Download presentation
Presentation is loading. Please wait.

1
9 疾病与人类健康 9. 1 肿瘤与癌症 癌(cancer)是一群不受生长调控而繁殖的细胞,也称恶性肿瘤。良性肿瘤则是一群仅局限在自己的正常位置,且不侵染周围其它组织和器官的细胞。
是一群不受生长调控而繁殖的细胞,也称恶性肿瘤。良性肿瘤则是一群仅局限在自己的正常位置,且不侵染周围其它组织和器官的细胞。")
2
图9-1 肿瘤组织示意图

3
癌基因(oncogene)可分为两大类:
一类是病毒癌基因(viral oncogene,V-onc),编码病毒癌基因的主要有DNA病毒和RNA病毒。
,编码病毒癌基因的主要有DNA病毒和RNA病毒。")
4
DNA病毒包括乙型肝炎病毒、SV40和多瘤病毒、乳头瘤病毒、腺病毒、疱疹病毒和痘病毒。
RNA病毒主要是反转录病毒。反转录病毒致癌基因(retrovirus onc)可能是研究最多的病毒基因,它们能使靶细胞发生恶性转化。
可能是研究最多的病毒基因,它们能使靶细胞发生恶性转化。")
5
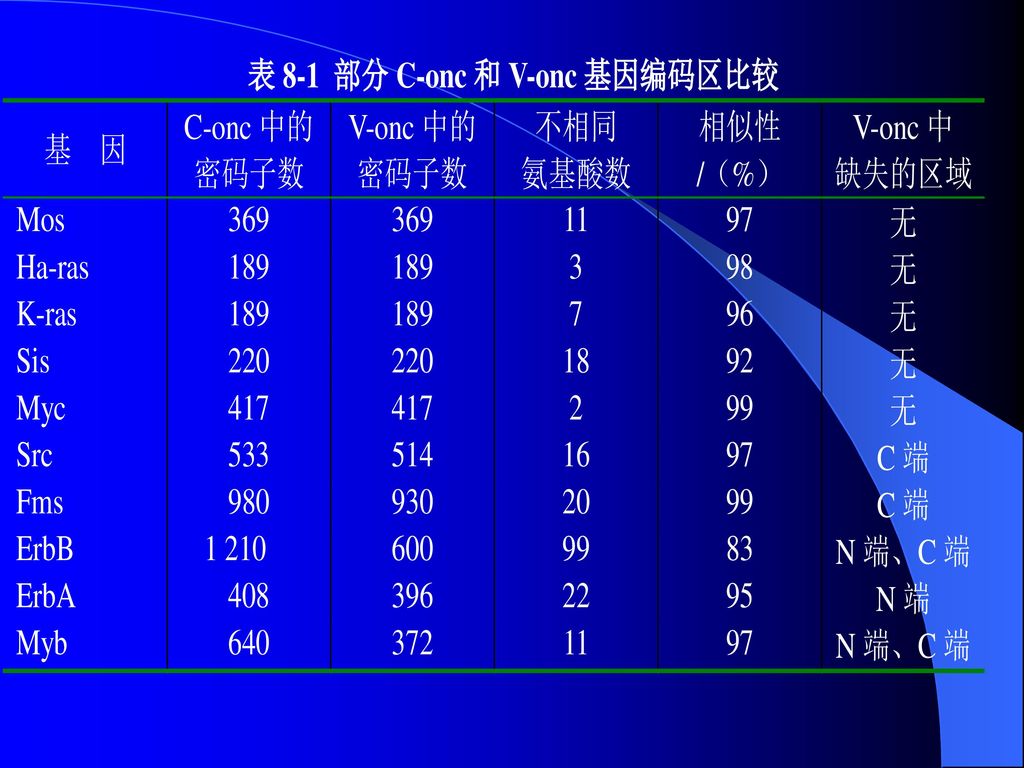
第二类是细胞转化基因(C-onc)(←原癌基因),它们能使正常细胞转化为肿瘤细胞,这类基因与病毒癌基因有显著的序列相似性。
(←原癌基因),它们能使正常细胞转化为肿瘤细胞,这类基因与病毒癌基因有显著的序列相似性。")
7
9.1.1 反转录病毒致癌基因 Rous在1910年发现带有单链RNA肉瘤病毒(一种反转录病毒)的鸡肉瘤无细胞滤液能在鸡体内诱发新的肉瘤。
的鸡肉瘤无细胞滤液能在鸡体内诱发新的肉瘤。")
8
这种病毒的基因组只有6-9kb。RNA上的基因数目很少,它们被包裹在由gag和env两个基因编码的蛋白质外壳中,其中env基因指导外壳蛋白的合成,gag基因则指导“鞘”蛋白的合成,这些“鞘”蛋白好像“道钉”一样“箍”在外壳的表面,维持外壳蛋白结构的稳定性。

9
图9-2 反转录病毒颗粒示意图

10
图9-3 反转录病毒基因结构示意图

11
反转录酶以病毒RNA为模板,转录出单链DNA分子,利用宿主DNA聚合酶指导合成第二条DNA链,以双链DNA形式整合到宿主细胞基因组中。
被整合的反转录病毒DNA分子称为原病毒(provirus),它能指导病毒mRNA的合成,并利用宿主细胞中的蛋白质合成机器,翻译生成病毒外壳蛋白等,最后组装成病毒颗粒。
,它能指导病毒mRNA的合成,并利用宿主细胞中的蛋白质合成机器,翻译生成病毒外壳蛋白等,最后组装成病毒颗粒。")
12
反转录病毒致癌机理: 反转录病毒中有许多致癌基因,其感染后激活宿主特定基因表达,从而破坏宿主细胞本身固有的平衡,导致细胞发生转化。

13
图9-4 反转录病毒插入引起C-myc基因活化的几种可能途径

14
V-onc基因的起源 研究发现,反转录病毒基因组中所带有的onc基因并非来自病毒本身,而是这些病毒在感染动物或人体之后获得的细胞原癌基因。 动物或人原癌基因经病毒修饰和改造后,成为病毒基因组的一部分并具有了致癌性,其作用的靶分子也往往发生改变。

15
原癌基因产物及其分类 除了病毒感染外,许多非病毒因子(如放射性物质、化学试剂亚硝酸、烷化剂等)也能诱导细胞转化。这些因子并没有把致癌基因或其他致癌的遗传信息带入细胞,而仅仅通过某种激活机制改变了细胞内原有的遗传信息(内源基因发生突变),使细胞发生恶性转化。
也能诱导细胞转化。这些因子并没有把致癌基因或其他致癌的遗传信息带入细胞,而仅仅通过某种激活机制改变了细胞内原有的遗传信息(内源基因发生突变),使细胞发生恶性转化。 .")
16
图9-7 黄曲霉素(Aflatoxin)导致细胞发生癌变的分子机制
导致细胞发生癌变的分子机制")
17
根据原癌基因产物在细胞中的位置可分为3类:
与膜结合的蛋白,主要有erbB、neu、fms、mas和src基因产物; 可溶性蛋白,包括mos、sis和fps基因产物; 第三类是核蛋白,包括myc、ets、jun和myb等。

18
根据这些蛋白的功能,它们又常被分为6个功能群类别,
蛋白激酶类 生长因子类 生长因子受体类 GTP结合蛋白类 核蛋白类和功能未知类

19
9. 1. 3 原癌基因的表达调控 原癌基因在正常细胞中通常以单拷贝形式存在,只有低水平的表达或根本不表达。
在很多情况下,原癌基因的结构发生了点突变或插入、重排、缺失及扩增等,改变其转录活性。

20
图8-8 细胞中原癌基因转变为癌基因的主要途径

21
1、点突变 研究发现,ras基因编码了一个分子量为2.1×104癌蛋白(p21),从人类膀胱癌细胞系T24 DNA中克隆的Ha-ras基因能够诱发NlH/3T3细胞转化,而从正常细胞DNA中克隆的该原癌基因没有这种功能。推测为点突变所致。
,从人类膀胱癌细胞系T24 DNA中克隆的Ha-ras基因能够诱发NlH/3T3细胞转化,而从正常细胞DNA中克隆的该原癌基因没有这种功能。推测为点突变所致。")
22
如人类肺癌细胞系Hs242的转化基因与Ha-ras高度相似,在这个基因中导致转化活性的遗传损伤是第二个外显子中引起p21蛋白第61位谷氨酰胺被亮氨酸所替代的一个点突变。

23
图9-9 ras基因的点突变及转化活性分析

24
2、LTR插入 LTR是逆转录病毒基因组两端的长末端重复序列(long terminal repeat),含有强启动子序列,当LTR插入原癌基因启动子区域或邻近部位后,可从根本上改变基因的正常调控规律。 LTR插入到c-myc 5’上游启动子附近,使c-myc的转录水平大大增加。

25
3、基因重排 正常情况下,c-myc定位于8q24,免疫球蛋白重链基因(IgH)定位于14q32,轻链λ基因(Igλ)定位于22q12,轻链基因(Igk)定位于2p11,。 在Burkitt淋巴瘤中,c-myc易位至IgH、Igk或Igλ的位点,使Ig基因与c-myc相连在一起,Ig基因启动子使原来不表达的c-myc高表达。

26
在正常人体细胞中,非受体型酪氨酸蛋白激酶基因abl位于第9号染色体上,表达量极低,不会诱发癌变。在慢性骨髓瘤病人细胞中,该基因却被转移到第22号染色体上,与bcr基因相融合,表达量大为提高。

27
图9-10 abl原癌基因通过选择性染色体重排转变成细胞癌基因

28
4、缺失。很多原癌基因5’上游区存在负调控序列,一旦该序列发生缺失或突变,就丧失抑制基因表达调控的能力。如Burkitt淋巴瘤中C-myc可因负调控序列的缺失或LTR插入破坏其结构而增强表达。
5、基因扩增。使每个细胞中基因拷贝数增加,从而直接增加可用的转录模板。

29
基因相互作用与癌基因表达 1、染色体构象对原癌基因表达的影响。 基因表达不仅取决于基因本身及其相邻区域的一级结构,也取决于其空间构象,即基因在染色体上的空间排列和染色质的结构。当两个基因相距太近时,往往不易形成有利于高效转录的空间结构。

30
基因与基因之间的间隔距离被定义为“基因领域”(gene territory)。同一DNA链上两个具有相同转录方向的基因间隔小于一定长度时,影响有效转录所必需的染色质结构的形成,从而使这两个基因中的一个或两个均不能转录或转录活性显著降低,产生所谓基因领域效应(gene territorial effect)。
。同一DNA链上两个具有相同转录方向的基因间隔小于一定长度时,影响有效转录所必需的染色质结构的形成,从而使这两个基因中的一个或两个均不能转录或转录活性显著降低,产生所谓基因领域效应(gene territorial effect)。")
31
正常人c-myc定位于第8号染色体,在其两侧分别存在强表达的基因,使c-myc处于两面受夹击的地位。在Burkitt淋巴瘤中,由于发生基因重排,使c-myc基因一侧的强表达基因消失,从而消除了对c-myc的基因领域效应,使后者的转录活性增强。

32
小鼠细胞中,c-myc的5’上游区域也存在一个强表达基因,全长15kb,距c-myc只有3kb。很显然,这一间隔距离太短,与基因有效转录应有的最小距离相差甚远,c-myc受基因领域效应的影响非常大,表达受抑制。 在小鼠乳腺癌细胞中,上述间隔距离被显著拉长,激活c-myc转录。

33
2、原癌基因终产物对基因表达的影响。 癌基因产物通过介质传递生长刺激信号的部位有3处:
①癌基因产物本身模拟了生长因子,因而与相应的受体作用,以自分泌的方式刺激细胞生长; ②癌基因产物模拟了已结合配体的生长因子受体,从而在无外源生长因子时提供了促进细胞分裂的信号;

34
③癌基因产物作用于细胞内生长控制途径,解除此途径对外源刺激信号的需求。
猴肉瘤病毒(SSV)的V-ras癌基因产物与人血小板衍生生长因子(PDGF), Src及V-erb癌基因产物与上皮生长因子(EGF)之间都存在着极高的相似性,表明生长因子与癌基因转化有关。
的V-ras癌基因产物与人血小板衍生生长因子(PDGF), Src及V-erb癌基因产物与上皮生长因子(EGF)之间都存在着极高的相似性,表明生长因子与癌基因转化有关。")
35
3、抑癌基因产物对原癌基因的调控。 因为抑癌基因产物能够抑制细胞的恶性增殖,所以它被认为是一种隐性癌基因。当细胞内由于某种原因造成这些基因的表达受抑制时,原癌基因就活跃表达,引起细胞癌变。

36
p53是通过杂合缺失鉴定的一个抑癌基因,在星形细胞癌、乳癌、肺癌、肠癌及骨肉瘤中都有高频率缺失现象。从癌细胞中得到的p53基因,其保守序列区有单一位点的突变,推测可能由于这一突变导致p53基因产物结构与功能的改变,失去抑癌活性。

37
4、外源信号对原癌基因表达的影响。细胞外信号(包括生长因子、激素、神经递质、药物等)作用于靶细胞后,通过细胞膜受体系统或其它直接途径被传递至细胞内,再通过多种蛋白激酶的活化,对转录因子进行修饰,然后激活一系列基因转录。
作用于靶细胞后,通过细胞膜受体系统或其它直接途径被传递至细胞内,再通过多种蛋白激酶的活化,对转录因子进行修饰,然后激活一系列基因转录。")
38
图9-11 许多原癌基因参与细胞信号转导过程

39
8.2人免疫缺损病毒——HIV 人免疫缺损病毒(HIV),俗称艾滋病毒(AIDS),诱发人类获得性免疫缺损综合症。该病毒在分类上属反转录病毒科(Retroviridae)慢病毒属中的灵长类免疫缺损病毒亚属。1983年,法国巴斯德研究所的Montaginer和美国国家卫生研究院癌症研究所Gallo等人首次证实HIV是艾滋病的病因。
,俗称艾滋病毒(AIDS),诱发人类获得性免疫缺损综合症。该病毒在分类上属反转录病毒科(Retroviridae)慢病毒属中的灵长类免疫缺损病毒亚属。1983年,法国巴斯德研究所的Montaginer和美国国家卫生研究院癌症研究所Gallo等人首次证实HIV是艾滋病的病因。")
40
HIV I是从欧洲和美洲分离的毒株,与猴艾滋病毒只有约45%的相似性,它的致病力很强,是引起全球艾滋病流行的主要病原。
HIV II与猴艾滋病毒的相似性高达75%,其毒力较弱,引起艾滋病的病程较长,症状较轻,且主要局限于西部非洲(表8-4)。 有关HIV的研究主要是针对HIV I进行的。
。 有关HIV的研究主要是针对HIV I进行的。")
42
HIV依靠血、血制品以及人体分泌的奶液和精液等传播,主要感染T淋巴细胞,也感染B淋巴细胞和单形核细胞等。
HIV感染形成多核巨细胞,并导致细胞死亡。HIV病毒可以通过所感染细胞扩散到全身,已在淋巴细胞、脑、胸腺、脾等组织发现了该病毒。 自然界广泛存在着突变株。

43
8.2.1 HIV病毒粒子的形态结构和传染 艾滋病病毒粒子是一种直径约为100nm的球状病毒,包被着由两层脂质组成的脂膜,这种结合有许多糖蛋白分子(主要是gp41和gp120)的脂质源于寄主细胞的外膜。蛋白质p24和p18组成其核心,内有基因组RNA链,链上附着有反转录酶。
的脂质源于寄主细胞的外膜。蛋白质p24和p18组成其核心,内有基因组RNA链,链上附着有反转录酶。")
44
图8-12 HIV I病毒粒子结构模式图。其中gp120和gp41为外膜蛋白,MA为内膜蛋白,CA为外壳蛋白,NC为核衣壳蛋白,RT为反转录酶。

45
HIV基因组及其编码的蛋白 1.HIV基因组结构 HIV基因组由两条单链正链RNA组成,每个RNA基因组约为9.7kb。在RNA5’端有一帽子结构(m7G5’GmpNp),3’端有多聚(A)尾巴。 主要由5’末端LTR、结构蛋白编码区(gag)、蛋白酶编码区(pro)、具有多种酶活性的蛋白编码区(pol)、外膜蛋白(env)和3’未端LTR组成。
、蛋白酶编码区(pro)、具有多种酶活性的蛋白编码区(pol)、外膜蛋白(env)和3’未端LTR组成。")
46
HIV I基因编码区有很多重叠,尤其在基因组的3’端。HIV基因组中的部分基因如Tat和Rev是不连续的,被插入的内含子分隔成两个外显子。

47
图8-13 HIV I基因结构和所编码的主要蛋白

48
2. HIV编码的蛋白质及其主要功能 HIV的结构蛋白主要包括3个基因。gag基因编码病毒的核心蛋白,翻译时先形成一个5.5×104的前体(p55),然后在HIV蛋白酶的作用下被切割成p17、p24、p15三个蛋白。P24和p17分别组成HIV颗粒的外壳(CA)和内膜蛋白(MA),p15进一步被切割成与病毒RNA结合的核衣壳蛋白(NC)p9和p7。
,然后在HIV蛋白酶的作用下被切割成p17、p24、p15三个蛋白。P24和p17分别组成HIV颗粒的外壳(CA)和内膜蛋白(MA),p15进一步被切割成与病毒RNA结合的核衣壳蛋白(NC)p9和p7。")
49
Pol基因编码病毒复制所需的酶类包括逆转录酶p66、整合酶p32。
从pol和gag基因重叠区内起始的一段序列为pro基因,它编码蛋白酶p22,在切割HIV蛋白前体的过程中起作用。 env编码8.8×104的蛋白质,经糖基化后其相对分子质量增至1.6×105,是HIV包膜糖蛋白gp160的前体,可进一步被切割成gp120和gp41。

50
gp120是外膜蛋白,感染细胞时可与细胞的CD4受体蛋白相结合,并与G蛋白偶联受体(GPCR)家族中的一个跨膜蛋白发生相互作用。
gp41是跨膜蛋白(TM),嵌入病毒包膜脂质中,对HIV的侵染和致病有非常重要的作用。
,嵌入病毒包膜脂质中,对HIV的侵染和致病有非常重要的作用。")
51
图8-14 HIV I感染小神经质细胞和巨噬细胞过程中信号传导及与其受体CD4和辅助受体相互作用示意图。

52
3.HIV膜蛋白主要功能区 (1)主要抗原决定簇:包括V3区(第 位的环状肽段)的主抗原决定簇及若干个较弱的决定簇,其中有两个分别位于gp41的 位和 位。 (2)T细胞决定簇:两个辅助性T细胞决定簇T2和T1分别在 位的C1区和 位的C4区,一个主要细胞毒T细胞决定簇位于第 位。另有3个较弱的细胞毒T细胞决定簇在gp41上。
T细胞决定簇:两个辅助性T细胞决定簇T2和T1分别在 位的C1区和 位的C4区,一个主要细胞毒T细胞决定簇位于第 位。另有3个较弱的细胞毒T细胞决定簇在gp41上。")
53
(3)CD4受体结合区:该区位于423-427位(C4区)。在上述功能区以外的某些氨基酸突变也能影响gp120的功能,说明上述各功能区发挥作用还依赖于整个分子特定的空间构象。
CD4受体结合区:该区位于 位(C4区)。在上述功能区以外的某些氨基酸突变也能影响gp120的功能,说明上述各功能区发挥作用还依赖于整个分子特定的空间构象。")
54
8.2.3 HIV的复制 HIV与受体结合后,病毒核心蛋白和两条RNA链进入细胞。反转录酶以病毒RNA为模板合成单链DNA,并由宿主细胞DNA聚合酶合成双链DNA(原病毒),经环化后进入细胞核并整合到染色体上,随细胞的分裂传代。可长期潜伏。
,经环化后进入细胞核并整合到染色体上,随细胞的分裂传代。可长期潜伏。")
55
主要过程如下: ①原病毒整合到宿主染色体上,无症状;②原病毒利用宿主细胞的转录和合成系统转录产生病毒mRNA,其中一部分编码病毒蛋白,与基因组RNA组装成新的病毒颗粒,从寄主细胞中释放出来侵染其它健康细胞;③寄主细胞瓦解死亡。

56
图8-15 HIV I在人体细胞内的复制和侵染过程示意图

57
HIV的感染及致病机理 HIV感染人体后立即大量复制和扩散,血清中出现HIV抗原,从外周血细胞、脑脊液和骨髓细胞中均能分离出HIV,是HIV原发感染的急性期。 大约70%以上的原发感染者在感染后2-4周内出现急性感染症状,包括发热、咽炎、淋巴结肿大、关节痛、中枢及外周神经系统病变、皮肤斑丘疹、粘膜溃疡等,持续1-2周后进入HIV感染的无症状潜伏期。

58
无症状潜伏期(可长达数年)。此时无任何临床症状,外周血中HIV抗原含量很低甚至检测不到。随感染时间的延长,HIV重新开始大量复制并造成免疫系统损伤。临床上病人感染逐步发展到持续性全身性淋巴腺病(PGL)、艾滋病相关综合症(APC)等,直至发展到艾滋病。
。此时无任何临床症状,外周血中HIV抗原含量很低甚至检测不到。随感染时间的延长,HIV重新开始大量复制并造成免疫系统损伤。临床上病人感染逐步发展到持续性全身性淋巴腺病(PGL)、艾滋病相关综合症(APC)等,直至发展到艾滋病。")
59
HIV除在细胞内大量繁殖造成细胞死亡外,还可通过以下几种途径导致免疫功能下降:
①HIV粒子表面的gp120蛋白脱落,与正常细胞膜上CD4受体结合,使该细胞被免疫系统误认为病毒感染细胞而遭杀灭; ②因T细胞CD4受体被gp120封闭,影响了其免疫辅助功能;

60
③HIV的gp120蛋白可刺激机体产生抗CD4结合部位的特异性抗体,阻断T细胞功能;
④带有病毒包膜蛋白的细胞可与其他细胞融合形成多核巨细胞而丧失功能。

61
8.2.5艾滋病的治疗及预防 迄今为止仍无任何药物可以完全抑制HIV在感染者体内的增殖并彻底治愈艾滋病。 已批准的抗HIV病毒药物主要如表8-7所示。

63
图8-21 核苷酸型和非核苷酸型反转录酶抑制剂的作用机制分析。a
图8-21 核苷酸型和非核苷酸型反转录酶抑制剂的作用机制分析。a.核苷酸型反转录酶抑制剂。由于它在结构上与脱氧核苷酸的相似性,掺入后使病毒DNA的合成不能进行。b.非核苷酸型反转录酶抑制剂。与反转录酶相结合,通过限制该酶的移动性而影响它的活性。

64
8.3 乙型肝炎病毒——HBV 病毒性肝炎是严重威胁人类健康的世界性传染病,引起肝炎的病毒通称肝炎病毒(hepatitis virus)。目前已经知道的至少有甲肝病毒(HAV)、乙肝病毒(HBV)、丙肝病毒(HCV)、丁肝病毒(HDV)和戊肝病毒(HEV)等5种病毒。
。目前已经知道的至少有甲肝病毒(HAV)、乙肝病毒(HBV)、丙肝病毒(HCV)、丁肝病毒(HDV)和戊肝病毒(HEV)等5种病毒。")
65
甲肝病毒属小RNA病毒科,乙肝病毒属嗜肝病毒科,丙肝病毒与黄热病毒和瘟病毒相似,为RNA病毒。
丁肝病毒基因组由单链环状RNA分子组成,其外壳能识别乙肝病毒的表面抗原,是一种与乙肝有关的缺陷型病毒,需要有HBV的辅助才能复制增殖。 戊肝病毒基因组为单链正链有多聚(A)RNA,球形无包膜。
RNA,球形无包膜。")
66
8.3.1肝炎病毒的分类及病毒粒子结构 1986年,国际病毒命名委员会正式将乙肝病毒定为嗜肝DNA病毒科成员,1990年又将该科病毒分为正嗜肝病毒属和禽嗜肝病毒属,乙肝病毒是正嗜肝病毒属成员。

67
HBV完整粒子的直径为42nm,由外膜和核壳组成,有很强的感染性。
其外膜由病毒的表面抗原、多糖和脂质构成;核壳直径27nm,由病毒的核心抗原组成,并含有病毒的基因组DNA、反转录酶和DNA结合蛋白等。

68
图8-22 HBV病毒粒子模式图

69
8.3.2 乙肝病毒基因组及其所编码的主要蛋白 乙肝病毒的基因组是一个有部分单链区的环状双链DNA分子,两条单链长度不同。 长链L(3.2kb)为负链,而短链S为正链,其长度不确定,约为负链的50%-80%左右。基因组依靠正链5’端约240bp的粘性末端与负链缺口部位的互补维持了环状结构。
为负链,而短链S为正链,其长度不确定,约为负链的50%-80%左右。基因组依靠正链5’端约240bp的粘性末端与负链缺口部位的互补维持了环状结构。")
70
在两条链的互补区两侧各有一个11碱基的直接重复序列(5’TTCAC-CTCTGC-3’),分别开始于第1842和1590核苷酸处,称为DR1和DR2。
,分别开始于第1842和1590核苷酸处,称为DR1和DR2。")
71
图8-23 HBV基因组结构(ayw株)。 :X启动子 :增强子 :核心启动子,增强子II :S启动子 :PreS1启动子 :多腺苷酸化信号
。 :X启动子 :增强子 :核心启动子,增强子II :S启动子 :PreS1启动子 :多腺苷酸化信号")
72
HBV的编码区及产物 (1)S编码区。S编码区编码乙肝表面抗原蛋白,分别编码由226个氨基酸残基的表面抗原主蛋白(SHBS)、 个氨基酸的原S1蛋白和55个氨基酸组成的原S2蛋白3部分组成。S蛋白和原S2蛋白组合在一起被称为中蛋白(MHBS),S蛋白和原S1、原S2蛋白组合在一起时被称为大蛋白(LHBS)。
S编码区。S编码区编码乙肝表面抗原蛋白,分别编码由226个氨基酸残基的表面抗原主蛋白(SHBS)、 个氨基酸的原S1蛋白和55个氨基酸组成的原S2蛋白3部分组成。S蛋白和原S2蛋白组合在一起被称为中蛋白(MHBS),S蛋白和原S1、原S2蛋白组合在一起时被称为大蛋白(LHBS)。")
73
SHBS是病毒外壳蛋白和22nm颗粒表面抗原的主要成分,占病毒蛋白的70%-90%,中蛋白和大蛋白则暴露于病毒颗粒表面。
表面抗原有很强的免疫原性,是乙肝疫苗的主要成分。

74
(2)P编码区。长2532碱基,约占全基因组3/4以上,是最长的编码区,包含全部S编码区并与C和X编码区有部分重叠。
P编码区由3个功能区和一个间隔区构成,分别为末端蛋白(又称引物酶)、间隔区、反转录酶/DNA聚合酶及RNaseH。末端蛋白是病毒进行反转录时的引物。P编码区可能先翻译成9.5×105多肽,然后加工成较小的功能型多肽。
、间隔区、反转录酶/DNA聚合酶及RNaseH。末端蛋白是病毒进行反转录时的引物。P编码区可能先翻译成9.5×105多肽,然后加工成较小的功能型多肽。")
75
(3)C编码区。该区长639碱基,翻译产物为病毒核心抗原(HBcAg)。其原初翻译产物是前核心蛋白,切除N端19肽和富含精氨酸的C末端后,成为E抗原(HBeAg),分子量为2.2×104蛋白,是核衣壳上唯一的结构蛋白。HBeAg是分泌型蛋白,可在血清中检测到。
C编码区。该区长639碱基,翻译产物为病毒核心抗原(HBcAg)。其原初翻译产物是前核心蛋白,切除N端19肽和富含精氨酸的C末端后,成为E抗原(HBeAg),分子量为2.2×104蛋白,是核衣壳上唯一的结构蛋白。HBeAg是分泌型蛋白,可在血清中检测到。")
76
(4)X编码区。是最小的编码区,编码X蛋白,覆盖了负链的缺口部位,虽然长度不等,但主要产物由154个氨基酸残基组成。X蛋白具有反式调控作用,能激活多个同源或异源启动子或增强子,与肝癌的发生有相关性。
X编码区。是最小的编码区,编码X蛋白,覆盖了负链的缺口部位,虽然长度不等,但主要产物由154个氨基酸残基组成。X蛋白具有反式调控作用,能激活多个同源或异源启动子或增强子,与肝癌的发生有相关性。")
77
HBV的复制 乙肝病毒基因组为双链DNA,但其复制并不通过半保留复制方式,而是通过反转录途径。

78
图8-24 HBV基因组复制模式图。

79
8.4 基因治疗 人类疾病的发生,其实都是人体细胞中自身基因的改变或由外源病原体的基因产物与人体基因相互作用的结果。 8.4.1 基因治疗的历史沿革 1990年,科学家第一次用反转录病毒为载体把腺苷脱氨酶基因(ADA)导入来自病人自身的T淋巴细胞,经扩增后输回患者体内,获得了成功。5年后,患者体内10%的造血细胞呈ADA基因阳性。
导入来自病人自身的T淋巴细胞,经扩增后输回患者体内,获得了成功。5年后,患者体内10%的造血细胞呈ADA基因阳性。")
80
基因治疗的两大途径 ex vivo与in vivo方式。 ex vivo途径 将含外源基因的载体在体外导入人体自身或异体(异种)被 “基因工程化的”细胞,经体外细胞扩增后输回人体。 这种方法易于操作,而且因为细胞扩增过程中对外源添加物质的大量稀释,不容易产生副作用。同时,治疗中用的是人体细胞,尤其是自体细胞,安全性好。

81
in vivo途径 将外源基因装配于特定的真核细胞表达载体上,直接导入人体内。 这种载体可以是病毒型或非病毒型,甚至是裸DNA,有利于大规模工业化生产。 这种方式导入的治疗基因以及其载体必须证明其安全性,而且导入体内之后必须能进入靶细胞,有效地表达并达到治疗目的。

82
基因治疗中的病毒载体 用于基因治疗的病毒载体应具备以下基本条件: 携带外源基因并能装配成病毒颗粒; 介导外源基因的转移和表达; 对机体没有致病力。

83
1、 病毒载体的产生 将适当长度的外源DNA插入病毒基因组的非必需区,包装成重组病毒颗粒。 将4.5kb的lacZ基因表达盒插入HSV-1病毒的UL44(编码糖蛋白C)基因的Xba I位点中,构建成重组HSV病毒。由于UL44基因产物对于HSV病毒在培养细胞中产毒性感染是非必需的,该重组病毒可以在细胞中增殖传代。
基因的Xba I位点中,构建成重组HSV病毒。由于UL44基因产物对于HSV病毒在培养细胞中产毒性感染是非必需的,该重组病毒可以在细胞中增殖传代。")
84
2、重组型病毒载体 在不改变病毒复制和包装所需的顺式作用元件的情况下,有选择性地删除病毒的某些必需基因尤其是前早期或早期基因以控制其表达,所缺失的必需基因的功能由同时导入细胞中的外源基因表达单位提供。一般通过同源重组方法将目的基因插入到病毒基因组中。

85
3、无病毒基因的病毒载体 在辅助系统的作用下,重组载体以特定形式(单链或双链DNA或RNA)被包装到不含有任何病毒基因的病毒颗粒中。这类载体的优点在于载体病毒本身安全性好,容量大。缺点在于往往需要辅助病毒参与载体DNA的包装,造成终产品中辅助病毒污染。
被包装到不含有任何病毒基因的病毒颗粒中。这类载体的优点在于载体病毒本身安全性好,容量大。缺点在于往往需要辅助病毒参与载体DNA的包装,造成终产品中辅助病毒污染。")
87
4、基因治疗中的问题 基因导入系统 基因表达的可控性 需要更多的治疗基因 1)基因导入系统。基因治疗中的关键问题是必须将治疗基因送入特定的靶细胞,并在该细胞中得到高效表达。这对于恶性肿瘤治疗尤为重要,如果不能有效地将治疗基因导入大多数肿瘤细胞,则至少要求它尽可能不进入或较少进入正常细胞。
基因导入系统。基因治疗中的关键问题是必须将治疗基因送入特定的靶细胞,并在该细胞中得到高效表达。这对于恶性肿瘤治疗尤为重要,如果不能有效地将治疗基因导入大多数肿瘤细胞,则至少要求它尽可能不进入或较少进入正常细胞。")
88
2)外源基因表达的可控性。 最理想的可控性是模拟人体内基因本身的调控形式,需要全基因或包括上下游的调控区及内含子序列,将对导入基因的载体系统产生严峻的挑战,因为今后设计的载体须有几十kb甚至上百kb的包装能力。
外源基因表达的可控性。 最理想的可控性是模拟人体内基因本身的调控形式,需要全基因或包括上下游的调控区及内含子序列,将对导入基因的载体系统产生严峻的挑战,因为今后设计的载体须有几十kb甚至上百kb的包装能力。")
89
在目的基因上游区接上gal 4的顺式元件,用带有疱疹病毒VP16/gal 4 DNA结合区的孕酮受体的变异体作为激活蛋白。

90
图8-26 导入基因的表达诱导框架图。

91
该变异体(PR-LBD△)只能与孕酮的颉抗剂(RU486)相结合而不与孕酮或其他衍生物结合。当体系中不存在RU486时,治疗基因的表达水平极低;而给予RU486后,RU486与PR-LBD△结合,激活VP16,杂种激活蛋白形成二体并通过gal 4 DNA结合区与治疗基因上游区的gal 4顺式元件结合,启动治疗基因的表达。
只能与孕酮的颉抗剂(RU486)相结合而不与孕酮或其他衍生物结合。当体系中不存在RU486时,治疗基因的表达水平极低;而给予RU486后,RU486与PR-LBD△结合,激活VP16,杂种激活蛋白形成二体并通过gal 4 DNA结合区与治疗基因上游区的gal 4顺式元件结合,启动治疗基因的表达。")
92
3)治疗基因过少。目前用于临床试验的治疗基因数量很少。绝大部分多基因疾病,如恶性肿瘤、高血压、糖尿病、冠心病、神经退行性疾病的致病基因还有待阐明,因此,可选择的靶基因不多。
治疗基因过少。目前用于临床试验的治疗基因数量很少。绝大部分多基因疾病,如恶性肿瘤、高血压、糖尿病、冠心病、神经退行性疾病的致病基因还有待阐明,因此,可选择的靶基因不多。")
Similar presentations



 的字头 AIDS 的译称,中文名为获得性免疫缺 陷综合症,是由一种人类免疫缺陷病毒 (Human.>")
,必须在活细胞内寄生并以复 制方式增殖的非细胞型微生物。 生物病毒微生物 原指一种动物来源的 毒素。 “virus” 一词源于 拉丁文。 病毒能增殖、遗传和演化,因而 具 有生命最基本的特征。>")



 ” 。 他的体内没有任何免疫系统, 没有任何抵御细菌、病毒的能力。>")



免疫缺陷病 2 )超敏反应(过敏反应) 3 )自身免疫病等.>")
 体积微小,可以通过细菌滤器 ; 结构简单,只含一种类型的核酸 ; 专性细胞内寄生,以复制方式增殖的一类非细 胞型微生物。 对抗生素不敏感,对干扰素敏感。>")

 (一)概述: ﹡ 细胞原癌基因 启动子插入 染色体移位 活化 点突变 基因扩增 癌基因 转录合成 生长因子 生长因子受体 癌基因产物(蛋白) 传导因子 核蛋白 转化蛋白 细胞恶性转化.>")



、乙型( HBV ) 丙型( HCV )、丁型( HDV )、戊型( HEV )。妊娠任 何时期都有被肝炎病毒感染的可能,其中乙型肝炎病毒 感染最常见。>")


