Download presentation
Presentation is loading. Please wait.

1
遺傳學重大的發現

2
遺傳法則 Laws of Inheritance
十九世紀末奧地利孟德爾(Gregor Mendel)神父,在教堂花圃中從事豌豆的雜交實驗,他是第一位有實驗設計(規劃如何配對雜交)、詳細記錄實驗結果,再利用數學方法分析實驗結果,歸納出遺傳法則而被尊稱為遺傳學之父。 孟德爾的研究起初沒有受到太大的重視,直到1900年才被重新發現。遺傳法則雖然無法完全解釋所有的遺傳現象,卻奠定了現代遺傳學的基礎。
神父,在教堂花圃中從事豌豆的雜交實驗,他是第一位有實驗設計(規劃如何配對雜交)、詳細記錄實驗結果,再利用數學方法分析實驗結果,歸納出遺傳法則而被尊稱為遺傳學之父。 孟德爾的研究起初沒有受到太大的重視,直到1900年才被重新發現。遺傳法則雖然無法完全解釋所有的遺傳現象,卻奠定了現代遺傳學的基礎。")
3
孟德爾的三大遺傳法則:(1)顯性法則 (2)分離法則 (3)自由分配法則。
顯性法則:控制生物遺傳性狀的基因有顯性與隱性之別,可分別用英文字母的大寫及小寫來代表。當顯性與隱性基因同時存在的情況下,只有顯性基因控制的性狀才會表現出來。 分離法則:成對的遺傳因子彼此獨立不混合,並在形成配子時彼此分離。

4
自由分配法則:種子各自的特點的遺傳方式沒有相互影響,每一項特徵都符合顯性法則以及分離法則,這被稱為自由分配法則。另外值得一提的是在孟德爾死後,發現這一法則只在一定的條件下方能成立。

5
基因在染色體上 Genes Are Located on Chromosomes
托馬斯·亨特·摩爾根(Thomas Hunt Morgan),美國遺傳學家、現代遺傳學之父。研究果蠅的遺傳突變,確立了染色體是基因的載體。發現位於同一染色體上的基因之間的鏈鎖遺傳特性,將多種突變基因定位在染色體上,製成染色體圖譜,即基因的連鎖圖。 繼承並發展孟德爾以豌豆雜交實驗為基礎的遺傳理論,為生物學發展為實驗科學奠定基礎。 獲1933年諾貝爾生理醫學獎。
,美國遺傳學家、現代遺傳學之父。研究果蠅的遺傳突變,確立了染色體是基因的載體。發現位於同一染色體上的基因之間的鏈鎖遺傳特性,將多種突變基因定位在染色體上,製成染色體圖譜,即基因的連鎖圖。 繼承並發展孟德爾以豌豆雜交實驗為基礎的遺傳理論,為生物學發展為實驗科學奠定基礎。 獲1933年諾貝爾生理醫學獎。")
6
基因控制生化事件 Genes Control Biochemical Events
1942年George Beadle與Edward Tatum 以麵包上的紅黴菌實驗證實,DNA上所帶遺傳訊息的功能是製造特定的酵素。 他們獲得1958年諾貝爾生理醫學獎。

7
轉位子 Transposons 轉位子又稱為『跳躍基因』,是美國科學家Brarbara MacClintock所發現的,也是1983年的諾貝爾生理醫學獎的唯一得主。 她研究玉米過程中發現轉位子是一種DNA序列,它們能夠在基因組中通過轉錄和逆轉錄,或在內切酶的作用下,在其他基因座上出現。 轉座子的發現,證明了基因組並不是一個靜態的集合,而是一個不斷在改變自身構成的動態有機體。

8
此發現對AIDS的研究有極大的貢獻。因為HIV病毒也是利用和轉位子類似的方式,將它們自己插入人類的基因體中。對於這些酵素的研究指出,它們的核心結構十分接近,這意味著它們極可能利用同樣的機制作用。
應用轉位子製造突變體是基因篩選工作重要的工具,例如,將某一藥用植物中已轉位子製造出不在產生該藥品的突變體,立刻可由該轉位子選殖出基因,許多植物抗病基因就是轉位子選殖出來的。

9
DNA攜帶遺傳物質 DNA Carries Genetic Material
主要功能是資訊儲存,其中包含的指令,是建構細胞內其他的化合物(如蛋白質與核糖核酸)所需的。 帶有遺傳訊息的去氧核糖核酸片段稱為基因。其他的去氧核糖核酸序列,有些直接以本身構造發揮作用,有些則參與調控遺傳訊息。
所需的。 帶有遺傳訊息的去氧核糖核酸片段稱為基因。其他的去氧核糖核酸序列,有些直接以本身構造發揮作用,有些則參與調控遺傳訊息。")
10
去氧核糖核酸是一種長鏈聚合物,組成單位稱為核苷酸(鹼基、糖類分子與磷酸三者的結合);糖類分子與磷酸藉由酯鍵相連,組成其長鏈骨架。
每個糖類分子都與四種鹼基(腺嘌呤(A)、胞嘧啶(C)、鳥嘌呤(G)與胸腺嘧啶(T))中的一種相接,這些鹼基沿著去氧核糖核酸長鏈所排列而成的序列,可組成遺傳密碼,是蛋白質胺基酸序列合成的依據。
、胞嘧啶(C)、鳥嘌呤(G)與胸腺嘧啶(T))中的一種相接,這些鹼基沿著去氧核糖核酸長鏈所排列而成的序列,可組成遺傳密碼,是蛋白質胺基酸序列合成的依據。")
11
在細胞內去氧核糖核酸能組成染色體結構;整組染色體則統稱為基因組。
對真核生物(如動物、植物及真菌)而言,染色體是存放於細胞核內;對於原核生物(如細菌)而言,則是存放在細胞質中的擬核裏。 人類細胞中最大的1號染色體中,就有2億2千萬個鹼基對。
而言,染色體是存放於細胞核內;對於原核生物(如細菌)而言,則是存放在細胞質中的擬核裏。 人類細胞中最大的1號染色體中,就有2億2千萬個鹼基對。")
12
重組去氧核糖核酸技術在現代生物學與生物化學中受到廣泛應用,所謂重組DNA,是指集合其他去氧核糖核酸序列所製成的人造去氧核糖核酸,由運載體將去氧核糖核酸轉型到受體細胞中。
經過遺傳改造處裡之後的生物體,可用來生產重組蛋白質,以供醫學研究使用,或是於農業上栽種。

13
刑事鑑定可利用犯罪現場遺留的血液、精液、皮膚、唾液或毛髮中的去氧核糖核酸,來辨識可能的加害人。此過程稱為遺傳指紋分析或去氧核糖核酸特徵測定。
此測定法於1984年由英國遺傳學家阿萊克·傑里斯發展出來的。 1988年英國的謀殺案嫌犯科林·皮奇福克,成為第一位因去氧核糖核酸特徵測定證據而遭定罪者。 去氧核糖核酸特徵測定也可用來辨識重大災害中的罹難者。

14
1869年瑞士醫生弗雷德里希·米歇爾最早分離出去氧核糖核酸。他從廢棄繃帶裡所殘留的膿液中發現。
1919年菲巴斯·利文辨識出組成去氧核糖核酸的鹼基、糖類以及磷酸核苷酸單元。 1937年威廉·阿斯特伯里完成了第一張X光繞射圖,闡明了去氧核糖核酸結構的規律性。 1953年阿弗雷德·赫希與瑪莎·蔡斯確認了去氧核糖核酸的遺傳功能。 1953年詹姆斯·沃森與佛朗西斯·克里克,提出核酸分子結構精確模型。1962年沃森、克里克以及威爾金斯共同獲得諾貝爾生理醫學獎。 1958年馬修·梅瑟生與富蘭克林·史達確認了去氧核糖核酸的複製機制。 1961年哈爾·葛賓·科拉納、羅伯特·W·霍利以及馬歇爾·沃倫·尼倫伯格解出遺傳密碼。1968年科拉納和尼倫伯格獲得諾貝爾生理醫學獎。 1990年代展開人類基因組計畫。 2001年多國合作的國際團隊與私人企業塞雷拉基因組公司,分別將人類基因組序列草圖發表於《自然》與《科學》兩份期刊。

15
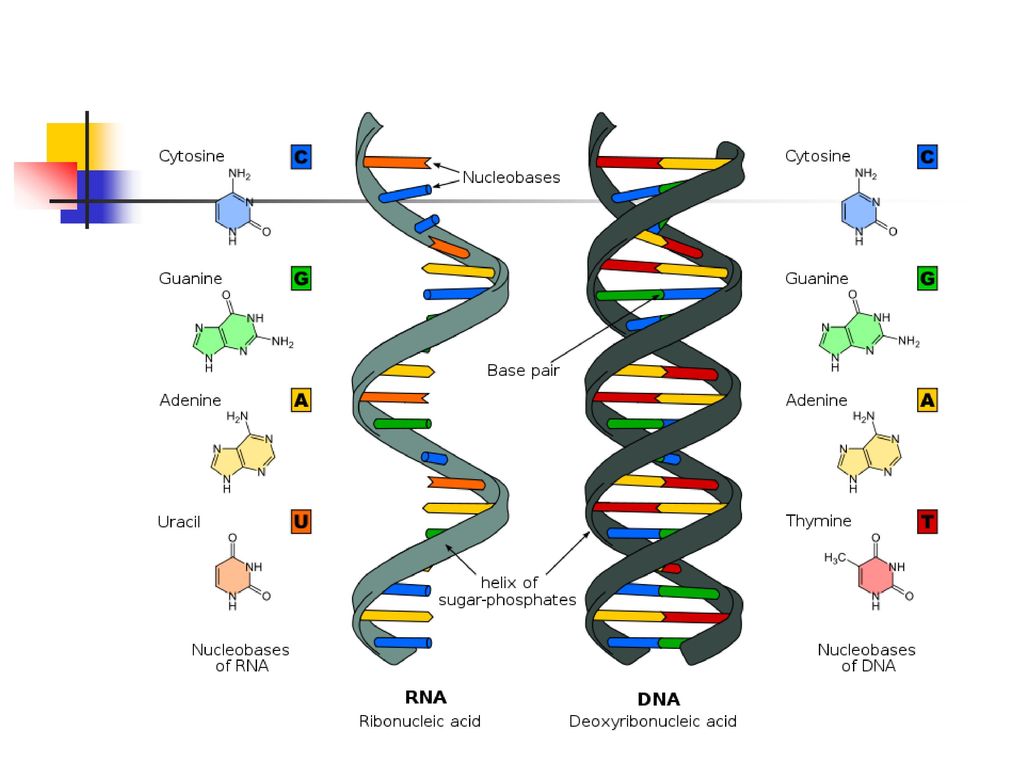
雙股螺旋狀的DNA DNA: Double Helix
去氧核糖核酸的結構是兩條互相配對並緊密結合的雙螺旋分子。 一股去氧核糖核酸上的各類鹼基,都只會與另一股上的一個特定類型鹼基產生鍵結。此種情形稱為互補性鹼基配對。 嘌呤與嘧啶之間藉由氫鍵聯結。 一般情況下,A只與T相連,而C只與G相連。

17
信使RNA Messenger RNA 信使RNA(mRNA),是由DNA經由轉錄而來,帶著相應的遺傳訊息,為下一步轉譯成蛋白質提供所需的訊息。 在許多真核生物中,反義RNA(anti-sense RNA)上的鹼基與基因的某段mRNA互補時,反義RNA可以抑制轉譯。反義RNA存在於細胞時,其互補的基因就不會合成蛋白質。
上的鹼基與基因的某段mRNA互補時,反義RNA可以抑制轉譯。反義RNA存在於細胞時,其互補的基因就不會合成蛋白質。")
18
這也許是對抗病毒複製的一種機制。 在生物化學的研究中,藉由使用一段反義mRNA使得標靶mRNA無法作用(RNA干擾),這種方法已經被廣泛用於基因研究工作。 例如利用RNA干擾技術,秀麗隱桿線蟲中所有基因的作用幾乎已經被研究完了。
,這種方法已經被廣泛用於基因研究工作。 例如利用RNA干擾技術,秀麗隱桿線蟲中所有基因的作用幾乎已經被研究完了。")
19
遺傳密碼 Genetic Code 自從發現了DNA的結構,科學家便開始致力研究有關製造蛋白質的秘密。首先伽莫夫指出需要以三個核酸一組才能為20個胺基酸編碼。 1961年,美國國家衛生院的Matthaei與馬歇爾·沃倫·尼倫伯格在無細胞系統環境下,把一條只由尿嘧啶(U)組成的RNA轉釋成一條只有苯丙氨酸(Phe)的多肽,由此破解了首個密碼子(UUU -> Phe)。
組成的RNA轉釋成一條只有苯丙氨酸(Phe)的多肽,由此破解了首個密碼子(UUU -> Phe)。")
20
隨後哈爾·葛賓·科拉納破解了其它密碼子,接著羅伯特·W·霍利發現負責轉錄過程的tRNA。
1968年,科拉納、霍利和尼倫伯格分享了諾貝爾生理醫學獎。 遺傳密碼是一組規則,將DNA或RNA序列以三個核苷酸為一組的密碼子轉譯為蛋白質的胺基酸序列,以用於蛋白質合成。 幾乎所有的生物都使用同樣的遺傳密碼,稱為標準遺傳密碼;即使是非細胞結構的病毒,它們也是使用標準遺傳密碼。但是也有少數生物使用一些稍微不同的遺傳密碼。

21
限制酶 Restriction Enzymes
限制酶又稱限制內切酶或限制性內切酶,全稱限制性核酸內切酶,是一種能將雙股DNA切開的酵素。 切割方法是將糖類分子與磷酸之間的鍵結切斷,進而於兩條DNA鏈上各產生一個切口,且不破壞核苷酸與鹼基。 切割形式有兩種,分別是可產生具有突出單股DNA的黏狀末端,以及末端平整無凸起的平滑末端。

22
由於斷開的DNA片段可由另一種稱為DNA連接酶的酵素黏合,因此染色體或DNA上不同的限制片段,得經由剪接作用而結合在一起。
限制酶在分子生物學與遺傳工程領域有廣泛的應用,此類酵素最早發現於某些品系的大腸桿菌體內,這些品系能夠「限制」噬菌體對其感染,因此得名。 科學家認為限制酶是細菌所演化出來對抗病毒感染,並幫助將已殖入的病毒序列移除的機制。是限制修飾系統的一部分。

23
約翰霍普金斯大學的丹尼爾·那森斯、漢彌爾頓·史密斯與伯克萊加州大學的沃納·亞伯因為限制酶的發現及研究,而共同獲得1978年的諾貝爾生理醫學獎。
此酵素最早的應用之一,是用來將胰島素基因轉殖到大腸桿菌,使其具備生產人類胰島素的能力。

24
RNA選擇性剪接 RNA Alternative Splicing
真核細胞的基因序列中,包含了內含子與外顯子,兩者交互穿插。其中內含子在基因轉錄成mRNA前體後會被RNA剪接體移除,剩下的外顯子才是能夠存在於成熟mRNA(之後再進一步轉譯成蛋白質)的片段。
的片段。")
25
選擇性剪接便是利用這樣的特性,一條未經剪接的RNA,含有的多種外顯子被剪成的不同組合,可轉譯出不同的蛋白質。
能將同一基因中的外顯子以不同的組合方式來表現,使一個基因在不同時間、不同環境中能夠製造出不同的蛋白質(基因表達調控),可增加生理狀況下系統的複雜性或適應性。
,可增加生理狀況下系統的複雜性或適應性。")
26
小衛星DNA Minisatellite DNA
小衛星DNA或小衛星序列是一段含有約10到100個鹼基對的DNA片段,在人類基因組中約出現在1000個以上的位置。 某些小衛星序列含有一個核心序列「GGGCAGGAXG」(X表示任何字母),其他大多則是一股皆嘌呤,另一股皆嘧啶。
,其他大多則是一股皆嘌呤,另一股皆嘧啶。")
27
在人體內,約90%的小衛星序列出現在靠近端粒的位置,端粒本身則主要是相連重複(tandem repeat)。
阿萊克·傑弗里斯爵士是英國遺傳學家,最早的DNA指紋分析及DNA特徵測定技術發展者。其技術首次在1983到1986年間的謀殺案應用,使嫌犯柯林·皮區佛克(Colin Pitchfork)因DNA證據而被定罪。
因DNA證據而被定罪。")
28
RNA干擾現象 RNA Interference
RNA干擾(RNAi)是指一種分子生物學上由雙鏈RNA誘發的基因沉默現象,其機制是通過阻礙特定基因的翻譯或轉錄來抑制基因表達。 當細胞中導入與內源性mRNA編碼區同源的雙鏈RNA時,該mRNA發生降解而導致基因表達沉默。與其它基因沉默現象不同的是,在植物和線蟲中,RNAi具有傳遞性,可在細胞之間傳播,此現象被稱作系統性RNA干擾。
是指一種分子生物學上由雙鏈RNA誘發的基因沉默現象,其機制是通過阻礙特定基因的翻譯或轉錄來抑制基因表達。 當細胞中導入與內源性mRNA編碼區同源的雙鏈RNA時,該mRNA發生降解而導致基因表達沉默。與其它基因沉默現象不同的是,在植物和線蟲中,RNAi具有傳遞性,可在細胞之間傳播,此現象被稱作系統性RNA干擾。")
29
在秀麗隱桿線蟲上實驗時還可使子一代產生基因突變,甚到於可用餵食細菌給線蟲的方式讓線蟲得以產生RNA干擾現象。
RNAi現象在生物中普遍存在。

30
1990年約根森(Jorgensen)研究小組發現RNA干擾現象。
1992年羅馬諾(Romano)和Macino在粗糙鏈孢霉中發現了外源導入基因可以抑制具有同源序列的內源基因的表達。 1995年Guo和Kemphues在線蟲中也發現RNA干擾現象。 1998年,安德魯·法厄(Andrew Z. Fire)等在秀麗隱桿線蟲中進行反義RNA抑制實驗時發現,作為對照加入的雙鏈RNA相比正義或反義RNA顯示出了更強的抑制效果。從與靶mRNA的分子量比考慮,加入的雙鏈RNA的抑制效果要強於理論上1:1配對時的抑制效果,因此推測在雙鏈RNA引導的抑制過程中存在某種擴增效應並且有某種酶活性參與其中。並將這種現象命名為RNA干擾。 2006年,安德魯·法厄與克雷格·梅洛(Craig C. Mello)由於在RNAi機制研究中的貢獻獲得諾貝爾生理醫學獎。
和Macino在粗糙鏈孢霉中發現了外源導入基因可以抑制具有同源序列的內源基因的表達。 1995年Guo和Kemphues在線蟲中也發現RNA干擾現象。 1998年,安德魯·法厄(Andrew Z. Fire)等在秀麗隱桿線蟲中進行反義RNA抑制實驗時發現,作為對照加入的雙鏈RNA相比正義或反義RNA顯示出了更強的抑制效果。從與靶mRNA的分子量比考慮,加入的雙鏈RNA的抑制效果要強於理論上1:1配對時的抑制效果,因此推測在雙鏈RNA引導的抑制過程中存在某種擴增效應並且有某種酶活性參與其中。並將這種現象命名為RNA干擾。 2006年,安德魯·法厄與克雷格·梅洛(Craig C. Mello)由於在RNAi機制研究中的貢獻獲得諾貝爾生理醫學獎。")
31
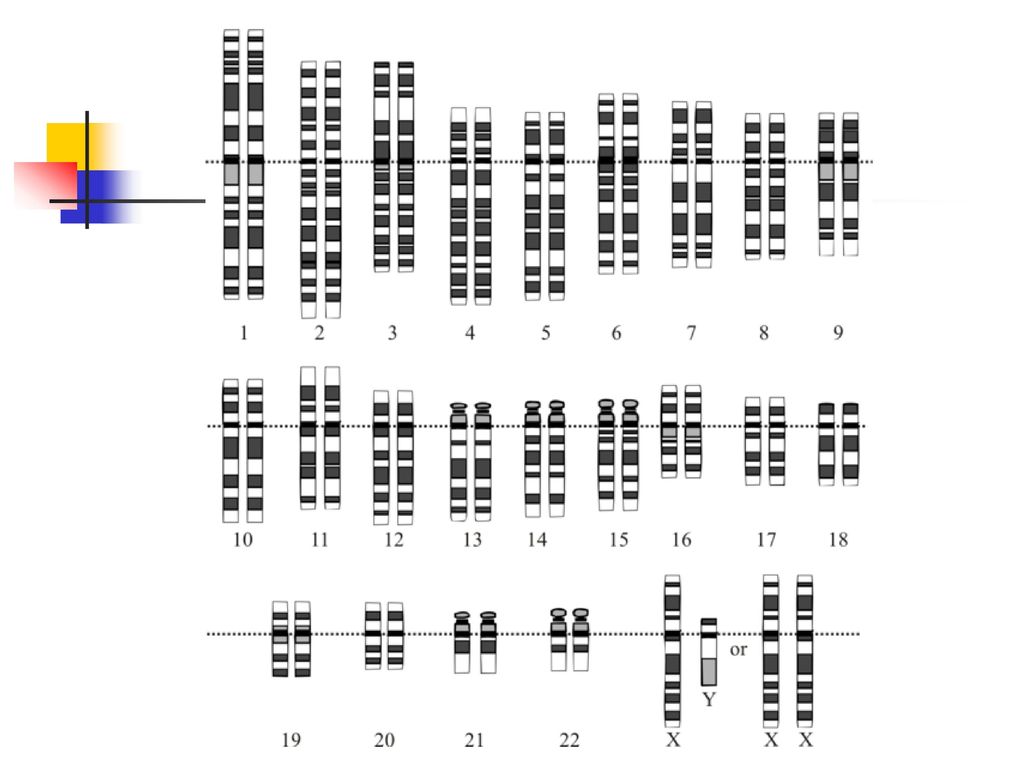
人類有25,000個基因 Humans Have 25,000 Genes
人類基因組又稱人類基因體,由23對染色體組成,其中包括22對體染色體、1條X染色體和1條Y染色體。 人類基因組含有約30億個DNA鹼基對,鹼基對是以氫鍵相結合的兩個含氮鹼基,以胸腺嘧啶(T)、腺嘌呤(A)、胞嘧啶(C)和鳥嘌呤(G)四種鹼基排列成鹼基序列,其中A與T之間由兩個氫鍵連接,G與C之間由三個氫鍵連接,鹼基對的排列在DNA中也只能是A對T,G對C。其中一部分的鹼基對組成了大約20000到25000個基因。
、腺嘌呤(A)、胞嘧啶(C)和鳥嘌呤(G)四種鹼基排列成鹼基序列,其中A與T之間由兩個氫鍵連接,G與C之間由三個氫鍵連接,鹼基對的排列在DNA中也只能是A對T,G對C。其中一部分的鹼基對組成了大約20000到25000個基因。")
32
人類擁有23種不同的染色體,其中有22個屬於體染色體,另外還有兩個能夠決定性別的性染色體,分別是X染色體與Y染色體。
1號到22號染色體的編號順序,大致符合他們由大到小的尺寸排列。 最大的染色體約含有2億5千萬個鹼基對,最小的則約有3800萬個鹼基對。 這些染色體通常以細絲狀存於細胞核內,若將單一細胞內的染色體拉成直線,那麼將大約有6英尺長。

33
在人類個體的體細胞中,通常含有來自親代的1到22對體染色體,再加上來自母親的X染色體,以及來自父親的X或Y染色體,總共是46個(23對)染色體。
科學家將染色體分為6組:1號到3號是A組;4號與5號是B組;X染色體以及6號到12號是C組;13號到15號是D組;16號到18號是E組;19號與20號是F組;21號、22號與Y染色體是G組。 對於一般人類來說,每個細胞核內只有兩套染色體。

34
人體內估計約有20000到25000個蛋白質編碼基因。 雖然人類的基因數量比起某些較為原始的生物(如線蟲與果蠅)更少,但是在人類細胞中使用了大量的選擇性剪接(alternative splicing;將穿插在內含子中的外顯子以選擇性的方式進行轉錄),這使得一個基因能夠製造出多種不同的蛋白質,且人類的蛋白質組規模也較前述的兩個物種更龐大。
更少,但是在人類細胞中使用了大量的選擇性剪接(alternative splicing;將穿插在內含子中的外顯子以選擇性的方式進行轉錄),這使得一個基因能夠製造出多種不同的蛋白質,且人類的蛋白質組規模也較前述的兩個物種更龐大。")
35
除了蛋白質編碼基因之外,人類的基因組還包含了數千個RNA基因(製造非編碼RNA),其中包括用來轉錄轉運RNA(tRNA)、核糖體RNA(rRNA)與信使RNA(mRNA)的基因。其中轉錄rRNA的基因稱為rDNA,分佈在許多不同的染色體上。
,其中包括用來轉錄轉運RNA(tRNA)、核糖體RNA(rRNA)與信使RNA(mRNA)的基因。其中轉錄rRNA的基因稱為rDNA,分佈在許多不同的染色體上。")
Similar presentations
 ” 。 他的体内没有任何免疫系统, 没有任何抵御细菌、病毒的能力。>")

:核蛋白体组成成分 转移 RNA ( tRNA ):转运氨基酸 信使 RNA ( mRNA ):蛋白质合成模板 不均一核 RNA ( hnRNA ):成熟 mRNA 的前体 小核 RNA ( snRNA ):>")

 染色體:細胞核上(細胞未分裂前稱為染色質) 原核生物:DNA>")








 A.细菌的遗传物质主要是DNA B.病毒的遗传物质主要是RNA>")





