Download presentation
Presentation is loading. Please wait.

1
核酸 ◆第一节 核酸通论 ◆第二节 核酸的组成 ◆第三节 核酸的结构 ◆第四节 核酸的性质

2
Nucleic Acid 第二章 核酸

3
第一节、核酸通论 一、核酸的发现和研究简史 1 、早期的发现 2 、核酸化学的研究 3、DNA 双螺旋结构模型的提出建立了统一的分子生物学
5 、当前生命科学已进入后基因组的时代

4
(一)、早期的发现 1868 - 69 F. Miescher从脓细胞核中提出含 磷量高的 核素(nuclein),其后从鲑鱼精子中提取出鱼精蛋白和核素。 Hertwig提出核素可能负责受精和遗传。 1889 R. Altmann从酵母和动物组织中提取出不含蛋白质的核酸。 Kossel和A. Neumann 从胸腺中提取 核酸,表明胸腺核酸与酵母核酸不同。 Hammars 证明酵母核酸中的糖是戊糖。

5
(二)、核酸化学的研究 1902 E. Fischer研究糖和嘌呤而获诺贝尔化学奖。 核酸中的嘌呤和嘧啶主要由Kossel等人所鉴 定。
P. A. Levene 和 W. A. 等人鉴定 D-核糖。 Kossel 因核酸化学研究中的成就而获诺贝尔 生理学奖。 1912 P. A. Levene 认为核酸由四种核苷酸等量聚 合而成。 1929 Levene 确定胸腺核酸中的糖是 D-2-脱氧核糖。

6
(三)、核酸化学的研究 Levene 在 “Nucleic 酸” 一书中仍主张核酸 分为动物核酸( zoonucleic 酸)和植物核 酸 ( phytonucleic 酸)。 1933 J. Brachet发展了组织化学的方法。 1938 T. Caspersson显微紫外分光光度技术。 1943 J. N. Davidson 用化学分析方法支持上述组 织化学和紫外分光光度法得出的结论:动、 植物组织均含有两类核酸。 E. Chargaff测定了DNA碱基,提出Chargaff 法则。

7
(四)、DNA 双螺旋结构模型的提出建立了统一的分子生物学
T. Avery 肺炎双球菌转化实验。 Hershey证明噬菌体感染时只有 DNA 进入细菌 细胞。 J. D. Watson 和 F。 H. C。 Crick提出DNA双螺旋结构模型。1962年与M. H. F. Wilkins 共获诺贝尔生理学奖。 1969 年噬菌体小组的 M. Delbrük、A. D. Hershey和S. E。 Luria也获诺贝尔奖。 M. Meselson和F. W. Stahl用超速离心同位素 标记的DNA,证明了DNA的双螺旋结构。 F. H. C.Crick提出中心法则。

8
(五).DNA 双螺旋结构模型的提出建立了统一的分子生物学
1959 S. Ochoa和A. Kornberg 由于酶促合成 RNA 和 DNA 而获诺贝尔奖。 1960 - 61 F. Jacob和J. Monod 发现 mRNA 并提出操纵子学说, 1965 年与 P. Rous一起获诺贝尔奖。 1965 M. W. Nirenberg 等破译了遗传密码 1968 年R. W. Holley .H. G. Khorana 和 M. W. Nirenburg 共获诺贝尔 奖。 1970 H. Temin和D. 巴尔的摩发现逆转录病毒。

9
(六)、DNA 重组技术带来生物技术重大突破
1970 H. O. 史密斯和 K. W. Wilcox 发现类型 Ⅱ限制 性内切酶。 1971 K. Danna和D. Nathans 作出 SV40 DNA 的限 制性图谱 1978 年诺贝尔化学奖授予发现。 限制酶的 W. Arber、H. O. Smith和D. Nathans 。 1972 P. Berg与其工作者将SV40 DNA与λ噬菌体 DNA重组。 1973 S. Cohen等获得第1个DNA重组体克隆。 1975 F. Sanger和A. R. Coulson发明酶法DNA快速 测序。

10
(七)、DNA 重组技术带来生物技术重大突破
1977. M. Maxam 和 W. Gilbert发明化学DNA测序。 1980年 Berg、Gilbert和Sanger共获诺贝尔化学奖。 1985 K Mullis发明PCR技术。1993年与发明定位诱变的M. 史密斯共获诺贝尔化学奖。 DNA 重组技术引起分子生物学第二次革命,使生命科学进入大科学工程的时代,并带动了生物技术产业的迅猛发展。

11
(八)、当前生命科学已进入后基因组的时代
1990 - 2005 年人类基因组计划提前完成,科学家们提出了后基因组时代(在 genome 之後的时代)的设想,于是产生了功能基因组学(功能的 genomics )和结构基因组学(结构的 genomics )的新学科由于基因产物需要经过一系列加工修饰才能执行功能,基因产物间的相互作用十分复杂,这些过程只从基因组本身进行研究是不足以揭示细胞功能的,因此又提出了蛋白质组学( proteomics )和 RNA 组学( RNomics )的计划。 生物化学与分子生物学发展的主要方向有 3 个:结构分子生物学、神经分子生物学、以及发育分子生物学 21 世纪这些领域将会取得巨大成果。
的设想,于是产生了功能基因组学(功能的 genomics )和结构基因组学(结构的 genomics )的新学科由于基因产物需要经过一系列加工修饰才能执行功能,基因产物间的相互作用十分复杂,这些过程只从基因组本身进行研究是不足以揭示细胞功能的,因此又提出了蛋白质组学( proteomics )和 RNA 组学( RNomics )的计划。 生物化学与分子生物学发展的主要方向有 3 个:结构分子生物学、神经分子生物学、以及发育分子生物学 21 世纪这些领域将会取得巨大成果。")
12
二、 核酸的分类和组成 (一)、核酸的分类 核酸分为两大类. 脱氧核糖核酸(DNA) Deoxyribonucleic Acid
核糖核酸(RNA) Ribonucleic Acid。
 Ribonucleic Acid。")
13
1、脱氧核糖核酸(DNA) DNA分子含有生物物种的所有遗传信息,分子量一般都很大。
病毒。

14
2、核糖核酸(RNA) RNA主要是负责DNA遗传信息的翻译和表达,分子量要比DNA小得多。RNA为单链分子。少数RNA病毒RNA为双链分子。主要存在于细胞核、细胞质。
 RNA主要是负责DNA遗传信息的翻译和表达,分子量要比DNA小得多。RNA为单链分子。少数RNA病毒RNA为双链分子。主要存在于细胞核、细胞质。")
15
RNA的类别 根据RNA的功能,可以分为 mRNA、tRNA、rRNA、hnRNA、snRNA、asRNA。

16
Messenger RNA (1)、mRNA (信使RNA) 约占总RNA的5%。 不同细胞的mRNA的链长和分子量差异很大。
它的功能是将DNA的遗传信息传递到蛋白质合成基地 – 核糖核蛋白体。

17
Transfer RNA (2)tRNA (转移RNA) 约占总RNA的10-15%。
它在蛋白质生物合成中起翻译氨基酸信息,并将相应的氨基酸转运到核糖核蛋白体的作用。 已知每一个氨基酸至少有一个相应的tRNA。 RNA分子的大小很相似,链长一般在73-78个核苷酸之间。

18
Ribosome RNA (3)rRNA (核糖体RNA) 约占全部RNA的80%, 是核糖核蛋白体的主要组成部分。
返回

19
第二节 核酸的组成 核酸(DNA和RNA)是一种线性多聚核苷酸,它的基本结构单元是核苷酸。 核苷酸本身由核苷和磷酸组成,
而核苷则由戊糖和碱基形成 DNA与RNA结构相似,但在组成成份上略有不同。

20
一.核苷酸 1、组成核酸的碱基 腺嘌呤Adenine

21
核苷酸 1、组成核酸的碱基 鸟嘌呤guanine

22
Structure A and G

23
核苷酸 1、组成核酸的碱基 尿嘧啶uracil

24
核苷酸 1、组成核酸的碱基 胞嘧啶cytosine

25
1.核苷酸 1、组成核酸的碱基 胸腺嘧啶thymine

26
Structure of T and C

27
核苷酸 2、碱基的结构特征 碱基都具有芳香环的结构特征。嘌呤环和嘧啶环均呈平面或接近于平面的结构。
碱基的芳香环与环外基团可以发生酮式—烯醇式或胺式—亚胺式互变异构。

28
胺式 亚胺式互变异构

29
酮式烯醇式互变异构

30
核苷酸 碱基的结构特征 嘌呤碱和嘧啶碱分子中都含有共轭双键体系,在紫外区有吸收(260 nm左右)。
。")
31
核苷酸 3、戊糖 组成核酸的戊糖有两种。DNA所含的糖为β-D-2-脱氧核糖;RNA所含的糖则为β-D-核糖。

32
1.核苷酸 4、核苷 nucleoside 糖与碱基之间的C-N键,称为C-N糖苷键。

33
核苷酸 5、核苷酸nucleotide 核苷酸是核苷的磷酸酯。作为DNA或RNA结构单元的核苷酸分别是5′-磷酸-脱氧核糖核苷和5′-磷酸-核糖核苷。

34
核苷酸 6、修饰成分 核酸中也存在一些不常见的稀有碱基。稀有碱基的种类很多,大部分是上述碱基的甲基化产物。

35
二.核苷酸的衍生物 1、 ATP (腺嘌呤核糖核苷三磷酸) ATP是生物体内分布最广和最重要的一种核苷酸衍生物。它的结构如下:
 ATP是生物体内分布最广和最重要的一种核苷酸衍生物。它的结构如下:")
36
ATP的性质 ATP 分子的最显著特点是含有两个高能磷酸键。ATP水解时, 可以释放出大量自由能。

37
2、GTP (鸟嘌呤核糖核苷三磷酸) GTP是生物体内游离存在的另一种重要的核苷酸衍生物。它具有ATP 类似的结构, 也是一种高能化合物。
GTP主要是作为蛋白质合成中磷酰基供体。在许多情况下, ATP 和 GTP 可以相互转换。

38
3、cAMP 和 cGMP cAMP(3’,5’- 环腺嘌呤核苷一磷酸)和 cGMP( 3’,5’-环鸟嘌呤核苷一磷酸)的主要功能是作为细胞之间传递信息的信使。 cAMP 和 cGMP 的环状磷酯键是一个高能键。在 pH 7.4 条件下, cAMP 和 cGMP 的水解能约为43.9 kj /mol,比 ATP 水解能高得多。

39
三.多聚核苷酸 多聚核苷酸是通过核苷酸的5’-磷酸基与另一分子核苷酸的C3’-OH形成磷酸二酯键相连而成的链状聚合物。
由脱氧核糖核苷酸聚合而成的称为DNA链; 由核糖核苷酸聚合而成的则称为RNA链。

40
多聚核苷酸的特点 在多聚核苷酸中,两个核苷酸之间形成的磷酸二酯键通常称为5′—3′磷酸二酯键。
多聚核苷酸链一端的C5′带有一个自由磷酸基,称为5′-磷酸端(常用5’-P表示);另一端C3’带有自由的羟基,称为3′-羟基端(常用3’-OH表示)。 多聚核苷酸链具有方向性,当表示一个多聚核苷酸链时,必须注明它的方向是5′→3′或是3′→5′。
;另一端C3’带有自由的羟基,称为3′-羟基端(常用3’-OH表示)。 多聚核苷酸链具有方向性,当表示一个多聚核苷酸链时,必须注明它的方向是5′→3′或是3′→5′。")
41
在多聚核苷酸(DNA或RNA)链中,由于构成核苷酸单元的戊糖和磷酸基是相同的,体现核苷酸差别的实际上只是它所带的碱基,所以多聚核苷酸链结构也可表示为:
方向性 A C G T 3' 3' 3' 3' P P P P P 5' 5' 5' 5'

42
在讨论有关核酸问题时,一般只关心其中碱基的种类和顺序,所以上式可以进一步简化为:
5′PAPCPGPCPTPGPTPA 3′ 或5′ ACGCTGTA 3′ 返回

43
第三节 核酸的结构 一、核酸的一级结构 多聚核苷酸是由四种不同的核苷酸单元按特定的顺序组合而成的线性结构聚合物,因此,它具有一定的核苷酸顺序,即碱基顺序。 核酸的碱基顺序是核酸的一级结构。 DNA的碱基顺序本身就是遗传信息存储的分子形式。生物界物种的多样性即寓于DNA分子中四种核苷酸千变万化的不同排列组合之中。 而mRNA(信息RNA)的碱基顺序,则直接为蛋白质的氨基酸编码,并决定蛋白质的氨基酸顺序。
的碱基顺序,则直接为蛋白质的氨基酸编码,并决定蛋白质的氨基酸顺序。")
44
(一)、RNA一级结构的特点 RNA一级结构研究最多的是tRNA、rRNA以及一些小分子的RNA。其中 tRNA分子具有以下特点: 分子量25000左右,大约由70-90个核苷酸组成,沉降系数为4S左右。 分子中含有较多的修饰成分。 3'-末端都具有CpCpAOH的结构。

45
1、mRNA一级结构的特点 真核细胞mRNA的3‘-末端有一段长达200个核苷酸左右的聚腺苷酸(polyA),称为 “尾结构” ,5’ -末端有一个甲基化的鸟苷酸,称为 “帽结构 ” 。
,称为 尾结构 ,5’ -末端有一个甲基化的鸟苷酸,称为 帽结构 。")
46
极大多数真核细胞mRNA在3‘-末端有一段长约200核苷酸的polyA。polyA是在转录后经polyA聚合酶的作用而添加上去的。原核生物的mRNA一般无polyA,但某些病毒mBNA也有3’-polyA,polyA可能有多方面功能,与mRNA从细胞核到细胞质的转移有关;与mRNA的半寿期有关,新合成的mRNA,.polyA链较长,而衰老的mRNA,polyA链缩短

47
动物细胞核糖体rRNA有四类:5SrRNA,5
动物细胞核糖体rRNA有四类:5SrRNA,5.8SrRNA,18SrRNA,28SRNA。许多rRNA的一级结构及由一级结构推导 出来的二级结构都已阐明,但是对许多rRNA的功能迄今仍不十分清楚。

48
2、RNA的高级结构特点 RNA是单链分子,因此,在RNA分子中,并不遵守碱基种类的数量比例关系,即分子中的嘌呤碱基总数不一定等于嘧啶碱基的总数。 RNA分子中,部分区域也能形成双螺旋结构,不能形成双螺旋的部分,则形成突环。这种结构可以形象地称为“发夹型”结构。

49
在RNA的双螺旋结构中,碱基的配对情况不象DNA中严格。G 除了可以和C 配对外,也可以和U 配对。G-U 配对形成的氢键较弱。不同类型的RNA, 其二级结构有明显的差异。
tRNA中除了常见的碱基外,还存在一些稀有碱基,这类碱基大部分位于突环部分.

50
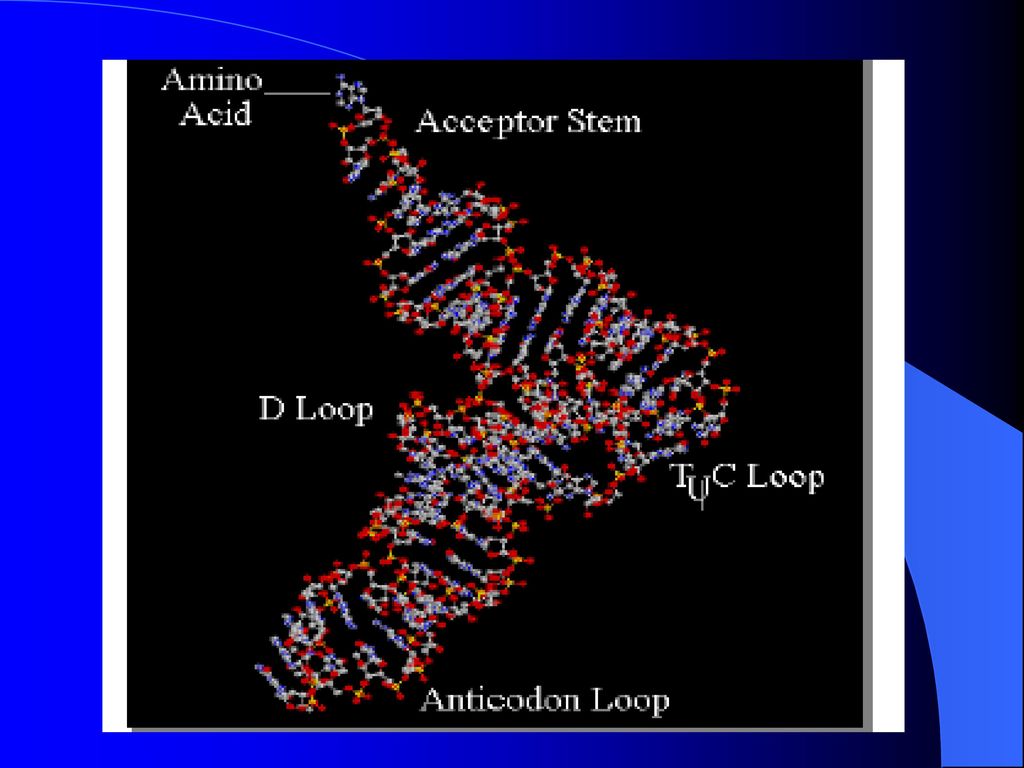
(1)、tRNA的二级结构 A、tRNA的二级结构
tRNA的二级结构都呈” 三叶草” 形状,在结构上具有某些共同之处,一般可将其分为五臂四环:包括氨基酸接受区、反密码区、二氢尿嘧啶区、TC区和可变区。除了氨基酸接受区外,其余每个区均含有一个突环和一个臂。

51
(1)氨基酸接受区 包含有tRNA的3’-末端和5’-末端, 3’-末端的最后3个核苷酸残基都是CCA,A为核苷。氨基酸可与其成酯,该区在蛋白质合成中起携带氨基酸的作用。 (2)反密码区 与氨基酸接受区相对的一般含有7个核苷酸残基的区域,其中正中的3个核苷酸残基称为反密码
氨基酸接受区 包含有tRNA的3’-末端和5’-末端, 3’-末端的最后3个核苷酸残基都是CCA,A为核苷。氨基酸可与其成酯,该区在蛋白质合成中起携带氨基酸的作用。 (2)反密码区 与氨基酸接受区相对的一般含有7个核苷酸残基的区域,其中正中的3个核苷酸残基称为反密码")
52
3)二氢尿嘧啶区 该区含有二氢尿嘧啶。 (4) TC区 该区与二氢尿嘧啶区相对, 假尿嘧啶核苷—胸腺嘧啶核糖核苷环(TC)由7个核苷酸组成,通过由5对碱基组成的双螺旋区(TC臂)与tRNA的其余部分相连。除个别例外,几乎所有tBNA在此环中都含有TC 。 (5)可变区 位于反密码区与TC区之间,不同的tRNA该区变化较大。
可变区. 位于反密码区与TC区之间,不同的tRNA该区变化较大。")
53
(2)、tRNA的三级结构 在三叶草型二级结构的基础上,突环上未配对的碱基由于整个分子的扭曲而配成对,目前已知的tRNA的三级结构均为倒L型
、tRNA的三级结构 在三叶草型二级结构的基础上,突环上未配对的碱基由于整个分子的扭曲而配成对,目前已知的tRNA的三级结构均为倒L型")
55
二、DNA的二级结构 1953年,J. Watson和F. Crick 在前人研究工作的基础上,根据DNA结晶的X-衍射图谱和分子模型,提出了著名的DNA双螺旋结构模型,并对模型的生物学意义作出了科学的解释和预测。

56
Watson和Crick的著名论文全文

57
1.DNA双螺旋结构的特点 DNA分子由两条DNA单链组成。 DNA的双螺旋结构是分子中两条DNA单链之间基团相互识别和作用的结果。

58
DNA双螺旋结构的要点 (1)DNA分子由两条多聚脱氧核糖核苷酸链(简称DNA单链)组成。两条链沿着同一根轴平行盘绕,形成右手双螺旋结构。螺旋中的两条链方向相反,即其中一条链的方向为5′→3′,而另一条链的方向为3′→5′。
DNA分子由两条多聚脱氧核糖核苷酸链(简称DNA单链)组成。两条链沿着同一根轴平行盘绕,形成右手双螺旋结构。螺旋中的两条链方向相反,即其中一条链的方向为5′→3′,而另一条链的方向为3′→5′。")
59
DNA双螺旋结构的要点 (2)嘌呤碱和嘧啶碱基位于螺旋的内侧,磷酸和脱氧核糖基位于螺旋外侧。碱基环平面与螺旋轴垂直,糖基环平面与碱基环平面成90°角。
嘌呤碱和嘧啶碱基位于螺旋的内侧,磷酸和脱氧核糖基位于螺旋外侧。碱基环平面与螺旋轴垂直,糖基环平面与碱基环平面成90°角。")
60
DNA双螺旋结构的要点 (3)螺旋横截面的直径约为2 nm,每条链相邻两个碱基平面之间的距离为3.4 nm,每10个核苷酸形成一个螺旋,其螺矩(即螺旋旋转一圈)高度为3.4 nm。
螺旋横截面的直径约为2 nm,每条链相邻两个碱基平面之间的距离为3.4 nm,每10个核苷酸形成一个螺旋,其螺矩(即螺旋旋转一圈)高度为3.4 nm。")
61
DNA双螺旋结构的要点 (4)两条DNA链相互结合以及形成双螺旋的力是链间的碱基对所形成的氢键。碱基的相互结合具有严格的配对规律,即腺嘌呤(A)与胸腺嘧啶(T)结合,鸟嘌呤(G)与胞嘧啶(C)结合,这种配对关系,称为碱基互补。A和T之间形成两个氢键,G与C之间形成三个氢键。 在DNA分子中,嘌呤碱基的总数与嘧啶碱基的总数相等。

62
2.DNA双螺旋的稳定性 DNA双螺旋结构在生理条件下是很稳定的。 维持这种稳定性的因素包括:两条DNA链之间形成的氢键;
由于双螺旋结构内部形成的疏水区,消除了介质中水分子对碱基之间氢键的影响;介质中的阳离子(如Na+、K+和Mg2+)中和了磷酸基团的负电荷,降低了DNA链之间的排斥力、范德华引力等。 改变介质条件和环境温度,将影响双螺旋的稳定性。
中和了磷酸基团的负电荷,降低了DNA链之间的排斥力、范德华引力等。 改变介质条件和环境温度,将影响双螺旋的稳定性。")
63
Francis Crick and James Watson
1962 Francis Crick, James Watson, and Maurice Wilkins receive the Nobel Prize for determining the molecular structure of DNA.

64
Structure of B-DNA and Z-DNA

66
DNA的三螺旋结构

67
DNA的三螺旋结构

68
Hoogsteen配对 在三股螺旋中,通常是一条同型寡聚核苷酸与寡聚嘧啶核苷酸—寡聚嘌呤核苷酸双螺旋的大沟结合,第三股的碱基可与Watson—Crick碱基对中的嘌呤碱形成Hoogsteen配对。第三股螺旋与寡聚嘌呤核苷酸同向平行。 Py.Pu*Py Py.Pu*Pu Py.Pu*rPu 返回

69
第四节、核酸的性质 一、含氮碱基的性质 嘌呤碱基和嘧啶碱基是核酸中最重要的组分。它们的性质对于核酸的性质和生物功能具有重要影响作用。
含氮碱基具有芳香环的结构特点。由于环上极性基团(如羰基、氨基等)的存在,碱基能够发生酮式—烯醇式或氨式—亚氨式的互变异构。因此,碱基既有芳香环的特性,也具有氨、酮和烯醇等相应的化学性质。
的存在,碱基能够发生酮式—烯醇式或氨式—亚氨式的互变异构。因此,碱基既有芳香环的特性,也具有氨、酮和烯醇等相应的化学性质。")
70
1、含氮碱基的碱性 嘌呤碱基和嘧啶碱基都具有弱碱性。 环内氨基的pKa值约为9.5。
碱基环外的氨基(存在于A、G和C)的碱性很弱,在生理pH条件下不能被质子化。这种情况与苯胺分子中的氨基相似。 因此嘌呤和嘧啶碱基的碱性主要是环内氨基的贡献。
的碱性很弱,在生理pH条件下不能被质子化。这种情况与苯胺分子中的氨基相似。 因此嘌呤和嘧啶碱基的碱性主要是环内氨基的贡献。")
71
2、碱基环氮原子的烷基化反应 在一定条件下,碱基环上的氮原子可以发生烷基化反应。
在同样条件下,U和T基本上不起反应。应用CH2N作为烷基化剂,则所有碱基都能发生上述反应。

72
二、核酸的性质 1.核酸的两性性质及等电点 与蛋白质相似,核酸分子中既含有酸性基团(磷酸基)也含有碱性基团(氨基),因而核酸也具有两性性质。
由于核酸分子中的磷酸是一个中等强度的酸,而碱性(氨基)是一个弱碱,所以核酸的等电点比较低。如DNA的等电点为4~4.5,RNA的等电点为2~2.5。 RNA的等电点比DNA低的原因,是RNA分子中核糖基2′-OH通过氢键促进了磷酸基上质子的解离。DNA没有这种作用。
是一个弱碱,所以核酸的等电点比较低。如DNA的等电点为4~4.5,RNA的等电点为2~2.5。 RNA的等电点比DNA低的原因,是RNA分子中核糖基2′-OH通过氢键促进了磷酸基上质子的解离。DNA没有这种作用。")
73
2、核酸的水解 (1)酸或碱水解 核酸分子中的磷酸二酯键可在酸或碱性条件下水解切断。
DNA和RNA对酸或碱的耐受程度有很大差别。例如,在0.1 mol/L NaOH溶液中,RNA几乎可以完全水解,生成2′-或3′-磷酸核苷;DNA在同样条件下则不受影响。这种水解性能上的差别,与RNA核糖基上2′-OH的邻基参与作用有很大的关系。在RNA水解时,2′-OH首先进攻磷酸基,在断开磷酯键的同时形成环状磷酸二酯,再在碱的作用形成水解产物。

74
(2)、酶水解 生物体内存在多种核酸水解酶。这些酶可以催化水解多聚核苷酸链中的磷酸二酯键。
以DNA为底物的DNA水解酶(DNases)和以RNA为底物的RNA水解酶(RNases)。 根据作用方式又分作两类:核酸外切酶和核酸内切酶。 核酸外切酶的作用方式是从多聚核苷酸链的一端(3′-端或5′-端)开始,逐个水解切除核苷酸;核酸内切酶的作用方式刚好和外切酶相反,它从多聚核苷酸链中间开始,在某个位点切断磷酸二酯键。 在分子生物学研究中最有应用价值的是限制性核酸内切酶。这种酶可以特异性的水解核酸中某些特定碱基顺序部位。
和以RNA为底物的RNA水解酶(RNases)。 根据作用方式又分作两类:核酸外切酶和核酸内切酶。 核酸外切酶的作用方式是从多聚核苷酸链的一端(3′-端或5′-端)开始,逐个水解切除核苷酸;核酸内切酶的作用方式刚好和外切酶相反,它从多聚核苷酸链中间开始,在某个位点切断磷酸二酯键。 在分子生物学研究中最有应用价值的是限制性核酸内切酶。这种酶可以特异性的水解核酸中某些特定碱基顺序部位。")
75
(3)、核酸的紫外吸收 在核酸分子中,由于嘌呤碱和嘧啶碱具有共轭双键体系,因而具有独特的紫外线吸收光谱,一般在260nm左右有最大吸收峰,可以作为核酸及其组份定性和定量测定的依据。
、核酸的紫外吸收 在核酸分子中,由于嘌呤碱和嘧啶碱具有共轭双键体系,因而具有独特的紫外线吸收光谱,一般在260nm左右有最大吸收峰,可以作为核酸及其组份定性和定量测定的依据。")
76
3、核酸的变性、复性与杂交 (1) 核酸的变性 核酸的变性是指核酸双螺旋区的多聚核苷酸链间的氢键断裂,变成单链结构的过程。变性核酸将失去其部分或全部的生物活性。核酸的变性并不涉及磷酸二酯键的断裂,所以它的一级结构(碱基顺序)保持不变。 能够引起核酸变性的因素很多。温度升高、酸碱度改变、甲醛和尿素等的存在均可引起核酸的变性。

77
RNA本身只有局部的双螺旋区,所以变性行为所引起的性质变化没有DNA那样明显。
利用紫外吸收的变化,可以检测核酸变性的情况。 例如,天然状态的DNA在完全变性后,紫外吸收(260 nm)值增加25-40%. 而RNA变性后,约增加1.1%。 这种现象称为增色效应.
值增加25-40%. 而RNA变性后,约增加1.1%。 这种现象称为增色效应.")
78
A、DNA变性的特征 DNA的变性过程是突变性的,它在很窄的温度区间内完成。因此,通常将引起DNA变性的温度称为融点,用Tm表示。
一般DNA的Tm值在70-85C之间。DNA的Tm值与分子中的G和C的含量有关。 G和C的含量高,Tm值高。因而测定Tm值,可反映DNA分子中G, C含量,可通过经验公式计算: (G+C)%=(Tm-69.3)X2.44
%=(Tm-69.3)X2.44.")
79
DNA变性

80
当DNA的稀盐溶液加热到80-100℃时,双螺旋结构即发生解体,两条链彼此分开,形成无规线团。
DNA变性后,它的一系列性质也随之发生变化,如紫外吸收(260 nm)值升高, 粘度降低等。
值升高, 粘度降低等。")
82
B、Tm vs GC Content

83
(2)、DNA复性
、DNA复性")
84
(3)、核酸的杂交 热变性的DNA单链,在复性时并不一定与同源DNA互补链形成双螺旋结构,它也可以与在某些区域有互补序列的异源DNA单链形成双螺旋结构。 这样形成的新分子称为杂交DNA分子。DNA单链与互补的RNA链之间也可以发生杂交。 核酸的杂交在分子生物学和遗传学的研究中具有重要意义。

85
核酸的杂交 返回

Similar presentations




:核蛋白体组成成分 转移 RNA ( tRNA ):转运氨基酸 信使 RNA ( mRNA ):蛋白质合成模板 不均一核 RNA ( hnRNA ):成熟 mRNA 的前体 小核 RNA ( snRNA ):>")








、DNA的一级结构 (二)、DNA的二级结构 (三)、DNA的三级结构.>")







