Download presentation
Presentation is loading. Please wait.

1
Yingzi Kang Ph.D Dept. of biochemistry, Tianjin Medical University
基因组的结构和功能 Yingzi Kang Ph.D Dept. of biochemistry, Tianjin Medical University

2
概 念 基因(gene) 是核酸的中贮存遗传信息的遗传单位,是贮存有功能的蛋白质多肽链或RNA序列信息及表达这些信息所必需的全部核苷酸序列(P27)。 从生化学上来说指的是一段DNA或RNA(病毒)顺序,该顺序可以产生或影响某种表型,可以由于突变生成等位基因变异体。 从遗传学上来说代表1个遗传单位、1个功能单位、1个交换单位或1个突变单位。

3
表型和基因型 某一机体可以观察到的特征称为表型(phenotype),与表型相应的基因组成称为基因型(genotype)。如细菌能否合成亮氨酸记为Leu+和Leu-,相应的基因型为Leu+和Leu-。
等位基因 (1)二倍体细胞有2套基因,一套来自父本,另一套来自母本,每个细胞的全部基因均由2套基因组成,每一对基因称做等位基因。 (2)由于突变作用引起DNA结构变异,所以某一基因可具有若干种不同的形式,这种同一基因不同的形式互称等信基因(allele)。
二倍体细胞有2套基因,一套来自父本,另一套来自母本,每个细胞的全部基因均由2套基因组成,每一对基因称做等位基因。 (2)由于突变作用引起DNA结构变异,所以某一基因可具有若干种不同的形式,这种同一基因不同的形式互称等信基因(allele)。")
4
基因的基本结构 5’、、、AGCCGACTATGTCGAAGCTT、、、、、、GCTTGACTATAAGACA、、、3’
3‘、、、TCGGCTGATACAGCTTCTAA、、、、、、CGAACTGATATTCTGT、、、5‘ 转录调控区 贮存RNA或蛋白质结构信息区 转录终止区

5
基因组(gencme) 细胞或生物中,一套完整单倍体遗传特质的总和(包括一种生物所需的全套基因及间隔序列)称为基因组(P27)。
基因组的结构主要指不同的基因功能区域在核酸分列中的分布和排布情况,基因组的功能是贮存和表达遗传信息。 人类基因组包含多染色体和XY两条性染色体上的全部遗传物质(核基因组)以及胞线粒体上的遗传物质(线粒体基因组)。(X免疫基因,男XY,女XX)。 * 1个配对(精子或卵子),1个单倍体细胞或1个病毒所包含的全套基因,称为基因组。
以及胞线粒体上的遗传物质(线粒体基因组)。(X免疫基因,男XY,女XX)。 * 1个配对(精子或卵子),1个单倍体细胞或1个病毒所包含的全套基因,称为基因组。")
6
第一节 原核基因组 Prokaryotic genome 以细菌为代表讲述,有称bacteria genome。细菌对医学分子生物学有重要贡献,是基因工程研究的主要材料之一。因为: 1.构造相对简单,基因结构也不复杂,取材便利,易于培养,可选择突变株进行研究,实验结果容易重复。(如选择DDDPI缺乏的Ecoli突变株,Ecoli仍可合成DNA,说明酸I对EcoliDNA合成不起作用。) 2.与人类有共同的要子生物学规律可,如: (1)遗传物质都是DNA; (2)主要的功能分子都是蛋白质; (3)基因密码是通用的,等等。 3.尤其是E.coli,是分子克隆是“明星“,基因工程主要原因的工程菌,因为基因工程的主要工作是克隆克核基因在原核系统中表达。
 2.与人类有共同的要子生物学规律可,如: (1)遗传物质都是DNA; (2)主要的功能分子都是蛋白质; (3)基因密码是通用的,等等。 3.尤其是E.coli,是分子克隆是 明星 ,基因工程主要原因的工程菌,因为基因工程的主要工作是克隆克核基因在原核系统中表达。")
7
一、原核生物基因组结构与功能的特点 1.基因组通常仅由一条环状双链DNA分子组成。 其DNA是与蛋白质结合,但并不形成染色体结构,只是习惯上将之称为染色体。细菌染色体DNA在胞内形成一个致密区域,即类核(nucleoid),类核无核膜将之与胞浆分开。 2.基因组中只有1个复制起点。 3.具有操纵子结构。 操纵子(operon) 是指数个功能相关的结构基因串联在一起,构成信息区,连同其上游的调控区(包括启动和操纵区)及其下游的转录终止信号构成的基因表达单位。(见第六章) 4.结构基因无重叠现象,基因组中任何一段DNA不会用于编码2种蛋白质。 5.基因序列是连续的,无内含子结构。
 是指数个功能相关的结构基因串联在一起,构成信息区,连同其上游的调控区(包括启动和操纵区)及其下游的转录终止信号构成的基因表达单位。(见第六章) 4.结构基因无重叠现象,基因组中任何一段DNA不会用于编码2种蛋白质。 5.基因序列是连续的,无内含子结构。")
8
6.编码区和非编码区(主要是调控序列)在基因组中约各占50%。(5%,95%)
7.基因组中的重复序列很少。编码蛋白质结构基因多为单拷贝,但编码rRNA的基因往往是多拷贝的,这有利于核糖体的快速组装。(15AA/秒,2AA/秒) 8.具有编码同功酶的基因(isogene) 这是一类结构不完全相同,而功能相同的基因。如E.coli含有2个编码乙酸乳酸合成酶的基因和2个编码分支酸变位酶同工酶的基因。 9.细菌基因组中存在可移动的DNA序列,包括插入序列和转座子。 10.原核基因的基本结构特点: 启动子(promoter)、操纵基因(operator)、调控序列、结构基因(structure gene)、终止子(terminator)。(见第六章)
 8.具有编码同功酶的基因(isogene) 这是一类结构不完全相同,而功能相同的基因。如E.coli含有2个编码乙酸乳酸合成酶的基因和2个编码分支酸变位酶同工酶的基因。 9.细菌基因组中存在可移动的DNA序列,包括插入序列和转座子。 10.原核基因的基本结构特点: 启动子(promoter)、操纵基因(operator)、调控序列、结构基因(structure gene)、终止子(terminator)。(见第六章)")
9
二、染色体外的遗传物质———质粒 (一)概念 1.质粒(plasmid) 是独立于许多细菌及某些真核细胞染色体外共价闭合环状的DNA分子(covalant closed circnlar,cccDNA),能独立复制的最小遗传单位。(P35-6) 2.质粒是双链的DNA分子,大小在1—200kb之间,和病毒不同,它们没有衣壳蛋白(裸DNA)。
。")
10
从生化学来说,除酵母的杀伤质粒(killer plasmid)是RNA外,其余质粒是染色体外的cccDNA分子。
是RNA外,其余质粒是染色体外的cccDNA分子。")
11
3.质粒与宿主细胞的关系 (1)质粒对宿主的生存不是必需的,只是“友好”的“借居”宿主细胞中,既不杀伤细胞,对宿主的代谢活动也无影响,宿主离开质粒照样的生存下去。 (2)质粒离开宿主就无法生存,只有依赖宿主细胞的(酶和蛋白质)帮助,才能完成自身的复制(扩增)、转录。 (3)质粒经常为宿主执行一些适当的遗传功能,作为对宿主细胞的补偿(“交房租”)。 (4)质粒赋于宿主各种有利的表型(质粒编码蛋白质或酶),使宿主获得生存优势,与我们基因工程实验紧密相关的,如抗生素抗性基因: Ampr 酶,水解β-内酰胺环,解除氨关毒性,使细菌抗氨关。 Tetr 膜蛋白,可阻止四环素进入细胞,使细菌抗四环素。
质粒经常为宿主执行一些适当的遗传功能,作为对宿主细胞的补偿( 交房租 )。 (4)质粒赋于宿主各种有利的表型(质粒编码蛋白质或酶),使宿主获得生存优势,与我们基因工程实验紧密相关的,如抗生素抗性基因: Ampr 酶,水解β-内酰胺环,解除氨关毒性,使细菌抗氨关。 Tetr 膜蛋白,可阻止四环素进入细胞,使细菌抗四环素。")
12
4.质粒发现和研究意义 1)理论意义 质粒能够复制、传递和表达遗传信息,从分子遗传学观点来看是一种有机体,是比病毒更原始的生命形式,是生命起源研究的起一块体重要基石。 2)实践意义 是基因工程的重要载体(vector),能把外源基因(目的基因)送到宿主细胞中去克隆扩增或克隆表达(见第八章)。 ①质粒是可以改造的,可以剪切、剪接的,基因工程的重要任务之一就是严格改造质粒的同时,控制质粒不传递,若一个致癌质粒可以传递就会传到外都是。
实践意义 是基因工程的重要载体(vector),能把外源基因(目的基因)送到宿主细胞中去克隆扩增或克隆表达(见第八章)。 ①质粒是可以改造的,可以剪切、剪接的,基因工程的重要任务之一就是严格改造质粒的同时,控制质粒不传递,若一个致癌质粒可以传递就会传到外都是。")
13
②作为基因工程载体的3个特点: A.都能独立自主的复制; B.都能便利的加以检测(抗生素抗性); C.都能容易引进宿主细胞中去,也易从宿主细胞中分离纯化(提质粒)。 质粒符合上述3个条件。 基因工程中主要使用人工构建的质粒。
; C.都能容易引进宿主细胞中去,也易从宿主细胞中分离纯化(提质粒)。 质粒符合上述3个条件。 基因工程中主要使用人工构建的质粒。")
14
(二)质粒的分类 1.按质粒的复制机理,分为2类: 1)严谨控制型(stringent contrd type) 2)松弛控制型(relaxed control type) (1)拷贝数少,一般<10个,分子量大; (1)拷贝数多,10-200个,分子量小; (2)复制受限,受细菌宿主DNA复制系 (2)复制不受细菌DNA复制系统限制, 统的控制; 仅需DDDPI,当宿主蛋白质合成受抑制 (3) 特点是这类质粒可以自传递; 时(如培养中加入氯要素时),其拷贝 (4)严谨控制机理(低拷贝原因),认 数可猛增至 之多,该性质对 为是该质粒可以产生阻逼蛋白,反馈 基因工程技术十分有利。 抑制自身DNA合成。 )分子量小,不具备自传递能力; 4)基因工程使用松弛型(高拷贝数) 质粒,以获得列多的基因产物。 *拷贝数(copy number)—细胞所含的按每—基因组计算的某种质粒或基因的数目。
复制受限,受细菌宿主DNA复制系 (2)复制不受细菌DNA复制系统限制, 统的控制; 仅需DDDPI,当宿主蛋白质合成受抑制. (3) 特点是这类质粒可以自传递; 时(如培养中加入氯要素时),其拷贝. (4)严谨控制机理(低拷贝原因),认 数可猛增至 之多,该性质对. 为是该质粒可以产生阻逼蛋白,反馈 基因工程技术十分有利。 抑制自身DNA合成。 3)分子量小,不具备自传递能力; 4)基因工程使用松弛型(高拷贝数) 质粒,以获得列多的基因产物。 *拷贝数(copy number)—细胞所含的按每—基因组计算的某种质粒或基因的数目。")
15
1)小型质粒,<15kb 2)大型质粒>15kb
2.按分子量大小,分为2类 1)小型质粒,<15kb )大型质粒>15kb 小型质粒,无接合和自传递能力,在按 多属接合型或自传递型,大型质粒只 合质粒协助也能转移,也可心通过转化 能通过细菌的接合作用人一个细菌 作用进入受体细胞,这类质粒种类较多, 传到另一个细菌。(如F质粒)。 几乎每种细菌都可以含有2种以上,基因 工程一般用小型质粒。
小型质粒,<15kb 2)大型质粒>15kb. 小型质粒,无接合和自传递能力,在按 多属接合型或自传递型,大型质粒只. 合质粒协助也能转移,也可心通过转化 能通过细菌的接合作用人一个细菌. 作用进入受体细胞,这类质粒种类较多, 传到另一个细菌。(如F质粒)。 几乎每种细菌都可以含有2种以上,基因. 工程一般用小型质粒。")
16
3.按质粒转移方式,分为3类 1)接合型质粒(conjugaative plasmid)带有效接触基因质粒,只能使细菌接合,本身不被传递.
2)可移动质粒(mobiliableplasmid)可以被传递,但不能使细菌接合型与可移动性共存时,能传递可移动质粒。 3)自传递质粒(selftran missible plasmid)兼具1)2)两种功能因而可以自传递,如F质粒。 *转化作用(transformation)—这是涉及细菌摄入外源DNA而实现基因转移一种机制。(见第八章)
可移动质粒(mobiliableplasmid)可以被传递,但不能使细菌接合型与可移动性共存时,能传递可移动质粒。 3)自传递质粒(selftran missible plasmid)兼具1)2)两种功能因而可以自传递,如F质粒。 *转化作用(transformation)—这是涉及细菌摄入外源DNA而实现基因转移一种机制。(见第八章)")
17
(三)质粒的功能 质粒的功能主要通过质粒本身携带的基因偏码蛋白质表现出来。携带质粒的宿主细胞可表现出相应表型。 1.性质粒 即雄性细菌F质粒,它本身转到F-宿主细胞时,使后者变成F+,改变宿主细菌性别。 2.抗生素抗性 抗药性(R)质粒使细菌产生抗生素抗性,这种抗药性抗性基因也可以转移到缺乏这种抗药基因的细菌体内,使之产生抗药性。 3.产生毒素的质粒 如col质粒能产生大肠杆菌素因子(colicin),杀死不合该毒素的亲缘细菌。 4.降解复杂的有机化合物作为能源质粒。 5.产生限制和修饰酶。(见第八章)
质粒使细菌产生抗生素抗性,这种抗药性抗性基因也可以转移到缺乏这种抗药基因的细菌体内,使之产生抗药性。 3.产生毒素的质粒 如col质粒能产生大肠杆菌素因子(colicin),杀死不合该毒素的亲缘细菌。 4.降解复杂的有机化合物作为能源质粒。 5.产生限制和修饰酶。(见第八章)")
18
(四)质粒的基本特性 1.自主复制 质粒的复制是自主调节的,不受染色体复制调节因素的影响。 复制调控系统由质粒上的复制起点(ori),质粒的rep基因和cop基因组成。 Rep蛋白启动质粒的复制,cop基因本身或其表达产物可抑制复制作用,从而控制质的拷贝数。 2.质粒的不相容性 利用相同复制系统的质粒不能共存于同一个细胞内。 PMB和COLEI是两个密切相关的复制调控系统,带有PMB和COLEI复制调控系统的质粒是不相容的。但它们与带有PSC101或P15A复制调控系统是完全相容的,可以共存于一个细胞内。不相容性使质粒能够很容易被克隆。 3.质粒的转移性 在自然条件下,在些质粒可以通过细菌接合作用在细菌细胞向传递。基因工程中常用的质粒载体缺乏转移所需的基因(mob基因),不能通过接合作用在细胞间传递,但可采用人工方法转化到细菌细胞中。
,不能通过接合作用在细胞间传递,但可采用人工方法转化到细菌细胞中。")
19
三.转位因子 转位因子(transposable element) 即可移动的基因成分(可移动基因,movable gene mob),是指能够在一个DNA分子内部或两上DNA分子之间移动的DNA片段。在细菌中指在质粒和染色体之间或在质粒和质粒之间移动的DNA片段(文献上有时形象地称其为是跳跃基因,jumping gene)。转位也是DNA重组的一种形式。
 即可移动的基因成分(可移动基因,movable gene mob),是指能够在一个DNA分子内部或两上DNA分子之间移动的DNA片段。在细菌中指在质粒和染色体之间或在质粒和质粒之间移动的DNA片段(文献上有时形象地称其为是跳跃基因,jumping gene)。转位也是DNA重组的一种形式。")
20
移动基因最早由美国冷泉港实验室(cold spring Harbor Laboratory)的女科学家B
移动基因最早由美国冷泉港实验室(cold spring Harbor Laboratory)的女科学家B.MClintock于上个世纪40年代晚期在玉米中首次发现的。60年代,为J.A.Shapirc研究大肠杆菌高效突变实验证实。1983年荣获诺贝尔生物学医学奖。
的女科学家B.MClintock于上个世纪40年代晚期在玉米中首次发现的。60年代,为J.A.Shapirc研究大肠杆菌高效突变实验证实。1983年荣获诺贝尔生物学医学奖。")
21
细菌的转位因子包插入序列,转座子及可转座的噬菌体。 1.插入序列(insertion sequence,IS)
(一)转位因子的种类及特征 细菌的转位因子包插入序列,转座子及可转座的噬菌体。 1.插入序列(insertion sequence,IS) IS的形体图 TS target site靶位点 Transposase gene 转位酶基因 IR inverted repeated 反向(倒转重复顺序) TS Transpcsase gene IR
转位因子的种类及特征. 细菌的转位因子包插入序列,转座子及可转座的噬菌体。 1.插入序列(insertion sequence,IS) IS的形体图. TS target site靶位点. Transposase gene 转位酶基因. IR inverted repeated 反向(倒转重复顺序) TS. Transpcsase gene. IR.")
22
(1)IS是一类较小的转位因子,长度约700-2000bp,按发现顺序IS1、IS2…命名,
只携带转移的必需基因,不含有其它偏码蛋白质结构基因,本身没有表型效应。 (2)IS两侧为反向(倒转)重复顺序(16-41bp),中间为转位酶基因,在插入新的位点侧有3~116p顺向重复顺序(directw repeated sequencedk),DR是靶位点序列复制的产物。 (3)IS到处活动,可以插入到E.coli染色体的各个位置上,也可以插入到质粒和某些噬菌体基因组上,甚至同一基因不同位点上。这种插入作用可以双向进行,可以是正向,也可以是反向插入IS这种移动方式称为转位作用(transposition)。 (4)在一个世代的107细菌中有1次插入。 *TR(反向倒转重复序列):GGAAGGT、、、ACCTTC CCTTCCA、、、TGGAAGG *DR(正同向重复序列):TACGTTACGT
IS两侧为反向(倒转)重复顺序(16-41bp),中间为转位酶基因,在插入新的位点侧有3~116p顺向重复顺序(directw repeated sequencedk),DR是靶位点序列复制的产物。 (3)IS到处活动,可以插入到E.coli染色体的各个位置上,也可以插入到质粒和某些噬菌体基因组上,甚至同一基因不同位点上。这种插入作用可以双向进行,可以是正向,也可以是反向插入IS这种移动方式称为转位作用(transposition)。 (4)在一个世代的107细菌中有1次插入。 *TR(反向倒转重复序列):GGAAGGT、、、ACCTTC. CCTTCCA、、、TGGAAGG. *DR(正同向重复序列):TACGTTACGT.")
23
(2)根据结构特征的不同,Tn可以分为2个亚类:
2.转座子(transposon,Tn) (1)Tn是一类较大的可移动成分,除mob gene外,尚含有其它基因,如抗药基因等。Tn是在研究抗药基因中发现的,由此知道抗药基因可在质粒之间,质粒与染色体之间或质粒与可转座的噬菌体之间来回移动,Tn的转位原理和Is基本相同,转位频率为10-3~10-6/拷贝。 (2)根据结构特征的不同,Tn可以分为2个亚类: ① 复合型Tn:转座酶由IS编码,IS可以是反向(或正向)重复构型。 ② TnA:数个结构基因(mob、mdr等)十IR组成。 IS1 Structural gene IS1 IR Structural genes IR
 (1)Tn是一类较大的可移动成分,除mob gene外,尚含有其它基因,如抗药基因等。Tn是在研究抗药基因中发现的,由此知道抗药基因可在质粒之间,质粒与染色体之间或质粒与可转座的噬菌体之间来回移动,Tn的转位原理和Is基本相同,转位频率为10-3~10-6/拷贝。 (2)根据结构特征的不同,Tn可以分为2个亚类: ① 复合型Tn:转座酶由IS编码,IS可以是反向(或正向)重复构型。 ② TnA:数个结构基因(mob、mdr等)十IR组成。 IS1. Structural gene. IS1. IR. Structural genes. IR.")
24
3.可转座的噬菌体(transposable phage)
(1)包括Mu和D108两种噬菌体,是一类温和噬菌体*。 (2)感染细菌后,可以整合到细菌染色体中,插入位点是随机的(而入phage插入位点是专一的),可以插到结构基因内部,引起突变,Mu即Mutator(突变子)因此得名。 (3)插入部位的2侧有短的DR,插入时,一个拷贝留在原位,新合成的拷贝插入新的部位。 (4)和IS,Tn相比,Mu末端不含IR,这是可转座成分的一个例外。
包括Mu和D108两种噬菌体,是一类温和噬菌体*。 (2)感染细菌后,可以整合到细菌染色体中,插入位点是随机的(而入phage插入位点是专一的),可以插到结构基因内部,引起突变,Mu即Mutator(突变子)因此得名。 (3)插入部位的2侧有短的DR,插入时,一个拷贝留在原位,新合成的拷贝插入新的部位。 (4)和IS,Tn相比,Mu末端不含IR,这是可转座成分的一个例外。")
25
*噬菌体(phage)—是侵袭细菌的病毒,主要由蛋白质和核酸组成。噬菌的生活周期分为①溶菌周期和②溶原周期。
溶原性噬菌体(温和噬菌体)→宿主菌→将自身DNA整合到细菌染色体中→和细菌染色体一起复制→随细菌增殖传到细菌子代中去,不产生子代Phage.
→宿主菌→将自身DNA整合到细菌染色体中→和细菌染色体一起复制→随细菌增殖传到细菌子代中去,不产生子代Phage.")
26
(二)转位作用的机理 1.复制性转位机理 共联体生成和解离,靶序列的切割与复制。 2.非复制型转位作用 转位将供体DNA转座因子两侧各切断一条单链并与靶序列的两个游离末端连接, 随后并没有复制过程,而是由转座酶将供体DNA转座因子的另一端也切断,因此在供体DNA留下一个致死性缺口。 转座子的两条游离单链在靶位点退火接合,DNA聚合酶项平缺口。

27
(三)转位的遗传效应 1.基因重排 可能产生1个新的蛋白分子等,基因重排是进化的动力。 2.基因突变 插入到基因内部,可引起插入失活。 3.插入位点引入新的基因 如引进抗药基因。
转位的遗传效应 1.基因重排 可能产生1个新的蛋白分子等,基因重排是进化的动力。 2.基因突变 插入到基因内部,可引起插入失活。 3.插入位点引入新的基因 如引进抗药基因。")
28
(四)细菌有限制—修饰系统 细菌的限制—修饰系统是分别由特定的基因编码的限制酶和修饰酶组成的二元系统。 1.防御外源性DNA入侵。 2.构成细菌种属和菌株之间交叉繁殖屏障,但又允许外源DNA有某些遗漏,利于物种进化。 3.基因工程重要的工具酶。(350/400) 甲基化酶 1.保护自身DNA不受限制酶切割(限制)。 2.影响DNA分子构象,利于基因表达调控。 限制酶和甲基化酶辩证关系。
 甲基化酶 1.保护自身DNA不受限制酶切割(限制)。 2.影响DNA分子构象,利于基因表达调控。 限制酶和甲基化酶辩证关系。")
29
第二节 真核生物基因组 一.真核生物基因组的结构 (一)真核基因的基本结构 1.结构基因、内含和外显子、断裂基因。
第二节 真核生物基因组 一.真核生物基因组的结构 (一)真核基因的基本结构 1.结构基因、内含和外显子、断裂基因。 (1)结构基因(structural gene)指能转录成为mRNA、rRNA或tRNA的DNA顺序。 (2)内含子和外显子 真核生物的结构基因是不连续的,编码序列被非编码序列打断,在编码序列之间的序列称为内含子(intron),编码序列称为外显子(extron)。 (3)断裂基因(split gene) 在真核类结构基因组中,编码顺序被许多称为内含子的非编码区分割成几段称之。
真核基因的基本结构. 1.结构基因、内含和外显子、断裂基因。 (1)结构基因(structural gene)指能转录成为mRNA、rRNA或tRNA的DNA顺序。 (2)内含子和外显子 真核生物的结构基因是不连续的,编码序列被非编码序列打断,在编码序列之间的序列称为内含子(intron),编码序列称为外显子(extron)。 (3)断裂基因(split gene) 在真核类结构基因组中,编码顺序被许多称为内含子的非编码区分割成几段称之。")
30
2.顺式调控元件 顺式调控元件(cis—acting elements) 指与结构基因表达调控相关,能够被基因调控蛋白特异性识别和结合的DNA序列。能与顺式作用元件结合调节基因转录活性的蛋白质因子称为反式作用因子(trans—acting factors)。顺式调控元件有:
 指与结构基因表达调控相关,能够被基因调控蛋白特异性识别和结合的DNA序列。能与顺式作用元件结合调节基因转录活性的蛋白质因子称为反式作用因子(trans—acting factors)。顺式调控元件有:")
31
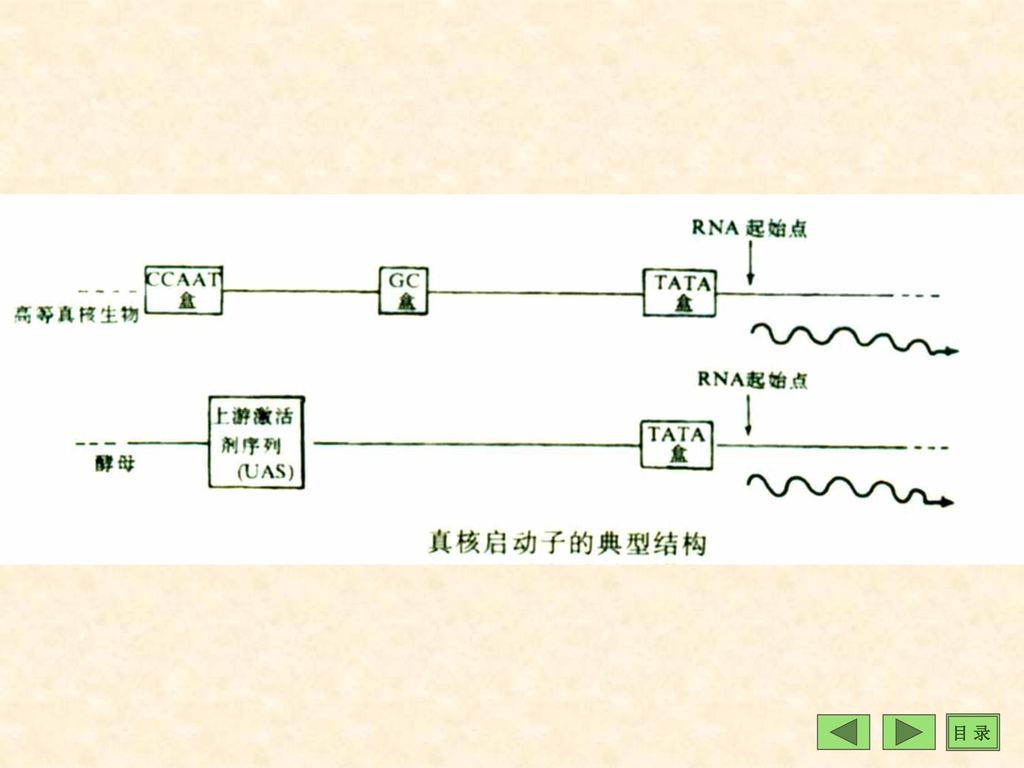
(1)启动子(promoter) ①概念:启动子是促进DNA转录的DNA序列,是DNA分子上可与RNA pol 特异性识别结合并使之转录的部位,但启动子本身不被转录。 ②功能特点:启动子位于结构基因上游启动子有方向性决定转录方向及那一条DNA链作模板转录(以信息链的互补链作模板转录,转录的mRNA与信息链一致)。 ③真核生物的启动子元件是TATA box TATA盒与TATA因子的转录因子结合后即成为完整的启动子。(见第六章)
。 ③真核生物的启动子元件是TATA box TATA盒与TATA因子的转录因子结合后即成为完整的启动子。(见第六章)")
32
(2)上游启动子元件(upstream promoter elements ups)
①UPS是TATA盒上游的一些特定的DNA序列。 ②反式作用因子可与这些元件结合,通过调节TATA因子与TATA box的结合、RNA pol与启动子结合及转录起始复合物形式来调控基因转录效率。

33
(3)反应元件(response elements)
一些信息分子的受体被细胞外信息分子激活后,能与特异的DNA序列结合,调控基因的表达。 这种DNA序列实际上也是顺式元件,由于能介导基因对细胞外的某种信号产生反应,被称为反应元件。 反应元件都具有较短的保守序列。这些无件通常位于启动子附近和增强子内,有不少是回文序列。

34
(4)增强子(enhancer)和沉默子(silencer)
增强子是一段DNA序列,其中含有多个能被反式作用因子识别与结合的顺式作用元件。反式作用因子与这些元件结合后,通常为增强邻近基因的转录。增强子一般位于转录起始点上游-100~-300bp处,但在基因之外或某些内含子中也有增强子序列(P42)。 增强子作用特点:①可在5’端或3’端发挥作用;②不受序列方向制约;③通过增强启动子发挥作用。沉默子 负调控序列、负增强子;
。 增强子作用特点:①可在5’端或3’端发挥作用;②不受序列方向制约;③通过增强启动子发挥作用。沉默子 负调控序列、负增强子;")
35
(5)加尾信号 在结构基因的最后一个外显子中有一个保守的AATAAA序列,此位点下游有一段GT丰富区或T丰富区,这两部分序列共同构成poly(A)加尾信号。 mRNA转录到此部位后,产生AAUAAA和随后的GU(或U)丰富区。与RNApol结合的延长因子可以识别这种结构并与之结合,然后在AAuAAA下游10-30个碱基的部位切断RNA,并加上poly(A)尾.
丰富区。与RNApol结合的延长因子可以识别这种结构并与之结合,然后在AAuAAA下游10-30个碱基的部位切断RNA,并加上poly(A)尾.")
36
(二)基因家族(gene family) 基因家族 是指核苷酸序列或编码产物具有一定程度同源性的一组基因. 基因家族 中各个基因之间的关系: 1.家族中各基因的核苷酸序列相同这些基因族也被称为单纯多基因家庭(如rRNA,tRNA家族)和复合多基因家族(如组蛋白基因家族). tRNA基因: 人类基因约有1300个tRNA基因,编码50多种tRNA。每种tRNA可有10-几百个基因拷贝。同种tRNA往往串联在一起形成基因簇,但基因间有非转录间隔区分隔,常常比结构基因长近10倍。

37
2.家族中各基因核苷酸序列高度同源 (1)人类生长激素基因家族 包括人生长激素(hGh)、人胎盘促乳素和催乳素(prolactin)。它们之间的同源性很高,尤其是hGh和hcs之间,蛋白质氨基酸序列有85%的同源性,mRNA上序列上有92%的同源性,说明它们是来自一个共同祖先基因。3种基因并不都排列在一起,hGh和hcs基因位于第17号染色体长臂,催乳素基因位于第6号染色体。 (2)α-株蛋白和β-株蛋白基因家族 这些基因家族的各个成员在DNA分子上的排列顺序按照发育的不同阶段先后次序排列,故也称“发育控制复合多基因家族”。
、人胎盘促乳素和催乳素(prolactin)。它们之间的同源性很高,尤其是hGh和hcs之间,蛋白质氨基酸序列有85%的同源性,mRNA上序列上有92%的同源性,说明它们是来自一个共同祖先基因。3种基因并不都排列在一起,hGh和hcs基因位于第17号染色体长臂,催乳素基因位于第6号染色体。 (2)α-株蛋白和β-株蛋白基因家族. 这些基因家族的各个成员在DNA分子上的排列顺序按照发育的不同阶段先后次序排列,故也称 发育控制复合多基因家族 。")
38
3.家族中各基因编码的蛋白质有高度的同源性,但基因的核苷酸序列可能不同。
如src癌基因家族 src,abl,fes,fgr,fps,fym,kck,lyn,ros,tkl,yes 此家族中各基因的DNA序列没有明显的同源性。 但每个基因产物都含有250个氨基酸顺序的同源蛋白激酶结构域。 4.家族各基因编码的蛋白质中具有很小的保守基序(conserved motif)。 如DEAD box基因家族。DEAD box:Asp-Glu-Ala-Asp. 此家族中各基因的DNA序列没有明显的同源性,但所有的表达产物都具有解旋酶的功能,都具有同样的保守基序(DEAD盒),DEAD是酶活性的关键结构。
。 如DEAD box基因家族。DEAD box:Asp-Glu-Ala-Asp. 此家族中各基因的DNA序列没有明显的同源性,但所有的表达产物都具有解旋酶的功能,都具有同样的保守基序(DEAD盒),DEAD是酶活性的关键结构。")
39
5.基因超家族(gene superfamily)
基因超家族 是指一组由多基因家族及单基因家族组成的更大的基因家族。它们的结构有程度不等的同源性,因此它们可能起源于相同的祖先基因,但是它们的功能并不一定相同,这一点正是与多基因家族的差别所在。这些基因在进化上也有亲缘关系,但亲缘关系较远,故将其称为基因超家族。如: (1)免疫球蛋白超基因家族 表达产物都有免疫球蛋白样的结构域结构。有2个微球蛋白、MHCI类抗原的α链,Ⅱ类抗原的α链和β链,Thy1、CD4、CD8等与免疫有关的分子。 在后又陆续发现了许多免疫系统内以及与免疫无关的家族成员。
免疫球蛋白超基因家族 表达产物都有免疫球蛋白样的结构域结构。有2个微球蛋白、MHCI类抗原的α链,Ⅱ类抗原的α链和β链,Thy1、CD4、CD8等与免疫有关的分子。 在后又陆续发现了许多免疫系统内以及与免疫无关的家族成员。")
40
(2)丝氨酸蛋白酶基因超家族 其基因产物都有一个特殊的功能区,具有酶的功能。功能区中丝氨酸是活性中心瓣关键氨基酸残基。
现有很多新成员加进去,如载脂蛋白(apolipoprotein),它们只是转移胆因醇蛋白颗粒中的成分而不具备任何水解蛋白质的酶功能。 (3)信号传递途径中的小GTP接合蛋白,现在称为ras超家族,包括ras家族、rho家族、rab家族,大约已有30个蛋白质基因被确定为该家族成员。
,它们只是转移胆因醇蛋白颗粒中的成分而不具备任何水解蛋白质的酶功能。 (3)信号传递途径中的小GTP接合蛋白,现在称为ras超家族,包括ras家族、rho家族、rab家族,大约已有30个蛋白质基因被确定为该家族成员。")
41
(三)假基因(pseudogene) 1.假基因 在多基因家族中某些与正常功能基因在核苷酸序列上相似,但不能转录或转录后生成无功能基因产物的DNA序列,被称为假基因。 DNA序列,被称为假基因。 2.假基因常用符号ψ表示,如ψα1 表示与α1 相似的假基因. 3.假基因与有功能的基因同源,原来也可以是有功能的基因,由于发生缺失(deletion)、例位(inversion)或点突变(point mutaion)等,成为无功能的基因,即形成了假基因,哺乳动物基因组中的1/4基因为假基因,可能为进化的痕迹。
、例位(inversion)或点突变(point mutaion)等,成为无功能的基因,即形成了假基因,哺乳动物基因组中的1/4基因为假基因,可能为进化的痕迹。")
42
(四)编码序列 真核生物基因组编码序列只占DNA总量的5%,但由于其基因组非常大,故其基因数量就原核生物基因组要大得多(几十倍)。
编码序列 真核生物基因组编码序列只占DNA总量的5%,但由于其基因组非常大,故其基因数量就原核生物基因组要大得多(几十倍)。")
43
(五)真核基因组中的转座子 在真核基因组中,编码序列在染色体中的位置相对比较稳定,但一些中度重复序列往往是可移动的。 在些可移动成分的结构与原核基因组的转位因子相似,是通过DNA介导的。 而另外一些中度重复序列的转移成分,要先转录成RNA,再逆转录生成cDNA,然后重新整合到基因组中,这种逆转录旁路的转移成分称为逆转录座子(retroposon),是RNA介导的。
,是RNA介导的。")
44
(六)重复序列(repeat sequence)
重复序列中,除了编码rRNA 、tRNA 、组蛋白及免疫球蛋白的结构基因外,大部分是非编码序列。 它们的功能 主要与基因组的结构稳定性,组织形式以及基因表达调控有关。 目前已发现一些重复序列的特征与遗传有密切联系,因此可以通过测定重复次数而协助遗传病的诊断。

45
据出现的频率不同可将DNA序列分为3类: 1.高度重复序列 在基因组中的重复次数 >105 2.中度重复序列 在基因组中的重复次数为 3.单拷贝序列 在整个基因组中出现1次或少数几次( )。
。 .")
46
(七)端粒(telommere) 以线性染色体形式存在的真核基因组DNA的末端都有一种特殊的结构,称为端粒。(形式上膨大成粒状而得名)。 结构 1.是染色体末端DNA和蛋白质构成复合体。仅在真核细胞染色体末端存在。 2.其DNA序列相当保守,一般由多个串联在一起的短寡核苷酸(5-86p)序列构成。 3.碱基成分因种属而异,重复次数在不同生物中变化较大,如小鼠的端粒DNA达150kb,人类的端粒DNA约5~15kb。 功能 1.保护线性DNA的完整复制 2.保护染色体末端及决定细胞的寿命等。
序列构成。 3.碱基成分因种属而异,重复次数在不同生物中变化较大,如小鼠的端粒DNA达150kb,人类的端粒DNA约5~15kb。 功能 1.保护线性DNA的完整复制. 2.保护染色体末端及决定细胞的寿命等。")
47
二.人类基因组中的重复顺序 (一)反向重复顺序(inverted repeats,IR) ATTAGC GCTAAT ATTAGC GGAT GCTAAT TAATCG CGATTA TAATCG CCTA CGATTA 1.连续的反向重复顺序,这种结构又称回文结构(palindrome),是指一段DNA顺序,在两条链上,正读与反读意义相同。 2.不连续的反向重复顺序之间含有间隔顺序。 1.2占人类基因组5%,可能与复制、转录调控有关。
,是指一段DNA顺序,在两条链上,正读与反读意义相同。 2.不连续的反向重复顺序之间含有间隔顺序。 1.2占人类基因组5%,可能与复制、转录调控有关。")
48
(二)串联重复顺序(tandem repeats)
1.编码区串联重复顺序 如组蛋白基因、5srRNA基因等。其意义在于快速大量合成相应基因的mRNA. 2.非编码区串联重复顺序 通常存在于间隔DNA*和内含子内,是组成卫得DNA*的基础。卫星DNA可分为三类,大卫星DNA、小卫星DNA和微卫星DNA。

49
*间隔DNA(spacer DNA)—真核基因组中,在基因之间也有一些非编码顺序将它们隔开,一般称为间隔DNA,是和内含子性质不同的插入顺序。
*卫星DNA(satellite DNA sat -DNA)—又称随体DNA。这部分DNA是在用cscl密度梯度离心时发现的。
—又称随体DNA。这部分DNA是在用cscl密度梯度离心时发现的。")
50
大肠杆菌DNA剪切成若干片段后离心只得到1个峰,而蟹DNA在主峰旁边还有1个小峰,其中所含DAN称sat-DNA.
Sat-DNA的A+T/G+C比值不同于主峰DNA的比值,因而其密度也不同于主峰DNA。比值改变原因是它们含有大量的重复顺序而使某段DNA分子(A+T)或(G+C) 偏低或偏高。(下图因比值远远大于主峰DNA,导致密度较低所致)。
或(G+C) 偏低或偏高。(下图因比值远远大于主峰DNA,导致密度较低所致)。")
51
(三)散在重复顺序(intersprersed repeats)
散在重复顺序是人类基因组中非串联非反向的重复顺序,包括少数活跃的转位因子,根据重复序列的长度可将该家族分为2个主要类型,短散在核元件和长散在核元件。 1.SINEs(shat interspersed nuclear elements) LINES(long interspersed nuclear elements) 代表 Alu家族(逆转录转座子) KpnI家族 (1)序列中含Alu限制酶位点 (1)序列中含KpnⅠ限制酶位点 (2)基因组中重复数3-5× (2)全长6-7kb (3)Alu顺序之间间隔:3-5kb (3)Knp Ⅰ消化后可见4条带 (4)相对集中在染色体R带 (4)集中分布在染色体G或Q带 逆转来转座子 RNA介导转座 ,合成cDNA→基因组
 2.LINES(long interspersed nuclear elements) 代表 Alu家族(逆转录转座子) KpnI家族. (1)序列中含Alu限制酶位点 (1)序列中含KpnⅠ限制酶位点. (2)基因组中重复数3-5×105 (2)全长6-7kb. (3)Alu顺序之间间隔:3-5kb (3)Knp Ⅰ消化后可见4条带. (4)相对集中在染色体R带 (4)集中分布在染色体G或Q带. 逆转来转座子 RNA介导转座 ,合成cDNA→基因组.")
52
三.人类基因组DNA多态性 某一机体可以观察到的特征,称为表现(phenotype),与表型相应的基因组成称为基因型(genotype).人类个体之间所以表现为千差万别,其物质基础在于基因组DNA的差异. DNA序列的多态性就是这些差异中十分重要的1种,DNA序列多态性可分为位点的多态性和串联重复顺序多态性,前者又可造成限制性长段长度多态性.

53
(一)DNA位点多态性(DNA site polymorphism)
DNA位点多态性 是由于等位基因间在特定位点上DNA序列存在差异造成的.在各种DNA位点多态性系统中,HLA(人类白细胞抗原)是最复杂的一种. 就其表型而言,从理论上来说,已远远超过了现有地球人口,人类为远交群体,因此要在无血缘关系人群找到HLA表型完全相同的个体几乎是不可能的.(见分子免疫学).
是最复杂的一种. 就其表型而言,从理论上来说,已远远超过了现有地球人口,人类为远交群体,因此要在无血缘关系人群找到HLA表型完全相同的个体几乎是不可能的.(见分子免疫学).")
54
(二)限制性中段长度多态性(restiction fragment length polymorplism,RFLP)
1.DNA位点的多态性可影响限制酶的切割位点,造成限制片段长度多态性,即用同一种限制酶消化不同个体DNA时,会得到长度各不相同的限片段类型。 是DNA多态性的一种特别有用的形式,当DNA分子由于中性突变,使某种限制酶切点数增加,减少或移位,导致限制性片段长度发生改变(也称为RFLP),就会得到不同限制性片段类型,这样的位点称为多态性位点。
,就会得到不同限制性片段类型,这样的位点称为多态性位点。")
55
(三)重复顺序长度多态性 1.小卫星DNA多态性很复杂,个体之间的差异极高。 2.微卫星DNA多态性,功能不太清楚. (四) DNA位点的多态性可以作为遗传标记,用于基因诊断或个体鉴定,其与疾病的相关研究正在进行之中。
 DNA位点的多态性可以作为遗传标记,用于基因诊断或个体鉴定,其与疾病的相关研究正在进行之中。 .")
56
本节概要地介绍了对人类基因组研究和认识,但显然还有大量的未知,比如仅弄清楚了极有限的功能基因,一小部分基因序列,距离彻底了解人类自身遗传背景还很遥远。正是存在这种科学内在的驱动力,人类在20世纪末开始了的探索自身为主要目的的伟大科学计划—人类基因组计划,其重要意义完全可以与原子弹计划和阿波罗登月计划相比。

57
第三节 病毒基因组 基因组(genome) 1个配(精子或卵子),1个单倍体细胞或1个病毒所包含的全套基因。病毒核酸或为DNA或为RNA,可以统称为病毒染色体。 完整的病毒颗粒具有蛋白质外壳,以保护病毒核酸不受核酸酶的破坏,并能识别和侵袭特定的宿主。类病毒例外,它是一类植物病毒,为很小的单链闭环RNA( 碱基)。
。")
58
病毒核酸与所有的原、真核生物的核酸组比较,最为突出的特点是每种病毒颗粒只含1种核酸,或DNA或RNA,两者不共存于1种病毒颗粒中,据此,病毒可以分成两组,即DNA病毒和RNA病毒。
病毒核酸分子量大小:RNA病毒小 D,DNA病毒大; D 基因数:3-几百个

59
一切烈性病毒(virulent virus)都具备参下几个时相的生活周期:(1)吸附;(2)核酸进入细胞;(3)转录、翻译和复制;(4)病毒颗粒成熟;(5)释放病毒颗粒。
典型的生活周期为6-48小时,噬菌体为20-60分钟。 病毒对细胞功能的干予,最典型的是噬菌体(phage),当其入菌后,立即接管了细菌的各种主要功能,细菌自身的DNA复制RNA转录,蛋白质合成,能量代谢等都停顿下来转变成合成病毒的机器。有些动物病毒也有类似的情况,但非普通规律。相对而言,动物病毒和宿主细胞可以共存一段时间至病毒生活周期的后期才造成宿主细胞的严重损害,这些损害有如下3个类型: (1)抑制宿主RNA和DNA的合成。 (2)核糖体合成受损。 (3)蛋白质合成受抑制是普遍现象。 与人类疾病有关的病毒属于动物病毒,它包括8个DNA病毒科和12个RNA病毒科,这些病毒基因组结构和功能已基本清楚(见分子病毒学HIV、没办法)。
,当其入菌后,立即接管了细菌的各种主要功能,细菌自身的DNA复制RNA转录,蛋白质合成,能量代谢等都停顿下来转变成合成病毒的机器。有些动物病毒也有类似的情况,但非普通规律。相对而言,动物病毒和宿主细胞可以共存一段时间至病毒生活周期的后期才造成宿主细胞的严重损害,这些损害有如下3个类型: (1)抑制宿主RNA和DNA的合成。 (2)核糖体合成受损。 (3)蛋白质合成受抑制是普遍现象。 与人类疾病有关的病毒属于动物病毒,它包括8个DNA病毒科和12个RNA病毒科,这些病毒基因组结构和功能已基本清楚(见分子病毒学HIV、没办法)。")
60
(二)病毒有(十)链RNA和(一)链RNA之分。
概念 极性和mRNA极性相同, 与mRNA极性相反或与(十)RNA 碱基顺序也相同。 互补的RNA。 作用 1.作为模板(与mRNA一样) 必须依赖 翻译出病毒多肽或蛋白质。 RNA的RNA 2.作为模板,合成(一)RNA, pol合成(十) 可以以(一)RNA为模板 RNA(mRNA) 合成子代病毒RNA。 才能翻译出 3.作为模板合成cDNA 病毒 (即以mRNA为模板反转录合成DNA)。 蛋白质。
RNA. 碱基顺序也相同。 互补的RNA。 作用 1.作为模板(与mRNA一样) 必须依赖. 翻译出病毒多肽或蛋白质。 RNA的RNA. 2.作为模板,合成(一)RNA, pol合成(十) 可以以(一)RNA为模板 RNA(mRNA) 合成子代病毒RNA。 才能翻译出. 3.作为模板合成cDNA. 病毒. (即以mRNA为模板反转录合成DNA)。 蛋白质。")
61
二.病毒基因组结构功能特点(P28) (一)病毒核酸可以是ssDNA、dsDNA或RNA分子,分子结构有发夹、环状、线型、节段型。以SSDNA,dsRNA最为突出。 (二)基因重叠 即同一段基因可以编码2种或以上的基因产物这种现象在其它生物细胞仅见于线粒体和质体DNA.所以是病毒核酸较为独特结构能使小小病毒携带较多的遗传信息,原因是病毒基因阅读框可以错位(SV40)。
基因重叠 即同一段基因可以编码2种或以上的基因产物这种现象在其它生物细胞仅见于线粒体和质体DNA.所以是病毒核酸较为独特结构能使小小病毒携带较多的遗传信息,原因是病毒基因阅读框可以错位(SV40)。")
62
(三)连续的和不连续的基因 病毒基因结构特征往往与其宿主细胞基因结构相似。
原核病毒(如噬菌体)基因是连续的,没有内含子; 真核病毒(如多瘤病毒)基因是不连续的,有内含子。 有意思的是,有些真核病毒的内含子或其中的一部分对某一基因来说是内含子,对另一基因却是外显子。 如SV40和多瘤病毒的早期区域就是这样的。 除了(+)RNA病毒外,真核病毒基因都是先转成mRNA前体,再经过剪接等步骤能成为成熟的mRNA.
基因是连续的,没有内含子; 真核病毒(如多瘤病毒)基因是不连续的,有内含子。 有意思的是,有些真核病毒的内含子或其中的一部分对某一基因来说是内含子,对另一基因却是外显子。 如SV40和多瘤病毒的早期区域就是这样的。 除了(+)RNA病毒外,真核病毒基因都是先转成mRNA前体,再经过剪接等步骤能成为成熟的mRNA.")
63
(四)节段性基因 大部分病毒核酸都是由一条双链或单链构成,少数病毒核酸由数个片段构成。这类病毒一般度RNA病毒。如flu-v由6-7个片段构成,各段在天然状态下不连接,而且可以转录成6-7个片段相应的 mRNA。单独的片段没有感染性,感染要一起感染才发挥作用。 (五)除了retro-v外,所有的病毒基因都是单倍体基因。即每个基因在某个病毒颗粒中只出现一次,即只有1套基因。 (六)编码区>非编码区(95%/5%)。病毒核酸大多数顺序都用来编码蛋白质。
除了retro-v外,所有的病毒基因都是单倍体基因。即每个基因在某个病毒颗粒中只出现一次,即只有1套基因。 (六)编码区>非编码区(95%/5%)。病毒核酸大多数顺序都用来编码蛋白质。")
64
(七)基因常常成簇排列,没有间隔序列或间隔序列很小。功能相关蛋白质基因在基因组的1个或几个特定部位,丛集成簇被转录成多顺反子,然后加工成各种蛋白质的mRNA模板。如腺病毒晚期基因。
(八)不规则的结构基因 1.几个结构基因的编码区不规则,因此有些结构基因无翻译起始序列。 2.有的mRNA(=gene)没有5’帽子,但有翻译增强子。 (九)DNA或RNA 1种病毒基因组只是1种核酸。
不规则的结构基因. 1.几个结构基因的编码区不规则,因此有些结构基因无翻译起始序列。 2.有的mRNA(=gene)没有5’帽子,但有翻译增强子。 (九)DNA或RNA 1种病毒基因组只是1种核酸。")
65
三、典型病毒基因组 (一)SV40病毒基因组 SV40(simian sarcom avirus40 猴肉瘤病毒)属乳多空病毒类,是最小的DNA病毒之一。它可长期潜伏于猴肾细胞,对新生仓鼠有致癌性,体外试验还可使多种属细胞恶性转化。
属乳多空病毒类,是最小的DNA病毒之一。它可长期潜伏于猴肾细胞,对新生仓鼠有致癌性,体外试验还可使多种属细胞恶性转化。 .")
66
1、基因组结构特点 (1)双链环状DNA分子、MW3×103KD,长52436p,与4种组蛋白(H2A、H2B、H3及H4)综合,与真核染色质区别在于不含H1。 (2)基因组由早期基因、调控区和晚期基因构成。 ①早期基因含有2个重叠基因,编码T和t(T=tumor)抗原 ②晚期基因含3个重叠基因,编码VP1(主),VP2和VP3(次)3种衣壳蛋白 ③调控区位早、晚期基因之间,长约4006p包括复制起点,启动子和增强子,可调节基因组的复制及早、晚期基因的转录。
抗原. ②晚期基因含3个重叠基因,编码VP1(主),VP2和VP3(次)3种衣壳蛋白. ③调控区位早、晚期基因之间,长约4006p包括复制起点,启动子和增强子,可调节基因组的复制及早、晚期基因的转录。")
67
2、SV40病毒基因的表达 (1)启动子位于调控区复制起点上游,其中含有RNA pdII识别位点位于-21-26bp之间的TATA盒可启动RNA转录起始准确位置,一般不相差10个核苷酸。 (2)增强子 在 位是2个串联的含有72bp的完全重复顺序,即为增强子,能增强SV40及异源基因的表达。 (3)早期基因转录的mRNA前体选择剪接形成不同的mRNA(TmRNA、t-mRNA)
增强子 在 位是2个串联的含有72bp的完全重复顺序,即为增强子,能增强SV40及异源基因的表达。 (3)早期基因转录的mRNA前体选择剪接形成不同的mRNA(TmRNA、t-mRNA)")
68
3、病毒基因组的复制及基因表达调控 T抗原控制病毒的复制,启动晚期基因转录。 (1)早期基因T及t的表达受T抗原的反馈阻遇,表现一种自我调节作用。 (2)晚期基因表达需要T抗原及SV40DNA本身的复制。 一般过程是,感染早期合成T、t抗原→T-方面抑制早期mRNA进一步合成;另一方激活多种与DNA复制有关的酶,引发病毒的复制→进而推动晚期RNA的合成,产生Vp蛋白(12h)→V-DNA+VP123→病毒颗粒。
→V-DNA+VP123→病毒颗粒。")
69
(二)HBV基因组 HBV是乙肝的病原体,它可造成持续性感染,引起急、慢性肝炎,部分患者可发生肝硬化,HBV原发性肝癌发生密切相关,我国HBSAg携带者高达10%,因而我国特别重视HBV的研究。
HBV基因组 HBV是乙肝的病原体,它可造成持续性感染,引起急、慢性肝炎,部分患者可发生肝硬化,HBV原发性肝癌发生密切相关,我国HBSAg携带者高达10%,因而我国特别重视HBV的研究。")
70
1、HBV基本结构 (1)电镜下观察 ①呈园球形,直径约42nm,称Dane颗粒(以发现者命名); ②中间含1个致密的内核(core),直径约27nm; ③外为1层约14nm外壳(capsid)。 (2)①表面有HBsAg(hepatitis B virhs surface antigen); ②核心有HBcAg和HBeAg; ③含DNA pol、prorein kinase(蛋白激酶)和reverse transcriptase(逆转录酶),后者与病毒复制特点有关。
①表面有HBsAg(hepatitis B virhs surface antigen); ②核心有HBcAg和HBeAg; ③含DNA pol、prorein kinase(蛋白激酶)和reverse transcriptase(逆转录酶),后者与病毒复制特点有关。")
71
2、HBV基因组结构 (1) ds(±)DNA组成的环状结构,开环不封闭。侵入细胞后借助胞内DNA修复机制转变成cccDNA分子(共价闭合环状超螺旋)。 (2)基因组双链DNA长短不一,外长内短,长负(链)短正(链),长链2/3是双链,1/3是单链。两链间隙由DNApol填充(上(1)述)。 (3)长链(负链)基因重叠是转录mRNA的模板链.
长链(负链)基因重叠是转录mRNA的模板链.")
72
不同毒株均含有4个开放性阅读框(openreading frame, ORF见第五章)S. C. P. X,其中P区与S. C. X
不同毒株均含有4个开放性阅读框(openreading frame, ORF见第五章)S.C.P.X,其中P区与S.C.X.互相重叠50%以上,故多个ORF可读码2次以上,编码2个以上蛋白质。其中: S区 编码主要蛋白为HBsAg C区 编码HBcAg等。 P区 编码DNApol是依赖RNA的DNApol(逆转录酶)。 X蛋白功能不详。
S.C.P.X,其中P区与S.C.X.互相重叠50%以上,故多个ORF可读码2次以上,编码2个以上蛋白质。其中: S区 编码主要蛋白为HBsAg. C区 编码HBcAg等。 P区 编码DNApol是依赖RNA的DNApol(逆转录酶)。 X蛋白功能不详。")
73
4、HBV与肝癌的关系 现在普遍认为,HBV感染与原发性肝癌关系密切,流行病学及分子病毒学的证据如下: (1)原发性肝癌(肝细胞性肝癌)HBSAg高达85%,其他肿瘤患者仅为15%; (2)培养肝癌细胞株,不断释放出HBsAg(+); (3)cDNA 探针检查到HBV已整合到肝癌细胞DNA中去; (4)肝癌病人的癌基因,如Ki-ras、Ha-ras、myc被激活。
cDNA 探针检查到HBV已整合到肝癌细胞DNA中去; (4)肝癌病人的癌基因,如Ki-ras、Ha-ras、myc被激活。")
74
(三)脊髓灰质炎病毒基因组(ss(+)RNA)
1、基因组结构特点 (1)ss(+)RNA,MW2.5X106。 (2)5’端无帽,3’端有poly(A)尾。 5’端2个uu总是 VPG蛋白,病毒入胞后被水解,5’端形成翻译增强子结构,结合核糖体开始翻译。 (3)基因组7个基因编码区无间隔,编码RNA复制酶、蛋白酶和衣壳蛋白。 连接
ss(+)RNA,MW2.5X106。 (2)5’端无帽,3’端有poly(A)尾。 5’端2个uu总是 VPG蛋白,病毒入胞后被水解,5’端形成翻译增强子结构,结合核糖体开始翻译。 (3)基因组7个基因编码区无间隔,编码RNA复制酶、蛋白酶和衣壳蛋白。 连接.")
75
(1)RNA复制的引物不是RNA或DNA,而是VPG-uu (+)RNA (一)RNA 新的(+)RNA
2、基因组信息传递的特点 (1)RNA复制的引物不是RNA或DNA,而是VPG-uu (+)RNA (一)RNA 新的(+)RNA (2)新合成(+)RNA3种功用(新合成poly(A)尾是直接复制出来的): ①作为mNA翻译病毒蛋白质; ②作为模板复制(一)RNA; ③作为子代病毒RNA,与蛋白质包装成病毒颗粒。 RNA复制酶
RNA复制的引物不是RNA或DNA,而是VPG-uu. (+)RNA (一)RNA 新的(+)RNA. (2)新合成(+)RNA3种功用(新合成poly(A)尾是直接复制出来的): ①作为mNA翻译病毒蛋白质; ②作为模板复制(一)RNA; ③作为子代病毒RNA,与蛋白质包装成病毒颗粒。 RNA复制酶.")
76
(四)逆转录病毒 1.逆转录病毒一般分类 逆转录病毒可以说就是RNA肿瘤病毒。 第一个被发现的RNA肿瘤病毒是劳氏肉瘤病毒(Rous Sarcoma vivus, RSV, 1911)。至今为止所发现的retro-v都能使动物致癌,如下。 (1)肉瘤病毒 (2)白血病病毒(leuremia virus) (3)乳腺瘤病毒(mammazy tumor virus)和淋巴瘤病毒,如鸟类髓细胞瘤病毒(avian myelocytoma, AMV) (4)人类嗜T细胞逆转录病毒(human T-lymphotropic retro-v, HTLV)HTLV-III现称人类免疫缺陷综合症病毒(human immunodeficiency virus, HIV)或AIDS病毒。
肉瘤病毒. (2)白血病病毒(leuremia virus) (3)乳腺瘤病毒(mammazy tumor virus)和淋巴瘤病毒,如鸟类髓细胞瘤病毒(avian myelocytoma, AMV) (4)人类嗜T细胞逆转录病毒(human T-lymphotropic retro-v, HTLV)HTLV-III现称人类免疫缺陷综合症病毒(human immunodeficiency virus, HIV)或AIDS病毒。")
77
2.逆转录病毒基结构特征 (1)是RNA的复合体 ①2条(+)RNA单体正向平行,携带所有的遗传信息,而且完全相同,为什么要求2条相同正链,意义不太清楚(怕基因丢失或损伤?)。 ②2条tRNA较小,是宿主tRNA(RSV是tRNAtrp),+RNA对于复制位置是至关重要。 (2)5’端有m7Gppp帽,3’端有Poly(A)尾。
5’端有m7Gppp帽,3’端有Poly(A)尾。")
78
(3)编码区(反式功能区)从5’→3’ 非编码区(顺序功能区,调控区)5’,3’都有
①gag(groupantigen)编码衣壳蛋 ①R区 逆转录合成cDNA必须区域 ②Pov(Polyrmerase) 逆转录酶 ②PB区 引物结合区 ③env(envelop) 包膜蛋白 ③U区 U3含启动子,U5与转录终止加尾有关 *④ONC(oncogene) 癌基因 ④ψ(包装信号),DLS(氢链结合位点) 如RSV是SRC C为调控区 *有的retro-V有ONC。
编码衣壳蛋 ①R区 逆转录合成cDNA必须区域. ②Pov(Polyrmerase) 逆转录酶 ②PB区 引物结合区. ③env(envelop) 包膜蛋白 ③U区 U3含启动子,U5与转录终止加尾有关. *④ONC(oncogene) 癌基因 ④ψ(包装信号),DLS(氢链结合位点) 如RSV是SRC C为调控区. *有的retro-V有ONC。")
79
3.逆转录病毒生活周期 (1)吸附 可能所有的动物病毒受体都是表面糖蛋白(MLV的受体是左旋氨基酸转运蛋白,受体与基因治疗)。 (2)入胞 逆转录,转录,翻译。 ①逆转录——前病毒基因组(cDNA)的形成 RNA+逆转录酶→宿主胞浆→逆转录→(RNA为模板,+RNA为引物)→cDNA(见第四章)→进入核→随机整合宿主染色体→形成前病毒。(phag仅注入核酸) cDNA形成新的重复(U3-R-U5)称LTR(long terminal repeat 长末端重复顺序)。cDNA含有RNA全部信息且较RNA长。
→cDNA(见第四章)→进入核→随机整合宿主染色体→形成前病毒。(phag仅注入核酸) cDNA形成新的重复(U3-R-U5)称LTR(long terminal repeat 长末端重复顺序)。cDNA含有RNA全部信息且较RNA长。")
80
逆转录酶除了逆转活性外,还至少还有3种活性:即RNaseH活性,内切核酸酶活性(特异地切断基因组3’端RNA,作为“+”链DNA的引物)和以“-”链DNA为模板合成“+”链DNA的活性。
和以 - 链DNA为模板合成 + 链DNA的活性。")
81
②转录——病毒基因组的形成 U3区启动子启动前病毒基因组转录出从5’区至3’R区的RNA。 ③翻译gag、env、pol gag —— 全长RNA能直接作为gag基因mRNA,进行翻译。但有90%的mRNA只译出gag多蛋白质,经肽链加工裂解为衣壳蛋白。 pol —— RNA序列有干扰gag终止码的序列,使约10%mRNA在翻译过程中,直至pol的终止码,经翻译后加工裂解,形成衣壳蛋白,逆转录酶和整合酶(integrase)。 env —— 一部分全长mRNA经过转录后的剪接,使env编码区与病毒RNA5’端帽子连接在一起→env mRNA →包膜糖蛋白。
。 env —— 一部分全长mRNA经过转录后的剪接,使env编码区与病毒RNA5’端帽子连接在一起→env mRNA →包膜糖蛋白。")
82
(3)病毒颗粒成熟←包装← ψ(基因治疗) RNA5’端PB-区与宿主tRNA以氢链相连形成复合体→包装→病毒颗粒。 (4)释放子病毒→感染其它细胞。
病毒颗粒成熟←包装← ψ(基因治疗) RNA5’端PB-区与宿主tRNA以氢链相连形成复合体→包装→病毒颗粒。 (4)释放子病毒→感染其它细胞。")
83
4.逆转录病毒引起白血病的3种机理.(以此说明RNA肿瘤病毒的致癌作用与基因组类型.)
(1)急性逆转录病毒 (2)慢性逆转录病毒 (3)HTlV(人类嗜T细胞逆转录病毒) ①含onc* ①不含onc ①含tat的HTLV ②可在数周内使培养 ②不能转化,但V感染 ②含使培养细胞转化,机理和1、2 细胞发生转化 整体动物潜伏数月致癌. 不同。 ③onc插到宿主DNA中 ③前V整合到proto-onc ③反式调控作用,tot插入→ 去产生致癌mRNA癌基 前,病毒LTR启动子增强 转录mRNA→tat蛋白作用于 因产物转化细胞致癌.子启动增强c-onc表达, TGF-β1等→使细胞增殖永生 改变细胞生长特性 化。
急性逆转录病毒 (2)慢性逆转录病毒 (3)HTlV(人类嗜T细胞逆转录病毒) ①含onc* ①不含onc ①含tat的HTLV. ②可在数周内使培养 ②不能转化,但V感染 ②含使培养细胞转化,机理和1、2. 细胞发生转化. 整体动物潜伏数月致癌. 不同。 ③onc插到宿主DNA中 ③前V整合到proto-onc ③反式调控作用,tot插入→ 去产生致癌mRNA癌基 前,病毒LTR启动子增强 转录mRNA→tat蛋白作用于. 因产物转化细胞致癌.子启动增强c-onc表达, TGF-β1等→使细胞增殖永生. 改变细胞生长特性. 化。")
84
(五)人类免疫缺陷病毒基因组(HIV) 获得性免疫缺陷综合征(AIDS)1981年在美国首先发现,其病原是一种破坏人免疫系统的retro-V.1986年命名为HIV。HIV特异性侵犯并损耗T细胞造成机体免疫缺陷,最后并发各种严重感染和恶性肿瘤,成为艾滋病(谈“艾”色变)。因该病无特效治疗,病死率几乎为100%。(洁身自好) 1.HIV感染的流行病学(节选) (1)我国流行特征: ①发现较早,流行仍属低下,但呈明显上升趋势。 ②分布地区广,表现形式多。 ③年龄范围广,以青壮年为主,男女比为11.5/1。 ④感染途径多,但通过性传播日显重要。 ⑤感染者的发现比重明显从外籍人员转向国内居民。
1981年在美国首先发现,其病原是一种破坏人免疫系统的retro-V.1986年命名为HIV。HIV特异性侵犯并损耗T细胞造成机体免疫缺陷,最后并发各种严重感染和恶性肿瘤,成为艾滋病(谈 艾 色变)。因该病无特效治疗,病死率几乎为100%。(洁身自好) 1.HIV感染的流行病学(节选) (1)我国流行特征: ①发现较早,流行仍属低下,但呈明显上升趋势。 ②分布地区广,表现形式多。 ③年龄范围广,以青壮年为主,男女比为11.5/1。 ④感染途径多,但通过性传播日显重要。 ⑤感染者的发现比重明显从外籍人员转向国内居民。")
85
(2)HIV感染的危险因素增加 ①性传播疾病发生率有增无减. ②流行人口增加,估计全国流动人口达12000万,年龄20-29,正处于性活跃期。 ③性观念,性行为改变. ④吸毒、贩毒剧增,吸毒者往往有性乱,女性则卖淫。 ⑤医源性传播控制工作及宣传防治工作薄弱。 专家认为中国大陆HIV疫情持续上升,如超过一定数量,将出现暴发流行。HIV为分为HIV-1和HIV-2两个型。 HIV-1是感染的主要毒株,一旦感染,终身带毒,进展快,预后差。 HIV-2感染的潜伏期长,致病性低。 中国存在HIV-1A、B、B‘、C、E五个亚型。

86
2.HIV的生物学特性(见分子病毒学) 3.HIV基因组结构和功能 (1)HIV基因组为SS(+)RNA,HIV-1 9.3kb,HIV-2 9.7kb,5’端有帽,3’端poly(A)尾含3个结构基因,至少6个调节基因。 (2)LTR与结构基因。 ①LTR 含启动子、增强子、负调节元件及哺乳动物组胺转录因子结合位点等。 ②gag 编码病毒核心结构蛋白 ③pol 编码逆转录酶、整合酶、蛋白酶。 ④env 编码包膜蛋白
LTR与结构基因。 ①LTR 含启动子、增强子、负调节元件及哺乳动物组胺转录因子结合位点等。 ②gag 编码病毒核心结构蛋白. ③pol 编码逆转录酶、整合酶、蛋白酶。 ④env 编码包膜蛋白.")
87
(3)HIV调节基因 ①tat 反式激活转录基因 ②rev 病毒蛋白表达调节因子 ③vif 病毒侵染性因子 ④nef 负调节子 ⑤vpr 病毒蛋白R ⑥vpu病毒蛋白U ⑦vpx病毒蛋白X 仅存于HIV-2中.

88
Yingzi Kang Ph.D Dept. of biochemistry, Tianjin Medical University
医学分子生物学 Yingzi Kang Ph.D Dept. of biochemistry, Tianjin Medical University

89
基本内容 第一篇 基因 第二篇 蛋白质 第三篇 细胞信号转导的分子机制(略) 第四篇 细胞增殖、分化与细胞凋亡的分子机制
第五篇 分子生物学实验技术

90
基本概念 分子生物学( molecular biology)是从分子水平(基因水平)去研究或解释一切生物学现象,并在分子水平上改造和利用生物的一门学科。 基因工程、细胞工程、生物工程、核酸与蛋白质相互作用。 基因工程是指在人工可以控制的条件下将基因剪切或重新组合,再导入到生物体内,使这些基因在其中表达并遗传下去。

91
注释 分子生物学是从分子水平研究生命现象、生命本质、生命活动及其规 的科学。因此分子生物学必须研究分子,但是其研究的侧重点不是化学,而是生物学,是从分子水平研究生命及其规 。 病毒 单细胞生物 多细胞生物 分子水平生命活动主要是通过核酸和蛋白质这两类生物大分子的活动来实现的。

92
基因的概念 遗传学概念:1865年孟德尔在提出“遗传”的概念时提出的,即基因代表了生物体内任何一种遗传物质(生物的某一种性状)。
分子生物学概念: 1868年生化学家发现了核酸(核内酸性物质) 1925年 荚膜(光滑、Ⅲ型)Ⅲ型活菌体致病;无荚膜(粗糙、Ⅱ型) 1944年美国科学家用提出的核酸代替菌体,取得同样效果。核酸 → 基因 生物大分子结构研究概念:应用物理学和化学研究核酸大分子结构和功能。
 1925年 荚膜(光滑、Ⅲ型)Ⅲ型活菌体致病;无荚膜(粗糙、Ⅱ型) 1944年美国科学家用提出的核酸代替菌体,取得同样效果。核酸 → 基因. 生物大分子结构研究概念:应用物理学和化学研究核酸大分子结构和功能。")
93
基因与基因组的概念 基因是生物体的遗传物质,一般是指与生物体某些性状有关的核酸(结构基因),以及负责调节控制基因活动的调控基因。
生物体内全部基因称为基因组(genome),包括了任何染色体体内中的任何一个基因,是一个很庞大的范围。 基因工程技术是要从基因组中分离、纯化、扩增某一个目的基因。
,包括了任何染色体体内中的任何一个基因,是一个很庞大的范围。 基因工程技术是要从基因组中分离、纯化、扩增某一个目的基因。")
94
分子克隆技术 是利用单细胞(如大肠杆菌)进行无性繁殖的技术 基因的剪切、重组、转化、增殖、分离
重组DNA的转化需要载体,最常用的为质粒,很多质粒本身就是大肠杆菌中提取出来的,是大肠杆菌中染色体外可以复制的环状DNA,可以出入于大肠杆菌。 重组后的质粒被引入到大肠杆菌中的过程为转化。

95
限制性内切酶 没有限制性内切酶的发现和应用就没有分子生物学的发展。
“核酸限制性内切酶”是一类能识别双链DNA中特定碱基顺序的核酸水解酶,基本上都是从原核生物中发现提取的。其原本的功能是生物抗击外来DNA的侵袭,犹似高等动物的免疫系统。主要原理是以内切方式水解核酸链中磷酸二酯键 5’-P 3’-OH Ⅰ型:复合功能酶,兼有修饰、切割两种特性,识别位点和切割位点不一致,不能形成特异片段。 Ⅲ型:与Ⅰ型相似。 Ⅱ型:通常所说的限制性内切酶,识别序列为4~6个bp。

96
载体(vector) 载体是携带靶DNA片段进入宿主细胞进行扩增和表达的工具。
质粒(plasmid)为最常用的载体。细菌染色体外小型双链环状DNA复制子,对细菌的某些代谢活动和抗药性表型有一些作用。质粒载体是在天然质粒基础上人工改造拼接而成,其不仅可以在细菌中复制,同时在添加真核细胞复制信号和启动子后,可构建成真核细胞复制表达的穿梭质粒,用途非常广泛。 其他载体:λ噬菌体、粘粒、病毒等
为最常用的载体。细菌染色体外小型双链环状DNA复制子,对细菌的某些代谢活动和抗药性表型有一些作用。质粒载体是在天然质粒基础上人工改造拼接而成,其不仅可以在细菌中复制,同时在添加真核细胞复制信号和启动子后,可构建成真核细胞复制表达的穿梭质粒,用途非常广泛。 其他载体:λ噬菌体、粘粒、病毒等.")
97
基因研究的几种重要分析方法 限制性内切酶酶谱分析 核酸分子杂交 核苷酸序列分析

98
限制性内切酶谱 利用内切酶的功能对基因组或单个基因进行酶切,经过多种酶(至少三个)的多个切点的反复比较,可以得到某一基因的酶切图谱。
如在某些疾病(特别是遗传病)时,基因结构或碱基序列发生变化(突变、缺失、插入),此时酶切图谱会有改变。利用限制性片段长度多态性(RFLP)进行分析。
时,基因结构或碱基序列发生变化(突变、缺失、插入),此时酶切图谱会有改变。利用限制性片段长度多态性(RFLP)进行分析。")
99
核酸分子杂交(hybridization)
具有一定同源性的两条单链在一定条件下(适宜的温度和离子强度等),可按碱基互补原则退火形成双链,杂交过程是高度特异性的。 杂交的双方是待测序列(靶序列)和探针,通常探针是经过标记的(同位素、生物素、化学发光物质),靶序列可以是克隆的基因片段,也可以是基因组,可以在体外进行(杂交膜上),也可以在细胞内进行(原位杂交)。 cDNA芯片为杂交检测的一个发展。
,可按碱基互补原则退火形成双链,杂交过程是高度特异性的。 杂交的双方是待测序列(靶序列)和探针,通常探针是经过标记的(同位素、生物素、化学发光物质),靶序列可以是克隆的基因片段,也可以是基因组,可以在体外进行(杂交膜上),也可以在细胞内进行(原位杂交)。 cDNA芯片为杂交检测的一个发展。")
100
核苷酸序列分析 (sequencing) 测序分析是基因研究中最精确的分析,所以又称为一级结构分析。一级结构的了解是进一步分析基因结构和功能关系的前提,同时对基因表达、调控的研究也是非常重要的。 主要方法是sanger提出的酶法(末端终止法)和Maxam、Gilbert提出的化学降解法。 现在在此基础上将测序工作自动化,大大加快了测序的速度,正是有了自动测序技术,才能提前完成了人类基因组的测序工作。

101
第 十 三 章 基因表达调控 Regulation of Gene Expression 目 录

102
Basic Conceptions and Principle
第 一 节 基本概念与原理 Basic Conceptions and Principle

103
一、基因表达的概念 基因表达是受调控的 * 基因组(genome) 一个细胞或病毒所携带的全部遗传信息或整套基因。
* 基因表达(gene expression) 基因经过转录、翻译,产生具有特异生物学功能的蛋白质分子的过程。 基因表达是受调控的 目 录
 基因经过转录、翻译,产生具有特异生物学功能的蛋白质分子的过程。 基因表达是受调控的. 目 录.")
104
二、基因表达的时间性及空间性 (一)时间特异性
按功能需要,某一特定基因的表达严格按特定的时间顺序发生,称之为基因表达的时间特异性(temporal specificity)。 多细胞生物基因表达的时间特异性又称阶段特异性(stage specificity)。 目 录
。 多细胞生物基因表达的时间特异性又称阶段特异性(stage specificity)。 目 录.")
105
(二)空间特异性 在个体生长全过程,某种基因产物在个体按不同组织空间顺序出现,称之为基因表达的空间特异性(spatial specificity)。 基因表达伴随时间顺序所表现出的这种分布差异,实际上是由细胞在器官的分布决定的,所以空间特异性又称细胞或组织特异性(cell or tissue specificity)。 目 录
。 目 录.")
106
三、基因表达的方式 (一)组成性表达 按对刺激的反应性,基因表达的方式分为:
某些基因在一个个体的几乎所有细胞中持续表达,通常被称为管家基因(housekeeping gene)。
。")
107
无论表达水平高低,管家基因较少受环境因素影响,而是在个体各个生长阶段的大多数或几乎全部组织中持续表达,或变化很小。区别于其他基因,这类基因表达被视为组成性基因表达(constitutive gene expression)。
。")
108
(二)诱导和阻遏表达 在特定环境信号刺激下,相应的基因被激活,基因表达产物增加,这种基因称为可诱导基因。
可诱导基因在特定环境中表达增强的过程,称为诱导(induction)。 如果基因对环境信号应答是被抑制,这种基因是可阻遏基因。可阻遏基因表达产物水平降低的过程称为阻遏(repression)。
。 如果基因对环境信号应答是被抑制,这种基因是可阻遏基因。可阻遏基因表达产物水平降低的过程称为阻遏(repression)。")
109
在一定机制控制下,功能上相关的一组基因,无论其为何种表达方式,均需协调一致、共同表达,即为协调表达(coordinate expression),这种调节称为协调调节(coordinate regulation)。
,这种调节称为协调调节(coordinate regulation)。")
110
四、基因表达调控的生物学意义 (一)适应环境、维持生长和增殖 (二)维持个体发育与分化
适应环境、维持生长和增殖 (二)维持个体发育与分化")
111
五、基因表达调控的基本原理 (一)基因表达的多级调控 基因激活 转录起始 转录后加工 mRNA降解 转录起始 蛋白质降解等 蛋白质翻译
翻译后加工修饰

112
(二)基因转录激活调节基本要素 1. 特异DNA序列和调节蛋白质
基因表达的调节与基因的结构、性质,生物个体或细胞所处的内、外环境,以及细胞内所存在的转录调节蛋白有关。 1. 特异DNA序列和调节蛋白质

113
原核生物 —— 操纵子(operon) 机制 蛋白质因子 特异DNA序列 编码序列 启动序列 操纵序列 其他调节序列 (promoter)
(operator) 蛋白质因子
 蛋白质因子.")
114
是RNA聚合酶结合并启动转录的特异DNA序列。 1) 启动序列
-35区 -10区 TTGACA TTAACT TTTACA TATGAT TATGTT TTGATA TATAAT CTGACG TACTGT N17 N16 N7 N6 A trp tRNATyr lac recA Ara BAD 共有序列

115
共有序列(consensus sequence) 决定启动序列的转录活性大小。
某些特异因子(蛋白质)决定RNA聚合酶对一个或一套启动序列的特异性识别和结合能力。
决定RNA聚合酶对一个或一套启动序列的特异性识别和结合能力。")
116
pol 启动序列 编码序列 操纵序列 2 操纵序列 ——阻遏蛋白(repressor)的结合位点
当操纵序列结合有阻遏蛋白时,会阻碍RNA聚合酶与启动序列的结合,或是RNA聚合酶不能沿DNA向前移动 ,阻碍转录。 pol 阻遏蛋白 启动序列 编码序列 操纵序列

117
3) 其他调节序列、调节蛋白 例如 激活蛋白(activator)可结合启动序列邻近的DNA序列,促进RNA聚合酶与启动序列的结合,增强RNA聚合酶活性。 有些基因在没有激活蛋白存在时,RNA聚合酶很少或完全不能结合启动序列。
可结合启动序列邻近的DNA序列,促进RNA聚合酶与启动序列的结合,增强RNA聚合酶活性。 有些基因在没有激活蛋白存在时,RNA聚合酶很少或完全不能结合启动序列。 .")
118
真核生物 编码序列 1) 顺式作用元件(cis-acting element) ——可影响自身基因表达活性的DNA序列 转录起始点 B A
不同真核生物的顺式作用元件中也会发现一些共有序列 ,如TATA盒、CAAT盒等,这些共有序列是RNA聚合酶或特异转录因子的结合位点。

119
2) 真核基因的调节蛋白 反式作用因子(trans-acting factor) 由某一基因表达产生的蛋白质因子,通过与另一基因的特异的顺式作用元件相互作用,调节其表达。 这种调节作用称为反式作用。 还有蛋白质因子可特异识别、结合自身基因的调节序列,调节自身基因的表达,称顺式作用。

120
a DNA 反式调节 A mRNA 蛋白质A A B C c DNA mRNA C 蛋白质C 顺式调节

121
2. DNA - 蛋白质 蛋白质-蛋白质 的相互作用 指的是反式作用因子与顺式作用元件之间的特异识别及结合。通常是非共价结合,被识别的DNA结合位点通常呈对称、或不完全对称结构。 绝大多数调节蛋白质结合DNA前,需通过蛋白质-蛋白质相互作用,形成二聚体(dimer)或多聚体(polymer)。
或多聚体(polymer)。")
122
⑴ 原核启动序列/真核启动子与RNA聚合酶活性
一些特异调节蛋白在适当环境信号刺激下表达,然后通过DNA-蛋白质、蛋白质-蛋白质相互作用影响RNA聚合酶活性。

123
Regulation of Prokaryotic
第 二 节 原核基因转录调节 Regulation of Prokaryotic Gene Transcription

124
一、原核基因转录调节特点 (一)σ因子决定RNA聚合酶识别特异性 (二)操纵子模型的普遍性 (三)阻遏蛋白与阻遏机制的普遍性
——调节的主要环节在转录起始 一、原核基因转录调节特点 (一)σ因子决定RNA聚合酶识别特异性 (二)操纵子模型的普遍性 (三)阻遏蛋白与阻遏机制的普遍性
σ因子决定RNA聚合酶识别特异性. (二)操纵子模型的普遍性. (三)阻遏蛋白与阻遏机制的普遍性.")
125
二、乳糖操纵子调节机制 Z Y A O P (一)乳糖操纵子(lac operon)的结构 调控区 结构基因 DNA 操纵序列 启动序列
CAP结合位点 启动序列 操纵序列 结构基因 Z: β-半乳糖苷酶 Y: 透酶 A:乙酰基转移酶 Z Y A O P DNA

126
(二)阻遏蛋白的负性调节 阻遏基因 pol I DNA Z Y A O P mRNA 阻遏蛋白 没有乳糖存在时
阻遏蛋白的负性调节 阻遏基因 pol I DNA Z Y A O P mRNA 阻遏蛋白 没有乳糖存在时")
127
pol I DNA Z Y A O P mRNA 阻遏蛋白 mRNA 启动转录 β-半乳糖苷酶 乳糖 半乳糖 有乳糖存在时

128
Z Y A O P (三)CAP的正性调节 + + + + 转录 DNA CAP 无葡萄糖,cAMP浓度高时 CAP CAP
CAP的正性调节 转录 DNA CAP 无葡萄糖,cAMP浓度高时 CAP CAP")
129
(四)协调调节 ※当阻遏蛋白封闭转录时,CAP对该系统不能发挥作用; ※如无CAP存在,即使没有阻遏蛋白与操纵序列结合,操纵子仍无转录活性。
单纯乳糖存在时,细菌利用乳糖作碳源; 若有葡萄糖或葡萄糖/乳糖共同存在时,细菌首先利用葡萄糖。 葡萄糖对 lac 操纵子的阻遏作用称分解代谢阻遏(catabolic repression)。
。")
130
低半乳糖时 高半乳糖时 葡萄糖低 cAMP浓度高 RNA-pol O O mRNA 葡萄糖高cAMP浓度低 O O

131
三、其他转录调节机制 ? (一)转录衰减 P O 色氨酸操纵子 调节区 结构基因 Trp 低时 Trp 高时 Trp RNA聚合酶 trpR
mRNA Trp 高时 ? 色氨酸操纵子 Trp

132
O P 前导序列 trp 密码子 trpR 1 2 3 4 调节区 结构基因 衰减子区域 UUUU…… 前导mRNA 终止密码子
14aa前导肽编码区: 包含序列1 衰减子结构 第10、11密码子为trp密码子 UUUU…… UUUU…… 形成发夹结构能力强弱: 序列1/2>序列2/3>序列3/4

133
转录衰减机制 3 4 前导DNA UUUU 3’ 前导mRNA 终止 核糖体 UUUU 3’ UUUU…… trp 密码子 前导肽
衰减子结构 就是终止子 可使转录 3 4 终止 核糖体 UUUU…… 3 4 1 2 5’ UUUU 3’ trp 密码子 前导肽 1.当色氨酸浓度高时

134
3 4 Trp合成酶系相关 结构基因被转录 前导DNA 结构基因 前导mRNA 核糖体 UUUU…… UUUU……
2 3 UUUU…… 核糖体 UUUU…… 3 4 2 1 5’ trp 密码子 序列3、4不能形成衰减子结构 前导肽 2.当色氨酸浓度低时

135
(二)基因重组 沙门菌鞭毛素基因的调节 hin I hin I H2 DNA 启动序列 H1 Hin重组酶 H2鞭毛素 阻遏蛋白 H2 H1
转位片段 H1鞭毛素

136
SOS基因 (三)SOS反应 DNA 操纵序列 Lex A阻遏蛋白 Lex A阻遏蛋白 基因表达 Rec A
激活 Rec A 与DNA 损伤修复有关的酶和蛋白质 紫外线

137
Regulation of Eukaryotic
第 三 节 真核基因转录调节 Regulation of Eukaryotic Gene Transcription

138
一、真核基因组结构特点 (一)真核基因组结构庞大 哺乳类动物基因组 DNA 约 3 × 10 9 碱基对
编码基因约 有 个,占总长的6 % rDNA等重复基因约 占 5% ~ 10%

139
(二)单顺反子 (三)重复序列 (四)基因不连续性 单顺反子(monocistron)
即一个编码基因转录生成一个mRNA分子,经翻译生成一条多肽链。 (三)重复序列 单拷贝序列(一次或数次) 高度重复序列(106 次) 中度重复序列(103 ~ 104次) 多拷贝序列 (四)基因不连续性
重复序列. 单拷贝序列(一次或数次) 高度重复序列(106 次) 中度重复序列(103 ~ 104次) 多拷贝序列. (四)基因不连续性.")
140
二、真核基因表达调控特点 (一)RNA聚合酶 (二)活性染色体结构变化 1. 对核酸酶敏感 活化基因常有超敏位点,位于调节蛋白结合位点附近。
RNA聚合酶 (二)活性染色体结构变化 1. 对核酸酶敏感 活化基因常有超敏位点,位于调节蛋白结合位点附近。")
141
2. DNA拓扑结构变化 正超螺旋 负超螺旋 3. DNA碱基修饰变化 天然双链DNA均以负性超螺旋构象存在; 基因活化后 RNA-pol
转录方向 3. DNA碱基修饰变化 真核DNA约有5%的胞嘧啶被甲基化, 甲基化范围与基因表达程度呈反比。

142
4. 组蛋白变化 (三)正性调节占主导 (四)转录与翻译分隔进行 (五)转录后修饰、加工 ① 富含Lys组蛋白水平降低
② H2A, H2B二聚体不稳定性增加 ③ 组蛋白修饰 ④ H3组蛋白巯基暴露 (三)正性调节占主导 (四)转录与翻译分隔进行 (五)转录后修饰、加工
正性调节占主导. (四)转录与翻译分隔进行. (五)转录后修饰、加工.")
143
三、真核基因转录激活调节 (一)顺式作用元件 1. 启动子
真核基因启动子是RNA聚合酶结合位点周围的一组转录控制组件,至少包括一个转录起始点以及一个以上的功能组件。 TATA盒 GC盒 CAAT盒

145
2. 增强子(enhancer) 3. 沉默子(silencer)
指远离转录起始点、决定基因的时间、空间特异性、增强启动子转录活性的DNA序列。 3. 沉默子(silencer) 某些基因的负性调节元件,当其结合特异蛋白因子时,对基因转录起阻遏作用。
 某些基因的负性调节元件,当其结合特异蛋白因子时,对基因转录起阻遏作用。")
146
(二)反式作用因子 1. 转录调节因子分类(按功能特性) * 基本转录因子(general transcription factors)
是RNA聚合酶结合启动子所必需的一组蛋白因子,决定三种RNA(mRNA、tRNA及rRNA)转录的类别。
转录的类别。")
147
* 特异转录因子(special transcription factors)
为个别基因转录所必需,决定该基因的时间、空间特异性表达。 转录激活因子 转录抑制因子

148
2. 转录调节因子结构 DNA结合域 转录激活域 TF 谷氨酰胺富含域 酸性激活域 脯氨酸富含域 蛋白质-蛋白质结合域 (二聚化结构域)
")
149
最常见的DNA结合域 1. 锌指(zinc finger) 常结合GC盒 C —— Cys H —— His
 常结合GC盒 C —— Cys H —— His")
150
Cys Zn His

151
2. α-螺旋 常结合CAAT盒

152
(三)mRNA 转录激活及其调节 polⅡ TFⅡF TAF TAF TAF TFⅡH TFⅡA TBP TFⅡB TBP相关因子 DNA
TATA DNA 真核RNA聚合酶Ⅱ在转录因子帮助下,形成的转录起始复合物

153
真核基因转录调节是复杂的、多样的 *不同的DNA元件组合可产生多种类型的转录调节方式; *多种转录因子又可结合相同或不同的DNA元件。 *转录因子与DNA元件结合后,对转录激活过程所产生的效果各异,有正性调节或负性调节之分。

Similar presentations
 的字头 AIDS 的译称,中文名为获得性免疫缺 陷综合症,是由一种人类免疫缺陷病毒 (Human.>")
,必须在活细胞内寄生并以复 制方式增殖的非细胞型微生物。 生物病毒微生物 原指一种动物来源的 毒素。 “virus” 一词源于 拉丁文。 病毒能增殖、遗传和演化,因而 具 有生命最基本的特征。>")

免疫缺陷病 2 )超敏反应(过敏反应) 3 )自身免疫病等.>")
染色体结构不同; ( 2 )原核生物具有正调控和负调控并重的特点,真核 生物目前已知的主要是正调控; ( 3 )原核生物的转录和翻译是相偶联的,真核生物的.>")



>")
>")






 蕭伊玲 18號 (510919)>")


