Download presentation
Presentation is loading. Please wait.

1
重组DNA技术 Recombinant DNA Technology
第二十四章 重组DNA技术 Recombinant DNA Technology (工具酶 目的基因的获得 载体 基因克隆 外源基因的原核表达 真核表达)
")
2
克隆(clone) 来自同一始祖的相同副本或拷贝的集合。 获取同一拷贝的过程称为克隆化(cloning),即无性繁殖。 技术水平:分子克隆(molecular clone) 即DNA 克隆(DNA cloning) 细胞克隆 器官(或组织)克隆 个体克隆(动物或植物)
克隆. 个体克隆(动物或植物)")
3
重组DNA技术(recombinant DNA technology)
又称分子克隆(molecular cloning)或DNA克隆(DNA cloning)或基因克隆(gene cloning) 技术,是指在体外利用酶学方法将目的DNA片段与能自主复制的遗传元件(又叫载体)连接,形成重组DNA分子,进而在受体细胞中复制、扩增,从而获得单一DNA分子的大量拷贝。 克隆目的基因后,针对该基因进行特定蛋白质或多肽制备以及定向改造基因结构所用的方法及相关的工作统称为基因工程(genetic engineering)。因此,重组DNA技术又称为基因工程技术。
或DNA克隆(DNA cloning)或基因克隆(gene cloning) 技术,是指在体外利用酶学方法将目的DNA片段与能自主复制的遗传元件(又叫载体)连接,形成重组DNA分子,进而在受体细胞中复制、扩增,从而获得单一DNA分子的大量拷贝。 克隆目的基因后,针对该基因进行特定蛋白质或多肽制备以及定向改造基因结构所用的方法及相关的工作统称为基因工程(genetic engineering)。因此,重组DNA技术又称为基因工程技术。")
4
* 重组DNA技术的发展简史 1865年 G.J.Mendel的豌豆杂交试验 1944年 O.T.Avery的肺炎球菌转化实验
1972年 P. Berg 构建第一个重组DNA分子(猿猴病毒DNA和 λ噬菌体DNA) 1973年 S. Cohen首次将DNA片段与质粒连接,并转化入E.coli 1977年 美国南旧金山由博耶和斯旺森建立世界上第一家遗传工, 程公司,专门应用重组DNA技术制造医学上重要的药物 1980年 开始建造第一家应用重组DNA技术生产胰岛素的工厂 1997年 英国罗林研究所成功地克隆了“多莉”羊 Mendel Paul Berg Ian Wilmut and “Dolly”
 1973年 S. Cohen首次将DNA片段与质粒连接,并转化入E.coli. 1977年 美国南旧金山由博耶和斯旺森建立世界上第一家遗传工, 程公司,专门应用重组DNA技术制造医学上重要的药物. 1980年 开始建造第一家应用重组DNA技术生产胰岛素的工厂. 1997年 英国罗林研究所成功地克隆了 多莉 羊. Mendel. Paul Berg. Ian Wilmut and Dolly")
5
Frequently Used Enzymes in Recombinant DNA Technology
第 一 节 重组DNA技术中常用的 工具酶 Frequently Used Enzymes in Recombinant DNA Technology

6
工具酶在重组DNA技术中具有 特殊的用途 限制性核酸内切酶 DNA连接酶 碱性磷酸酶 反转录酶 末端脱氧核苷酰转移酶(末端转移酶)
DNA聚合酶I大片段 Taq DNA聚合酶 多核苷酸激酶

7
重组DNA技术中常用的工具酶 工 具 酶 功 能 限制性核酸内切酶 识别特异序列,切割DNA DNA连接酶
功 能 限制性核酸内切酶 识别特异序列,切割DNA DNA连接酶 催化DNA中相邻的5´磷酸基和3´羟基末端之间形成磷酸二酯键,使DNA切口封合或使两个DNA分子或片段连接 碱性磷酸酶 切除末端磷酸基 反转录酶 合成cDNA;替代DNA聚合酶I进行填补,标记或DNA序列分析 末端脱氧核苷酰转移酶 在3´羟基末端进行同聚物加尾 DNA聚合酶Ⅰ 合成双链cDNA分子或片段连接;缺口平移制作高比活性探针;DNA序列分析;填补3´末端 Klenow片段 又名DNA聚合酶I大片段,具有完整DNA聚合酶I的53聚合、35外切活性,而无53外切活性。常用于cDNA第二链合成,双链DNA 3末端标记等 Taq DNA聚合酶 常用于PCR;产物的3′末端带有A,可用于T-A克隆 多核苷酸激酶 催化多聚核苷酸5´羟基末端磷酸化,或标记探针

8
一、限制性核酸内切酶用于切割DNA + 定义:
限制性核酸内切酶(restriction endonuclease, RE)是识别DNA的特异序列, 并在识别位点或其周围切割双链DNA的一类内切酶。限制性核酸内切酶是重组DNA技术中重要的工具酶。 Bam HⅠ GGATCC CCTAGG G CCTAG + GATCC G
是识别DNA的特异序列, 并在识别位点或其周围切割双链DNA的一类内切酶。限制性核酸内切酶是重组DNA技术中重要的工具酶。 Bam HⅠ. GGATCC. CCTAGG. G. CCTAG. + GATCC. G.")
9
分类:根据酶的识别切割特性、催化条件及是否具有修饰酶活性可分为Ⅰ、Ⅱ、Ⅲ型三大类(基因工程技术中常用Ⅱ型)
★Ⅰ型:属于复合功能酶,兼有修饰和切割DNA两种特性,需要Mg2+、ATPs-腺苷酰甲硫氨酸作为催化的辅因子,在DNA降解时伴随有ATP的水解。即它具有核酸内切酶、甲基化酶、ATP酶和DNA解旋酶四种活性。若识别位点上两条DNA链均未甲基化,则行使内切酶功能,并在切割DNA同时或之后转变为ATP酶;若位点上只有1条链被甲基化则发挥修饰功能,使另一条链也甲基化;若位点上两条链均甲基化就与位点解离。其显著特点是识别位点和切割部位不一致,无固定切割位点,一般在识别位点以外的1kb(不少于400bp)到几kb(可多达7kb)处随机切割,不产生特异片段,故在基因重组中没有应用价值。 ★ Ⅲ型:与Ⅰ型酶特性类似,也有甲基化功能,但无ATP酶和DNA解旋酶活力;不同的是它能在DNA链上的特异位点切割,其切割位点在识别位点以外,对基因工程的意义也不大。
到几kb(可多达7kb)处随机切割,不产生特异片段,故在基因重组中没有应用价值。 ★ Ⅲ型:与Ⅰ型酶特性类似,也有甲基化功能,但无ATP酶和DNA解旋酶活力;不同的是它能在DNA链上的特异位点切割,其切割位点在识别位点以外,对基因工程的意义也不大。")
10
与甲基化酶共同构成细菌的限制修饰系统,限制外源DNA, 保护自身DNA,犹似高等动物的免疫系统。
★Ⅱ型:就是通常指的DNA限制性内切酶,在基因工程技术中常用。特点:分子量小,仅需Mg2+作为催化反应辅助因子,它们能识别双链DNA的特异顺序,并在这个顺序内进行切割,产生特异的DNA片段。 Ⅱ类酶识别序列特点为回文结构,一般为4~6bps(有些为8或8个以上碱基序列) ,且富含GC。 GGATCC CCTAGG 限制酶的作用 与甲基化酶共同构成细菌的限制修饰系统,限制外源DNA, 保护自身DNA,犹似高等动物的免疫系统。
 ,且富含GC。 GGATCC. CCTAGG. 限制酶的作用. 与甲基化酶共同构成细菌的限制修饰系统,限制外源DNA, 保护自身DNA,犹似高等动物的免疫系统。")
11
(一)重组DNA技术中通常使用Ⅱ型限制酶
命名 Hin dⅢ Haemophilus influenzae d株 流感嗜血杆菌d株的第三种酶 属 种 株 序 第一个字母取自产生该酶的细菌属名,用大写斜体; 第二、第三个字母是该细菌的种名,用小写斜体; 第四个字母(有时无)代表株(可大写或小写); 用罗马数字表示发现的先后次序。
代表株(可大写或小写); 用罗马数字表示发现的先后次序。")
12
EcoRⅠ 命名 属 种 株 序 E=Escherichia,埃希氏菌属 co=coli,大肠杆菌菌种 R=RY13,菌株名
属 种 株 序 E=Escherichia,埃希氏菌属 co=coli,大肠杆菌菌种 R=RY13,菌株名 I=第一个被分离到的内切酶

13
(二)Ⅱ型限制性内切酶具有一些共性和特性
1. 具有特定的识别和切割位点 II型限制性内切酶的识别位点举例('示切割位点) 限制性内切酶 识别位点 Apa I GGGCC’C C’CCGGG Sma I CCC’GGG GGG’CCC BamH I G’GATCC CCTAG’G Sau 3A I GATC’ ’CTAG Pst I CTGCA’G G’ACGTC Not I GC’GGCCGC CGCCGG’CG EcoR I G’AATTC CTTAA’G Sfi I GGCCNNNN’NGGCC CCGGN’NNNNCCGG
 限制性内切酶. 识别位点. Apa I. GGGCC’C. C’CCGGG. Sma I. CCC’GGG. GGG’CCC. BamH I. G’GATCC. CCTAG’G. Sau 3A I. GATC’ ’CTAG. Pst I. CTGCA’G. G’ACGTC. Not I. GC’GGCCGC. CGCCGG’CG. EcoR I. G’AATTC. CTTAA’G. Sfi I. GGCCNNNN’NGGCC. CCGGN’NNNNCCGG.")
14
2. 识别的核苷酸序列通常为回文结构(palindrome)
")
16
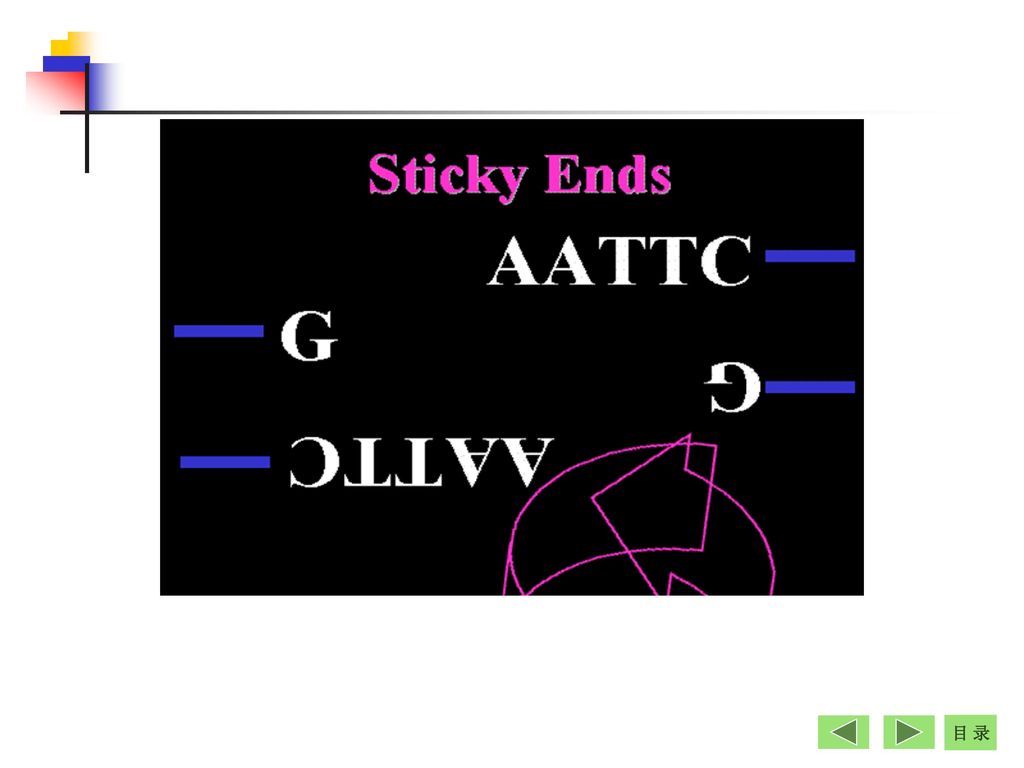
+ + 平端切口 黏端切口 (blunt end) (sticky end) 3. 切割双链DNA分子后产生黏端或平端 GTCGAC
HindⅡ Bam HⅠ GTCGAC CAGCTG GGATCC CCTAGG GTC CAG GAC CTG + GATCC G + G CCTAG 平端切口 黏端切口 (blunt end) (sticky end)
 (sticky end)")
18
4. 不同的酶可以产生同样的末端 同尾酶(isocaudarner)
有些限制性内切酶虽然识别序列不完全相同,但切割DNA后,产生相同的黏端,这样的酶彼此互称为同尾酶。这两个相同的黏端称为配伍末端(compatible end)。 Bam HⅠ Bg lⅡ GGATCC CCTAGG AGATCT TCTAGA A TCTAG + GATCT A GATCC G G CCTAG +
。 Bam HⅠ. Bg lⅡ. GGATCC. CCTAGG. AGATCT. TCTAGA. A. TCTAG. + GATCT. A. GATCC. G. G. CCTAG. +")
19
5. 不同的酶可以识别同一个序列 能识别同一序列(切割位点可同或不同)但来源不同的两种酶互称同工异源酶(isoschizomer)或同裂酶。
Bam HⅠ GGATCC CCTAGG G CCTAG GATCC G + BstⅠ GGATCC CCTAGG GATCC G G CCTAG + XmaⅠ CCCGGG GGGCCC C CCCGG CCGGG G + SmaⅠ GGG CCC CCCGGG GGGCCC CCC GGG +

20
6. 切割会受其他因素的影响 限制性内切酶通常不能切割在识别位点内有特异碱基甲基化的序列。 缓冲液或环境温度也会影响一些酶的特异性。例如,EcoR I在正常情况下识别“-GAATTC-”序列,但在甘油浓度大于5%(v/v)或反应温度较低时识别序列可变为“-AATT-”或“-嘌呤嘌呤AT嘧啶嘧啶-”,这种现象称为星号活性(star activity),以EcoR I*表示。
或反应温度较低时识别序列可变为 -AATT- 或 -嘌呤嘌呤AT嘧啶嘧啶- ,这种现象称为星号活性(star activity),以EcoR I*表示。")
21
二、DNA连接酶用于催化DNA片段连接 连接酶的作用机制
DNA连接酶(DNA ligase):催化两个相邻的3′-OH和5′-磷酸基团形成3′,5′-磷酸二酯键,从而使DNA片段或单链断裂形成的缺口连接起来。 连接酶的作用机制
:催化两个相邻的3′-OH和5′-磷酸基团形成3′,5′-磷酸二酯键,从而使DNA片段或单链断裂形成的缺口连接起来。 连接酶的作用机制.")
22
BamH I

23
DNA连接酶的来源 1. 大肠杆菌染色体编码的 DNA连接酶 需NAD+作为辅因子 2. 大肠杆菌T4噬菌体DNA编码的T4 DNA连接酶 需ATP作为辅因子

24
三、重组DNA 技术中还需使用其他常用工具酶
(一) 碱性磷酸酶能去除核酸分子末端的磷酸基团 (二)反转录酶以RNA为模板合成cDNA (三)末端脱氧核苷酰转移酶用于给DNA末端加尾以构成黏性末端 (四)DNA聚合酶I主要用于合成DNA (五)DNA聚合酶I大片段保留了有用的酶活性 (六)Taq DNA聚合酶常用于PCR (七)多核苷酸激酶可使寡核苷酸链5´羟基末端磷酸化
 碱性磷酸酶能去除核酸分子末端的磷酸基团. (二)反转录酶以RNA为模板合成cDNA. (三)末端脱氧核苷酰转移酶用于给DNA末端加尾以构成黏性末端. (四)DNA聚合酶I主要用于合成DNA. (五)DNA聚合酶I大片段保留了有用的酶活性. (六)Taq DNA聚合酶常用于PCR. (七)多核苷酸激酶可使寡核苷酸链5´羟基末端磷酸化.")
25
(一)碱性磷酸酶能去除核酸分子末端的磷酸基团
来自于大肠杆菌(bacterial alkaline phosphatase,BAP)或小牛肠(calf intestinal alkaline phosphatase,CIP)。用于脱去DNA(RNA)5′末端的磷酸基团,使5′ -P成为5′ -OH,该过程称核酸分子的脱磷酸作用。 P 5′ OH 3′ OH 5′ 3′
或小牛肠(calf intestinal alkaline phosphatase,CIP)。用于脱去DNA(RNA)5′末端的磷酸基团,使5′ -P成为5′ -OH,该过程称核酸分子的脱磷酸作用。 P. 5′ OH. 3′ OH. 5′ 3′")
26
(二)反转录酶以RNA为模板合成cDNA (reverse transcriptase)
5′ 3′ 5′ 3′ AAAAAA 反转录酶 (reverse transcriptase) Oligo-dT 随机引物 AAAAAA TTTTTT cDNA cDNA 反转录酶催化的cDNA合成
 Oligo-dT. 随机引物. AAAAAA. TTTTTT. cDNA. cDNA. 反转录酶催化的cDNA合成.")
27
(三)末端脱氧核苷酰转移酶用于给DNA末端加尾以构成黏端
3′OH 5′ 5′ 3′ dGTP GGG GG 末端转移酶 3′ 5′ 5′ 3′

28
(四)DNA聚合酶I主要用于合成DNA 大肠杆菌DNA聚合酶I(DNA polymerase I)是基因工程中常用的DNA聚合酶。其除具有5′→3′聚合酶活性外,还有3′→5′及5′→3′核酸外切酶活性。因其具有5′→3′核酸外切酶活性而常用于DNA探针的缺口平移法(nick translation)标记。
是基因工程中常用的DNA聚合酶。其除具有5′→3′聚合酶活性外,还有3′→5′及5′→3′核酸外切酶活性。因其具有5′→3′核酸外切酶活性而常用于DNA探针的缺口平移法(nick translation)标记。")
29
(五)DNA聚合酶I大片段保留了有用的酶活性
DNA聚合酶I大片段(large fragment of DNA polymerase I)为DNA聚合酶I用枯草杆菌蛋白酶(subtilisin)裂解后产生的大片段,这个片段也称为Klenow片段(Klenow fragment)。其保留了5′→3′聚合酶活性及3′→ 5′外切酶活性,失去了5′ →3′外切酶活性。它具有的3′→5′外切酶活性能保证DNA复制的准确性,即把DNA合成过程中错配的核苷酸去除,再把正确的核苷酸接上去(该活性称为校对活性)。 Klenow片段的主要用途有:① 补齐双链DNA的3′末端,使其转变成平端;或通过补齐3′端而使3′末端标记;② 在cDNA克隆中,用于第二股链的合成;③ 用于DNA序列分析。
为DNA聚合酶I用枯草杆菌蛋白酶(subtilisin)裂解后产生的大片段,这个片段也称为Klenow片段(Klenow fragment)。其保留了5′→3′聚合酶活性及3′→ 5′外切酶活性,失去了5′ →3′外切酶活性。它具有的3′→5′外切酶活性能保证DNA复制的准确性,即把DNA合成过程中错配的核苷酸去除,再把正确的核苷酸接上去(该活性称为校对活性)。 Klenow片段的主要用途有:① 补齐双链DNA的3′末端,使其转变成平端;或通过补齐3′端而使3′末端标记;② 在cDNA克隆中,用于第二股链的合成;③ 用于DNA序列分析。")
30
G GA A T T C T T A A C T T A A 将黏端转变成平端 OH 3′ 5′ 5′ 3′ DNA聚合酶 3′ 5′ 5′
EcoRⅠ 对5′-黏端片段,利用大肠杆菌DNA聚合酶I的Klenow片段,在3′-OH端延伸,可填平末端的凹陷并将其转变成平端。

31
C T G C A C G G 将黏端转变成平端 PstⅠ 5′ 3′ 5′ 3′ 核酸外切酶 3′ 5′ 5′ 3′
对3′黏端的片段,应用如T4 DNA聚合酶的3′→5′的核酸外切酶活性可切去未配对的序列,将其转变成平端。

32
(六)Taq DNA聚合酶常用于PCR Taq DNA聚合酶具有良好的聚合活性和热稳定性,常用于PCR。该酶具有5′→3′聚合酶活性及5′→3′外切酶活性,但没有3′→5′外切酶活性,因而无3′→5′校对活性,故在PCR反应中如果发生碱基错配,该酶没有校正功能。针对此不足,目前研发出多种高保真耐高温DNA聚合酶,大大降低了PCR过程中的碱基错配率。 另外,Taq DNA聚合酶具有末端转移酶作用,能在所合成DNA链的3′末端加上一个单独的腺苷酸残基(A)。这样的PCR产物可直接与带有3′-T的线性化载体(T载体)连接(即T-A克隆)。
。这样的PCR产物可直接与带有3′-T的线性化载体(T载体)连接(即T-A克隆)。")
33
(七)多核苷酸激酶可使寡核苷酸链5´羟基末端磷酸化
常用的是T4多核苷酸激酶(T4 polynucleotide kinase),该酶含5′多核苷酸激酶和3′磷酸酶活性,能催化ATP的γ-位磷酸向DNA和RNA的5′-羟基转移。 该酶常用于:① DNA或RNA的5′末端标记;② 使没有5′-末端磷酸的DNA片段磷酸化,以供连接和克隆之用。
,该酶含5′多核苷酸激酶和3′磷酸酶活性,能催化ATP的γ-位磷酸向DNA和RNA的5′-羟基转移。 该酶常用于:① DNA或RNA的5′末端标记;② 使没有5′-末端磷酸的DNA片段磷酸化,以供连接和克隆之用。")
34
Preparation of Interested DNA
第二节 目的DNA的获取 Preparation of Interested DNA

35
一、化学合成法可直接合成目的DNA 要求:已知目的基因的核苷酸序列或其产物的氨基酸序列 应用:一般用于小分子肽类基因的合成

36
* 化学合成法获取目的DNA 色 苯丙 蛋 赖 由已知氨基酸序列推测可能的DNA序列

37
二、从基因组DNA文库中获取目的DNA 第一步:构建基因组DNA文库(是指包含某一个生物细胞或组织全部基因组DNA序列的随机克隆群体,以DNA片段的形式贮存了所有的基因组DNA信息)。 第二步:筛选含有目的基因的克隆。
。 第二步:筛选含有目的基因的克隆。")
38
* 从基因组DNA文库获取目的基因 组织或细胞染色体DNA 限制性内切酶或机械剪切 基因片段 克隆载体 基因组DNA文库 存在于转化细胞内由克隆载体所携带的所有基因组DNA片段的集合 重组DNA分子 受体菌 含重组分子的转化菌

39
三、从cDNA文库中获取目的DNA 第一步:构建cDNA文库(包含某一组织细胞在一定条件下所表达的全部mRNA经反转录而合成的cDNA序列的克隆群体,它以cDNA片段的形式贮存了全部的基因表达信息)。 第二步:筛选含有目的cDNA的克隆。

40
cDNA文库 存在于转化细胞内由克隆载体所携带的所有cDNA片段的集合

41
基因组DNA文库和cDNA文库的构建

42
从基因组DNA文库或cDNA文库中筛选目的DNA的策略
1. 菌落或噬斑原位杂交 2. 针对表达文库,可用抗原-抗体或配体-受体结合的原理(亲和筛选法)筛选
筛选.")
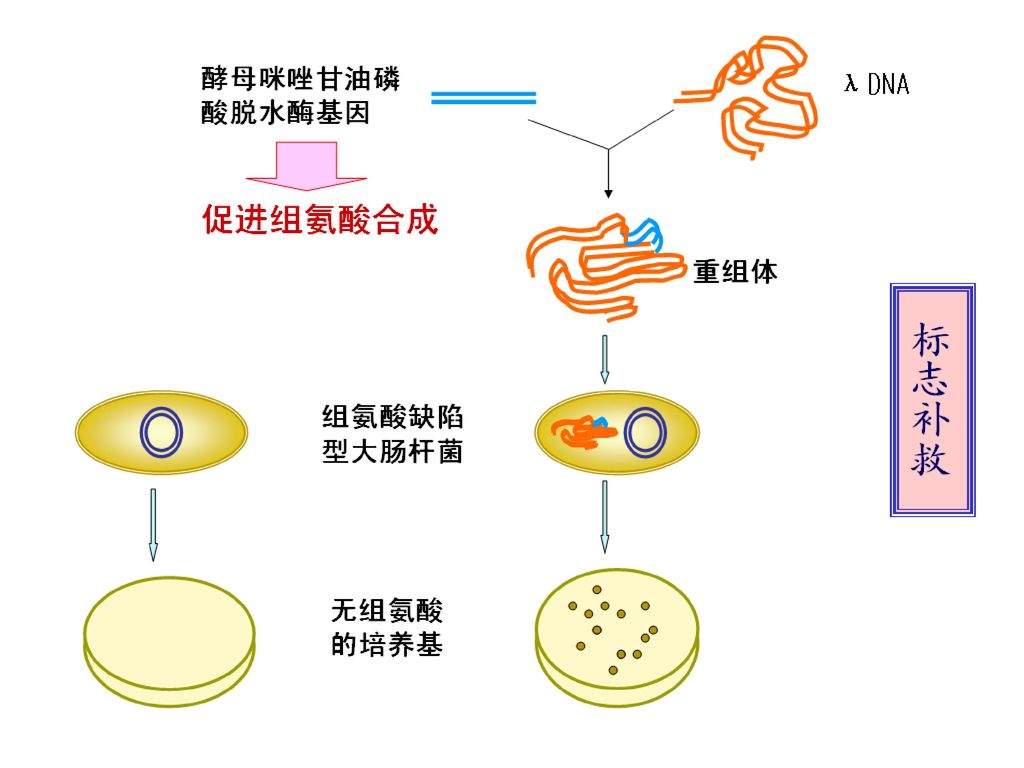
43
*从基因组DNA文库获取目的基因 菌落原位杂交法等方法,可从文库中筛选含有目的基因的噬菌体

44
AAAAAA 从cDNA文库获取目的基因 cDNA synthesis cDNA library Infect cells

45
四、经PCR获取目的DNA 如果已经知道目的基因的序列,就能很方便地用PCR反应,从基因组DNA或cDNA中获得目的基因,可不必要经过复杂的DNA文库构建过程。 PCR是一种高效快速的体外DNA聚合程序

46
PCR 体系 模板 引物 耐热DNA聚合酶 dNTPs 含Mg2+的缓冲液

47
PCR的过程 变性(Denaturing): 经加热使模板DNA 从 dsDNA 变成ssDNA
退火(Annealing): 引物与模板DNA杂交 延伸(Extension): DNA 合成
: 引物与模板DNA杂交. 延伸(Extension): DNA 合成.")
48
ing

49
PCR的头三个循环

50
五、通过特异杂交系统获取某些转录因子的编码DNA
报告基因 “诱饵”DNA序列 酵母单杂交系统 B. 有转录因子:报告基因表达 . 转录因子 报告基因 C. 有转录因子 AD/DNA结合蛋白-融合蛋白:报告基因表达 转录因子的 AD DNA结合蛋白 报告基因

51
. 酵母双杂交系统 A. 无相互作用的蛋白质,报告基因不表达 报告基因 启动子 B. 有相互作用的蛋白质,报告基因表达 报告基因 启动子
“猎物” “诱饵” AD 酵母双杂交系统 BD 报告基因 启动子 上游激活序列 B. 有相互作用的蛋白质,报告基因表达 “诱饵” “猎物” . AD BD 启动子 报告基因 上游激活序列

52
DNA Vectors in Recombinant DNA Technology
第三节 重组DNA技术中的DNA载体 DNA Vectors in Recombinant DNA Technology

53
载体概述 载体(vector)是为携带感兴趣的外源DNA,实现外源DNA在受体细胞中的无性繁殖或表达有意义的蛋白质所采用的一些DNA分子
克隆载体(cloning vector) 按功能分 表达载体(expression vector) 质粒载体 噬菌体载体 黏粒载体 病毒载体 人工染色体载体等 按基本元件的来源不同
 按功能分. 表达载体(expression vector) 质粒载体. 噬菌体载体. 黏粒载体. 病毒载体. 人工染色体载体等. 按基本元件的来源不同.")
54
克隆载体(cloning vector) 为使插入的外源DNA序列被扩增而特意设计的载体称为克隆载体。 表达载体(expression vector) 为使插入的外源DNA序列可转录和翻译成多肽链而特意设计的载体称为表达载体。
 为使插入的外源DNA序列被扩增而特意设计的载体称为克隆载体。 表达载体(expression vector) 为使插入的外源DNA序列可转录和翻译成多肽链而特意设计的载体称为表达载体。")
55
一、克隆载体用于外源DNA的克隆和无性繁殖
(一)克隆载体应具备的主要特点 至少有一个复制起点(origin of replication, ori) 至少有一个选择性标志(selection marker) 有适宜的限制性内切酶的单一切点:称为多克隆位点(multiple cloning sites,MCS) 应有较高的拷贝数。 pBR322质粒图谱
克隆载体应具备的主要特点. 至少有一个复制起点(origin of replication, ori) 至少有一个选择性标志(selection marker) 有适宜的限制性内切酶的单一切点:称为多克隆位点(multiple cloning sites,MCS) 应有较高的拷贝数。 pBR322质粒图谱.")
56
(二) 常用克隆载体有多种 1. 质粒 (plasmid)
特点:能在宿主细胞内独立自主复制;带有某些遗传信息, 会赋予宿主细胞一些遗传性状。

57
严紧型质粒:质粒与细菌染色体同步复制,处于严密控制之下,其拷贝数较低。
松弛型质粒:质粒的复制快于细菌染色体复制(即质粒复制不受严格控制),其拷贝数很高。重组DNA技术中使用的质粒通常是松弛型。 不同的质粒必须兼容才能共存于同一细胞中,这对复合转染(或转化)有参考价值。 目前,已有一系列商品化质粒克隆载体可供选择,如pUC系列、pGEM系列等。 质粒载体通常所能容纳的外源DNA长度在10 kb以内,外源DNA片段越长,质粒越不稳定。
,其拷贝数很高。重组DNA技术中使用的质粒通常是松弛型。 不同的质粒必须兼容才能共存于同一细胞中,这对复合转染(或转化)有参考价值。 目前,已有一系列商品化质粒克隆载体可供选择,如pUC系列、pGEM系列等。 质粒载体通常所能容纳的外源DNA长度在10 kb以内,外源DNA片段越长,质粒越不稳定。")
58
pUC系列质粒图谱

59
2. 噬菌体(phage) λ噬菌体DNA改造系统 M13噬菌体DNA改造系统(含lacZ基因) M13mp系列
λgt系列(插入型,适用于cDNA克隆) EMBL系列(置换型,适用于基因组DNA克隆) cos位点:λ噬菌体DNA为线性双链分子,两端各有一条由12个碱基组成、彼此完全互补且5′单链突出的序列(即粘性末端),称cos位点 M13噬菌体DNA改造系统(含lacZ基因) M13mp系列
 EMBL系列(置换型,适用于基因组DNA克隆) cos位点:λ噬菌体DNA为线性双链分子,两端各有一条由12个碱基组成、彼此完全互补且5′单链突出的序列(即粘性末端),称cos位点. M13噬菌体DNA改造系统(含lacZ基因) M13mp系列.")
60
3. 穿梭载体(shuttle vector) 人工构建,含有不止一个复制起点,能携带外源DNA序列在不同种类宿主细胞中复制、扩增。
 人工构建,含有不止一个复制起点,能携带外源DNA序列在不同种类宿主细胞中复制、扩增。")
61
二、表达载体用于外源DNA的表达 表达载体是指用来在宿主细胞中表达外源基因的载体 依据其宿主细胞的不同可分为:
原核表达载体(prokaryotic expression vector) 真核表达载体(eukaryotic expression vector)
 真核表达载体(eukaryotic expression vector)")
62
R:调节序列; P:启动子; SD:SD序列;TT:转录终止序列
(一) 原核表达载体用于在原核细胞中表达外源基因 从克隆载体发展而来,其除了具备克隆载体的一般特点外,还需有调控外源基因有效转录和翻译的序列,如启动子、核糖体结合位点、转录终止序列等。目前应用最广泛的原核表达载体是大肠杆菌表达载体。原核表达载体的基本组成如下: R:调节序列; P:启动子; SD:SD序列;TT:转录终止序列 大肠杆菌表达载体
 原核表达载体用于在原核细胞中表达外源基因. 从克隆载体发展而来,其除了具备克隆载体的一般特点外,还需有调控外源基因有效转录和翻译的序列,如启动子、核糖体结合位点、转录终止序列等。目前应用最广泛的原核表达载体是大肠杆菌表达载体。原核表达载体的基本组成如下: R:调节序列; P:启动子; SD:SD序列;TT:转录终止序列. 大肠杆菌表达载体.")
63
1. 启动子是启动外源基因表达的必需元件,其能被RNA聚合酶所识别:目前原核表达载体常使用的启动子有以下几种
Lac启动子(乳糖启动子) Trp启动子(色氨酸启动子) Tac启动子(乳糖和色氨酸的杂合启动子) PL和PR启动子(λ噬菌体的左向和右向启动子) T7启动子
 Trp启动子(色氨酸启动子) Tac启动子(乳糖和色氨酸的杂合启动子) PL和PR启动子(λ噬菌体的左向和右向启动子) T7启动子.")
64
2. SD序列提供核糖体结合位点 mRNA在细菌中的翻译严格依赖于核糖体结合位点(ribosome binding site)的存在。SD序列(Shine-Dalgarno sequence)位于转录起始位点上游8~13 bp处,为富含嘌呤的短片段,其能与核糖体30S小亚基中的16S rRNA 3′端的部分序列互补结合。SD序列作为核糖体结合位点,保证了翻译起始复合物的形成。
的存在。SD序列(Shine-Dalgarno sequence)位于转录起始位点上游8~13 bp处,为富含嘌呤的短片段,其能与核糖体30S小亚基中的16S rRNA 3′端的部分序列互补结合。SD序列作为核糖体结合位点,保证了翻译起始复合物的形成。")
65
3. 转录终止序列有助于外源基因的高效表达 尽管载体中没有转录终止序列,也可表达某些外源蛋白,但表达效果通常不理想。因此,多数原核表达载体中都带有转录终止序列。 转录终止序列长短不一,短的只有几十bp,长的可达几百bp。 转录终止序列有助于使RNA聚合酶重点转录克隆的外源基因,控制所转录RNA的长度,提高RNA稳定性。 位于启动子上游的转录终止序列可阻止其他启动子的通读,降低本底;位于多克隆位点下游的转录终止序列可防止因外源基因表达而干扰载体的稳定性。

66
4. 几种常见的原核表达载体各有特点 依据表达目的蛋白的方式,可将原核表达载体分为 (1)非融合型表达载体 (2)融合型表达载体
(3)分泌型表达载体
分泌型表达载体.")
67
非融合蛋白是指不与细菌的任何蛋白或多肽融合在一起进行表达的蛋白。表达该类蛋白的载体称为非融合型表达载体。
(1)非融合型表达载体用于表达非融合蛋白 非融合蛋白是指不与细菌的任何蛋白或多肽融合在一起进行表达的蛋白。表达该类蛋白的载体称为非融合型表达载体。 使用强启动子tac,紧接tac启动子的是多克隆位点,目的基因克隆于启动子和SD序列下游。在多克隆位点下游包含一个很强的rrnB核糖体RNA的转录终止子,其作用是为了稳定载体系统,因为上游强的tac启动子控制的转录必须由强终止子抑制,才不至于干扰与载体本身稳定性有关的基因表达。载体的其余部分来自pBR322。 使用pKK223-3时,应配套使用LacIq宿主菌,在无IPTG诱导时,tac启动子受阻遏,当加入IPTG后,便可去阻遏,从而使外源基因表达。
非融合型表达载体用于表达非融合蛋白. 非融合蛋白是指不与细菌的任何蛋白或多肽融合在一起进行表达的蛋白。表达该类蛋白的载体称为非融合型表达载体。 使用强启动子tac,紧接tac启动子的是多克隆位点,目的基因克隆于启动子和SD序列下游。在多克隆位点下游包含一个很强的rrnB核糖体RNA的转录终止子,其作用是为了稳定载体系统,因为上游强的tac启动子控制的转录必须由强终止子抑制,才不至于干扰与载体本身稳定性有关的基因表达。载体的其余部分来自pBR322。 使用pKK223-3时,应配套使用LacIq宿主菌,在无IPTG诱导时,tac启动子受阻遏,当加入IPTG后,便可去阻遏,从而使外源基因表达。")
68
(2)融合型表达载体用于表达融合蛋白 将一段特殊的蛋白质(或短肽)的基因构建入载体,使之与目的蛋白融合表达可形成融合蛋白(fusion protein)。表达该类蛋白的载体称为融合型表达载体,如pGEX系统。 pGEX系统包括多种载体,如pGEX-lλT、pGEX-2T、pGEX-3X、pGEX-4T、pGEX-5X、pGEX-6P等。该系统载体使用强启动子tac,紧接启动子的是SD序列及其下游的GST基因,然后是多克隆位点。用IPTG可诱导目的基因的表达,表达产物与GST融合,故可利用该标签对蛋白进行纯化。

69
(3)分泌型表达载体用于外源基因的分泌表达
该类载体的优点是能把在受体细菌内表达的外源蛋白有效地分泌到周质腔,甚至穿过细胞外膜进入培养基中。 在构建该类载体时,除有一般表达载体应有的启动子等元件外,还需含有编码信号肽的序列(通常置于SD序列下游)。 分泌型载体既可用于非融合蛋白的表达,也可用于融合蛋白的表达。 pIN Ⅲ系列载体是较常用的分泌型表达载体,可使所表达的蛋白分泌到宿主菌的周质腔中。该系列载体以pBR322为基础构建而成,带有大肠杆菌中最强的启动子之一,即lpp(脂蛋白基因)启动子,其中编码信号肽的序列取自大肠杆菌中分泌蛋白的基因ompa (外膜蛋白基因)。 pIN Ⅲ系列载体有3种,即pIN Ⅲ-ompA1、pIN Ⅲ-ompA2和pIN Ⅲ-ompA3,分别适用于使用不同密码子框架的目的基因的插入,克隆位点分别为EcoRⅠ、HindⅢ和BamHⅠ。
。 分泌型载体既可用于非融合蛋白的表达,也可用于融合蛋白的表达。 pIN Ⅲ系列载体是较常用的分泌型表达载体,可使所表达的蛋白分泌到宿主菌的周质腔中。该系列载体以pBR322为基础构建而成,带有大肠杆菌中最强的启动子之一,即lpp(脂蛋白基因)启动子,其中编码信号肽的序列取自大肠杆菌中分泌蛋白的基因ompa (外膜蛋白基因)。 pIN Ⅲ系列载体有3种,即pIN Ⅲ-ompA1、pIN Ⅲ-ompA2和pIN Ⅲ-ompA3,分别适用于使用不同密码子框架的目的基因的插入,克隆位点分别为EcoRⅠ、HindⅢ和BamHⅠ。")
70
(二)真核表达载体用于在真原核细胞中表达外源基因
真核表达载体包括在大肠杆菌中起作用的复制起始位点、抗生素抗性基因、多克隆位点等。 真核表达载体中还具有: ① 真核表达调控元件:包括启动子、增强子、转录终止序列、poly A 加尾信号等 ② 真核细胞复制起始序列 ③ 真核细胞药物抗性基因

71
真核表达载体的基本组成 OriPro:原核复制起始序列;P:启动子;MCS:多克隆位点;TT:转录终止序列;orieuk:真核复制起始序列。注:不是所有真核表达载体都有整合序列

72
真核启动子和增强子在不同类型细胞中的活性差别很大。某些来源于病毒的启动子和增强子,其真核宿主细胞范围较广,如Rous肉瘤病毒基因组长末端重复序列(long terminal repeats,LTR)、SV40病毒早期基因的启动子和增强子、人类巨细胞病毒(CMV)启动子等。这些启动子和增强子组合可在广泛的宿主细胞中起作用,故在真核表达载体中被普遍使用。
、SV40病毒早期基因的启动子和增强子、人类巨细胞病毒(CMV)启动子等。这些启动子和增强子组合可在广泛的宿主细胞中起作用,故在真核表达载体中被普遍使用。")
73
真核表达载体带有的poly A 加尾信号可保证新转录的mRNA能有效地加上poly A。
为mRNA加上poly A依赖于mRNA 3′末端的AAUAAA和其下游的GU或U富含区。 尽管全长cDNA克隆可能已带有AAUAAA序列和一段poly A,但这些序列仍不足以保证poly A的有效形成。因此,载体中必须带有poly A 加尾信号。 常用的来自SV40的poly A加尾信号为237 bp,其中同时含有针对早期和晚期转录子的切割和poly A 加尾信号。两套信号作用的方向相反,并分别位于不同的DNA链上,对mRNA的加工均很有效。

74
根据真核宿主细胞的不同,真核表达载体可主要分为 :
酵母表达载体 昆虫表达载体 哺乳类细胞表达载体

75
(1)酵母表达载体适于在酵母细胞中表达外源蛋白
根据载体在酵母细胞中复制方式的不同,可将酵母载体分为五类, YIp:酵母整合型质粒 YRp:酵母复制型质粒 YCp:酵母着丝粒型质粒 YEp:酵母游离型质粒 YLp:酵母线性质粒 除YLp外,其余各类既可在大肠杆菌,又可在酵母细胞中复制与扩增。

76
根据在转化酵母细胞中的复制机制,又可将酵母载体分为两个基本类型:
① 整合型载体,如YIp包含一个酵母ura3标志基因、大肠杆菌复制起始序列以及抗生素抗性基因。该质粒缺乏在酵母细胞中进行自主复制的元件,其需通过质粒DNA与酵母DNA同源序列重组而整合至酵母基因组中 。 ② 自主复制型载体,这类载体因在酵母细胞中有自我复制能力而得名,属于这类载体的有:YRp、YEp和YCp。

77
YRp含有酵母自主复制序列,但转化后缺乏有效的分离,故在有丝分裂后,质粒在亲代细胞分布极多而在子代细胞较少或丢失,在遗传学方面极不稳定。
YEp具有可诱导型的启动子、信号肽编码序列(常用交配因子α序列)和酵母质粒的2μm环片段,此类质粒可在染色体外自主复制(约50~100拷贝/细胞),其转化效率高且质粒稳定,因此常用该类载体在酵母细胞中高效表达外源基因。 YCp是由自主复制序列与着丝点序列连接而成,该类载体的特征是单拷贝(1~2拷贝/细胞),遗传性极其稳定。着丝点序列存在于酵母细胞每条染色体中,该序列在有丝分裂和减数分裂中能使染色体稳定而准确地复制,这是保持该类载体稳定的关键因素。
和酵母质粒的2μm环片段,此类质粒可在染色体外自主复制(约50~100拷贝/细胞),其转化效率高且质粒稳定,因此常用该类载体在酵母细胞中高效表达外源基因。 YCp是由自主复制序列与着丝点序列连接而成,该类载体的特征是单拷贝(1~2拷贝/细胞),遗传性极其稳定。着丝点序列存在于酵母细胞每条染色体中,该序列在有丝分裂和减数分裂中能使染色体稳定而准确地复制,这是保持该类载体稳定的关键因素。")
78
(2)昆虫表达载体可在昆虫细胞中表达真核基因
昆虫表达载体主要有两类: ①杆状病毒表达载体。载体的启动子为多角体蛋白(polyhedrin)基因的启动子,所使用的外泌信号序列来自蜜蜂的milittin序列,其可在宿主细胞(如Sf9或Sf20)中高水平表达外源蛋白。 ②果蝇表达载体。载体上的启动子有Ac5(果蝇黑素激动蛋白5C基因)启动子和MT(果蝇黑素金属硫蛋白基因)启动子,所使用的外泌信号序列为Bip序列,其可连续表达或诱导表达外源蛋白。
基因的启动子,所使用的外泌信号序列来自蜜蜂的milittin序列,其可在宿主细胞(如Sf9或Sf20)中高水平表达外源蛋白。 ②果蝇表达载体。载体上的启动子有Ac5(果蝇黑素激动蛋白5C基因)启动子和MT(果蝇黑素金属硫蛋白基因)启动子,所使用的外泌信号序列为Bip序列,其可连续表达或诱导表达外源蛋白。")
79
(3)哺乳类细胞表达载体适宜在哺乳类细胞中 表达真核基因
据载体进入宿主细胞的方式分为病毒载体和质粒载体 据载体在宿主细胞内是否整合于细胞染色体DNA,可将其分为整合型和非整合型载体 整合型载体一般是随机整合入染色体,但所携带外源基因的表达受插入位点的影响,同时还可能会改变宿主细胞的生长特性,整合型载体通常用于外源基因的稳定表达;非整合型载体通常用于外源基因的瞬时表达 最常用的哺乳类细胞表达载体是病毒载体

80
一个理想的病毒表达载体,通常应具备以下条件:
① 使用安全,对宿主细胞无毒副效应; ② 能容纳较大的外源DNA片段且转染效率高; ③ 所产生的病毒效价高; ④ 外源基因在靶组织或靶细胞中能长期、稳定表达 ⑤ 病毒的靶向性强; ⑥ 外源基因的表达具有可控性。 然而,遗憾的是,到目前为止,还没有任何一个病毒载体能满足以上全部理想条件。

81
病毒表达载体由各种病毒DNA衍生而来,其构建时一般都把细菌质粒复制起点放置其中,使病毒载体及其所携带的目的DNA能方便地在细菌中繁殖和克隆,然后再转入真核细胞。
经过质粒化改建的病毒载体通常都包括病毒启动子、包装元件、遗传标记和质粒复制起点等几个部分。

82
目前,常用的病毒表达载体包括如下多种: ① 反转录病毒(retrovirus,RV)载体 RV属ssRNA病毒。 构建RV载体时,将RV的DNA插入pBR322质粒,同时删除RV的三个编码基因,保留两端的LTR和病毒颗粒包装序列Ψ,再加入供筛选的标记基因。5′LTR具启动子和增强子活性并具转录起始信号,3′LTR具转录终止信号。 RV载体具有较高整合和表达外源基因的能力,广泛用于转基因动物和基因治疗研究,但因其随机整合,其安全性问题需加以考虑。

83
② 慢病毒(lentivirus,LV)载体
LV载体是以HIV-1(I型人类免疫缺陷病毒)为基础发展起来的表达载体; 与一般RV载体不同的是,它对分裂细胞和非分裂细胞均具有感染能力; 该载体可将外源基因有效地整合到宿主染色体上,从而获得持久表达; 该载体宿主细胞较广,在表达目的蛋白和小干扰RNA方面都有广泛应用 。
为基础发展起来的表达载体; 与一般RV载体不同的是,它对分裂细胞和非分裂细胞均具有感染能力; 该载体可将外源基因有效地整合到宿主染色体上,从而获得持久表达; 该载体宿主细胞较广,在表达目的蛋白和小干扰RNA方面都有广泛应用 。")
84
③ 腺病毒(Adenovirus,AV)载体
AV属线性dsDNA病毒; 目前,普遍使用的是AV第三代载体,载体中去除了所有AV的编码基因,仅保留了5′和3′末端反向重复序列(inverted terminal repeats,ITR)及病毒包装序列Ψ,病毒载体外壳的包装需要辅助病毒提供编码序列; 第三代AV载体能容纳较大的DNA片段,能较长时间地表达外源基因,其毒性和免疫原性都大大降低。
及病毒包装序列Ψ,病毒载体外壳的包装需要辅助病毒提供编码序列; 第三代AV载体能容纳较大的DNA片段,能较长时间地表达外源基因,其毒性和免疫原性都大大降低。")
85
④ 腺相关病毒(adenovirus-associated virus,AAV)载体
AAV属于线性ssDNA病毒。 构建AAV载体时,用外源基因及其调节序列取代病毒的rep和cap编码区,仅保留两端145 bp的ITRs,负责病毒的获救、复制、包装与整合;而辅助质粒则保留所有编码序列而无ITRs。两种质粒间无重复序列,重组AAV复制和壳化所需的rep和cap基因,由辅助质粒直接以蛋白表达方式提供,故不会发生野生型病毒序列的重组、复制和包装。当AAV载体与辅助质粒共转染辅助病毒(AV)感染的细胞时,即能获救、复制并包装成重组AAV 颗粒。 AAV载体具有能稳定表达外源基因、特异整合至人的19号染色体长臂末端、病毒稳定且宿主范围广、不引发免疫反应等优点 。
感染的细胞时,即能获救、复制并包装成重组AAV 颗粒。 AAV载体具有能稳定表达外源基因、特异整合至人的19号染色体长臂末端、病毒稳定且宿主范围广、不引发免疫反应等优点 。")
86
⑤ 痘苗病毒(vaccinia virus,VV)载体
VV属dsDNA病毒。 构建VV载体时,通常以胸苷激酶(thymidine kinase, tk)基因作为筛选标志。tk序列中插有该病毒的早期启动子和下游的多克隆位点。含有外源DNA的重组VV载体的tk基因不表达,当与野生型VV共转染宿主细胞并经tk同源序列重组,便可获得既有外源基因又有tk基因的重组VV颗粒,进而导入tk缺陷的细胞后,可在含有次黄嘌呤、氨蝶呤和胸苷的培养基中筛选,只有tk表达的细胞才能生长。 VV载体具有克隆容量大(可插入25~40 kb的外源DNA片段)、可插入多个外源基因、病毒宿主细胞广且使用安全等优点。
基因作为筛选标志。tk序列中插有该病毒的早期启动子和下游的多克隆位点。含有外源DNA的重组VV载体的tk基因不表达,当与野生型VV共转染宿主细胞并经tk同源序列重组,便可获得既有外源基因又有tk基因的重组VV颗粒,进而导入tk缺陷的细胞后,可在含有次黄嘌呤、氨蝶呤和胸苷的培养基中筛选,只有tk表达的细胞才能生长。 VV载体具有克隆容量大(可插入25~40 kb的外源DNA片段)、可插入多个外源基因、病毒宿主细胞广且使用安全等优点。")
87
⑥ 猿猴空泡病毒40(simian vacuolating virus 40,SV40)载体
SV40的基因组为双链环状DNA 人工构建的SV40载体主要有重组病毒质粒载体和取代型重组病毒载体两种类型,前者是将SV40基因组复制起点序列插入质粒载体中,从而构建成病毒-质粒载体;后者是用外源基因取代病毒基因组中与其大小相当的一个片段,从而形成取代型重组病毒载体

88
⑦ 单纯疱疹病毒(herpes simplex virus,HSV)载体
HSV为线性DNA病毒 构建HSV载体时,通常由Ⅰ型HSV基因组改建而成 目前,HSV载体主要有两类,一类是即刻早期(immediate early,IE)基因缺陷型载体,通过缺失这些IE基因,降低了毒性,从而使外源基因表达时间延长;另一类是利用该病毒的扩增子(amplicons)制备的载体,该类载体仅含有HSV复制起点和包装序列以及调节序列。辅助质粒为装配型质粒,其除了缺失包装序列外,包含所有HSV基因组。两种质粒共转染宿主细胞后,产生感染性重组病毒颗粒,其中仅含有HSV基因组序列,所有与毒性相关的HSV蛋白均以低水平表达 HSV载体具有克隆容量大(可插入长达50 kb的外源DNA)、外源基因可获得稳定表达、整合入宿主基因组后无致癌危险、病毒宿主范围广且有较高的转移效率和效价
基因缺陷型载体,通过缺失这些IE基因,降低了毒性,从而使外源基因表达时间延长;另一类是利用该病毒的扩增子(amplicons)制备的载体,该类载体仅含有HSV复制起点和包装序列以及调节序列。辅助质粒为装配型质粒,其除了缺失包装序列外,包含所有HSV基因组。两种质粒共转染宿主细胞后,产生感染性重组病毒颗粒,其中仅含有HSV基因组序列,所有与毒性相关的HSV蛋白均以低水平表达. HSV载体具有克隆容量大(可插入长达50 kb的外源DNA)、外源基因可获得稳定表达、整合入宿主基因组后无致癌危险、病毒宿主范围广且有较高的转移效率和效价.")
89
⑧ 其他病毒载体 除上述病毒载体外,目前用于构建真核表达载体的病毒还有人巨细胞病毒、脊髓灰质炎病毒等。此外,还有人试图利用乙型肝炎病毒等构建真核表达载体。

90
三、特殊载体具有特定用途 (一)黏粒兼具质粒和噬菌体DNA的特性
黏粒(cosmid):又称粘性质粒或装配型质粒,是由λ DNA的cos区与质粒重新构建而成的杂合双链环状DNA载体。 黏粒的特点:① 含有质粒的抗性标记基因及复制起点和多克隆位点,可按质粒的方式在宿主菌中进行自主复制;② 带有λ噬菌体DNA的cos序列,可像λ噬菌体DNA一样进行体外包装;③ 黏粒本身较小,通常只有几kb,但能容纳高达50 kb的外源DNA片段;④ 非重组体黏粒很小,故不能在体外进行包装,所以有利于后续阳性克隆的筛选;⑤ 黏粒因缺乏全部λ基因,不会产生噬斑,但在选择培养基平板上形成菌落,这一点也与质粒载体一致。 黏粒pJB8图谱
:又称粘性质粒或装配型质粒,是由λ DNA的cos区与质粒重新构建而成的杂合双链环状DNA载体。 黏粒的特点:① 含有质粒的抗性标记基因及复制起点和多克隆位点,可按质粒的方式在宿主菌中进行自主复制;② 带有λ噬菌体DNA的cos序列,可像λ噬菌体DNA一样进行体外包装;③ 黏粒本身较小,通常只有几kb,但能容纳高达50 kb的外源DNA片段;④ 非重组体黏粒很小,故不能在体外进行包装,所以有利于后续阳性克隆的筛选;⑤ 黏粒因缺乏全部λ基因,不会产生噬斑,但在选择培养基平板上形成菌落,这一点也与质粒载体一致。 黏粒pJB8图谱.")
91
(二)BAC、PAC和YAC用于大片段DNA的克隆
细菌人工染色体(bacterial artificial chromosome,BAC)基于F质粒构建而成。BAC能容纳高达400 kb(一般为50 kb~250 kb)的外源DNA。 基于大肠杆菌P1噬菌体的载体(E.coli bacteriophage P1-based vector, PAC)与BAC的克隆容量相当。 酵母人工染色体(yeast artificial chromosome,YAC)载体能携带更大的DNA片段,其由酵母线性质粒(YLp)发展而来,含有染色体的两个端粒片段,能复制出线性DNA,其克隆容量可高达3 Mb(一般为500 kb~2Mb)。利用重组线性DNA转化细胞,可建成包含人全部基因组DNA的巨型YAC文库。 BAC、PAC和YAC的使用为人类基因组物理图谱和序列测定提供了强有力的工具,大大促进了人类基因组计划的完成。
基于F质粒构建而成。BAC能容纳高达400 kb(一般为50 kb~250 kb)的外源DNA。 基于大肠杆菌P1噬菌体的载体(E.coli bacteriophage P1-based vector, PAC)与BAC的克隆容量相当。 酵母人工染色体(yeast artificial chromosome,YAC)载体能携带更大的DNA片段,其由酵母线性质粒(YLp)发展而来,含有染色体的两个端粒片段,能复制出线性DNA,其克隆容量可高达3 Mb(一般为500 kb~2Mb)。利用重组线性DNA转化细胞,可建成包含人全部基因组DNA的巨型YAC文库。 BAC、PAC和YAC的使用为人类基因组物理图谱和序列测定提供了强有力的工具,大大促进了人类基因组计划的完成。")
92
端粒重复序列(telomeric repeat, TEL)
着丝粒(centromere, CEN) 自主复制序列(autonomously replication sequences, ARS) BamHⅠ切割后形成微型酵母染色体→EcoRⅠ切割形成染色体的两臂→外源DNA 插入该切点形成人工酵母染色体→转化入酵母菌→通过复制、分裂,克隆大片段DNA
 自主复制序列(autonomously replication sequences, ARS) BamHⅠ切割后形成微型酵母染色体→EcoRⅠ切割形成染色体的两臂→外源DNA 插入该切点形成人工酵母染色体→转化入酵母菌→通过复制、分裂,克隆大片段DNA.")
93
(三)启动子探针型载体用于分离和分析基因启动子
启动子探针型载体(即通常所说的报告基因载体)是一种快速分离和鉴定基因启动子的工具型载体 包含两个基本部分:转化单元和检测单元。其中,转化单元包含质粒克隆载体必备的转化系统(复制起点和抗生素抗性基因,用于选择被转化的细胞);检测单元则包括一个已失去转录功能且易于检测的遗传标志基因以及多克隆位点。 例如,pGL3-basic就是一个典型的启动子探针型载体,它不含启动子,含有一个荧光素酶(luciferase,luc)标志基因。如果插入片段中含有启动子序列,则luc表达,否则,luc不表达,故根据luc表达与否和表达水平的高低,即可判断插入片段中启动子序列的有无和活性强弱。 启动子探针型载体pGL3-basic图谱
是一种快速分离和鉴定基因启动子的工具型载体. 包含两个基本部分:转化单元和检测单元。其中,转化单元包含质粒克隆载体必备的转化系统(复制起点和抗生素抗性基因,用于选择被转化的细胞);检测单元则包括一个已失去转录功能且易于检测的遗传标志基因以及多克隆位点。 例如,pGL3-basic就是一个典型的启动子探针型载体,它不含启动子,含有一个荧光素酶(luciferase,luc)标志基因。如果插入片段中含有启动子序列,则luc表达,否则,luc不表达,故根据luc表达与否和表达水平的高低,即可判断插入片段中启动子序列的有无和活性强弱。 启动子探针型载体pGL3-basic图谱.")
94
在启动子探针型载体的基础上,还发展出了用于分离和分析其他基因表达调控元件(如增强子或衰减子)的载体。这类载体与启动子探针型载体的区别,就是自身含有启动子,故将空载体转入细胞,就有标记基因(如luc)表达。当在启动子上游或下游(乃至标记基因下游)插入外源DNA片段后,便可根据标记基因表达水平的改变来判断所插入的DNA片段中是否含有增强或抑制基因表达的调控元件。
的载体。这类载体与启动子探针型载体的区别,就是自身含有启动子,故将空载体转入细胞,就有标记基因(如luc)表达。当在启动子上游或下游(乃至标记基因下游)插入外源DNA片段后,便可根据标记基因表达水平的改变来判断所插入的DNA片段中是否含有增强或抑制基因表达的调控元件。")
95
(四)组织特异性表达载体使外源基因在特定组织表达
把某种组织特异性的调控序列(如启动子)构建于表达载体,使外源基因的表达严格受控于该调控序列,就形成了组织特异性表达载体 该类载体为研究特定组织的发育和针对特定组织或器官的基因治疗提供了有效手段 目前研究较多的有乳腺组织特异性表达载体,它是基因工程药物的生物反应器;肿瘤细胞特异性表达载体能在肿瘤细胞内有效表达毒性蛋白,从而特异性杀伤肿瘤细胞;神经组织特异性表达载体,对治疗早老性痴呆具有积极作用
构建于表达载体,使外源基因的表达严格受控于该调控序列,就形成了组织特异性表达载体. 该类载体为研究特定组织的发育和针对特定组织或器官的基因治疗提供了有效手段. 目前研究较多的有乳腺组织特异性表达载体,它是基因工程药物的生物反应器;肿瘤细胞特异性表达载体能在肿瘤细胞内有效表达毒性蛋白,从而特异性杀伤肿瘤细胞;神经组织特异性表达载体,对治疗早老性痴呆具有积极作用.")
96
(五)双启动子和串联启动子表达载体各有特殊用途
一般的表达载体只有一个启动子,为了特殊需要,可给表达载体组装两个启动子,构成双启动子表达载体,旨在方便地研究不同条件下的基因表达情况或分别启动不同基因的表达。 有时为了提高目的基因的表达水平,可将两个或两个以上的相同启动子串联在一起,构成串联启动子表达载体。

97
(六)某些表达载体用于表达非蛋白分子 有些表达载体不用于表达蛋白质,而是用于表达小分子干扰RNA(small interfering RNA,siRNA)或微小RNA(micro RNA,miRNA)等。 例如,目前,用于表达siRNA的载体已有多种(如pSilencer2.1-U6 neo,mU6pro等)。
。")
98
四、载体上的常用标志或标签 载体上的标志(marker):通常用于重组子或阳性克隆的筛选、鉴定 常见的标志有: ①抗生素抗性基因
b.用于哺乳类细胞阳性克隆筛选的抗生素抗性基因 ② 酶的编码基因 ③ 营养缺陷选择标志

99
标签(tag):其编码序列通常构建于表达载体,与目的基因位于同一阅读框内,这样就可使所表达的蛋白上带上标签肽。
标签肽大小不等,小的只有几个氨基酸残基,大的有几十kD。 标签肽常用于表达产物的分离、纯化与鉴定。 常用的tag序列包括 His:6~8个组氨酸 FLAG:八肽序列 Strep:链亲和素,八肽序列 HA:血凝素,九肽序列 Myc:十肽序列 T7:11肽序列 S:14肽序列 HSV:11肽序列 VSV-G:11肽序列 GST SPA:葡萄球菌蛋白质A MBP:麦芽糖结合蛋白 GFP:绿荧光蛋白 RFP:红荧光蛋白 VSV:vesicular stomatitis virus,口蹄疫病毒

100
第四节 DNA克隆的基本过程 Procedure of DNA Cloning

101
目的DNA的分离获取(分) 载体的选择与准备(择) 目的DNA与载体连接(接) 重组DNA转入受体细胞(转) 重组体的筛选与鉴定(筛)
 载体的选择与准备(择) 目的DNA与载体连接(接) 重组DNA转入受体细胞(转) 重组体的筛选与鉴定(筛)")
102
一、分离获取目的DNA有多种方法(分) DNA克隆的第一步是分离获取目的DNA。
目的DNA的来源有多种,包括基因组DNA、经mRNA反转录的cDNA、PCR扩增产物、化学合成法获得的DNA等。

103
二、根据DNA克隆的目的选择与准备 适宜载体(择)
进行DNA克隆的目的主要有二:① 获得目的DNA片段;② 获得目的DNA片段所编码的蛋白质 针对第一种目的,通常选用克隆载体;针对第二种目的,需选用表达载体 选择载体时,还要考虑目的DNA的大小、受体细胞的种类和来源等因素 选择载体时还需注意载体内应有适宜的多克隆位点

104
不同载体的克隆容量及适宜宿主细胞 载体 插入DNA片段 宿主细胞 质粒 <5~10 kb 细菌,酵母 λ噬菌体载体 细菌 黏粒
BAC ~400 kb YAC ~3Mb 酵母

105
三、目的DNA与适宜载体连接形成重组DNA(接)
(一)黏端连接为最适连接 1. 单一相同黏端连接会产生载体自连 配伍末端的连接情况与单一相同黏端连接相似
黏端连接为最适连接. 1. 单一相同黏端连接会产生载体自连. 配伍末端的连接情况与单一相同黏端连接相似.")
106
单一相同黏端连接产生载体自连 载体和目的DNA用EcoR I切割 DNA连接酶 粘性末端 质粒载体 目的DNA 重组 DNA 错误连接 +

107
BamH I

108
单一相同黏端连接存在如下缺点: 容易出现载体自身环化 目的DNA双向插入载体(即正向和反向插入) 多拷贝现象 采用碱性磷酸酶预处理线性化载体DNA,使之去磷酸化,可有效减少载体自身环化 目的DNA如果反向插入载体,虽不影响基因克隆,但却影响外源基因的表达

109
2. 定向克隆可有效避免载体自连和DNA片段的反向插入
Eco RⅠ切割位点 Bg lⅡ切割位点 + EcoRⅠ+ Bg lⅡ 双酶切 Eco RⅠ+ Bg lⅡ T4 DNA连接酶 重组体 定向克隆:将待克隆DNA分子和载体分子采用同一对限制性核酸内切酶切割后连接起来,可实现外源DNA片段的定向插入,此即定向克隆

110
3. 通过其他措施产生黏端进行连接 ① 人工接头法 ② 同聚物加尾法 ③ PCR法 ④T-A克隆法

111
人工接头(adaptor或linker)连接
由平端加上新的酶切位点,再用限制酶切割产生粘性末端,而后进行黏端连接。 人工接头(adaptor/linker ):是借助化学合成[和(或)结合退火的方法]而得到的含有一种或一种以上限制性内切酶切点的平端双链寡核苷酸片段。 5′---GGTG▼ AATTCACC---3′ 3′---CCACTTAA ▲GTGG---5′
:是借助化学合成[和(或)结合退火的方法]而得到的含有一种或一种以上限制性内切酶切点的平端双链寡核苷酸片段。 5′---GGTG▼ AATTCACC---3′ 3′---CCACTTAA ▲GTGG---5′")
112
人工接头(adaptor/linker)连接
Eco RⅠ 5´- 3´- CCGAATTCG GGCTTAAGC

113
5′---GGTG AAAAAAACACC---3′ 3′---CCACTTTTTTT GTGG---5′
同聚物加尾连接 在末端转移酶(terminal transferase)的作用下,在DNA片段末端加上同聚物序列,制造出黏端,再进行黏端连接。 加同聚物尾 5′---GGTG AAAAAAACACC---3′ 3′---CCACTTTTTTT GTGG---5′
的作用下,在DNA片段末端加上同聚物序列,制造出黏端,再进行黏端连接。 加同聚物尾. 5′---GGTG AAAAAAACACC---3′ 3′---CCACTTTTTTT GTGG---5′")
114
同聚物加尾连接 重组体 目的基因 载体DNA T4 DNA连接酶 5´ 3´ 限制酶或机械剪切 限制酶 3´ T(T)nT
A(A)nA 3´ λ-核酸外切酶 末端转移酶 + dATP + dTTP T(T)nT A(A)nA A(A)nA T(T)nT T4 DNA连接酶 重组体 3´ A(A)nA T(T)nT 3 ´
nA 3´ λ-核酸外切酶. 末端转移酶. + dATP. + dTTP. T(T)nT. A(A)nA. A(A)nA T(T)nT. T4 DNA连接酶. 重组体. 3´ A(A)nA. T(T)nT 3 ´")
115
PCR法产生黏端 针对目的DNA的5′ 和3′ 端,设计一对特异引物,在每条引物的5′ 端分别加上不同的限制性内切酶位点,然后以目的DNA为模板,经PCR 扩增便可得到带有引物序列的目的DNA,进而用相应限制性内切酶切割PCR产物,产生黏端,随后便可与带有相同黏端的线性化载体进行有效连接。

116
T-A克隆法 在使用Taq DNA聚合酶进行PCR时,扩增产物的3′ 末端可加上一个单独的腺苷酸残基(A)而成为黏端,这样的PCR产物可直接与带有3′-T的线性化载体(T载体)连接,此即T-A克隆。
而成为黏端,这样的PCR产物可直接与带有3′-T的线性化载体(T载体)连接,此即T-A克隆。")
117
(二)平端连接效率较低 载体 目的基因 限制性内切酶 限制性内切酶 重组体 T4 DNA连接酶 载体自连 目的基因 自连
平端连接效率较低 载体 目的基因 限制性内切酶 限制性内切酶 重组体 T4 DNA连接酶 载体自连 目的基因 自连")
118
为了提高连接效率,可采用提高连接酶用量、延长连接时间、降低反应温度、增加DNA片段与载体的摩尔比等措施

119
(三)粘-平末端的连接效率介于黏端和平端连接之间
粘-平末端连接是指目的DNA和载体之间通过一端为黏端、另一端为平端的方式进行连接 以该方式连时,目的DNA为定向插入载体(即定向克隆) 该连接方式的连接效率介于黏端和平端连接之间 可采用提高平端连接效率的措施来提高该方式的连接效率
 该连接方式的连接效率介于黏端和平端连接之间. 可采用提高平端连接效率的措施来提高该方式的连接效率.")
120
(四)快速连接法无需纯化DNA片段 该方法从低融点琼脂糖凝胶中直接切下所需的DNA片段,无需纯化,连同凝胶一起与载体DNA进行连接。本法省时省力,对DNA黏端和平端的连接均适用,但其连接效率明显低于常规连接,且DNA连接酶用量大。
快速连接法无需纯化DNA片段 该方法从低融点琼脂糖凝胶中直接切下所需的DNA片段,无需纯化,连同凝胶一起与载体DNA进行连接。本法省时省力,对DNA黏端和平端的连接均适用,但其连接效率明显低于常规连接,且DNA连接酶用量大。")
121
四、重组DNA转入受体细胞(转) 转化(transformation) 质粒或黏粒→细菌 (感受态细胞,competent cell)
质粒→酵母菌(真核细胞) 转染(transfection) 外源DNA→真核细胞(酵母除外) 噬菌体DNA→感受态细菌 感染(infection) 噬菌体颗粒→细菌 病毒颗粒→哺乳细胞
 转染(transfection) 外源DNA→真核细胞(酵母除外) 噬菌体DNA→感受态细菌. 感染(infection) 噬菌体颗粒→细菌. 病毒颗粒→哺乳细胞.")
122
受体菌条件 安全宿主菌 限制酶和重组酶缺陷 处于感受态(competent)
细菌的感受态( competent bacterium):细菌易于接纳外源物质的一种天然状态,在细菌的对数生长前期,为了分裂增殖的需要,细菌胞膜上会表达一些特殊的蛋白质,利于外源物质穿透胞膜,帮助细菌吸收外源营养物质。基因工程操作中,通过物理化学的方法也可使细菌处于感受态。处于该状态的细菌被称为感受态细胞。
:细菌易于接纳外源物质的一种天然状态,在细菌的对数生长前期,为了分裂增殖的需要,细菌胞膜上会表达一些特殊的蛋白质,利于外源物质穿透胞膜,帮助细菌吸收外源营养物质。基因工程操作中,通过物理化学的方法也可使细菌处于感受态。处于该状态的细菌被称为感受态细胞。")
123
将重组DNA转入受体细胞的常用方法 化学法: 氯化钙化学转化 物理法: 电击法 显微注射法 等

124
用氯化钙化学转化

125
电击法

126
显微注射法

127
五、筛选与鉴定重组DNA有多种方法(筛)
一般一个载体只携带某一段外源DNA,一个细胞只接受一个重组DNA分子。最后培养出来的细胞群中只有一部分、甚至只有很小一部分是含有目的序列的重组体。将目的重组体筛选出来就等于获得了目的序列的克隆 筛选与鉴定程序主要包括:先筛选出带有载体的克隆;然后筛选鉴定出带有重组载体的克隆;最后筛选鉴定出带有目的DNA的克隆 主要筛选和鉴定方法有遗传标志筛选法、序列特异性筛选法、亲和筛选法等

128
(一)借助载体上的遗传标志可快捷方便地进行筛选
1. 利用抗生素抗性标志可筛选出带有载体的重组子 将含有某种抗生素抗性基因的载体转入宿主细胞后,将细胞在含有相应抗生素的培养基中培养,无载体转入的细胞将被杀死,生长的细胞即是含有载体的细胞。至于细胞中的载体是否为含有目的DNA的重组载体,尚需进一步鉴定。

129
抗药性标志筛选 抗性平板

130
2. 利用标志补救筛选带有载体的重组子 标志补救(marker rescue)是指当载体上的标志基因在宿主细胞中表达时,通过互补宿主细胞的相应缺陷而使细胞在相应选择培养基中存活。利用该策略可初步筛选带有载体的重组子。 标志补救也可用于外源基因导入哺乳动物细胞后的阳性克隆的初筛。例如,当把带有dhfr标志基因的真核表达载体导入dhfr缺陷的哺乳类细胞后,则可使细胞在无胸腺嘧啶的培养基中存活,从而筛选出带有载体的克隆。要确定是否为带有重组载体的阳性克隆,还需进一步鉴定。

132
3. 利用不同遗传标志可筛选出带有重组载体的克隆
(1)利用抗性基因的插入失活可筛选带有重组载体的克隆 诸多载体都带有抗生素抗性基因,当外源DNA插入某一抗性基因内,便可使该抗性基因失活。借助该抗性标志及载体上的其他标志,便可筛选出带有重组载体的克隆。
利用抗性基因的插入失活可筛选带有重组载体的克隆. 诸多载体都带有抗生素抗性基因,当外源DNA插入某一抗性基因内,便可使该抗性基因失活。借助该抗性标志及载体上的其他标志,便可筛选出带有重组载体的克隆。")
133
插入失活筛选带有重组载体的克隆

134
(2)利用抗性基因的插入表达也可筛选带有重组载体的克隆 例如:质粒pTR262携带来自λ噬菌体的CI基因,该基因在正常情况下表达产生阻遏蛋白,从而使tetR基因不能表达。当在CI基因中插入外源DNA后,将导致CI基因失活,从而使tetR基因去阻遏而表达。如果细菌内含有插入表达的重组体,则可在含有四环素的培养基上生长。
利用抗性基因的插入表达也可筛选带有重组载体的克隆 例如:质粒pTR262携带来自λ噬菌体的CI基因,该基因在正常情况下表达产生阻遏蛋白,从而使tetR基因不能表达。当在CI基因中插入外源DNA后,将导致CI基因失活,从而使tetR基因去阻遏而表达。如果细菌内含有插入表达的重组体,则可在含有四环素的培养基上生长。")
135
(3)α-互补筛选利用了β-半乳糖苷酶功能互补的基因片段
α-互补:pUC、pGEM及M13mp等载体系列均携带β-半乳糖苷酶N端146个氨基酸残基(α片段)的编码序列,在该序列中含有多克隆位点;经过改造的宿主菌基因组DNA含有β-半乳糖苷酶C端(ω片段)的编码序列。只有当载体与宿主细胞同时共表达该酶的α和ω片段时,才能形成有活性的β-半乳糖苷酶,该现象称为α-互补(alpha complementation) 。 β-半乳糖苷酶可在诱导剂IPTG存在的情况下,使底物X-gal转变为蓝色产物,使细菌克隆或噬斑呈蓝色。当外源DNA片段插入载体的多克隆位点后,便不能表达α片段,转化细菌后不能分解底物,结果使菌落或噬斑呈现白色。因此,α-互补筛选又称蓝-白筛选 。
的编码序列,在该序列中含有多克隆位点;经过改造的宿主菌基因组DNA含有β-半乳糖苷酶C端(ω片段)的编码序列。只有当载体与宿主细胞同时共表达该酶的α和ω片段时,才能形成有活性的β-半乳糖苷酶,该现象称为α-互补(alpha complementation) 。 β-半乳糖苷酶可在诱导剂IPTG存在的情况下,使底物X-gal转变为蓝色产物,使细菌克隆或噬斑呈蓝色。当外源DNA片段插入载体的多克隆位点后,便不能表达α片段,转化细菌后不能分解底物,结果使菌落或噬斑呈现白色。因此,α-互补筛选又称蓝-白筛选 。")
136
α-互补筛选(蓝-白筛选)
")
137
α-互补(蓝-白筛选) 载体上有β-半乳糖苷酶N端编码序列, 细菌染色体DNA有β-半乳糖苷酶C端编码序列 Lac Z基因 MCS
X-Gal 蓝色 Lac Z基因 MCS α-互补(蓝-白筛选) 载体上有β-半乳糖苷酶N端编码序列, 细菌染色体DNA有β-半乳糖苷酶C端编码序列
 载体上有β-半乳糖苷酶N端编码序列, 细菌染色体DNA有β-半乳糖苷酶C端编码序列.")
138
4. 利用噬菌体的包装特性可初筛带有重组载体的克隆
λ噬菌体的一个重要遗传特性就是其在包装时对λ DNA大小有严格要求,只有重组λ DNA的长度达到其野生型长度的75%~105%时,方能包装形成有活性的噬菌体颗粒,进而在培养基上生长时呈现清晰的噬斑,而不含外源DNA的单一噬菌体载体DNA因其长度太小而不能被包装成有活性的噬菌体颗粒,故不能感染细菌形成噬斑。

139
(二)根据序列特异性可筛选出特定的重组DNA
根据序列特异性进行筛选的方法包括: 限制性内切酶法 PCR法 核酸杂交法 DNA测序法等

140
1. 限制性内切酶法是根据限制酶切图谱筛选特定DNA
针对初筛为阳性的克隆,提取其重组DNA,以合适的限制性内切酶进行消化,经琼脂糖凝胶电泳便可判断有无DNA片段的插入及插入片段的大小。同时,根据酶切位点在插入片段内部的不对称分布,可用该方法鉴定DNA片段的插入方向;进而可用多种限制酶制作和分析插入片段的酶切图谱。

141
限制性内切酶图谱分析 目的序列插入载体会使载体DNA限制性内切酶图谱(restriction map)发生变化,如插入的目的序列中有其他限制性内切酶位点,也能在酶切电泳图谱上观察到。
发生变化,如插入的目的序列中有其他限制性内切酶位点,也能在酶切电泳图谱上观察到。")
142
2. PCR法可直接鉴定目的DNA的存在 利用序列特异性引物,经PCR进行扩增,可鉴定出阳性克隆。 如果利用克隆位点两侧序列设计引物进行PCR,再结合序列分析,便能可靠地证实插入片段的方向、序列和阅读框的正确性。

143
3. 核酸杂交法常用于从文库中筛选目的DNA 常用方法是将转有外源DNA的菌落或噬斑影印到硝酸纤维膜上,细菌裂解后所释放出的DNA将吸附在膜上,将膜与标记的核酸探针杂交,通过检测探针的存在即可鉴定出含有重组DNA的克隆。该方法又称为菌落或噬斑原位杂交(in situ hybridizaiton)。根据核酸探针标记物的不同,可通过放射自显影、化学发光、酶作用于底物显色等方法来显示探针的存在位置(即阳性克隆的存在位置)。
。根据核酸探针标记物的不同,可通过放射自显影、化学发光、酶作用于底物显色等方法来显示探针的存在位置(即阳性克隆的存在位置)。")
144
菌落或噬斑原位杂交

145
Southern印迹(Southern Blot)
")
146
4. 核苷酸序列测定是最准确的鉴定目的DNA的方法
测定核苷酸序列的方法主要有双脱氧链末端终止法和化学降解法,尤其以前者最为常用。 针对已知序列,通过测序可明确序列和阅读框的正确性;针对未知序列,可明确序列顺序,为进一步研究提供依据。

147
戊 糖 (RNA) 1´ 2´ 3´ 4´ 5´ 核糖(ribose) (DNA) 脱氧核糖(deoxyribose)
 1´ 2´ 3´ 4´ 5´ 核糖(ribose) (DNA) 脱氧核糖(deoxyribose)")
148
5′端 3′端 C G A DNA的 序 列 测 定

149
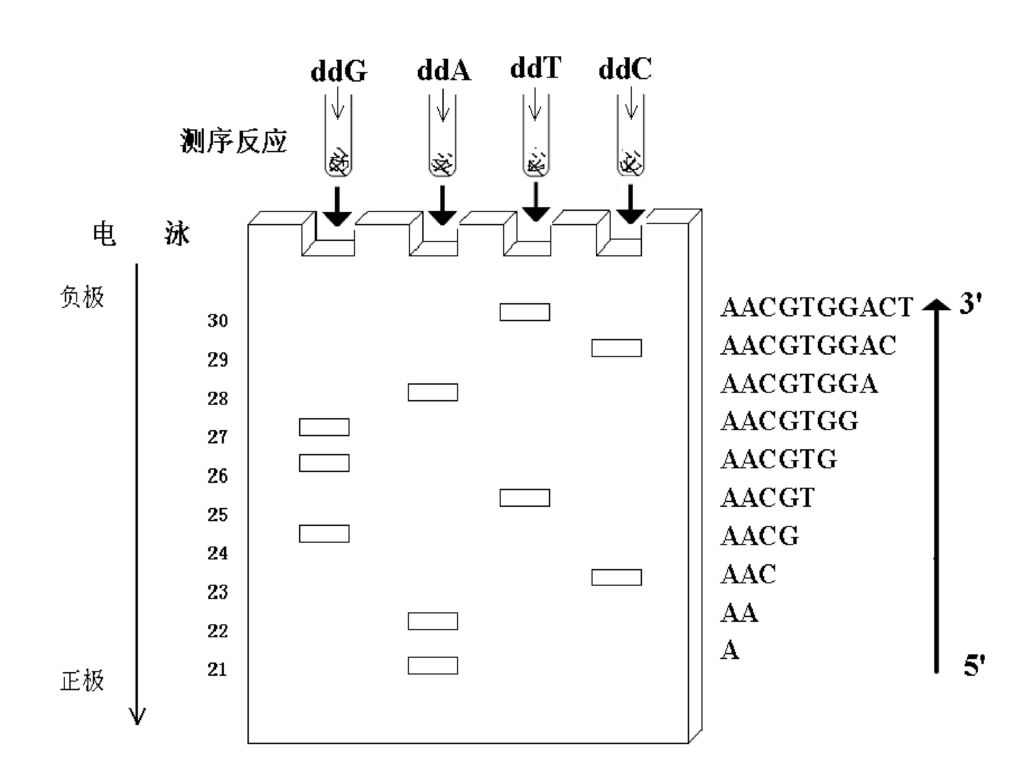
DNA链末端合成终止法

151
DNA自动测序 采用荧光替代放射性核素标记是实现DNA序列分析自动化的基础。用不同荧光分子标记四种双脱氧核苷酸(或同一测序引物),然后进行Sanger测序反应,反应产物经电泳(平板电泳或毛细管电泳)分离后,通过四种激光激发不同大小DNA片段上的荧光分子使之发射出四种不同波长荧光,检测器采集荧光信号,并依此确定DNA碱基的排列顺序。
,然后进行Sanger测序反应,反应产物经电泳(平板电泳或毛细管电泳)分离后,通过四种激光激发不同大小DNA片段上的荧光分子使之发射出四种不同波长荧光,检测器采集荧光信号,并依此确定DNA碱基的排列顺序。")
152
DNA自动测序结果举例

153
(三)亲和筛选法借助相关表达产物而筛选阳性克隆
亲和筛选法的前提是重组DNA进入受体细胞后能够表达出其编码产物。 常用的亲和筛选法的原理是基于抗原-抗体反应或配体-受体反应。一般做法同上述菌落或噬斑原位杂交相似,只是被检测的靶分子换成了吸附于硝酸纤维膜上的蛋白质,检测探针换成了抗体/抗原或配体/受体。 Ab

154
亚克隆(subcloning)及其目的 通过以上分、择、接、转、筛五个步骤,便完成了一次DNA克隆过程。有时为了达到某种新的目的,需要对已克隆的DNA进行再次克隆,该过程称为亚克隆。 亚克隆的目的有多种,常见的有: ① 实现目的DNA的高效扩增 ② 表达目的DNA编码的蛋白质 ③ 对目的DNA序列进行分析 ④ 对目的DNA进行修饰(如突变、缺失、引入新的限制酶切位点等) ⑤ 获得单链DNA或RNA ⑥ 鉴定目的DNA是否具有某些特殊功能
 ⑤ 获得单链DNA或RNA. ⑥ 鉴定目的DNA是否具有某些特殊功能.")
155
Frequently Used Prokaryotic andEukaryotic Expression Systems
第五节 常用的原核、真核细胞 表达体系 Frequently Used Prokaryotic andEukaryotic Expression Systems

156
一、原核细胞表达体系主要利用细菌 表达外源蛋白
原核表达体系主要以细菌作为宿主细胞,包括大肠杆菌、枯草杆菌、乳酸菌、沙门氏菌、苏云金杆菌等,其中又以大肠杆菌表达体系应用最为广泛和最成熟。原核表达体系既可用于原核基因表达,也可用于真核基因表达。

157
大肠杆菌 (Escherichia coli)
遗传背景清楚,基因工程操作方 便,商品化表达载体种类齐全,表达效率高 不具备真核细胞的蛋白质复性系统,真核基因在大肠杆菌中会合成错误空间构象的多肽链,易形成包涵体(无正确折叠的立体结构) 无真核生物的蛋白质加工系统,如糖基化磷酸化修饰等
 无真核生物的蛋白质加工系统,如糖基化磷酸化修饰等.")
158
枯草杆菌 (Bacillus subtilis)
分泌蛋白质能力强,一般有天然立体结构 无糖基化修饰功能,培养液中蛋白酶活性高,重组蛋白易受蛋白酶的水解 质粒不稳定,尚无商品化的表达载体

159
其他宿主菌 乳酸菌 (Lactic acid bacteria) 沙门氏菌 (Salmonella typhimurium) 苏云金杆菌(Bacillus thuringiensis) (可杀灭农林害虫,也可杀灭蚊、蝇等的幼虫,不伤害害虫天敌,对人和动物安全无害——微生物杀虫剂)
 沙门氏菌 (Salmonella typhimurium) 苏云金杆菌(Bacillus thuringiensis) (可杀灭农林害虫,也可杀灭蚊、蝇等的幼虫,不伤害害虫天敌,对人和动物安全无害——微生物杀虫剂)")
160
外源基因在原核表达体系中 表达的必要条件:正常的转录与翻译
(1)将外源基因置于强启动子和SD顺序(核糖体结合位点)控制下 (2)删除内含子和5′非编码区 (3)维持正确开放阅读框架(ORF) (4)mRNA稳定且可被有效翻译和折叠,形成的蛋白质不被降解
将外源基因置于强启动子和SD顺序(核糖体结合位点)控制下. (2)删除内含子和5′非编码区. (3)维持正确开放阅读框架(ORF) (4)mRNA稳定且可被有效翻译和折叠,形成的蛋白质不被降解.")
161
(一)影响原核表达体系有效表达外源基因的因素有多个
1. 目的基因的组成序列是决定外源基因表达的关键因素 由于原核细胞缺乏转录后加工机制,无法切除内含子序列,故针对来自真核细胞的基因,只能选择cDNA,不能使用基因组DNA 真核基因mRNA无结合细菌核糖体的SD序列,cDNA起始密码子上游的非编码序列必须删除 目的基因mRNA中的密码子应是原核宿主细胞的偏嗜密码子,否则,表达效率会降低 要让目的基因恰好插在启动子最佳作用的距离,才有利于基因的表达

162
2. 表达载体是决定外源基因表达的另一关键因素
常用的原核表达载体有非融合表达载体、融合表达载体和分泌型表达载体 通常一个理想的原核表达载体应具备强而可控的启动子、核糖体识别位点和转录终止信号三个特征 应容易转入宿主菌 通过试用不同的载体,可选出对某一基因表达最理想者 有时,通过调整载体上SD序列与起始密码子之间的距离,可提高蛋白表达效率 选择表达载体时还要考虑目的蛋白的理化性质。对于分子量小、容易降解的蛋白质,通常采用融合表达载体,反之则可采用非融合表达载体。

163
3. 受体菌能明显影响目的基因的表达 受体菌通常是工程菌,即经过改造,使限制修饰系统和重组系统缺陷的细菌,同时不能感染寄生于其他种群而造成危害 最常用的受体菌是大肠杆菌工程菌 同一蛋白质在不同工程菌中的表达水平可能会有差异,通常需试用几种工程菌,从中选出最佳者 根据表达载体上启动子的类型和特性来选择受体菌,例如,若使用带有T7启动子的载体,受体菌需能表达T7噬菌体RNA聚合酶 有时,还要利用一些温度敏感基因或药物诱导基因来协调宿主菌的生长周期和目的基因的表达周期,以获得目的蛋白的最佳表达

164
4. 目的蛋白的存在形式也能明显影响目的基因的表达效率
在原核细胞(如大肠杆菌)中,所表达的重组外源蛋白的存在形式由表达载体和宿主菌的特性共同决定 主要存在形式包括:以包涵体(不溶性)形式存在于细菌胞质中;以可溶性形式存在于细菌胞质中;通过运输或分泌方式以可溶形式定位于细菌周质腔,甚至穿过细胞外膜进入培养基中 通常,以包涵体形式存在时,其表达量最高,但蛋白无活性,需先经变性步骤将蛋白纯化出来,进而经复性才能得到有活性的蛋白;以可溶形式或分泌形式表达时,其表达量较低,但蛋白有活性,可直接在天然条件下进行纯化而得到有功能的蛋白
中,所表达的重组外源蛋白的存在形式由表达载体和宿主菌的特性共同决定. 主要存在形式包括:以包涵体(不溶性)形式存在于细菌胞质中;以可溶性形式存在于细菌胞质中;通过运输或分泌方式以可溶形式定位于细菌周质腔,甚至穿过细胞外膜进入培养基中. 通常,以包涵体形式存在时,其表达量最高,但蛋白无活性,需先经变性步骤将蛋白纯化出来,进而经复性才能得到有活性的蛋白;以可溶形式或分泌形式表达时,其表达量较低,但蛋白有活性,可直接在天然条件下进行纯化而得到有功能的蛋白.")
165
① 更换启动子 提高原核表达水平及目的蛋白溶解性的策略: ② 改变诱导条件 ③ 使用偏嗜性密码子 ④ 使用蛋白酶缺陷的宿主菌
⑤ 进行分泌表达 ⑥ 采用融合表达 ⑦ 与分子伴侣或折叠酶共表达 ⑧ 调整SD序列与起始密码子之间的距离,使mRNA在核糖体上定位后,翻译起始密码子正好进入核糖体的P位

166
在天然状态下,呈低限转录水平(以乳糖操纵子的启动子Plac为例)
I DNA O P pol 阻遏基因 外源基因 mRNA 阻遏蛋白 无诱导物(IPTG)时
时.")
167
I P O pol 可通过IPTG诱导增强其转录水平 DNA 外源基因 启动转录 mRNA mRNA 阻遏蛋白 IPTG

168
IPTG诱导目标蛋白的表达

169
(二)原核表达体系既有优点也有不足 原核表达体系有以下优势: ① 宿主菌遗传背景和生理特性清楚,有多种工程菌株可供选择
② 原核细胞繁殖能力强,操作简便、省时 ③ 大规模发酵成本低,生产潜力大 ④ 表达水平一般较真核系统高,且下游工艺简单、易于控制

170
原核表达系统的主要缺点: ① 原核细胞缺乏真核细胞所特有的翻译后加工修饰系统(如糖基化、磷酸化等),因此,针对这些需经修饰的蛋白,无法用原核表达体系获得有活性的产物 ② 目的基因不能是含有内含子的DNA ③ 细菌本身产生的内毒素等热原不易去除干净,为产品纯化增加了难度 ④ 蛋白的高水平表达常形成包涵体,提取和纯化步骤繁琐,而且蛋白复性较困难,容易出现肽链的错误折叠等问题

171
包涵体及其特点 包涵体(inclusion body):存在大肠杆菌胞浆中的由不溶性蛋白质聚集折叠形成的晶状结构物 包涵体的特点
大小0.5~1.0µm 比重在1.2左右 内含复杂成分:错误折叠的表达蛋白质, RNA聚合酶的四个亚基,外膜蛋白,16sRNA,质粒DNA,脂类,糖类等 不溶于水,加入强的蛋白质变性剂方可溶解,例如6~8mol/L盐酸胍,9~10mol/L尿素

172
包涵体形成的原因 形成机制: 蛋白质合成速率 蛋白质的折叠状态 蛋白质本身的性质
真核来源的蛋白质在缺乏分子伴侣(molecular chaperone)时无法形成天然三维结构 大肠杆菌内部的还原环境不利于二硫键的形成 蛋白质本身的性质 电荷平均数 形成转角的氨基酸残基组分 蛋白质合成速率 在强启动子的作用下,外源蛋白质过快的翻译使得其无法产生正确的折叠,而形成包涵体蛋白
时无法形成天然三维结构. 大肠杆菌内部的还原环境不利于二硫键的形成. 蛋白质本身的性质. 电荷平均数. 形成转角的氨基酸残基组分. 蛋白质合成速率. 在强启动子的作用下,外源蛋白质过快的翻译使得其无法产生正确的折叠,而形成包涵体蛋白.")
173
包涵体形成的原因 具体因素 温度 表达水平 细菌遗传性状 异源蛋白质氨基酸序列 温度低更利于蛋白质的折叠 过量表达的异源蛋白质容易形成包涵体
某些大肠杆菌染色体上的热休克基因(Hsp、GroEL、PPIase等)表达产物有助于重组异源蛋白质的正确折叠 异源蛋白质氨基酸序列
表达产物有助于重组异源蛋白质的正确折叠. 异源蛋白质氨基酸序列.")
174
形成包涵体的优缺点 优点 缺点 易于分离;产量高;载体构建较简单;可保护蛋白质被蛋白酶降解;可保护宿主菌免受伤害。
蛋白质无活性,复性后也较难恢复其天然功能。无法剪切肽链N端的甲硫氨酸,从而使蛋白真实性受影响。

175
包涵体蛋白的复性(蛋白质的体外复性) 复性:使形成包涵体的表达蛋白恢复其天然结构与活性的过程称为包涵体蛋白质的复性。包括包涵体的溶解变性与蛋白质重折叠(refolding)两大操作。
 复性:使形成包涵体的表达蛋白恢复其天然结构与活性的过程称为包涵体蛋白质的复性。包括包涵体的溶解变性与蛋白质重折叠(refolding)两大操作。")
176
包涵体的溶解变性 原因: 包涵体中的蛋白质大部分以分子间和分子内错配的二硫键形成可逆性聚集体。因此,必须完全将这些错配蛋白质溶解并变性。
方法: 利用变性剂,包括清洗剂、促溶剂(强阳离子或强阴离子)、极端pH溶液、或混合溶剂等种类。实验室常用盐酸胍和尿素。
、极端pH溶液、或混合溶剂等种类。实验室常用盐酸胍和尿素。")
177
蛋白质重折叠 复性的原理:在适宜的蛋白质纯度、浓度,pH,离子强度和温度等条件下,逐渐降低变性剂的浓度,并在一些辅助因子的作用下,恢复蛋白质的天然结构与功能。 复性方法:稀释法、透析法、层析法等。 由大肠杆菌产生的包涵体中,重组异源蛋白质的复性率个别可达34%,一般不超过20%。

178
二、真核细胞表达体系利用多种 真核细胞表达外源蛋白
二、真核细胞表达体系利用多种 真核细胞表达外源蛋白 与原核表达体系相比,真核表达体系(特别是以哺乳类细胞作为宿主细胞的表达体系)具有如下优点: ① 目的基因既可是基因组DNA,也可是cDNA,转录后能进行加工 ② 目的基因的表达可受到更严格地调控,能对所表达的蛋白质进行加工修饰,确保二硫键的精确形成,能正确进行糖基化、磷酸化、寡聚体的形成等加工 ③ 可使蛋白进行分泌表达,从而有利于蛋白的分离纯化 ④ 所表达的目的蛋白不易被降解,且对宿主细胞的影响较小
具有如下优点: ① 目的基因既可是基因组DNA,也可是cDNA,转录后能进行加工. ② 目的基因的表达可受到更严格地调控,能对所表达的蛋白质进行加工修饰,确保二硫键的精确形成,能正确进行糖基化、磷酸化、寡聚体的形成等加工. ③ 可使蛋白进行分泌表达,从而有利于蛋白的分离纯化. ④ 所表达的目的蛋白不易被降解,且对宿主细胞的影响较小.")
179
与原核表达体系相比,真核表达体系也存在诸多不足:
① 宿主细胞繁殖速度慢,培养条件要求高 ② 蛋白表达水平低 ③ 整个操作过程复杂、费时、成本高

180
根据目的蛋白表达的时空差异,可将真核表达体系分为瞬时、稳定和诱导表达体系
在瞬时表达体系中,载体DNA不能整合到细胞基因组中,其随细胞分裂而逐渐丢失,目的蛋白的表达时限短暂 在稳定表达体系中,载体DNA因整合到细胞基因组中而稳定存在于细胞内,目的蛋白能持久、稳定表达 在诱导表达体系中,目的基因的转录受外源小分子诱导后才得以开放

181
常用的真核表达体系 酵母表达体系 昆虫细胞表达体系 哺乳类细胞表达体系 各种表达体系各有其优缺点,应根据具体需要进行选择

182
(一)酵母表达体系是最为成熟的真核细胞 表达体系
酵母表达体系兼具原核和真核表达体系的优点: ① 酵母是最简单的真核生物,其在某些方面与原核细胞类 似,繁殖速度快,培养和发酵等操作也较为简单,利于工业化生产 ② 多种酵母的遗传背景较清楚,基因表达调控机制研究得较为透彻 ③ 酵母具有真核细胞的特点,能识别内含子,能对蛋白质进行一定程度地翻译后修饰,并且有分泌功能,从而方便了蛋白的纯化 ④ 不产生毒素,安全性较好

183
① 酿酒酵母广泛用于面包和乳酪工业,被FDA列为安全菌
酿酒酵母表达系统的优点包括: ① 酿酒酵母广泛用于面包和乳酪工业,被FDA列为安全菌 ② 酿酒酵母的遗传背景完全清楚,便于操作与改造 ③ 基因工程中使用的酿酒酵母为营养缺陷型,通过标志补救可筛选出含有表达载体的重组细胞 酿酒酵母表达体系的不足: ① 发酵时产生的乙醇会影响酵母本身的生长以及蛋白质的产量和活性,故难于进行高密度发酵 ② 蛋白质的分泌效率低,大于30 kD的蛋白质几乎不分泌 ③ 重组蛋白常发生过度糖基化,从而丧失蛋白的正常功能 ④ 表达载体YEp不稳定,易丢失,YIp传代不高

184
毕赤酵母为甲醇型酵母(即能在以甲醇为唯一碳源和能源的培养基上生长),毕赤酵母表达体系具有如下优势:
诱导型强启动子AOX1 高密度发酵产量很高 多拷贝整合表达量大 高表达性 高稳定性 外源基因可稳定传代 高分泌性 分泌蛋白的能力强 可进行胞内或胞外表达 表达的外源蛋白易提纯 表达的外源蛋白质量高 高适用性 简单易行 操作简单方便、成本较低

185
毕赤酵母表达体系也有其不足: ① 对毕赤酵母的遗传背景尚不完全清楚,因此对其进行遗传改造的难度较大 ② 发酵周期长 ③ 因密码子的偏嗜性问题或转录提前终止等原因,致使某些蛋白低表达或不表达 ④ 所表达的某些外源蛋白仍然会遭受蛋白酶的降解 ⑤ 所表达的某些外源蛋白会发生过度糖基化

186
多形汉逊酵母也是一类甲醇型酵母 多形汉逊酵母表达体系的优点包括:
① 该酵母是一种耐热酵母,最适生长温度为37~43℃(在49℃仍能生长),因其生长温度范围宽而易于培养和控制 ② 该酵母具有特殊的甲醇代谢途径 ③ 有多种营养缺陷株可供利用,阳性克隆筛选方便 ④ 可在所表达的外源蛋白C端加上一固定序列(S/A/C- K/R/H-L),从而将蛋白定位于过氧化物酶体,这样既避免了蛋白酶对蛋白的降解,同时也减少了外源蛋白对细胞的毒害 ⑤ 表达载体可通过同源或非同源重组的方式,以多拷贝整合于宿主细胞基因组,从而提高表达量
,因其生长温度范围宽而易于培养和控制. ② 该酵母具有特殊的甲醇代谢途径. ③ 有多种营养缺陷株可供利用,阳性克隆筛选方便. ④ 可在所表达的外源蛋白C端加上一固定序列(S/A/C- K/R/H-L),从而将蛋白定位于过氧化物酶体,这样既避免了蛋白酶对蛋白的降解,同时也减少了外源蛋白对细胞的毒害. ⑤ 表达载体可通过同源或非同源重组的方式,以多拷贝整合于宿主细胞基因组,从而提高表达量.")
187
裂殖酵母也可用于外源基因的表达。该酵母是一类不能出芽生殖而只能以分裂和产孢子的方式繁殖的酵母(因此定名为裂殖酵母)。与前面几种酵母相比,它具有更多的与高等真核生物相似的特性。该系统使用的表达载体有游离型和整合型载体,其中以游离型载体使用更多。该系统可以表达胞内蛋白、膜蛋白和分泌蛋白,但分泌蛋白的表达较困难,尚需进一步探索。
。与前面几种酵母相比,它具有更多的与高等真核生物相似的特性。该系统使用的表达载体有游离型和整合型载体,其中以游离型载体使用更多。该系统可以表达胞内蛋白、膜蛋白和分泌蛋白,但分泌蛋白的表达较困难,尚需进一步探索。")
188
其他酵母表达体系也在尝试之中,如乳克鲁维氏酵母表达体系已用于人血清白蛋白、人白介素1β的表达;arxula adeninivorams酵母表达体系也已建立。
总之,酵母种类繁多,特色各异,目前,已经开发出来作为外源蛋白表达体系的只是其中极小部分,更多的还有待于深入研究。

189
(二)昆虫细胞表达体系能比较理想地表达 真核蛋白
昆虫是高等真核生物,其与哺乳类细胞类似,具有蛋白的翻译后修饰、加工以及转移外源蛋白的能力。与哺乳类细胞相比,昆虫细胞生长速度快,可不需CO2,甚至不需血清,易于悬浮培养,能较高水平地表达外源蛋白,可以胞内表达,也可以进行分泌性表达。 目前,昆虫细胞表达体系主要有杆状病毒表达体系和果蝇表达体系。

190
杆状病毒表达体系 以Sf9和Sf21细胞系以及家蚕为表达宿主。 表达载体的启动子采用多角体蛋白(polyhedrin)基因的强启动子。
由于杆状病毒的基因组很大,限制性内切酶位点多,难于直接进行常规克隆,所以一般采用将外源基因克隆于专门设计的转移载体上,与病毒DNA共转染细胞后,经同源重组和筛选而得到重组病毒。经重组病毒感染宿主细胞后,即可高水平表达目的蛋白。 一般情况下,杆状病毒表达体系所表达的外源蛋白只有少部分是分泌性的,大部分为非分泌性。为了解决这个问题,将热休克蛋白70与外源蛋白共表达可明显提高重组蛋白的分泌水平。

191
果蝇细胞表达体系 结合了昆虫细胞高水平表达和哺乳类细胞稳定表达且蛋白很少被降解的优点 以S2细胞为宿主细胞,该细胞可在无CO2和无血清条件下高密度培养,内源性表达的果蝇细胞蛋白不与哺乳类细胞蛋白相互作用,从而为蛋白功能研究提供了一个空白背景 所用表达载体上的启动子有Ac5启动子和MT启动子,以便重组蛋白能稳定或瞬时高水平表达;表达载体可以多拷贝整合于宿主细胞基因组,从而提高目的蛋白的表达量 表达载体可连续表达,也可诱导表达(常采用CuSO4作为诱导剂

192
(三)哺乳类细胞表达体系所表达的蛋白 最接近天然蛋白
哺乳类细胞表达体系的优势 能使外源蛋白进行更好地分泌表达;表达产物不易降解 能使蛋白更加精确地折叠成正确构象 能进行复杂的N型糖基化和准确的O型糖基化等多种翻译后加工,因而表达产物在分子结构、理化特性和生物学功能方面最接近于天然的高等生物蛋白质 哺乳类细胞表达体系的缺陷 外源蛋白表达效率低 技术操作困难 细胞培养成本和条件要求高 难以大规模生产

193
CHO细胞是应用最广泛的哺乳类基因表达受体细胞之一
常用的CHO细胞包括野生型和dhfr缺失突变株 该细胞的特点是:能进行准确的转录和翻译后修饰、加工;很少分泌自身的内源蛋白,适合蛋白质的分泌表达和胞内表达,便于下游产物分离纯化;对培养基的要求低,既可贴壁生长也可悬浮生长;外源基因被整合入宿主细胞染色体后能稳定保持,具有重组基因的高效扩增和表达能力

194
COS细胞是利用复制起点缺失的SV40基因组DNA转化非洲绿猴肾细胞系CV-1得到的细胞系
该细胞广泛用于外源基因的瞬时表达、基因表达调控和蛋白质结构与功能等的研究 COS细胞的优点是:能在无血清的条件下表达外源蛋白,易培养和转染;能使转染到该细胞中的带有SV40复制子的质粒快速扩增;能瞬时表达大量的外源蛋白

195
其他哺乳类宿主细胞 小仓鼠肾(BHK)细胞 犬肾的高分化内皮细胞株(MDCK) 人胚肾细胞293-HEK 小鼠皮肤成纤维细胞NIH/3T3
细胞 犬肾的高分化内皮细胞株(MDCK) 人胚肾细胞293-HEK 小鼠皮肤成纤维细胞NIH/3T3")
196
细胞名称 来源 已投放产品 BHK-21 地鼠幼鼠肾 凝血Ⅷ因子 CHO-K1 中国仓鼠卵巢 tPA,Ⅷ因子,EPO,G-CSF等 C127 小鼠乳腺肿瘤 GH MDCK 长耳狗肾脏 兽用疫苗 Namalwa Burkitt淋巴瘤病人的类淋巴母细胞 EPO,LT,tPA,IFN Vero 成年非洲绿猴肾 HBsAg,GH 鼠骨髓瘤细胞 骨髓瘤 tPA,抗体 COS SV40转化的绿猴肾细胞 CD34,CD69,TGF

197
Applications of Recombinant DNA Technology
第六节 重组DNA技术的应用 Applications of Recombinant DNA Technology

198
利用重组DNA技术生产有应用价值的药物是当今医药发展的一个重要方向,其有望成为21世纪的支柱产业之一
该技术一方面可用于改造传统的制药工业,例如,使用该技术可改造制药所需要的工程菌种或创建新的工程菌种,从而提高抗生素、维生素、氨基酸等药物的产量 另一方面就是利用该技术生产有药用价值的蛋白质/多肽与疫苗等产品。目前经该技术发展的蛋白质/多肽类药物与疫苗已逾百种

199
一、重组DNA技术广泛应用于发展蛋白质/多肽类药物与疫苗
产 品 名 称 主 要 功 能 组织纤溶酶原激活剂 抗凝、溶解血栓 凝血因子Ⅷ、Ⅸ 促进凝血、治疗血友病 粒细胞-巨噬细胞集落剌激因子 剌激白细胞生成 促红细胞生成素 促进红细胞生成,治疗贫血 多种生长因子 刺激细胞生长与分化 生长素 治疗侏儒症 胰岛素 治疗糖尿病 多种干扰素 抗病毒、抗肿瘤、免疫调节 多种白细胞介素 免疫调节、调节造血 肿瘤坏死因子 杀伤肿瘤细胞、免疫调节、参与炎症 骨形态形成蛋白 修复骨缺损、促进骨折愈合 超氧化物歧化酶 清除自由基、抗组织损伤 单克隆抗体 利用其结合特异性进行诊断、肿瘤靶向治疗 乙肝疫苗 预防乙肝 口服重组B亚单位霍乱菌苗 预防霍乱

200
二、重组DNA技术应用于真核细胞 转基因和基因打靶
转基因动物技术是指将外源基因导入到动物的组织细胞内,并使导入的基因通过遗传传给子代 转基因动物的制备步骤主要包括基因表达载体的构建、转基因动物的建立和鉴定、转基因动物品系的繁育 目前,转基因技术已成功用于建立多种人类疾病的动物模型(如肿瘤、糖尿病、高血压、关节炎等模型)、制备生产药用蛋白的转基因动物,即生物反应器(如可生产凝血因子Ⅸ的转基因绵羊等)、改良动物品种、基因治疗和器官移植研究等 在转基因动物中,以转基因小鼠(transgenic mouse)最为常见
、制备生产药用蛋白的转基因动物,即生物反应器(如可生产凝血因子Ⅸ的转基因绵羊等)、改良动物品种、基因治疗和器官移植研究等. 在转基因动物中,以转基因小鼠(transgenic mouse)最为常见.")
201
建立转基因小鼠的常用方法 直接将目的DNA显微注射到受精卵的雄性原核,然后植入假孕母鼠体内,使之发育成幼仔 将带有目的基因的胚胎干细胞注射到囊胚,然后在小鼠体内发育成幼仔

202
制备转基因小鼠和小鼠基因打靶的基本过程

203
(二)重组DNA技术可用于基因打靶 基因打靶(gene targeting)是利用同源重组原理,使靶基因失活或置换,从而观察目的基因的功能
通过同源重组失活或剔除某一基因,称基因敲除(gene knock-out) 通过同源重组使突变基因被置换,称基因敲入(gene knock-in) 基因打靶技术的发展趋势是:① 通过条件性基因敲除技术,在时间和空间上对基因敲除进行调控;② 发展满足大规模基因功能研究所需要的随机基因敲除技术;③ 通过定位引入技术在基因组上引入精细突变以研制精确模仿人类疾病的动物模型
 通过同源重组使突变基因被置换,称基因敲入(gene knock-in) 基因打靶技术的发展趋势是:① 通过条件性基因敲除技术,在时间和空间上对基因敲除进行调控;② 发展满足大规模基因功能研究所需要的随机基因敲除技术;③ 通过定位引入技术在基因组上引入精细突变以研制精确模仿人类疾病的动物模型.")
204
制备转基因小鼠和小鼠基因打靶的基本过程

205
三、重组DNA技术还应用于其他诸多方面 (一)重组DNA技术是提前完成人类基因组计划的主要手段
人类基因组计划(human genome project,HGP)的主要目标是揭示人类遗传信息的组成和表达,具体主要通过完成四张图谱(即遗传图、物理图、DNA序列图和转录图)来实现 重组DNA技术的快速发展和完善加速了HGP的完成,其中包括大片段DNA克隆、DNA的大尺度分析、全自动DNA序列测定等
的主要目标是揭示人类遗传信息的组成和表达,具体主要通过完成四张图谱(即遗传图、物理图、DNA序列图和转录图)来实现. 重组DNA技术的快速发展和完善加速了HGP的完成,其中包括大片段DNA克隆、DNA的大尺度分析、全自动DNA序列测定等.")
206
(二)该技术广泛应用于基因诊断,并在基因治疗方面具有广阔前景
可参照第二十八章学习

207
(三)遗传疾病的预防 1. 产前诊断 2. 携带者测试 3. 证候前诊断 4. 遗传病易感性
遗传疾病的预防 1. 产前诊断 2. 携带者测试 3. 证候前诊断 4. 遗传病易感性")
Similar presentations


:核蛋白体组成成分 转移 RNA ( tRNA ):转运氨基酸 信使 RNA ( mRNA ):蛋白质合成模板 不均一核 RNA ( hnRNA ):成熟 mRNA 的前体 小核 RNA ( snRNA ):>")




>")










>")

