Download presentation
1
第七章 微生物的遗传变异和育种

2
本章的学习目的及要求 1. 利用微生物的三个经典实验:转化实验、噬菌体感染实验和植物病毒的重组实验证明遗传变异的物质基础是核酸。了解遗传物质(DNA/RNA)在微生物细胞内的存在部位(核或核区、核糖体、质粒等)和功能特性,熟练掌握原核生物的质粒定义及几种典型质粒的特点。 2. 理解基因和基因组的概念;了解真核生物和原核生物在基因组结构、基因结构及遗传过程中的主要差别;了解微生物基因表达调控的相关元件及其功能,了解原核微生物基因表达调控的分子机制。 3. 掌握基因突变类型的定义、基因突变的特点以及基因突变自发性和不对应性的实验证明。

3
4. 掌握诱发突变、自发突变的机制,熟练掌握紫外线对DNA的损伤和修复机制。
5. 了解诱变育种的基本环节、原则,熟练掌握产量突变株、抗药性突变株、营养缺陷型突变株的筛选方法和基本原理,掌握艾姆氏实验的原理。 6. 掌握原核生物基因重组方式的种类及各类型的基本机制、相关概念等,灵活运用这些知识来解决一些实际问题。掌握准性杂交的定义、过程和生物学意义。 7. 掌握基因工程的定义和相关技术术语,熟悉基因工程的基本操作步骤。 8. 了解菌种衰退的原因;掌握复壮的定义和方法;了解菌种保藏原理;掌握常见菌种保藏方法、特点以及国际著名菌种保藏机构的名称。

4
1、遗传:指上一代生物如何将自身的一整套遗传基因稳定地传递给下一代的行为或功能。
2、遗传型:即基因型,指某一生物个体所含有的全部遗传因子即基因组所携带的遗传信息。 3、表型:指某一生物体所具有的一切外表特征和内在特性的总和,是其遗传型在合适环境条件下通过代谢和发育而得到的具体体现。 4、变异:指生物体在某种外因或内因的作用下所引起的遗传物质结构或数量的改变,即遗传型的改变。 5、饰变:指外表的修饰性改变,即一种不涉及遗传物质结构改变而只发生在转录、转译水平上的表型变化。

5
微生物的遗传和变异的特点: ①、微生物的代谢作用旺盛,有极高的繁殖速度;生活史周转快;环境因素可在短期内重复影响其生长和繁殖,易发生变异,又能迅速产生大量后代,有利于自然选择和人工选择。 ②、微生物细胞体积小,面积大,与外界环境能直接接触。当环境条件变化剧烈时,大多数个体易死亡而淘汰,个别细胞则发生变异而适应新环境。 ③、大多数微生物均进行无性繁殖,而且营养体多为单倍体,因而便于建立纯系及长期保存大量品系。如果一旦发生变异,也能够迅速在性状上反映出来。

6
第一节 遗传变异的物质基础 第二节 基因突变和诱变育种 第三节 基因重组和杂交育种 第四节 菌种的衰退、复壮和保藏

7
第一节 遗传变异的物质基础 一、证明遗传物质基础的三个经典实验 1、经典转化实验 创立人:
英国人Griffith于1928年首次发现这一现象。 研究对象: 肺炎链球菌S型和R型。 过程: ①动物实验; ②细菌培养试验; ③S型菌的无细胞抽提液试验; ④离体转化实验。

8
①动物实验

9
②细菌培养试验

10
③S型菌的无细胞抽提液试验 活R菌+S菌无细胞抽提液 长出大量R菌和少量S菌

11
④离体转化实验 1)从活的S菌中抽提各种细胞成分(DNA,蛋白质,荚膜多糖等); 2)对各组分进行转化实验:
从活的S菌中抽提各种细胞成分(DNA,蛋白质,荚膜多糖等); 2)对各组分进行转化实验:")
12
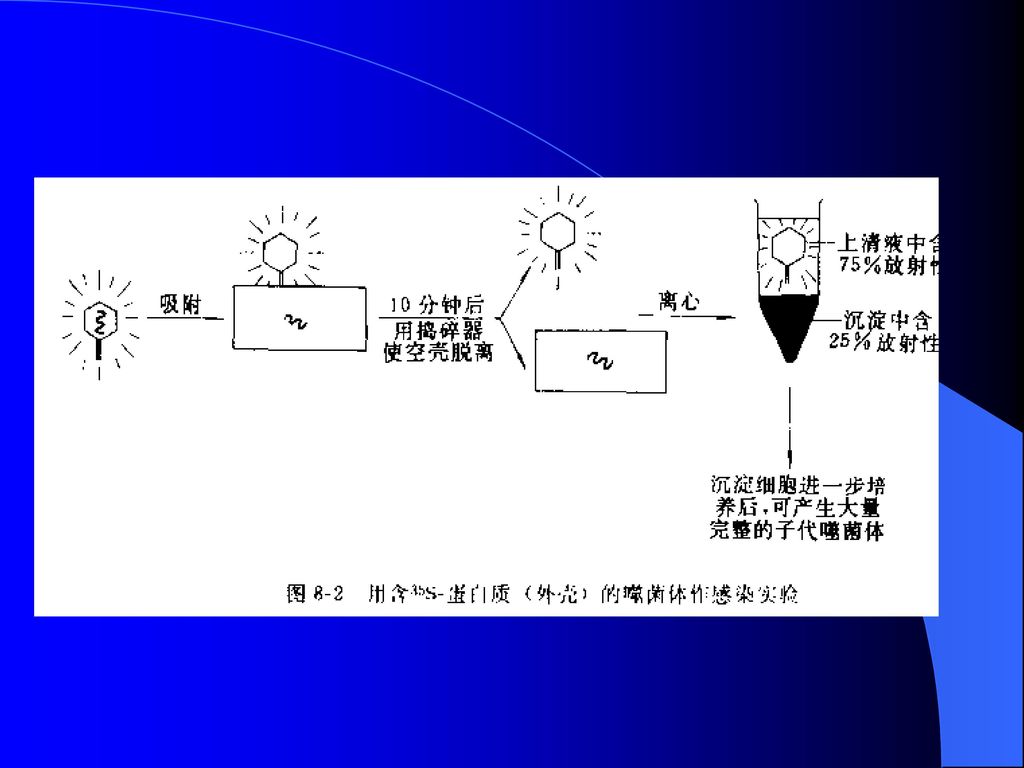
2、噬菌体感染实验 创立人:美国人Hershey AND Chase于1952年 研究对象:噬菌体 过程:

14
3、植物病毒的重建实验 创立人:Conrat AND Singer于1956年创立 研究对象:TMV AND HRV 过程:

15
关于朊病毒: 朊病毒(又称毒朊)是一类侵染动物、并在宿主细胞内复制的小分子无免疫性的疏水蛋白质。纯化的感染因子称为朊病毒蛋白( PrP )。 PrP是具有传染性的蛋白质致病因子,迄今未发现蛋白内有核酸,但已知的传染性疾病的传播必须有核酸组成的遗传物质,才能感染宿主并在宿主体内自然繁殖。那么这是生命界的又一特例呢?还是因为目前人们的认识和技术所限而尚未揭示的生命之谜呢?

16
这种传染性蛋白质颗粒是宿主细胞基因组基因编码的蛋白质分子的异构体,具有促使正常蛋白质分子变构成异构体的催化活性,从而造成朊病毒可以在宿主细胞中自身增殖的现象。

17
二、遗传物质在微生物细胞内存在的部位和方式
(一)细胞水平 集中在细胞核或核区。 (二)细胞核水平 1、核基因组 又称染色体基因组或基因组,是一个物种的单倍体的所有染色体及其所包含的遗传信息的总称。 (1)原核生物的基因组 1)小,为双链的DNA分子,一般具有单一的复制起点; 2)基因组上遗传信息具有连续性; 3)功能相关的基因构成操纵子或形成重叠基因,且常转录为多顺反子mRNA。 4)基因组的重复序列少而短; 5)结构基因的单拷贝及rRNA基因的多拷贝。
细胞水平. 集中在细胞核或核区。 (二)细胞核水平. 1、核基因组. 又称染色体基因组或基因组,是一个物种的单倍体的所有染色体及其所包含的遗传信息的总称。 (1)原核生物的基因组. 1)小,为双链的DNA分子,一般具有单一的复制起点; 2)基因组上遗传信息具有连续性; 3)功能相关的基因构成操纵子或形成重叠基因,且常转录为多顺反子mRNA。 4)基因组的重复序列少而短; 5)结构基因的单拷贝及rRNA基因的多拷贝。")
18
(2)真核微生物的基因组 1)典型的染色体结构,大,具有许多复制起点; 2)一个基因组包含若干染色体,一般不呈环状; 3)功能上密切相关的基因集中程度不如原核生物,很少有关于操纵子的报道,一般为单顺反子; 4)基因不连续,分外显子和内含子; 外显子:DNA序列中被转录成为mRNA的片段称为外显子。 内含子:在成熟mRNA上未反应出的DNA区段称为内含子。 5)有大量重复序列,有很多是不编码序列。
基因不连续,分外显子和内含子; 外显子:DNA序列中被转录成为mRNA的片段称为外显子。 内含子:在成熟mRNA上未反应出的DNA区段称为内含子。 5)有大量重复序列,有很多是不编码序列。")
19
2、核外染色体 (1)原核细胞的染色体外染色体:主要指质粒 ①定义: 游离于核基因组外,具有独立复制能力的小型共价闭合环状DNA分子,即cccDNA称为质粒。 ②性质 1)不相容性:亲缘关系较近的质粒在同一寄主细胞中不能稳定复制。 2)可传递性:质粒可在不同菌株间发生转移,与核染色体整合、脱离等。 3)可被消除性:在一些理化因子作用下,质粒的复制受到抑制而核染色体的复制仍继续进行,从而使子代细胞中的质粒消除。 4)可重组性:在质粒与质粒之间、质粒与染色体之间可发生重组。
可传递性:质粒可在不同菌株间发生转移,与核染色体整合、脱离等。 3)可被消除性:在一些理化因子作用下,质粒的复制受到抑制而核染色体的复制仍继续进行,从而使子代细胞中的质粒消除。 4)可重组性:在质粒与质粒之间、质粒与染色体之间可发生重组。")
20
③质粒的分子结构: 1) 具有麻花状的超螺旋结构,大小相当于核基因组的1%。 2) 质粒携带着某些染色体上所没有的基因,使细菌等原核生物被赋予了某些对其生存并非必不可少的特殊功能。 3)质粒是一种独立存在于细胞内的复制子。 a.严紧型复制控制:其复制行为与核染色体的复制同步。 b.松弛型复制控制:其复制行为与核染色体的复制不同步。

21
④质粒的种类 a. 致育质粒:控制宿主致育性的质粒。 b. 抗性质粒:是使其宿主微生物对抗生素、化学药物或重金属离子等杀菌剂表现出抗性的质粒。 c. 细菌素质粒 细菌素:是细菌产生的一般只能抑制或杀死种内不同亚种或菌株中敏感细菌的特殊多肽类代谢产物。 d. 降解质粒:是与微生物降解某些特殊的难分解的有机物质的能力有关的质粒。 e. 致病性质粒:是与宿主微生物对人、动物或植物的致病性有关的质粒。 f. 共生固氮质粒:是存在于根瘤菌中能控制它与相应豆科植物进行共生固氮作用的质粒。 g. 产生毒素的质粒:编码细菌毒素的质粒。 h. 代谢型质粒:系控制微生物的某一特殊代谢过程的质粒。 i. 隐蔽质粒:系指已经检测并从微生物细胞内分离到,但其表型效应尚未查清的质粒。

22
⑤典型质粒简介 1)F因子 (fertility factor):又称致育因子或性因子。 功能:决定细菌的性别,同时还具有转移能力。 结构: 转座因子:可整合到宿主染色体的一定部位,导致基因重组。 tra转移区:与质粒转移和性菌毛合成有关。

23
2)R因子(resistance factor):又称R质粒
a. 结构:由两个相连的DNA片段组成——RTF和r决定因子。 RTF功能:RTF即抗性转移因子。含调节DNA复制和拷贝数的基因以及转移基因,具有转移功能。 r决定因子功能:r决定因子即抗性决定因子。无转移功能,含各种抗性因子。 b. 作用: 引起致病菌对多种抗生素的抗性,对传染病的防治造成危害;R质粒可用做菌株筛选时的选择性标记或改造成外源基因的克隆载体。

24
3)Col因子(colicinogenic factors):
即产大肠杆菌素因子。 4)Ti质粒(tumor inducing plasmid): 即诱癌质粒。是植物遗传工程的主要载体。 5)巨大质粒(mega 质粒): 发现于根瘤菌属中,上面有一系列固氮基因。
Ti质粒(tumor inducing plasmid): 即诱癌质粒。是植物遗传工程的主要载体。 5)巨大质粒(mega 质粒): 发现于根瘤菌属中,上面有一系列固氮基因。")
25
(2)真核生物核外DNA 1)细胞质基因: 指线粒体和叶绿体等DNA,其结构与原核细胞的DNA相似,亦能编码结构蛋白。 2)共生生物: 如卡巴颗粒。 3)2u质粒: 存在于酿酒酵母的细胞核中,不能与核基因组整合。
真核生物核外DNA 1)细胞质基因: 指线粒体和叶绿体等DNA,其结构与原核细胞的DNA相似,亦能编码结构蛋白。 2)共生生物: 如卡巴颗粒。 3)2u质粒: 存在于酿酒酵母的细胞核中,不能与核基因组整合。")
26
(三)染色体水平 1、染色体数 不同物种,其染色体数目差别很大,但对同一物种来说,染色体的数目是恒定的。
染色体水平 1、染色体数 不同物种,其染色体数目差别很大,但对同一物种来说,染色体的数目是恒定的。")
27
2、染色体倍数:指同一细胞中相同染色体的套数。
(1)整倍体:指染色体的数目变异呈成套数目的改变,改变后的染色体数目是整倍数的(或者说细胞内含有完整即成套的染色体)。 ①单倍体:细胞核中含有一个完整染色体组的称为单倍体。 ②二倍体:细胞核中含有二个完整染色体组的称为二倍体。 ③多倍体:细胞核中含有超过两个染色体组的称为多倍体。 (2)非整倍体:是整倍体中缺少或额外增加一条或 n条染色体的变异型。
整倍体:指染色体的数目变异呈成套数目的改变,改变后的染色体数目是整倍数的(或者说细胞内含有完整即成套的染色体)。 ①单倍体:细胞核中含有一个完整染色体组的称为单倍体。 ②二倍体:细胞核中含有二个完整染色体组的称为二倍体。 ③多倍体:细胞核中含有超过两个染色体组的称为多倍体。 (2)非整倍体:是整倍体中缺少或额外增加一条或 n条染色体的变异型。")
28
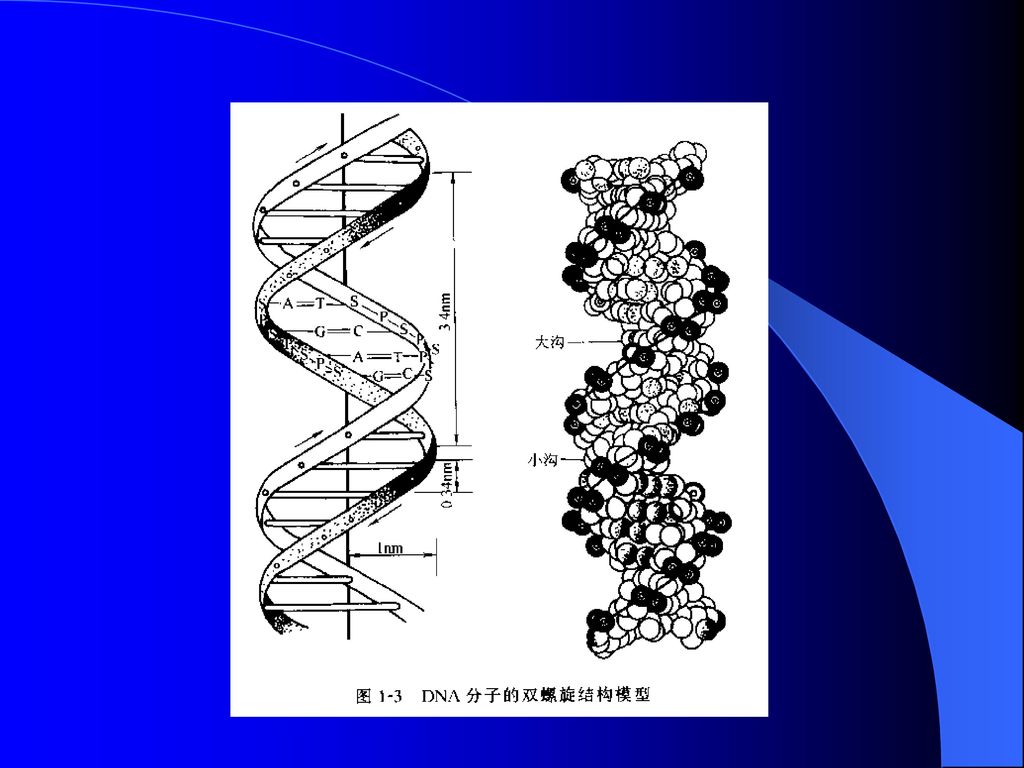
(四)核酸水平 1、 核酸种类:大多DNA,少数RNA。 2、DNA结构: (1)一级结构: 四种脱氧核糖核苷酸即脱氧腺嘌呤核苷酸、脱氧鸟嘌呤核苷酸、脱氧胞嘧啶核苷酸和脱氧胸腺嘧啶核苷酸,通过3’、5’—磷酸二酯键连接起来的直线形或环形多聚体。 (2)二级结构:指两条多核苷酸链反向平行盘绕而生成的双螺旋结构。 ①DNA分子是由两条互相平行的脱氧核苷酸长链盘绕而成的; ②DNA中脱氧核糖和磷酸交替连接,排在外侧,构成基本骨架,碱基排列在内侧; ③两条链上的碱基通过氢键相结合,形成碱基对,其组成按碱基互补配对原则即A与T结合,G与C结合。
二级结构:指两条多核苷酸链反向平行盘绕而生成的双螺旋结构。 ①DNA分子是由两条互相平行的脱氧核苷酸长链盘绕而成的; ②DNA中脱氧核糖和磷酸交替连接,排在外侧,构成基本骨架,碱基排列在内侧; ③两条链上的碱基通过氢键相结合,形成碱基对,其组成按碱基互补配对原则即A与T结合,G与C结合。")
30
(3)高级结构: 指DNA双螺旋进一步扭曲盘绕所形成的特定空间结沟。超螺旋结构是DNA高级结构的主要形式,可分为正超螺旋与负超螺旋两大类。 3、DNA长度:即基因组的大小用bp、kb、Mb表示。
高级结构: 指DNA双螺旋进一步扭曲盘绕所形成的特定空间结沟。超螺旋结构是DNA高级结构的主要形式,可分为正超螺旋与负超螺旋两大类。 3、DNA长度:即基因组的大小用bp、kb、Mb表示。")
31
(五)基因水平 1、基因概念 基因是一个含有特定遗传信息的核苷酸序列,是遗传物质的最小功能单位。 2、种类 (1)结构基因和调节基因:编码蛋白质的基因 结构基因→各种结构蛋白和催化各种生化反应的酶; 调节基因→阻遏蛋白或激活蛋白。 (2)核糖体RNA基因(rDNA)与转译RNA基因(tDNA):无翻译产物的基因 (3) 启动子与操纵基因:不转录的DNA区段
核糖体RNA基因(rDNA)与转译RNA基因(tDNA):无翻译产物的基因. (3) 启动子与操纵基因:不转录的DNA区段.")
32
3、原核生物基因调控系统 (1) 操纵子:功能相关的几个基因前后相连,在加上一个共同的调节基因和一组共同的控制位点(启动子、操纵基因等)在基因转录时协同动作,包含这些结构基因和控制区的整个核苷酸序列就称为操纵子。 1) 启动子:是一种能被依赖于DNA的RNA聚合酶的结合部位,也是转录的起始点。 2) 操纵基因:是位于启动基因和结构基因之间的一段碱基顺序,能与阻遏物相结合,以此来决定结构基因的转录是否能进行。 3) 结构基因:是决定某一多肽的DNA模板,可根据其上的碱基顺序转录出对应的mRNA,然后再可通过核糖体而翻译出相应的酶。 4) 调节基因:一般位于相应操纵子的附近,用于编码组成型调节蛋白。
 启动子:是一种能被依赖于DNA的RNA聚合酶的结合部位,也是转录的起始点。 2) 操纵基因:是位于启动基因和结构基因之间的一段碱基顺序,能与阻遏物相结合,以此来决定结构基因的转录是否能进行。 3) 结构基因:是决定某一多肽的DNA模板,可根据其上的碱基顺序转录出对应的mRNA,然后再可通过核糖体而翻译出相应的酶。 4) 调节基因:一般位于相应操纵子的附近,用于编码组成型调节蛋白。")
33
(2) 操纵子模型的基本内容 在调节基因产物的作用下,通过操纵基因控制结构基因的转录,从而发生酶的诱导或阻遏。
 操纵子模型的基本内容 在调节基因产物的作用下,通过操纵基因控制结构基因的转录,从而发生酶的诱导或阻遏。")
34
4、基因的命名——三字命名法 (1)基因名称一般用三个小写字母表示,三个英文字母取自表示该基因特性的一个或一组英文单词的前三个字母; 例:赖氨酸基因: lysine——lys——Lys (基因) (基因产物) 与核糖体中较大蛋白质亚基有关的基因 ribosomal protein large——rpl (2)产生同一突变型表型的不同基因,用三个字母后面所加的斜体大写字母表示; 例:组氨酸基因——his; 各组氨酸基因——hisA;hisB
产生同一突变型表型的不同基因,用三个字母后面所加的斜体大写字母表示; 例:组氨酸基因——his; 各组氨酸基因——hisA;hisB.")
35
(3)同一基因的不同突变位点用基因符号后面的阿拉伯数字表示;
例:色氨酸基因——trp 各个色氨酸基因——trpA;trpB 基因trp各位点的突变型——trpA23;trpA46 (4)抗性基因,一般把“抗”用大写R注在基因符号右上角。 (5)突变型基因的表示方法是在基因符号的右上角加-。 (6)当染色体上存在有缺失时,可用△表示,缺失部分放在△符号的括号中。 例: △(lac,pro)表示从乳糖发酵基因到脯氨酸基因这一段染色体发生了缺失。
抗性基因,一般把 抗 用大写R注在基因符号右上角。 (5)突变型基因的表示方法是在基因符号的右上角加-。 (6)当染色体上存在有缺失时,可用△表示,缺失部分放在△符号的括号中。 例: △(lac,pro)表示从乳糖发酵基因到脯氨酸基因这一段染色体发生了缺失。")
36
(六)密码子水平 1、三联子密码——遗传的信息单位 mRNA上每三个核苷酸翻译成蛋白质多肽链上的一个氨基酸,这三个核苷酸就叫密码,又称三联子密码。 2、遗传密码的性质 1)密码的简并性: 指一种以上密码子编码同一个氨基酸的现象。 2)密码的通用性: 所有的生物共用一套密码。 3)密码子与反密码子的相互作用——摆动学说(或称变偶假说) 在密码子和反密码子的配对中,前两对严格遵守碱基配对原则,第三对碱基有一定的自由度可以“摆动”,因而使某些 tRNA可以识别一个以上的密码子。
密码的通用性: 所有的生物共用一套密码。 3)密码子与反密码子的相互作用——摆动学说(或称变偶假说) 在密码子和反密码子的配对中,前两对严格遵守碱基配对原则,第三对碱基有一定的自由度可以 摆动 ,因而使某些 tRNA可以识别一个以上的密码子。")
37
3、密码子的使用规律 1)原核生物中大部分以AUG为起始密码子,少数使用GUG;真核生物则全部为AUG;终止密码子为UAA、UAG、UGA。 2)密码不重叠。 3)密码间无间隔。
原核生物中大部分以AUG为起始密码子,少数使用GUG;真核生物则全部为AUG;终止密码子为UAA、UAG、UGA。 2)密码不重叠。 3)密码间无间隔。")
38
第二节 基因突变和诱变育种 一、基因突变 简称突变,指细胞内(或病毒粒内)遗传物质的分子结构或数量突然发生的可遗传的变化。 (一)类型
1、根据表型划分

39
选择性突变—具有选择性标记,可通过某种环境条件使它们得到优势生长,从而取代原始菌株。
非选择性突变—无选择性标记,而只有一些性状的数量差别,如菌落大小、颜色深浅、代谢物产量等。 (1) 营养缺陷型:某一野生型菌株由于发生基因突变而丧失合成一种或几种生长因子的能力,因而无法在基本培养基上正常生长繁殖的变异类型,称为营养缺陷型。它们可在加有某生长因子的基本培养基平板上选出。 (2) 抗性突变型:由于基因突变而使原始菌株产生了对某种化学药物或致死物理因子抗性的变异类型。它们可在加有相应药物或用相应物理因子处理的培养基平板上选出。
 营养缺陷型:某一野生型菌株由于发生基因突变而丧失合成一种或几种生长因子的能力,因而无法在基本培养基上正常生长繁殖的变异类型,称为营养缺陷型。它们可在加有某生长因子的基本培养基平板上选出。 (2) 抗性突变型:由于基因突变而使原始菌株产生了对某种化学药物或致死物理因子抗性的变异类型。它们可在加有相应药物或用相应物理因子处理的培养基平板上选出。")
40
(3) 条件致死突变型:某菌株或病毒经基因突变后,在某种条件下可正常地生长、繁殖并实现其表型,而在另一种条件下却无法生长、繁殖的突变类型,称为条件致死突变型。
Ts突变株(温度敏感突变株):是一类典型的条件致死突变株,指可在某一温度下生长而在另一温度下不生长的突变株。 (4) 形态突变型:指由于突变而产生的个体或菌落形态所发生改变。 (5) 抗原突变型: 指由于基因突变而引起的抗原结构发生突变的变异类型 。 (6) 产量突变型:通过基因突变而获得的在有用代谢产物产量上明显有别于原始菌株的突变株,称为产量突变型。若产量高于原始菌株者,称为正变株,反之则称负变株。
:是一类典型的条件致死突变株,指可在某一温度下生长而在另一温度下不生长的突变株。 (4) 形态突变型:指由于突变而产生的个体或菌落形态所发生改变。 (5) 抗原突变型: 指由于基因突变而引起的抗原结构发生突变的变异类型 。 (6) 产量突变型:通过基因突变而获得的在有用代谢产物产量上明显有别于原始菌株的突变株,称为产量突变型。若产量高于原始菌株者,称为正变株,反之则称负变株。")
41
2、根据突变起因分

42
(1) 自发突变: 1) 自发突变:是指在没有人工参与下生物体自然发生的突变。称它为“自发”,决不意味着这种突变是没有原因的,而只是说明人们对它还没有很好的或很具体的认识而已。 2)自发突变的原因: ① 背景辐射和环境因素引起; ② 微生物自身有害代谢产物引起; ③ DNA复制中的错误;这种错误的概率约10-6。 (2) 诱发突变 简称诱变,是指通过人为的方法,利用物理、化学或生物因素显著提高基因自发突变频率的手段。
 诱发突变. 简称诱变,是指通过人为的方法,利用物理、化学或生物因素显著提高基因自发突变频率的手段。")
43
(二)突变率 1、概念:指某一个细胞(或病毒粒)每一个世代中发生某一性状突变的概率。也是突变在每个细胞生存的单位生物学时间(世代)内发生的概率。 2、测定: 应用选择培养法来直接测定突变率 。
突变率 1、概念:指某一个细胞(或病毒粒)每一个世代中发生某一性状突变的概率。也是突变在每个细胞生存的单位生物学时间(世代)内发生的概率。 2、测定: 应用选择培养法来直接测定突变率 。")
44
(三)基因突变的特点 1、自发性:各种性状的突变,可以在没有人为的诱变因素处理下自发地产生。 2、不对应性:指突变的性状与引起突变的原因间无直接的对应关系。 3、稀有性:自发突变虽可随时发生,但其突变率却是极低和稳定的,一般在l0-6~10-9之间。 4、独立性:突变的发生一般是独立的,某一基因的突变率不受它种基因突变率的影响。 5、可诱变性:自发突变的频率可因诱变剂的影响而大为提高。 6、稳定性:基因突变后的新性状是稳定的。 7、可逆性:由原始的野生型基因变异为突变型基因的过程,称为正向突变,相反的过程则称为回复突变或回变。任何性状都可发生正向突变,也都可发生回复突变。

45
突变是通过适应而产生的,突变的原因与性状间是相对应的。
(四)基因突变的自发性和不对应性的证明 突变是通过适应而产生的,突变的原因与性状间是相对应的。 PK 突变是自发的,与环境是不相对应的。
基因突变的自发性和不对应性的证明. 突变是通过适应而产生的,突变的原因与性状间是相对应的。 PK. 突变是自发的,与环境是不相对应的。")
46
1、变量实验 (1)实验内容: E.coli对于phageT1的抗性变异波动实验。
实验内容: E.coli对于phageT1的抗性变异波动实验。")
47
(2)原理: 通过小试管中抗性细菌数的波动情况证明细菌抗性的来历。如果抗药性的出现是由于细菌对于药物所发生的适应性的反应,那么分装在许多小试管中的样品将出现大致相等数目的细菌;如果抗药性的出现是由于基因突变,由于突变在时间上的随机性,不同试管中的抗性细菌数将会表现高度波动。
原理: 通过小试管中抗性细菌数的波动情况证明细菌抗性的来历。如果抗药性的出现是由于细菌对于药物所发生的适应性的反应,那么分装在许多小试管中的样品将出现大致相等数目的细菌;如果抗药性的出现是由于基因突变,由于突变在时间上的随机性,不同试管中的抗性细菌数将会表现高度波动。")
48
(3)结果: 从来自同一个培养物的大试管取样测定的抗性菌落数比较接近,而来自不同培养物(20支小试管)的样品的抗性菌落数的数值却相差很大。比较二者的平均值虽然相近,但后者的方差远远大于前者。 (4)结论: E.coli抗噬菌体性状的突变,不是由环境因素——噬菌体诱导出来的,而是在它接触到噬菌体前,在某一次细胞分裂过程中随机地自发产生的。这种自发突变发生得越早,则抗性菌落出现得越多,反之则越少。噬菌体在这里仅起着淘汰原始的未突变的敏感菌和甄别抗噬菌体突变型的作用。

49
2、涂布实验 (1)实验内容:
实验内容:")
50
(2)实验原理: 若抗性是在接触噬菌体以后产生的,那么喷噬菌体以前的重新涂布无非使细菌所处的位置发生改变,而不会因为涂布大大地改变细菌对于噬菌体的反应,因此两组培养皿上出现的抗性菌落数应相等;若抗性发生在接触噬菌体以前,那么在喷上噬菌体以前某些菌落中的抗性突变型细菌已经分裂若干次,在不经重新涂布的培养皿上形成一个菌落,在重新涂布的培养皿上出现若干菌落。 (3)结果:重新涂布的培养皿上出现的抗性菌落多余未经重新涂布的。 (4)结论:抗性突变的发生在接触噬菌体之前。
结果:重新涂布的培养皿上出现的抗性菌落多余未经重新涂布的。 (4)结论:抗性突变的发生在接触噬菌体之前。")
51
3、影印培养实验 (1)实验内容:
实验内容:")
52
(2)实验原理: 通过盖印章的方式,在未接触抗性因素的条件下筛选出的抗性突变株。由于实验过程中没有接触过抗性因素,所以直接证明了基因突变的自发性。 (3)结果:得到越来越多的抗性菌落,最终甚至可以得到纯的抗性菌株细胞群。。 (4)结论:基因突变的自发性。
结论:基因突变的自发性。")
53
(五)突变机制 1、诱发突变的机制 (1)点突变:即狭义的基因突变,指DNA的单个碱基发生替换所引起的突变。 1)碱基置换:一对碱基被另一对碱基所置换。 转换:即DNA链中的一个嘌呤被另一个嘌呤或是一个嘧啶被另一个嘧啶所置换 颠换:即DNA链中的一个嘌呤被另一个嘧啶或是一个嘧啶被另一个嘌呤所置换。

54
2) 移码突变: 指DNA分子中的一个或少数几个核苷酸的增添(插入)或缺失,从而使该部位后面的全部遗传密码发生转录和转译错误的一类突变。由移码突变所产生的突变株,称为移码突变株。 a. 由于在DNA分子的编码区插入或缺失非3的整数倍个(1个、2个或4个)核苷酸而导致阅读框架位移,导致翻译出来的蛋白质的氨基酸序列与野生型完全不同。 b. 如果插入或缺失的碱基正好是3个或其整倍数,那么在翻译出的多肽上可能是多一个、几个或少一个、几个氨基酸,而不完全打乱整个氨基酸序列。
核苷酸而导致阅读框架位移,导致翻译出来的蛋白质的氨基酸序列与野生型完全不同。 b. 如果插入或缺失的碱基正好是3个或其整倍数,那么在翻译出的多肽上可能是多一个、几个或少一个、几个氨基酸,而不完全打乱整个氨基酸序列。")
57
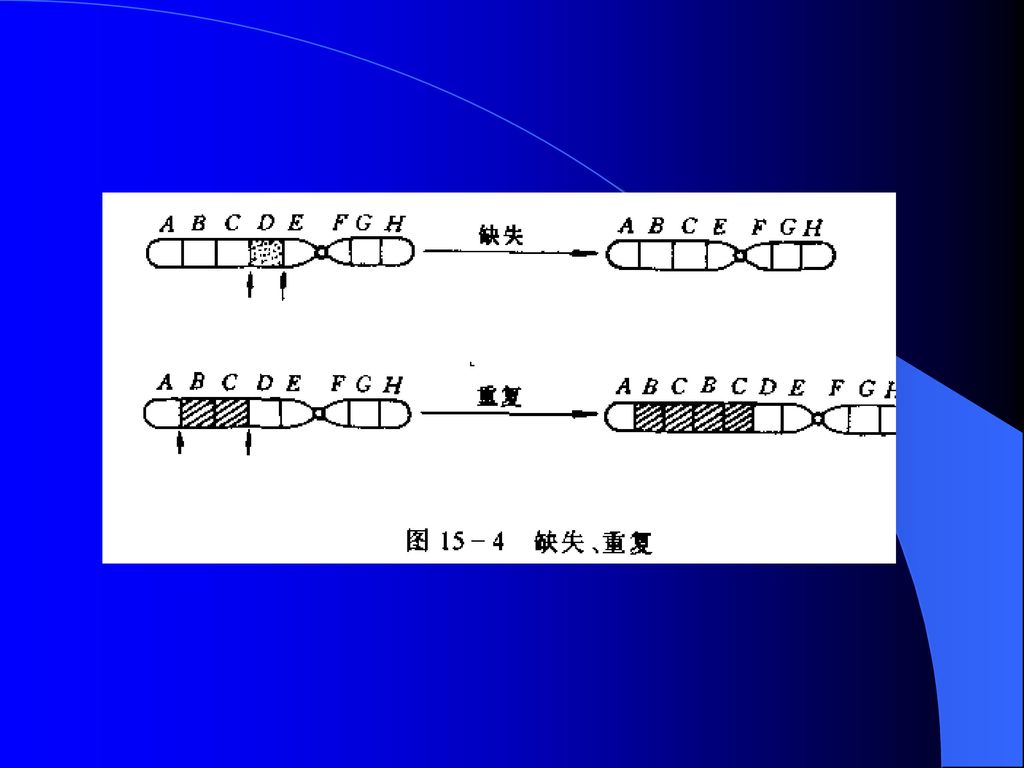
(2)染色体畸变 指染色体的结构和数目的改变。是DNA分子大范围的损伤。 1)染色体结构变异 缺失 指染色体丢失了一个片段,使之位于这个片段上的基因也随之丢失。 重复: 一个染色体上某一片段出现两份或两份以上,使位于这些片段上的基因多了一份或几份。 缺失和重复都起因于染色体上基因数目的变化。

59
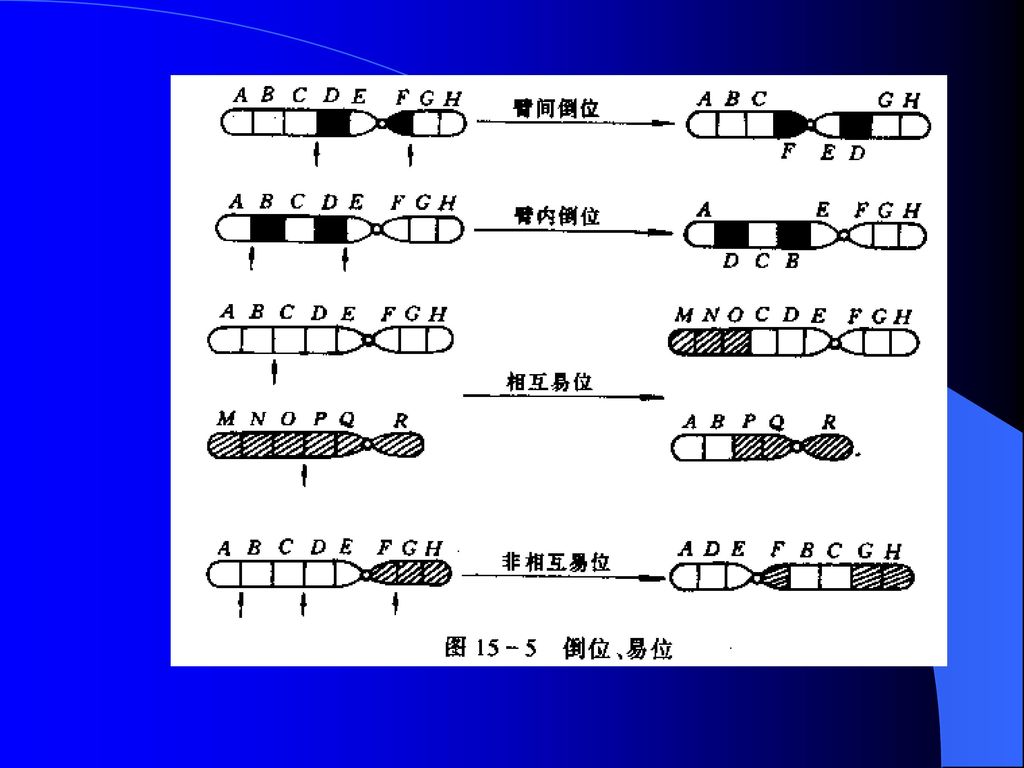
倒位 在同一染色体上某一个片段作180°的颠倒,造成染色体上基因顺序的重排。如果颠倒的片段包括着丝粒在内的倒位称为臂间倒位,不包括着丝粒的倒位称为臂内倒位。 易位 一个染色体臂的一段移接到另一非同源染色体的臂上的结构变异。 最常见的是相互易位,非同源染色体间相互交换染色体片段,造成染色体间基因的重排。 倒位和易位都起因于染色体上基因排列位置的变化。

61
转座因子和转座 转座因子:存在于细胞内,位于染色体或质粒上的一段特殊、可移动的DNA序列。 转座:DNA序列通过非同源重组的方式,从染色体某一部位转移到同一染色体上另一部位或其他染色体上某一部位的现象,称为转座。 转座因子的种类: 插入序列 IS: kb,只能引起转座效应,不含其它基因。 转座子 Tn :2-2.5 kb,含有几个至十几个基因。 转座噬菌体:37 kb,含有20多个基因。

62
2)染色体数目改变 a. 倍数性改变 b.非整倍性改变
染色体数目改变 a. 倍数性改变 b.非整倍性改变")
63
2、自发突变的机制 1)DNA复制中的错误——互变异构和环出效应 ①互变异构效应:由于碱基T、G发生了酮式至烯酵式或C、A发生了氨基式至亚氨基式的互变异构效应而引起碱基置换,造成突变。 ②环出效应:DNA上个别碱基对的局部解离和错误退火而导致环状突出,引起移码突变。

64
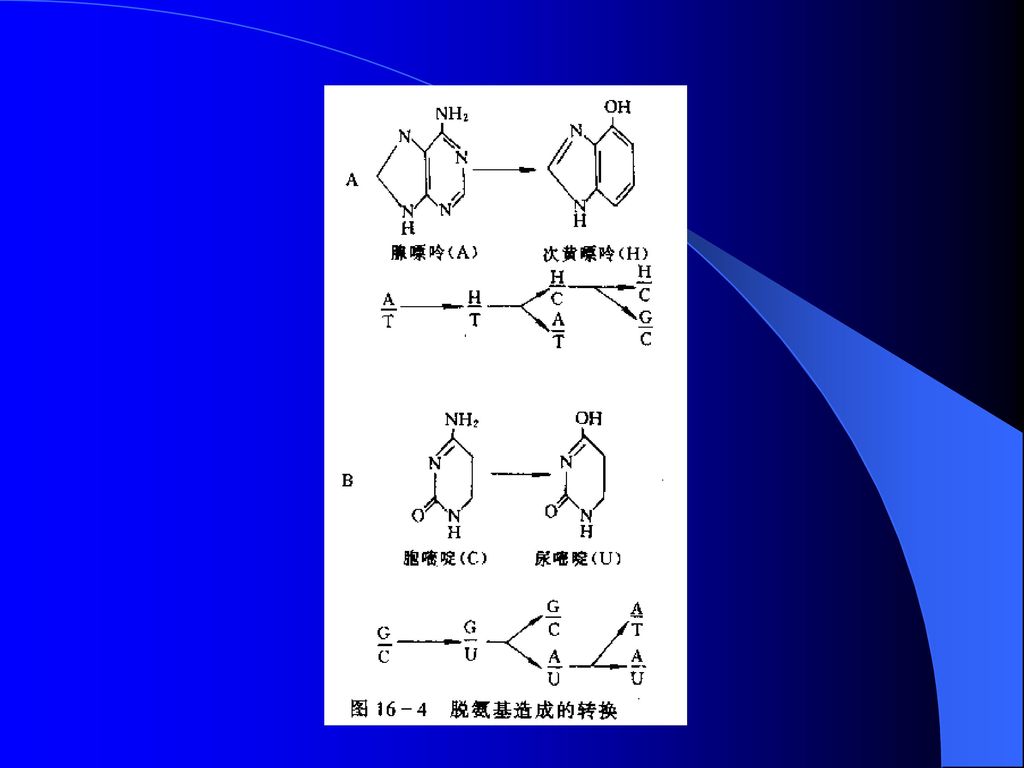
2)自发损伤 ① 脱嘌呤: 由于碱基和脱氧核糖间的糖苷键受到破坏,从而引起一个鸟嘌呤或腺嘌呤从DNA分子上脱落下来,在DNA复制过程中,在一个无嘌呤位点对面插入一个碱基,引起碱基错配,造成突变。 ② 脱氨基: 腺嘌呤脱氨基变成次黄嘌呤,胞嘧啶脱氨基变成尿嘧啶,引起转换。

66
③ 氧化性损伤: 指活泼氧化物对DNA前体及DNA本身造成的氧化性损伤,从而引起突变。 3)可转移因子的作用:由微生物自身所携带的转座遗传因子的转移和插入所造成的突变。
可转移因子的作用:由微生物自身所携带的转座遗传因子的转移和插入所造成的突变。")
67
(六)紫外线对DNA的损伤及机体对损伤DNA的修复
紫外线作用:同链DNA的相邻T间形成共价T二聚体,阻碍碱基间正常配对,从而引起突变或死亡。 (1) 光复活 (2) 切除修复 (3) 重组修复 (4) SOS修复
 光复活. (2) 切除修复. (3) 重组修复. (4) SOS修复.")
68
(1) 光复活: 光复活是专一地针对紫外线引起的DNA损伤而形成的嘧啶二聚体在损伤部位就地修复的修复途径,是在可见光(300一600 nm)的活化之下,由光复活酶(PR酶)又称光解酶,催化嘧啶二聚体分解成为单体的过程。
 光复活: 光复活是专一地针对紫外线引起的DNA损伤而形成的嘧啶二聚体在损伤部位就地修复的修复途径,是在可见光(300一600 nm)的活化之下,由光复活酶(PR酶)又称光解酶,催化嘧啶二聚体分解成为单体的过程。")
69
(2) 切除修复: 又称核苷酸外切修复,是紫外线等辐射物质所造成的损伤部位被取代的暗修复系统。此系统是在几种酶的协同作用下,先在损伤的任一端打开磷酸二酯键,然后外切掉一段寡核苷酸;留下的缺口由修复性合成来填补,再由连接酶将其连接起来。
 切除修复: 又称核苷酸外切修复,是紫外线等辐射物质所造成的损伤部位被取代的暗修复系统。此系统是在几种酶的协同作用下,先在损伤的任一端打开磷酸二酯键,然后外切掉一段寡核苷酸;留下的缺口由修复性合成来填补,再由连接酶将其连接起来。")
70
(3) 重组修复: 是一种越过损伤部位而进行修复的途径,它必须在DNA进行复制的情况下进行,故又称复制后修复。发生在DNA复制过程或复制之后,不切除DNA损伤部位的修复。DNA链在复制时,受损的模板作用消失,互补单链(新链)里留下空隙,产生诱导信号,recA基因被诱导,产生大量重组蛋白,与新链缺口结合,引起 子链和母链交换。交换后母链缺口通过聚合作用,以对侧子链为模板合成DNA 片段填充,连接酶连接新旧链完成复制。
里留下空隙,产生诱导信号,recA基因被诱导,产生大量重组蛋白,与新链缺口结合,引起 子链和母链交换。交换后母链缺口通过聚合作用,以对侧子链为模板合成DNA 片段填充,连接酶连接新旧链完成复制。")
71
(4) SOS修复 是在DNA受损伤的范围较大而且复制受到抑制时出现的一种修复作用。 一种能够引起误差修复的紧急呼救修复,是在无模板DNA 情况下合成酶的诱导修复。正常情况下有关酶系无活性,DNA受损伤而复制又受到抑制情况下发出信号,激活有关酶系,对DNA损伤进行修复,其中DNA多聚酶起重要作用,在无模板情况下,进行DNA修复再合成,并将DNA片段插入受损DNA空隙处。

72
二、突变与育种 菌种工作包括三方面:选种、育种、复壮和保藏。 选种—从自然界和生产中选择符合需要菌种。 育种—进一步提高已有菌种某种性能,使更符合要求。 选育新菌种可从几方面着手: 1)从原有菌株入手进行各种遗传改造工作。 2)根据文献资料报导的微生物种属,向国内外菌种保藏机构索取同种或同属菌株,从中寻求符合要求者。 3)根据所需菌种特性、嗜好或工艺要求,从特定的生态环境中以特定方法重新分离自然菌株。
根据文献资料报导的微生物种属,向国内外菌种保藏机构索取同种或同属菌株,从中寻求符合要求者。 3)根据所需菌种特性、嗜好或工艺要求,从特定的生态环境中以特定方法重新分离自然菌株。")
73
(一)自发突变与育种 1、生产育种 在利用微生物进行大生产的过程中由于微生物会以10-6左右的突变率进行自发突变,从而分离出生产性状优于原菌株的优良菌株。 2、定向育种 用某一特定环境长期处理某一微生物群体,同时不断地进行移种传代,以达到积累和选择合适的自发突变体的一种古老育种方法。 (二)诱变育种 利用物理或化学诱变剂处理均匀分散的微生物细胞群,促进其突变率大幅度提高,然后设法采用简便、快速、高效的筛选方法,从中挑选少数符合目的的突变株,以供生产科研之用。
诱变育种. 利用物理或化学诱变剂处理均匀分散的微生物细胞群,促进其突变率大幅度提高,然后设法采用简便、快速、高效的筛选方法,从中挑选少数符合目的的突变株,以供生产科研之用。")
74
诱变育种基本步骤 基本步骤: 原种 (出发菌株) → 纯化→斜面→同步培养→离心洗涤→ 振荡打散→过滤→菌悬液→诱变处理 → 平板分离 → 斜面 → (活菌计数) (计数) (变异率) 斜面 → 斜面 →保藏及扩大试验 (初筛) (复筛) (再复筛)
 (复筛) (再复筛)")
75
1、出发菌株的选择 (1) 对出发菌株的要求 生长快,营养要求粗放,发育早,产孢子多,对诱变剂敏感性高,已能积累少量产品或前体物的菌株。 (2) 出发菌株的选择 1) 选取自然界新分离的野生型菌株,它们对诱变因素敏感,容易发生变异; 2) 选取生产中由于自发突变或长期在生产条件下驯化而筛选得到的菌株,与野生型菌株较相象,容易达到较好的诱变效果; 3) 选取每次诱变处理都有一定提高的菌株,往往多次诱变可能效果迭加,积累更多的提高。
 选取生产中由于自发突变或长期在生产条件下驯化而筛选得到的菌株,与野生型菌株较相象,容易达到较好的诱变效果; 3) 选取每次诱变处理都有一定提高的菌株,往往多次诱变可能效果迭加,积累更多的提高。")
76
2、出发菌株的纯化 (1) 目的:获得遗传性状基本一致的,并且稳定的变种。 (2) 原因:遗传背景复杂的菌种诱变后负变率将增加;诱变史长的菌株,采用强烈诱变剂处理,又不进行纯化分离,诱变效果差。 (3) 纯种分离方法:常用划线分离法和稀释分离法。
 目的:获得遗传性状基本一致的,并且稳定的变种。 (2) 原因:遗传背景复杂的菌种诱变后负变率将增加;诱变史长的菌株,采用强烈诱变剂处理,又不进行纯化分离,诱变效果差。 (3) 纯种分离方法:常用划线分离法和稀释分离法。")
77
3、同步培养(前培养) (1)目的:获得生理状态一致的培养物。 (2) 原因:突变率高,重现性也好。 (3) 方法: 1) 细菌一般要求培养至对数生长期; 2) 霉菌处理使用分生孢子,应该将分生孢子在液体培养基中短时间培养,使孢子孵化,处于活化状态,并恰好未形成菌丝体。
 (1)目的:获得生理状态一致的培养物。 (2) 原因:突变率高,重现性也好。 (3) 方法: 1) 细菌一般要求培养至对数生长期; 2) 霉菌处理使用分生孢子,应该将分生孢子在液体培养基中短时间培养,使孢子孵化,处于活化状态,并恰好未形成菌丝体。")
78
4、单细胞(或单孢子)悬液的制备 (1) 目的 获得单细胞(或单孢子) 、均匀的悬液。 (2) 原因 一方面分散状态的细胞可以均匀地接触诱变剂,还可减少分离现象发生。另一方面又可避免长出不纯菌落。 (3) 方法 1) 菌龄:对数期细胞、刚成熟的孢子或活化的孢子 。 2) 菌悬液浓度:一般真菌孢子或酵母菌细胞悬浮液的浓度为106个/mL、放线菌或细菌的浓度为108个/mL左右。 3) 菌悬液的配制方法: 离心洗涤前培养物,用冷生理盐水或缓冲液制备菌悬液,放在盛有玻璃珠的三角瓶内振荡10min,令其分散,用无菌脱脂棉或滤纸过滤。通过菌体计数,调整菌悬液的浓度供诱变处理。
 方法. 1) 菌龄:对数期细胞、刚成熟的孢子或活化的孢子 。 2) 菌悬液浓度:一般真菌孢子或酵母菌细胞悬浮液的浓度为106个/mL、放线菌或细菌的浓度为108个/mL左右。 3) 菌悬液的配制方法: 离心洗涤前培养物,用冷生理盐水或缓冲液制备菌悬液,放在盛有玻璃珠的三角瓶内振荡10min,令其分散,用无菌脱脂棉或滤纸过滤。通过菌体计数,调整菌悬液的浓度供诱变处理。")
79
5、诱变处理 (1) 诱变剂种类的选择 常用诱变剂有两大类:物理诱变剂和化学诱变剂。 常用的物理诱变剂: 紫外线、x射线、γ射线(如Co60等)、等离子、快中子、α射线、β射线、超声波等。 常用的化学诱变剂: 碱基类似物、烷化剂、羟胺、吖定类化合物等。

80
(2) 最适诱变剂量的选择 1) 诱变剂剂量的表示方式 a.UV的剂量指强度与作用时间的乘积。 b.化学诱变剂常以在一定外界条件下诱变剂的浓度和作用时间的乘积来表示。 c.在育种实践中,常以杀菌率来作诱变剂的相对剂量。

81
2) 最适诱变剂量的确定 a.确定依据 诱变的最适剂量,应该使所希望得到的突变株在存活群体中占有最大的比例。 b. 确定方法 通过比较剂量—存活率曲线和剂量—诱变率曲线,找到某诱变剂的剂量—存活率—诱变率三者的最佳结合点,即为诱变剂的最适剂量。 ① 剂量—存活率曲线 以诱变剂的剂量为横坐标,以细胞存活率的对数值为纵坐标绘制的曲线。 ② 剂量—诱变率曲线 以诱变剂的剂量为横坐标,以诱变后获得的突变细胞数为纵坐标绘制的曲线。

82
c.紫外诱变的方法 ①紫外灯应先预热20~30min ②将10m1菌悬液放在直径为9cm的培养皿中,液层厚度约为2mm,启动磁力搅拌器,使用15w功率紫外灯管,照射距离为30cm左右,照射时间以几秒至数十分钟为宜,具芽孢的菌株需处理10min左右。 ③设计一个照射不同时间梯度的实验,根据不同时间照射的死亡率,作出照射时间与死亡率的曲线,这样就可以选择适当的照射剂量。 实验中避免光复活现象: 实验时,为了避免光复活现象,处理过程应在暗室的红光下操作,处理完毕后,将盛菌悬液的器皿用黑布包起来培养,然后再进行分离筛选。

83
(3)诱变剂的处理方式 1) 单因子处理 采用单一诱变剂处理出发菌株。 2) 复合因子处理 指两种以上诱变因子共同诱发菌体突变。 ① 两种或多种诱变剂先后使用。 ② 同一种诱变剂的重复使用。 ③ 两种或多种诱变剂的同时使用。

84
(4)诱变剂的处理方法 1) 直接处理方法 将菌悬液用物理、化学因子处理,然后涂平板分离突变株。 2) 生长过程处理 适用于诱变作用强的而杀菌率较低的诱变剂,或在分裂过程中只对DNA起作用的诱变剂,如NTG、LiCl、秋水仙碱等。 ① 将诱变剂加入培养基涂平板。 ② 先把培养基制成平板,将一定浓度的诱变剂和菌体加入平板。 ③ 摇瓶振荡培养处理

85
6、后培养 后培养是指诱变后的菌悬液不直接分离于平板,而是立即转移到营养丰富的培养基中培养数代。 (1)方法 将诱变处理后的菌体转移到适宜的培养条件下,培养几小时,让突变细胞繁殖几代。 (2) 目的 1) 诱变处理后发生的突变,通过修复、繁殖,即DNA的复制,才能形成一个稳定突变体。 2) 根据突变体表型延迟现象,通过后培养,使表型都得到充分表达。 (3) 后培养的培养基:一般培养基中加入足量的酪素水解物、酵母膏等富含特种氨基酸、生长因子和ATP的营养物质。
 诱变处理后发生的突变,通过修复、繁殖,即DNA的复制,才能形成一个稳定突变体。 2) 根据突变体表型延迟现象,通过后培养,使表型都得到充分表达。 (3) 后培养的培养基:一般培养基中加入足量的酪素水解物、酵母膏等富含特种氨基酸、生长因子和ATP的营养物质。")
86
7、突变株的分离与筛选 (1) 筛选方案:
 筛选方案:")
87
(2)筛选方法 1) 随机筛选 也称摇瓶筛选。该法主要是随机挑选的平板菌落进行摇瓶筛选。 2) 平板菌落预筛 是在培养皿或特制玻璃框平板上进行的,是用于诱变后从试样小检出突变体的一种琼脂平板筛选法。 常用的方法有纸片培养显色法、透明圈法、琼脂块培养法等。

88
(3)产物活性测定 1)琼脂平板空洞法 该法是在特制的玻璃框琼脂平板上进行的,以检验菌或底物与一定量的琼脂制成平板,用专制的打孔器取出琼脂块,在留下的圆孔中加入发酵液,或用圆滤纸片浸透发酵液直接覆于琼脂平板上,在适合温度下培养一定时间,测量圆孔或滤纸片周围形成的抑菌圈或水解圈,根据活性团的大小挑选高产菌株。 2) 纸片法
 纸片法.")
89
(4) 摇瓶数据的调整和有关菌株特性的观察分析
对摇瓶发酵液的测试数据要尽量应用生物学统计方法来处理 。 在分离、筛选的整个试验过程中,每个阶段都要周密地观察菌株特性,诱变后分离在平板上的菌落生长情况,如生长快慢、菌落形态、大小、颜色等,要一一详细记录。 菌落移入试管斜面后的生长情况、接入到摇瓶种子和发酵培养基后,其培养过程中的糖、氮利用、生长速度、pH变化、颜色、强度等,都要及时观察,结合镜检、测定产物活性,进行详细记录。 综合菌落的形态特征、培养特征及生化特征作为初筛时挑选菌落的依据。 对筛选过程中摇瓶产量数据的分布作全面分析,以便帮助判断诱变剂、诱变剂量及筛选条件的选择是否恰当。

90
(5) 培养基和培养条件的调整 利用单因子法、正交实验法、均匀设计、响应面法等方法进行培养基和培养条件的优化,以充分发挥高产菌株的高产性能。
 培养基和培养条件的调整 利用单因子法、正交实验法、均匀设计、响应面法等方法进行培养基和培养条件的优化,以充分发挥高产菌株的高产性能。")
91
(三)三类突变株的筛选方法 1、产量突变株的筛选——琼脂块培养法
三类突变株的筛选方法 1、产量突变株的筛选——琼脂块培养法")
92
2、抗性突变株的筛选——梯度平板法 梯度平板法是定向筛选抗药性突变株的一种有效方法。通过制备琼脂表面存在药物浓梯度的平板、在其上涂布诱变处理后的细胞悬液、经培养后再从其上选取抗药性菌落等步骤,就可定向筛选到相应抗药性突变株。在筛选抗代谢药物的抗性菌株以取得相应代谢物的高产菌株方面,此法能达到定向培育的效果。 实例:定向培育抗异烟肼的吡多醇高产突变株

93
3、营养缺陷型突变株的筛选 营养缺陷型突变株在科学研究和生产实践上具有极其重要的意义: 在科学实验中: ①它们既可作为研究代谢途径和杂交(包括半知菌的准性杂交、细菌的接合和各种细胞的融合等)、转化、转导、转座等遗传规律所不可缺少的遗传标记菌种; ②可作为氨基酸、维生素或碱基等物质生物测定中的试验菌种; 在生产实践中: ①它们既可直接用作发酵生产氨基酸、核苷酸等有益代谢产物的菌种; ②也可作为对生产菌种进行杂交育种、重组育种和利用基因工程育种时所不可缺少的亲本遗传标志和杂交种的选择性标志。

94
(1) 与筛选营养缺陷型突变株有关的三类培养基
1)基本培养基(MM[—]) 仅能满足某微生物的野生型菌株生长所需要的最低成分的组合培养基,称基本培养基。 2)完全培养基(CM[+]) 凡可满足一切营养缺陷型菌株营养需要的天然或半组合培养基,称完全培养基。 3)补充培养基(SM[A]或[B]) 凡只能满足相应的营养缺陷型突变株生长需要的组合或半组合培养基,称补充培养基。
基本培养基(MM[—]) 仅能满足某微生物的野生型菌株生长所需要的最低成分的组合培养基,称基本培养基。 2)完全培养基(CM[+]) 凡可满足一切营养缺陷型菌株营养需要的天然或半组合培养基,称完全培养基。 3)补充培养基(SM[A]或[B]) 凡只能满足相应的营养缺陷型突变株生长需要的组合或半组合培养基,称补充培养基。")
95
(2) 与营养缺陷型突变有关的三类遗传型个体
1)野生型[A+B+] 指从自然界分离到的任何微生物在其发生人为营养缺陷突变前的原始菌株。 2)营养缺陷型[A-B+]、[A+B-] 野生型菌株经诱变剂处理后,由于发生了丧失某酶合成能力的突变,因而只能在加有该酶合成产物的培养基中才能生长,这类突变菌株称为营养缺陷型突变株,或简称营养缺陷型。 3)原养型[A+B+] 一般指营养缺陷型突变株经回复突变或重组后产生的菌株,其营养要求在表型上与野生型相同,遗传型均用[A+B+]表示。
野生型[A+B+] 指从自然界分离到的任何微生物在其发生人为营养缺陷突变前的原始菌株。 2)营养缺陷型[A-B+]、[A+B-] 野生型菌株经诱变剂处理后,由于发生了丧失某酶合成能力的突变,因而只能在加有该酶合成产物的培养基中才能生长,这类突变菌株称为营养缺陷型突变株,或简称营养缺陷型。 3)原养型[A+B+] 一般指营养缺陷型突变株经回复突变或重组后产生的菌株,其营养要求在表型上与野生型相同,遗传型均用[A+B+]表示。")
97
(3) 营养缺陷型的筛选方法——四个环节:诱变、淘汰野生型、检出和鉴定营养缺陷型。
1)诱变剂处理 2)浓缩缺陷型 ① 抗生素法 青霉素法:主要适用于细菌 原理:青霉素能抑制细菌细胞壁的生物合成,因而能杀死生长繁殖着的细菌,但不能杀死处于休止状态的细菌。在只能使野生型正常生长而不能使营养缺陷型生长的基本培养基中,野生型则被杀死而缺陷型则不被杀死,于是缺陷型就得到了浓缩。
诱变剂处理. 2)浓缩缺陷型. ① 抗生素法. 青霉素法:主要适用于细菌. 原理:青霉素能抑制细菌细胞壁的生物合成,因而能杀死生长繁殖着的细菌,但不能杀死处于休止状态的细菌。在只能使野生型正常生长而不能使营养缺陷型生长的基本培养基中,野生型则被杀死而缺陷型则不被杀死,于是缺陷型就得到了浓缩。")
98
方法步骤: i. 细菌诱变; ii. 将处理后的细菌放在完全培养基中培养以使突变型充分表现; iii. 将细菌转入不含氮源物质的基本培养基中培养,以使营养缺陷型菌株内所吸收的完全培养基成分充分消耗,以便停止生长繁殖; iv. 转入含有无机氮源和青霉素的基本培养基中培养,野生型正常生长而营养缺陷型不能生长,从而使缺陷型得到浓缩。

99
制霉菌素法:适用于真菌。 原理:制霉菌素属于大环内酯类抗生素,可与真菌细胞膜上的甾醇作用,从而引起膜的损伤。因为它只能杀死生长繁殖着的酵母菌或霉菌,故也可用于淘汰相应的野生型菌株和“浓缩”营养缺陷型菌株。

100
② 菌丝过滤法 菌丝过滤法适用于丝状生长的真菌或放线菌。 原理:在基本培养基中、野生型的孢子能发芽成菌丝,而营养缺陷型的孢子则不能。因此,将诱变剂处理后的孢子在基本培养基中培养一段时间后,再用擦镜纸等合适滤纸过滤。如此重复数次后,就可去除大部分野生型个体,同样也就达到了“浓缩”营养缺陷型的目的。 差别杀菌法: 芽孢远比营养体耐热,利用温度的差异可以浓缩缺陷型。此方法仅适用于形成芽孢的细菌。 饥饿法: 当微生物发生某一营养型缺陷时,在不补充该缺陷物质的培养条件下会自行死亡。可是如果该细胞在发生一种缺陷型突变的同时又发生了另一种类型的突变,这一细胞反而有可能因暂时避免死亡而被检出。

101
3)检出缺陷型 ①夹层培养法
检出缺陷型 ①夹层培养法")
102
②限量补充培养法 把诱变处理后的细胞接种在含有微量(0.01%以下)蛋白胨的基本培养基上,野生型细胞就迅速长成较大的菌落,而营养缺陷型则生长缓慢,并只能形成微小的菌落。 ③逐个检出法 细胞诱变处理 平板涂布在完全培养基上 接种针或灭菌牙签转接 培养观察

103
④影印平板法

104
4)鉴定缺陷型 可借生长谱法进行。 生长谱法是指在混有供试菌的平板表面点加微量营养物,视某营养物的周围有否长菌来确定该供试菌的营养要求的一种快速、直观的方法。
鉴定缺陷型 可借生长谱法进行。 生长谱法是指在混有供试菌的平板表面点加微量营养物,视某营养物的周围有否长菌来确定该供试菌的营养要求的一种快速、直观的方法。")
105
(四)Ames实验 利用细菌营养缺陷型的回复突变检出环境或食品中是否存在化学致癌剂的一种简便有效方法。 1、诱变的生物化学统一性 任何能改变核酸结构的因素,都可引起核酸生物学功能的改变;凡能引起核酸其他功能改变的因素,一般也能引起突变。

106
2、筛选简单模型研究高等生物的突变(癌症)
(1) 化学物质对核酸结构的改变和对生物的三致作用 (2) 致突变因素,一般都有致突致癌致畸效应(三致作用),反之亦然 (3) 对原核生物有效的因素,则对非细胞生物和真核生物也有效 (4) 能引起易鉴出的选择性突变的因素,也能引起难以检出的非选择性突变 (5) 凡能引起负变的因素,也可引起正变 (6) 凡能引起回复突变的因素,一般也能引起正向突变
 化学物质对核酸结构的改变和对生物的三致作用. (2) 致突变因素,一般都有致突致癌致畸效应(三致作用),反之亦然. (3) 对原核生物有效的因素,则对非细胞生物和真核生物也有效. (4) 能引起易鉴出的选择性突变的因素,也能引起难以检出的非选择性突变. (5) 凡能引起负变的因素,也可引起正变. (6) 凡能引起回复突变的因素,一般也能引起正向突变.")
107
3、Ames实验原理: 根据诱变剂的共性原则,即:化学试剂对细菌的诱变率与其对动物的致癌性成正比,利用细菌营养缺陷型的回复突变来检测食品或环境中是否存在致癌剂。鼠伤寒沙门氏菌的组氨酸缺陷型(his-)菌株在基本培养基[-]的平板上不能生长,如发生回复突变则能生长。
![3、Ames实验原理: 根据诱变剂的共性原则,即:化学试剂对细菌的诱变率与其对动物的致癌性成正比,利用细菌营养缺陷型的回复突变来检测食品或环境中是否存在致癌剂。鼠伤寒沙门氏菌的组氨酸缺陷型(his-)菌株在基本培养基[-]的平板上不能生长,如发生回复突变则能生长。](http://slidesplayer.com/slide/11398682/61/images/107/3%E3%80%81Ames%E5%AE%9E%E9%AA%8C%E5%8E%9F%E7%90%86%EF%BC%9A+%E6%A0%B9%E6%8D%AE%E8%AF%B1%E5%8F%98%E5%89%82%E7%9A%84%E5%85%B1%E6%80%A7%E5%8E%9F%E5%88%99%EF%BC%8C%E5%8D%B3%EF%BC%9A%E5%8C%96%E5%AD%A6%E8%AF%95%E5%89%82%E5%AF%B9%E7%BB%86%E8%8F%8C%E7%9A%84%E8%AF%B1%E5%8F%98%E7%8E%87%E4%B8%8E%E5%85%B6%E5%AF%B9%E5%8A%A8%E7%89%A9%E7%9A%84%E8%87%B4%E7%99%8C%E6%80%A7%E6%88%90%E6%AD%A3%E6%AF%94%EF%BC%8C%E5%88%A9%E7%94%A8%E7%BB%86%E8%8F%8C%E8%90%A5%E5%85%BB%E7%BC%BA%E9%99%B7%E5%9E%8B%E7%9A%84%E5%9B%9E%E5%A4%8D%E7%AA%81%E5%8F%98%E6%9D%A5%E6%A3%80%E6%B5%8B%E9%A3%9F%E5%93%81%E6%88%96%E7%8E%AF%E5%A2%83%E4%B8%AD%E6%98%AF%E5%90%A6%E5%AD%98%E5%9C%A8%E8%87%B4%E7%99%8C%E5%89%82%E3%80%82%E9%BC%A0%E4%BC%A4%E5%AF%92%E6%B2%99%E9%97%A8%E6%B0%8F%E8%8F%8C%E7%9A%84%E7%BB%84%E6%B0%A8%E9%85%B8%E7%BC%BA%E9%99%B7%E5%9E%8B%28his-%29%E8%8F%8C%E6%A0%AA%E5%9C%A8%E5%9F%BA%E6%9C%AC%E5%9F%B9%E5%85%BB%E5%9F%BA%5B-%5D%E7%9A%84%E5%B9%B3%E6%9D%BF%E4%B8%8A%E4%B8%8D%E8%83%BD%E7%94%9F%E9%95%BF%EF%BC%8C%E5%A6%82%E5%8F%91%E7%94%9F%E5%9B%9E%E5%A4%8D%E7%AA%81%E5%8F%98%E5%88%99%E8%83%BD%E7%94%9F%E9%95%BF%E3%80%82.jpg "3、Ames实验原理: 根据诱变剂的共性原则,即:化学试剂对细菌的诱变率与其对动物的致癌性成正比,利用细菌营养缺陷型的回复突变来检测食品或环境中是否存在致癌剂。鼠伤寒沙门氏菌的组氨酸缺陷型(his-)菌株在基本培养基[-]的平板上不能生长,如发生回复突变则能生长。")
108
4、方法

109
第三节 基因重组和杂交育种 基因重组:凡把两个不同性状个体内的遗传基因转移到起,经过遗传分子间的重新组合,形成新遗传型个体的方式,称为基因重组或遗传重组。 杂交:两个性状不同的菌株或变种之间进行细胞结合,遗传物质交换重新组合成新的性状。

110
一、原核生物的基因重组 特点:(1)片段性 (2)单向性 (3)转移机制独特而多样
片段性 (2)单向性 (3)转移机制独特而多样")
111
(一)转化 1、转化的发现 F. Grifith所做肺炎球菌对小鼠的感染实验以及10年后Averry等在体外实现转化。 2、定义 受体菌直接吸收了来自供体菌的DNA片段,通过交换,把它组合到自己的基因组中,从而获得供体菌部分遗传性状的现象。转化后的受体菌,称转化子,供体菌的DNA片段称为转化因子。

112
感受态即受体菌最易接收外源DNA片段并实现转化的生理状态。
3、感受态 (1)定义 感受态即受体菌最易接收外源DNA片段并实现转化的生理状态。 (2)特点 1)表示细胞具有摄取外源DNA的能力; 2)自然转化的感受态细胞受感受态因子调节; 是调节感受态的一类特异蛋白,它可以催化外来DNA片段的吸收或降解细胞表面某种成分,让细胞表面的DNA受体显露出来。它包括三种主要成分:与膜相关的DNA结合蛋白、细胞壁自溶素和几种核酸酶。 3)是细胞群体在某个生长阶段的一种特性,大多与生长周期有关 。 4)是反映细胞个体之间遗传信息交流的一种程度。
定义. 感受态即受体菌最易接收外源DNA片段并实现转化的生理状态。 (2)特点. 1)表示细胞具有摄取外源DNA的能力; 2)自然转化的感受态细胞受感受态因子调节; 是调节感受态的一类特异蛋白,它可以催化外来DNA片段的吸收或降解细胞表面某种成分,让细胞表面的DNA受体显露出来。它包括三种主要成分:与膜相关的DNA结合蛋白、细胞壁自溶素和几种核酸酶。 3)是细胞群体在某个生长阶段的一种特性,大多与生长周期有关 。 4)是反映细胞个体之间遗传信息交流的一种程度。")
113
4、转化机制 (1)转化发生的条件 1) 受体细胞处于感受态。 2) 受体细胞吸附的转化因子,必须是双链的DNA,且DNA分子的相对分子质量不小于3×105 。 转化不需两个细胞直接接触
转化发生的条件 1) 受体细胞处于感受态。 2) 受体细胞吸附的转化因子,必须是双链的DNA,且DNA分子的相对分子质量不小于3×105 。 转化不需两个细胞直接接触")
114
(2)转化的过程 1)感受态的出现 2)DNA的结合与进入 ①G+
转化的过程 1)感受态的出现 2)DNA的结合与进入 ①G+")
115
②G- 3) DNA的整合(重组) 进入细胞的外源DNA通过同源重组以置换的方式整合进受体染色体DNA,经复制和细胞分裂后形成重组体。
 DNA的整合(重组) 进入细胞的外源DNA通过同源重组以置换的方式整合进受体染色体DNA,经复制和细胞分裂后形成重组体。")
117
(3)影响转化效率的因素: 1) 受体细胞的感受态——决定转化因子能否被吸收进入受体细胞; 2) 受体细胞的限制酶系统和其他核酸酶——决定转化因子在整合前是否被分解; 3) 受体和供体染色体的同源性——决定转化因子的整合。
影响转化效率的因素: 1) 受体细胞的感受态——决定转化因子能否被吸收进入受体细胞; 2) 受体细胞的限制酶系统和其他核酸酶——决定转化因子在整合前是否被分解; 3) 受体和供体染色体的同源性——决定转化因子的整合。")
118
6、转染 (1)定义:把噬菌体或其他病毒DNA(RNA)提取出来,用它去感染感受态的宿主细胞,并产生正常噬菌体或病毒后代。 (2)转染和转化:作为转染的病毒核酸,并不是作为供体基因的功能,被感染的宿主也决不是能形成转化子的受体菌。 (3)转染和病毒感染: 1)转染中进入宿主细胞的是DNA,而在病毒感染中则是完整的病毒颗粒; 2)转染是转化途径,而病毒感染是复制途径。
转染和病毒感染: 1)转染中进入宿主细胞的是DNA,而在病毒感染中则是完整的病毒颗粒; 2)转染是转化途径,而病毒感染是复制途径。")
119
B菌株:bio+met+thr-leu-(需要苏氨酸和异亮氨酸)
(二)接合 1、细菌基因重组的发现和证实 A菌株:bio-met-thr+leu+(需要生物素和硫氨酸) B菌株:bio+met+thr-leu-(需要苏氨酸和异亮氨酸)
接合. 1、细菌基因重组的发现和证实. A菌株:bio-met-thr+leu+(需要生物素和硫氨酸) B菌株:bio+met+thr-leu-(需要苏氨酸和异亮氨酸)")
120
(1)转化作用的排除——Davis的U形管试验
结果:基本培养基平板上未长出原养型菌落。 结论:混合培养得到的重组子不是转化的结果;重组子的出现需要两亲本细胞的直接接触。

121
用双重营养缺陷型菌株A-B-C+D+和A+B+C-D-混合培养,结果仍出现原养型菌落。
(2)互养的排除 将基因型A-B+Tls和A+B-Tlr两种细菌在基本培养基上混合培养,接触较短的一段时间以后,喷上噬菌体Tl,把A-B+Tls细菌杀死。经培养以后仍有原养型菌落出现,这说明原养型菌落的出现并非由于互养。 (3)回复突变的排除 用双重营养缺陷型菌株A-B-C+D+和A+B+C-D-混合培养,结果仍出现原养型菌落。 两个基因同时回复突变,则其可能性只有<10-16(10-8×10-8),这种几率在平板上是很难检测到的,所以混合培养能出现10-5——10-6频率的菌落一定是重组的结果。
互养的排除. 将基因型A-B+Tls和A+B-Tlr两种细菌在基本培养基上混合培养,接触较短的一段时间以后,喷上噬菌体Tl,把A-B+Tls细菌杀死。经培养以后仍有原养型菌落出现,这说明原养型菌落的出现并非由于互养。 (3)回复突变的排除. 用双重营养缺陷型菌株A-B-C+D+和A+B+C-D-混合培养,结果仍出现原养型菌落。 两个基因同时回复突变,则其可能性只有<10-16(10-8×10-8),这种几率在平板上是很难检测到的,所以混合培养能出现10-5——10-6频率的菌落一定是重组的结果。")
122
(4)基因重组的证实 1)原养型细菌的三种可能性:异核体、杂合二倍体、单倍重组体。 异核体和杂合二倍体在继续培养过程中或多或少会出现分离现象,也就是说在原养型的后代中或多或少会出现缺陷型细菌。可是,营养缺陷型细菌的混合培养中出现的原养型菌落的基因是稳定的,所以这种原养型可能就是单倍重组体。
基因重组的证实 1)原养型细菌的三种可能性:异核体、杂合二倍体、单倍重组体。 异核体和杂合二倍体在继续培养过程中或多或少会出现分离现象,也就是说在原养型的后代中或多或少会出现缺陷型细菌。可是,营养缺陷型细菌的混合培养中出现的原养型菌落的基因是稳定的,所以这种原养型可能就是单倍重组体。")
123
2)杂交子代类型的分析 A-B-C+D+1ac-smrT1s×A+B+C-D-lac+smsTlr杂交
杂交子代类型的分析 A-B-C+D+1ac-smrT1s×A+B+C-D-lac+smsTlr杂交")
124
2、接合的定义:供体菌(“雄性”)通过性菌毛与受体菌(“雌性”)直接接触,把F质粒或其携带的不同长度的核基因组片段传递给后者,使后者获得获得若干新遗传性状的现象,称为接合。通过接合而获得新遗传性状的受体细胞称为接合子。 3、F质粒 (1)F质粒的特性 1)是一种附加体 2)可传递 3)可消除 4)是一种遗传性状 5)是合成性菌毛基因的载体,也是决定细菌性别的物质基础。
F质粒的特性. 1)是一种附加体. 2)可传递. 3)可消除. 4)是一种遗传性状. 5)是合成性菌毛基因的载体,也是决定细菌性别的物质基础。")
125
(2)E.coliF质粒的4种存在方式及相互联系
1)根据大肠杆菌细胞内有无F质粒及F质粒在细胞内的存在状态可将细胞分为4种类型:F-、F+、Hfr及F’。 ①F+菌株: 即“雄性”菌株,指细胞内含有一至几个F质粒,而且F质粒以自主复制形式存在。F+菌株细胞表面着生一至几条性菌毛。 ②F-菌株:即“雌性”菌株,指细胞内不含F质粒,细胞表面也无性菌毛的菌株。 ③Hfr菌株:即高频重组菌株,是指F质粒通过同源重组整合到宿主的染色体上,随着宿主染色体的复制而复制。 ④F’菌株:携带了宿主的一部分染色体的F质粒称为F’质粒。 凡携带F’质粒的菌株称为初生F’菌株;通过F’菌株与F-菌株的接合可使后者也成为F’菌株,即为次生F’菌株。
根据大肠杆菌细胞内有无F质粒及F质粒在细胞内的存在状态可将细胞分为4种类型:F-、F+、Hfr及F’。 ①F+菌株: 即 雄性 菌株,指细胞内含有一至几个F质粒,而且F质粒以自主复制形式存在。F+菌株细胞表面着生一至几条性菌毛。 ②F-菌株:即 雌性 菌株,指细胞内不含F质粒,细胞表面也无性菌毛的菌株。 ③Hfr菌株:即高频重组菌株,是指F质粒通过同源重组整合到宿主的染色体上,随着宿主染色体的复制而复制。 ④F’菌株:携带了宿主的一部分染色体的F质粒称为F’质粒。 凡携带F’质粒的菌株称为初生F’菌株;通过F’菌株与F-菌株的接合可使后者也成为F’菌株,即为次生F’菌株。")
126
2)E.coli四种接合型菌株的联系
E.coli四种接合型菌株的联系")
127
① F+x F-杂交→2F+

128
② Hfr×F- a.过程: 结果仍是Hfr细胞和F-细胞

129
b.意义——接合中断法 接合中断法(即中断杂交):就是将两个菌株在培养液中进行通风培养,每隔一定时间取样,把菌液放入组织捣碎器里搅拌以中断杂交,经过稀释接种到鉴别培养基上,待形成菌落后鉴定它们的基因型。 原理: Hfr×F-杂交中的DNA转移过程存在着严格的顺序性,所以,在实验室中可以每隔一定时间利用强烈搅拌等措施,使接合细胞对中断其接合,以获得呈现不同数量Hfr性状的F-接合子。根据这一原理,就可选用几种有特定整合位点的Hfr菌株,使其与F-菌株进行接合,并在不同时间使接合中断,最后根据F-中出现Hfr菌株中各种性状的时间早晚(用分钟表示),画出一幅比较完整的环状染色体图。
,画出一幅比较完整的环状染色体图。")
130
③ F’×F-杂交——性导 性导: F’菌株和F-菌株杂交时会将其带有的染色体基因带入受体细胞,实现基因重组。这种以F’质粒来传递供体基因的方式称为性导,又叫F质粒转导或F因子转导。

131
(三)转导 1、转导的发现: (1) 实验 鼠伤寒沙门菌的一个突变菌株LT22(trp-)和另一个突变菌株LT2(his-)在基本培养基上进行混合培养,结果在107细胞中得到大约100个原养型菌落。
转导 1、转导的发现: (1) 实验 鼠伤寒沙门菌的一个突变菌株LT22(trp-)和另一个突变菌株LT2(his-)在基本培养基上进行混合培养,结果在107细胞中得到大约100个原养型菌落。")
132
(2)解释 1) U形管实验 ——沙门菌的基因重组并不是通过细胞接合,而是通过某些可过滤因子而发生的。 2)可过滤因子的特性 ①可过滤因子并不由于DNA酶的处理而失活; ②可过滤因子和从溶源性的LT22菌株得来的噬菌体(称为P22)具有相同的大小和质量; ③可过滤因子加热后失活,用抗P22血清处理后也失活; ④把抗P22的LT2和LT22菌株混合培养,在基本培养基上不出现原养型菌落。 ——可过滤因子是温和噬菌体P22。

133
2、定义: 通过缺陷噬菌体的媒介,把供体细胞的小片段DNA携带到受体细胞中,通过交换与整合,使后者获得前者部分遗传性状的现象,称为转导。由转导作用而获得部分新性状的重组细胞,称为转导子。

134
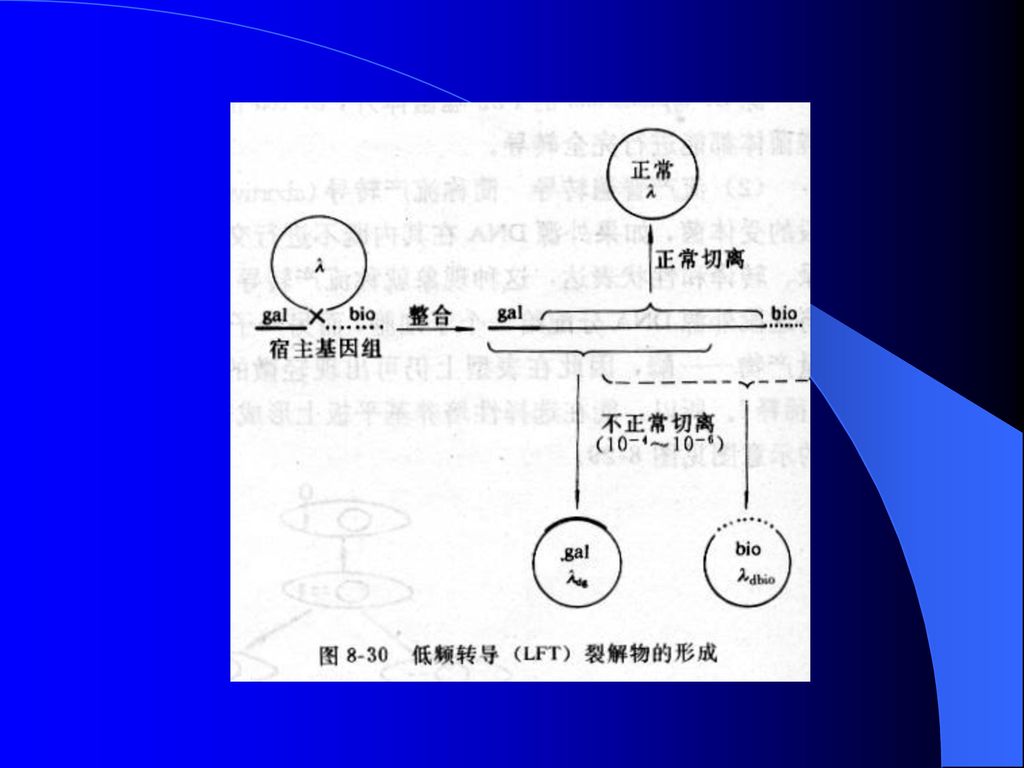
3、转导噬菌体的形成 (1)局限性转导噬菌体(部分缺陷噬菌体): 1)通过错误切离得到——λ phage 坎贝尔模型:
局限性转导噬菌体(部分缺陷噬菌体): 1)通过错误切离得到——λ phage 坎贝尔模型:")
136
2)由于双重溶源菌的裂解而形成: ①双重溶源菌:同时感染有正常噬菌体和缺陷噬菌体的受体菌称为双重溶源菌。 ②过程: 当双重溶源菌被紫外线等诱导时,其中的正常噬菌体的基因可补偿缺陷噬菌体所缺失的部分基因功能,因而两种噬菌体就同时获得复制的机会。由双重溶源菌所产生的裂解物中,含有等量的正常和缺陷噬菌体粒子。

137
(2)普遍性转导噬菌体(完全缺陷噬菌体):通过裂解反应得到——P22 phage
是噬菌体在供体细胞中包装了宿主不同长度的DNA片段所得到的。

138
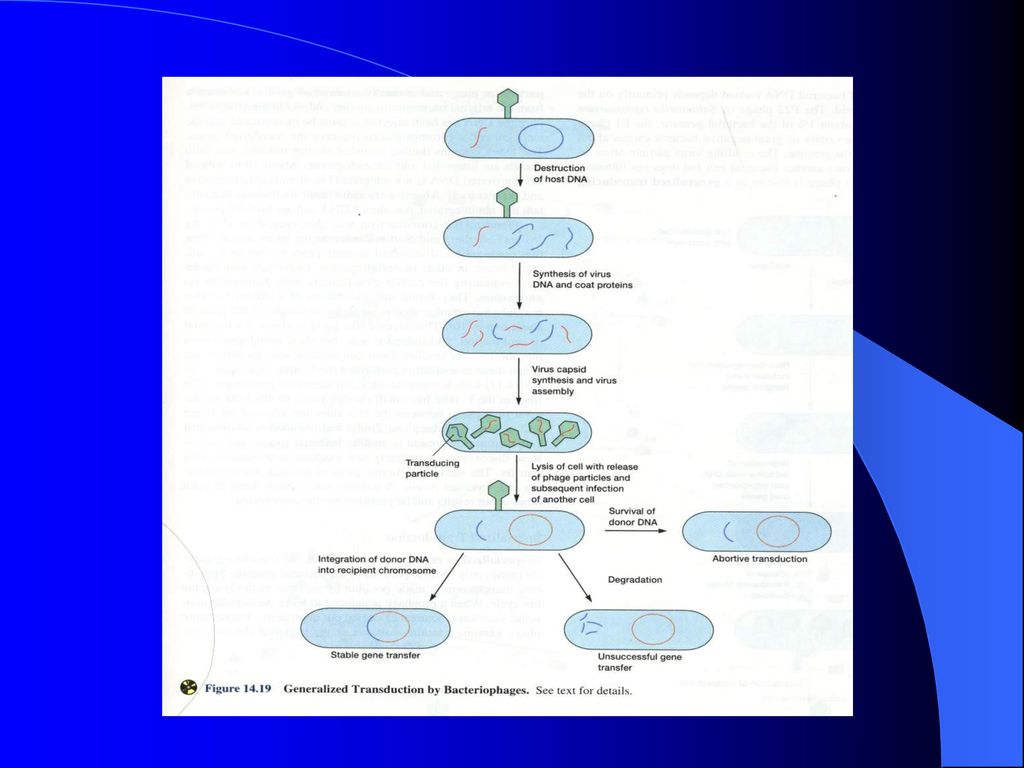
4、转导过程(种类) (1) 普遍转导 1)定义:通过极少数完全缺陷噬菌体对供体菌基因组上任何小片段DNA进行“误包”,而将其遗传性状传递给受体菌的现象,称为普遍转导。
 (1) 普遍转导 1)定义:通过极少数完全缺陷噬菌体对供体菌基因组上任何小片段DNA进行 误包 ,而将其遗传性状传递给受体菌的现象,称为普遍转导。")
139
2)普遍转导中外源DNA的三种后果 ①进入受体的外源DNA通过与细胞染色体的重组交换而形成稳定的转导子——完全普遍转导。 ② 流产转导 ③ 外源DNA被降解,转导失败,在选择平板上无菌落形成
普遍转导中外源DNA的三种后果 ①进入受体的外源DNA通过与细胞染色体的重组交换而形成稳定的转导子——完全普遍转导。 ② 流产转导 ③ 外源DNA被降解,转导失败,在选择平板上无菌落形成")
141
①进入受体的外源DNA通过与细胞染色体的重组交换而形成稳定的转导子——完全普遍转导。
经转导而获得了供体菌DNA片段的受体菌,导入的外源DNA若与受体细胞核染色体组上的同源区段配对,再经过双交换而整合到染色体组上,从而使后者成为一个遗传性状稳定的重组体的现象称为完全普遍转导。形成的转导子称为普遍转导子。

142
b. 过程: 包含着基因转移和基因重组。

143
② 流产转导 a. 定义: 经转导而获得了供体菌DNA片段的受体菌,如果外源DNA在其内既不进行交换、整合和复制,也不迅速消失,而仅进行转录、转译和性状表达,这种现象就称流产转导。

144
b. 过程

145
(2) 局限转导 1)定义:指通过部分缺陷的温和噬菌体把供体菌的少数特定基因携带到受体菌中,并与后者的基因组整合、重组,形成转导子的现象。 2)特点 ① 只能转导供体菌的个别特定基因; ② 该特定基因由部分缺陷的噬菌体携带; ③ 缺陷噬菌体是由于其在形成过程中所发生的低频率 “误切”,或由于双重溶源菌的裂解而形成; ④局限转导噬菌体的产生要通过UV等因素对溶源菌的诱导并引起裂解后才产生。

146
3)局限转导类型: a. 低频转导: 指通过一般溶源菌释放的噬菌体所进行的转导,因其只能形成极少数( )转导子,故称低频转导。 b. 高频转导: 指通过双重溶源菌所产生的裂解物所进行的转导,因其含有等量的正常噬菌体和缺陷噬菌体,具有高频率的转导功能,故称高频转导。
局限转导类型: a. 低频转导: 指通过一般溶源菌释放的噬菌体所进行的转导,因其只能形成极少数( )转导子,故称低频转导。 b. 高频转导: 指通过双重溶源菌所产生的裂解物所进行的转导,因其含有等量的正常噬菌体和缺陷噬菌体,具有高频率的转导功能,故称高频转导。")
147
(3)局限性转导与普遍性转导的主要区别: 1)局限性转导中,被转导的基因共价地与噬菌体DNA连接,与噬菌体DNA一起进行复制、包装以及被导入受体细胞中。而完全转导包装的可能全部是宿主菌的基因; 2)局限性转导颗粒携带特定的染色体片段并将固定的个别基因导入受体,故称为局限性转导。
局限性转导与普遍性转导的主要区别: 1)局限性转导中,被转导的基因共价地与噬菌体DNA连接,与噬菌体DNA一起进行复制、包装以及被导入受体细胞中。而完全转导包装的可能全部是宿主菌的基因; 2)局限性转导颗粒携带特定的染色体片段并将固定的个别基因导入受体,故称为局限性转导。")
148
5、转导和溶源转变 (1)溶源转变:当正常的温和噬菌体感染其宿主而使其发生溶源化时,因噬菌体的基因整合到宿主的核基因组上,而使后者获得了除免疫性以外的新性状的现象,称为溶源转变。 (2)溶源转变与转导有着本质的不同。 首先,这种温和噬菌体并不携带任何来自供体菌的外源基因,使宿主带来新性状的正是噬菌体本身的基因; 其次,这种温和噬菌体是完整的,而不是缺陷的;第三,获得新性状的是溶源化的宿主细胞,而不是什么转导子; 第四,获得的性状可随噬菌体的消失而同时消失。

149
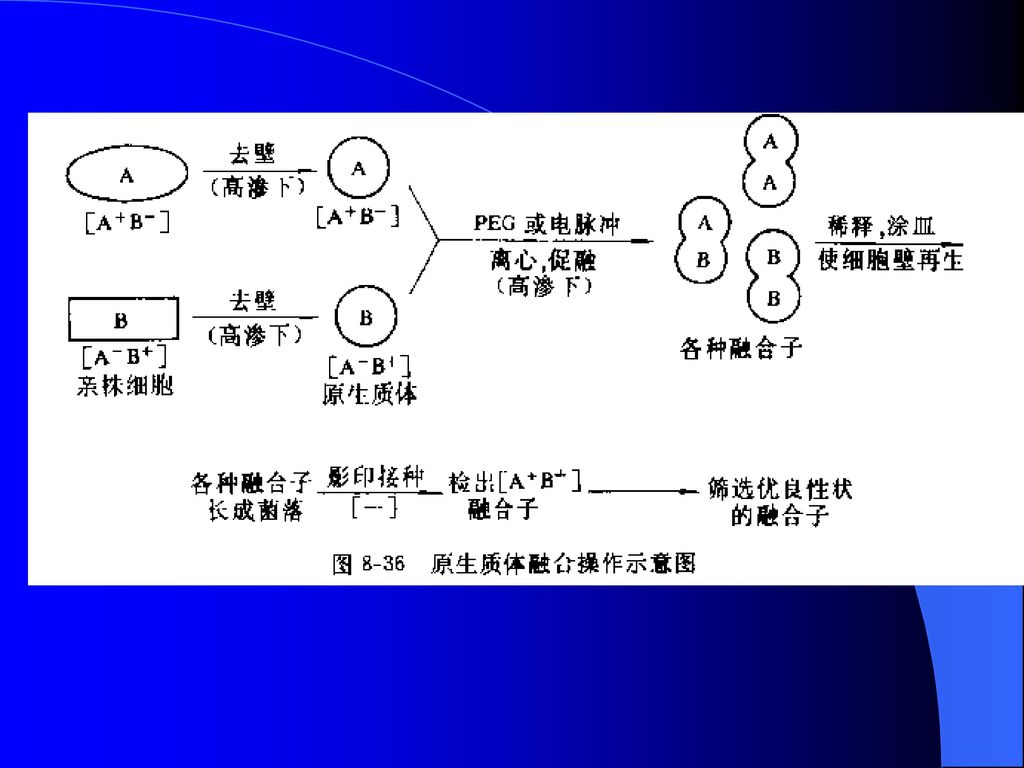
(四)原生质体融合 1、定义 通过人为方法,使遗传性状不同的两细胞的原生质体发生融合,并产生重组子的过程,又称细胞融合。 2、原生质体融合的主要步骤 (1)原生质体制备: 亲本菌株须: ①具有良好生产性状; ②具有一些稳定的明显的遗传标记。 离心收集对数后期菌体,破壁,使用渗透压稳定剂(甘露醇、山梨醇、蔗糖、氯化钾、氯化钠等)进行保护,离心收集原生质体。
进行保护,离心收集原生质体。")
150
(2)原生质体融合: 加入促融合剂-聚乙二醇(PEG)及Ca2+、Mg2+,使原生质体表面形成电极性,相互易于吸引,形成聚集物。UV、电场、激光等技术可应用。 (3)再生成正常细胞: 融合后的原生质体不具细胞壁,不能在普通培养基上增殖,无法表现,必须使其重新形成壁。再生培养基必须具有与原生质体体内相同渗透压,常用含Ca2+、Mg2+及渗透压稳定剂的完全培养基。 (4)检出融合细胞;通过选择性培养基 (5)筛选优良性状的融合子。进行生物测定
检出融合细胞;通过选择性培养基. (5)筛选优良性状的融合子。进行生物测定.")
152
三、真核微生物的基因重组 真核微生物的基因重组方式有有性杂交、准性杂交、原生质体融合和遗传转化等。 杂交:两个性状不同的菌株或变种之间进行细胞结合,遗传物质交换重新组合成新的性状。 (一)有性杂交 1、有性生殖 指由两个生殖细胞或两个体细胞结合造成受精作用,而产生有性孢子的生殖方式。 2、有性杂交 有性杂交是指在微生物的有性繁殖过程中,两个性细胞相互接合,通过质配、核配后形成双倍体的合子,随之合子进行减数分裂,部分染色体可能发生交换而进行随机分配,由此而产生重组染色体及新的遗传型,并把遗传性状按一定的规律性遗传给后代的过程。

153
(二)准性杂交 1、准性生殖: 在同种而不同株的体细胞间发生的融合,不借减数分裂而导致低频率基因重组产生重组子。 2、准性生殖过程 (1)菌丝联结、质配。 (2)形成异核体。 (3)核配。 (4)体细胞交换和单倍体化。
体细胞交换和单倍体化。")
154
3、准性生殖和有性生殖的比较

155
第四节 菌种的衰退、复壮和保藏 一、菌种的退化与复壮 (一) 菌种的退化现象 1、菌种衰退
第四节 菌种的衰退、复壮和保藏 一、菌种的退化与复壮 (一) 菌种的退化现象 1、菌种衰退 生产菌株生产性状的劣化或遗传研究菌株遗传标记的丢失称为菌种衰退。 2、菌种衰退的表现 (1)形态性状 (2)生长速度 (3)代谢能力 (4)侵染力 (5)不良条件抗性
 菌种的退化现象. 1、菌种衰退. 生产菌株生产性状的劣化或遗传研究菌株遗传标记的丢失称为菌种衰退。 2、菌种衰退的表现. (1)形态性状. (2)生长速度. (3)代谢能力. (4)侵染力. (5)不良条件抗性.")
156
(二) 菌种退化的原因 1、自发突变 菌种退化的主要原因是有关基因的负突变。 2、通过诱变获得的高产菌株本身不纯 高产突变只发生在一个核上,随着核的分离,原来未变异的低产性状逐渐恢复。单核微生物由于高产突变只发生在一条DNA链上,也往往发生分离回复的现象。 3、培养、保藏条件 可以通过对自发突变率的影响来表现,也可在不改变基因的情况下表现。

157
(三)防止退化的措施 1、合理的育种 : (1)选育菌种时所处理的细胞应使用单核的,避免使用多核细胞; (2)合理选择诱变剂的种类和剂量或增加突变位点,以减少分离回复;在诱变处理后进行充分的后培养及分离纯化,以保证保藏菌种纯粹。 2、选用合适的培养基; 3、创造良好的培养条件; 4、控制传代次数; 5、利用不同类型的细胞进行移种传代; 6、采用有效的菌种保藏方法。

158
(四)退化菌种的复壮 狭义的复壮 是在菌种已发生衰退的情况下,通过纯种分离和 测定生产性能等方法,从衰退的群体中找出尚未衰退的个体,以达到恢复该菌原有性状的一种措施; 广义的复壮 指在菌种的生产性能尚未衰退前,有意识地进行纯种分离和生产性能的测定工作,以期菌种的生产性能逐步有所提高。 1、纯种分离; 2、通过寄主进行复壮; 3、淘汰已退化个体 。

159
二、微生物菌种保藏 (一)微生物菌种保藏的目的 妥善保藏,达到菌不死、不衰、不乱以及便于研究、交换和使用的目的。 (二)菌种的保藏原则: 1、挑选典型优良纯种(最好是它们的休眠体); 2、创造一个有利于休眠的环境条件; 3、方法的通用性、操作的简便性及设备的普及性。

160
(三)常用的微生物菌种保藏技术 1、斜面传代保藏 将菌种定期在新鲜琼脂斜面培养基上、液体培养基中或穿刺培养,然后在低温条件下保存。它可用于实验室中各类微生物的保藏,此法简单易行,且不要求任何特殊的设备。但此方法易发生培养基干枯、菌体自溶、基因突变、菌种退化、菌株污染等不良现象。

161
2、矿物油中浸没保藏 可用于丝状真菌、酵母、细菌和放线菌的保藏。特别对难于冷冻干燥的丝状真菌和难以在固体培养基上形成孢子的担子菌等的保藏更为有效。是将琼脂斜面或液体培养物或穿刺培养物浸入矿物油中于室温下或冰箱中保藏。 3、干燥-载体保藏 适用于产孢子或芽孢的微生物的保藏。是将菌种接种于适当的载体上,如河砂、土壤、硅胶、滤纸及麸皮等,以保藏菌种。以沙土保藏用得较多。

162
4、冷冻保藏 是指将菌种于-20℃以下的温度保藏。 (1)普通冷冻保藏技术(-20℃): (2)超低温冷冻保藏技术:要求长期保藏的微生物菌种,一般都应在-60℃以下的超低温冷藏柜中进行保藏。 (3)液氮冷冻保藏技术: 在液氮中保藏的菌种的存活率远比其他保藏方法高且回复突变的发生率极低。液氮保藏巳成为工业微生物菌种保藏的最好方法。

163
5、真空冻干保藏 此法是微生物菌种长期保藏的最为有效的方法之一,大部分微生物菌种可以在冻干状态下保藏10年之久而不丧失活力。而且经冻干后的菌株无需进行冷冻保藏,便于运输。但操作过程复杂,并要求一定的设备条件。 6、寄主保藏 适用于一些难于用常规方法保藏的动植物病原菌和病毒。 7、基因工程菌的保藏 基因工程菌最好应保藏在含低浓度选择剂的培养基中。









:核蛋白体组成成分 转移 RNA ( tRNA ):转运氨基酸 信使 RNA ( mRNA ):蛋白质合成模板 不均一核 RNA ( hnRNA ):成熟 mRNA 的前体 小核 RNA ( snRNA ):>")










 A.染色体 B.质体 C.线粒体 D.核糖体>")




