Download presentation
1
RNA的合成与加工 生物化学

2
第十三章 RNA 的代谢 §13.1 DNA指导下的RNA 的合成:转录 §13.2转录后RNA的加工
§13.4 RNA指导下的RNA的合成:RNA复制 §13.3 RNA指导下DNA的合成:逆转录 §13.2转录后RNA的加工

3
§13.1 DNA指导下的RNA 的合成:转录 本节主要内容 一、转录与DNA复制比较 六、RNA生物合成抑制剂 五、真核生物的转录过程 四、启动子与终止子 三、大肠杆菌的转录过程 二、DNA指导下的RNA聚合酶

4
§13.1 DNA指导下的RNA 的合成:转录 一、转录与DNA复制的比较 1、相同点:
转录(transcription):以DNA为模板,根据碱基配对的原则合成与DNA互补的RNA的过程。 一、转录与DNA复制的比较 1、相同点: ●合成方向都是5 ’ →3’ ●都以三磷酸核苷酸为底物(NTP或dNTP) ●都需要模板
:以DNA为模板,根据碱基配对的原则合成与DNA互补的RNA的过程。 一、转录与DNA复制的比较. 1、相同点: ●合成方向都是5 ’ →3’ ●都以三磷酸核苷酸为底物(NTP或dNTP) ●都需要模板.")
5
§13.1 DNA指导下的RNA 的合成:转录 转录不需引物; 2、转录与DNA复制的不同点
DNA全程复制,而转录只选择DNA的一个片断进行转 录;可连续独立转录成RNA的片断称为“转录单位” 或“操纵子”,可转录的基因称为“结构基因”。 选择哪个基因转录与特定的时间、空间和特定的生理状 态有关。 转录不需引物;

6
双链 DNA 中只有一条链具有转录活性(称为模板链);
模板链( watson链):双链DNA中具有转录活性的的链称 为模板链,又称反义链(或负链)。 编码链( crick链) :双链DNA中无转录活性的链称为编 码链,又称有义链(或正链)。 并不是所有的模板链都在一条链上;
:双链DNA中具有转录活性的的链称. 为模板链,又称反义链(或负链)。 编码链( crick链) :双链DNA中无转录活性的链称为编. 码链,又称有义链(或正链)。 并不是所有的模板链都在一条链上;")
7
§13.1 DNA指导下的RNA 的合成:转录 转录时双链局部解开,新生RNA链与模板链形成杂螺

8
二、DNA指导下RNA聚合酶(DDRP)
●四种核苷三磷酸(ATP、GTP、CTP、UTP)为底物; ●以DNA为模板,模板可以是单链DNA、双链DNA,但 双链DNA状态下活性最大; ●聚合酶无校对功能(即外切酶功能),转录的错误率为 1/104 ~ 1/105 ●聚合速度(E.coli)为 50 ~ 90 nt/min
为底物; ●以DNA为模板,模板可以是单链DNA、双链DNA,但. 双链DNA状态下活性最大; ●聚合酶无校对功能(即外切酶功能),转录的错误率为. 1/104 ~ 1/105. ●聚合速度(E.coli)为 50 ~ 90 nt/min.")
9
§13.1 DNA指导下的RNA 的合成 核心酶 全酶 2、大肠杆菌的RNA聚合酶 核心酶(β’βα2):催化链的延长
起始因子:σ :功能未知 核心酶 全酶

10
§13.1 DNA指导下的RNA 的合成:转录 大肠杆菌RNA聚合酶各亚基的功能 亚基 基因 Mr 亚基数目 功能 α rpoA 2
40,000 2 酶的装配,与启动子上 游元件和活化因子结合 β rpoB 155,000 1 结合核苷酸底物,催 化磷酸二酯键形成 β’ rpoC 160,000 与模板结合 σ rpoD 32,000 ~ 92,000 识别启动子,促进转录的起始 9,000 未知

11
§13.1 DNA指导下的RNA 的合成:转录 3、真核细胞的RNA聚合酶 同样,真核 RNA 聚合酶无校对功能。 - 酶类 分布 产物
α-鹅膏蕈碱 分子量 I 核仁 rRNA (5.8S、18S、28S) 对酶的作用 不抑制 ~ Ⅱ 核质 hnRNA snRNA 低浓度抑制 Ⅲ tRNA 5SrRNA 高浓度抑制 - 同样,真核 RNA 聚合酶无校对功能。
 对酶的作用. 不抑制 ~ Ⅱ. 核质. hnRNA snRNA. 低浓度抑制. Ⅲ. tRNA 5SrRNA. 高浓度抑制. - 同样,真核 RNA 聚合酶无校对功能。")
12
§13.1 DNA指导下的RNA 的合成:转录 三、大肠杆菌的转录过程 1、识别阶段:RNA聚合酶在σ亚基的引导下结合于启动子上;
6、RNA 和 RNA 聚合酶从 DNA 上脱落。

13
§13.1 DNA指导下的RNA 的合成:转录 RNA 的 合 成 过 程 起始 离开 双链DNA局部解开 磷酸二酯键形成 5
延长阶段 3 5 解链区到达基因终点 5 3 RNA 终止阶段 5 3

14
§13.1 DNA指导下的RNA 的合成:转录 转录泡 编码链 恢复螺旋 解螺旋 模板链

15
●利用足迹法(footprint)和DNA测序法可
四、启动子与终止子 1、启动子(promotor) ●指RNA聚合酶识别、结合和开始转录的 一段特定的DNA序列。 ●利用足迹法(footprint)和DNA测序法可 以确定启动子的序列结构.
 ●指RNA聚合酶识别、结合和开始转录的. 一段特定的DNA序列。 ●利用足迹法(footprint)和DNA测序法可. 以确定启动子的序列结构.")
16
足迹法 §13.1 DNA指导下的RNA 的合成:转录 将DNA起始转录的限制性片段分离; 用RNA聚合酶与之结合,再用DNA酶部分水解。
与酶结合的部位被保护而不被水解,其余部分被 水解成彼此只相差一个核苷酸的大小不等的片段 经凝胶电泳可测得酶结合部位的的序列结构。

17
足迹法示意图

18
§13.1 DNA指导下的RNA 的合成

19
§13.1 DNA指导下的RNA 的合成:转录 大肠杆菌RNA聚合酶的启动子

20
§13.1 DNA指导下的RNA 的合成 大肠杆菌不同的σ因子识别不同启动子

21
§13.1 DNA指导下的RNA 的合成:转录 图中所示:-35区是RNA聚合酶最先识别并结合的部位 ,形成所谓“闭合复合物”,然后向下游移动至富含AT的-10区域,这是转录时开始解链的部位,此时形成“开放复合物”。

22
不依赖因子 的终止子 富含GC 富含AT 依赖因子 不富含GC 不富含AT
§13.1 DNA指导下的RNA 的合成:转录 两类终止子的结构特点 终止子类型 回文结构序列 回文结构后序列 不依赖因子 的终止子 富含GC 富含AT 依赖因子 不富含GC 不富含AT

23
通读:RNA聚合酶越过终止子继续转录的现象称为通读,
§13.1 DNA指导下的RNA 的合成:转录 提供转录终止信号的DNA序列称为终止子。 2、终止子(terminator) 两类 不依赖ρ-因子的终止子 依赖ρ-因子的终止子 抗终止因子:引起抗终止作用的蛋白质称为抗终止因子。 通读:RNA聚合酶越过终止子继续转录的现象称为通读, 通读是由于终止子被特异性的因子抑制产生的。 终止因子:协助RNA聚合酶识别终止信号的辅助因子 或蛋白质。
 两类. 不依赖ρ-因子的终止子. 依赖ρ-因子的终止子. 抗终止因子:引起抗终止作用的蛋白质称为抗终止因子。 通读:RNA聚合酶越过终止子继续转录的现象称为通读, 通读是由于终止子被特异性的因子抑制产生的。 终止因子:协助RNA聚合酶识别终止信号的辅助因子. 或蛋白质。")
24
§13.1 DNA指导下的RNA 的合成:转录 含富GC的回文序 列和寡聚U序列 (1)不依赖ρ-因子的终止子
不依赖ρ-因子的终止子")
25
§13.1 DNA指导下的RNA 的合成:转录 不依赖r -因子的终 止子的终止反应

26
§13.1 DNA指导下的RNA 的合成:转录 (2)依赖ρ -因子的终止子: r -因子:含六聚体的寡聚蛋白,具有:
依赖RNA的NTP酶活性,它结合于新生RNA上,借助于 水解NTP产生能量推动RNA聚合酶向前移动, RNA-DNA解螺旋酶活性,当酶遭遇终止子时暂停前 进, ρ-因子追上酶,与酶相互作用释放DNA-RNA 杂螺旋中的RNA.

27
§13.1 DNA指导下的RNA 的合成 依赖ρ -因子的终止子的终止反应:

28
§13.1 DNA指导下的RNA 的合成:转录 3、真核生物RNA聚合酶的启动子 真核生物的启动子非常复杂,其识别由转录因子而不 是σ因子;
转录因子的种类繁多; 增强子(initiator) :能增强转录的顺式作用元件(DNA 序列),但没有启动子功能,存在的位置不定; 顺式作用元件:存在于DNA分子上的调控转录的序列; 反式作用元件:能与顺式作用元件结合而调控转录的 蛋白因子。
 :能增强转录的顺式作用元件(DNA. 序列),但没有启动子功能,存在的位置不定; 顺式作用元件:存在于DNA分子上的调控转录的序列; 反式作用元件:能与顺式作用元件结合而调控转录的. 蛋白因子。")
29
真核生物RNA聚合酶Ⅱ的启动子 调节序列 -30区,解链位置,决定转录起点 称上游因子,影 响转录起始频率

30
§13.1 DNA指导下的RNA 的合成:转录 真核RNA聚合酶Ⅱ需要许多其它蛋白因子

31
五、真核生物的转录过程 §13.1 DNA指导下的RNA 的合成:转录 1. 转录的起始:
首先TATA结合蛋白(TBP)结合于TATA盒,然后 TFⅡB、TFⅡF、TFⅡE、polⅡ、TFⅡH也依次结 合在启动子处形成闭合复合物; TFⅡH作用于起始子(Inr)位置开始解螺旋形成开 放复合物; TFⅡH具有激酶活性,它将RNA聚合酶的C-端结构域 的多个位点磷酸化,使起始复合物的构象改变而促 进转录的起始阶段过渡进入延伸阶段。
结合于TATA盒,然后. TFⅡB、TFⅡF、TFⅡE、polⅡ、TFⅡH也依次结. 合在启动子处形成闭合复合物; TFⅡH作用于起始子(Inr)位置开始解螺旋形成开. 放复合物; TFⅡH具有激酶活性,它将RNA聚合酶的C-端结构域. 的多个位点磷酸化,使起始复合物的构象改变而促. 进转录的起始阶段过渡进入延伸阶段。")
32
真核RNA聚合酶Ⅱ和转录因子在启动子上的装配
§13.1 DNA指导下的RNA 的合成:转录 真核RNA聚合酶Ⅱ和转录因子在启动子上的装配 起始复合物装配完成,TFⅡH使CTD磷酸化

33
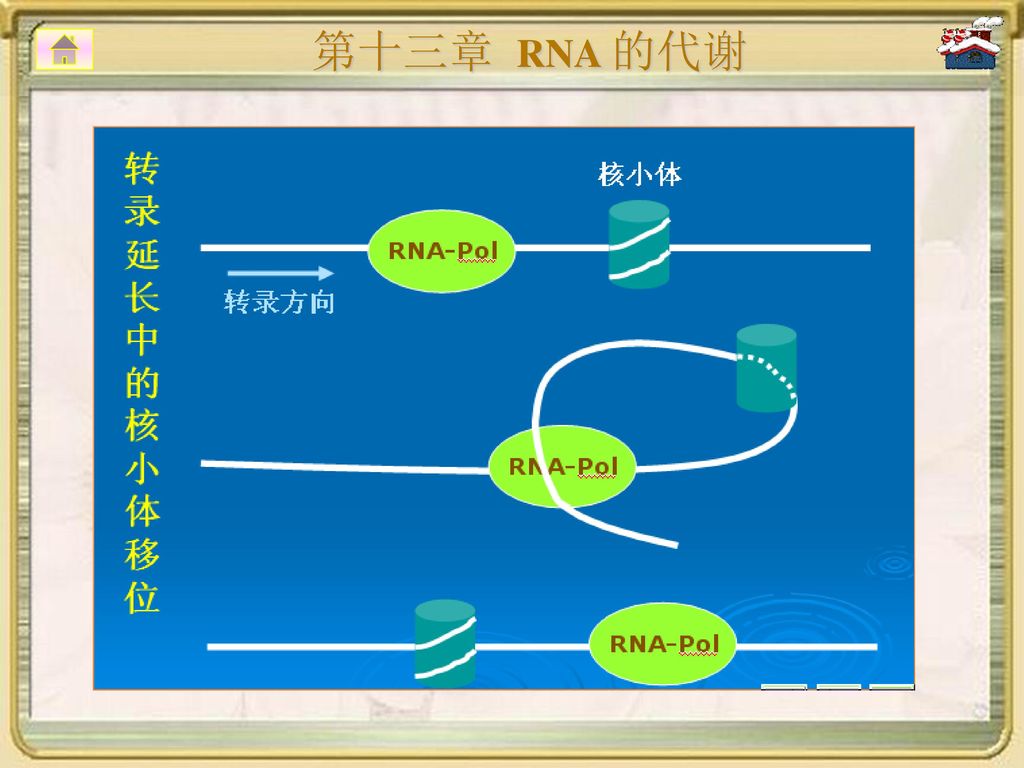
延长反应开始后,TFⅡE、TFⅡH释放;
§13.1 DNA指导下的RNA 的合成 2.转录的延伸过程: 延长反应开始后,TFⅡE、TFⅡH释放; 延长因子加入,与RNA聚合酶、TFⅡF形成延伸 复合物,使RNA聚合酶的延长效率大大促进; 真核生物转录延伸过程与原核生物相似,但因核 膜相隔,没有原核生物的转录与翻译同步现象。 RNA聚合酶前移时会遭遇核小体,电镜可观察到核 小体移位和解聚现象。

35
§13.1 DNA指导下的RNA 的合成:转录 3.转录终止 终止过程与RNA转录后的加工紧密联系;
下一个转录的起始。

36
烷化剂:使DNA发生烷基化,发生在G-N7、A-N1或N3 或
§13.1 DNA指导下的RNA 的合成 1、模板抑制剂 六、RNA生物合成的抑制剂:三类 烷化剂:使DNA发生烷基化,发生在G-N7、A-N1或N3 或 N7 、C-N1,易引起嘌呤的水解,在DNA上留下空隙干扰 复制或转录;或引起碱基错配。 嵌入染料:可插入双链DNA相邻的碱基对之间,一般具有芳 香族发色团。溴化乙锭(EB)是一种高灵敏的荧光试剂, 常用来检测DNA和RNA。与DNA结合后抑制其复制和转录。 放线菌素D:放线菌素D与DNA形成非共价的复合物,抑制 其模板功能(低浓度抑制转录,较高浓度抑制复制)。具 有类似作用的还有色霉素A3 、橄榄霉素、光神霉素。
是一种高灵敏的荧光试剂, 常用来检测DNA和RNA。与DNA结合后抑制其复制和转录。 放线菌素D:放线菌素D与DNA形成非共价的复合物,抑制. 其模板功能(低浓度抑制转录,较高浓度抑制复制)。具. 有类似作用的还有色霉素A3 、橄榄霉素、光神霉素。")
37
§13.1 DNA指导下的RNA 的合成:转录 放线菌素D的结构与作用 放线菌素 D:模板抑制剂 (丫啶) 口
 口")
38
§13.1 DNA指导下的RNA 的合成:转录 2、嘌呤和嘧啶类似物:人工合成的碱基类似物能够抑制
和干扰核酸的合成或直接掺到核酸分子中,形成异常 RNA或DNA。 SH NH2 OH NH2 NH2 NH2 SH F

39
§13.1 DNA指导下的RNA 的合成:转录 3、RNA聚合酶抑制剂 (1)利福霉素:抑制细菌RNA聚合酶活性。利福平可以高
效抑制结核杆菌,杀死麻疯杆菌,在体外有抗病毒作 用。 (2)利链霉素:与细菌RNA聚合酶亚基结合,抑制转录过 程中RNA链的延长反应。 (3)-鹅膏蕈碱:抑制真核生物RNA聚合酶活性。
利链霉素:与细菌RNA聚合酶亚基结合,抑制转录过. 程中RNA链的延长反应。 (3)-鹅膏蕈碱:抑制真核生物RNA聚合酶活性。")
40
§13.2 转录后RNA的加工 本节主要内容 一、原核生物RNA的加工 四、RNA的编辑 三、RNA的拼接机理 二、真核生物RNA的加工

41
一、原核生物RNA的加工 (一)mRNA前体的加工: 不需要加工,通常一边转 录一边翻译,即转录未结 束,翻译已经开始。
返回

42
§13.2 转录后RNA的加工 2、少数多顺反子mRNA需要内切酶切成更小单位。 加工时即将两部分切开。
核糖体大亚基蛋白基因 L10/ L7/ L12 RNA聚合酶亚基基因: β-亚基、β’-亚基 处于同一转录单位,同时转录,单独翻译 加工时即将两部分切开。 细胞内RNA聚合酶的翻译水平远低于核糖体蛋白的翻 译水平,将两者切开,有利于各自的翻译的调控。 类似的加工在某些噬菌体中也很常见。

43
§13.2 转录后RNA的加工 (二)、rRNA前体的加工 甲基化 核酸外切酶 6500个核苷酸(30 S) 内切酶RNaseⅢ、P 外切酶
、rRNA前体的加工 甲基化 核酸外切酶 6500个核苷酸(30 S) 内切酶RNaseⅢ、P 外切酶")
44
§13.2 转录后RNA的加工 E.coli 30SrRNA前体的结构

45
(三)、tRNA的加工 内切酶将tRNA两端切断; 外切酶从两端向内切去 多余序列:修剪; 在3’-端加CCAOH序列;切 除居间序列;
核苷酸的修饰与异构化。 在3’-端加CCAOH序列;切 除居间序列; 外切酶从两端向内切去 多余序列:修剪;

46
§13.2 转录后RNA的加工 tRNAPhe的加工过程

47
§13.2 转录后RNA的加工 二、真核生物RNA的一般加工 (一)mRNA前体的加工 卵清蛋白基因与成熟mRNA的比较 其中:
A、B、C、D、E、 F、G 为内含子, 1、2、3、4、5、 6、7 为外显子。

48
§13.2 转录后RNA的加工 真核生物的结构基因通常是断裂的 断裂基因(spilit gene):真核生物的结构基因中常被
一些不能表达成蛋白质的核苷酸序列-内含子隔开成若 干部分,这样的基因称为断裂基因。 外显子(exon):断裂基因中能表达成mRNA的序列 (即可为蛋白质编码的序列)称为外显子。 内含子(intron):断裂基因中不能为mRNA编码的 序列。 原核生物的结构基因一般是连续的,不含内含子; 真核生物的结构基因一般含多个内含子不等。
:断裂基因中能表达成mRNA的序列. (即可为蛋白质编码的序列)称为外显子。 内含子(intron):断裂基因中不能为mRNA编码的. 序列。 原核生物的结构基因一般是连续的,不含内含子; 真核生物的结构基因一般含多个内含子不等。")
49
§13.2 转录后RNA的加工 因发现断裂基因而获1993年诺贝尔生理学奖的Richard J Roberts and Phllip A Sharp Richard J Roberts 1943 Phllip A Sharp 1944

50
§13.2 转录后RNA的加工 卵清蛋白hnRNA的加工过程

51
真核mRNA前体的加工步骤: §13.2 转录后RNA的加工 加工包括: 1、5’端连接“帽子”结构(m7G5ppp5NmpNp-);
3、hnRNA被剪接,把内含子转录序列剪掉,把外 显子转录序列拼接上。 4、分子内部的核苷酸甲基化修饰。 1、5’端连接“帽子”结构(m7G5ppp5NmpNp-); 2、3’端添加polyA “尾巴”;
; 2、3’端添加polyA 尾巴 ;")
52
§13.2 转录后RNA的加工

53
§13.2 转录后RNA的加工 一种真核mRNA的5’-“帽子”加工

54
§13.2 转录后RNA的加工 真核生物mRNA3’-“尾巴”的加工 内切酶

55
§13.2 转录后RNA的加工 (二)真核rRNA前体的加工 (45S)
真核rRNA前体的加工 (45S)")
56
§13.2 转录后RNA的加工 (三)真核tRNA前体的加工
真核tRNA前体的加工")
57
§13.2 转录后RNA的加工 三、RNA的拼接机理 ●大多真核生物基因是断裂基因,即含有内含子。
●断裂基因的转录产物必需通过拼接,去除内含 子,使编码区(外显子,exon)成为连续序列, 这是基因表达的重要环节。 ●内含子的结构多种多样,拼接机制也是多种 多样的。
成为连续序列, 这是基因表达的重要环节。 ●内含子的结构多种多样,拼接机制也是多种. 多样的。")
58
(一)内含子的分类 根据基因的类型和剪接的方式不同,通常把内含子分 为四类: Ⅰ类:主要存在核、线粒体、叶绿体的rRNA基因,也有
线粒体、叶绿体的mRNA的基因; Ⅱ类:主要存在真菌、藻类和植物线粒体、叶绿体的 mRNA的基因中; Ⅲ类:大多数mRNA的基因所含的内含子,常以形成套索 结构而剪接; Ⅳ类:是tRNA基因及其初级转录产物中的内含子,剪接 过程需要酶与ATP。

59
§13.2 转录后RNA的加工 (二)RNA的四种拼接方式 核蛋白体(snRNP)的作用(类型Ⅲ内含子的切除)。
类型Ⅰ内含子的自我拼接:核酶的作用 类型Ⅱ内含子的自我拼接:核酶的作用 hnRNA的拼接:核内小RNA(SnRNA)与蛋白质组成 核蛋白体(snRNP)的作用(类型Ⅲ内含子的切除)。 核tRNA的拼接:酶促拼接
与蛋白质组成. 核蛋白体(snRNP)的作用(类型Ⅲ内含子的切除)。 核tRNA的拼接:酶促拼接.")
60
§13.2 转录后RNA的加工 背景知识: 核酶的发现 1981年Cech T在研究四膜虫(Tetrahymena thermophila)
背景知识: 核酶的发现 1981年Cech T在研究四膜虫(Tetrahymena thermophila) rRNA前体拼接过程中发现,此类的拼接无需蛋白质的酶 参与作用,它可自我催化完成。Cech称具有催化功能的 RNA为核酶(ribozyme)。 随后Altman S发现RNaseP中的M1RNA单独也具有催 化功能 。 1989年cech T和Altman S共同获诺贝尔化学奖。 核酶的发现改变了酶的化学本质是蛋白质的传统观 念,被认为是近三十年来生物化学领域中最令人鼓 舞的成就之一。
 rRNA前体拼接过程中发现,此类的拼接无需蛋白质的酶. 参与作用,它可自我催化完成。Cech称具有催化功能的. RNA为核酶(ribozyme)。 随后Altman S发现RNaseP中的M1RNA单独也具有催. 化功能 。 1989年cech T和Altman S共同获诺贝尔化学奖。 核酶的发现改变了酶的化学本质是蛋白质的传统观. 念,被认为是近三十年来生物化学领域中最令人鼓. 舞的成就之一。")
61
发现四膜虫rRNA前体的中的内含子是自我剪接的thomas Cech
发现核糖核酸酶P的M1RNA具有催化活性的Altman 1939 发现四膜虫rRNA前体的中的内含子是自我剪接的thomas Cech 1947 因发现核酶而获1989年诺贝尔奖化学的两位科学家

62
借助与鸟苷酸或鸟苷的作用,内含子自我催化,
§13.2 转录后RNA的加工 1、类型Ⅰ内含子的自我拼接(P333图13-11) 借助与鸟苷酸或鸟苷的作用,内含子自我催化, 通过两次转酯反应完成拼接,切下L19RNA。 L19RNA 四膜虫rRNA的剪接方式
 借助与鸟苷酸或鸟苷的作用,内含子自我催化, 通过两次转酯反应完成拼接,切下L19RNA。 L19RNA. 四膜虫rRNA的剪接方式.")
63
§13.2 转录后RNA的加工 四膜虫rRNA前体中内含子Ⅰ(L19RNA)的二级结构 5’-端的核苷酸序列
的二级结构 5’-端的核苷酸序列")
64
§13.2 转录后RNA的加工 L19 RNA体外催化的反应 反应时间 pG9 pG8 pG7 pG6 pG5 pG4 pG3

65
也是由内含子自 2、类型Ⅱ内含子的自我拼接 §13.2 转录后RNA的加工 我催化完成,但转 酯反应无鸟苷的参 与,两次转酯反应
后内含子形成套索 结构而切下。

66
3. hnRNA的加工:依赖snRNP的剪接作用
小核RNA(Small nuclear RNA,snRNA): 细胞核内存在的小分子RNA,大小约100~300个核苷酸, 因尿嘧啶含量较高得名,称之为U系列的snRNA 。 snRNA与多种蛋白质结合形成snRNP(核蛋白体)参 与hnRNA 的加工。 U1、U2、U4、U5、U6等snRNA结合形成的snRNP参与 hnRNA的拼接 U3 snRNP参与rRNA的拼接。
: 细胞核内存在的小分子RNA,大小约100~300个核苷酸, 因尿嘧啶含量较高得名,称之为U系列的snRNA 。 snRNA与多种蛋白质结合形成snRNP(核蛋白体)参. 与hnRNA 的加工。 U1、U2、U4、U5、U6等snRNA结合形成的snRNP参与. hnRNA的拼接. U3 snRNP参与rRNA的拼接。")
67
§13.2 转录后RNA的加工 hnRNA的拼接过程

68
4、核内tRNA前体的酶促拼接(内含子Ⅳ的切除)
核酸内切酶 环磷酸二酯酶 激酶 连接酶 磷酸酯酶

69
§13.2 转录后RNA的加工 (三)RNA拼接的意义 RNA拼接现象给生物学家一系列令人困惑的疑问:
为什么生物机体要先转录内含子,然后将其切除? RNA合成后再拼接是一个非常耗能的过程,对细胞 其收益是什么? 内含子由何而来? 内含子有无生物功能?

70
围绕以上问题,提出一些看法或观点,但争议很大,
目前尚无定论: 1、RNA的拼接是生物机体在进化历史中形成的,是进 化的结果; 2、RNA的拼接是基因表达的重要环节。RNA转录后通过 拼接而抽取有用信息,形成连续的编码序列,并可通 过选择性拼接而控制生物机体生长发育。因此,这是 真核生物遗传信息精确调节和控制的方式之一。

71
§13.2 转录后RNA的加工 3、基因由模块装配而成,模块间的间隔序列也就演变 4、RNA的拼接主要存在于真核生物,原核生物极为少见,
成内含子,因此外显子和内含子有着同样的古老历 史。而现今存在的几类内含子也各自有其起源和进 化历史,从它们的拼接方式和分布可以大致推测其 起源时间。 4、RNA的拼接主要存在于真核生物,原核生物极为少见, 但并非完全没有。一种合理的解释是原核生物为适应 快速生长的需要,在进化中已将内含子丢掉。事实上, 快速生长的单细胞如酵母,其编码的基因也几乎没有 内含子。然而内含子的存在和拼接作用对生物机体的 进化十分重要。

72
§13.2 转录后RNA的加工 5、内含子和外显子是相对的,有些内含子具有编码序 列,能产生蛋白质和功能RNA。如Ⅰ型内含子能产生
核酸内切酶,Ⅱ型内含子能产生核酸内切酶和逆转 录酶,以帮助内含子的转移。

73
§13.2 转录后RNA的加工 四、RNA的编辑 1、不同RNA拼接方式导致一个基因多种产物 降钙素 甲状腺 脑 降钙素基 因相关肽

74
真核生物复杂转录产物的两种不同的剪接方式
§13.2 转录后RNA的加工 真核生物复杂转录产物的两种不同的剪接方式

75
一、逆转录(reverse transcription)
§13.3 RNA指导下DNA的合成:逆转录 一、逆转录(reverse transcription) 定义: 以RNA为模板,按照RNA中的核苷酸顺序合成DNA 的过程称为逆转录,由逆转录酶催化进行。 1970年Temin等和Baltimore分别在前人研究劳氏肉瘤 病毒(ASV)和小白鼠白血病病毒(MLV) 的基础上发现: 用特异抑制物(放线菌素D)能抑制ASV病毒和MLV病毒 的复制,而对一般RNA病毒的复制无影响。已知放线菌素D 专门抑制以DNA为模板的反应,可见AVS病毒和MLV病毒的 复制过程必然涉及到DNA。所以Temin于1964年提出前病毒 的假说。
 定义: 以RNA为模板,按照RNA中的核苷酸顺序合成DNA. 的过程称为逆转录,由逆转录酶催化进行。 1970年Temin等和Baltimore分别在前人研究劳氏肉瘤. 病毒(ASV)和小白鼠白血病病毒(MLV) 的基础上发现: 用特异抑制物(放线菌素D)能抑制ASV病毒和MLV病毒. 的复制,而对一般RNA病毒的复制无影响。已知放线菌素D. 专门抑制以DNA为模板的反应,可见AVS病毒和MLV病毒的. 复制过程必然涉及到DNA。所以Temin于1964年提出前病毒. 的假说。")
76
2.前病毒学说:致癌RNA病毒基因组RNA复制时必须经过
§13.3 RNA指导下DNA的合成:逆转录 2.前病毒学说:致癌RNA病毒基因组RNA复制时必须经过 一个DNA中间体即前病毒,此前病毒再整合到宿主细 胞中,随细胞的增殖而传递给子细胞,导致细胞的恶 性转化,即致癌RNA病毒的遗传信息可以由RNA传递至 DNA. 1970年Temin等和Baltimore分别从劳氏肉瘤病毒和小白鼠 白血病病毒等致病RNA病毒中分离出逆转录酶,迄今已知 的致癌RNA病毒都含有逆转录酶.

77
核糖核酸酶H的活性,专一水解RNA-DNA杂交分子 DNA指导的DNA聚合酶活性; RNA指导的DNA聚合酶活性;
3.逆转录酶是多功能酶,兼有3种酶的活性: 无校对功能,错误率高,易产生变异。 核糖核酸酶H的活性,专一水解RNA-DNA杂交分子 中的RNA,可沿5’3’方向起核酸外切酶的作用; DNA指导的DNA聚合酶活性; RNA指导的DNA聚合酶活性;

78
§13.3 RNA指导下DNA的合成:逆转录 致癌RNA病毒的逆转录过程 逆转录酶也和DNA聚合酶一样,沿53’方向合成
细胞恶性转化; +RNA -DNA RNA-DNA杂交分子 +RNA -DNA +DNA 双链DNA(前病毒) 单链病毒RNA +RNA 逆转录酶也和DNA聚合酶一样,沿53’方向合成 DNA,底物为四种dNTP,并要求短链RNA作引物。
 单链病毒RNA. +RNA. 逆转录酶也和DNA聚合酶一样,沿53’方向合成. DNA,底物为四种dNTP,并要求短链RNA作引物。")
79
§13.3 RNA指导下DNA的合成:逆转录 逆转录病毒的生活史

80
4.逆转录病毒的基因组结构 §13.3 RNA指导下DNA的合成:逆转录 劳氏肉瘤病毒基因组
LTR gag pol env src LTR:长末端重复序列; gag:病毒粒子结构蛋白基因,编码基质蛋白、衣壳蛋白 gag+pol:编码逆转录酶、整合酶、蛋白酶 evn:编码病毒外膜糖蛋白前体 src:癌基因,导致细胞癌变,是一种酪蛋白激酶

81
§13.3 RNA指导下DNA的合成:逆转录 HIV病毒基因组: gag vif tat vpu env pol rev nef LTR
rve vpr nef gag tat vif tat vpu env pol LTR LTR HIV病毒:感染T淋巴细胞后杀死细胞,造成宿主机 体免疫系统损伤,引起艾滋病(AIDS)
")
82
§13.3 RNA指导下DNA的合成:逆转录 5.逆转录酶发现的理论与实践意义: 理论上: 逆转录酶催化的逆转录和遗传信息的流向是对传统
的中心法则的一种补充和发展。 致癌RNA病毒和肝炎DNA病毒都含逆转录酶,但前 者遗传信息按RNA→DNA→RNA流向,后者按DNA→ RNA→DNA流向,这为两类的病毒防治或治疗提供 了必要的理论依据。 真核生物细胞中也发现逆转录酶存在,如DNA端粒 中、网织红细胞和正在分裂的淋巴细胞中,这可能 与细胞分化和抗体的形成有关。

83
§13.3 RNA指导下DNA的合成:逆转录 1983年,发现人类免疫缺陷病毒(human immune
deficiency virus,HIV),感染T淋巴细胞后即杀死 细胞,造成宿主机体免疫系统损伤,引起艾滋病 (acquired immunodeficiency syndrome,AIDS), 该病毒为逆转录病毒。 逆转录酶的实践意义:构建cDNA文库、艾滋病、癌症 药物设计等。 cDNA:互补DNA ,几乎所有真核生物mRNA分子的3‘末端 都有一段polyA,当加入寡聚dT作为引物时,mRNA就可作为模 板,在逆转录酶催化下在体外合成与该mRNA互补的DNA,称为 cDNA。
,感染T淋巴细胞后即杀死. 细胞,造成宿主机体免疫系统损伤,引起艾滋病. (acquired immunodeficiency syndrome,AIDS), 该病毒为逆转录病毒。 逆转录酶的实践意义:构建cDNA文库、艾滋病、癌症. 药物设计等。 cDNA:互补DNA ,几乎所有真核生物mRNA分子的3‘末端. 都有一段polyA,当加入寡聚dT作为引物时,mRNA就可作为模. 板,在逆转录酶催化下在体外合成与该mRNA互补的DNA,称为. cDNA。")
84
因发现逆转录病毒的遗传物质而获1975年 诺贝尔奖的三位科学家 Howard Martin Temin USA
§13.3 RNA指导下DNA的合成:逆转录 因发现逆转录病毒的遗传物质而获1975年 诺贝尔奖的三位科学家 Daviel Baltimore USA 1938 Howard Martin Temin USA Renoto Dulbecco USA 1914

85
§13.3 RNA指导下DNA的合成:逆转录 5、端粒酶是一种逆转录酶 端粒(telomere):真核染色体末端的特殊结构,它
一般含许多串联重复的寡聚核苷酸序列,通常是 TxGy,x、y常为1-4。 端粒结构提出了一个特别的生物学问题。DNA复制需 要引物,但在线形DNA分子中,DNA复制结束后,引 物去除留下的链的短缺无法进行补缺,若没有特殊 的机制解决末端复制,DNA就会因为复制而不断变短。 这个问题是由一种特殊的酶—端粒酶(telomerase) 解决的。
 解决的。")
86
端粒酶是一种逆转录酶 §13.3 RNA指导下DNA的合成:逆转录 端粒酶:端粒的合成酶
含有一段RNA和蛋白质,其中的RNA成分为150个核 苷酸,并含1.5个拷贝的与端粒互补的重复序列 CyAx,这是端粒TxGy合成的模板。 端粒酶是以该段RNA为模板的逆转录酶,端粒合成时 以该段RNA为引物和模板,合成因引物切除而产生的 DNA末端缺口,使染色体的端粒达到正常的长度。

87
§13.3 RNA指导下DNA的合成:逆转录 端粒的合成

88
§13.3 RNA指导下DNA的合成:逆转录

89
§13.4 RNA指导下的RNA的合成:RNA复制
两个阶段: (1)病毒RNA可充当mRNA,利用寄主中的核糖体合成外壳 蛋白和复制酶的β亚基。 (2)复制酶的β亚基与来自宿主细胞的亚基α’ σ自动 装配成RNA复制酶,进行RNA复制,通常以分子中单链 RNA为模板(正链),复制出一条新的RNA链(负 链),然后以负链为模板复制出大量正链,再与外壳 蛋白组装成新的噬菌体颗粒。
病毒RNA可充当mRNA,利用寄主中的核糖体合成外壳. 蛋白和复制酶的β亚基。 (2)复制酶的β亚基与来自宿主细胞的亚基α’ σ自动. 装配成RNA复制酶,进行RNA复制,通常以分子中单链. RNA为模板(正链),复制出一条新的RNA链(负. 链),然后以负链为模板复制出大量正链,再与外壳. 蛋白组装成新的噬菌体颗粒。")
90
二、RNA复制酶的催化性质 §13.4 RNA指导下的RNA的合成:RNA复制 以四种NTP为底物; 专一性地选择病毒RNA为模板;
错误率高,易变异。 按5’ →3’的方向合成病毒RNA; 专一性地选择病毒RNA为模板;

91
病毒RNA的复制方式 §13.4 RNA指导下的RNA的合成:RNA复制 1、病毒含正链RNA:进入宿主细胞后先进行病毒RNA复制酶
和有关病毒蛋白质的合成(借助于宿主细胞的蛋白质合 成体系),然后进行RNA的复制,再装配病毒颗粒。如: 噬菌体、Qβ和灰质炎病毒。 2、病毒含负链RNA和复制酶:这类病毒进入宿主细胞后, 先进行RNA的复制合成正链RNA,再以正链RNA为模板合 成病毒蛋白质、负链RNA,然后装配病毒颗粒。如:狂 犬病病毒、马水苞性口炎病毒。
,然后进行RNA的复制,再装配病毒颗粒。如: 噬菌体、Qβ和灰质炎病毒。 2、病毒含负链RNA和复制酶:这类病毒进入宿主细胞后, 先进行RNA的复制合成正链RNA,再以正链RNA为模板合. 成病毒蛋白质、负链RNA,然后装配病毒颗粒。如:狂. 犬病病毒、马水苞性口炎病毒。")
92
§13.4 RNA指导下的RNA的合成:RNA复制

93
§13.4 RNA指导下的RNA的合成:RNA复制
(±)双链DNA (-)DNA (+)RNA (-)RNA RNA (+)链 (±)双链RNA (+)RNA
双链DNA. (-)DNA. (+)RNA. (-)RNA. RNA. (+)链. (±)双链RNA. (+)RNA.")
94
本章完


 防治宣導.>")
 的字头 AIDS 的译称,中文名为获得性免疫缺 陷综合症,是由一种人类免疫缺陷病毒 (Human.>")
,必须在活细胞内寄生并以复 制方式增殖的非细胞型微生物。 生物病毒微生物 原指一种动物来源的 毒素。 “virus” 一词源于 拉丁文。 病毒能增殖、遗传和演化,因而 具 有生命最基本的特征。>")

 ” 。 他的体内没有任何免疫系统, 没有任何抵御细菌、病毒的能力。>")





>")







 蕭伊玲 18號 (510919)>")
