Download presentation
Presentation is loading. Please wait.

1
第一節 肌肉組織的特性與分類 第二節 骨骼肌 第三節 平滑肌 第四節 心肌
4 肌肉系統 第一節 肌肉組織的特性與分類 第二節 骨骼肌 第三節 平滑肌 第四節 心肌

2
學習目標 說明骨骼肌的構造。 說明肌肉收縮的肌絲滑動學說。 說明肌肉收縮的能量來源。
閱讀本章之後,讀者應能達到以下目標: 說明骨骼肌的構造。 說明肌肉收縮的肌絲滑動學說。 說明肌肉收縮的能量來源。 明白在興奮及收縮聯合中,鈣離子、旋轉素、旋轉肌球素所扮演的角色。 解釋引起肌肉疲乏的原因。 說明運動單位及其如何影響肌肉收縮。 說明白肌與紅肌的差異。 說明動作電位如何由神經傳導至肌肉纖維中,並引起肌肉的收縮。 描述平滑肌的構造與其收縮的調節。 說明肌纖維收縮的全或無定律。

3
第一節 肌肉組織的特性與分類 肌肉組織的特性 肌肉組織的分類

4
肌肉組織的特性 肌肉組織具下列四項基本生理特性: 肌肉具有收縮或肌小節變短的能力,即收縮性 (contractility)。
肌肉受到神經衝動興奮而引起反應的能力,即興奮性 (excitability)。 肌肉具有擴大伸展的能力,即伸展性 (extensibility)。 肌肉在收縮變短或擴大伸展後能恢復其原來大小形狀的能力,即為彈性 (elasticity)。
。 肌肉具有擴大伸展的能力,即伸展性 (extensibility)。 肌肉在收縮變短或擴大伸展後能恢復其原來大小形狀的能力,即為彈性 (elasticity)。")
5
由這些特性可知肌肉的基本功能之一-運動,而骨骼是提供運動時的槓桿原理,關節的活動度則影響著骨骼與骨骼之間的移動能力,移動的力量則來自於肌肉組織。

6
肌肉組織的分類 肌肉組織是由肌細胞(即肌纖維)所構成,可依構造上、功能位置以及神經控制來分類。 依構造分類 依功能位置分類 依神經控制分類
所構成,可依構造上、功能位置以及神經控制來分類。 依構造分類 依功能位置分類 依神經控制分類")
7
依構造分類 依構造上肌細胞可分為橫紋肌與平滑肌: 橫紋肌:骨骼肌與心肌的粗、細肌絲排列規律,形成明暗相間的橫紋。
平滑肌:內臟肌不具橫紋,故稱為平滑肌。

8
依功能位置分類 依功能位置可分為骨骼肌、心肌及平滑肌:
骨骼肌:幾乎附著於骨頭上,當步行或跑步時,下肢部分骨骼肌收縮可使腳踝、膝蓋與臀部移動。當站立或坐著時,肌肉收縮幫助維持身體的姿勢。而藉由骨骼肌的收縮亦可產生大量的熱,以維持體溫的恆定。

9
心肌:構成心臟壁,負責心臟的收縮,將血液由心臟唧出到大動脈,供給身體的各部位。
平滑肌:形成各種內臟壁,負責內臟的收縮,如:胃腸道的蠕動收縮使食物通過。

10
依神經控制分類 依神經控制可分為隨意肌與不隨意肌: 隨意肌:骨骼肌受體運動神經系統的支配,亦即受意識的控制,所以又稱為隨意肌。
不隨意肌:心肌和平滑肌(內臟肌)受自主神經系統的支配,不受意識的控制,所以稱為不隨意肌。
受自主神經系統的支配,不受意識的控制,所以稱為不隨意肌。")
11
第二節 骨骼肌 骨骼肌的構造 運動單位 肌絲滑動學說 收縮肌絲的分子特徵 肌絲滑動的橫橋循環 初始肌肉纖維長度與張力的關係 興奮及收縮聯合
第二節 骨骼肌 骨骼肌的構造 運動單位 肌絲滑動學說 收縮肌絲的分子特徵 肌絲滑動的橫橋循環 初始肌肉纖維長度與張力的關係 興奮及收縮聯合 興奮及收縮聯合 肌肉收縮的能量來源 肌肉收縮的型式 肌纖維收縮的全或無定律 骨骼肌纖維的種類 骨骼肌的神經控制

12
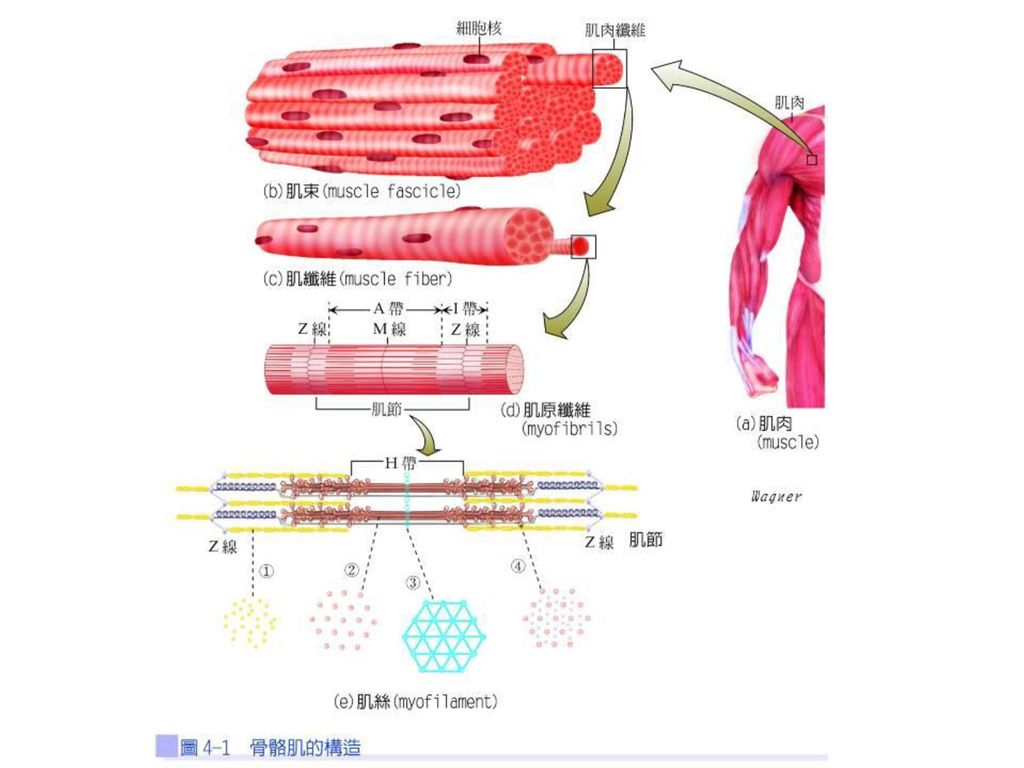
骨骼肌的構造 骨骼肌是由成束的肌束(muscle fascicle)構成,而肌束是由肌纖維(muscle fiber)構成,肌纖維是由肌原纖維(myofibrils)組成,肌原纖維則是由粗、細肌絲(myofilaments)組成(圖4-1)。
構成,而肌束是由肌纖維(muscle fiber)構成,肌纖維是由肌原纖維(myofibrils)組成,肌原纖維則是由粗、細肌絲(myofilaments)組成(圖4-1)。")
14
骨骼肌的肌纖維有許多細胞核(為多核細胞),肌纖維的長度平均為3公分,有的甚至可超過30公分,有的則短於0
骨骼肌的肌纖維有許多細胞核(為多核細胞),肌纖維的長度平均為3公分,有的甚至可超過30公分,有的則短於0.1公分,長度視部位而定。直徑一般約在10~100µm。肌纖維被一層特化的細胞膜包圍著,稱為肌漿膜(sarcolemma),其內的細胞質稱為肌漿(sarcoplasm),肌漿內含有許多細胞核、粒線體及平行排列的肌原纖維,直徑約1~2µm,長度則與肌纖維相同。
,肌纖維的長度平均為3公分,有的甚至可超過30公分,有的則短於0.1公分,長度視部位而定。直徑一般約在10~100µm。肌纖維被一層特化的細胞膜包圍著,稱為肌漿膜(sarcolemma),其內的細胞質稱為肌漿(sarcoplasm),肌漿內含有許多細胞核、粒線體及平行排列的肌原纖維,直徑約1~2µm,長度則與肌纖維相同。")
15
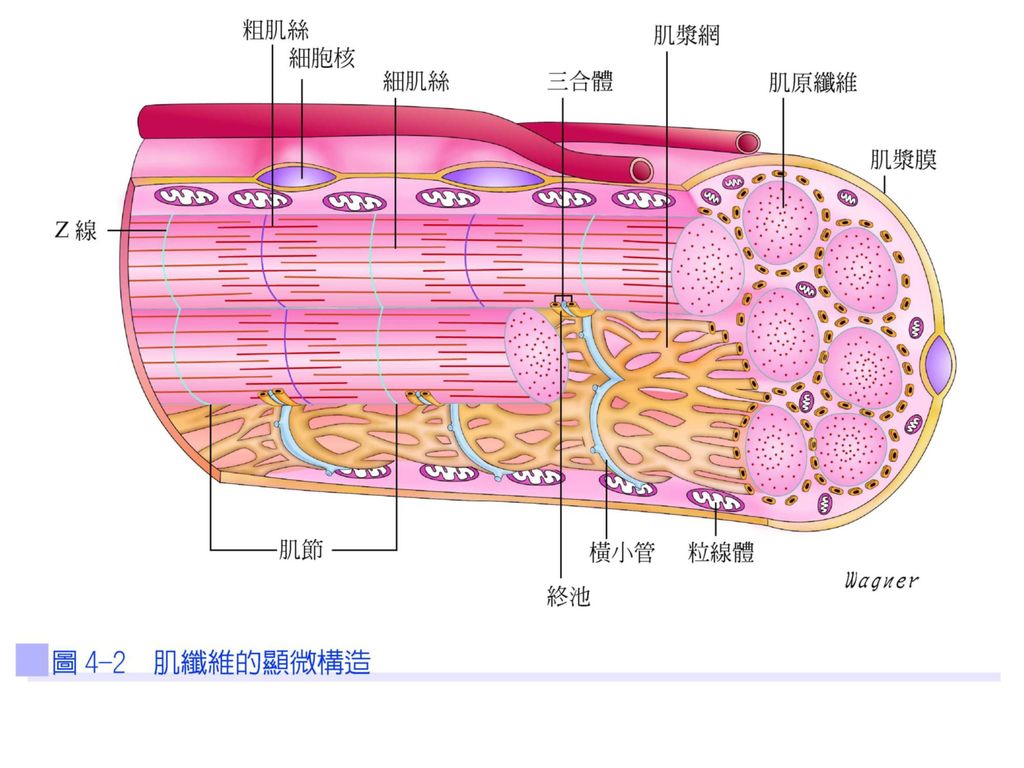
肌漿內的肌漿網(sarcoplasmic reticulum)為平行圍繞著每條肌原纖維的網狀組織,內儲存大量的鈣離子,動作電位會促使肌漿網釋放鈣離子來引起肌纖維的收縮。以垂直方向橫過肌漿網的是橫小管或稱T小管(transverse tubule;T tubule),是由肌漿膜延伸到肌纖維外。一個橫小管與其兩旁肌漿網的終池(側囊)(terminal cisterna)(lateral sac)組成三合體(triad)(圖4-2)。
為平行圍繞著每條肌原纖維的網狀組織,內儲存大量的鈣離子,動作電位會促使肌漿網釋放鈣離子來引起肌纖維的收縮。以垂直方向橫過肌漿網的是橫小管或稱T小管(transverse tubule;T tubule),是由肌漿膜延伸到肌纖維外。一個橫小管與其兩旁肌漿網的終池(側囊)(terminal cisterna)(lateral sac)組成三合體(triad)(圖4-2)。")
17
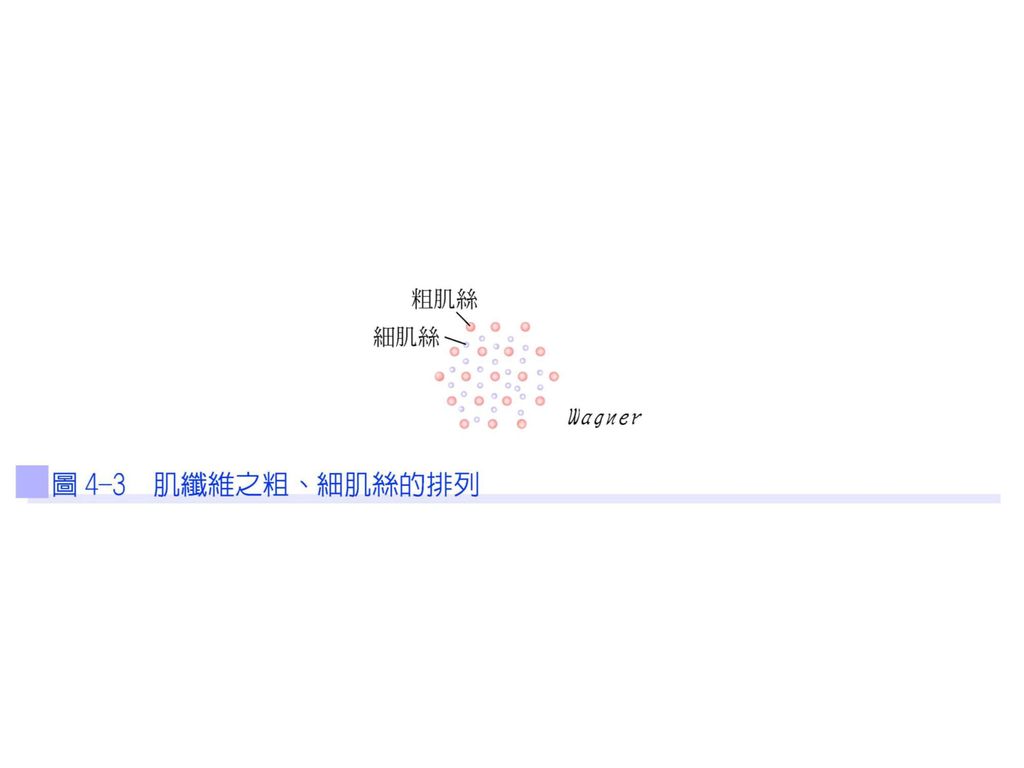
肌原纖維是由粗、細肌絲以規律的方式排列,故在顯微鏡下呈現明暗相間的橫紋,若從橫切面來看,每一條粗肌絲是由6條細肌絲以六角形狀包圍;每一條細肌絲是由3條粗肌絲以三角形狀包圍(圖4-3)。粗肌絲(thick myofilaments)是由肌凝蛋白(myosin)構成;細肌絲(thin myofilaments)是由肌動蛋白(actin)構成。
。粗肌絲(thick myofilaments)是由肌凝蛋白(myosin)構成;細肌絲(thin myofilaments)是由肌動蛋白(actin)構成。")
19
粗肌絲與細肌絲重疊而呈現較暗的部分,稱為A帶(A bands)或暗帶,A帶的長度等於粗肌絲的長度;只含細肌絲呈現較亮的部分稱為I帶(I bands)或稱亮帶,亦即位於兩條粗肌絲之間。穿過I帶的黑線稱為Z線(Z line),兩條Z線之間即為一個肌節(sarcomere),肌節為肌肉收縮的基本功能單位。A帶中只含粗肌絲的部分稱為H區(H zone),亦即位於兩條細肌絲之間;穿過H區的細線稱為M線(M line),M線為連接粗肌絲的中央部位(圖4-1)。
或暗帶,A帶的長度等於粗肌絲的長度;只含細肌絲呈現較亮的部分稱為I帶(I bands)或稱亮帶,亦即位於兩條粗肌絲之間。穿過I帶的黑線稱為Z線(Z line),兩條Z線之間即為一個肌節(sarcomere),肌節為肌肉收縮的基本功能單位。A帶中只含粗肌絲的部分稱為H區(H zone),亦即位於兩條細肌絲之間;穿過H區的細線稱為M線(M line),M線為連接粗肌絲的中央部位(圖4-1)。")
20
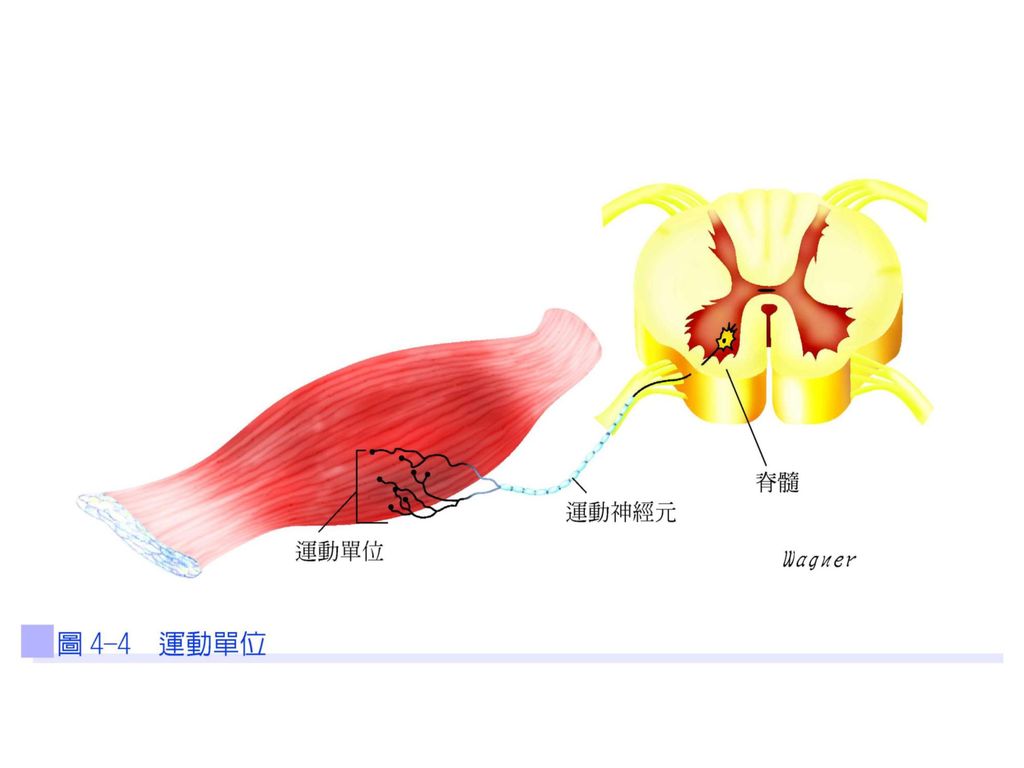
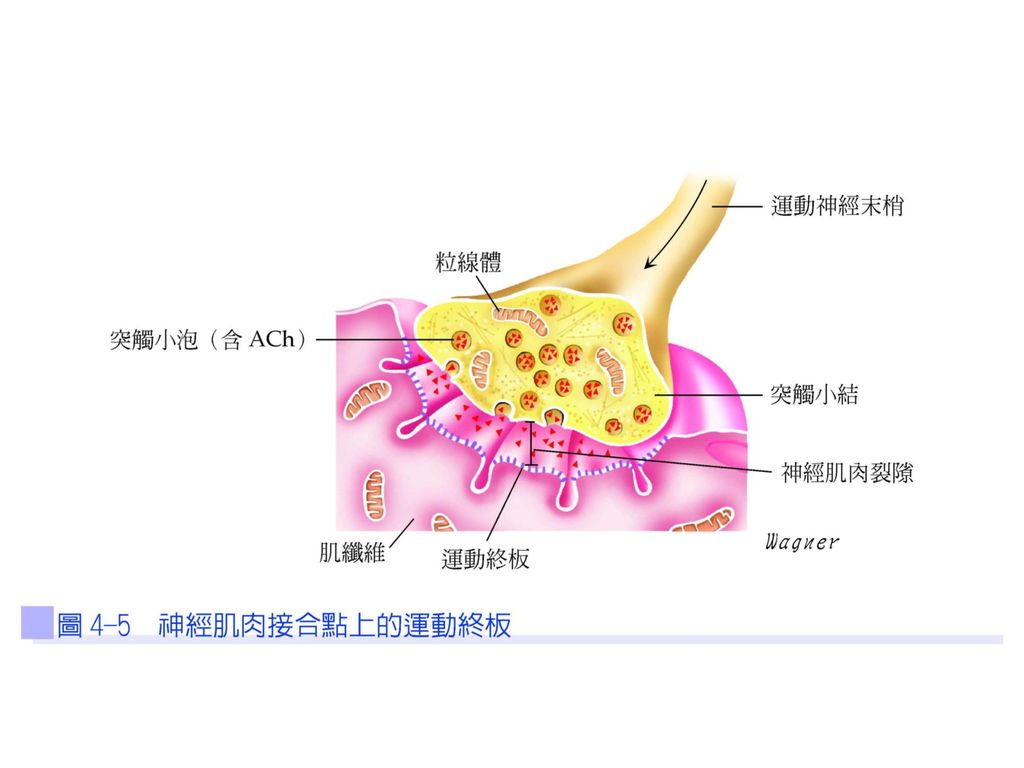
運動單位 同一運動神經細胞體、其神經軸突與所有其所支配的肌纖維,稱為一個運動單位(圖4-4)。當動作電位傳到運動神經元的軸突末梢時,突觸小泡會釋放出乙醯膽鹼(acetylcholine;ACh)到運動終板(圖4-5),亦即神經肌肉接合處,然後與肌纖維上的乙醯膽鹼接受器結合,並打開接受器所控制的離子通道,因而造成運動終板的局部去極化,稱為終板電位(end-plate potential;EPP)。
。當動作電位傳到運動神經元的軸突末梢時,突觸小泡會釋放出乙醯膽鹼(acetylcholine;ACh)到運動終板(圖4-5),亦即神經肌肉接合處,然後與肌纖維上的乙醯膽鹼接受器結合,並打開接受器所控制的離子通道,因而造成運動終板的局部去極化,稱為終板電位(end-plate potential;EPP)。")
23
肌肉收縮的強度與參與收縮的運動單位數目多寡有關。小的運動單位有較小的運動神經細胞體、較細的軸突,且支配較少的肌肉纖維,可控制精細動作,如眼部周圍控制眼球轉動的肌肉,其神經支配比例(innervation ratio)(運動神經元數目:肌肉纖維數目)平均約為1:23,可以控制眼球的轉動。
(運動神經元數目:肌肉纖維數目)平均約為1:23,可以控制眼球的轉動。")
24
但是大的運動單位有較大的運動神經細胞體、較粗的軸突,且支配較多的肌肉纖維,負責粗動作,如肱二頭肌,其神經支配平均約為1:1,000。

25
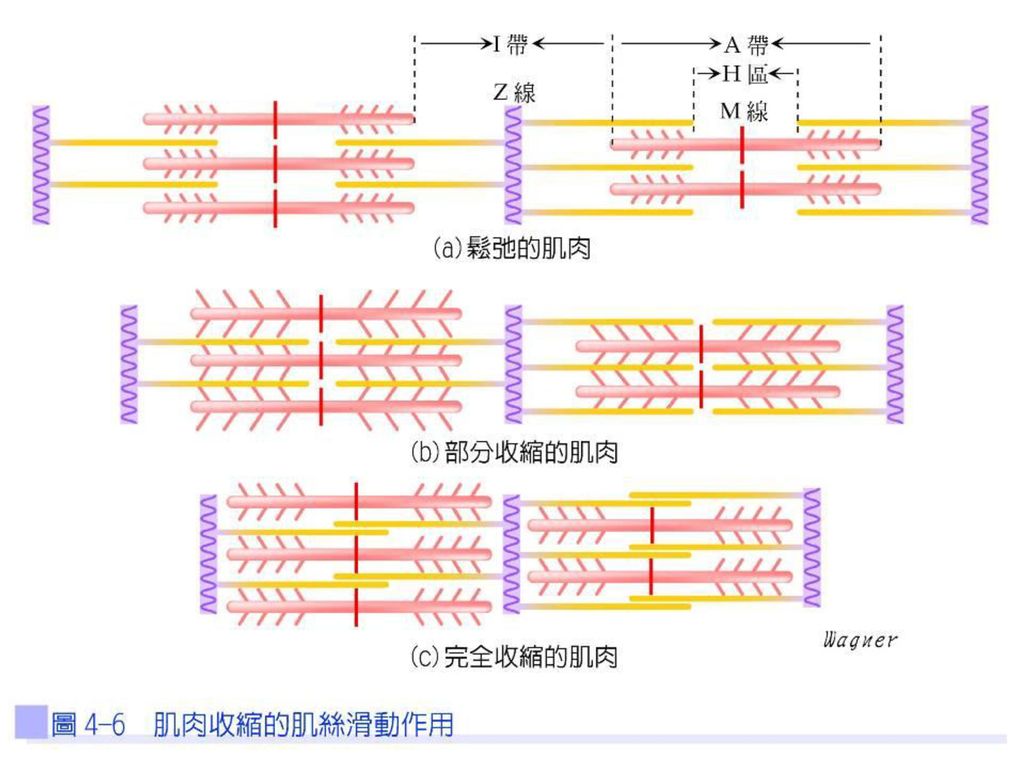
肌絲滑動學說 肌纖維收縮時,雖然長度變短,但是粗、細肌絲的長度並沒有改變,因為直接影響肌原纖維長度的,便是肌節的長短,也就是Z線之間的距離。當肌纖維收縮時,和粗肌絲相重疊的兩側細肌絲會滑向M線,使得肌節的長度變短,當每一肌節變短時,整條肌纖維也就跟著變短了。

26
粗、細肌絲在肌肉收縮時,長度皆沒有改變,造成肌節收縮的原因是因肌動蛋白絲彼此向內靠近,重疊部分較多;Z線也被肌動蛋白絲向內拉,使之接近肌凝蛋白絲末端,所以A帶的長度不變。相鄰的A帶互相靠近,介於其間之I帶則縮短,細肌絲向M線滑動的結果,H區也會跟著縮短(圖4-6)。
。")
28
收縮肌絲的分子特徵 粗肌絲 細肌絲

29
粗肌絲 粗肌絲亦稱肌凝蛋白絲(myosin filament),粗肌絲是由許多分子量約為480,000的肌凝蛋白分子所組成的,而肌凝蛋白分子則是由6個多胜肽鏈(polypeptide chain)所組成,包括:2個重鏈及4個輕鏈。
,粗肌絲是由許多分子量約為480,000的肌凝蛋白分子所組成的,而肌凝蛋白分子則是由6個多胜肽鏈(polypeptide chain)所組成,包括:2個重鏈及4個輕鏈。")
30
肌凝蛋白分成兩個部位: 尾部(tail):向著中央,形成粗肌絲的長軸。
頭部(head)向著兩端,形成球狀,又稱為橫橋(cross bridge),橫橋的移動為肌絲之間滑動的力量來源,具有2個重要位置
向著兩端,形成球狀,又稱為橫橋(cross bridge),橫橋的移動為肌絲之間滑動的力量來源,具有2個重要位置.")
31
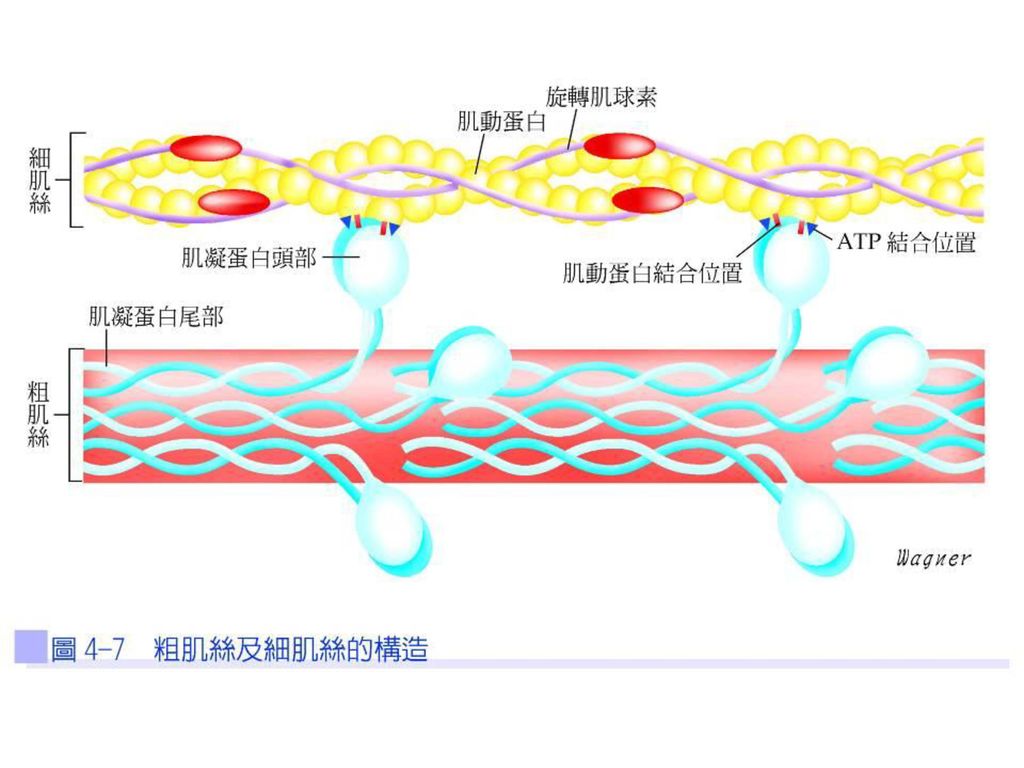
(1)ATP的結合位置(ATP binding site):可與ATP結合。橫橋頭部也是肌凝蛋白ATP水解酶(myosin ATPase),能將ATP分解為ADP和Pi(磷酸根),所釋放的能量可以活化橫橋使其能夠連接於肌動蛋白之上,這個反應是發生在橫橋與肌動蛋白結合之前;(2)肌動蛋白結合位置(actin binding site):可與肌動蛋白結合(圖4-7)。
ATP的結合位置(ATP binding site):可與ATP結合。橫橋頭部也是肌凝蛋白ATP水解酶(myosin ATPase),能將ATP分解為ADP和Pi(磷酸根),所釋放的能量可以活化橫橋使其能夠連接於肌動蛋白之上,這個反應是發生在橫橋與肌動蛋白結合之前;(2)肌動蛋白結合位置(actin binding site):可與肌動蛋白結合(圖4-7)。")
33
細肌絲 細肌絲又稱肌動蛋白絲(actin filament),其由三種蛋白質構成(圖4-8):
第一種為肌動蛋白(actin):主要為雙鏈的F型肌動蛋白分子(F-actin protein molecular)所構成,而每個雙鏈F型肌動蛋白螺旋體中的二個鏈是由300~400個球狀肌動蛋白(globular subunit)組成,又稱G-actin。
:主要為雙鏈的F型肌動蛋白分子(F-actin protein molecular)所構成,而每個雙鏈F型肌動蛋白螺旋體中的二個鏈是由300~400個球狀肌動蛋白(globular subunit)組成,又稱G-actin。")
35
第二種為旋轉肌球素(tropomyosin):其每一分子的旋轉肌球素分子量是70,000。在肌肉放鬆時,細肌絲上的肌凝蛋白結合位置會被一旋轉肌球素分子所遮蓋,造成細肌絲無法與粗肌絲結合,肌肉便無法收縮。
:其每一分子的旋轉肌球素分子量是70,000。在肌肉放鬆時,細肌絲上的肌凝蛋白結合位置會被一旋轉肌球素分子所遮蓋,造成細肌絲無法與粗肌絲結合,肌肉便無法收縮。")
36
第三種蛋白質為旋轉素(troponin):附著於旋轉肌球素之上,而非直接附著於肌動蛋白上,其由三個結合疏鬆的次單位所構成:(1)旋轉素I (troponin I)對肌動蛋白有很強的親和力;(2)旋轉素T (troponin T)對旋轉肌球素有很強的親和力;(3)旋轉素C (troponin C)對鈣離子有親和力。
:附著於旋轉肌球素之上,而非直接附著於肌動蛋白上,其由三個結合疏鬆的次單位所構成:(1)旋轉素I (troponin I)對肌動蛋白有很強的親和力;(2)旋轉素T (troponin T)對旋轉肌球素有很強的親和力;(3)旋轉素C (troponin C)對鈣離子有親和力。")
37
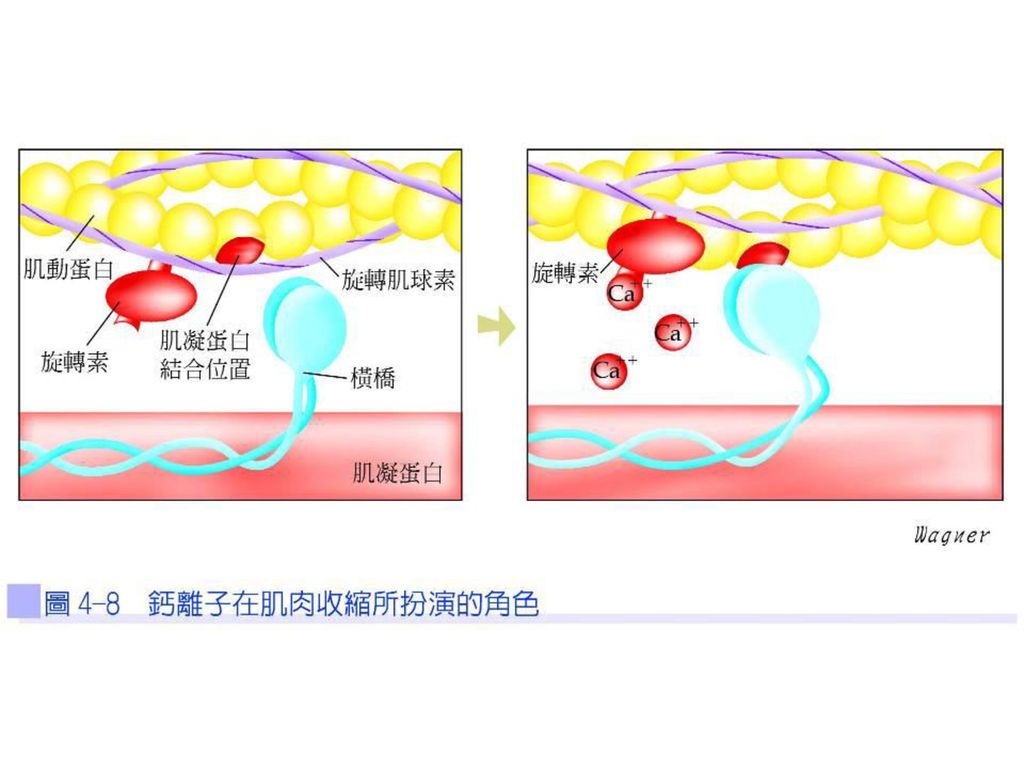
肌絲滑動的橫橋循環 肌纖維對於鬆弛(休息)狀態時,旋轉肌球素的位置遮蓋了肌動蛋白上的肌凝蛋白結合位置(myosin binding site),阻止了橫橋與肌動蛋白的結合,此時肌漿質中的鈣離子濃度是相當低的,但當肌纖維受到刺激時(即受到神經的支配而產生動作電位時),會使鈣離子濃度急速上升。
狀態時,旋轉肌球素的位置遮蓋了肌動蛋白上的肌凝蛋白結合位置(myosin binding site),阻止了橫橋與肌動蛋白的結合,此時肌漿質中的鈣離子濃度是相當低的,但當肌纖維受到刺激時(即受到神經的支配而產生動作電位時),會使鈣離子濃度急速上升。")
38
鈣離子與旋轉素C (troponin C)結合,導致旋轉素與旋轉肌球素產生結構上的改變並側移,便露出肌動蛋白的肌凝蛋白結合位置,橫橋才能與肌動蛋白結合(圖4-8),並產生力擊(power stroke)。
結合,導致旋轉素與旋轉肌球素產生結構上的改變並側移,便露出肌動蛋白的肌凝蛋白結合位置,橫橋才能與肌動蛋白結合(圖4-8),並產生力擊(power stroke)。")
39
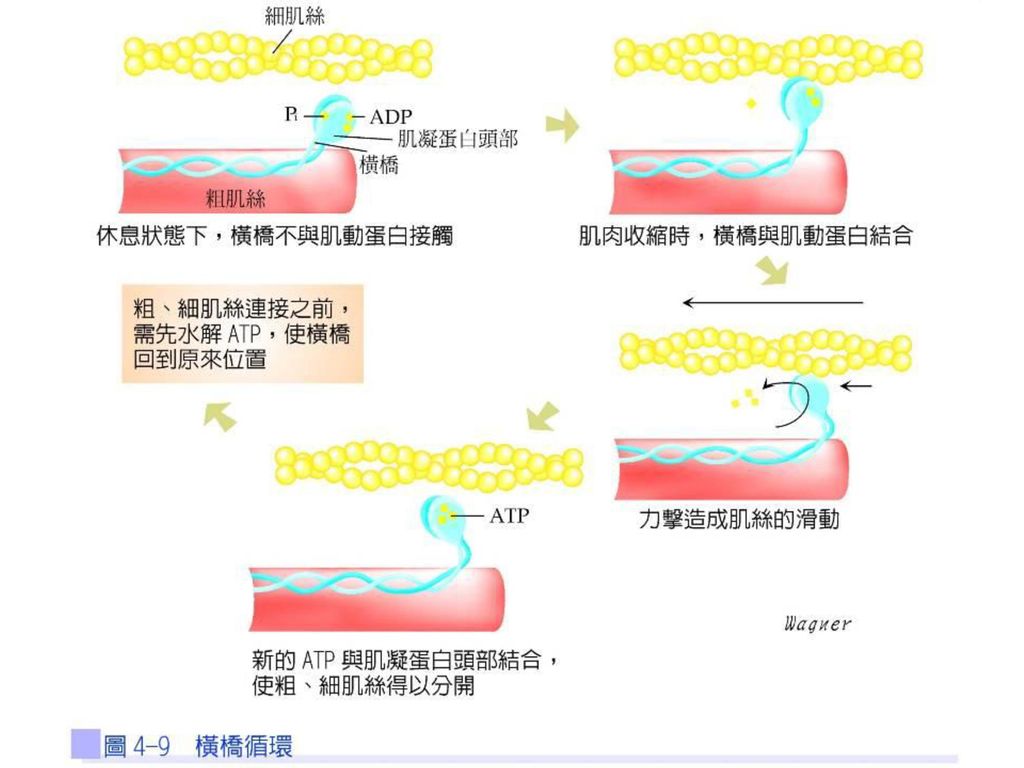
所謂力擊是當橫橋與肌動蛋白活化位置結合後,使頭部往肌凝蛋白絲中央傾斜,肌動蛋白絲受到牽引隨之同向移動,造成橫橋頭部從90度角傾斜成45度角。每一個橫橋都會與肌動蛋白絲結合,在任一瞬間與細肌絲結合的橫橋數目越多,則產生力擊的力量越大(圖4-9)。
。")
41
在產生力擊之前,必須先有ATP與肌凝蛋白的頭部結合。ATP水解成ADP+Pi後,肌凝蛋白始得與肌動蛋白結合(橫橋),接著Pi自肌凝蛋白脫離而產生力擊。此時,必須再有新的ATP與肌凝蛋白頭部結合,才能使粗、細肌絲分開,進入下一次的橫橋循環。
,接著Pi自肌凝蛋白脫離而產生力擊。此時,必須再有新的ATP與肌凝蛋白頭部結合,才能使粗、細肌絲分開,進入下一次的橫橋循環。")
42
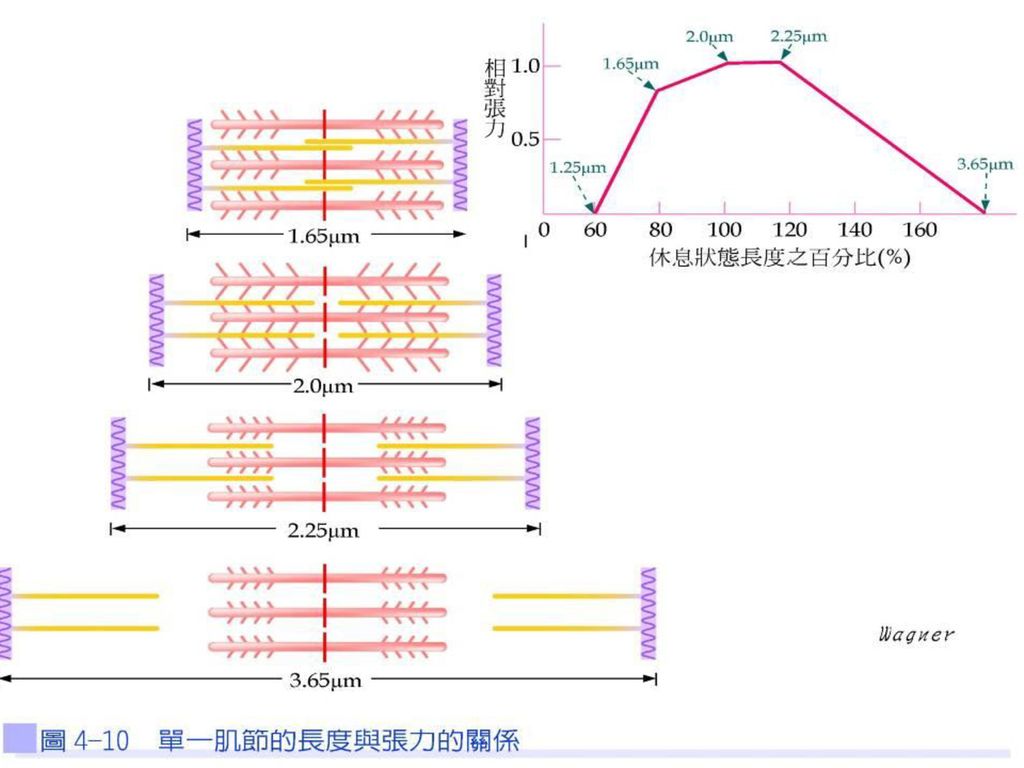
初始肌肉纖維長度與張力的關係 被刺激收縮的肌纖維數目、肌纖維的粗細以及肌纖維在鬆弛狀態時的長度等因素,會影響肌肉收縮強度。就橫紋肌而言,在最理想的鬆弛狀態長度時引發收縮,可使肌肉產生最大的收縮強度。若鬆弛狀態肌節過長,肌動蛋白與橫橋連接的數目減少,引發的收縮力相對變小。

43
反之,若肌節被縮短至原長度的60%時,會造成Z線與粗肌絲互相接觸,即粗、細肌絲已達最大重疊程度,此時肌肉無法再做進一步收縮力(圖4-10)。
。")
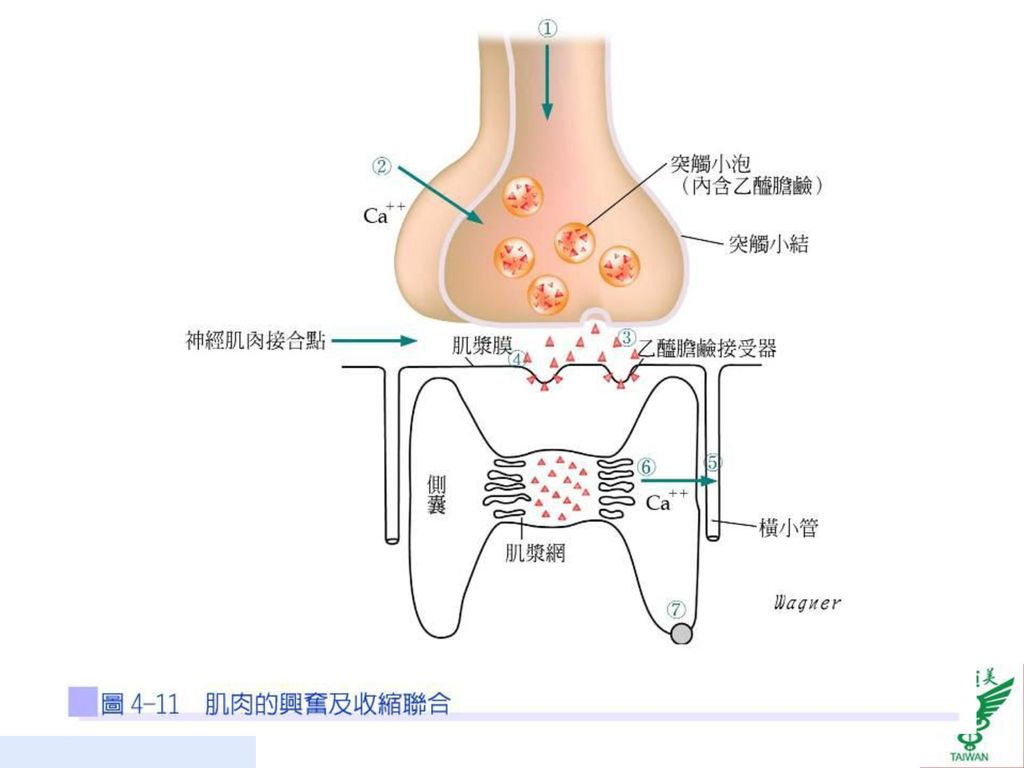
45
興奮及收縮聯合 骨骼肌纖維的靜止膜電位約為-90mV,動作電位持續時間約為4毫秒(msec),當運動神經元的動作電位抵達軸突末梢時,鈣離子進入突觸小結(synaptic knob),造成突觸小泡(synaptic vesicle)藉由胞吐作用(exocytosis)釋放出乙醯膽鹼,再與肌細胞膜上的乙醯膽鹼接受器結合,打開接受器所控制的離子通道,因而造成運動終板的局部去極化,稱為終板電位(end-plate potential;EPP),約為-50~-75 mV。
,當運動神經元的動作電位抵達軸突末梢時,鈣離子進入突觸小結(synaptic knob),造成突觸小泡(synaptic vesicle)藉由胞吐作用(exocytosis)釋放出乙醯膽鹼,再與肌細胞膜上的乙醯膽鹼接受器結合,打開接受器所控制的離子通道,因而造成運動終板的局部去極化,稱為終板電位(end-plate potential;EPP),約為-50~-75 mV。")
46
若其強度超過閾值,則引發肌肉的動作電位,動作電位經由T小管傳播到肌纖維的內部,肌漿網的終池釋放鈣離子到肌原纖維內,並與旋轉素C結合,使旋轉素及旋轉肌球素側移,因而露出肌動蛋白之肌凝蛋白結合位置(myosin binding site),粗肌絲的肌凝蛋白頭部之肌動蛋白結合位置與細肌絲的肌動蛋白之肌凝蛋白結合位置結合形成橫橋,肌凝蛋白頭部的ATP水解酶活化並水解ATP而產生能量,導致肌凝蛋白頭部擺動,造成粗、細肌絲滑動而產生肌肉收縮(圖4-11)。
,粗肌絲的肌凝蛋白頭部之肌動蛋白結合位置與細肌絲的肌動蛋白之肌凝蛋白結合位置結合形成橫橋,肌凝蛋白頭部的ATP水解酶活化並水解ATP而產生能量,導致肌凝蛋白頭部擺動,造成粗、細肌絲滑動而產生肌肉收縮(圖4-11)。")
49
肌肉收縮的能量來源 肌肉收縮的能量來源有五種方式,介紹如下:

50
在肌肉細胞中,磷酸肌酸(creatine phosphate)的濃度是ATP的三倍,可作為一個儲存高能磷酸根的儲備槽。在運動時,磷酸肌酸會在肌凝蛋白頭端和肌動蛋白結合位置發生水解,使ADP再形成ATP,而使收縮得以持續。
的濃度是ATP的三倍,可作為一個儲存高能磷酸根的儲備槽。在運動時,磷酸肌酸會在肌凝蛋白頭端和肌動蛋白結合位置發生水解,使ADP再形成ATP,而使收縮得以持續。")
51
肝醣為肝臟和骨骼肌細胞中含量特別豐富的碳水化合物,為細胞內葡萄糖的來源。當氧供應不足時,葡萄糖會被還原成乳酸(lactic acid),稱為無氧醣分解(anaerobic glycolysis),其產生的能量比高能磷酸根所產生的少許多,且乳酸會造成肌肉疲勞,但其不需氧的提供即可進行。
,稱為無氧醣分解(anaerobic glycolysis),其產生的能量比高能磷酸根所產生的少許多,且乳酸會造成肌肉疲勞,但其不需氧的提供即可進行。")
53
當有足夠氧存在時,葡萄糖可分解為二氧化碳和水,並釋出能量,稱為有氧醣分解(aerobic glycolysis)。約有80%(4/5)的乳酸會從骨骼肌擴散出,然後被送至肝臟,最後合成葡萄糖或肝醣,等待再使用。可是卻有20%(1/5)的乳酸累積在肌肉中,而造成肌肉疲勞,最後在骨骼肌中的乳酸亦會分解成二氧化碳及水,而這個轉化作用必須使用額外的氧,即氧債 (oxygen debt)。
。約有80%(4/5)的乳酸會從骨骼肌擴散出,然後被送至肝臟,最後合成葡萄糖或肝醣,等待再使用。可是卻有20%(1/5)的乳酸累積在肌肉中,而造成肌肉疲勞,最後在骨骼肌中的乳酸亦會分解成二氧化碳及水,而這個轉化作用必須使用額外的氧,即氧債 (oxygen debt)。")
55
骨骼肌也可自血液中攝取游離的脂肪酸(fatty acid),而將其氧化為二氧化碳及水。
當你做中度運動到激烈運動時,因為尚未有足夠的時間來增加氧氣的運輸,所以前45~90秒主要進行無氧呼吸,也稱為乳酸代謝。如果運動是中度的,在運動初期兩分鐘之後,有氧細胞呼吸開始提供肌肉所需要的能量。

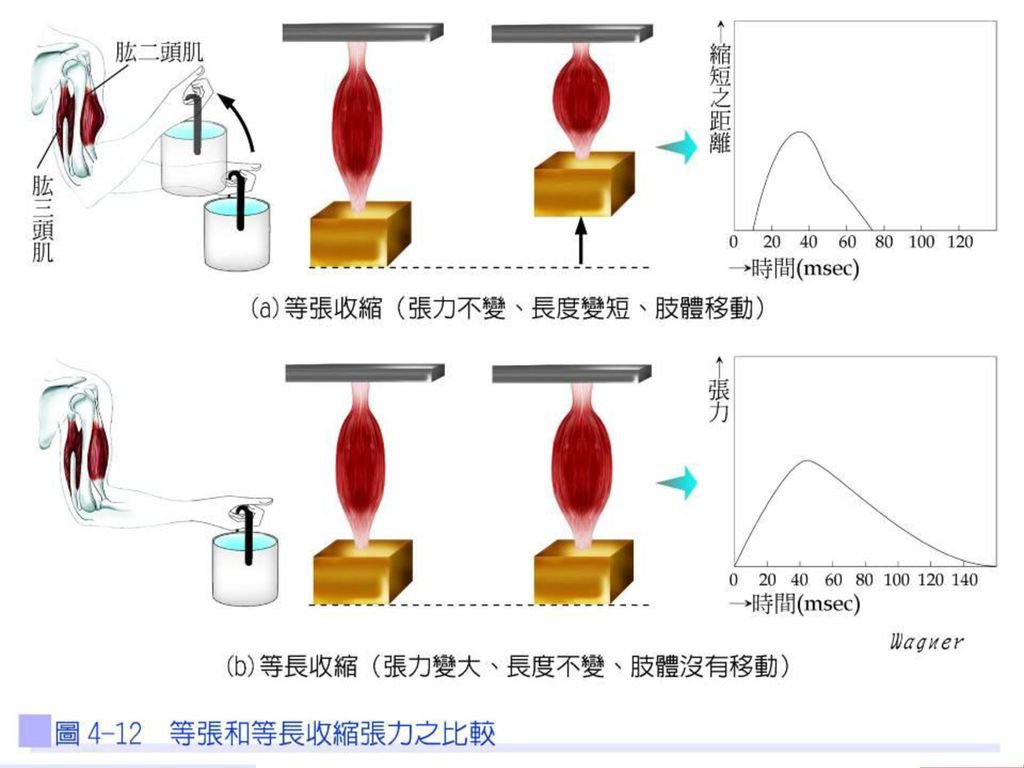
56
肌肉收縮的型式 依肌纖維長度是否改變可分成等長收縮及等張收縮(圖4-12)。
。")
58
等長收縮 等張收縮 抽動收縮 加成作用 強直 階梯收縮 肌肉疲乏

59
等長收縮 肌肉收縮所作的功(work)等於收縮力量(force)與位移(distance)的乘積。等長收縮(isometric contraction)張力改變但長度無改變,亦即無位移,所以無法作功,肌肉收縮消耗ATP,100%形成熱能。例如:比腕力,雙方僵持不下時,上肢肌肉的收縮狀態,即為等長收縮。
等於收縮力量(force)與位移(distance)的乘積。等長收縮(isometric contraction)張力改變但長度無改變,亦即無位移,所以無法作功,肌肉收縮消耗ATP,100%形成熱能。例如:比腕力,雙方僵持不下時,上肢肌肉的收縮狀態,即為等長收縮。")
60
等張收縮 等張收縮(isotonic contraction)力量不變,長度改變,亦即有位移,呈現運動狀態,等張收縮消耗之ATP,40%作功,60%形成熱能。例如:當用手舉起水桶,並保持手臂於半空中的位置時,即呈僵硬靜止狀態,手臂肌肉便產生了等長收縮,如再增加收縮的強度,則可導致肌肉長度的縮短,即呈運動狀態,此時等長收縮便轉變成等張收縮。
力量不變,長度改變,亦即有位移,呈現運動狀態,等張收縮消耗之ATP,40%作功,60%形成熱能。例如:當用手舉起水桶,並保持手臂於半空中的位置時,即呈僵硬靜止狀態,手臂肌肉便產生了等長收縮,如再增加收縮的強度,則可導致肌肉長度的縮短,即呈運動狀態,此時等長收縮便轉變成等張收縮。")
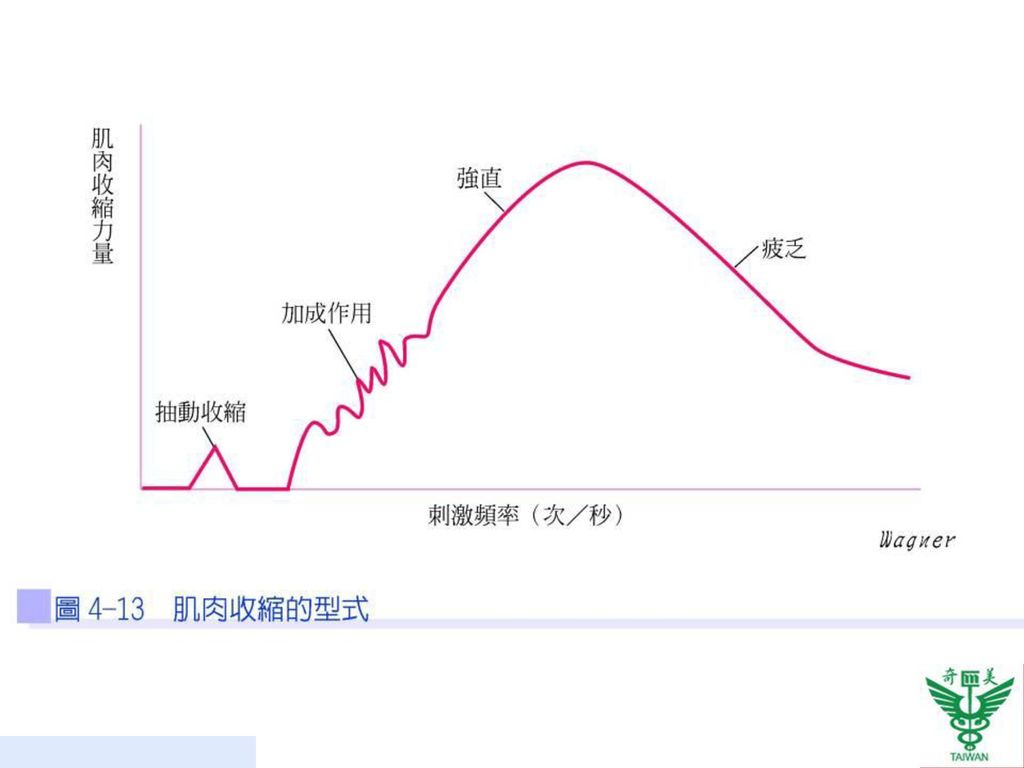
61
依刺激頻率的快慢可分成抽動收縮、加成作用、強直、階梯收縮及肌肉疲乏(圖4-13)。
。")
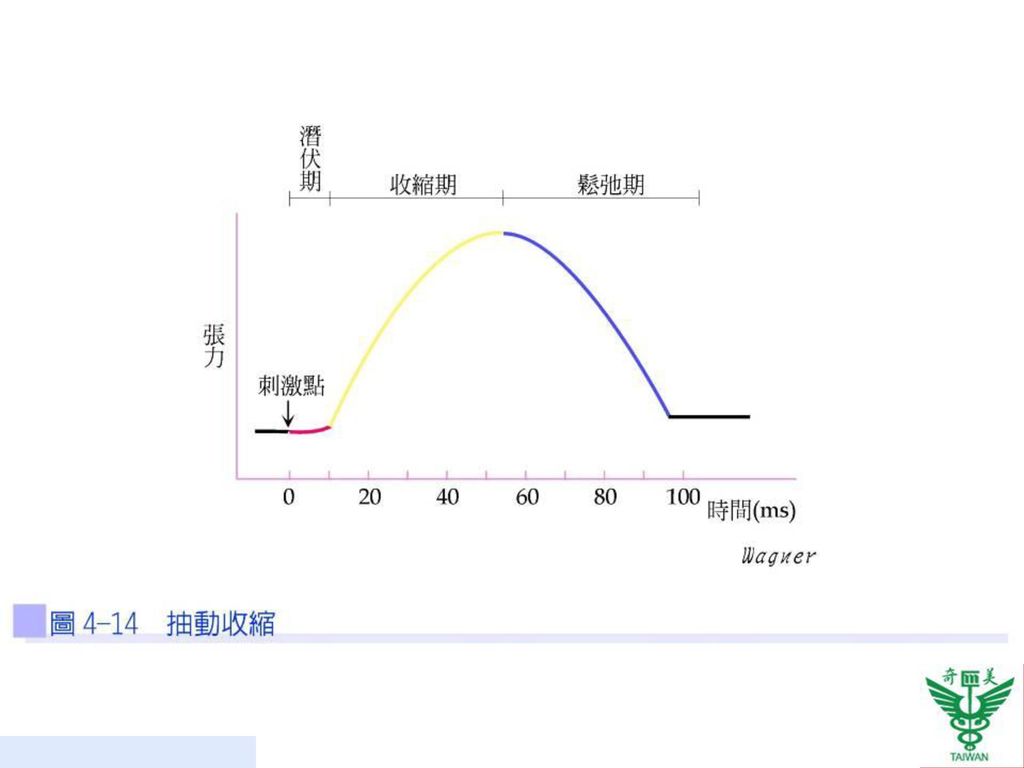
63
抽動收縮 當肌肉接受足夠強度的單一電擊刺激時,產生快速的痙攣反應稱為抽動收縮(twitch contraction)。整個過程可分三個時期(圖4-14): 第一期為潛伏期(latent period):指從刺激到肌纖維開始收縮所需的時期,為三個時期時間最短,起因於鈣離子由肌漿網釋放出來直到收縮所花費的時間。
:指從刺激到肌纖維開始收縮所需的時期,為三個時期時間最短,起因於鈣離子由肌漿網釋放出來直到收縮所花費的時間。")
65
第二期為收縮期(contraction period):指肌漿內高濃度的鈣離子引起肌纖維長度縮短的時期。
第三期為舒張期(relaxation period):為鈣離子被主動運輸至肌漿網,造成肌纖維長度回復的時間。
:為鈣離子被主動運輸至肌漿網,造成肌纖維長度回復的時間。")
66
加成作用 骨骼肌收縮的強度是有層級性的(graded),可依據不同的強度來決定其收縮的強度。當在肌肉鬆弛期尚未結束,給予第二個刺激,造成第二次收縮與第一次收縮重疊而強度增加的情形,稱為肌肉收縮的加成作用(summation)(圖4-13)。
,可依據不同的強度來決定其收縮的強度。當在肌肉鬆弛期尚未結束,給予第二個刺激,造成第二次收縮與第一次收縮重疊而強度增加的情形,稱為肌肉收縮的加成作用(summation)(圖4-13)。")
67
強直 強直 (tetanus)指連續快速的刺激,肌肉不停的收縮(圖4-13)。可分成兩種:
不完全強直(incomplete tetanus):若持續收縮加成,未達最高強度之前,即每兩個刺激間肌肉只有部分鬆弛期。 完全強直(complete tetanus):肌肉持續處於收縮狀態,最後無法分辨出兩次抽動收縮之間的放鬆期,稱為完全強直。
:若持續收縮加成,未達最高強度之前,即每兩個刺激間肌肉只有部分鬆弛期。 完全強直(complete tetanus):肌肉持續處於收縮狀態,最後無法分辨出兩次抽動收縮之間的放鬆期,稱為完全強直。")
68
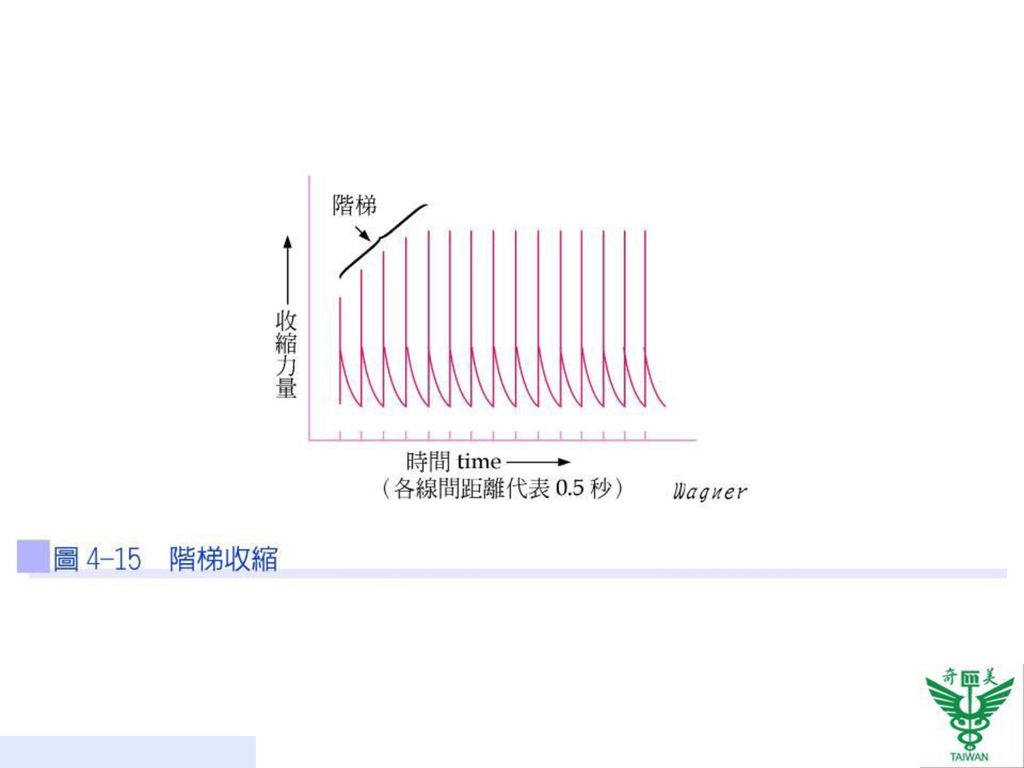
階梯收縮 一塊肌肉在受到相同刺激數次之後,產生更強而有力的收縮稱之為階梯收縮(treppe;staircase effect)。其機轉為:(1)增加肌纖維對鈣的利用率;(2)防止旋轉肌球素的阻隔作用。運動員賽前的暖身運動便是利用此原理(圖4-15)。
。其機轉為:(1)增加肌纖維對鈣的利用率;(2)防止旋轉肌球素的阻隔作用。運動員賽前的暖身運動便是利用此原理(圖4-15)。")
70
肌肉疲乏 肌肉疲乏(muscle fatigue)指肌肉受一段長時間的連續刺激,其收縮程度會漸弱,終至不反應為止,通常於完全強直一段時間後所產生(圖4-13)。其原因包括:
指肌肉受一段長時間的連續刺激,其收縮程度會漸弱,終至不反應為止,通常於完全強直一段時間後所產生(圖4-13)。其原因包括:")
71
肌肉細胞外鉀離子的堆積,影響動作電位的形成。
運動時乳酸堆積,造成氫離子濃度增加及pH值的下降,對肌肉的興奮及收縮聯合造成不良影響,而引起肌肉酸痛。 肌漿網釋出鈣離子的能力減弱。 肌肉收縮所產生的乳酸,其所分解的二氧化碳,會產生二氧化碳之毒效應。 可利用的氧減少。

72
肌纖維收縮的全或無定律 一運動單位分離出來,最初以弱電流刺激,小於閾值的刺激不會造成肌纖維的收縮,當電流增強至某一強度時,亦即超過閾值的刺激,則造成最大的收縮,並以恆定大小傳遞下去,稱為全或無定律。

73
因肌纖維的收縮程度全靠肌漿網所釋出的鈣離子量而定,而鈣離子能否釋出又全靠T小管是否有動作電位傳入,所以當刺激低於閾值時,動作電位便無法產生,T小管就不會有動作電位傳入,肌漿網也不會釋出鈣離子,肌纖維就不會收縮。

74
反之,一旦刺激超過閾值時,動作電位的大小皆相同,所以傳入T小管的動作電位大小也相同,肌漿網釋出的鈣離子量亦相同,因此肌纖維的收縮程度也會相同。
簡而言之,當給予超過閾值的刺激時,同一運動單位內的所有纖維就會完全收縮,若低於閾值的刺激則完全不收縮,稱為全或無定律(all-or-none principle)。
。")
75
骨骼肌纖維的種類 骨骼肌纖維的種類可依收縮速率的快慢及能量來源分別分成快肌(fast fiber)與慢肌(slow fiber)或稱有氧肌(oxidative fiber)和醣解肌(glycolytic fiber)。有氧肌纖維內含大量粒線體,可行有氧呼吸,但行有氧呼吸必須有足夠的氧氣,所以有氧肌纖維有較多的微血管供應氧氣。
與慢肌(slow fiber)或稱有氧肌(oxidative fiber)和醣解肌(glycolytic fiber)。有氧肌纖維內含大量粒線體,可行有氧呼吸,但行有氧呼吸必須有足夠的氧氣,所以有氧肌纖維有較多的微血管供應氧氣。")
76
此外,含有大量的肌紅素 (myoglobin),所以肌肉看起來比較暗紅,因此有氧肌纖維又稱紅肌纖維(red muscle fiber)。反之,無氧肌含有大量的肝醣及代謝肝醣的酵素,而只有少許的粒線體,所以無需太多的微血管輸送氧氣來行有氧呼吸,無氧肌所含的肌紅素也較少,所以看起來比較蒼白,因此無氧肌又名白肌纖維(white muscle fiber)。
,所以肌肉看起來比較暗紅,因此有氧肌纖維又稱紅肌纖維(red muscle fiber)。反之,無氧肌含有大量的肝醣及代謝肝醣的酵素,而只有少許的粒線體,所以無需太多的微血管輸送氧氣來行有氧呼吸,無氧肌所含的肌紅素也較少,所以看起來比較蒼白,因此無氧肌又名白肌纖維(white muscle fiber)。")
77
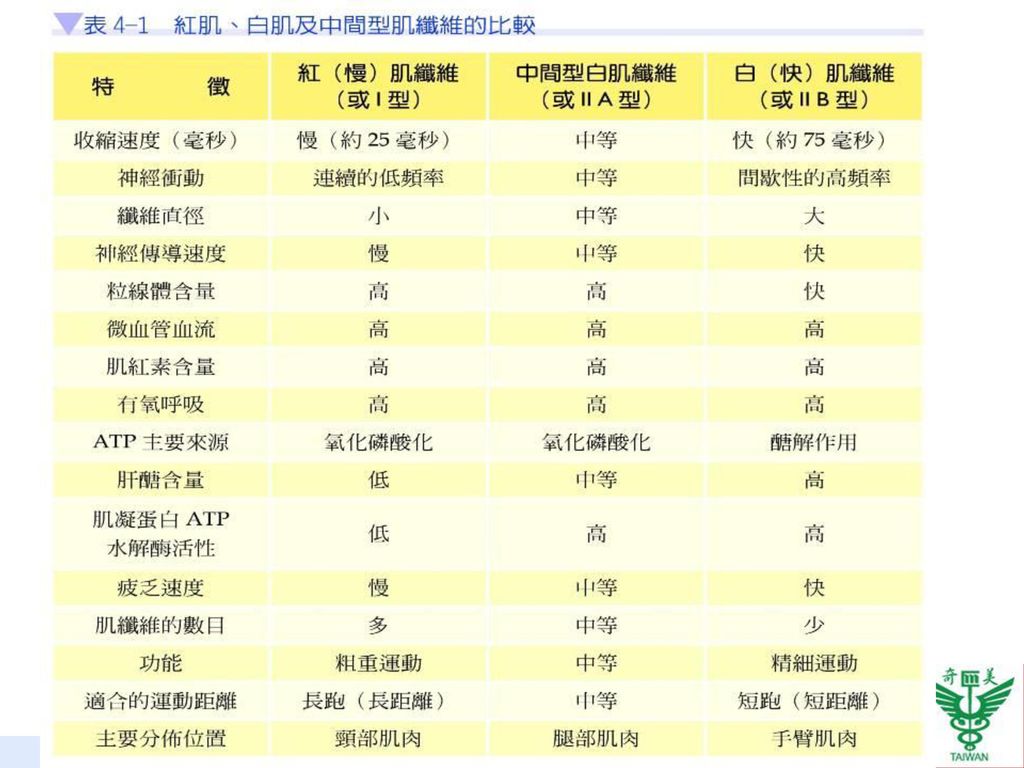
除了慢肌及快肌兩種肌纖維之外,體內尚含其他中間型的肌纖維,例如:II A型快肌纖維(type II A fiber),特性介於快肌與慢肌之間,不但收縮快速,並具有高度氧化容量,行有氧呼吸,故較不易產生疲乏(表4-1)。
,特性介於快肌與慢肌之間,不但收縮快速,並具有高度氧化容量,行有氧呼吸,故較不易產生疲乏(表4-1)。")
79
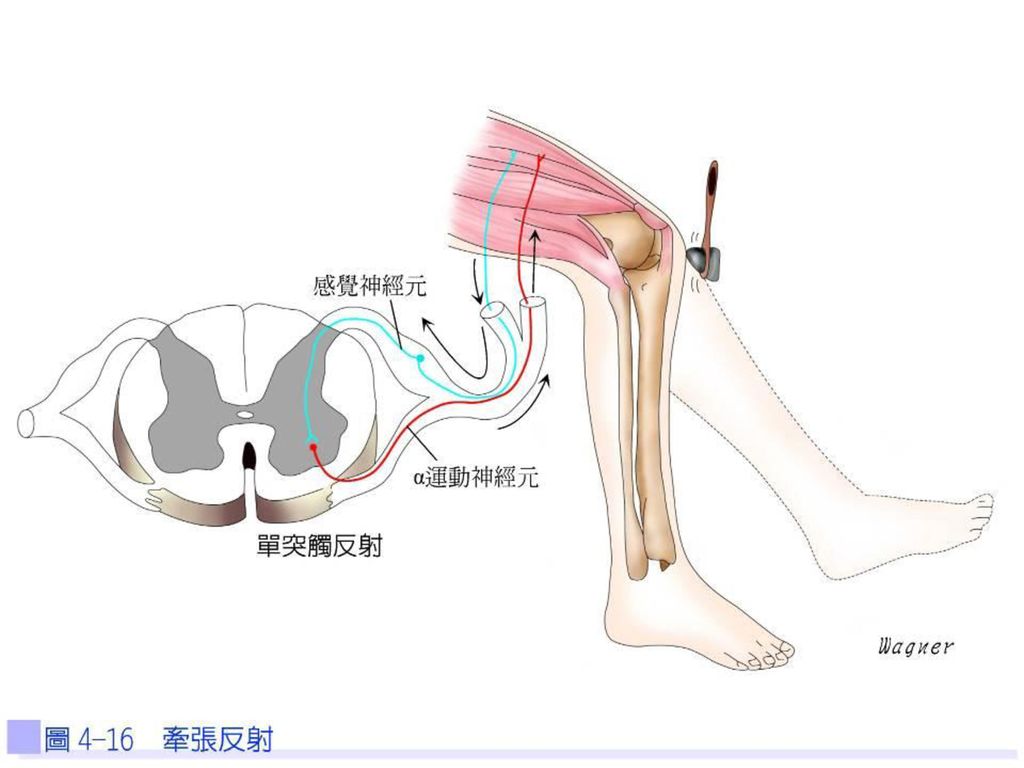
骨骼肌的神經控制 骨骼肌中含有肌梭,是肌肉接受牽張刺激的感受器,為肌肉中的長度監測器。當肌肉受到拉扯時,肌梭可刺激感覺神經元產生神經衝動,由脊神經傳入脊髓的背根,感覺神經元直接在脊髓中與 α運動神經元連接而形成單突觸反射。α運動神經元刺激梭外肌纖維的收縮,並解除了原有的牽張,稱為牽張反射。γ運動神經元將肌梭拉緊,加強肌梭對牽張的敏感度,即使當肌肉處於縮短狀態時,也能有效監控肌肉的長度(圖4-16)。
。")
81
第三節 平滑肌 平滑肌的構造 平滑肌的類型 平滑肌收縮機制
第三節 平滑肌 平滑肌(又稱內臟肌)構成內臟器官的管壁肌肉,與心肌一樣只受自主神經而不受意識的控制,故又名不隨意肌。 平滑肌的構造 平滑肌的類型 平滑肌收縮機制
構成內臟器官的管壁肌肉,與心肌一樣只受自主神經而不受意識的控制,故又名不隨意肌。 平滑肌的構造. 平滑肌的類型. 平滑肌收縮機制.")
82
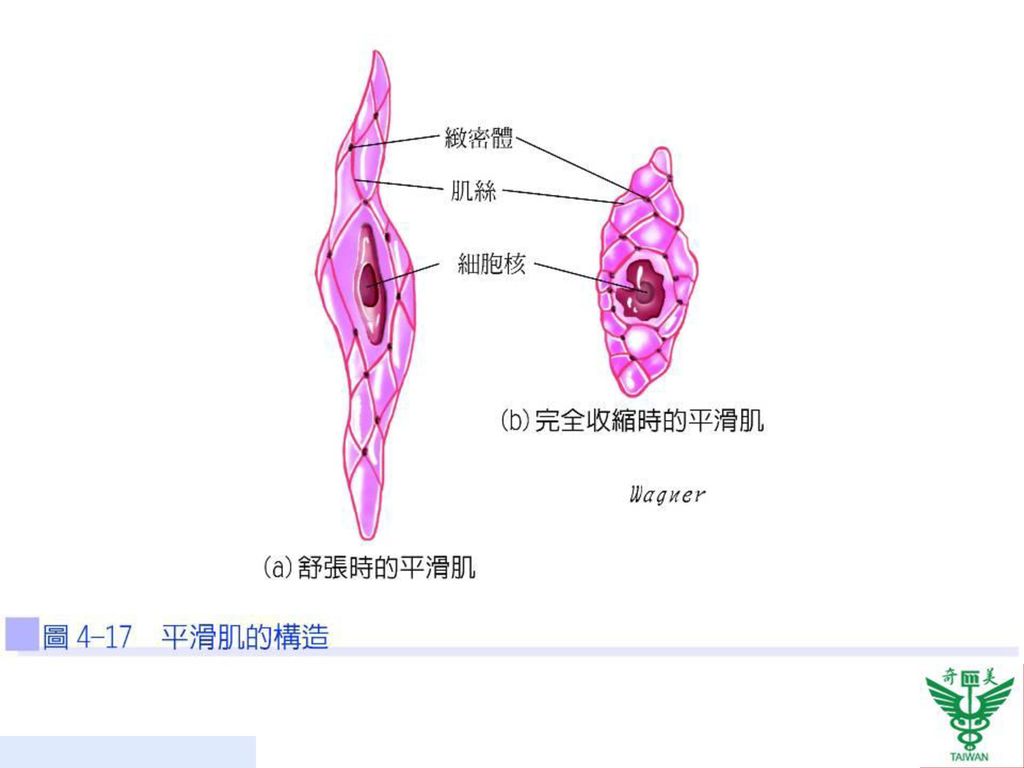
平滑肌的構造 平滑肌的長度約為50~400 µm,直徑在2~10 µm之間的紡綞型肌纖維,單一細胞核位於細胞中央,且具有分裂能力。細胞內粗、細肌絲排列不整齊,且附著於細胞膜或細胞質的緻密體(dense body)上。於功能上,緻密體類似橫紋肌中的Z線,其分佈於肌漿膜上,彼此之間亦以中間絲相連(圖4-17)。因平滑肌纖維沒有肌節,也沒有A帶或I帶,因此不具橫紋,故稱為平滑肌。平滑肌中細肌絲與粗肌絲的比例約16:1(在橫紋肌中為2:1)。
上。於功能上,緻密體類似橫紋肌中的Z線,其分佈於肌漿膜上,彼此之間亦以中間絲相連(圖4-17)。因平滑肌纖維沒有肌節,也沒有A帶或I帶,因此不具橫紋,故稱為平滑肌。平滑肌中細肌絲與粗肌絲的比例約16:1(在橫紋肌中為2:1)。")
84
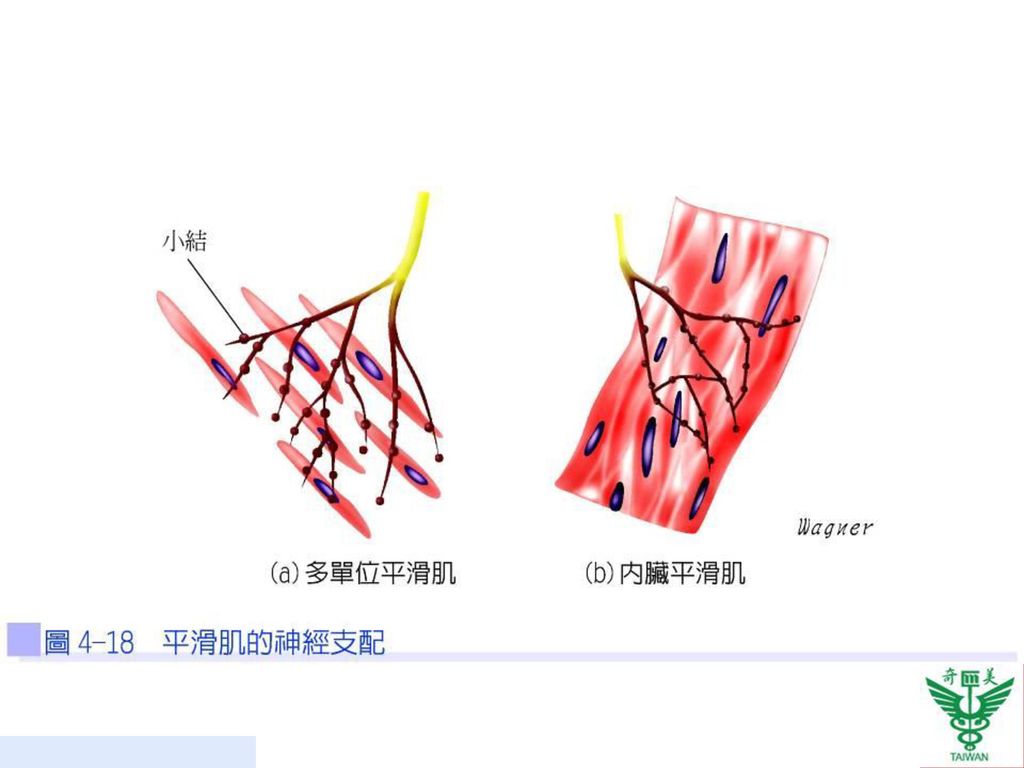
在平滑肌中並不如橫紋肌一般具有周全的肌漿質網系統,所以鈣離子的部分來源須靠細胞外鈣離子以擴散的方式進入肌纖維中來達成。支配平滑肌纖維的軸突,亦不具有骨骼肌的運動終板,而是形成許多小結(varicosities),內含有類似骨骼肌終板的囊泡,其中含有神經傳遞物質(圖4-18)。
,內含有類似骨骼肌終板的囊泡,其中含有神經傳遞物質(圖4-18)。")
86
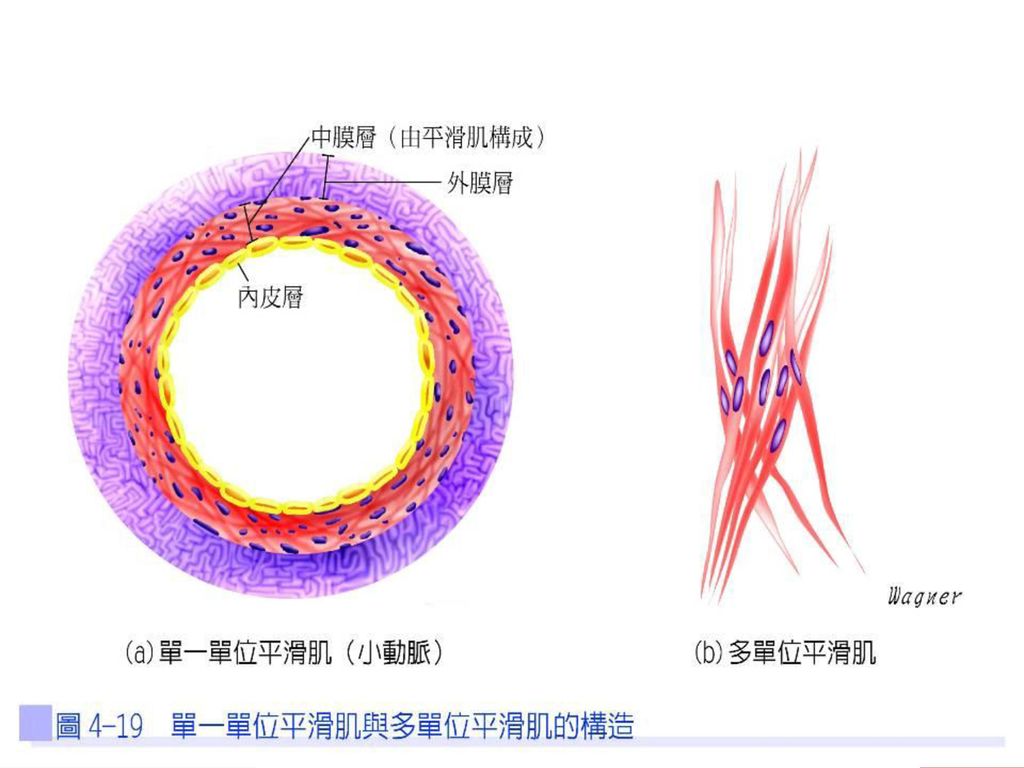
平滑肌的類型 身體內因神經支配及細胞間相互作用,可將平滑肌分成單一單位及多單位兩類型(表4-2及圖4-19)。 單一單位或內臟平滑肌
多單位平滑肌

89
單一單位或內臟平滑肌 單一單位或內臟平滑肌(single-unit or visceral smooth muscle)位於中空的內臟壁上,如消化道、生殖道、尿道等。肌纖維之間彼此以間隙接合(gap junction)而形成一大片內臟肌,當動作電位傳來時,可透過低阻力的細胞間接合,快速地將訊息傳至其他細胞,所以又稱為合體平滑肌(syncytial smooth muscle)。
位於中空的內臟壁上,如消化道、生殖道、尿道等。肌纖維之間彼此以間隙接合(gap junction)而形成一大片內臟肌,當動作電位傳來時,可透過低阻力的細胞間接合,快速地將訊息傳至其他細胞,所以又稱為合體平滑肌(syncytial smooth muscle)。")
90
多單位平滑肌 多單位平滑肌(multi-unit smooth muscle)位於眼球的虹膜、睫狀肌等,此類平滑肌纖維之間無細胞接合,所以每個平滑肌必須受自主軸突刺激。
位於眼球的虹膜、睫狀肌等,此類平滑肌纖維之間無細胞接合,所以每個平滑肌必須受自主軸突刺激。")
91
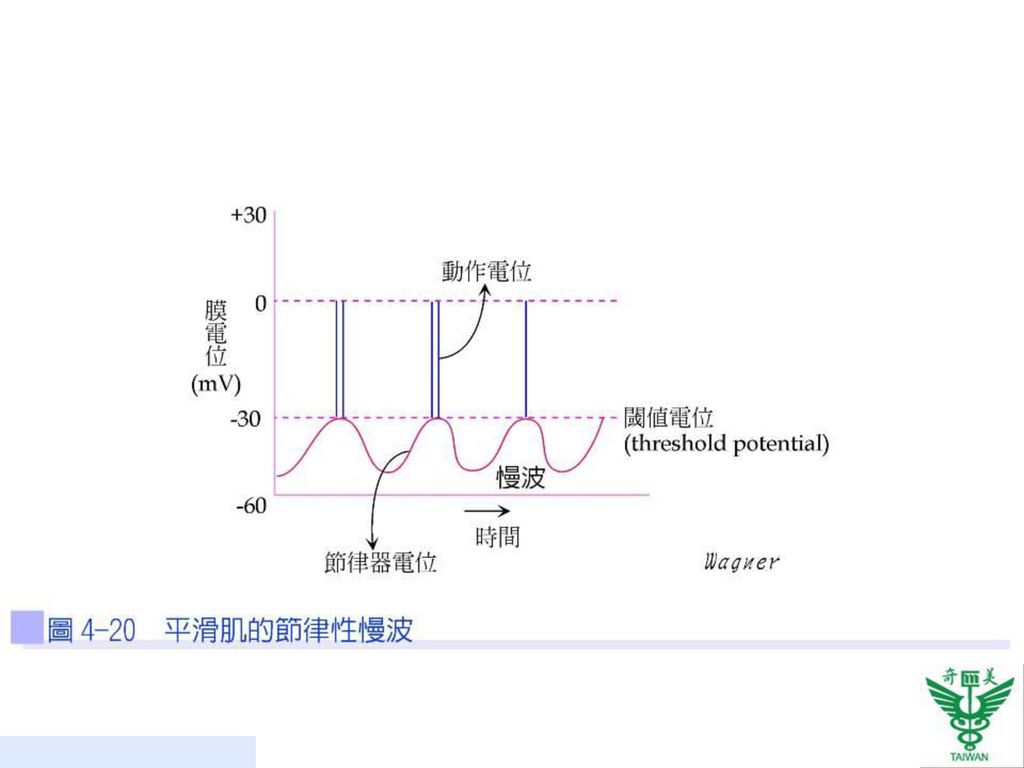
平滑肌收縮機制 平滑肌纖維的靜止膜電位約為-50mV,平滑肌能被多種不同訊號刺激而產生收縮,例如:神經訊號、荷爾蒙刺激、肌肉牽扯,部分則具有自我興奮性,亦即無需外來的刺激,平滑肌本身可自行產生動作電位,持續時間約為50毫秒(50msec),其通常伴隨著膜電位的慢波節律(slow wave rhythm)。慢波並非動作電位,所以不會造成肌肉收縮,但當慢波的電位高於閾值,即可產生動作電位,並傳至整個內臟平滑肌而引起收縮(圖4-20)。
,其通常伴隨著膜電位的慢波節律(slow wave rhythm)。慢波並非動作電位,所以不會造成肌肉收縮,但當慢波的電位高於閾值,即可產生動作電位,並傳至整個內臟平滑肌而引起收縮(圖4-20)。")
93
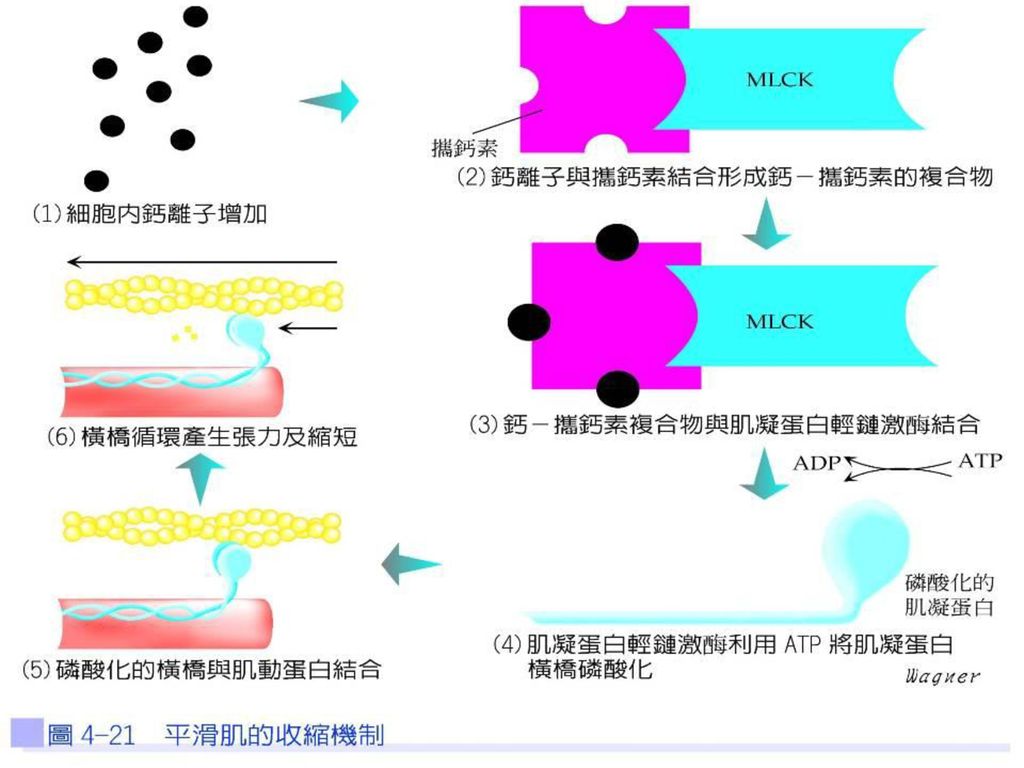
鈣離子通道開啟,鈣自細胞外進入細胞內,使細胞內鈣離子增加,因平滑肌並無旋轉素,鈣離子與攜鈣素(calmodulin)結合,其所形成的複合物活化肌凝蛋白輕鏈激酶(myosin light-chain kinase;MLCK),而催化橫橋頭部的磷酸化反應,在平滑肌中,橫橋的頭部必須先被磷酸化,才可與肌動蛋白結合,而產生肌肉收縮(圖4-21)。
結合,其所形成的複合物活化肌凝蛋白輕鏈激酶(myosin light-chain kinase;MLCK),而催化橫橋頭部的磷酸化反應,在平滑肌中,橫橋的頭部必須先被磷酸化,才可與肌動蛋白結合,而產生肌肉收縮(圖4-21)。")
95
因平滑肌及心肌收縮時鈣離子的來源主要為細胞外液,所以若將此兩種細胞放在不含鈣離子的溶液中,給予電刺激也無法引起收縮。但因骨骼肌收縮所需之鈣全部來自肌漿網,即使將骨骼肌細胞外液完全去除,其收縮仍不受影響。

96
第四節 心肌 心肌的構造 心肌收縮機制

97
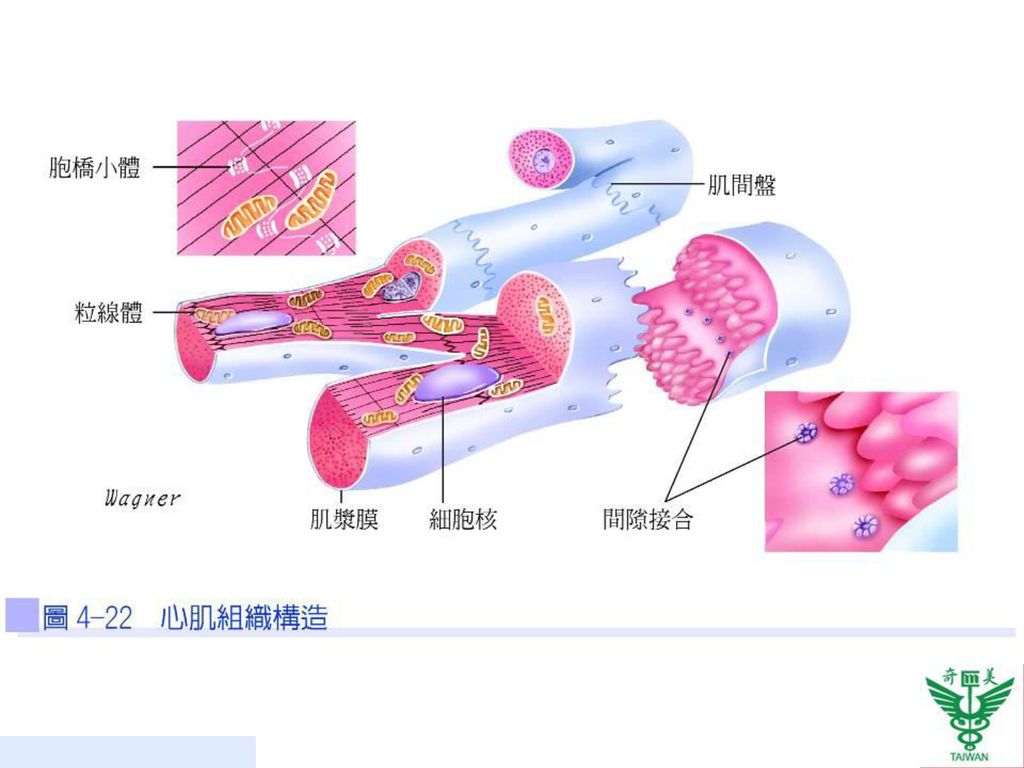
心肌的構造 心肌組織只存在心臟,心肌和骨骼肌一樣皆是由粗肌絲與細肌絲規則的排列成有橫紋的外觀,所以也是屬於橫紋肌。心肌的肌纖維較短,長約100~150µm,直徑約10~20µm,呈分叉狀,相鄰的心肌纖維之間藉著肌間盤(intercalated disks)互相連結形成網狀結構,肌間盤處有胞橋小體(desmosome)(圖4-22),將兩個肌細胞即心房合體細胞(atrial syncytium)及心室合體細胞(ventricular syncytium)彼此接合在一起。
互相連結形成網狀結構,肌間盤處有胞橋小體(desmosome)(圖4-22),將兩個肌細胞即心房合體細胞(atrial syncytium)及心室合體細胞(ventricular syncytium)彼此接合在一起。")
99
相鄰的肌絲有間隙接合(gap junction)亦即電性突觸,可打開離子通道,讓離子自由進出,而使電性衝動由一個細胞擴散到其他細胞,幫助更多的心室細胞產生同步收縮。這種心肌細胞物理性和電性的相互連結稱為心肌組織(myocardium),運作如同一個單一功能的單位。
亦即電性突觸,可打開離子通道,讓離子自由進出,而使電性衝動由一個細胞擴散到其他細胞,幫助更多的心室細胞產生同步收縮。這種心肌細胞物理性和電性的相互連結稱為心肌組織(myocardium),運作如同一個單一功能的單位。")
100
心肌組織傳導動作電位如一個單位(如同一個細胞),而且會一起收縮。因此,不像骨骼肌是分級收縮,心肌收縮是單位內所有心肌細胞一起參與;因此,心肌組織使整個心臟收縮。
,而且會一起收縮。因此,不像骨骼肌是分級收縮,心肌收縮是單位內所有心肌細胞一起參與;因此,心肌組織使整個心臟收縮。")
101
心肌部分的肌絲與骨骼肌不同,心肌為單核細胞,細胞核位於中央,不具運動終板,由於具有節律點(pacemaker),可自發性地產生動作電位,自主神經系統可調整節律點自發去極化的速率,亦即心跳速率(有關心臟自發性收縮及心跳速率的調節詳見第八章介紹)。
,可自發性地產生動作電位,自主神經系統可調整節律點自發去極化的速率,亦即心跳速率(有關心臟自發性收縮及心跳速率的調節詳見第八章介紹)。")
102
心肌收縮機制 心肌的收縮機制和骨骼肌大同小異,由於心肌組織的收縮,將血液送至體循環與肺循環,因此必須含有大量的粒線體,心肌細胞的粒線體約佔細胞的35%,骨骼肌細胞的粒線體則約佔2%而已,因為心肌細胞的能量來源特別需要依賴有氧代謝。

103
正常心肌的靜止膜電位約為-90mV,動作電位持續時間約為200毫秒(msec),約相當於絕對不反應期,為防止心肌發生強直收縮,所以心肌並無加成作用。
,約相當於絕對不反應期,為防止心肌發生強直收縮,所以心肌並無加成作用。")
104
由於心肌的肌漿網沒有骨骼肌那麼發達,無法儲存足夠的鈣離子,所以心肌收縮所需的鈣離子除了來自肌漿網外,還有一部分是經由橫小管的鈣通道直接進入細胞,因橫小管的末端開口於心肌的細胞外液,因此橫小管的鈣離子來源取決於細胞外液中鈣離子的濃度。

105
心臟的傳導系統及心房、心室肌肉皆是由特化的心肌組織組成,只受自主神經而不受意識的控制,所以心肌是不隨意肌。
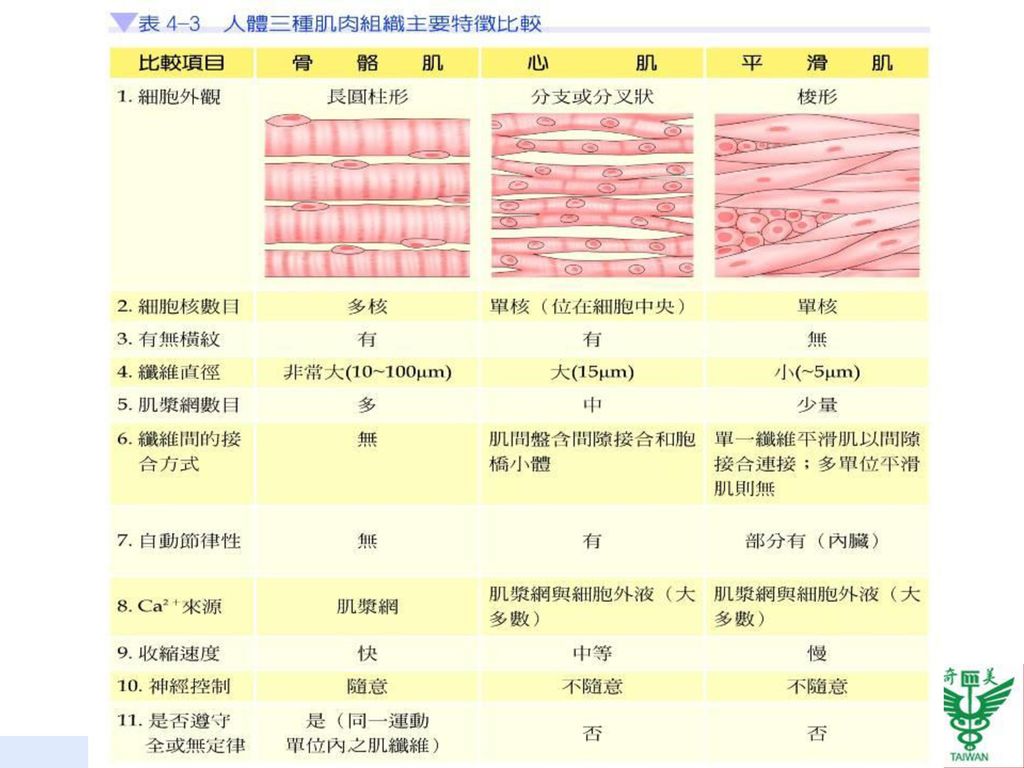
表4-3為人體之骨骼肌、心肌、平滑肌三種肌肉組織特徵之比較。

107
~ THE END ~ 回目錄

Similar presentations





 2. 脊髓發出: 31 對脊髓神經 ( spinal nerve ) 圖 9-24 人體 12 對腦 神經及其分布圖(圖 中之Ⅰ、Ⅱ、Ⅲ …… 分 別代表第 1 ~ 12 對腦神 經)。>")
 P.1 目錄 P.1 主題 P.2 目錄 P.3 類別 P.4 飲食金字塔 P.2.>")
 二、一維自由落體運動的特性 範例 1 自由落體( v 0 =0 ) 範例 2 自由落體的函數圖 範例 3 鉛直上拋 範例 4 自由落體運動公式.>")
 。 *以二次元元素 ( 透視、 重疊、色彩明暗 ) 表達 三次元的立體或空間感.>")



: 未刺激時的膜電位>")







:>")

細胞衰老(senescence) (2)基因體的不穩定(genomic instability)>")
