Download presentation
Presentation is loading. Please wait.

1
第10章 酶的作用机制和 酶的调节

2
内容提要 酶的活性部位 酶催化的独特性质 影响酶催化效率的有关原因 酶活性的调节 同工酶

3
10.1 酶的活性部位 酶活性部位的特点 酶分子中直接和底物结合,并和酶催化作用直接有关的部位称为酶的活性部位(active site)或活性中心(active center)
或活性中心(active center)")
4
酶的活性部位 一般是特异的氨基酸残基比较集中的区域,即与酶活力直接相关的区域。 活性部位又分为:
1)结合部位(Binding site) ----负责与底物 的结合,决定 酶的专一性。 2)催化部位(Catalytic site) ----负责催化底物键的断裂形成新键,决定酶的催化能力。
结合部位(Binding site) ----负责与底物 的结合,决定 酶的专一性。 2)催化部位(Catalytic site) ----负责催化底物键的断裂形成新键,决定酶的催化能力。")
5
酶的活性部位-结合部位 左图:二氢叶酸还原酶底物结合部位与两个底物: NADP+ 四氢叶酸

6
酶的活性部位-结合部位 Gly216 Gly226 Ser189 Val216 Thr226 酶的底物结合部位决定酶的专一性

7
酶的活性部位-结合部位 右图:胰凝乳蛋白酶的催化部位(由His 57, Asp 102, Ser 195组成) Asp 102 His 57
 Asp 102 His 57")
8
酶的活性部位-结合部位 酶的活性部位的共同点: 1)活性部位通常只占整个酶分子体积的1%-- 2%。酶分子的催化部位一般只有2—3个氨
基酸残基组成。 2)酶的活性部位是一个三维实体。 3)酶的活性部位并不是和底物的形状正好互 补的,而是在酶和底物结合的过程中,底物 分子或酶分子,有时是两者的构象同时发生 了一定的变化后才互补的。
酶的活性部位是一个三维实体。 3)酶的活性部位并不是和底物的形状正好互. 补的,而是在酶和底物结合的过程中,底物. 分子或酶分子,有时是两者的构象同时发生. 了一定的变化后才互补的。")
9
酶的活性部位 4)酶的活性部位是位于酶分子表面的一个裂 缝(crevice)内。是相当疏水的区域。在裂 缝内底物有效浓度可达到很高。
5)底物通过次级键较弱的力结合到酶上。 6)活性部位具有柔性或可运动性。
底物通过次级键较弱的力结合到酶上。 6)活性部位具有柔性或可运动性。")
10
酶活性部位的特点

11
研究酶活性部位的方法 酶分子侧链基团的化学修饰法

12
研究酶活性部位的方法 选择一种化合物,当其与被研究的酶作用时能专门与活性部位氨基酸残基侧链基团共价结合,然后将这个带标记化合物的酶水解,肽键被打开,但标记化合物共价键不被打开,因此可以分离得到带有标签的肽段,即可分析出活性部位的氨基酸残基。 巯基、羟基、咪唑基、氨基、羧基和胍基 化学修饰法有一定的缺陷:化学修饰有可能使活性部位之外的某个氨基酸残基的侧链改变,而影响酶分子的正常结构,因而导致酶活性的丧失。

13
研究酶活性部位的方法 A.非特异性共价修饰 化学试剂已和活性部位基团结合的鉴别: 一.酶活力的丧失程度和修饰剂浓度成一定的比例关系。
二. 底物或与活性部位结合的可逆抑制剂可保 护共价修饰剂的抑制作用,此法不但可以肯定某种基团是必需基团,还可以确信此基团 位于酶的活性部位。

14
研究酶活性部位的方法 B.特异性共价修饰 某一化学试剂专一地修饰酶活性部位的某一氨基酸残基,使酶失活。
二异丙基氟磷酸(DFP)能专一性地与酶活性部位的丝氨酸残基的羟基共价结合,使酶活力丧失。 二异丙基磷酰化酶(DIP—酶)
能专一性地与酶活性部位的丝氨酸残基的羟基共价结合,使酶活力丧失。 二异丙基磷酰化酶(DIP—酶)")
15
研究酶活性部位的方法 DFP一般不与蛋白质反应,也不与胰凝乳蛋白酶原和变性的胰凝乳蛋白酶反应,它也不和天然胰凝乳蛋白酶活性部位Ser195以外的27个Ser结合,可见活性部位Ser处于一个特殊的结构中,对DFP敏感。 对含有DFP集团的片段进行分析,不仅知道该酶活性部位附近的氨基酸顺序,也可得知DIP标记在该酶分子的Ser195残基上。

16
研究酶活性部位的方法 C. 亲和标记(affinity labeling)法 合成与底物结构相似的共价修饰剂。
一.可以较专一地引入酶的活性部位,接近底物结合位点。 二.具有活泼的化学基团可以与活性部位的某一基团结合形成稳定的共价键。利用酶对底物的特殊亲和力将酶加以修饰标记,故称亲和标记。试剂称“活性部位指示试剂”。

17
胰凝乳蛋白酶 对甲苯磺酰—L—苯丙氨酸乙酯(TPE)是底物 对甲苯磺酰—L—苯丙氨酰氯甲基酮(TPCK)是它的亲和试剂 可使组氨酸咪唑基烷化。 邹承鲁 图p387
是底物 对甲苯磺酰—L—苯丙氨酰氯甲基酮(TPCK)是它的亲和试剂 可使组氨酸咪唑基烷化。 邹承鲁 图p387")
18
科学家有责任说明真相 科学道德败给了广告 中科院邹承鲁:
中国科学院资深院士、第三世界科学院院士,我国著名生物化学家邹承鲁11月23日凌晨5点在北京逝世,享年83岁。

19
邹承鲁1923年5月17日生于山东省青岛市,祖籍江苏无锡。
1941年,他毕业于由天津迁到重庆的南开中学高中部;同年,考入设在昆明的由北京大学、清华大学、南开大学三校联合组成的西南联合大学,就读于化学系。 1946年,在招考英庚款公费出国留学生的考试中,他以第一名的优异成绩被录取。赴英后,师从英国剑桥大学著名生物化学家D基林(Keilin)教授。 1951年,邹承鲁获英国剑桥大学生物化学博士学位。
教授。 1951年,邹承鲁获英国剑桥大学生物化学博士学位。")
20
1951年回国后与王应睐及汪静合作纯化了琥珀酸脱氢酶,并发现其辅基为与蛋白部分共价结合的FAD。此外,他们对呼吸链及其他酶系也进行了一系列的工作,为我国酶学及呼吸链的研究奠定了良好基础。
1958年,他参加发起人工合成胰岛素工作,并负责胰岛素A和B链的拆合。这项工作的完成确定了胰岛素全合成的路线,为胰岛素的人工合成做出了重要贡献。

21
定点诱变法 利用定点突变技术(site directed mutagenesis),改变编码蛋白质基因中的DNA序列,研究酶活性部位的氨基酸。
,改变编码蛋白质基因中的DNA序列,研究酶活性部位的氨基酸。")
22
定点诱变法 1987年,Craik将胰蛋白酶Asp102诱变为Asn102。 酶活性降低5000倍。
羧肽酶A中Tyr248原认为催化所必须,Tyr248 的密码子(TAT)→Phe (TTT),kcat 不变,Km高6倍。
→Phe (TTT),kcat 不变,Km高6倍。")
23
10.2 酶催化反应的独特性质 1)酶反应可分成两类:一类反应仅仅涉及到电子的转移,反应速率或转换数在108S-1数量级;另一类反应涉及到电子和质子两者或其他基团的转移,反应速率在103S-1数量级。 2)酶的催化作用是由氨基酸侧链上的功能基团和辅酶为媒介的。主要的是His、 Ser、Cys、Lys、Glu和Asp。
酶的催化作用是由氨基酸侧链上的功能基团和辅酶为媒介的。主要的是His、 Ser、Cys、Lys、Glu和Asp。")
24
酶催化反应的独特性质 3)酶催化反应的最适pH范围通常是狭小的。 4)与底物相比较,酶分子很大,而活性部位通常只比底物大一点。
5)酶除了具有催化反应所必须的活性基团外,还有别的特性,使反应进行得更有力。
酶除了具有催化反应所必须的活性基团外,还有别的特性,使反应进行得更有力。")
25
10.3 影响酶催化效率的有关因素 10.3.1酶和底物的邻近效应与定向效应
酶和底物复合物的形成过程既是专一性的识别过程,更重要的是分子间反应变为分子内反应的过程。

26
酶和底物的邻近效应与定向效应 邻近效应是指酶与底物结合形成中间复合物以后,使底物和底物(如双分子反应)之间,酶的催化基团与底物之间结合于同一分子而使有效浓度得以极大的升高,从而使反应速率大大增加的一种反应。 如乙酸对硝基苯脂以咪唑催化水解的反应。 底物有效浓度增加24倍,反应速率加快24倍。

27
酶和底物的邻近效应与定向效应 定向反应 是指反应物的反应基团之间和酶的催化基团与底物的反应基团之间的正确取位产生的效应。
羧基和酯之间,自由度愈小,愈能使他们邻近,并有一定的取向,反应速率就愈大。 如邻羟苯丙酸的内酯形成,当两个甲基取代了苯环邻近的碳原子上的氢,反应速率可提高2.5×1011。

28
底物和酶的邻近效应与定向效应

29
底物和酶的邻近效应与定向效应 咪唑催化对-硝基苯酚乙酸酯水解的反应: 将咪唑和对-硝基苯酚乙酸酯结合到一个分子上后,反应速率增加24倍。

30
10.3.2底物的形变和诱导契合 当酶遇到其专一性底物时,酶中某些基团或离子可以使底物分子内敏感键中的某些基团的电子云密度增高或降低,产生“电子张力”,使敏感键的一端更加敏感,底物底物分子发生形变。 底物和酶结合时构象变化示意图

31
诱导契合学说 (1958, Daniel E. Koshland):
该学说认为酶表面并没有一种与底物互补的固定形状,而只是由于底物的诱导才形成了互补形状。

32
底物的形变和诱导契合

33
10.3.3 酸碱催化 酸碱催化是通过瞬时的向反应物提供质子或从反应物接受质子以稳定过渡态,加速反应的一类催化机制。
酸碱催化 酸碱催化是通过瞬时的向反应物提供质子或从反应物接受质子以稳定过渡态,加速反应的一类催化机制。 专一的酸碱催化或狭义的酸碱催化。 化学反应 无催化剂 催化剂(H+) 蔗糖水解 kJ/mol 104.7 kJ/mol
 蔗糖水解 kJ/mol kJ/mol.")
34
酸碱催化 总酸碱催化或广义的酸碱催化。 在生理条件下,因H+和OH-的浓度甚低,故体内的酶反应以总酸碱催化作用较为重要。
氨基、羧基、巯基、酚羟基及咪唑基。

35
酸碱催化 His 是酶的酸碱催化作用中最活泼的氨基酸残基。

36
共价催化 共价催化又称亲核催化或亲电子催化,在催化时,亲核催化剂或亲电子催化剂能分别放出电子或汲取电子并作用于底物的缺电子中心或负电中心,迅速形成不稳定的共价中间复合物,降低反应活化能,使反应加速。

37
共价催化 底物中典型的亲电中心,包括磷酰基、酰基和糖基 现已知100多种酶在催化过程中形成共价中间物。
底物与酶分子中亲核基团分别形成酰基—丝氨酸、酰基—半胱氨酸、磷酸丝氨酸、磷酸组胺酸和西佛碱。 磷酰基 酰基 糖基

38
金属离子催化 1/3的酶催化需要金属离子: ①金属酶:含紧密结合的金属离子 ②金属-激活酶: 含松散结合的金属离子

39
10.3.6 多元催化和协同效应 多种作用配合在一起起作用: 胰凝乳蛋白酶: Asp102 /His57/Ser159:
多元催化和协同效应 多种作用配合在一起起作用: 胰凝乳蛋白酶: Asp102 /His57/Ser159: 电荷中继网(亲核催化和碱催化)
")
40
活性部位微环境的影响 非极性环境中两个带电集团之间的静电作用比在极性环境中高,疏水微环境有利于酶的作用。

41
酶活性的调节控制 别构调节 酶原的激活 可逆的共价修饰

42
别构调节 酶分子的非催化部位与某些化合物可逆地非共价地结合后发生构像的改变,进而改变酶的活性状态,称为酶的别构调节(allosteric regulation) 别构酶 (allosteric enzyme) 正效应物(postitive effector)别构激活剂 负效应物(negative effector)别构抑制剂 同促效应 异促效应
别构激活剂. 负效应物(negative effector)别构抑制剂. 同促效应. 异促效应.")
43
别构调节 天冬氨酸转氨甲酰酶

44
别构调节

45
ATCase 的结构 调节亚基 c: 催化肽链;r:调节肽链 催化亚基

46
与底物结合后,ATCase两个催化肽链 的三级与四级结构变化图解

47
别构调控 CTP 是酶的抑制剂 ATP 是酶的激活剂
异促效应(heterotropic effect):非底物分子的调节物对别构酶的调节作用。
:非底物分子的调节物对别构酶的调节作用。")
48
别构酶的性质 一般都是寡聚酶 别构酶的动力学 协同指数(cooperativity index,CI):鉴别不同的协同作用及协同的程度。 又称饱和比值Rs
:鉴别不同的协同作用及协同的程度。 又称饱和比值Rs")
49
位点被90%饱和时的底物浓度 CI=Rs= ——————————————— 位点被10%饱和时的底物浓度 =811/n n协同系数(Hill系数) 米氏类型的酶 Rs=81 正协同效应的酶Rs<81 负协同效应的酶Rs>81

50
别构酶的性质

51
别构模型

52
别构模型

53
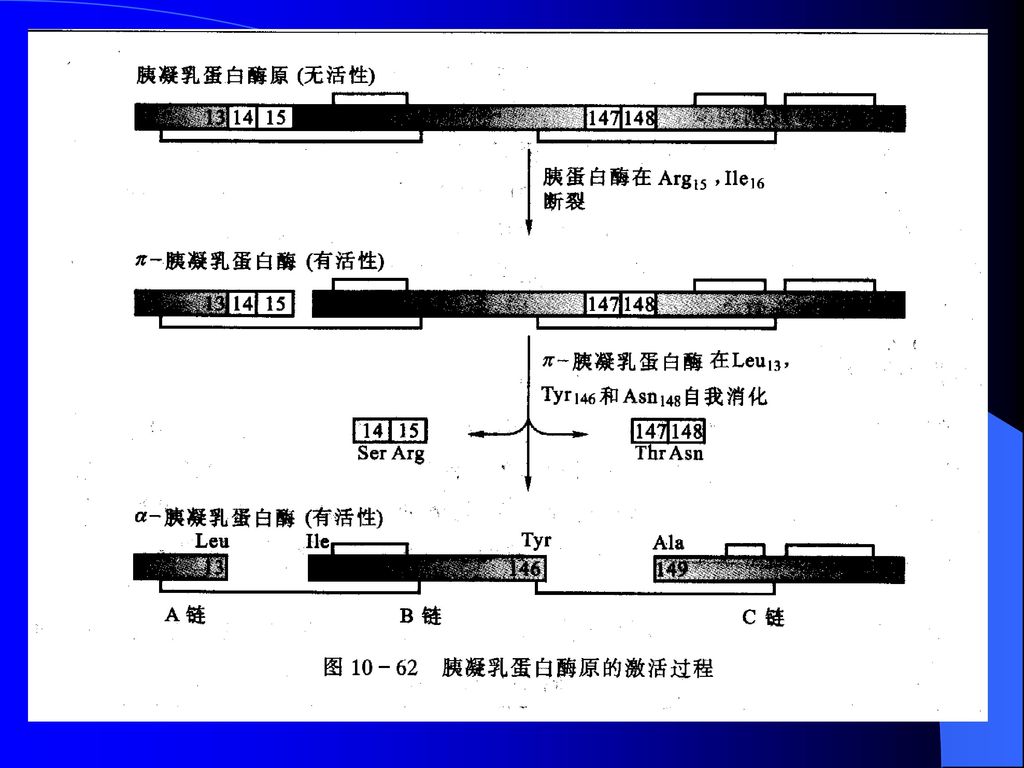
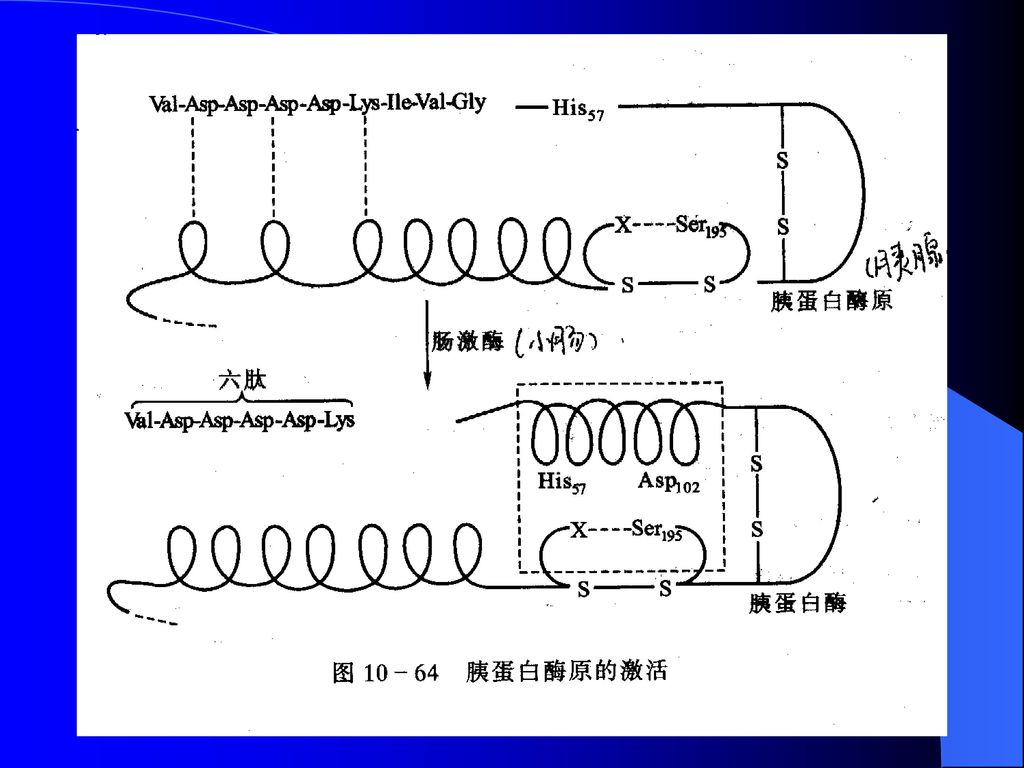
酶原的激活 消化系统蛋白酶原的激活 胰凝乳蛋白酶原(紫)与胰凝乳蛋白酶(绿)的构象比较
与胰凝乳蛋白酶(绿)的构象比较")
56
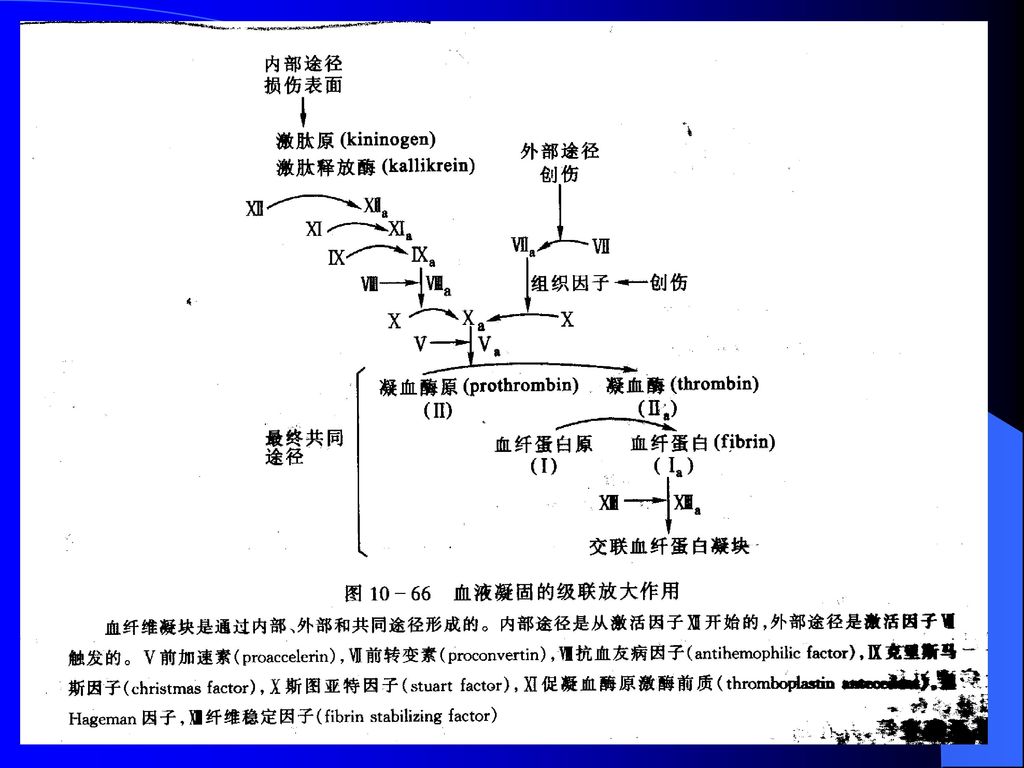
凝血机制 凝血机制: ※被伤害的血管收缩以减少血液的流失 ※血小板粘聚形成栓塞堵住伤口 ※通过一连串酶原激活反应和凝血因子 的作用使血液凝聚

58
可逆的共价修饰 通过共价调节酶进行 已知的修饰类型: ☆磷酸化 ☆腺苷酰化 ☆尿苷酰化 ☆ ADP-核糖基化 ☆甲基化

59
可逆的共价修饰

60
蛋白质的磷酸化 糖原磷酸化酶(无活性) 糖原磷酸化酶-P(有活性) 底物被蛋白质磷酸化的氨基酸有 Lys, Ser, Tyr
 糖原磷酸化酶-P(有活性) 底物被蛋白质磷酸化的氨基酸有 Lys, Ser, Tyr")
61
10.6 同工酶 乳酸脱氢酶

62
乳酸脱氢酶 LDH 同工酶 亚基组 LDH的活性百分数(%) 心肌 肝 肾 骨骼肌 血清 73 2 43 0 27.1
心肌 肝 肾 骨骼肌 血清 LDH1 LDH2 LDH3 LDH4 LDH5 H4 H3M H2M 2 HM 3 M 4

63
10 酶的作用机制和酶的调节 作业 名词解释 酶的活性中心,酶原,酶原激活,共价修饰,同工酶(举例) 回答问题
试述DFP对胰凝乳蛋白酶的修饰作用。 酶的活性中心有何特点? 影响酶催化效率的因素有哪些? 试述别构酶的特点。 试述乳酸脱氢酶的特点。 酶共价修饰的类型有哪些?

64
第十章完

Similar presentations



。 2、发酵工程在医药工业和食品工业中的 应用(A:知道)。>")










蛋白质的降解: 外源蛋白的消化 内源性蛋白的选择性降解 (2)氨基酸的分解代谢:>")




