Download presentation
Presentation is loading. Please wait.

1
欢 迎

2
是什么在修复你的DNA ------2015诺贝尔化学奖 http://www.guokr.com/article/440793/
瑞典科学家托马斯-林达尔、美国保罗-莫德里奇、土耳其阿齐兹-桑贾尔、 Lindahl, Tomas. "My journey to DNA repair." Genomics, proteomics & bioinformatics 11.1 (2013): 2-7.
:")
3
第二节 DNA损伤和修复

4
Transposons 转座子 Chemicals 化学物质 Radiation 辐射 Replication errors 复制错误
DNA is an unusually stable molecule, BUT it is not invincible. Transposons 转座子 Chemicals 化学物质 Radiation 辐射 Replication errors 复制错误 . 4

5
二、 损伤原因 (一)内源性损伤 (1) DNA复制错误 (2) 碱基互变异构体 (3) 自发的化学变化:脱嘌呤;脱氨基
(4) 氧化作用损伤碱基
 氧化作用损伤碱基.")
6
(1) DNA复制错误 复制过程中,A与C配对,这样在复制后该位点产生A-T和G-C两种配对模式,产生突变。
聚合酶漏校的结果,出现的频率较低。 碱基配对的错误频率约为 ; DNA聚合酶本身具有校对作用:将不正确插 入的核苷酸切除掉,重新加上正确的核苷酸。 这样,每掺入一个核苷酸,发生错误的机会 有 。

7
1.1 Mismatched Base Mistakes during DNA replication are a common source of transitions and transversions. DNA polymerase occasionally pairs a base on the template strand with a non-complementary base. Usually, the mistake is recognized and removed using the protein’s 3’5’ exonuclease activity. However, the error is occasionally left unfixed, creating an abnormality called a mismatched base or a DNA mismatch. DNA复制过程中发生错误是转换和颠换的常见来源。DNA聚合酶偶尔会配上一个与模板链上的碱基不互补的碱基。通常,这样的错误能被该酶的3’5’外切核酸酶活性识别并加以去除。然而,这样的错误偶尔会留在DNA上从而造成一种异常情况,这被称为错配碱基或DNA错配。 d 7

8
Mismatched base → Point mutation
如果这样的错配碱基没有被修复,最终它将发展成一种突变。在下一轮复制中,这一不正确的碱基出现在一条模板链上,与它互补的碱基会被加到新链中去。虽然就这一碱基对而言没有任何结构上的错误,但它已经与基因中原先那一正常的碱基对不同了。 8

9
(2) 碱基互变异构体 碱基异构式引起DNA复制过程的错误 碱基异构式: A(amino 氨基) A(imino 亚氨基)
C(a) C(i) G(keto 酮式) G(enol 烯醇式) G(k) G(e,i) T(keto) T(enol-2’) or T(enol-4’)
 C(i) G(keto 酮式) G(enol 烯醇式) G(k) G(e,i) T(keto) T(enol-2’) or T(enol-4’)")
10
Tautomeric forms of the DNA bases
我们所熟悉的含氮碱基恰巧也是互变异构体。每种碱基更稳定的形式是我们前面学习过的结构。然而,它们也可以互相转换成其它结构。A和C可以互变异构化成它们的亚氨基式,通常写成A*和C*。T和G可以互变异构化成它们的烯醇式T*和G*。这些另类碱基具有非常不一样的氢键成键特性:A*与C结合,C*与A结合,T*与G结合,G*与T结合。虽然这些碱基的正常形式以大约10,000:1的比例得到偏爱,但是如果这种互变异构化发生在复制过程中,那么它不管是发生在模板链上也好还是发生在将要加上去的碱基中也好,它都会导致新合成链中整合进一个不正确碱基。当这一碱基回复到它的正常形式后,将出现一个DNA错配。 10

11
(3) 自发的化学变化 碱基的脱氨基(deamination)作用
碱基的环外氨基有时会自发脱落,从而胞嘧啶会变成尿嘧啶、腺嘌呤会变成次黄嘌呤(H)、鸟嘌呤会变成黄嘌呤(X)等。 胞嘧啶自发脱氨基的频率约为每个细胞每天190个。
、鸟嘌呤会变成黄嘌呤(X)等。 胞嘧啶自发脱氨基的频率约为每个细胞每天190个。")
13
G – C → A – T Replication C→U U
脱氨基即从一个核苷酸上去除一个氨基,是一种与水发生的简单反应。当脱氨基发生在胞嘧啶上时,产生的是尿嘧啶。如同一个错配的碱基那样,如果这样的变化没有改正过来,那么每个尿嘧啶将在下一轮复制中引起一个突变。因为尿嘧啶与胸腺嘧啶非常相似,在DNA复制中它将与腺嘌呤配对。而在原始基因中,胞嘧啶与鸟嘌呤配对。这样,在基因中就引入了一个点突变。 13

14
脱嘌呤(depurination)与脱嘧啶
自发的水解可使嘌呤和嘧啶从DNA链的核糖磷酸骨架上脱落下来。 一个哺乳类细胞37℃,20h内DNA链自发脱落的嘌呤约1000个、嘧啶约500个, 长寿命不复制繁殖的哺乳类细胞(如神经细胞)在整个生活期间自发脱嘌呤数约为108,约占细胞DNA中总嘌呤数的3%。
在整个生活期间自发脱嘌呤数约为108,约占细胞DNA中总嘌呤数的3%。")
15
Depurinate ( > 5000 nt) Repair ( most )
G G C C Depurinate ( > 5000 nt) T T A A Depurination is the detachment of a purine base from the DNA backbone. Every day, approximately 5,000-10,000 nucleotides in a human cell’s genome lose a purine. 脱嘌呤即从DNA骨架上脱下一个嘌呤碱,一个人类细胞基因组每天大约有5,000-10,000个核苷酸失去它们的嘌呤。 Repair ( most ) C C G A A T T 15
 T. T. A. A. Depurination is the detachment of a purine base from the DNA backbone. Every day, approximately 5,000-10,000 nucleotides in a human cell’s genome lose a purine. 脱嘌呤即从DNA骨架上脱下一个嘌呤碱,一个人类细胞基因组每天大约有5,000-10,000个核苷酸失去它们的嘌呤。 Repair. ( most ) C. C. G. A. A. T. T. 15.")
16
Depurination → Mutation
G – C → A – T → C – G → T - A A C “How can I continue my work?” A 同样,这也能在下一轮复制中造成突变和其它困难。如果DNA聚合酶到达了一个没有碱基的核苷酸位置,它很可能会将一个错误的碱基加到合成链上,因为模板链不能为它提供任何信息。另一种可能结果是:复制随即停止并等待细胞采取旁路机理,而这也会引入突变。 DNA pol A T 16

17
4. 氧化作用损伤碱基 细胞呼吸的副产物O2、H2O2等会造成DNA损伤,能产生胸腺嘧啶乙二醇、羟甲基尿嘧啶等碱基修饰物,引起DNA单链断裂等损伤 每个哺乳类细胞每天DNA单链断裂发生的频率约为5万次。 DNA的甲基化、结构的其他变化等,这些损伤的积累可能导致老化。

18
(二)外源性损伤 物理因素:紫外线(ultra violet, UV)、 各种辐射 2. 化学诱变剂
外源性损伤 物理因素:紫外线(ultra violet, UV)、 各种辐射 2. 化学诱变剂")
19
1. 物理因素:由紫外线、电离辐射、X射线等引起的DNA损伤。其中,X射线和电离辐射常常引起DNA链的断裂,而紫外线常常引起嘧啶二聚体的形成,如TT,TC,CC等二聚体。这些嘧啶二聚体由于形成了共价键连接的环丁烷结构,因而会引起复制障碍。

20
人皮肤因受紫外线照射而形成二聚体的频率可达每小时5×104/细胞,只局限在皮肤中。
微生物受紫外线照射后,会影响其生存。 紫外线照射还能引起DNA链断裂等损伤。

21
电离辐射引起的DNA损伤 直接效应是DNA直接吸收射线能量而遭损伤,
主要是由OH-自由基引起,包括DNA链上的碱基氧化修饰、过氧化物的形成、碱基环的破坏和脱落等。 一般嘧啶比嘌呤更敏感。

22
b.脱氧核糖变化 脱氧核糖上的每个碳原子和羟基上的氢都能与OH-反应,导致脱氧核糖分解,引起DNA链断裂。 c.DNA链断裂 射线的直接和间接作用都可能使脱氧核糖破坏或磷酸二酯键断开。 单链断裂(single strand broken), 双链断裂(double strand broken)。 单链断裂发生频率为双链断裂的10-20倍,但比较容易修复;对单倍体细胞来说(如细菌)一次双链断裂就是致死事件。
。 单链断裂发生频率为双链断裂的10-20倍,但比较容易修复;对单倍体细胞来说(如细菌)一次双链断裂就是致死事件。")
23
2. 化学因素: 常见的化学诱变剂 化合物类别 分子改变 碱基类似物 (如:5-BU) A→5-BU →G 羟胺类 (NH2OH) T →C
亚硝酸盐 (NO2-) C →U 烷化剂 (如:氮芥类) G →mG
 C →U. 烷化剂 (如:氮芥类) G →mG.")
24
1、碱基类似物(Base analog) 5-溴尿嘧啶 5-Bromine Uracil 2-氨基嘌呤 2-Amino purine O
NH2

25
: G : A 酮式5-BrU的渗入 烯醇式enol 第一轮复制 酮式到烯醇式的转变 第二轮复制 酮式Keto A·TG·C 转变
AGCTTCCTA TCGAAGGAT AGCTBCCTA 酮式5-BrU的渗入 烯醇式enol Br OH H O : G AGCTBCCTA TCGAGGGAT AGCTTCCTA TCGAAGGAT 第一轮复制 酮式到烯醇式的转变 Br 酮式Keto H O AGCTBCCTA TCGAAGGAT AGCTCCCTA TCGAGGGAT 第二轮复制 : A A·TG·C 转变 5-BrU 烯醇式渗入为 G·CA·T 转变

26
2、 碱基的化学修饰导致突变 又称化学突变剂: 亚硝酸(nitrous acid HNO2) 羟氨(hydroxylamine HA)
2、 碱基的化学修饰导致突变 又称化学突变剂: 亚硝酸(nitrous acid HNO2) 羟氨(hydroxylamine HA) 甲磺酸乙酯(ethyl mathanesulfonate EMS) N-甲基-N’-硝基-N-亚硝基胍 (N-mathyl-N’-nitro-N-nitrosoguanidion NNG) 产生---错义突变
 羟氨(hydroxylamine HA) 甲磺酸乙酯(ethyl mathanesulfonate EMS) N-甲基-N’-硝基-N-亚硝基胍. (N-mathyl-N’-nitro-N-nitrosoguanidion NNG) 产生---错义突变.")
27
亚硝酸与亚硝酸盐 AT GC 可加速C脱氨基生成U,A脱氨基生成H。 GC AT

28
Nitrous Acid Pickled food Nitrous acid Cancer
A fairly simple chemical mutagen is nitrous acid. It has been discovered that nitrous acid has some relationship with higher incidence of cancer. What is the reason? 一种相当简单的化学诱变剂是亚硝酸,研究发现,亚硝酸与某些癌症高发事件有关,原因是什么呢? Nitrous acid Cancer 28

29
烷化剂的诱变机制

30
EMS: ethylmethane sulfonate
具有诱变作用的烷化剂的一个实例是乙基甲磺酸(EMS)。EMS能够将乙基(─CH2CH3)加到含氮碱基的多个位置上。其后果取决于乙基被加到哪个位置。当乙基被加到鸟嘌呤的酮上时能发生很强的诱变修饰作用。此时,鸟嘌呤的碱基配对特性被改变,它会与胸腺嘧啶配对而不与胞嘧啶配对。同样,如果这样的改变没有得到修复的话就会在下一轮复制产生的DNA中出现一个完全的突变。 EMS Replication G : C mG : C mG : T G – C → A – T 30
。EMS能够将乙基(─CH2CH3)加到含氮碱基的多个位置上。其后果取决于乙基被加到哪个位置。当乙基被加到鸟嘌呤的酮上时能发生很强的诱变修饰作用。此时,鸟嘌呤的碱基配对特性被改变,它会与胸腺嘧啶配对而不与胞嘧啶配对。同样,如果这样的改变没有得到修复的话就会在下一轮复制产生的DNA中出现一个完全的突变。 EMS. Replication. G : C. mG : C. mG : T. G – C → A – T. 30.")
31
3、嵌合剂的致突变作用 吖啶橙 (Acridine Orange AO) 扁平染料分子
溴化乙锭 (Ethidium Bromide EB ) 分子插入 -A TTTCG - -T AAAGC- TAO T -ATTTCG - -TAAAGC- AO EB -ATTTTTCG - -TAAAAAGC- 吖啶橙插入导致碱基的缺失突变 结合前面错义、移框的抑制突变-----因此遗传学上鉴定突变型(突变类型)可用分别使用产生错义、移框突变的突变剂二次处理,看其回复突变的有无 -AT EB TTTTCG- -TA -ATX’TTTTCG- -TAX AAAAGC- AAAAGC- X 结果产生---移框突变
 分子插入. -A TTTCG - -T AAAGC- TAO T. -ATTTCG - -TAAAGC- AO. EB. -ATTTTTCG - -TAAAAAGC- 吖啶橙插入导致碱基的缺失突变. 结合前面错义、移框的抑制突变-----因此遗传学上鉴定突变型(突变类型)可用分别使用产生错义、移框突变的突变剂二次处理,看其回复突变的有无. -AT EB TTTTCG- -TA. -ATX’TTTTCG- -TAX AAAAGC- AAAAGC- X. 结果产生---移框突变.")
32
4、 突变热点(Mutation Hotpoint)
基因自发突变的频率是一定的 DNA分子上某些位点的突变频率大大平均数,这样的位点称为----

33
a) 重复序列 (repetitive seq. )/ palindromic seq.
e.g. Lac. Operon ---CTGGCTGGCTGG--- 复制时,模板与新生链间滑动错配 缺失,插入突变 b) m5C C m5C T 脱氨氧化 U 复制期发生在新生链上不发生突变, 若发生在模板链上很快突变 非复制期突变频率50% m5C脱氨氧化成T若发生在DNA复制期间的新生链上,就可以被修复;但若发生在复制期间的模板链上,则很快引起突变 c) 不同诱变剂作用位点不同,转座子插入位点不同
 m5C. C. m5C. T. 脱氨氧化. U. 复制期发生在新生链上不发生突变, 若发生在模板链上很快突变. 非复制期突变频率50% m5C脱氨氧化成T若发生在DNA复制期间的新生链上,就可以被修复;但若发生在复制期间的模板链上,则很快引起突变. c) 不同诱变剂作用位点不同,转座子插入位点不同.")
34
第三节 DNA的修复系统 一、复制修复 二、损伤修复 三、复制后修复

35
一、复制过程中的错配的修复

36
1、错配修复碱基来源:校正活性所漏校的碱基 使复制的保真性提高102~103倍
错配修复系统(MMR Mismatch Repair) DNA mismatch A----- C--- 错配碱基的修复会使复制的保真性提高102~103倍 DNApol (ξ= 10-8) 经第二次校正ξ= 10-11
 DNA mismatch A C--- 错配碱基的修复会使复制的保真性提高102~103倍. DNApol (ξ= 10-8) 经第二次校正ξ=")
37
2、错配修复系统(Mismatch repair )
(1)组成 dam 甲基化酶 DNA腺嘌呤甲基化酶(m6A甲基化酶) DNA polymerase Helicase SSB 外切核酸酶 (Ⅰ和Ⅶ) 连接酶 MCE (mismatch correct enzyme) 3 subunits mutH, L, S 扫描新生链中错配碱基 识别新生链中非 m6A 的GATC序列 酶切含错配碱基的新生DNA区段
组成. dam 甲基化酶. DNA腺嘌呤甲基化酶(m6A甲基化酶) DNA polymerase. Helicase SSB 外切核酸酶 (Ⅰ和Ⅶ) 连接酶. MCE (mismatch correct enzyme) 3 subunits mutH, L, S. 扫描新生链中错配碱基. 识别新生链中非 m6A 的GATC序列. 酶切含错配碱基的新生DNA区段.")
38
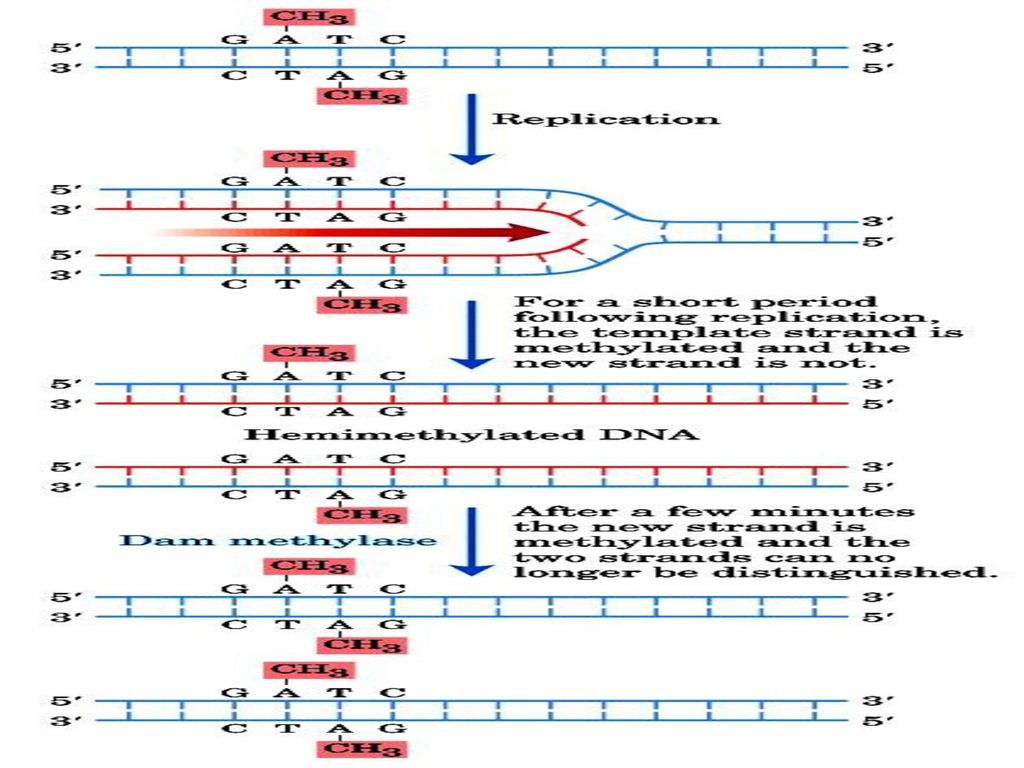
★DNA合成过程中的甲基化变化 平均每2kb左右有一GATC seq. 错配修复系统受甲基化的引导
DNA中的GATC(palindromic seq.) 为m6A甲基化敏感位点 平均每2kb左右有一GATC seq. 错配修复系统受甲基化的引导
 为m6A甲基化敏感位点. 平均每2kb左右有一GATC seq. 错配修复系统受甲基化的引导.")
40
a、MutH/MutS 扫描识别错配 碱基和邻近的GATC序列
(2) 修复过程 a、MutH/MutS 扫描识别错配 碱基和邻近的GATC序列 切点--甲基化GATC中 G的5’侧 甲基化程度的差异 DNA helicase II, SSB, exonuclease I去除包括错 配碱基的片段 Immediate methylation of the A in GATC of the parental strand but not of the daughter strand right after replication Expensive to keep the accuracy DNA polymerase III 和 DNA ligase 填充缺口 昂贵的代价用于保证DNA的准确性
 修复过程. a、MutH/MutS 扫描识别错配. 碱基和邻近的GATC序列. 切点--甲基化GATC中. G的5’侧. 甲基化程度的差异. DNA helicase II, SSB, exonuclease I去除包括错. 配碱基的片段. Immediate methylation of the A in GATC of the parental strand but not of the daughter strand right after replication. Expensive to keep the accuracy. DNA polymerase III 和. DNA ligase 填充缺口. 昂贵的代价用于保证DNA的准确性.")
41
Mismatch Repair 41

42
MMR: Mismatch Repair (1/2)
在MMR的第一步,蛋白质MutS和MutL结合到错配位置。同时,一种称为MutH的蛋白质结合到一定距离以外的GATC序列上。MutS- MutL和MutH之后形成一个复合体,在GATC位置产生一个单链缺口。 42

43
MMR: Mismatch Repair (2/2)
之后,称为UvrD的解旋酶从缺口的位置解开双螺旋到错配位点稍微后面一点。这使外切核酸酶得以摧毁含有错误的单链DNA区域。最后,DNA聚合酶III通过合成新的DNA修补这一空缺,假定不会再次出错。新DNA片段然后由连接酶与原先的链连接起来。这样,虽然错配和IDL一般来说只是影响到一个或几个碱基,但细胞的策略是移去错配位置旁边相对较大一块DNA,然后全部重新合成它。 43

44
3、 尿嘧啶-N-糖苷酶系统 ( ung system )
修复尿嘧啶的来源:dUTP的渗入 胞嘧啶的自发脱氨氧化

46
---TAGC--- ---A CG--- ung-ase ① ---TAGC--- A U C G ---TAGC---
---ATCG--- U AP内切酶(Apurinase) ② DNApolⅠ Ligase ③ ---TAGC--- ---A CG--- ---TAGC--- ---ATCG---
 ②. DNApolⅠ. Ligase. ③. ---TAGC A CG TAGC ATCG---")
47
二、DNA损伤的修复

48
一、嘧啶二聚体的产生 类型:TT二聚体 ( )、CC二聚体( )、 CT二聚体( ) TT CC CT C C 相邻的胸腺嘧啶
胸腺嘧啶二聚体

49
(一 ) 光复活(photoreactivation)
二、二聚体修复的机制 (一 ) 光复活(photoreactivation) 1. 概念:在可见光存在的条件下,在光复活酶作用下将UV引起嘧啶二聚体分解为单体的过程。 2. 条件:可见光(300~600nm)、PR酶、嘧啶二聚体 3. 作用过程: ①光复活酶与T=T结合形成复合物; ②复合物吸收可见光切断T=T之间的C-C共价键,使二聚体变成单体; ③光复酶从DNA链解离. *光复活是原核生物中的一种主要修复形式。
 光复活(photoreactivation) 1. 概念:在可见光存在的条件下,在光复活酶作用下将UV引起嘧啶二聚体分解为单体的过程。 2. 条件:可见光(300~600nm)、PR酶、嘧啶二聚体. 3. 作用过程: ①光复活酶与T=T结合形成复合物; ②复合物吸收可见光切断T=T之间的C-C共价键,使二聚体变成单体; ③光复酶从DNA链解离. *光复活是原核生物中的一种主要修复形式。")
50
光修复过程 ◆╥ 结构在DNA螺旋结构上形成一个巨大的凸起或扭曲,这对DNA分子好象是个“赘瘤”。这个“瘤”被一种特殊的巡回酶(patroling enzyme),例如光激活酶(photoreacting enzyme)所辨认,在有蓝色光波的条件下,二聚体被切开,DNA回复正常。这种经过解聚作用使突变回复正常的过程叫做光修复(light repair)
,例如光激活酶(photoreacting enzyme)所辨认,在有蓝色光波的条件下,二聚体被切开,DNA回复正常。这种经过解聚作用使突变回复正常的过程叫做光修复(light repair)")
51
光复活(photo reactivation )
复制前、不容易出错 400 nm 蓝光、PR 酶 (photo-reactivation enzyme) 光敏裂合酶(photolyase) ----TT---- ----AA---- ----TT---- ----AA---- 可见光激活 变形的DNA吸收与发色团特征波长相当的光波后,酶-DNA复合物利用这一能量使二聚体裂解为单链,酶随即脱落 PR ----TT---- ----AA---- ----TT---- ----AA----
 光敏裂合酶(photolyase) ----TT AA TT AA---- 可见光激活. 变形的DNA吸收与发色团特征波长相当的光波后,酶-DNA复合物利用这一能量使二聚体裂解为单链,酶随即脱落. PR. ----TT AA TT AA----")
52
(二) 切除修复 核苷酸切除修复,碱基切除修复
(二) 切除修复 核苷酸切除修复,碱基切除修复 1.概念:(核苷酸外切修复、暗修复)先在损伤的任何一端打开磷酸二酯键,然后外切掉一段寡核苷酸;留下的缺口由修复性合成来填补,再由连接酶将其连接起来。2.特点:消除由UV引起的损伤,也能消除由电离辐射和化学诱变剂引起的其他损伤。切除的片段可由几十到上万bp,分别称短补丁修复、长补丁修复。 3.过程: ①内切酶的作用在DNA损伤的一端,切开形成一个切口; ②外切酶的作用将损伤部位切除; ③聚合酶的作用将切口补齐,留下一个切口; ④连接酶的作用将DNA连接形成完整的DNA链。
 切除修复 核苷酸切除修复,碱基切除修复. 1.概念:(核苷酸外切修复、暗修复)先在损伤的任何一端打开磷酸二酯键,然后外切掉一段寡核苷酸;留下的缺口由修复性合成来填补,再由连接酶将其连接起来。2.特点:消除由UV引起的损伤,也能消除由电离辐射和化学诱变剂引起的其他损伤。切除的片段可由几十到上万bp,分别称短补丁修复、长补丁修复。 3.过程: ①内切酶的作用在DNA损伤的一端,切开形成一个切口; ②外切酶的作用将损伤部位切除; ③聚合酶的作用将切口补齐,留下一个切口; ④连接酶的作用将DNA连接形成完整的DNA链。")
53
在原核生物中,与DNA切除修复有关的蛋白质包括UvrA、UvrB、UvrC,以及DNA pol Ⅰ和DNA连接酶。

54
核苷酸切除修复 NER: Nucleotide Excision Repair
核苷酸切除修复(NER)表面上看与错配修复相似。大肠杆菌中NER开始于蛋白质UvrA和UvrB结合到DNA的损伤部位。UvrA很快就离开了,但UvrB仍旧保持接触。之后,UvrC结合到UvrB上并在损伤碱基的两边各产生一个单链切口。一般来说两个切口之间相距12个核苷酸。 54
表面上看与错配修复相似。大肠杆菌中NER开始于蛋白质UvrA和UvrB结合到DNA的损伤部位。UvrA很快就离开了,但UvrB仍旧保持接触。之后,UvrC结合到UvrB上并在损伤碱基的两边各产生一个单链切口。一般来说两个切口之间相距12个核苷酸。 54.")
55
NER: Nucleotide Excision Repair (2/2)
然后,解旋酶UvrD彻底去除含有错误的单链片段。空缺的部分由DNA聚合酶重新合成。 55

56
真核生物XP切除修复系统 1.识别 4.修复 2.组装 3.切除

57
(xeroderma pigmentosis,XP)
着色性干皮病 (xeroderma pigmentosis,XP) 是一种切除修复有缺陷的遗传性疾病。 在研究其发病机制时,发现一些相关的基 因,称为XPA、XPB、XPC等。这些基因 的表达产物与Uvr类蛋白有同源序列,也是 起辨认和切除损伤DNA作用的。XP病人是 由于XP基因有缺陷,不能修复紫外线照射 引起的DNA损伤,因此易发生皮肤癌。
 是一种切除修复有缺陷的遗传性疾病。 在研究其发病机制时,发现一些相关的基. 因,称为XPA、XPB、XPC等。这些基因. 的表达产物与Uvr类蛋白有同源序列,也是. 起辨认和切除损伤DNA作用的。XP病人是. 由于XP基因有缺陷,不能修复紫外线照射. 引起的DNA损伤,因此易发生皮肤癌。")
59
碱基切除修复 Base Excision Repair
Damage to C (e.g. deamination) 碱基切除修复(BER)用于取代经历了常见的和较小修饰(如脱氨基)的碱基。 59
 碱基切除修复(BER)用于取代经历了常见的和较小修饰(如脱氨基)的碱基。 59.")
60
碱基切除修复有关的酶 1) DNA糖基化酶(DNA glycosylase): 识别DNA中损伤的或错误的碱基;水解N-糖苷键去除碱基。 2) AP内切酶(Endonucleases) : 有3’5’外切酶活性,切除无碱基位点 3) 脱氧核糖磷酸二酯酶(dRpase): 切除AP位点的脱氧核糖磷酸,产生一个核苷酸的缺口。 4) DNA聚合酶: 在缺口处进行修复,加入一个核苷酸。在AP位点5’端进行广泛的修复合成。 5) DNA连接酶:连接最后一个切口。
 脱氧核糖磷酸二酯酶(dRpase): 切除AP位点的脱氧核糖磷酸,产生一个核苷酸的缺口。 4) DNA聚合酶: 在缺口处进行修复,加入一个核苷酸。在AP位点5’端进行广泛的修复合成。 5) DNA连接酶:连接最后一个切口。")
62
BER: Base Excision Repair (1/2)
从机理上看,BER与NER和MMR有很大不同。BER开始的时候,称为DNA糖基化酶的蛋白质结合到发生损伤的碱基上并切断它与脱氧核糖的连接而将其去除。细胞含有几种不同的DNA糖基化酶,分别识别并去除不同种类的损伤碱基。下一步,原来与损伤碱基连接的糖和磷酸也被去除,在DNA上留下一个小的空隙。称为AP内切核酸酶和DNA磷酸二酯酶的蛋白质负责完成这一步。 62

63
BER: Base Excision Repair (2/2)
最后,DNA聚合酶I修补这一空隙,在原先是损伤碱基的位置加上一个正常的核苷酸。实际上,DNA聚合酶I还会用它独特的5’3’外切核酸酶功能同步去除并合成空隙下游几个核苷酸。在真核生物和原核生物中,BER是相似的。 63

64
复制后修复 有时DNA损伤后没有被及时修复,通过机体细胞内一套复制后修复系统,可使危害减到最小。 重组修复 SOS修复

65
重组修复 1.概念: 通过对DNA的复制和同源链的重组,来完成对损伤部位的修复,又称复制后修复。 2.特点: ① 修复过程伴随DNA的复制和重组; ② 仅修复新合成的不完整的单链,原先的损伤单链仍然保留; ③部分重组蛋白的精确性差,修复的出错率较高。 3.重组修复过程: (1)复制:以损伤单链为模板复制时,越过损伤部位,对应位点留下缺口;未损伤单链复制成完整双链。 (2)重组:缺口单链与完整同源单链重组,缺口转移到完整链,使损伤单链的互补链完整,损伤单链仍然保留。 (3)再合成:转移后的缺口以新互补链为模板聚合补齐。
复制:以损伤单链为模板复制时,越过损伤部位,对应位点留下缺口;未损伤单链复制成完整双链。 (2)重组:缺口单链与完整同源单链重组,缺口转移到完整链,使损伤单链的互补链完整,损伤单链仍然保留。 (3)再合成:转移后的缺口以新互补链为模板聚合补齐。")
66
重组修复(recombination repairing):
这是DNA的复制过程中所采用的一种有差错的修复方式。

67
重组修复(Recombinative—Repair)
后复制修复、E.coli的挽回系统 该系统存在的实验证据 E.coli 存活% U.V 计量 w.t. UvrA+ RecA+ rec a- uvr a-

68
(四) SOS修复 3.特点: 1. 概念:是在DNA分子受损伤的范围较大而且复制受到抑制时出现的一种应急修复作用。 2. 过程
①当DNA损伤较大时(如产生很多的T=T),正常的DNA多聚酶复制到损伤位点时,其活性受到抑制; ②短暂抑制后产生一种新的DNA多聚酶,催化损伤部位DNA的复制,由于新的DNA多聚酶的修复校正功能较低,新合成的碱基错配频率较高,易引起突变。 3.特点: ①修复系统需要在DNA分子受损伤的范围较大而且复制受到抑制时才能够启动。 ②修复系统对错配碱基的修复校正功能低下,从而增加突变的频率。
,正常的DNA多聚酶复制到损伤位点时,其活性受到抑制; ②短暂抑制后产生一种新的DNA多聚酶,催化损伤部位DNA的复制,由于新的DNA多聚酶的修复校正功能较低,新合成的碱基错配频率较高,易引起突变。 3.特点: ①修复系统需要在DNA分子受损伤的范围较大而且复制受到抑制时才能够启动。 ②修复系统对错配碱基的修复校正功能低下,从而增加突变的频率。")
69
③在紧急情况下,细胞通过一定水平的变异来换取细胞的幸存,有利于细胞逃生。 4
③在紧急情况下,细胞通过一定水平的变异来换取细胞的幸存,有利于细胞逃生。 4.SOS系统的启动: 通过操纵子(结构基因、启动子、操纵基因、调节基因)来实现: A. SOS基因:recA基因、UvrA、UvrB、UmuC等,也称din基因 (damage inducible gene),为操纵子的结构基因; B. lex基因:阻遏蛋白基因,正常情况下结合在操纵基因上; C. recA基因:重组蛋白基因,应急状态下启动蛋白质水解酶活性,水解阻遏蛋白,使din基因高效表达,从而启动SOS修复系统。
来实现: A. SOS基因:recA基因、UvrA、UvrB、UmuC等,也称din基因 (damage inducible gene),为操纵子的结构基因; B. lex基因:阻遏蛋白基因,正常情况下结合在操纵基因上; C. recA基因:重组蛋白基因,应急状态下启动蛋白质水解酶活性,水解阻遏蛋白,使din基因高效表达,从而启动SOS修复系统。")
70
SOS修复 当DNA损伤广泛难以继续复制时,由此而诱发出一系列复杂的反应。
在E. coli,各种与修复有关的基因,组成一个称为调节子(regulon)的网络式调控系统。 这种修复特异性低,对碱基的识别、选择能力差。通过SOS修复,复制如能继续,细胞是可存活的。然而DNA保留的错误较多,导致较广泛、长期的突变。
的网络式调控系统。 这种修复特异性低,对碱基的识别、选择能力差。通过SOS修复,复制如能继续,细胞是可存活的。然而DNA保留的错误较多,导致较广泛、长期的突变。")
71
SOS系统诱导,错误潜伏的复制超越二聚体而进行
SOS 修复--无模板指导的DNA复制 大剂量的紫外线照射,大量的二聚体产生 SOS系统诱导,错误潜伏的复制超越二聚体而进行 SOS修复酶只有在细胞受到损伤时才存在(正常细胞中不存在), 机制--a、SOS系统以某种方式对polⅢ进行修饰(改变校对亚基功能)b、由polⅡ负责超越创伤 复制结果大量的没有被错配修复系统和切除修复系统纠正的错误碱基导致突变--错误倾向性----存活下来总比死亡好 错误碱基
, 机制--a、SOS系统以某种方式对polⅢ进行修饰(改变校对亚基功能)b、由polⅡ负责超越创伤. 复制结果大量的没有被错配修复系统和切除修复系统纠正的错误碱基导致突变--错误倾向性----存活下来总比死亡好. 错误碱基.")
72
SOS repair 是一种错误倾向性极强的修复机制
当DNA复制度过难关后 RecA-p很快消失 LexA gene on SOS off SOS repair 是一种错误倾向性极强的修复机制 是进化中形成的“ 竭尽全力,治病救人” 的措施 (正常状态下,SOS是关闭的) SOS repair 是一种error-prone 极强的修复机制
 SOS repair 是一种error-prone 极强的修复机制.")
73
真核生物DNA损伤修复缺陷疾病 病名 灵敏性 肿瘤易感性 症状 着色性 干皮病 UV辐射烷化作用 皮肤癌黑色素瘤 皮肤及眼对光敏感 毛细血管
扩张性失调 γ射线 淋巴瘤 不稳定的步态(运动失调)皮肤及腿血管扩张(毛细血管扩张)染色体失常 Fanconi贫血 交联剂 白血病 血细胞数减少,先天畸形 Bloom综合征 UV辐射 光敏感,DNA连接酶缺陷
皮肤及腿血管扩张(毛细血管扩张)染色体失常. Fanconi贫血. 交联剂. 白血病. 血细胞数减少,先天畸形. Bloom综合征. UV辐射. 光敏感,DNA连接酶缺陷.")
74
基因突变 (一)、突变概念 突变(mutation)是指DNA发生可遗传的永久性的突变。 自发突变:自然发生的突变
诱发突变:理化因素引起的突变 突变可发生在染色体水平或基因水平,本节主要介绍基因突变。

75
(二)、突变类型 (一)点突变:DNA分子上的碱基错配称点突变(point mutation)。 (1)转换:发生在同型碱基之间,即嘌呤代替另一嘌呤,或嘧啶代替另一嘧啶。 (2)颠换:发生在异型碱基之间,即嘌呤变嘧啶或嘧啶变嘌呤。
、突变类型 (一)点突变:DNA分子上的碱基错配称点突变(point mutation)。 (1)转换:发生在同型碱基之间,即嘌呤代替另一嘌呤,或嘧啶代替另一嘧啶。 (2)颠换:发生在异型碱基之间,即嘌呤变嘧啶或嘧啶变嘌呤。")
76
N-val · his · leu · thr · pro · glu · glu · · · · · · C 肽链
正常成人Hb (HbA)β亚基 N-val · his · leu · thr · pro · glu · glu · · · · · · C 肽链 CTC GAG 基因 镰形红细胞贫血病人Hb (HbS) β亚基 N-val · his · leu · thr · pro · val · glu · · · · · · C 肽链 CAC GTG 基因
β亚基. N-val · his · leu · thr · pro · glu · glu · · · · · · C. 肽链. CTC GAG. 基因. 镰形红细胞贫血病人Hb (HbS) β亚基. N-val · his · leu · thr · pro · val · glu · · · · · · C. 肽链. CAC GTG. 基因.")
77
(二)移码突变,缺失 (deletion)、插入 (insertion) 1. 缺失:一个碱基或一段核苷酸链从DNA大分子上消失。 2
(二)移码突变,缺失 (deletion)、插入 (insertion) 1.缺失:一个碱基或一段核苷酸链从DNA大分子上消失。 2.插入:原来没有的一个碱基或一段核苷酸链插入到DNA大分子中间。 3.缺失或插入都可导致移框 (frame-shift)突变。框移突变是指三联体密码的阅读方式改变,造成蛋白质氨基酸排列顺序发生改变。
移码突变,缺失 (deletion)、插入 (insertion) 1.缺失:一个碱基或一段核苷酸链从DNA大分子上消失。 2.插入:原来没有的一个碱基或一段核苷酸链插入到DNA大分子中间。 3.缺失或插入都可导致移框 (frame-shift)突变。框移突变是指三联体密码的阅读方式改变,造成蛋白质氨基酸排列顺序发生改变。")
78
“The cat ate its rat.” “Txh eca tat eit sra t.” “The cat ate its rat.”
hpcsc.appstate.edu/index.php? module=pagemaste... 78

79
缺失引起框移突变 正常 缺失C 5’ ……G C A G U A C A U G U C …… 丙 缬 组 缬
谷 酪 蛋 丝 5’ ……G C A G U A C A U G U C …… 丙 缬 组 缬 正常 5’ ……G A G U A C A U G U C …… 缺失C

80
Frameshift mutation 移码突变在翻译中会产生剧烈的影响。位于突变位点上游的密码子没有受到影响。然而。突变位点下游的大多数密码子将变成彻底不同的密码子,它们编码完全不同的氨基酸或变成终止密码子。如果蛋白质还是被生产出来了,那么它基本上是没有功能的。 80

81
2. 根据对遗传信息的改变情况分类 (1)同义突变:指没有改变产物氨基酸序列的密码子变化。 密码子的简并性。AAA,AAG-----赖氨酸
(2)错义突变:指碱基序列的改变引起了表达产物氨基酸序列的改变。 (3)无义突变:指某个碱基的改变使代表某种氨基酸的密码子变成终止密码子。
错义突变:指碱基序列的改变引起了表达产物氨基酸序列的改变。 (3)无义突变:指某个碱基的改变使代表某种氨基酸的密码子变成终止密码子。")
82
82

83
Missense mutation 83

84
Nonsense mutation 84

85
3.根据突变表现型对环境的敏感性分类 (1)非条件性突变(nonconditional mutation) (2)条件性突变(conditional mutation)根据研究目的选择一定的突变,但能正常生长或近乎正常。最常用的条件是温度,如温度敏感突变。
非条件性突变(nonconditional mutation) (2)条件性突变(conditional mutation)根据研究目的选择一定的突变,但能正常生长或近乎正常。最常用的条件是温度,如温度敏感突变。")
86
4.从突变效应背离或回到野生型可分为 (1)正向突变(forward mutation):指野生型变为突变型
(2)回复突变(back mutation)基因发生突变得到方向是可逆的,突变株经过又一次突变可恢复为野生型的性状,这种第二次点突变称为回复突变。
回复突变(back mutation)基因发生突变得到方向是可逆的,突变株经过又一次突变可恢复为野生型的性状,这种第二次点突变称为回复突变。")
87
本章结束!!

Similar presentations





 發現遺傳因子。 1. 基因的不同等位基因解釋了諸多的遺傳性狀。 2. 對每一種性狀而言,一種生物體遺傳有兩個等位基因, 每一個等位基因得自於一方親代.>")






>")





 染色體:細胞核上(細胞未分裂前稱為染色質)>")



进行了著名的肺炎双球菌转化实验。加热杀死的S型肺炎球菌可以使无害的R型肺炎球菌转化为有害的S型肺炎球菌,为什么呢?在美国纽约洛克菲勒研究所工作的艾弗里(Avery)立刻敏感地抓住了这一问题,进行了“转化因子”实验,艾弗里等人的研究工作表明:DNA是遗传物质。>")
