Download presentation
1
第五章 屠宰后肉的变化 孔保华

2
引言 热鲜肉→肉的尸僵→解僵成熟→自体酶解→腐败变质 muscle to meat 动物刚屠宰后,肉温还没有散失,柔软具有较小弹性,这种处于生鲜状态的肉称作热鲜肉。 经过一定时间,肉的伸展性消失,肉体变为僵硬状态,这种现象称为死后僵直(rigor mortis),此时肉加热食用是很硬的,而且持水性也差,因此加热后重量损失很大,不适于加工。
,此时肉加热食用是很硬的,而且持水性也差,因此加热后重量损失很大,不适于加工。")
3
继续贮藏,其僵直情况会缓解,经过自身解僵,肉又变得柔软起来,同时持水性增加,风味提高,所以在利用肉时,一般应解僵后再使用,此过程称作肉的成熟(conditioning)。
成熟肉在不良条件下贮存,经酶和微生物作用分解变质称作肉的腐败(putrefaction)。 屠宰后肉的变化,即包括上述肉的尸僵、肉的成熟、肉的腐败三个连续变化过程。在肉品工业生产中,要控制尸僵、促进成熟、防止腐败。
。 屠宰后肉的变化,即包括上述肉的尸僵、肉的成熟、肉的腐败三个连续变化过程。在肉品工业生产中,要控制尸僵、促进成熟、防止腐败。")
4
第一节 肌肉收缩的机制 第二节 肉的僵直 第三节 肉的成熟 第四节 肉的腐败变质

5
第一节 肌肉收缩的机制 肌肉收缩的基本单位 肌肉收缩与松弛的生物化学机制

6
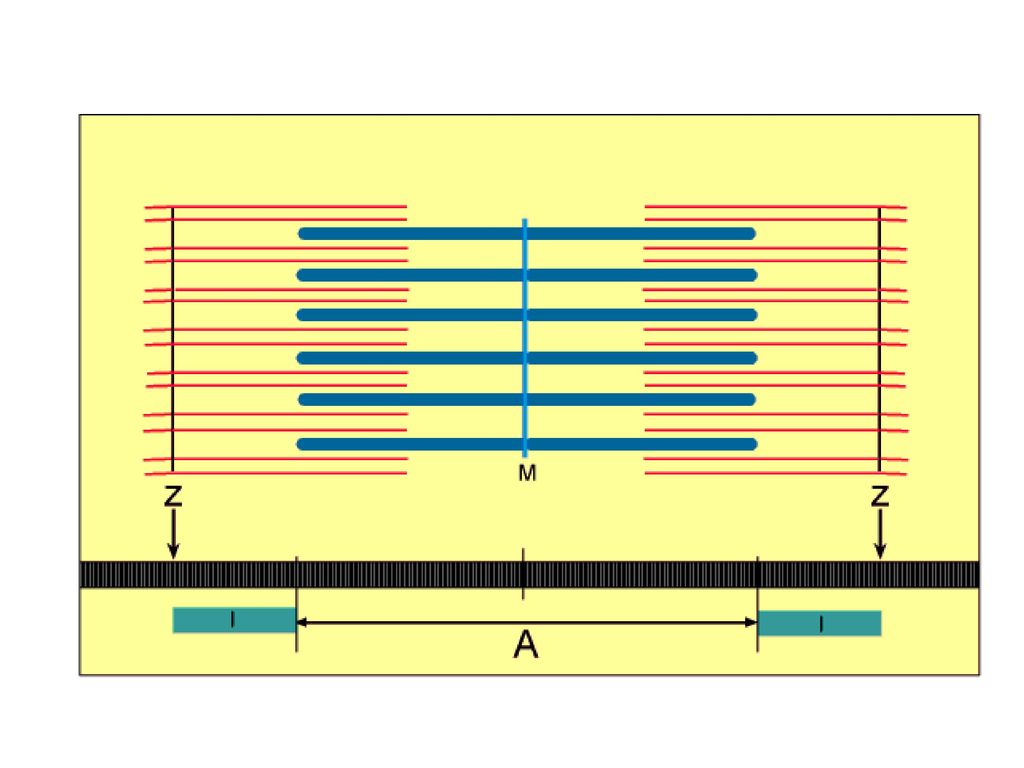
肌肉收缩的基本单位

7
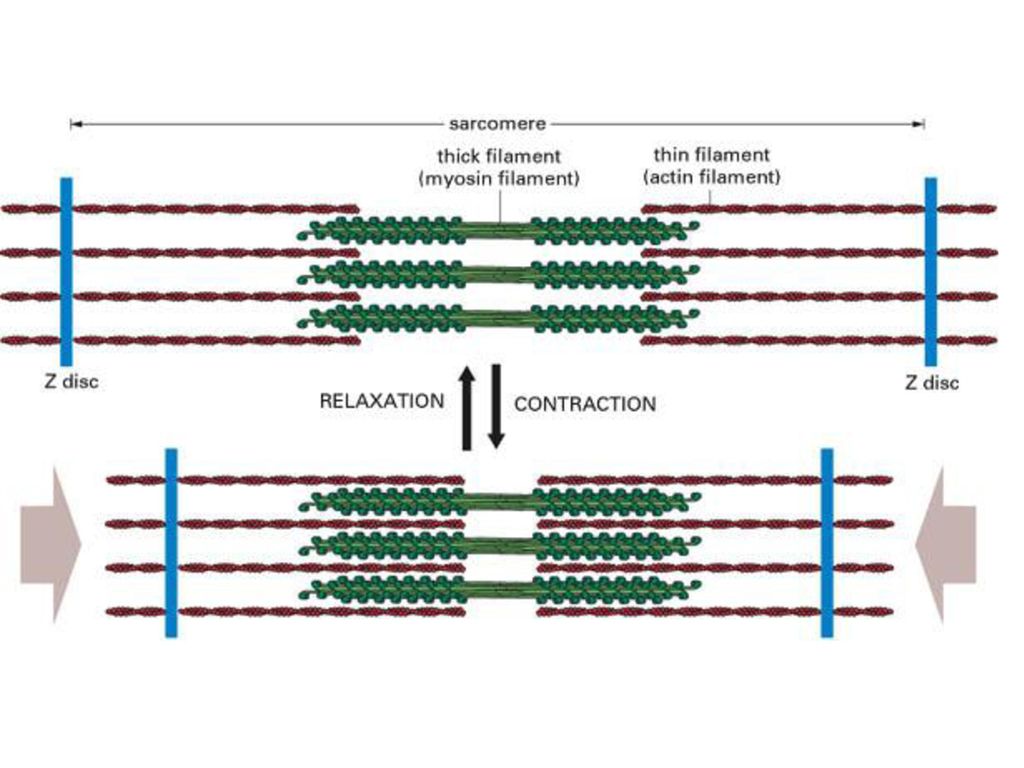
在每一条肌球蛋白粗丝的周围,有六对肌动蛋白纤丝,围绕排列而构成六方格状结构。在每个肌球蛋白粗丝的周围,有放射状的突起,这些突起呈螺旋状排列,每六个突起排列位置恰好旋转一周。在突起上含有ATP酶的活性中心的重酶解肌球蛋白,并能和F─肌动蛋白结合。粗丝和细丝不是永久性结合的,由于某些因素会产生离合状态,便产生肌肉的伸缩。即所说的肌肉收缩和松弛。

8
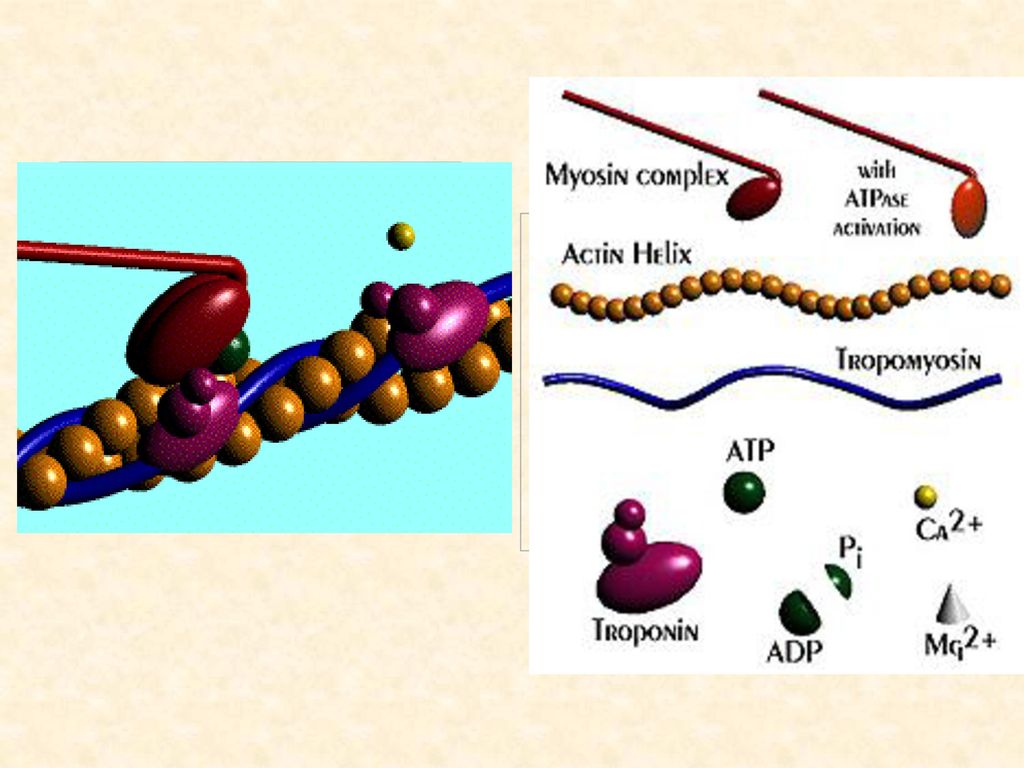
肌肉收缩包括以下四种主要因子 (1)收缩因子 肌球蛋白(myosin)、肌动蛋白(actin)、原肌球蛋白(tropomyosin)和肌原蛋白(troponin)。 (2)能源 ATP (3)调节因子 初级调节因子─钙离子,次级调节因子─原肌球蛋白和肌原蛋白。 (4)疏松因子 肌质网系统(sarcoplasmic reticulum system)和钙离子泵。
疏松因子 肌质网系统(sarcoplasmic reticulum system)和钙离子泵。")
14
生活的肌肉处于静止状态时,由于Mg和ATP形成复合体的存在,防碍了肌动蛋白与肌球蛋白粗丝突起端的结合。
肌球蛋白头是一种ATP酶,这种酶的激活需要Ca2+的激活。

15
神经冲动→肌内膜→肌质网释放Ca2+ → →使肌动蛋白暴露与肌球蛋白结合位点 →使ATP酶活化→ATP分解产生能量 →肌动蛋白与肌球蛋白结合→收缩

16
★肌肉收缩时首先由神经系统(运动神经)传递信号,来自大脑的信息经神经纤维传到肌原纤维膜产生去极化作用,神经冲动沿着T小管进入肌原纤维,可促使肌质网将Ca2+释放到肌浆中。
★进入肌浆中的Ca2+浓度从10-7 Mol增高到10-5 Mol时,钙离子即与细丝的肌原蛋白钙结合亚基(TnC)结合,引起肌原蛋白三个亚单位构型发生变化,使原肌球蛋白更深地移向肌动蛋白的螺旋槽内,从而暴露出肌动蛋白纤丝上能与肌球蛋白头部结合的位点。
结合,引起肌原蛋白三个亚单位构型发生变化,使原肌球蛋白更深地移向肌动蛋白的螺旋槽内,从而暴露出肌动蛋白纤丝上能与肌球蛋白头部结合的位点。")
17
★钙离子可以使ATP从其惰性的Mg--ATP复合物中游离出来,并刺激肌球蛋白的ATP酶,使其活化。
★ ATP酶被活化后,将ATP分解为ADP + Pi + 能量,同时肌球蛋白纤丝的突起端点与肌动蛋白纤丝结合,形成收缩状态的肌动球蛋白。

18
★ 当神经冲动产生的动作电位消失,通过肌质网钙泵作用,肌浆中的钙离子被收回。肌原蛋白钙结合亚基(TN-C)失去Ca2+,肌原蛋白抑制亚基(TN-l)又开始起控制作用。
★ ATP与Mg形成复合物,且与肌球蛋白头部结合。而细丝上的原肌球蛋白分子又从肌动蛋白螺旋沟中移出,挡往了肌动蛋白和肌球蛋白结合的位点,形成肌肉的松驰状态。 如果ATP供应不足,则肌球蛋白头部与肌动蛋白结合位点不能脱离,使肌原纤维一直处于收缩状态,这就形成尸僵。

19
第二节 肉的僵直 一、屠宰后肌肉糖原的酵解 二、死后僵直的机制 三、死后僵直的过程 四、死后僵直的类型 五、冷收缩及解冻僵直收缩
六、尸僵和保水性的关系 七、尸僵开始和持续时间 第二节 肉的僵直

20
尸僵的定义 屠宰后的肉尸(胴体)经过一定时间,肉的伸展性逐渐消失,由弛缓变为紧张,无光泽,关节不活动,呈现僵硬状态,叫作尸僵。
尸僵的肉硬度大,加热时不易煮熟,有粗糙感,肉汁流失多,缺乏风味,不具备可食肉的特征。这样的肉从相对意义上讲不适于加工和烹调。

21
一、屠宰后肌肉糖原的酵解 糖酵解作用 酸性极限pH值

22
糖酵解作用 动物屠宰以后,糖原的含量会逐渐减少,动物死后血液循环停止,供给肌肉的氧气也就中断了,其结果促进糖的无氧酵解过程,糖原形成乳酸,直至下降到抑制糖酵解酶的活性为止。 有氧代谢,一个葡萄糖可产生39 个ATP,而糖酵解作用,一个葡萄糖可产生3 个ATP,使能量的产生大大减少。

24
酸性极限pH值 一般活体肌肉的pH值保持中性(7.0~7.2),死后由于糖原酵解生成乳酸,肉的pH值逐渐下降,一直到阻止糖原酵解酶的活性为止,这个pH值称极限pH值。
,死后由于糖原酵解生成乳酸,肉的pH值逐渐下降,一直到阻止糖原酵解酶的活性为止,这个pH值称极限pH值。")
25
哺乳动物肌肉的极限pH值为5.4~5.5之间,达到极限pH值时大部分糖原已被消耗,这时即使残留少量糖原,由于糖酵解酶的钝化,也不能继续分解了。肉的pH值下降对微生物,特别是对细菌的繁殖有抑制作用,所以从这个意义来说,死后肌肉pH值的下降,对肉的加工质量有十分重要的意义。

26
宰后极限pH值的影响因素 与宰前状况有关 如果在屠宰前剧烈运动,或注射肾上腺素,那么这时的糖原含量就会减少。如果这时候屠宰,死后继续消耗糖原,肉就会产生高极限pH值。 与牲畜的种类、不同的部位及个体的差异等内在因素有关, 受屠宰前是否注射药物、环境的温度等外界因素影响。环境温度越高,pH值变化越快。

27
二、死后僵直的机制 ※ATP减少 动物死之后,呼吸停止了,供给肌肉的氧气也就中断了,此时其糖原不再像有氧时最终氧化成CO2和H2O,而是在缺氧情况下经糖酵解产生乳酸。在正常有氧条件下,每个葡萄糖单位可氧化生成39个分子ATP,而经过糖酵解只能生成3分子ATP,ATP的供应受阻。然而体内ATP的消耗,由于肌浆中ATP酶的作用却在继续进行,因此动物死后,ATP的含量迅速下降。

28
※ 肌质网破裂 ATP的减少及pH值的下降,使肌质网功能失常,发生崩解,肌质网失去钙泵的作用,内部保存的钙离子被放出,致使Ca2+浓度增高,促使粗丝中的肌球蛋白ATP酶活化,更加快了ATP的减少,结果肌动蛋白和肌球蛋白结合形成肌动球蛋白,引起肌肉收缩表现出肉尸僵硬。 ※反应不可逆 这种情况下由于ATP不断减少,所以反应是不可逆的,则引起永久性的收缩。

29
三、死后僵直的过程 ◆急速期:随着弹性的迅速消失出现僵硬阶段叫急速期
动物死后僵直的过程分为三个阶段: ◆迟滞期:从屠宰后到开始出现僵直现象为止,即肌肉的弹性以非常缓慢的速度进展阶段,称为迟滞期; ◆急速期:随着弹性的迅速消失出现僵硬阶段叫急速期 ◆僵硬后期:最后形成延伸性非常小的一定状态而停止叫僵硬后期。到最后阶段肌肉的硬度可增加到原来的10~40倍,并保持较长时间。

30
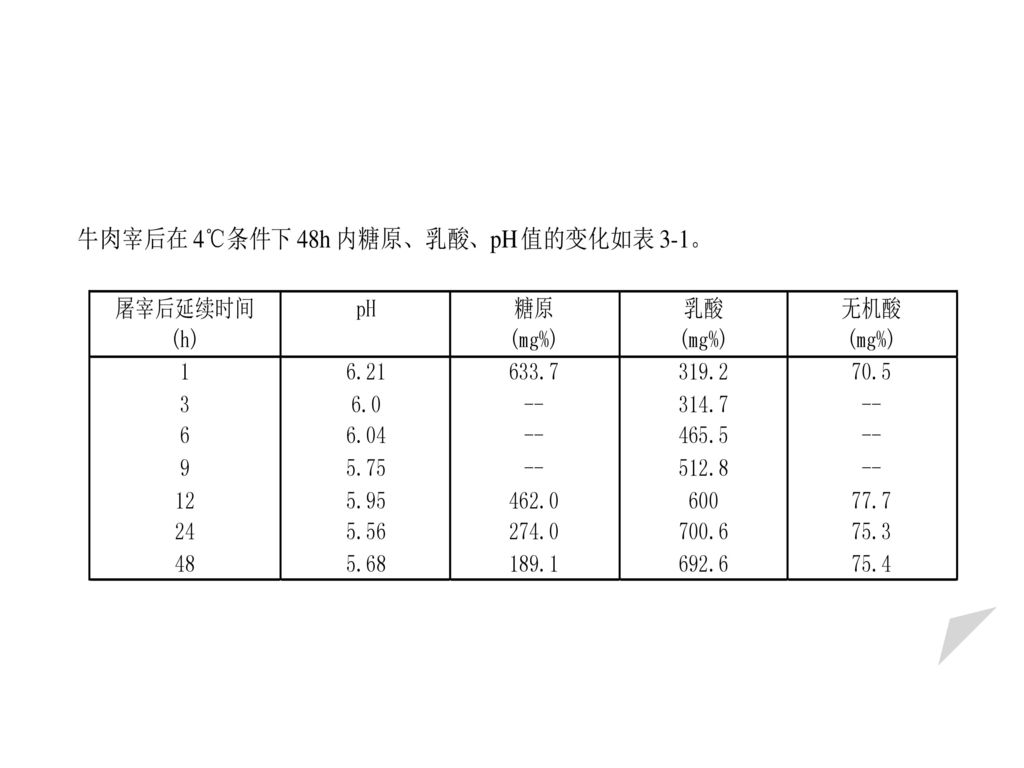
图3─7 死后僵直期肌肉物理和化学的变化 (牛肉37℃下)
图3─7 死后僵直期肌肉物理和化学的变化 (牛肉37℃下)
")
31
肌肉死后僵直过程与肌肉中的ATP下降速度有着密切的关系。在迟滞时期,肌肉中ATP的含量几乎恒定,这是由于肌肉中还存在另一种高能磷酸化合物──磷酸肌酸(CP),在磷酸激酶的作用下,由ADP再合成ATP,而磷酸肌酸变成肌酸: ADP + CP = 肌酸 + ATP 在此时期,细丝还能在粗丝中滑动,肌肉比较柔软,这一时期与ATP的贮量及磷酸肌酸的贮量有关。 随着磷酸肌酸的消耗殆尽,使ATP的形成主要依赖糖酵解,使ATP迅速下降而进入急速期。当ATP降低至原含量的15%~20%时,肉的延伸性消失而进入僵直后期。

32
动物屠宰之后磷酸肌酸与pH值迅速下降,而ATP在磷酸肌酸降到一定水平之前尚维持相对的恒定,此时肌肉的延伸性几乎没有变化,只有当磷酸肌酸下降到一定程度时,ATP开始下降,并以很快的速度进行,由于ATP的迅速下降,肉的延伸性也迅速消失,迅速出现僵直现象。 因此处于饥饿状态下或注入胰岛素情况下屠宰的动物肉,肌肉中糖原的贮备少,ATP的生成量则更少,这样在短时间内就会出现僵直,即僵直的迟滞期短。

33
图3-8 僵直时肌肉柔软性的变化曲线

34
从图中的五条曲线可以看出,屠宰时动物的生理状态不同,则第一阶段迟滞期长短不同,从1. 5h(第Ⅴ条曲线)到9h(第Ⅰ条曲线),而急速期从0
从图中的五条曲线可以看出,屠宰时动物的生理状态不同,则第一阶段迟滞期长短不同,从1.5h(第Ⅴ条曲线)到9h(第Ⅰ条曲线),而急速期从0.5h(第Ⅲ条曲线)到2h(第Ⅳ条曲线)。
到9h(第Ⅰ条曲线),而急速期从0.5h(第Ⅲ条曲线)到2h(第Ⅳ条曲线)。")
35
图3-9僵直过程中肌肉弹性系数的变化曲线

36
通过上述现象可以证明,引起死后僵直过程的变化,与肌肉中ATP的消失有直接的关系。随着ATP的消失,肌肉的肌球蛋白与肌动蛋白立即结合,生成肌动球蛋白,因而失去弹性。所以最初阶段迟滞期的长短是由ATP含量决定的。

37
四、冷收缩(cold shortening) 解冻僵直收缩(thaw shortening)
肌肉宰后有三种短缩或收缩形式,即热收缩(heat shortening)、冷收缩和解冻僵直收缩。 热收缩是指一般的尸僵过程,缩短程度和温度有很大关系,这种收缩是在尸僵后期,当ATP含量显著减少以后会发生,在接近零度时收缩的长度为开始长度的5%,到40℃时,收缩为开始的50%。
、冷收缩和解冻僵直收缩。 热收缩是指一般的尸僵过程,缩短程度和温度有很大关系,这种收缩是在尸僵后期,当ATP含量显著减少以后会发生,在接近零度时收缩的长度为开始长度的5%,到40℃时,收缩为开始的50%。")
38
4-1、冷收缩(cold shortening)
")
39
由冷收缩可知,死后肌肉的收缩速度未必温度越高,收缩越快,牛、羊、鸡在低温条件下也可产生急剧收缩,该现象红肌肉比白肌肉出现得更多一些,尤以牛肉明显。
从刚屠宰后的牛屠体上切下一块牛头肌肉片,立刻分别在1~37℃的温度中放置,结果表明:在1℃中贮藏的肉收缩最快、最急剧。在15℃中贮藏的肉收缩得最慢,而且也最小。

40
温度越高,ATP的消耗越大。低温收缩与ATP减少产生的僵直收缩是不一样的。最近一项结果表明,冷收缩不是由肌质网的作用产生,而是由线粒体释放出来的Ca2+产生的,含有大量线粒体的红色肌肉,在死后厌氧的低温条件下放置,线粒体机能下降而释放出钙,Ca2+再被在低温条件下机能下降的肌质网回收而引起收缩。

41
防止冷收缩的方法 为了防止冷收缩带来的不良效果,采用电刺激的方法,使肌肉中ATP迅速消失,pH值迅速下降,使尸僵迅速完成,即可改善肉的质量和外观色泽。 去骨的肌肉易发生冷收缩,硬度较大,带骨肉则可在一定程度上抑制冷收缩,所以目前普遍使用的屠体直接成熟是不太会出现冷收缩的。对于猪胴体,一般不会发生冷收缩。

42
4-2、解冻僵直收缩 肌肉在僵直未完成前进行冻结,仍含有较高的ATP,在解冻时由于ATP发生强烈而迅速的分解而产生的僵直现象,称为解冻僵直。解冻时肌肉产生强烈的收缩,收缩的强度较正常的僵直剧烈的多,并有大量的肉汁流出。

43
解冻僵直发生的收缩严重有力,可缩短50%,这种收缩可破坏肌肉纤维的微结构,而且沿肌纤维方向收缩不够均一。
在尸僵发生的任何一点进行冷冻,解冻时都会发生解冻僵直,但随肌肉中ATP浓度的下降,肌肉收缩力也下降。在刚屠宰后立刻冷冻,然后解冻时,这种现象最明显。 因此要在形成最大僵直之后再进行冷冻,以避免这种现象的发生。

44
五、尸僵和保水性的关系 尸僵阶段除肉的硬度增加外,肉的保水性减少,在最大尸僵期时最低。肉中的水分最初时渗出到肉的表面,呈现湿润状态,并有水滴流下。 刚宰后的肉保水性好,几小时以后保水性降低,到48~72h(最大尸僵期)肉的保水性最低。宰后24h有45%的肉汁游离。
肉的保水性最低。宰后24h有45%的肉汁游离。")
45
尸僵降低保水性的原因 pH降低:屠宰后的肌肉,随着糖酵解作用的进行,肉的pH值下降至极限值5.4~5.5,此pH值正是肌原纤维多数蛋白质的等电点附近。 由于ATP的消失和肌动球蛋白形成,肌球蛋白纤丝和肌动蛋白纤丝之间的间隙减少了,故而肉的保水性大为降低。 蛋白质某种程度的变性,肌浆中的蛋白质在高温低pH值作用下沉淀变性,不仅失去了本身的保水性,而且由于沉淀到肌原纤维蛋白质上,也进一步影响到肌原纤维的保水性。

46
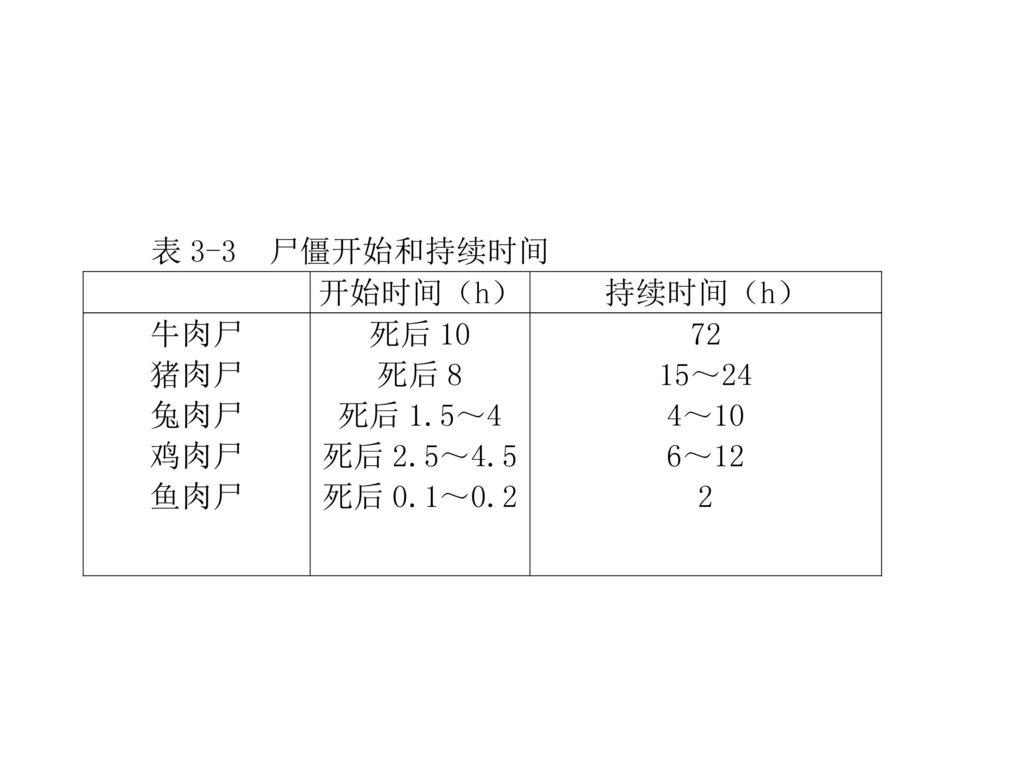
六、尸僵开始和持续时间 因动物的种类、品种、宰前状况,宰后肉的变化及不同部位而异。一般鱼类肉尸发生早,哺乳类动物发生较晚,不放血致死较放血致死发生的早,温度高发生的早持续时间短,温度低则发生的晚,持续时间长。 表3─3为不同动物尸僵时间。肉在达到最大尸僵时以后,即开始解僵软化进入成熟阶段。

48
一、肉成熟的条件及机制 二、成熟肉的物理变化 三、成熟肉的化学变化 四、促进肉成熟的方法 第三节 肉的成熟

49
肉的成熟定义 尸僵持续一定时间后,即开始缓解,肉的硬度降低,保水性有所恢复,使肉变得柔嫩多汁,具有良好的风味,最适于加工食用,这个变化过程即为肉的成熟。肉的成熟包括尸僵的解除及在组织蛋白酶作用下进一步成熟的过程。

50
一、肉成熟的条件及机制 (一)死后僵直的解除 (二)组织蛋白酶的作用 尸僵时肉的僵硬是肌纤维收缩的结果,可以认为
成熟时又恢复伸长而变为柔软。肌肉死后僵直达 到顶点之后,并保持一定时间,其后又逐渐变软, 解除僵直状态。 解除僵直所需时间由动物的种类、 肌肉的部位以及其它外界条件不同而异。在2~ 4℃条件贮存的肉类,对鸡肉需3~4h达到僵直的 顶点,而解除僵直需2d,其它牲畜完成僵直约需 1~2d,而解除僵直猪、马肉需3~5d,牛约需1周 到10d左右。

51
解僵实 质的几 种假说 肌原 纤维 小片化 死后肌肉 中肌动蛋 白和肌球 蛋白纤维 之间结合 变弱 肌肉中结构弹性 网状蛋白的变化 蛋白酶说

52
1.肌原纤维小片化 刚宰后的肌原纤维和活体肌肉一样,是由数十到数百个肌节沿长轴方向构成的纤维,而在肉成熟时则断裂成1~4个肌节相连的小片状。这种肌原纤维断裂现象被认为是肌肉软化的直接原因。这时在相邻肌节的Z线变得脆弱,受外界机械冲击很容易断裂。

53
产生小片化的原因 (1) 死后僵直肌原纤维产生收缩的张力,使Z线在持续的张力作用下发生断裂,张力的作用越大,小片化的程度越大。
(2) Ca2+作用引起的。死后肌质网功能破坏,Ca2+从网内释放,使肌浆中的Ca2+ 浓度增高,刚宰后肌浆中Ca2+,浓度为1×10-6Mol,成熟时为1×10-4M,比原来增高100倍。高浓度的Ca2+长时间作用于Z线,使Z线蛋白变性而脆弱给予物理力的冲击和牵引而发生断裂。
 Ca2+作用引起的。死后肌质网功能破坏,Ca2+从网内释放,使肌浆中的Ca2+ 浓度增高,刚宰后肌浆中Ca2+,浓度为1×10-6Mol,成熟时为1×10-4M,比原来增高100倍。高浓度的Ca2+长时间作用于Z线,使Z线蛋白变性而脆弱给予物理力的冲击和牵引而发生断裂。")
54
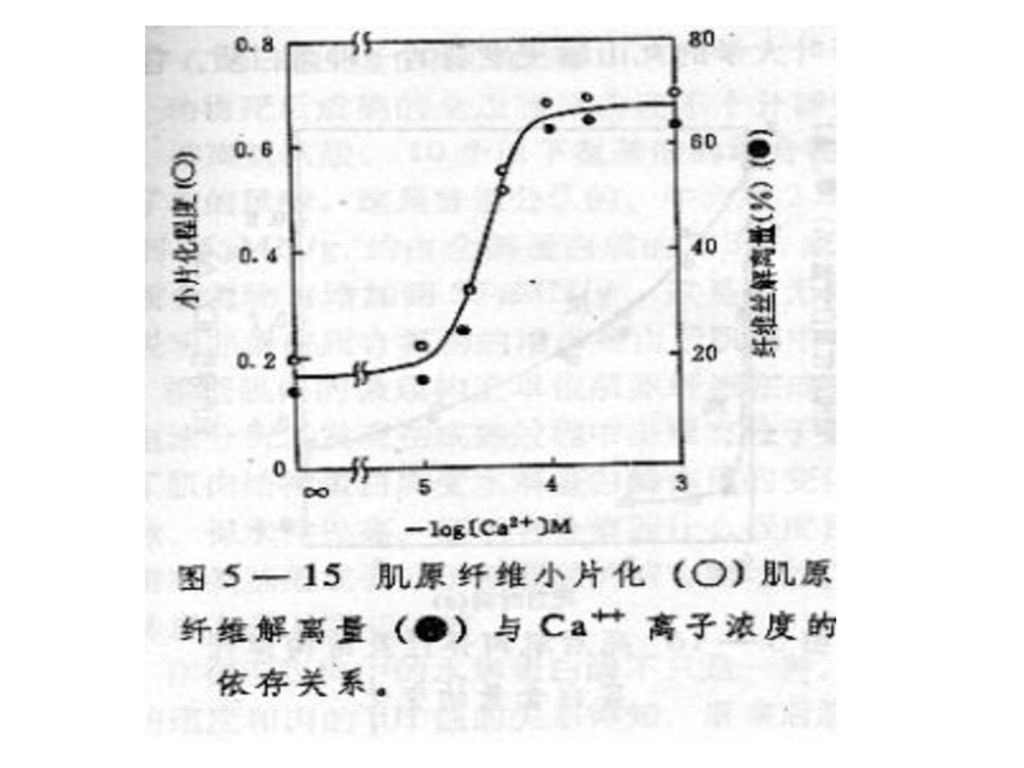
肌原纤维小片化与Ca2+ 浓度的关系如图5-12。从图中看出当Ca2+ 浓度在1×10-5Mol以下时,对小片化无显著影响,而当超过1×10-5 Mol数量时,肌原纤维小片化程度忽然增加,Ca2+浓度达到1×10-4Mol时达到最大值。

56
2.死后肌肉中肌动蛋白和肌球蛋白纤维之间结合变弱
随着保藏时间的延长,肌原纤维的分解量逐渐增加,如家兔肌肉在10℃条件下保藏2d的肌原纤维只分解5%,而到6d时近50%的肌原纤维被分离,当加入ATP时分解量更大。肌原纤维分离的原因,恰与肌原纤维小片化是一致的。小片化是从肌原纤维的Z线处崩解,正表明肌球蛋白和肌动蛋白之间的结合减弱了。

57
3.肌肉中结构弹性网状 蛋白的变化 结构弹性网状蛋白是肌原纤维中除去粗丝、细丝及Z线等蛋白质后,不溶性的并具有较高弹性的蛋白质。贯穿于肌原纤维的整个长度,连续的构成网状结构。 随着保藏时间的延长和弹性的消失而减少,结构弹性蛋白的含量也达到最低值。肉类在成熟软化时结构弹性蛋白质的消失,导致肌肉弹性的消失。

58
钙激活酶与肉的成熟 肌肉成熟变化的反应是在多种酶的协同作用下完成的.其中一个起主要作用的酶就是钙激活酶,因为只有它才能启动肌原纤维蛋白的降解,破坏Z线,释放肌纤丝,从而引起其它蛋白酶的作用,促进肌纤维的降解

59
钙激活酶与肌肉成熟的关系 使肌原纤维I带和Z盘结合变弱或断裂 使连接蛋白Costameres的降解

60
二、成熟肉的物理变化 pH值的变化 保水性的变化 嫩度的变化 风味的变化

61
pH值的变化 肉在成熟过程中pH值发生显著的变化。刚屠宰后肉的pH值在6~7之间,约经1h开始下降,尸僵时达到最低5.4~5.6之间,而后随保藏时间的延长开始慢慢地上升。

62
保水性的变化 (1)保水性的回升和pH值变化有关,随着解僵,pH值逐渐增高,偏离了等电点,蛋白质静电荷增加,使结构疏松,因而肉的持水性增高。
肉在成熟时保水性又有回升。 (1)保水性的回升和pH值变化有关,随着解僵,pH值逐渐增高,偏离了等电点,蛋白质静电荷增加,使结构疏松,因而肉的持水性增高。 (2)此外随着成熟的进行,蛋白质分解成较小的单位,从而引起肌肉纤维渗透压增高。 保水性恢复只能部分恢复,不可能恢复到原来状态,因肌纤维蛋白结构在成熟时发生了变化。
保水性的回升和pH值变化有关,随着解僵,pH值逐渐增高,偏离了等电点,蛋白质静电荷增加,使结构疏松,因而肉的持水性增高。 (2)此外随着成熟的进行,蛋白质分解成较小的单位,从而引起肌肉纤维渗透压增高。 保水性恢复只能部分恢复,不可能恢复到原来状态,因肌纤维蛋白结构在成熟时发生了变化。")
63
嫩度的变化 随着肉成熟的发展,肉的柔软性产生显著的变化。刚屠宰之后生肉的柔软性最好,而在2昼夜之后达到最低的程度。如热鲜肉的柔软性平均值为74%,保藏六昼夜之后又重新增加,平均可达鲜肉时的83%。

64
风味的变化 肉在成熟过程中由于蛋白质受组织蛋白酶的作用,游离的氨基酸含量有所增加,主要表现在浸出物质中。新鲜肉中酪氨酸和苯丙氨酸等很少,而成熟后的浸出物中有酪氨酸、苯丙氨酸、苏氨酸、色氨酸等存在,其中最多的是谷氨酸、精氨酸、亮氨酸、缬氨酸、甘氨酸,这些氨基酸都具有增强肉的滋味和香气的作用,所以成熟后的肉类,肉的风味提高,与这些氨基酸成分有一定的关系。此外,肉在成熟过程中,ATP分解产生次黄嘌呤核苷酸(IMP),它为味质增强剂。
,它为味质增强剂。")
65
三、成熟肉的化学变化 (一)蛋白水解 (二)次黄嘌呤核苷酸(IMP)的形成 (三)肌浆蛋白溶解性的变化
(四)构成肌浆蛋白的N─端基的数量增加 (五)金属离子的增减
构成肌浆蛋白的N─端基的数量增加. (五)金属离子的增减.")
66
(一)蛋白水解 在成熟中,水溶性非蛋白质态含氮化合物会增加。
如果将处于极限pH值5.5~5.8的家兔的背最长肌,在37℃条件下无菌贮藏5~6个月,那么三氯乙酸可溶性氮,即非蛋白态氮就会从初期总氮的10%~30%上升到37%(家兔)和31%(牛)。开始贮藏后的10天增长速度很快。在低温条件下贮藏,其增长速度较慢。牛背最长肌在2℃条件下贮藏30天其非蛋白态氮增长到45μmol/g肉,家兔背最长肌在3~4℃条件下贮藏7天,增长到55μmol/g肉。
和31%(牛)。开始贮藏后的10天增长速度很快。在低温条件下贮藏,其增长速度较慢。牛背最长肌在2℃条件下贮藏30天其非蛋白态氮增长到45μmol/g肉,家兔背最长肌在3~4℃条件下贮藏7天,增长到55μmol/g肉。")
67
(二)次黄嘌呤核苷酸(IMP)的形成 死后肌肉中的ATP在肌浆中ATP酶作用下迅速转变为ADP,而ADP又在肌激活酶的作用下进一步水解为AMP,再由脱氨酶的作用下形成AMP,反应过程如下:
次黄嘌呤核苷酸(IMP)的形成 死后肌肉中的ATP在肌浆中ATP酶作用下迅速转变为ADP,而ADP又在肌激活酶的作用下进一步水解为AMP,再由脱氨酶的作用下形成AMP,反应过程如下:")
69
根据分析结果表明,僵直前的肌肉中ADP很少,而ATP的含量较多;但僵直以后IMP的数量特多,其中肌苷、次黄嘌呤、次黄苷、ADP、ATP、IDP(次黄苷二磷酸)、ITP(次黄苷三磷酸)等较少。
、ITP(次黄苷三磷酸)等较少。")
70
(三)肌浆蛋白溶解性的变化 屠宰后接近24h肌浆蛋白溶解度降到最低程度。表3─6是在4℃条件下成熟过程随时间的延续,肌浆蛋白溶解性的变化,从表中数字看出,刚屠宰之后的热鲜肉,转入到浸出物中的肌浆蛋白最多,6h以后肌浆蛋白的溶解性就显著减少而呈不溶状态,直到第一昼夜终了,达到最低限度,只是最初热鲜肉的19%。到第四昼夜可增加到开始数量36%,相当第一昼夜的2倍,以后仍然继续增加。对盐的溶解性也是新鲜肉最高,经1~2昼夜溶解性开始降低,为鲜肉的65%,以后又继续增加,当达到四昼夜时达73%。

72
(四)构成肌浆蛋白的 N─端基的数量增加 随着肉成熟的发现,蛋白质结构发生变化,使肌浆蛋白质氨基酸和肽链的N─端基(氨基)的数量增多。而相应的氨基酸如二羧酸、谷氨酸、甘氨酸、亮氨酸等都增加,显然伴随着肉成熟进行,构成肌浆蛋白质的肽链被打开,形成游离N─端基的增多。所以成熟后的肉类,柔软性增加,水化程度增加,热加工时保水能力增高,这些都与N─端基的增多有一定的关系。
的数量增多。而相应的氨基酸如二羧酸、谷氨酸、甘氨酸、亮氨酸等都增加,显然伴随着肉成熟进行,构成肌浆蛋白质的肽链被打开,形成游离N─端基的增多。所以成熟后的肉类,柔软性增加,水化程度增加,热加工时保水能力增高,这些都与N─端基的增多有一定的关系。")
73
(五)金属离子的增减 在成熟中可看到水提取的金属离子增减情况。Na和Ca增加,K减少。在活体肌肉中,Na和K大部分以游离形态存在于细胞内,一部分与蛋白质等结合。Mg几乎全部处于游离状态,ATP变成Mg-ATP,成为肌球蛋白的基质。Ca基本不以游离的形态存在,而与肌质网、线粒体、肌动蛋白等结合。增加的钙可能是某种变化游离出来的。

74
四、促进肉成熟的方法 (一)物理因素的控制 (二)化学因素 (三)生物学因素
物理因素的控制 (二)化学因素 (三)生物学因素")
75
(一)物理因素的控制 1.温度 2.电刺激可加快死后僵直的发展 3.力学因素
物理因素的控制 1.温度 2.电刺激可加快死后僵直的发展 3.力学因素")
76
1.温度 温度高成熟的快,Wilson等试验45.5rad γ射线照射牛肉,结合防腐进行高温成熟,43℃24h即完成,它和低温1.7℃成熟14d获得嫩度效果相同,缩短时间10多倍,但这样的肉颜色、风味都不好。 高温和低pH值环境下,不易形成僵直肌动球蛋白。中温成熟时,尸僵硬度是在中温域引起,此时肌肉缩短度小,因而成熟的时间短。为了防止尸僵时短缩,可把不剔骨肉在中温域进入尸僵。

77
2.电刺激可加快死后僵直的发展 电刺激主要用于牛、羊肉中,这个方法可以防止冷收缩。
所谓电刺激是家畜屠宰放血后,在一定的电压电流下,对胴体进行通电,从而达到改善肉质的目的。 电刺激使用的电压变化非常显著,有高电压和低电压之分。高电压可以达到300V以上,甚至600V以上,低电压则为100V以下。

78
屠宰后的机体用电流刺激可以加快生化反应过程和pH值的下降速度,促进尸僵过程的进行。对像牛肉和羊肉含有较多红色肌肉的家畜肉,在冷却的时候,随着肉的温度下降ATP没有完全消失,因而肌浆网内摄取Ca2+的能力降低了,同时Ca2+也从线粒体被游离到肌浆中,使肌浆中的Ca2+ 浓度急剧地增加,这样使肌肉发生强烈收缩,而电刺激可以预防这种现象。

79
电刺激不单可以促进ATP的消失和pH值下降,而且对促进肉质色泽鲜明、肉质软化有明显作用;特别是经过电刺激加工的热鲜肉,易于施行热剔骨,因此可以节省30%~50%的冷却能量,节省70%~80%冷库容积,对提高肉类加工企业的经济效益有很大意义。

80
屠宰后的机体用电流刺激可以加快生化反应过程和pH值的下降速度,促进尸僵过程的进行。
近20年来人们对电刺激的应用不仅是为了改善肉的质量,而且也是为了解决现代化肉类冷加工中出现的实际问题。 生产实践中,要求屠宰后的牛羊胴体在24h以内,肉体中心温度降低到7℃以下,必须以相当快的速度冷却,然而快速冷却易产生肌肉寒冷收缩现象,损害了肉的嫩度。 实际证明,当死后肌肉的pH值迅速降低到6以下,就可以防止产生冷收缩。而电刺激可以加快肌肉中的生物化学反应,迅速生成乳酸,使pH值下降。

81
如200V电压,频率为12.5HZ,刺激时间为2min,使pH值迅速降低,最初pH值越高,下降的幅度越大,刺激后的肌肉在35℃条件经3~4h降到6.2以下。

82
从上述数字可以说明,最初pH值越高,下降幅度越大,而且刺激的敏感性随死后僵直时间的延长而减弱,反之则相反。根据新西兰资料,对羊胴体,屠宰后30min内进行刺激,牛不超过45min。

83
在工业上使用电刺激方法首先是在新西兰。新西兰牛羊生产量很大,在生产旺季,一些屠宰厂日宰量达1万头,因此要在屠宰流水线后对胴体进行快速冷却,使用通常方法,很容易产生冷收缩,这就使新西兰的研究者们最先着手解决使用电刺激所存在的问题,以保证肉的质量。 电刺激可改善肉的嫩度,防止寒冷收缩。根据法国的资料介绍,刺激后的牛肉可提高嫩度15%~16%。刺激后的肉类在肉温降低到10℃以下之前,pH值可降低到6.0以下,防止寒冷收缩,为实现热剔骨创造了前提条件。

84
电刺激促进肌肉嫩化的机理: ①电刺激加快尸僵过程,减少了冷收缩,这一点是由于电刺激加快了肌肉中ATP的降解,促进糖原分解速度,使胴体pH值很快下降到6以下,这时再对牛、羊肉进行冷加工,就可防止冷收缩,提高肉的嫩度; ②电刺激激发强烈的收缩,使肌原纤维断裂,肌原纤维间的结构松弛,可以容纳更多的水分,使肉的嫩度增加; ③电刺激使肉的pH值下降,还会促进酸性蛋白酶的活性,蛋白酶分解蛋白质,大分子分解为小分子,使嫩度增加。

85
3.力学因素 尸僵时带骨肌肉收缩,这时以相反的方向牵引,可使僵硬复合体形成最少。通常成熟时,将跟腱用钩挂起,此时主要是腰大肌受牵引。如果将臂部用钩挂起,不但腰大肌短缩被抑制,而半腱肌、半膜肌、背最长肌均受到拉伸作用,可以得到较好的嫩度。

86
(二)化学因素 (1)屠宰前注射肾上腺激素、胰岛素等,使动物在活体时加快糖的代谢过程,肌肉中糖原大部分被消耗或从血液中排出。宰后肌肉中糖原和乳酸含量极少,肉的pH值较高,在6.4~6.9的水平,肉始终保持柔软状态。
化学因素 (1)屠宰前注射肾上腺激素、胰岛素等,使动物在活体时加快糖的代谢过程,肌肉中糖原大部分被消耗或从血液中排出。宰后肌肉中糖原和乳酸含量极少,肉的pH值较高,在6.4~6.9的水平,肉始终保持柔软状态。")
87
(2)注入Ca2+可以促进软化 Ca2+可以激活钙激活酶(Calpain),钙激活酶为巯基内切酶,有两种活性形式,一种可被μmol水平的Ca2+激活称μ一Calpain,另一种可被mmol水平的Ca2+激活,称m一calpain,另外在肉中还存在一种Calpain抑制蛋白Calpastatin。
注入Ca2+可以促进软化 Ca2+可以激活钙激活酶(Calpain),钙激活酶为巯基内切酶,有两种活性形式,一种可被μmol水平的Ca2+激活称μ一Calpain,另一种可被mmol水平的Ca2+激活,称m一calpain,另外在肉中还存在一种Calpain抑制蛋白Calpastatin。")
88
钙激活酶激活需要细胞内有足够的钙离子(浓度达mM),在胴体进入僵直阶段后,由于ATP的减少,肌浆网体失去钙泵作用而破裂,钙离子释放,细胞内游离钙离子浓度增加,最高能达0.lmM.但这个浓度不足以使钙激活酶全部表现活性,因此可以从外源增加细胞内钙离子浓度,以激活钙激活酶,从而促进肉的嫩化。
,在胴体进入僵直阶段后,由于ATP的减少,肌浆网体失去钙泵作用而破裂,钙离子释放,细胞内游离钙离子浓度增加,最高能达0.lmM.但这个浓度不足以使钙激活酶全部表现活性,因此可以从外源增加细胞内钙离子浓度,以激活钙激活酶,从而促进肉的嫩化。")
89
从表中可以看出六偏磷酸钠(Ca 螯合剂)、柠檬酸钠(糖解阻抑剂)、氯化镁(肌动球蛋白形成阻抑剂)等,各自显出对尸僵硬度的抑制作用
、柠檬酸钠(糖解阻抑剂)、氯化镁(肌动球蛋白形成阻抑剂)等,各自显出对尸僵硬度的抑制作用")
90
(三)生物学因素 基于肉内蛋白酶活性可以促进肉质软化考虑,也有从外部添加蛋白酶强制其软化的可能。用微生物和植物酶,可使固有硬度和尸僵硬度都减少,常用的有木瓜酶。方法可以采用临屠宰前静泳注射或刚宰后肌肉注射,宰前注射能够避免脏器损伤和休克死亡。木瓜酶的作用最适温度≥50℃,低温时也有作用。为了预防羊肉的寒冷收缩,在每kg肉中注入30mg,在70℃加热后,可收到软化的效果。

91
第四节 肉的腐败变质 一、肉类腐败的原因和条件 二、肌肉组织的腐败 三、脂肪的氧化和酸败

92
肉的腐败变质定义 肉的腐败变质--指肉在组织酶和微生物作用下发生质的变化,最终失去食用价值。
肉的自家溶解--肉在自溶酶作用下的蛋白质分解过程。 肉的腐败--由微生物作用引起的蛋白质分解过程。 酸败--肉中脂肪的分解过程。 如果说肉的成熟的变化主要是糖酵解过程(也有核蛋白的分解,脂肪不分解),那么肉变质时的变化主要是蛋白质和脂肪分解过程。 污染源---毛皮、土地、粪便、空气、水、工具、包装容器、操作工人等。
,那么肉变质时的变化主要是蛋白质和脂肪分解过程。 污染源---毛皮、土地、粪便、空气、水、工具、包装容器、操作工人等。")
93
一、肉类腐败的原因和条件 肉类腐败是成熟过程的加深,动物死后由于血液循环的停止,吞噬细菌的作用停止了,这就使得细菌有可能繁殖和传播。但在正常条件下屠宰的肉类,肌肉中含有相当数量的糖原,死后由于糖原的酵解,形成乳酸,使肌肉的pH值从最初的7.0左右,下降到5.4~5.6,对腐败细菌的繁殖生长是极为不利的条件,起抑制腐败作用。

94
健康动物血液和肌肉通常是无菌的,肉类的腐败,实际上主要是由于在屠宰、加工、流通等过程受外界微生物的感染所致。由于微生物作用的结果,不仅改变了肉的感官性质、颜色、弹性、气味等,使肉的品质发生严重的恶化,而且破坏了肉的营养价值,或由于微生物生命活动代谢产物形成有毒物质,因此这一条件下腐败的肉类,能引起人们的食物中毒。

95
肉类的腐败,通常由外界环境中好气性微生物污染肉表面开始,然后又沿着结缔组织向深层扩散,特别是临近关节、骨骼和血管地方,最容易腐败。并且由生物分泌的胶原蛋白酶使结缔组织的胶原蛋白水解形成粘液,同时产生气体,分解成氨基酸、水、二氧化碳、氨气;有糖原存在下发酵,形成醋酸和乳酸。因此形成恶臭的气味。

96
微生物对脂肪进行两种酶促反应,一是由于分泌的脂肪酶分解脂肪,产生游离脂肪酸和甘油;另一种是氧化酶通过β─氧化作用,氧化脂肪,产生氧化的酸败气味。但肉类及其制品发生严重的腐败并不单纯是由于微生物所引起,而是由空气中氧,在光线、温度以及金属离子的共同作用的结果。

97
新鲜肉发生腐败的外观特征主要表现为色泽、气味的恶化和表面发粘。
表面发粘是微生物作用产生腐败的主要标志。在流通中,当肉表面的细菌达107个/cm2,就有粘液出现,并有不良的气味。达到这种状态所需的日数与最初污染细菌的个数有关,污染的细菌数越多,则腐败越快。也受环境的温度和湿度影响,温度越高,湿度越大,越易产生发粘的现象。

98
温度是决定微生物生长繁殖的重要因素,温度越高繁殖发育越快。
水分是仅次于温度决定肉食品上微生物繁殖的重要因素,一般霉菌和酵母比细菌耐受较高的渗透压。 pH值对细菌的繁殖极为重要,所以肉的最终pH值对防止肉的腐败具有十分重要意义。 多数细菌在pH值7左右最适于繁殖,在pH值4以下、9以上繁殖就困难。

99
二、肌肉组织的腐败 肌肉组织的腐败就是蛋白质受微生物作用的分解过程。天然蛋白质通常不能被微生物所同化,这是因为天然蛋白质是高分子的胶体粒子,它不能通过细胞膜而扩散,因此大多数微生物都是在蛋白质分解产物上才能迅速发展,所以肉成熟或自溶为微生物的繁殖准备了条件。

101
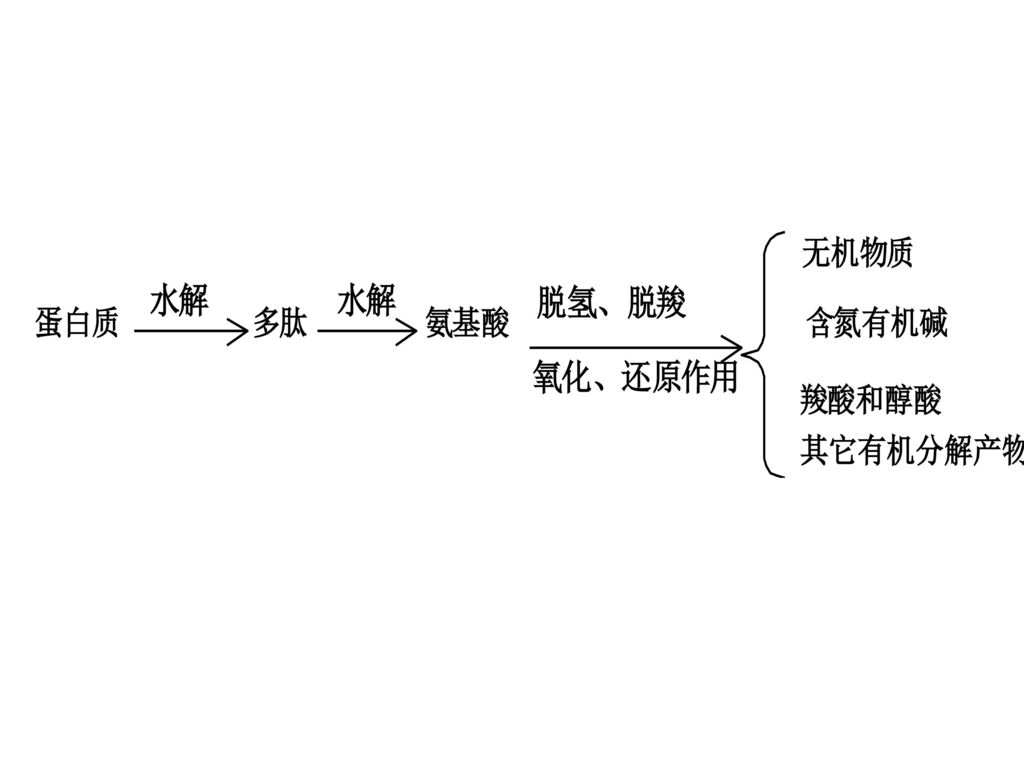
微生物对蛋白质的腐败分解,通常是先形成蛋白质的水解初步产物─多肽,再水解成氨基酸。有时也可直接由蛋白质分子分离出来。多肽与水形成粘液,附在肉的表面。它与蛋白质不同,能溶于水,煮制时转入肉汤中,使肉汤变得粘稠混浊,利用这点可鉴定肉的新鲜程度。 蛋白质腐败分解形成的氨基酸,在微生物分泌酶的作用下,发生复杂的生物化学变化,产生多种物质:有机酸、有机碱、醇及其它各种有机物质,分解的最终产物为CO2、H2O、NH3、H2S、P等。

102
三、脂肪的氧化和酸败 脂肪 酸败 水解 光、热、氧、催化剂 光、水、催化剂 酶、微生物 酶、微生物 过氧化物 高 甘 二 一
酸败 水解 光、热、氧、催化剂 光、水、催化剂 酶、微生物 酶、微生物 过氧化物 高 甘 二 一 醇 聚 缩 低 醛 酮 气 级 油 酸 酸 酸 合 合 级 类 类 体 脂 甘 甘 物 物 脂 产 肪 油 油 肪 体 酸 脂 脂

103
屠宰后,肉在贮藏中,最易变化的成分之一为脂肪。此变化最初为脂肪组织本身所含酶的作用,其次为细菌产生酶的酸败。另一方面因空气中氧的作用,而发生氧化作用。前者属于加水分解(Hydrolysis),后者称之氧化作用(Oxidation)。水解是由脂肪酶(Lipase)的作用。
,后者称之氧化作用(Oxidation)。水解是由脂肪酶(Lipase)的作用。")
104
能产生脂肪酶的细菌,可使脂肪分解为脂肪酸和甘油,一般说来,有强力分解蛋白能力的需 氧细菌的大多数菌种都能分解脂肪。细菌中具有分解脂肪特性的菌种不同。如假单孢菌属,其中解脂能力最强的是荧光假单孢菌。其它如:黄杆菌属、无色杆菌属、产碱杆菌属、赛氏杆菌属、小球菌属、葡萄球菌属、芽孢杆菌属中许多都具有分解脂肪的特性。 能分解脂肪的霉菌比细菌多,常见的霉菌有黄曲霉、黑曲霉、灰绿青霉等。

105
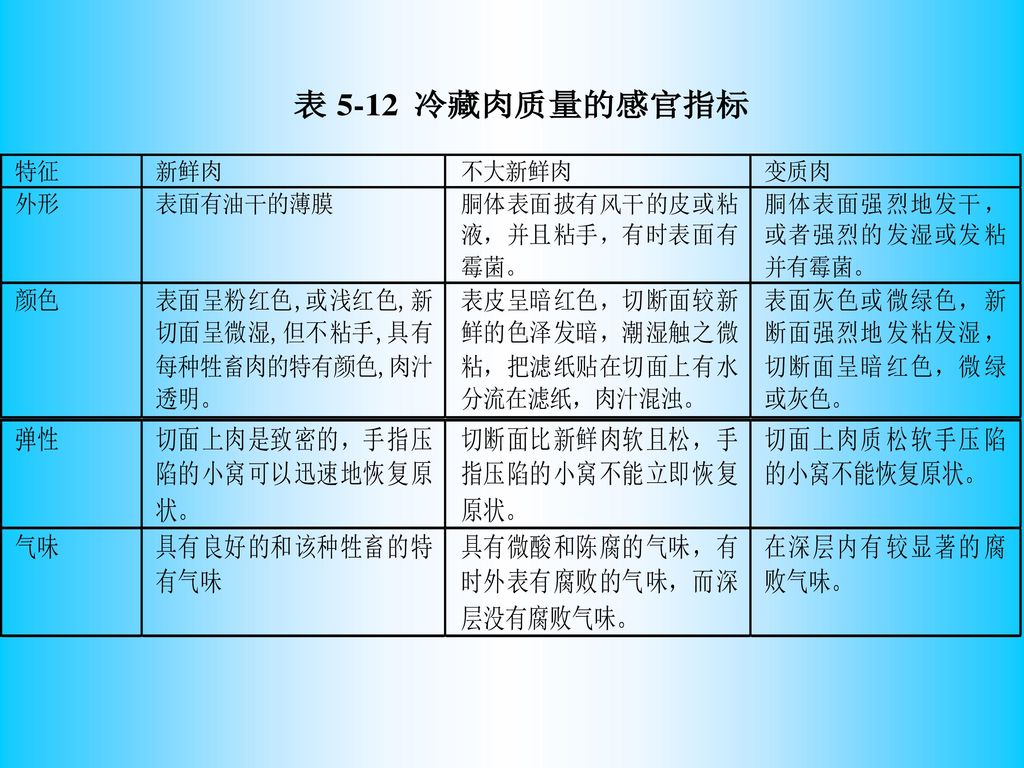
四、腐败肉的感官特征 感官是指人的视觉、嗅觉、触觉、及听觉的综合反应。 视觉─肉的组织状态,粗嫩、粘滑、干湿、色调、光泽等。
嗅觉─气味有无,强弱、香、臭、腥、膻等。 味觉─滋味的鲜美、香甜、苦涩、酸臭。 触觉─坚实、松弛、弹性、拉力等。 听觉─检查冻肉、罐头的声音清脆、混浊。









 导致胎儿缺氧的母体因素有 ①微小动脉供血不足:如妊高征等 ②红细胞携氧量不足:如重度贫血、一氧化碳中毒等;>")








的過濾作用 腎小管的再吸收作用 腎小管的分泌作用 尿液的組成>")






9:00~10:30 地 點:校史室 報告人:學務主任鐘勻鈴>")

