Download presentation
Presentation is loading. Please wait.

1
11 糖代谢中的其它途径

2
11.1 戊糖磷酸途径能提供核糖-5-磷酸和NADPH
11.2 葡萄糖醛酸途径可以生成糖醛酸和抗坏血酸 11.3 饮食中的其它糖可以经酵解途径降解 1 . 果糖可以转换为甘油醛-3-磷酸 2 . 半乳糖可被转换为葡萄糖-1-磷酸 3 . 甘露糖可转换为果糖-6-磷酸 11.4 糖原的降解需要磷酸化酶、转移酶和去分支酶 11.5 糖原合成的底物是UDP-葡萄糖 11.6 葡萄糖可以通过糖异生途径由非糖物质合成

3
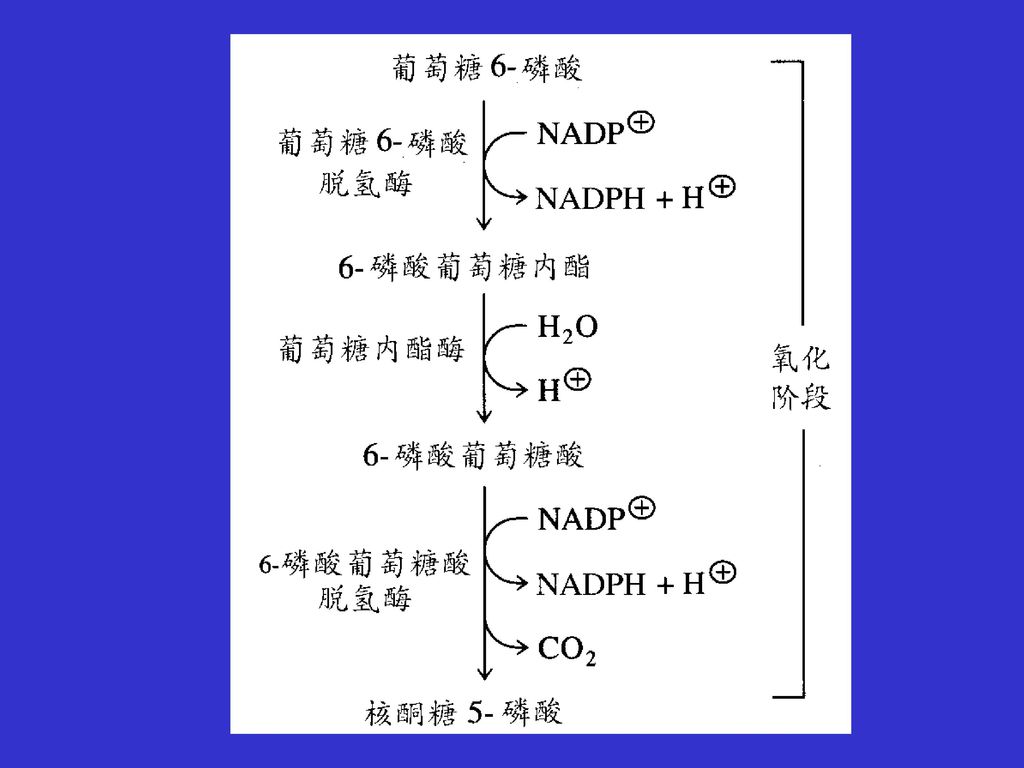
11.1 戊糖磷酸途径能提供核糖-5-磷酸和NADPH+H+
戊糖磷酸途径也称为己糖磷酸支路,提供核糖-5-磷酸和NADPH+H+。核糖-5-磷酸用于核酸的生物合成;而NADPH+H+主要用于需要还原力的生物合成中。 磷酸戊糖途径可以分为氧化阶段和非氧化阶段,在氧化反应阶段中,当葡萄糖-6-磷酸转换为五碳核酮糖-5-磷酸时,生成NADPH+H+ 。 葡萄糖-6-磷酸+2 NADP++H2O 核酮糖-5-磷酸+2NADPH+2H++CO2 如果细胞需要大量的NADPH+H+和核苷酸,则所有的核酮糖-5-磷酸都可异构化形成核糖-5-磷酸,戊糖磷酸途径就会终止于氧化阶段。通常需要的NADPH+H+要比核糖-5-磷酸多,所以大多数核糖-5-磷酸都转换为糖酵解的中间产物。

4
戊糖磷酸途径

7
磷酸戊糖途径氧化阶段反应 磷酸戊糖途径氧化阶段的第一个反应是葡萄糖-6-磷酸脱氢转化成6-磷酸葡萄糖酸内酯的反应,反应由葡萄糖-6-磷酸脱氢酶催化,反应中NADP+被还原生成NADPH+H+。这步反应是整个戊糖磷酸途径的主要调节部位,葡萄糖-6-磷酸脱氢酶受NADPH的别构抑制,通过这一简单调节,戊糖磷酸途径可以自我限制NADPH+H+的生产。 氧化阶段的第二个酶是葡萄糖酸内酯酶,它催化6-磷酸葡萄糖酸内酯水解生成6-磷酸葡萄糖酸,最后6-磷酸葡萄糖酸在第三个酶6-磷酸葡萄糖酸脱氢酶的作用下氧化脱羧生成核酮糖-5-磷酸、CO2和另一分子的NADPH+H+ 。氧化阶段的最重要的功能是提供NADPH。

8
氧化阶段生成NADPH+H+反应

9
3核酮糖-5-磷酸 2果糖-6-磷酸+甘油醛-3-磷酸
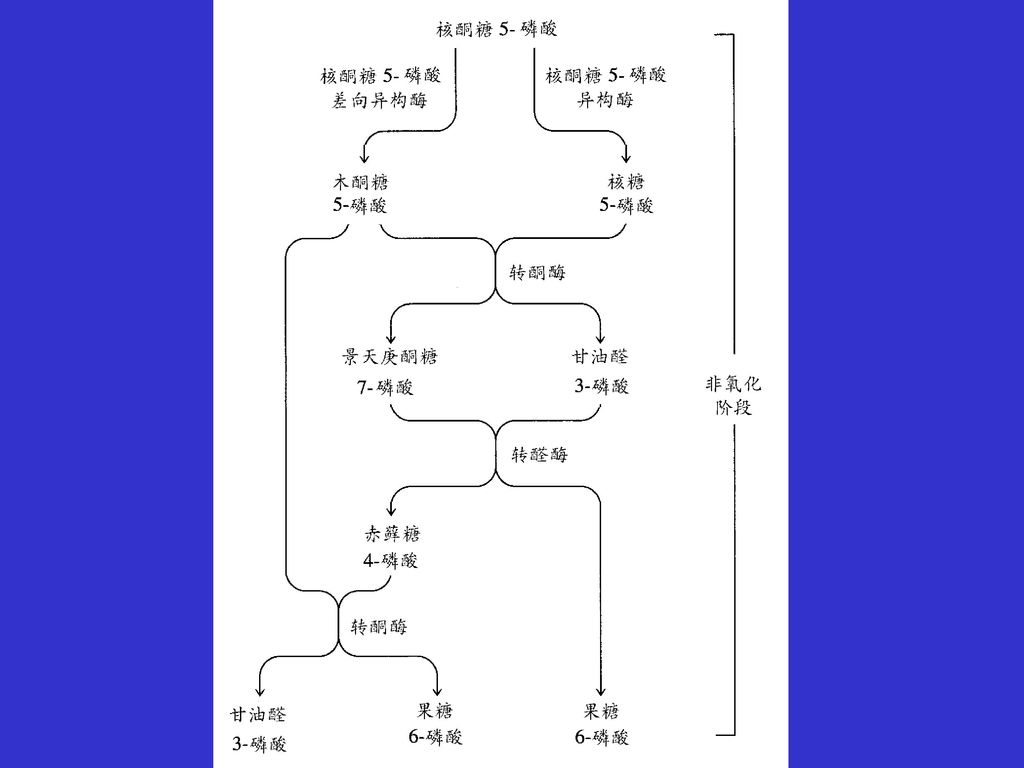
磷酸戊糖途径非氧化阶段反应 非氧化阶段是一条转换途径,通过氧化阶段产生的核酮糖-5-磷酸转换为糖酵解的中间产物果糖-6-磷酸和甘油醛-3-磷酸。如果所有的戊糖磷酸都转换为酵解的中间产物,3分子的戊糖分子可以转换为2分子的己糖和1分子的丙糖。 3核酮糖-5-磷酸 果糖-6-磷酸+甘油醛-3-磷酸 在差向异构酶和异构酶的催化下,核酮糖-5-磷酸转换为木酮糖-5-磷酸和核糖-5-磷酸; 木酮糖-5-磷酸和核糖-5-磷酸经转酮酶催化形成7碳产物景天庚酮糖-7- 磷酸和3碳产物甘油醛-3-磷酸; 景天庚酮糖-7-磷酸和3碳产物甘油醛-3-磷酸再经转醛酶催化转换为果糖-6-磷酸和赤藓糖-4-磷酸, 赤藓糖-4-磷酸再与另一分子的木酮糖-5-磷酸经转酮酶催化生成果糖-6-磷酸和甘油醛-3-磷酸。

10
非氧化阶段反应

11
转酮反应 转醛反应

12
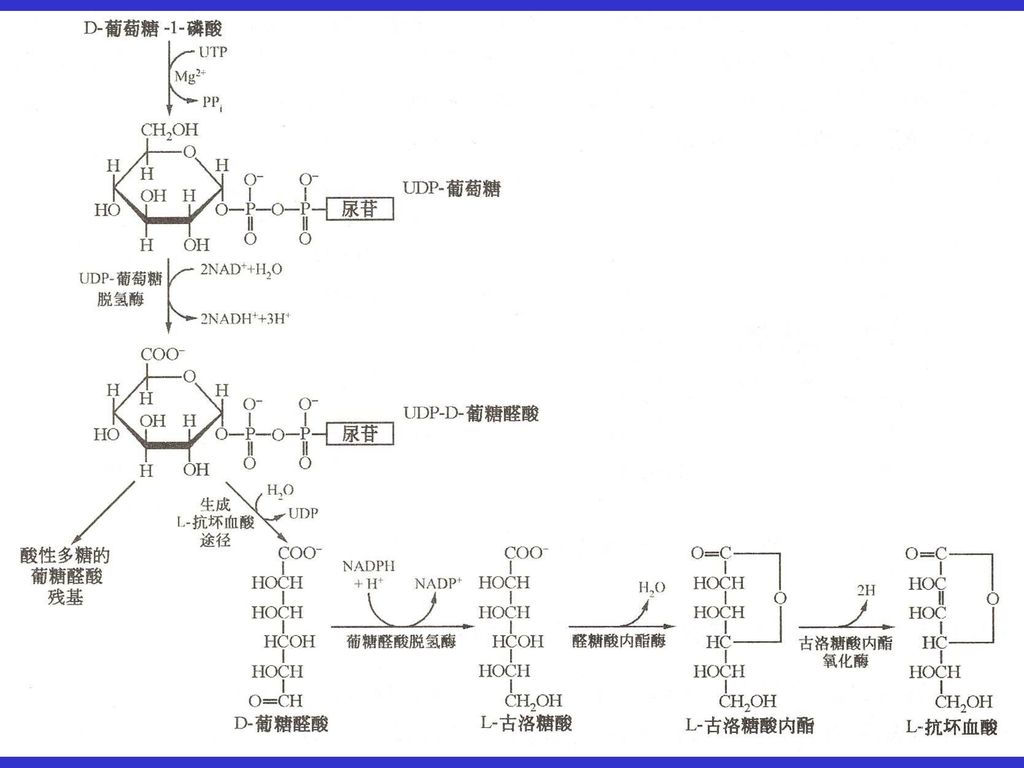
11.2葡萄糖醛酸途径可以生成糖醛酸和抗坏血酸 葡萄糖-1-磷酸首先与UTP反应生成UDP-葡萄糖,
葡萄糖醛酸途径是葡萄糖氧化的另一条次要途径,葡萄糖可以转换为两个特殊的产物:D-糖醛酸和L-抗坏血酸。 葡萄糖-1-磷酸首先与UTP反应生成UDP-葡萄糖, UDP-葡萄糖脱氢形成UDP-葡萄糖醛酸。 UDP-葡萄糖醛酸水解生成D-葡萄糖醛酸并释放出UDP。 D-葡萄糖醛酸经NADPH还原生成L-古洛糖酸,L-古洛糖酸在内酯酶的作用下形成L-古洛糖酸内酯,L-古洛糖酸内酯经古洛糖酸内酯氧化酶催化脱氢生成L-抗坏血酸。 包括人在内的某些动物,如豚鼠、猴、一些鸟和一些鱼等由于缺少古洛糖酸内酯氧化酶,不能生物合成抗坏血酸,所以必须从食物中摄取。人如果不能获得足够的维生素C,将发展为坏血病。

14
11.3 饮食中的其它糖可以经酵解途径降解 1 . 果糖可以转换为甘油醛-3-磷酸
在肝脏中,果糖激酶催化果糖磷酸化生成果糖-1-磷酸,反应需要ATP。 果糖-1-磷酸醛缩酶催化果糖-1-磷酸裂解生成甘油醛和磷酸二羟丙酮,后者经丙糖磷酸异构酶催化转换为甘油醛-3-磷酸, 甘油醛则是在丙糖激酶的作用下,消耗一分子ATP后生成甘油醛-3-磷酸。 总的转化结果是一分子果糖转化为二分子甘油醛-3-磷酸,同时消耗了两分子ATP。富含果糖或蔗糖的饮食由于丙酮酸的过量生成可能会导致脂肪肝,丙酮酸是脂肪和胆固醇生物合成的前体。

15
一分子果糖转化为二分子甘油醛-3-磷酸的过程

16
2 .半乳糖可被转换为葡萄糖-1-磷酸 乳糖酶催化乳糖水解为葡萄糖和半乳糖,半乳糖是葡萄糖的C-4差向异构体,可以通过尿苷二磷酸葡萄糖(UDP-葡萄糖)再循环途径转化为葡萄糖-1-磷酸。
再循环途径转化为葡萄糖-1-磷酸。")
17
作业:设计一条技术路线,解决亚洲人乳糖不耐症的问题,以实现国民素质的提高!
喂食奶制品的婴幼儿依赖于半乳糖代谢途径。患有半乳糖血症(不能正常代谢半乳糖)的婴幼儿都是缺乏半乳糖-1-磷酸尿苷酰基转移酶。缺少这种酶会造成细胞内半乳糖-1-磷酸的堆积,有可能损害肝的功能,这可通过使皮肤发黄的黄疸的出现来确认。 另外还可能损伤中枢神经系统。在婴儿出生时,通过检测脐带红细胞中的半乳糖-1-磷酸尿苷酰基转移酶可以确定是否患有半乳糖血症。如果在饮食中去掉乳糖可以避免这种遗传病带来的严重后果。 作业:设计一条技术路线,解决亚洲人乳糖不耐症的问题,以实现国民素质的提高!
的婴幼儿都是缺乏半乳糖-1-磷酸尿苷酰基转移酶。缺少这种酶会造成细胞内半乳糖-1-磷酸的堆积,有可能损害肝的功能,这可通过使皮肤发黄的黄疸的出现来确认。 另外还可能损伤中枢神经系统。在婴儿出生时,通过检测脐带红细胞中的半乳糖-1-磷酸尿苷酰基转移酶可以确定是否患有半乳糖血症。如果在饮食中去掉乳糖可以避免这种遗传病带来的严重后果。 作业:设计一条技术路线,解决亚洲人乳糖不耐症的问题,以实现国民素质的提高!")
18
2 .甘露糖可被转换为果糖-6-磷酸 甘露糖主要来自糖蛋白和某些多糖。甘露糖中己糖激酶催化下,转化为甘露糖-6-磷酸,然后在甘露糖异构酶催化下转化为果糖-6-磷酸。

19
11.4 糖原的降解需要磷酸化酶、转移酶和去分支酶
糖原磷酸化酶可以从糖原的非还原端连续地进行磷酸解,磷酸解直至距 -1,6 糖苷键的分支点还剩下4个葡萄糖单位的部位停止,剩下的底物称为极限糊精,极限糊精可以通过糖原去分支酶作用进一步降解。去分支酶具有葡聚糖转移酶和淀粉-1,6-葡糖苷酶两种催化活性。 葡聚糖转移酶催化支链上的3个葡萄糖残基转移到糖原分子的一个游离的4′端上,形成一个新的 -1,4 糖苷键,而淀粉-1,6-葡糖苷酶催化转移后剩下的通过 -1,6 糖苷键连接的葡萄糖残基的水解,释放出一分子的葡萄糖。因此对于原来糖原聚合物中的每个分支点都可释放出一分子葡萄糖。

20
糖原磷酸化酶可以从糖原的非还原端连续地进行磷酸解
变位反应 葡萄糖-1-磷酸在磷酸葡萄糖变位酶的作用下可以转换为葡萄糖-6-磷酸

21
糖原中大约90%的葡萄糖残基通过糖酵解却可以获得三分子ATP。
分支酶的葡聚糖转移酶活性 分支酶的淀粉-1,6-葡糖苷酶活性

22
11.5 糖原合成的底物 是UDP-葡萄糖 葡萄糖在己糖激酶催化下磷酸化生成葡萄糖-6-磷酸。
将一个葡萄糖-6-磷酸中的葡萄糖基结合到糖原上(糖原的延伸)都需要进行3步酶促反应。 P183
都需要进行3步酶促反应。 P183.")
23
1、 磷酸葡萄糖变位酶将葡萄糖-6-磷酸转换为葡萄糖-1-磷酸;
2、葡萄糖-1-磷酸在UDP-葡萄糖焦磷酸化酶作用下被UTP活化,生成UDP-葡萄糖和无机焦磷酸(PPi); 3、在糖原合酶(Synthase)催化下UDP-葡萄糖中的葡萄糖基通过-(1→4)糖苷键结在已合成的糖原(相当于合成的引物)的非还原端。 植物和某些细菌合成淀粉或糖原时,使用的底物是ADP-葡萄糖,而不是UDP-葡萄糖。 合成糖原或淀粉还需要另一种淀粉-(1,4→1,6)-转葡糖苷酶,催化糖原支链的形成,所以该酶又称为糖原分支酶,它从延伸的葡萄糖链的非还原端除去至少含有6个葡萄糖基的寡糖,然后通过-(1→6)连接酶再把该寡糖连接到离最近的-(1→6)分支点至少有4个葡萄糖基的位置。
; 3、在糖原合酶(Synthase)催化下UDP-葡萄糖中的葡萄糖基通过-(1→4)糖苷键结在已合成的糖原(相当于合成的引物)的非还原端。 植物和某些细菌合成淀粉或糖原时,使用的底物是ADP-葡萄糖,而不是UDP-葡萄糖。 合成糖原或淀粉还需要另一种淀粉-(1,4→1,6)-转葡糖苷酶,催化糖原支链的形成,所以该酶又称为糖原分支酶,它从延伸的葡萄糖链的非还原端除去至少含有6个葡萄糖基的寡糖,然后通过-(1→6)连接酶再把该寡糖连接到离最近的-(1→6)分支点至少有4个葡萄糖基的位置。")
24
糖原合酶

25
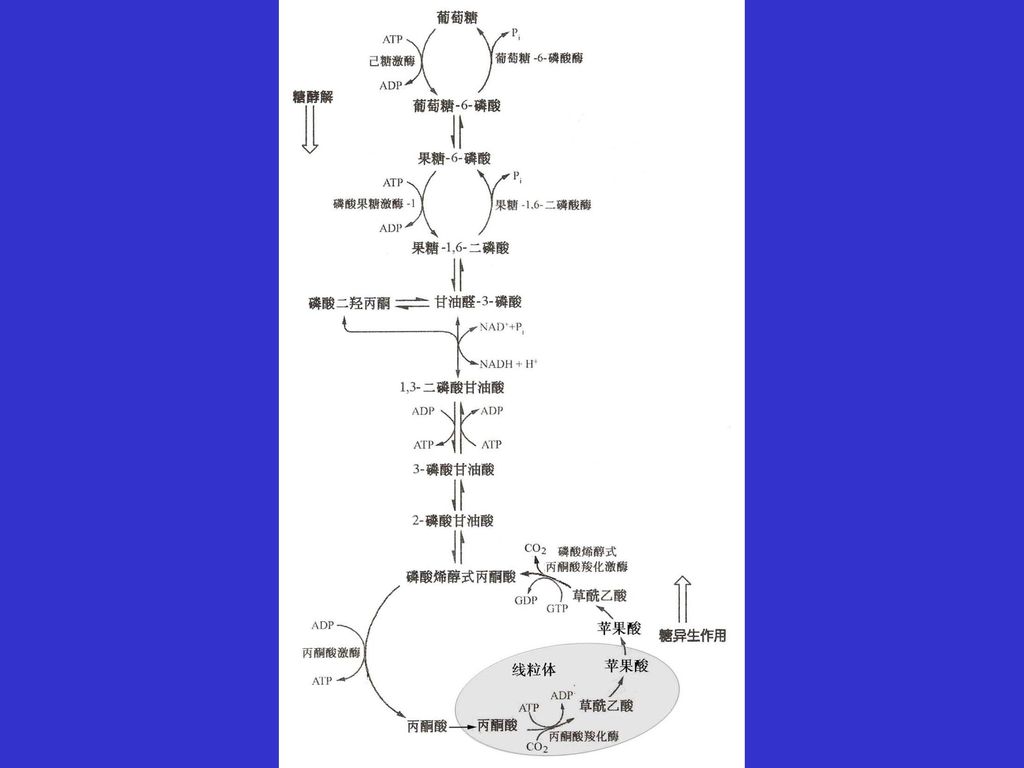
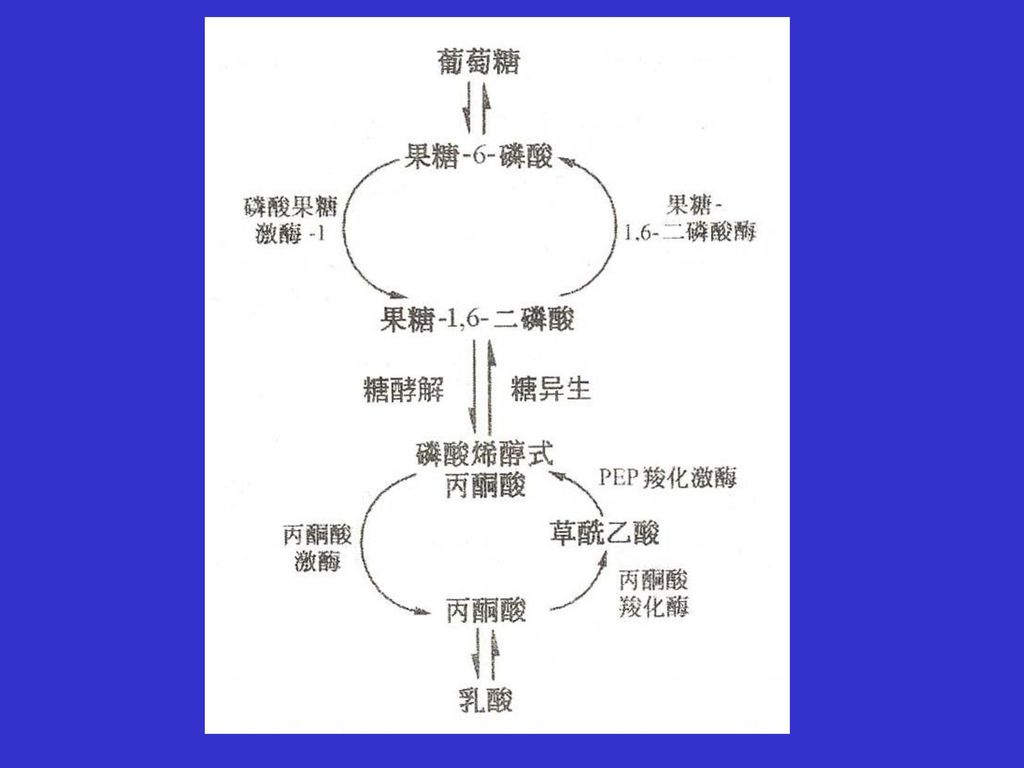
11.6 葡萄糖可以通过糖异生途径由非糖物质合成 大多数生物都有一个生物合成葡萄糖的途径。哺乳动物的某些组织,主要是肝脏、肾脏可以由非糖前体物质,例如乳酸和丙氨酸从头合成葡萄糖,由非糖前体物质合成糖的过程称为糖异生。下图比较了由丙酮酸生成葡萄糖的糖异生过程与葡萄糖的酵解过程。 从图中可以看出,糖异生和酵解两个过程中的许多中间代谢物是相同的,一些反应以及催化反应的酶也是一样的。酵解途径的七步可逆反应只要改变反应的方向就变成了糖异生中的反应了。 但糖异生并非是糖酵解的逆转,其中由丙酮酸激酶、磷酸果糖激酶和己糖激酶催化的三个高放能反应就是不可逆转的,需要消耗能量走另外途径,或由其它的酶催化,来克服这三个不可逆反应带来的能障。

27
1、丙酮酸羧化生成草酰乙酸 在丙酮酸羧化酶(生物素作为辅基)的催化下,丙酮酸羧化生成草酰乙酸,反应消耗一分子的ATP。丙酮酸羧化酶催化的反应是不可逆反应,反应受乙酰CoA别构抑制。
的催化下,丙酮酸羧化生成草酰乙酸,反应消耗一分子的ATP。丙酮酸羧化酶催化的反应是不可逆反应,反应受乙酰CoA别构抑制。")
28
2、草酰乙酸转化为磷酸烯醇式丙酮酸 丙酮酸羧化生成的草酰乙酸经磷酸烯醇式丙酮酸羧化激酶催化生成磷酸烯醇式丙酮酸。在体内该反应是不可逆的,但在体外,分离的磷酸烯醇式丙酮酸羧激酶却可以催化该反应的逆反应。

29
3、果糖-1,6-二磷酸水解生成果糖-6-磷酸 磷酸烯醇式丙酮酸和果糖-1,6-二磷酸之间的糖异生反应都是糖酵解途径中的逆反应。 但在糖异生途径中果糖-1,6-二磷酸不能通过酵解逆反应生成果糖-6-磷酸,而是使用另一个果糖-1,6-二磷酸酶催化果糖-1,6-二磷酸水解生成果糖-6-磷酸,反应释放出大量的自由能,反应也是不可逆的。

30
4、葡萄糖-6-磷酸水解生成葡萄糖 果糖-6-磷酸沿酵解的逆反应异构化生成葡萄糖-6-磷酸,但在糖异生途径中,葡萄糖-6-磷酸水解为葡萄糖和无机磷酸则需要另一个葡萄糖-6-磷酸酶,葡萄糖-6-磷酸水解反应是不可逆的。

31
2丙酮酸+4ATP+2GTP+2NADH+2H++6H2O→ 葡萄糖+4ADP+2GDP+6Pi+2NAD+
糖异生等于用了4分子ATP克服由2分子丙酮酸形成2分子高能磷酸烯醇式丙酮酸的能障,用了2分子ATP进行磷酸甘油激酶催化反应的可逆反应。葡萄糖经糖酵解转化为两分子丙酮酸净生成2分子ATP,而由两分子丙酮酸经糖异生途径合成一分子葡萄糖却消耗了6个ATP,糖异生比酵解净生成的ATP多用了4分子ATP。

32
果糖-6-磷酸+ATP―――→果糖-1,6-二磷酸+ADP
许多哺乳动物组织含有的糖异生途径的酶不完全,例如肌肉就是这样的组织,不能进行糖异生,但一些肌肉细胞含有高活性的果糖-1,6-二磷酸酶,却可以进行一种所谓的“无效循环”(也称为底物循环)反应。从下图中可以看出,磷酸果糖激酶和果糖-1,6-二磷酸酶同时作用的反应结果是ATP水解为ADP和Pi。 果糖-6-磷酸+ATP―――→果糖-1,6-二磷酸+ADP +) 果糖-1,6-二磷酸+H2O―――→果糖-6-磷酸+Pi _____________________________________________________ ATP + H2O―――→ADP+Pi
反应。从下图中可以看出,磷酸果糖激酶和果糖-1,6-二磷酸酶同时作用的反应结果是ATP水解为ADP和Pi。 果糖-6-磷酸+ATP―――→果糖-1,6-二磷酸+ADP. +) 果糖-1,6-二磷酸+H2O―――→果糖-6-磷酸+Pi _____________________________________________________. ATP + H2O―――→ADP+Pi.")
34
要点归纳 1. 所有生物中都存在戊糖磷酸途径,在细胞质中戊糖磷酸途径产生NADPH和核糖-5-磷酸,NADPH用于还原性生物合成,而核糖-5-磷酸用于RNA、DNA和核苷酸辅酶的合成中。 2. 戊糖磷酸途径开始于葡萄糖-6-磷酸脱氢形成6-磷酸葡糖酸内酯,内酯水解形成6-磷酸葡糖酸,然后氧化脱羧生成核酮糖-5-磷酸,在上述2步氧化反应中NADP+为电子受体,被还原为NADPH。在接下来的反应中核酮糖-5-磷酸(酮糖)异构化形成核糖-5-磷酸(醛糖)。当细胞需要的NADPH比核糖-5-磷酸更多时,核糖-5-磷酸通过转酮酶和转醛酶作用转换为甘油醛-3-磷酸和果糖-6-磷酸。转酮酶和转醛酶将戊糖磷酸途径和酵解联系起来。
异构化形成核糖-5-磷酸(醛糖)。当细胞需要的NADPH比核糖-5-磷酸更多时,核糖-5-磷酸通过转酮酶和转醛酶作用转换为甘油醛-3-磷酸和果糖-6-磷酸。转酮酶和转醛酶将戊糖磷酸途径和酵解联系起来。")
35
3. 糖原分子在糖原磷酸化酶催化下每次从糖原的非还原端磷酸解生成一分子葡萄糖-1-磷酸,非还原端降解生成的葡萄糖-1-磷酸经葡萄糖变位酶作用转换为葡萄糖-6-磷酸,在肌肉组织中葡萄糖-6-磷酸经酵解和柠檬酸循环进一步代谢,完全氧化为CO2和H2O,而在肝脏中,葡萄糖-6-磷酸水解生成葡萄糖,葡萄糖可进入血液补充血糖。而糖原分支点降解涉及到分支酶的葡聚糖糖转移酶和α-1,6-糖苷酶活性。 4. 在糖原合成中,首先在葡萄糖焦磷酸化酶催化下由葡萄糖-1-磷酸和GTP形成激活中间产物UDP-葡萄糖。然后糖原合酶催化UDP-葡萄糖中的葡萄糖基转移至生长着的糖原分子的末端葡萄糖残基的C-4羟基。分支酶将一些α-1,4-糖苷键转为α-1,6-糖苷键增加非还原末端数目,以便合成多分支的糖原。多分支有利于糖原的快速降解。

36
5. 糖异生是一个由非糖物质,例如乳酸、氨基酸和甘油合成葡萄糖的途径。糖异生中的许多反应都是糖酵解反应的逆反应,但酵解中的磷酸烯醇式丙酮酸→丙酮酸、果糖-6-磷酸→果糖-1,6-二磷酸和葡萄糖→葡萄糖-6-磷酸三步不可逆反应是不能利用的。糖异生途径是通过另外四个反应克服这三个高能障不可逆反应的。 在线粒体中在丙酮酸羧化酶催化下丙酮酸羧化生成草酰乙酸,然后经苹果酸进入到细胞质后重新转化为草酰乙酸,在磷酸烯醇式丙酮酸羧化激酶催化下草酰乙酸脱羧和磷酸化生成磷酸烯醇式丙酮酸。另外两个反应是分别由果糖-1,6-二磷酸酶和葡萄糖-6-磷酸酶催化的果糖-1,6-二磷酸和葡萄糖-6-磷酸的水解反应。

37
糖代谢 Over 了! [希望大家认真掌握其要点]
![糖代谢 Over 了! [希望大家认真掌握其要点]](http://slidesplayer.com/slide/11533957/62/images/37/%E7%B3%96%E4%BB%A3%E8%B0%A2+Over+%E4%BA%86%EF%BC%81+%5B%E5%B8%8C%E6%9C%9B%E5%A4%A7%E5%AE%B6%E8%AE%A4%E7%9C%9F%E6%8E%8C%E6%8F%A1%E5%85%B6%E8%A6%81%E7%82%B9%5D.jpg "糖代谢 Over 了! [希望大家认真掌握其要点]")
Similar presentations
 反应部位 器官,细胞内定位 生理意义 代谢调节 主要调节点,主要变构抑制剂、变构激活剂 各代谢途径之间的联系和调控.>")
 CO.>")




 合成代谢------需要能量 物质代谢 能量代谢>")






的作用下进行的。 重点:物质代谢途径的相互联系,酶活性的调节。>")





