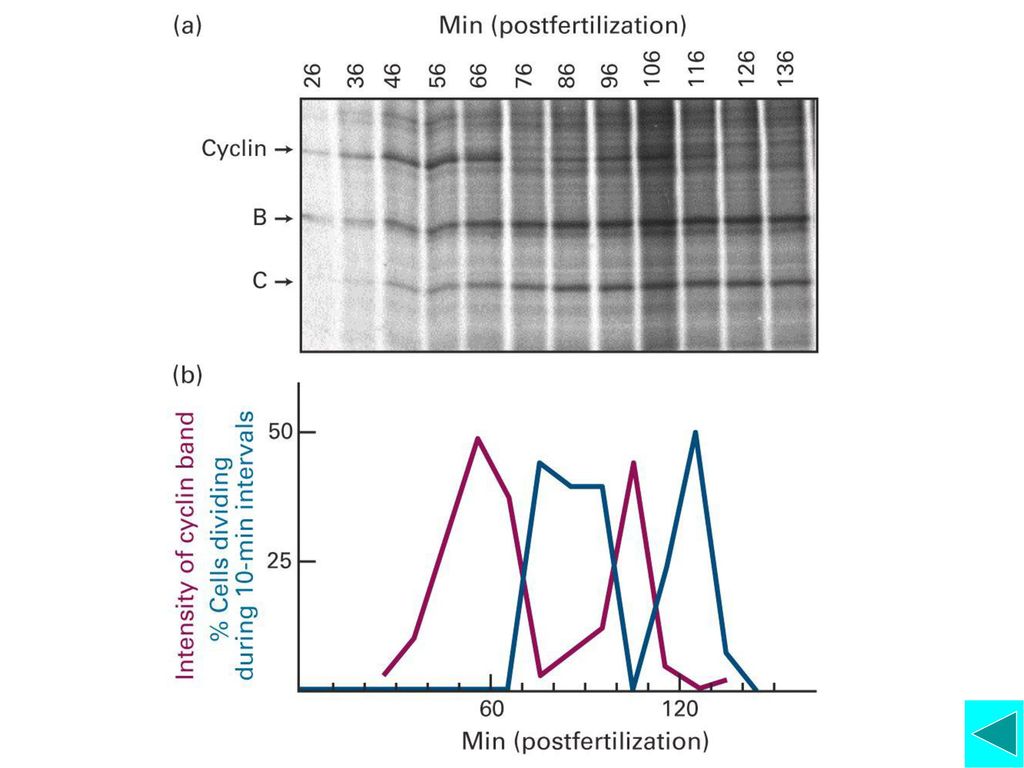
对间期的细胞进行同步化后进行细胞融合 G1+S S+G G1+G2 结论:在S期的细胞质中存在有刺激G1期细胞进入S期的某种因子,定名为SPF。 将处于间期不同时期的细胞与M期细胞进行融合,观察到特异的提前凝集染色体PCC 结论:在M期细胞中存在有诱导染色体凝集的因子,称为M期促分裂因子。

69
P. Nurse利用裂殖酵母进行的实验 他进行的实验和Hartwell的实验相似,只是实验材料换成了裂殖酵母的温度敏感的突变株。
他从中分离得到了第一个cdc 基因--- cdc 2。它的表达产物为p34cdc2。 cdc 2基因的突变可使细胞停在G2/M交界处。 P34 cdc 2 (或 P34 cdc 28)必须与p56 cdc 13结合后才表现出激酶的活性。
必须与p56 cdc 13结合后才表现出激酶的活性。")
70
Tim Hunt利用海胆卵细胞进行的实验 他在实验中发现,在海胆的卵细胞中存在两种特殊类型的蛋白质,这两种蛋白质的含量随细胞周期的进程变化而变化,这种蛋白质被命名为细胞周期蛋白(cyclin)。
。")
72
比较这几个实验得出的结论 MPF的p32亚基与酵母中的p34cdc2为同源产物,而MPF中的p45与p56 cdc 13和周期蛋白B是同源物。 p32= p34cdc2=CDK(催化亚基) MPF p45=cyclin= p56 cdc 13 (调节亚基)
")
73
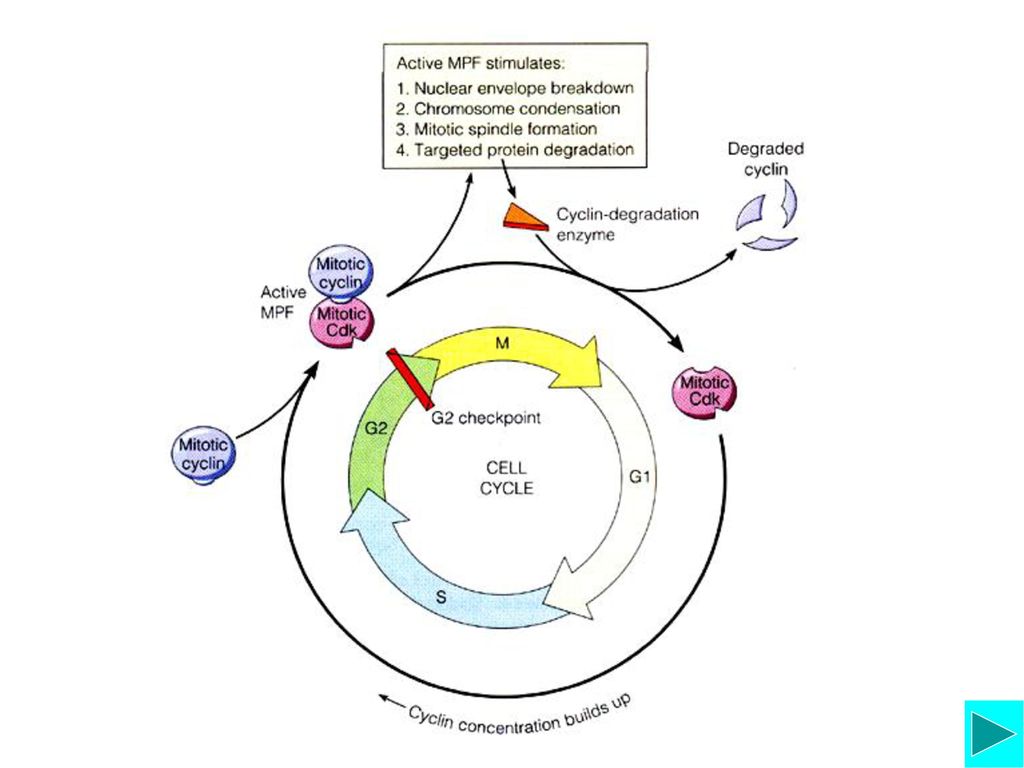
MPF是一种使多种底物蛋白磷酸化的蛋白激酶;由M期Cyclin-Cdk ( Cyclin-dependent protein kinase)形成的复合物。MPF=(CDK1=p34cdc2)+cyclinB
形成的复合物。MPF=(CDK1=p34cdc2)+cyclinB")
74
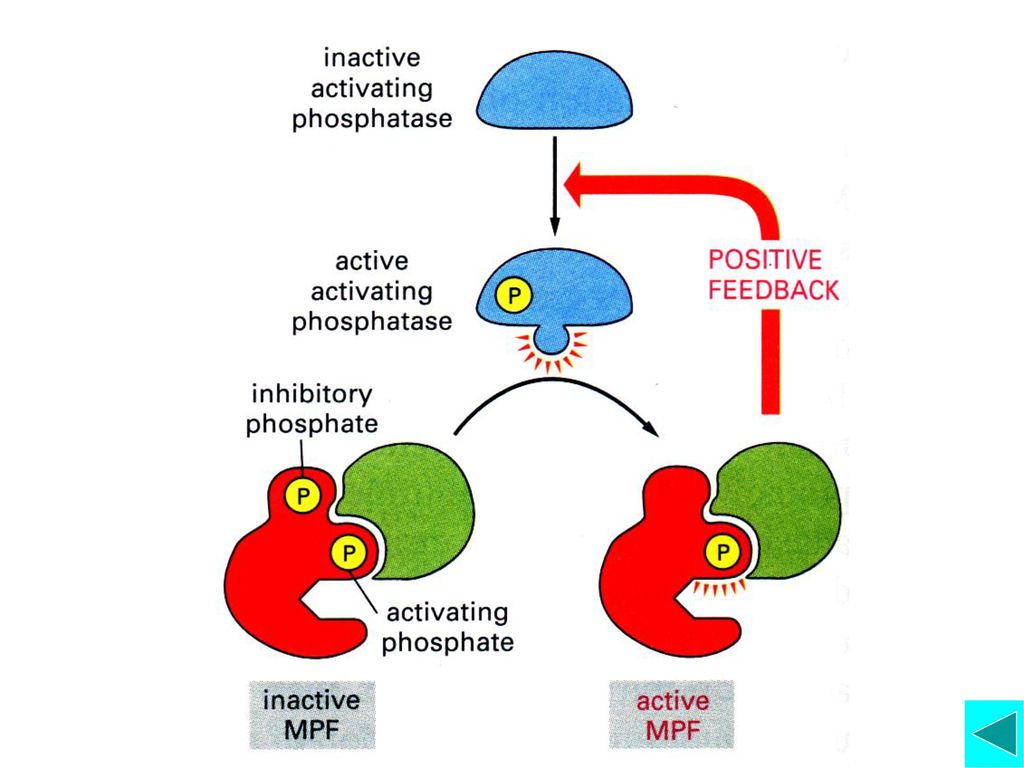
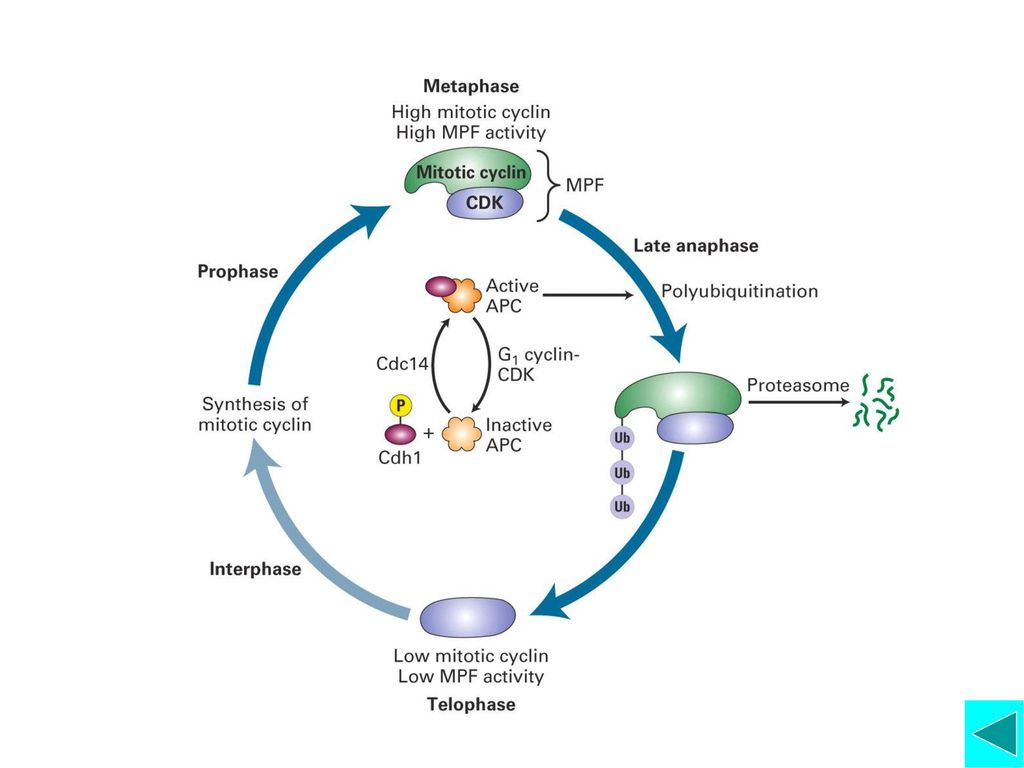
Mitotic Cyclin-Cdk(MPF)的活化与功能
激酶与磷酸酶的调节 活化的MPF可使更多的MPF活化 Cyclin的降解 CDK激酶抑制蛋白的作用 功能:启动细胞从G2期进入M期的相关事件

75
四、Cyclin-Cdk复合物的多样性及细胞周期运转
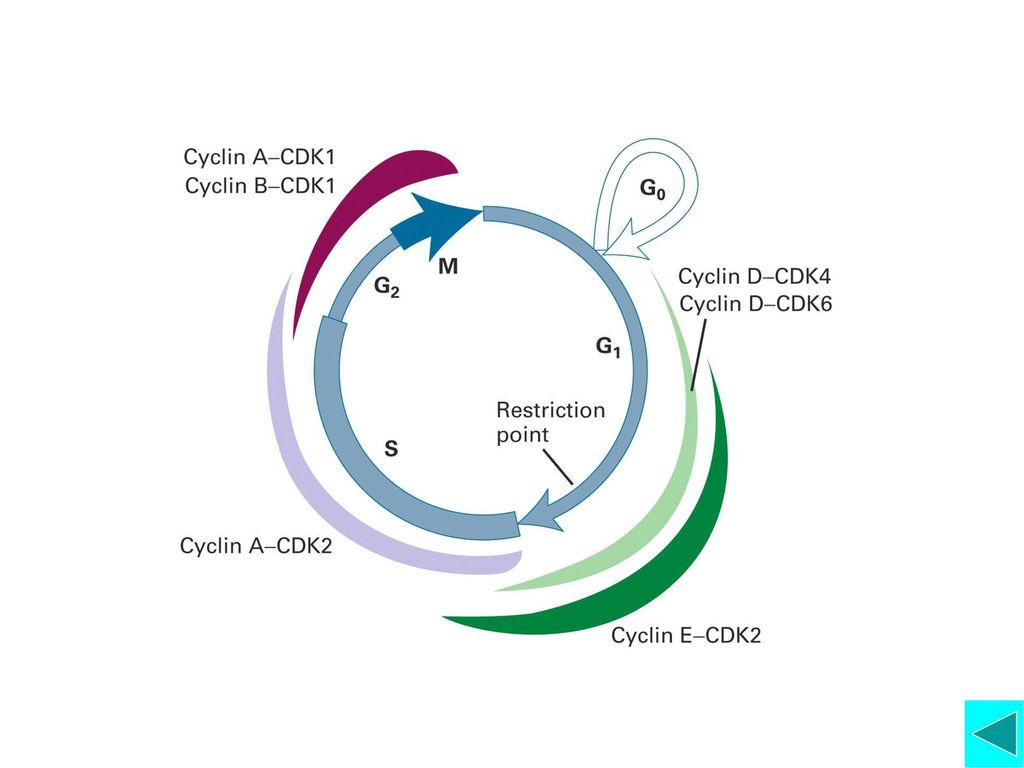
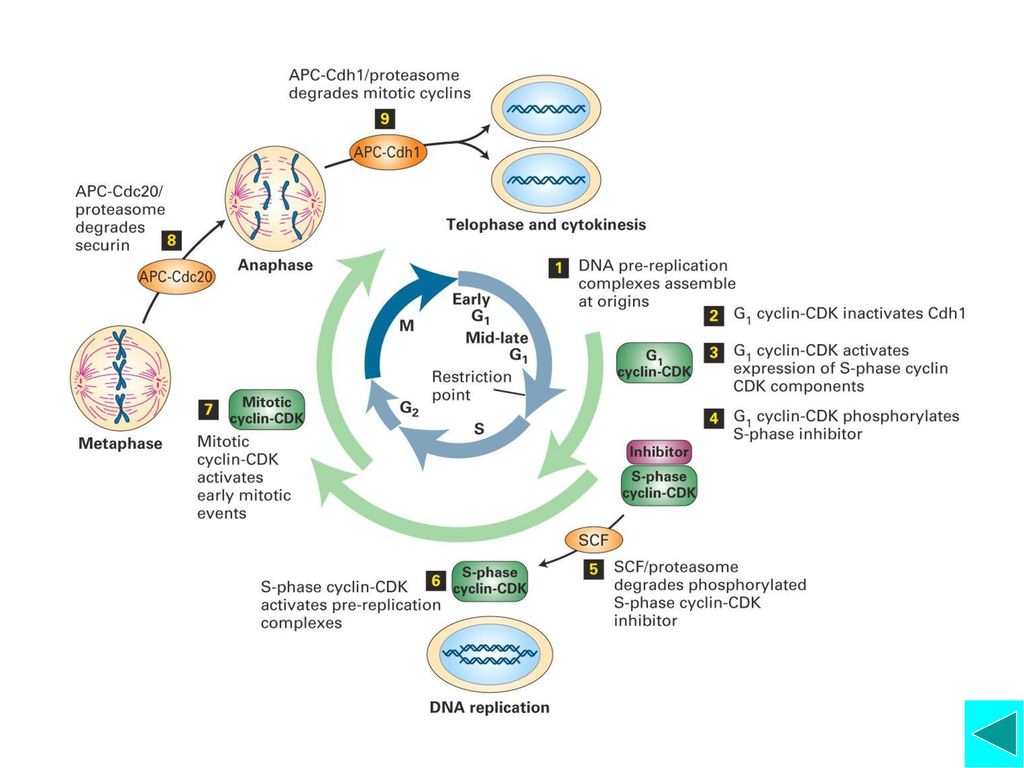
Cyclin-Cdk是调控细胞周期的引擎:不同的周期蛋白与不同的CDK结合,构成不同的Cyclin-Cdk;不同的Cyclin-Cdk在不同的时相表现活性,影响不同的下游事件。 G1/S转化 G2/M转化(MPF的作用) M期周期蛋白与中期向后期的转化 Anaphase Promoting Complex (APC), 细胞周期调控模型总结
 M期周期蛋白与中期向后期的转化. Anaphase Promoting Complex (APC), 细胞周期调控模型总结.")
77
Cyclin-Cdk复合物的多样性 G1 S G2/M Cyclin-Cdk Cyclin-Cdk Cyclin-Cdk
Budding Yeast CLN1,2,3-CDC CLB5,,(3,4)-CDC28 CLB1,2(3,4)-CDC28 Fission Yeast CIG1-CDC CIG2-CDC CIG13-CDC2 Higher Eukaryotes CyclinD1,2,3-CDK4/6 CyclinA-CDK CyclinB-cdc2/CDK1 CyclinE1,2-CDK2 G1 Substrates S Substrates G2/M Substrates Growth and Morphogenesis DNA Replication Mitosis
-CDC28 CLB1,2(3,4)-CDC28. Fission Yeast CIG1-CDC2 CIG2-CDC2 CIG13-CDC2. Higher Eukaryotes CyclinD1,2,3-CDK4/6 CyclinA-CDK2 CyclinB-cdc2/CDK1. CyclinE1,2-CDK2. G1 Substrates. S Substrates. G2/M Substrates. Growth and. Morphogenesis. DNA Replication. Mitosis.")
78
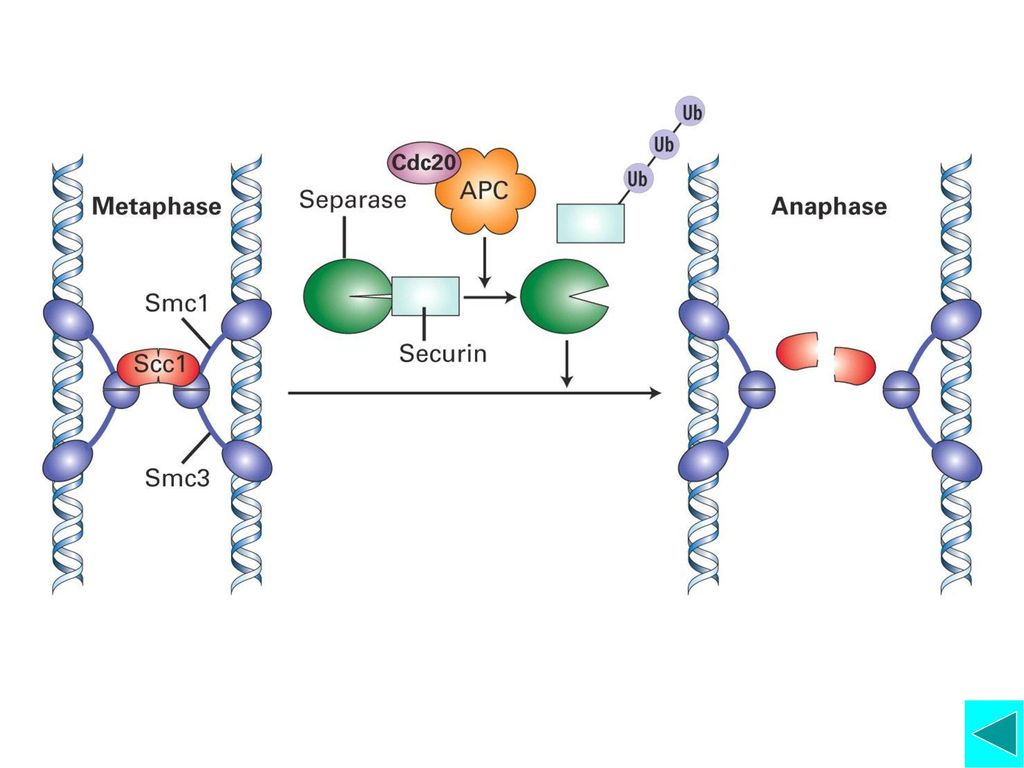
APC的活化:Cdc20是APC的活化因子。Cdc20和Mad2蛋白位于动粒上,在染色体结合有丝分裂纺锤体前将不能从动粒上释放,由于Mad2与Cdc20结合而抑制APC的活性。所以只有所有染色体都与纺锤体结合后,APC才有活性,才启动细胞向后期转换。 APC主要介导两类蛋白降解: Anaphase Inhibitors(后期抑制因子)和Mitotic Cyclin。前者维持姐妹染色单体粘连, 抑制后期启动;后者的降解意味着有丝分裂即将结束,即染色体开始去凝集,核膜重建。 APC介导选择性降解的靶蛋白与Ubiquitin结合
和Mitotic Cyclin。前者维持姐妹染色单体粘连, 抑制后期启动;后者的降解意味着有丝分裂即将结束,即染色体开始去凝集,核膜重建。 APC介导选择性降解的靶蛋白与Ubiquitin结合.")
80
五、细胞周期运转的阻遏 细胞至少可通过两种不同机制阻遏细胞周期的运转:Cdk抑制蛋白(CDI)阻止Cyclin-Cdk复合物的装配或活 性;周期调控系统组分停止合成。 CDI包括CIP/KIP家族和INK4家族,其作用是抑制Cyclin-Cdk复合物的装配或活性,而将细胞阻止在不同的检验点。如DNA受损后,细胞将停留于G1 Checkpoint 让DNA修复或者凋亡 周期调控系统组分停止合成,如G0细胞,大部分Cyclin和Cdk都消失,这在多细胞生物尤其明显。

82
细胞类型 细胞周期时间 两栖类早期胚胎细胞 30分钟 酵母细胞 1.5---3小时 小肠上皮细胞 约12小时 体外培养的哺乳类 上皮细胞
约20小时 人类肝细胞 约1年

83
Experimental demonstration of the coordinated
Synthesis of DNA and hitones.

93
.

102
a. Electron micrograph of SC of human pachytene bivalent
a. Electron micrograph of SC of human pachytene bivalent. (K: kinetochore; arrow: recombination nodules); b. Schematic diagram of SC; c. Electron micrograph of SC after treatment with DNase to remove chromasomal fibers.
; b. Schematic diagram of SC; c. Electron micrograph of SC after treatment with DNase to remove chromasomal fibers.")
104
Visible evidence of crossing over

107
人M期细胞与袋鼠(Ptk)G1、S、G2期细胞融合诱导PCC:提示M期细胞存在诱导PCC的因子;
G1、S、G2期细胞融合诱导PCC:提示M期细胞存在诱导PCC的因子;")
108
处于第六期的爪蟾卵母细胞(RD前期I),具GV。
,具GV。")
109
注射实验表明:孕酮诱导卵母细胞成熟;成熟卵细胞质中,含有卵母细胞成熟的因子,称做MPF。 1988年Maller实验室分离纯化了MPF,证明它主要含有p32和p45两种蛋白,二者结合后,表现出蛋白激酶的活性。

111
CAK=CDK1-Activiting Kinase

Similar presentations

























:核蛋白体组成成分 转移 RNA ( tRNA ):转运氨基酸 信使 RNA ( mRNA ):蛋白质合成模板 不均一核 RNA ( hnRNA ):成熟 mRNA 的前体 小核 RNA ( snRNA ):>")



 二中 锋.>")


 第四单元:细胞的增殖与分化 第十二讲:细胞的增殖.>")


的意义 ●细胞周期与细胞分裂 ●细胞周期调控.>")
 减数分裂(Meiosis)>")




