Download presentation
1
第九章 蛋白质的加工、易位及降解 1 蛋白质的加工 2 蛋白质易位 3 蛋白质的降解 4 小 结

2
1.1 N端fMet或Met切除 1.2 二硫键形成 1.3 特定氨基酸化学修饰 1.4 新生肽链中非功能片段切除
1 蛋白质的加工 1.1 N端fMet或Met切除 1.2 二硫键形成 1.3 特定氨基酸化学修饰 1.4 新生肽链中非功能片段切除 Back

3
1.1 N端fMet或Met的切除 细菌蛋白质N端甲酰基能被甲酰化酶水解。 不管原核生物或真核生物,N端Met常在肽链合成完前被切除。
有些病毒mRNA可翻译成很长多肽链,经蛋白酶水解后得到几个功能蛋白质分子。

5
Genomic structure of maize dwarf mosaic virus

6
1.2 二硫键形成 mRNA中没有胱氨酸密码子,而不少蛋白质含有二硫键.这是蛋白质合成后通过两个半胱氨酸的氧化作用生成的。

8
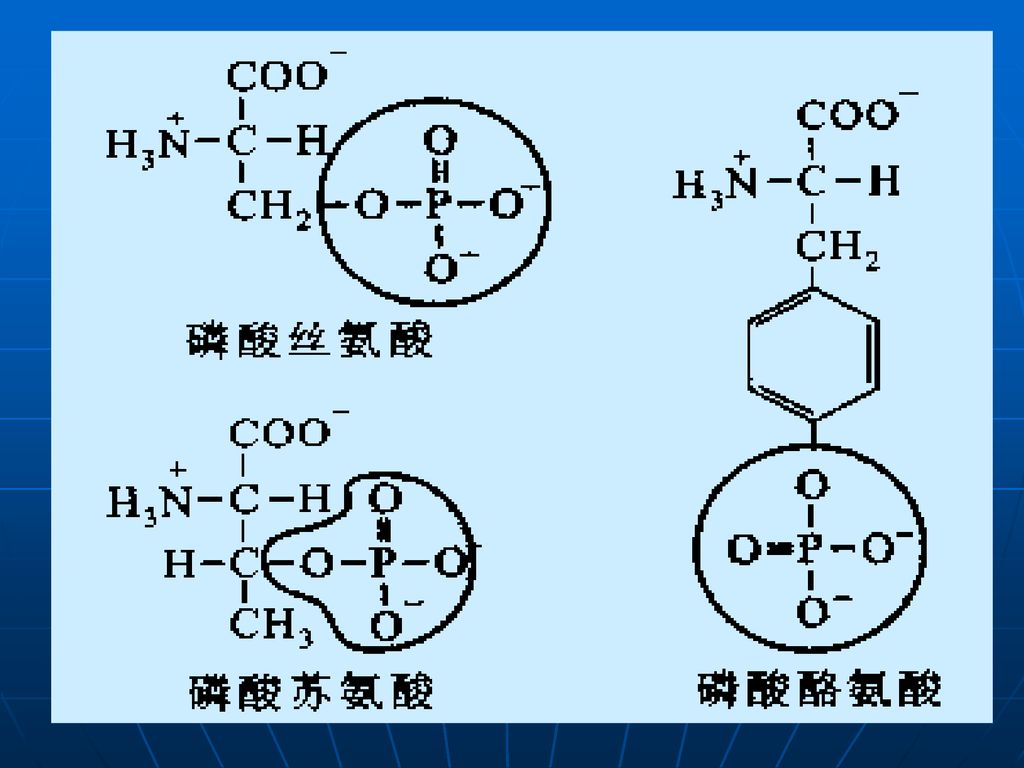
1.3 特定氨基酸的化学修饰 氨基酸侧链修饰作用包括磷酸化(如核糖体蛋白质)、糖基化(如各种糖蛋白)、甲基化(如组蛋白、肌肉蛋白质)、乙基化(如组蛋白)、羟基化(如胶原蛋白)和羧基化等。
、糖基化(如各种糖蛋白)、甲基化(如组蛋白、肌肉蛋白质)、乙基化(如组蛋白)、羟基化(如胶原蛋白)和羧基化等。")
13
小牛组蛋白H3前35个氨基体残基中的化学修饰

14
1.4 新生肽链中非功能片段切除 不少多肽类激素和酶的前体需要经过加工才能变为活性分子,如胰蛋白酶原经过加工切去部分肽段才能成为有活性的胰蛋白酶。

17
蜂毒毒蛋白的加工成熟 蜂毒蛋白只有经蛋白酶水解切除N端的22个 氨基酸以后才有生物活性,该胞外蛋白酶只 能特异切割X-Y 2肽。其中X是Ala,Asp和Glu; Y是Ala或Pro。

18
2 蛋白质易位 2.1 概 述 2.2 共翻译易位 2.3 翻译后易位 2.4 细菌蛋白质易位 分子伴侣 Back

19
2.1 概 述 细胞中蛋白质合成:绝大多数在细胞质中合成;小一部分在细胞器(叶绿体和线粒体)中合成。
2.1 概 述 细胞中蛋白质合成:绝大多数在细胞质中合成;小一部分在细胞器(叶绿体和线粒体)中合成。 定位于细胞器内的大部分在细胞质中合成,细胞器内合成的留在细胞器内。 蛋白质插入或穿过生物膜的过程称为蛋白质易位(protein translocation)。
中合成。 定位于细胞器内的大部分在细胞质中合成,细胞器内合成的留在细胞器内。 蛋白质插入或穿过生物膜的过程称为蛋白质易位(protein translocation)。")
20
蛋白质易位的两种方式 共翻译易位 co-translational translocation ◆是即将进入内质网的蛋白质的易位方式;
◆蛋白质正合成的时候就可与易位装置结合; ◆结果使核糖体定位于内质网表面,称膜结合核糖体(membrane-bound ribosome)。
。")
21
Proteins can enter the ER-Golgi pathway only by associating with the endoplasmic reticulum while they are being synthesized.

22
翻译后易位 post-translational translocation
◆蛋白质翻译完成后从核糖体上释放,然合扩散至合适靶膜并与易位装置结合。 ◆蛋白质合成时,其核糖体不与任何细胞器相连,称游离核糖体(free ribosome)
")
23
Proteins synthesized on free ribosomes in the cytosol are directed after their release to specific destinations by short signal motifs.

24
2.2 共翻译易位 2.2.1 信号肽假说 2.2.2 信号肽及其特性 2.2.3 信号肽与蛋白质转运的关系
2.2.4 蛋白质共翻译易位的基本过程

25
信号肽假说 共翻译定位蛋白质定位信息存在于该蛋白质自身结构中(信号肽),并可通过与特殊受体相互作用,使蛋白质定位于靶位。
,并可通过与特殊受体相互作用,使蛋白质定位于靶位。")
26
Proteins that enter the ER-Golgi pathway may flow through to the plasma membrane or may be diverted to other destinations by specific signals.

27
The endoplasmic reticulum consists of a highly folded sheet of membranes that extends from the nucleus. The small objects attached to the outer surface of the membranes are ribosomes.

28
2.2.2 信号肽及其特性 信号肽 (signal peptide): 能启动蛋白质运转的任何一段多肽。
绝大部分被运入内质网内腔蛋白质都带有一个信号肽。 信号肽结构特点: (1)常位于蛋白质N末端,可切割; (2)长度: 个残基; (3)序列内有一个全部或大部分疏水组成的疏水核心; (4)靠近该序列 N 端常有几个正电荷氨基酸; (5)其C-末端靠近切割处常带数个极性氨基酸,离切割点最近那个氨基酸常带很短侧链(Ala或Gly )
常位于蛋白质N末端,可切割; (2)长度: 个残基; (3)序列内有一个全部或大部分疏水组成的疏水核心; (4)靠近该序列 N 端常有几个正电荷氨基酸; (5)其C-末端靠近切割处常带数个极性氨基酸,离切割点最近那个氨基酸常带很短侧链(Ala或Gly )")
29
An N-terminal signal sequence is hydrophobic
The signal sequence of bovine growth hormone consists of the N-terminal amino acids and has a central highly hydrophobic region, preceded or flanked by regions containing polar amino acids.

30
2.2.3 信号肽与蛋白质转运的关系 (1)完整信号肽是保证蛋白质转运的必要条件; (2)仅有信号肽不足以保证蛋白质转运发生;
(3)信号序列切除并不是转运所必需; (4)并非所有运转蛋白质都有可降解的信号肽。
信号序列切除并不是转运所必需; (4)并非所有运转蛋白质都有可降解的信号肽。")
31
2.2.4 蛋白质共翻译易位的基本过程 蛋白质共翻译易位可分两个阶段: 首先:带有新生肽链的核糖体与膜结合;
蛋白质共翻译易位的基本过程 蛋白质共翻译易位可分两个阶段: 首先:带有新生肽链的核糖体与膜结合; 然合:新生肽链进入膜通道并易位。

32
核糖体与膜结合需要信号识别颗粒(signal recognition particle, SRP)。SRP有两种能力:
◆结合新合成的分泌型蛋白的信号序列; ◆可和位于膜上的 SRP 受体结合。

33
The signal sequence initiates membrance entry

34
2.3 翻译后易位 前导肽及其特性 蛋白质穿越不同器膜的过程

35
2.3.1 前导肽及其特性 翻译后跨膜易位的蛋白质,前体一般含前导肽(leader peptide),前导肽在跨膜运转中起重要作用。
过膜后,前导肽水解,蛋白质变为有功能的蛋白质。

37
前导肽的一般特性: (1)带正电荷碱性氨基酸(Arg)较丰富, 分散于不带电荷的氨基酸序列之间; (2)缺少带负电荷的酸性氨基酸;
(3)羟基氨基酸(Ser)含量较高; (4)有形成两亲(亲水和疏水)α- 螺旋能力。
羟基氨基酸(Ser)含量较高; (4)有形成两亲(亲水和疏水)α- 螺旋能力。")
38
The leader sequence of yeast cytochrome c oxidase subunit IV consists of 25 neutral and basic amino acids. The first 12 amino acids are sufficient to transport any attached polypeptide into the mitochondrial matrix.

39
Note 任何蛋白过膜须解决一个问题:蛋白质表面常是亲水性的,而生物膜是疏水性的。因此必须在膜上构建特殊的通道才能使蛋白质穿过膜。

40
蛋白质穿越不同器膜的过程 (1)穿越内质网(endoplasmic reticulum) 、线粒体(mitochondria)及叶绿体 (chloroplast) 膜 膜上含有镶嵌于膜的蛋白质样结构,它可以使蛋白质不与疏水性的类脂分子接触而穿过膜。

41
Proteins enter the ER, mitochondrion, chloroplast by binding to a translocon that transports them across the membrane.

42
(2)穿越过氧化物酶体膜 (peroxisome)
膜上也有类似的装置,但底物蛋白质并不直接与其结合。需载体蛋白协助穿过膜。

43
Proteins are transported into peroxisomes by a carrier protein that binds them in the cytosol, passes with them through the membrane channel, and releases them on the other side.

44
(3)穿越细胞核膜 向核内转运需更大更复杂的装置,即核孔(nuclear pore )。由载体蛋白与底物结合并进行跨核孔转运
穿越细胞核膜 向核内转运需更大更复杂的装置,即核孔(nuclear pore )。由载体蛋白与底物结合并进行跨核孔转运")
45
Proteins enter the nucleus by passage through very large nuclear pores.

46
Proteins are localized by specific pathways
proteins that are localized post-translationally are released into the cytosol after synthesis on free ribosomes. Some have signals for targeting to organelles such as the nucleus or mitochondria. Proteins that are localized cotranslationally associate with the ER membrane during synthesis. The proteins pass into the endoplasmic reticulum, along to the Golgi, and then through the plasma membrane, unless they have signals that cause retention at one of the steps on the pathway.

47
2.4 细菌蛋白质的易位 细菌蛋白通过共翻译或翻译后可被定位于细胞膜或细胞周质空间,或被分泌出去。

49
The Sec system has the SecYEG translocon embedded in the membrane, the SecA associated protein that pushes proteins through the channel, the SecB chaperone that transfers nascent proteins to SecA, and the signal peptidase that cleaves the N-terminal signal from the translocated protein.

50
SecB transfers a nascent protein to SecA, which inserts the protein into the channel. Translocation requires hydrolysis of ATP and a protonmotive force. SecA undergoes cycles of association and dissociation with the channel and provides the motive force to push the protein through.

51
2.5 分子伴侣 (chaperone) 定 义 生物学功能
 定 义 生物学功能")
52
2.5.1 定 义 分子伴侣:结合在一些不完全装配或不恰当折叠蛋白上,帮助它们折叠或防止它们聚集的蛋白质。
定 义 分子伴侣:结合在一些不完全装配或不恰当折叠蛋白上,帮助它们折叠或防止它们聚集的蛋白质。 如:Hsp70和与之相关的一些蛋白组成了一类主要分子伴侣,作用于许多目标蛋白质。

53
2.5.2 生物学功能 (1)帮助新生蛋白质正确折叠 (2)纠正错误折叠或介导其降解 1)帮助新生蛋白质的正确折叠:
生物学功能 (1)帮助新生蛋白质正确折叠 (2)纠正错误折叠或介导其降解 1)帮助新生蛋白质的正确折叠: 当蛋白质刚刚合成时,即离开核糖体进入细胞质时,它以未折叠的形式存在。分子伴侣通过控制表面的可接近性来影响折叠的过程,这一过程与最初正确构象的获得有关; 2)纠正错误折叠: 当蛋白质变性时,新的区域会被暴露并可与其它区域相互作用。而在蛋白质起始合成时会发生(短暂的)错误折叠,这两种相互作用是相似的。分子伴侣会识别这些错误折叠的蛋白质,并帮助其复性,或者介导其降解。
帮助新生蛋白质正确折叠. (2)纠正错误折叠或介导其降解. 1)帮助新生蛋白质的正确折叠: 当蛋白质刚刚合成时,即离开核糖体进入细胞质时,它以未折叠的形式存在。分子伴侣通过控制表面的可接近性来影响折叠的过程,这一过程与最初正确构象的获得有关; 2)纠正错误折叠: 当蛋白质变性时,新的区域会被暴露并可与其它区域相互作用。而在蛋白质起始合成时会发生(短暂的)错误折叠,这两种相互作用是相似的。分子伴侣会识别这些错误折叠的蛋白质,并帮助其复性,或者介导其降解。")
54
Hydrophobic regions of proteins are intrinsically interactive, and will aggregate with one another when a protein is synthesized (or denatured) unless prevented.
 unless prevented.")
55
Proteins emerge from the ribosome or from passage through a membrane in an unfolded state that attracts chaperones to bind and protect them from misfolding.

56
Chaperones bind to interactive regions of proteins as they are synthesized to prevent random aggregation. Regions of the protein are released to interact in an orderly manner to give the proper conformation.

57
3 蛋白质的降解 概 述 降解涉及到的三种重要酶 有序降解机理 Back

58
3.1 概 述 ◆生物体内蛋白的降解是一个有序的过程。 ◆不同蛋白质其半衰期差异较大。
3.1 概 述 ◆生物体内蛋白的降解是一个有序的过程。 ◆不同蛋白质其半衰期差异较大。 ◆蛋白质半衰期与N端氨基酸残基有一定关系(P145)。
。")
59
阿龙·切哈诺沃 (1947-)
")
60
3.2 降解涉及到的三种重要酶 泛素活化酶 (E1) (Ubiquitin-activating enzyme) 泛素联合酶(E2)
3.2 降解涉及到的三种重要酶 泛素活化酶 (E1) (Ubiquitin-activating enzyme) 泛素联合酶(E2) (Ubiquitin conjugating enzyme) 泛素连接酶 (E3) (Ubiquitin ligase) 泛素活化酶 (ubiquitin - activating enzyme,E1) 泛素结合酶 (ubiquitin - conjugating enzyme,E2) 泛素蛋白连接酶 (ubiquitin -proteinligating enzyme, E3)
 (Ubiquitin-activating enzyme) 泛素联合酶(E2) (Ubiquitin conjugating enzyme) 泛素连接酶 (E3) (Ubiquitin ligase) 泛素活化酶. (ubiquitin - activating enzyme,E1) 泛素结合酶. (ubiquitin - conjugating enzyme,E2) 泛素蛋白连接酶. (ubiquitin -proteinligating enzyme, E3)")
61
3.3 蛋白质有序降解机理 ▼ 酶E1激活泛素分子,此过程需要ATP能量。泛素分子被激活后被运送到E2上, E2负责将泛素绑在被降解的蛋白质上。 ▼ E3能识别被降解的蛋白质。当E2携带泛素分子在E3的指引下接近被降解蛋白质时,E2就把泛素分子绑在被降解蛋白质上。 ▼ 在这个过程中,酶E3所发挥的作用最大。一旦蛋白与多个泛素分子相连(每一个泛素分子都加在前一个泛素分子第46位Lys残基上),就形成了蛋白降解信号。
,就形成了蛋白降解信号。")
63
4 小 结 重点内容: ①几个重要概念:蛋白质易位、共翻译易位、翻译后易位、信号肽、导肽、分子伴侣;②前体蛋白质加工内容;③共翻译易位机制;④翻译后易位的机制;⑤信号肽的结构特点;⑥信号肽与蛋白质转运的关系;⑦前导肽的一般特性;⑧蛋白质穿越不同细胞器膜的不同方式;⑨分子伴侣的生物学功能;⑩蛋白质有序降解机理。 了解内容: ①细菌中蛋白质跨膜转运机理;②蛋白质半衰期与N端氨基酸残基的关系。 Back













染色体结构不同; ( 2 )原核生物具有正调控和负调控并重的特点,真核 生物目前已知的主要是正调控; ( 3 )原核生物的转录和翻译是相偶联的,真核生物的.>")










>")



 電子傳遞鏈 傳導概論>")

>")
