Download presentation
Presentation is loading. Please wait.

1
植物生理学电子教案 西南师范大学 2001

2
绪 论 一、植物生理学的定义和任务 1.定义: 植物生理学(plant physiology):是研究植物尤其是高等绿色植物生命活动规律的科学。 植物的生命活动:在水分代谢、矿质营养、光合作用和呼吸作用等基本代谢基础上,表现出种子萌发、生长、开花、结果等生长发育过程。 2.植物生理学的任务:研究和了解植物在各种环境条件下,进行生命活动的规律和机理,从而将这些成果应用于一切利用植物生产的事业中。

3
二、植物生理学的产生和发展 (一)我国古代关于植物生理学方面的论述 1.水分代谢 2.矿质营养 3.光合作用 4.呼吸储藏 5.植物生长物质 6.生长发育
我国古代关于植物生理学方面的论述 1.水分代谢 2.矿质营养 3.光合作用 4.呼吸储藏 5.植物生长物质 6.生长发育")
4
(二)植物生理学的产生与发展 1.研究开始时期(16-17世纪) 2.奠基与成长时期(18-19世纪) 3.飞跃发展时期(20世纪) (三)我国植物生理发展情况 起步晚,发展慢。 我国植物生理学起业人:钱崇澍(shu ) 我国植物生理学奠基人:李继侗、罗宗洛、汤佩松 现在一些有影响的研究人员:

5
(四)近年来植物生理学发展的特点 1.研究层次越来越广 2.学科之间相互渗透 3.理论联系实际 4.研究手段现代化
近年来植物生理学发展的特点 1.研究层次越来越广 2.学科之间相互渗透 3.理论联系实际 4.研究手段现代化")
6
三、植物生理学发展展望 研究重点:能量转变 研究焦点:膜的结构和功能 我国植生研究的主要任务: 1.深入基础理论研究(有所为,有所不为)
2.大力开展应用基础研究和应用研究

7
第二篇植物的物质生产和光能利用 代谢(metabolism):维持生命各种活动过程中的化学变化总称。
同化作用(assimilation):植物从环境中吸收简单无机物,经过各种化学变化形成各种复杂的有机物,综合成为自身的一部分,同时把太阳能转变为化学能储存于有机物中的过程。 异化作用(catabolism):植物将体内复杂的有机物分解为简单的无机物,并将储存于其中的能量释放,供生命活动的过程。 植物代谢的特点:能把环境中简单的无机物直接合成复杂的有机物。
:植物从环境中吸收简单无机物,经过各种化学变化形成各种复杂的有机物,综合成为自身的一部分,同时把太阳能转变为化学能储存于有机物中的过程。 异化作用(catabolism):植物将体内复杂的有机物分解为简单的无机物,并将储存于其中的能量释放,供生命活动的过程。 植物代谢的特点:能把环境中简单的无机物直接合成复杂的有机物。")
8
第一章植物的水分代谢 概 述

9
第一节植物对水分的需要 一、植物的含水量规律
第一节植物对水分的需要 一、植物的含水量规律 1.不同植物含水量不同; 2.同种植物生长环境不同,含水量也不同; 3.同株植物不同器官和组织含水量也不相同; 4.同一器官在不同生长期含水量不一样。

10
二、植物体内的水分存在状态 1.水分存在状态: 束缚水(bound water) 自由水(free water)
2.形成不同水分存在状态的原因:(图1) 3.两种水分存在状态与植物代谢的关系: 4.原生质的两种存在状态 溶胶(sol): 凝胶(gel): 5.粘性(viscosity)
 3.两种水分存在状态与植物代谢的关系: 4.原生质的两种存在状态. 溶胶(sol): 凝胶(gel): 5.粘性(viscosity)")
11
三、水分在植物生命活动中的作用 1.水分是构成原生质的主要成分 2.水分是代谢作用中的反应底物 3.水分是植物对物质吸收和运输的溶剂
4.水分能保持植株的固有姿态 5.水具有特殊的理化性质 (1)高比热利于体温稳定 (2)高气化热避免高温伤害 (3)具极性原生质交体稳定 (4)表面张力大利于吸附和运输 (5)透光性好利于光合。
高比热利于体温稳定. (2)高气化热避免高温伤害. (3)具极性原生质交体稳定. (4)表面张力大利于吸附和运输. (5)透光性好利于光合。")
12
第二节植物细胞对水分的吸收 一、细胞的渗透性吸水
植物的吸水方式 (一)自由能和水势 自由能 化学势 水势
自由能和水势. 自由能. 化学势. 水势.")
13
(二)渗透作用 渗透作用(osmosis): 渗透系统:
渗透作用 渗透作用(osmosis): 渗透系统:")
14
质壁分离:植物细胞由于液泡失水而使原生质体与细胞壁分离的现象。
(三)植物细胞是一个渗透系统 1.概念: 质壁分离:植物细胞由于液泡失水而使原生质体与细胞壁分离的现象。 质壁分离复原:把发生质壁分离的植物细胞放入清水或水势较高的溶液中,液泡变大,整个原生质体慢慢恢复原来状态的过程。
植物细胞是一个渗透系统. 1.概念: 质壁分离:植物细胞由于液泡失水而使原生质体与细胞壁分离的现象。 质壁分离复原:把发生质壁分离的植物细胞放入清水或水势较高的溶液中,液泡变大,整个原生质体慢慢恢复原来状态的过程。")
15
2.发生质壁分离的条件: (1)外界环境水势低于细胞水势; (2)原生质层具有选择性; (3)细胞壁与细胞质的收缩能力不同。 3.质壁分离说明以下问题: (1)原生质层具有半透膜的性质; (2)判断细胞的死活; (3)能测定细胞的渗透势。
能测定细胞的渗透势。")
16
(四)植物细胞的水势 1.典型植物细胞的水势:水势=衬质势+压力势+渗透势 2.形成液泡前植物细胞的水势:水势=衬质势 3.细胞吸水饱和时水势为0。 4.衬质势:细胞胶体物质亲水性和毛细管对自由水束缚而引起的水势降低值(实质是增加吸水力),为负值。 5.压力势:由于细胞壁压力的存在而增加的水势(它阻止吸水),一般为正值,但质壁分离时为0,剧烈蒸腾时为负。 6.膨压:细胞吸水膨胀而对细胞壁产生的压力。 7.渗透势:又叫溶质势,由于溶质颗粒的存在而使水势降低的部分(水的自由能降低),一般为负值。
,一般为正值,但质壁分离时为0,剧烈蒸腾时为负。 6.膨压:细胞吸水膨胀而对细胞壁产生的压力。 7.渗透势:又叫溶质势,由于溶质颗粒的存在而使水势降低的部分(水的自由能降低),一般为负值。")
17
(五)细胞间的水分移动 水势差异决定水流方向和速度 X Y
水势梯度:当多个细胞连在一起时,如果一端细胞的水势高,另一端的水势低,顺次下降就形成一个水势梯度。水分从水势高的地方流向水势低的地方。植物器官水分流动就遵循这一规律。 渗透势=-1.4Mpa 压力势=+0.8Mpa 水势=-0.6Mpa 渗透势=-1.2Mpa 压力势=+0.4Mpa 水势=-0.8Mpa

18
二、细胞的吸涨作用 吸涨作用(imbibition):亲水胶体吸水膨胀的现象。 大分子物质的亲水性强弱: 蛋白质>淀粉>纤维素
:亲水胶体吸水膨胀的现象。 大分子物质的亲水性强弱: 蛋白质>淀粉>纤维素")
19
三、细胞的代谢性吸水 代谢性吸水(metabolic absorption of water):利用细胞呼吸释放的能量,使水分经质膜而进入细胞的过程。 证据:呼吸与吸水正相关
:利用细胞呼吸释放的能量,使水分经质膜而进入细胞的过程。 证据:呼吸与吸水正相关")
20
第三节植物根系对水分的吸收 1.植物的吸水器官:根系 2.根系的吸水部位:根尖 3.根尖的吸水区域:根毛区(图)
4.根毛区为吸水的主要区域原因何在? (1)根毛多,吸收面积大; (2)细胞壁由果胶物质组成,亲水性强; (3)疏导组织发达。 5.其他区域: 细胞质浓厚,疏导组织不发达,对水阻力大。
根毛多,吸收面积大; (2)细胞壁由果胶物质组成,亲水性强; (3)疏导组织发达。 5.其他区域: 细胞质浓厚,疏导组织不发达,对水阻力大。")
21
一、根系吸水动力 (一)根压(root pressure): 根压:植物根系的生理活动使液流从根部上升的压力(主动吸水)。 证据:伤流、吐水
伤流(bleeding):从受伤或折断的植物组织溢出液体的现象。 伤流液(bleeding sap)成分: 吐水(guttation):从未受伤的叶片的尖端或边缘的水孔向外溢出液滴的现象。
:从受伤或折断的植物组织溢出液体的现象。 伤流液(bleeding sap)成分: 吐水(guttation):从未受伤的叶片的尖端或边缘的水孔向外溢出液滴的现象。")
22
产生根压的机理 (1)渗透理论 (2)代谢理论
渗透理论 (2)代谢理论")
23
(二)蒸腾拉力 大气——叶片——气孔下腔——下腔叶肉细胞——旁边细胞——导管——根 土壤
蒸腾拉力 大气——叶片——气孔下腔——下腔叶肉细胞——旁边细胞——导管——根 土壤")
24
二、影响根系吸收水分的主要外界条件 外界条件:大气因子和土壤因子 本节主要讲述土壤因子 (一)土壤可用水分
1.定义:永久萎蔫系数以外多余的水分。 2.永久萎蔫系数(permanent wilting coefficioent):植物叶片刚显示萎蔫之后,转到阴湿之处仍不能恢复原状,此时的土壤含水量与土壤干重的百分率(引起植物萎蔫不能因蒸腾的减弱而恢复的土壤最高百分含水量)。 3.土壤水分的分类:重力水、毛管水和束缚水。
:植物叶片刚显示萎蔫之后,转到阴湿之处仍不能恢复原状,此时的土壤含水量与土壤干重的百分率(引起植物萎蔫不能因蒸腾的减弱而恢复的土壤最高百分含水量)。 3.土壤水分的分类:重力水、毛管水和束缚水。")
25
1、高温: 加速根的老化和木质化减少吸收面积; 酶的活性下降甚至失活, 原生质流动缓慢。 2、低温: 水分粘性增大,扩散速率降低;
(二)通气状况 (三)土壤温度 1、高温: 加速根的老化和木质化减少吸收面积; 酶的活性下降甚至失活, 原生质流动缓慢。 2、低温: 水分粘性增大,扩散速率降低; 原生质粘性增大,水分不易透过; 呼吸速率下降,影响根压; 根系生长缓慢,影响吸水面积。 3、适温: (四)土壤溶液浓度
通气状况. (三)土壤温度. 1、高温: 加速根的老化和木质化减少吸收面积; 酶的活性下降甚至失活, 原生质流动缓慢。 2、低温: 水分粘性增大,扩散速率降低; 原生质粘性增大,水分不易透过; 呼吸速率下降,影响根压; 根系生长缓慢,影响吸水面积。 3、适温: (四)土壤溶液浓度.")
26
第四节蒸腾作用 蒸腾作用(transpiration):水分以气体状态,通过植物体的表面(主要是叶子)从体内散发到体外的现象。
一、蒸腾作用的生理意义和部位 1.生理意义: (1)植物吸水和运输的主要动力; (2)有利于矿质、盐类的吸收; (3)能降低叶片温度。 2. 植株蒸腾部位:幼体、成体 3.叶片蒸腾的方式:皮孔蒸腾、角质蒸腾、气孔蒸腾
植物吸水和运输的主要动力; (2)有利于矿质、盐类的吸收; (3)能降低叶片温度。 2. 植株蒸腾部位:幼体、成体. 3.叶片蒸腾的方式:皮孔蒸腾、角质蒸腾、气孔蒸腾.")
27
二、气孔运动 气孔结构特点: 1.细胞壁不均匀加厚; 2.细胞器与表皮细胞不同; 3.体积小于表皮细胞; 4.与表皮细胞间无胞间连丝。

28
(一)经过气孔的蒸腾速率 1. 气孔扩散速率特点: 比同面积自由水面的蒸发速率快50倍以上。 2
(一)经过气孔的蒸腾速率 1.气孔扩散速率特点: 比同面积自由水面的蒸发速率快50倍以上。 2.扩散速率快的原因——小孔扩散原理 (二)气孔运动 1.气孔周期性运动 2.气孔运动的原因 保卫细胞的吸水和失水
经过气孔的蒸腾速率 1.气孔扩散速率特点: 比同面积自由水面的蒸发速率快50倍以上。 2.扩散速率快的原因——小孔扩散原理 (二)气孔运动 1.气孔周期性运动 2.气孔运动的原因 保卫细胞的吸水和失水")
29
(三)气孔运动及机理: 1.淀粉-糖变化学说(starch-sugar conversion theory): 主要内容: 认为保卫细胞水势的变化是糖和淀粉互相转化的结果。关键酶为淀粉磷酸化酶(在PH 促进淀粉水解,PH 促进葡萄糖-1磷酸合成淀粉)。 在光照条件下由于保卫细胞具有叶绿体进行光合作用消耗二氧化碳细胞PH升高,促进淀粉水解,细胞内可溶物质增加,水势降低,气孔打开吸水。 在黑暗条件下, 。 存在问题: (1)对某些植物的保卫细胞无叶绿体,但气孔仍然开闭,难以解释; (2)气孔开/关在先,糖变化在后。
对某些植物的保卫细胞无叶绿体,但气孔仍然开闭,难以解释; (2)气孔开/关在先,糖变化在后。")
30
2.无机离子吸收学说(inorganic ion uptake theory):
细胞膜上有H-ATP酶,它可被蓝光和红光激活,利用ATP将H+从保卫细胞运到周围细胞,同时吸收K+和Cl-,降低了细胞的水势,气孔张开。 3.苹果酸生成学说(malate production theory): PEP+HCO-3——OAA+磷酸 OAA+NADH——苹果酸+NAD
: PEP+HCO-3——OAA+磷酸. OAA+NADH——苹果酸+NAD.")
31
(四)影响气孔运动的因素 (1)光照 为其主要因素,它促进糖、苹果酸的形成和K+、Cl-的积累。 (2)温度 影响气孔开度 (3)二氧化碳浓度 低浓度促进气孔开放,高浓度促进气孔关闭 (4)叶片含水量
影响气孔运动的因素 (1)光照 为其主要因素,它促进糖、苹果酸的形成和K+、Cl-的积累。 (2)温度 影响气孔开度 (3)二氧化碳浓度 低浓度促进气孔开放,高浓度促进气孔关闭 (4)叶片含水量")
32
三、影响蒸腾作用的内外条件 1.光照:最主要条件 2.大气的相对湿度 3.温度 4.外界空气流动速率 5.昼夜变化 气孔蒸腾水蒸气扩散过程
蒸腾速率=扩散力/扩散途径阻力=(气孔下腔蒸腾压-叶外蒸腾压)/(气孔阻力+扩散层阻力 (一)外界条件对蒸腾作用的影响 1.光照:最主要条件 2.大气的相对湿度 3.温度 4.外界空气流动速率 5.昼夜变化
/(气孔阻力+扩散层阻力 (一)外界条件对蒸腾作用的影响. 1.光照:最主要条件. 2.大气的相对湿度. 3.温度. 4.外界空气流动速率. 5.昼夜变化.")
33
(二)内部因素对蒸腾作用的影响 1.气孔频度 2.气孔大小 3.气孔开度 4.气孔下腔大小 5.气孔的特殊构造 6.叶片内部面积
内部因素对蒸腾作用的影响 1.气孔频度 2.气孔大小 3.气孔开度 4.气孔下腔大小 5.气孔的特殊构造 6.叶片内部面积")
34
(三)蒸腾作用的表示法 1. 蒸腾速率(transpiration rate): 植株在一定时间内单位叶面积蒸腾的水量。用克/平方分米
(三)蒸腾作用的表示法 1.蒸腾速率(transpiration rate): 植株在一定时间内单位叶面积蒸腾的水量。用克/平方分米.小时表示。 2.蒸腾比率(transpiration ratio): 植株每消耗1千克水所形成的干物质克数。用克表示。 3.蒸腾系数(transpiration coefficient): 又叫需水量,植株制造1克干物质所需水分的克数。用克表示。 蒸腾系数是蒸腾比率的倒数。
蒸腾作用的表示法 1.蒸腾速率(transpiration rate): 植株在一定时间内单位叶面积蒸腾的水量。用克/平方分米.小时表示。 2.蒸腾比率(transpiration ratio): 植株每消耗1千克水所形成的干物质克数。用克表示。 3.蒸腾系数(transpiration coefficient): 又叫需水量,植株制造1克干物质所需水分的克数。用克表示。 蒸腾系数是蒸腾比率的倒数。")
35
(四)减慢蒸腾速率的途径 1.减少叶面积 2.应用抗蒸腾剂 (1)抗蒸腾剂(antitranspiration):能阻碍蒸腾作用而对光合作用和生长影响不大的物质。 (2)抗蒸腾剂种类: 代谢型: 薄膜型: 反射型: 3.遮光遮风处理

36
第五节植物体内水分运输 一、水分运输的途径 1.水分从被吸收到蒸腾到体外经过的途径:
土壤溶液——根部——皮层薄壁细胞——木质部导管和管胞——茎或叶的木质部——叶片木质部膜端细胞——气孔下腔附近的叶肉细胞细胞壁——蒸腾 2.根据原生质的有无植物组织分类: 质外体(apoplast,非原生质体):没有原生质体的部分; 共质体(symplast):原生质体 3.水分在茎、叶细胞内运输的两种途径: (1)经过死细胞:长距离运输 (2)经过活细胞:适于短距离运输
:没有原生质体的部分; 共质体(symplast):原生质体. 3.水分在茎、叶细胞内运输的两种途径: (1)经过死细胞:长距离运输. (2)经过活细胞:适于短距离运输.")
37
二、水分沿导管或管胞上升的动力 1.水分沿导管、管胞上升的动力: (1)根压 (2)蒸腾拉力:主要动力 2.如何保证导管内的水柱不断?
内聚力学说(cohesion theory): 3.有关内聚力学说的争论的焦点: (1)水分上升是否需要活细胞参与; (2)木质部有气泡,水柱不可能连续,为什么水柱还能继续上升?
: 3.有关内聚力学说的争论的焦点: (1)水分上升是否需要活细胞参与; (2)木质部有气泡,水柱不可能连续,为什么水柱还能继续上升?")
38
三、水分运输的速度 1.活细胞原生质体对水流阻力很大,因亲水性物质存在形成水合膜。水流经过原生质的速度为每小时千分之一厘米;
2.水分在木质部运输速率比薄壁细胞快得多,为每小时3-45米。

39
第六节合理灌溉的生理基础 灌溉的基本任务是用最少量的水取得最大的效果。 一、作物的需水规律
作物需水量引种类而异;。如小麦生育期同一作物在不同生育期对水分的需要量也有很大的差别分五个阶段 1.萌芽到分蘖前期:需水量不大; 2.分蘖末期到抽穗期:耗水量增多,出现第一个水分临界期(critical period of water); 3.抽穗到开始灌浆:水分重要; 4.开始灌浆到乳熟期:第二个水分临界期; 5.乳熟末期到完熟期:减少水分供应。
; 3.抽穗到开始灌浆:水分重要; 4.开始灌浆到乳熟期:第二个水分临界期; 5.乳熟末期到完熟期:减少水分供应。")
40
二、合理灌溉的指标 1.从土壤湿度决定灌溉时期 2.灌溉的形态指标 长相、叶形、叶色 3.灌溉的生理指标
水势、细胞质液浓度、渗透势、气孔开度 注意具体问题具体分析

41
三、合理灌溉增产的原因 1.灌溉可改善各种生理作用,尤其是光合作用: (1)植株生长加强,叶面增大,增加光合面积;
(2)根系活动能力加强,叶片水分充足,加快光合速率,同时还能改善光合作用的午休现象; (3)茎、叶输导组织发达,提高水分和同化物的运输效率,改善光合产物分配作用,提高产量。 2.能改变栽培环境——“生态需水”
根系活动能力加强,叶片水分充足,加快光合速率,同时还能改善光合作用的午休现象; (3)茎、叶输导组织发达,提高水分和同化物的运输效率,改善光合产物分配作用,提高产量。 2.能改变栽培环境—— 生态需水")
42
矿质营养(mineral nutrition):植物对矿物质的吸收、转运和同化。
第二章 植物的矿质营养 矿质营养(mineral nutrition):植物对矿物质的吸收、转运和同化。
:植物对矿物质的吸收、转运和同化。")
43
第一节 植物必需的矿质元素 一、植物体内的元素
矿质元素(mineral element):植物燃烧后以氧化物形态存在于灰分中的元素,又称灰分元素。 氮不是矿质元素,但由于也是植物从土壤中吸收的所以也归入矿质元素来讨论。 植物体 干物质 (5-90%) 水分 (10-95%) 有机化合物 (90%) 无机化合物 (10%)
:植物燃烧后以氧化物形态存在于灰分中的元素,又称灰分元素。 氮不是矿质元素,但由于也是植物从土壤中吸收的所以也归入矿质元素来讨论。 植物体. 干物质. (5-90%) 水分. (10-95%) 有机化合物. (90%) 无机化合物. (10%)")
44
二、植物必需的矿质元素 1.如何确定何种元素为必需元素,何者不必要?
技术:溶液培养法(solution culture method)与砂培法( sand culture method) 确定方法: 2.常用培养液:Hoagland(荷格伦特)培养液 3.必需矿质元素具备的条件: (1)由于该元素缺乏,植物生育发生障碍,不能完成生活史; (2)除去该元素,则表现专一的缺乏症,而且这种缺乏是可以恢复和预防的;
与砂培法( sand culture method) 确定方法: 2.常用培养液:Hoagland(荷格伦特)培养液. 3.必需矿质元素具备的条件: (1)由于该元素缺乏,植物生育发生障碍,不能完成生活史; (2)除去该元素,则表现专一的缺乏症,而且这种缺乏是可以恢复和预防的;")
45
(3)该元素在植物营养生理上应表现出直接效果,决不是因土壤或培养基的物理、化学、微生物条件的改变而产生的间接效果。
4.植物必需的元素有碳、氢、氧、氮、硫、磷、钾、钙、镁、铁、锰、硼、锌、铜、钼、氯共16种。 大量元素(major element):植物体内含量占植物干重的0.1%以上的元素。碳、氢、氧、氮、硫、磷、钾、钙、镁9种; 微量元素(minor element):植物体内含量占植物干重的0.01%以下的元素。铁、锰、硼、锌、铜、钼、氯7种。 5.植物必需元素是一个动态概念,种类随研究深入可能会有新的元素。 思考:植物体内有多少种矿质元素?有多少种必需矿质元素?
:植物体内含量占植物干重的0.1%以上的元素。碳、氢、氧、氮、硫、磷、钾、钙、镁9种; 微量元素(minor element):植物体内含量占植物干重的0.01%以下的元素。铁、锰、硼、锌、铜、钼、氯7种。 5.植物必需元素是一个动态概念,种类随研究深入可能会有新的元素。 思考:植物体内有多少种矿质元素?有多少种必需矿质元素?")
46
三、植物必需矿质元素的生理作用 矿质元素在植物体内的的生理作用: (1)结构组成; (2)参与调节; (3)电化学作用。 (一)大量元素
1.氮 (1)吸收形式 (2)生理作用 (3)缺乏时症状
吸收形式. (2)生理作用. (3)缺乏时症状.")
47
2.磷 (1)吸收形式 (2)分布 (3)作用: A、核苷酸的组分 B、在糖类代谢中的作用 C、对氮代谢的作用 D、对脂肪转变的关系
b.糖类合成、分解和转变都需要ATP、磷酸和核苷二磷酸参加; c.磷促进糖类运输; d.光合作用需磷。 C、对氮代谢的作用 D、对脂肪转变的关系 (4)缺磷时症状
缺磷时症状.")
48
3.钾 (1)吸收状态 (2)分布 (3)作用 促进呼吸进程及核酸和蛋白质的合成 对糖类的合成运输有影响 提高抗旱性 (4)缺乏时的症状
吸收状态 (2)分布 (3)作用 促进呼吸进程及核酸和蛋白质的合成 对糖类的合成运输有影响 提高抗旱性 (4)缺乏时的症状")
49
4.硫 分布均匀 不足时蛋白质含量显著减少,叶绿素的形成也受到影响,缺乏时叶片呈黄绿色。 5.钙 主要存在于老器官 缺钙生长受抑制,严重时幼嫩器官(根尖、茎端)溃烂坏死。 6.镁 主要才能在于幼嫩器官和组织 缺镁时叶绿素不能合成,叶脉仍绿脉间变黄,有时呈红紫色,严重时形成褐斑坏死。

50
(二)微量元素 1.铁 (1)吸收形式 (2)生理作用 (3)缺乏时症状:如:黄叶病 2.锰 (1)生理作用
(2)缺乏时症状:叶绿体破坏解体(缺绿) 3.硼 (2)缺乏时症状:花而不实
缺乏时症状:叶绿体破坏解体(缺绿) 3.硼. (2)缺乏时症状:花而不实.")
51
4.锌 (1)生理作用 (2)缺乏时症状:“花白叶”病、“小叶病” 5.铜 作用 6.钼 7.氯
生理作用 (2)缺乏时症状: 花白叶 病、 小叶病 5.铜 作用 6.钼 7.氯")
52
四、作物缺乏矿质元素的诊断 (一)化学分析诊断法 待测株与正常植株比较 (二)病症诊断法 注意元素间的相互作用和元素之间的位置竞争
(三)加入诊断法 大量元素可以土壤施肥作追肥;微量元素可以用根外追肥或浸渗法。
加入诊断法. 大量元素可以土壤施肥作追肥;微量元素可以用根外追肥或浸渗法。")
53
第二节 植物细胞对矿质元素的吸收 一、生物膜 植物细胞的膜系统 (一)膜的特性和化学成分
1.特性:selective permeability 2.结构成分: 蛋白质(structural protein、functional protein)和脂质主要为磷脂(具有双亲媒性)
和脂质主要为磷脂(具有双亲媒性)")
54
(二)膜的结构 1.单位膜模型(Unit membtane model) 2. 流动镶嵌模型(fluid mosaic model) 外在蛋白(exfrinsic protein) 内在蛋白(integral protein)
 外在蛋白(exfrinsic protein) 内在蛋白(integral protein) .")
55
二、被动吸收 植物细胞对矿质元素吸收的方式:被动吸收、主动吸收和胞饮作用
被动吸收(passive absorption)指由于扩散作用或其它物理过程而进行的吸收,不需能量,又称为非代谢吸收。 (一)简单扩散(simple diffusion) 脂溶性物质和水分子顺浓度差扩散。 (二)杜南平衡(Donnan equilibrium) 细胞内可扩散正负离子浓度之积等于细胞外可扩散正负离子浓度之积的平衡。
指由于扩散作用或其它物理过程而进行的吸收,不需能量,又称为非代谢吸收。 (一)简单扩散(simple diffusion) 脂溶性物质和水分子顺浓度差扩散。 (二)杜南平衡(Donnan equilibrium) 细胞内可扩散正负离子浓度之积等于细胞外可扩散正负离子浓度之积的平衡。")
56
三、主动吸收 主动吸收(active absorption):又叫主动运输,代谢吸收,指细胞利用呼吸释放的能量做功而逆着浓度差吸收矿物质的过程。 (一)主动吸收与呼吸 关系密切,证据: 累积(accumulation):物质逆着浓度差进入细胞或组织的过程。
:物质逆着浓度差进入细胞或组织的过程。 .")
57
(二)主动吸收的机理 载体学说 1.内容: 生物膜上存在一些组分(载体R),能有选择地与外界物质(分子或离子,Me)结合,形成载体-物质复合体(MR),透过质膜,在质膜内方脱离,把物质(分子或离子,Mi )释放在细胞内。 Me+R MR Mi+R 2.有载体(carrier)参与的证据: (1)饱和效应(saturation effect) (2)离子竞争(ion competition)
参与的证据: (1)饱和效应(saturation effect) (2)离子竞争(ion competition)")
58
3.载体及作用方式 载体:指膜中专门运送物质的大分子蛋白质。又叫运输酶,透过酶。 载体作用方式: (1)扩散方式: 认为载体为透过酶,其在膜内可扩散,在扩散过程中把外界物质带入细胞内。 (2)变构方式: 认为载体为变构酶,其在膜内不可移动,它为膜的 一部分,横跨膜内外,通过变构作用,把膜外分子或离子运到细胞去。

59
质子泵学说 1.基本观点: 细胞对离子的吸收和运输是由膜上的载体电致质子泵(electrogenic proton pump)推动的。 起电致质子泵作用的是ATP酶。 2.运输过程: 3.运输方式:协同运输,主要分两类: (1)共向运输 (2)反向运输
共向运输. (2)反向运输.")
60
四、胞饮作用 1.胞饮作用(pinocytosis): 物质吸附在质膜上,通过膜的内折而转移到细胞内的攫取物质及液体的过程。 2.特点:
非选择性,可吸收大分子。 3.胞饮过程: 物质吸附——质膜内陷(物质进入)——质膜内折——小囊泡——两种去向: (1)囊泡溶解,物质留在细胞质内; (2)交给液泡。
——质膜内折——小囊泡——两种去向: (1)囊泡溶解,物质留在细胞质内; (2)交给液泡。")
61
五、溶质在液泡中的累积 思考:如何区分动植物细胞? 1. 液泡的作用: 2.积累方式:反向运输系统
3.液泡膜H+-ATP酶与质膜H+-ATP酶有何不同?

62
第三节 植物体对矿质元素的吸收 一、植物吸收矿质元素的特点 (一)对盐分和水分的相对吸收 既有关,又无关。不存在直接的依赖关系
(二)离子的选择吸收 离子选择性吸收(ion selective absorption)的表现: (1)不同植物吸收离子情况不同; (2)对同一盐的阴阳离子吸收有差异。 生理酸性盐: 生理碱性盐: 生理中性盐:
离子的选择吸收. 离子选择性吸收(ion selective absorption)的表现: (1)不同植物吸收离子情况不同; (2)对同一盐的阴阳离子吸收有差异。 生理酸性盐: 生理碱性盐: 生理中性盐:")
63
(三)单盐毒害和离子对抗 单盐毒害(toxicity of single salt):植物生长培养液中只有一种金属离子对植物起有害作用的现象。 离子对抗(ion antagonism):在发生单盐毒害的溶液中,如再加入少量的其它金属离子,即能减弱或消除单盐毒害离子之间这种作用叫离子对抗。 平衡溶液(balanced solution):对植物生长良好而无毒害作用的溶液。 思考:平衡溶液是否完全营养液?
:对植物生长良好而无毒害作用的溶液。 思考:平衡溶液是否完全营养液?")
64
二、根部对矿质元素的吸收 (一)根部吸收矿物质的区域 吸收部位为根尖,吸收区域为根尖。
根部吸收矿物质的区域 吸收部位为根尖,吸收区域为根尖。")
65
(二)根部吸收矿物质的过程 1.把离子吸附在根部表面 交换吸附(exchange absorption): 不需能,与温度无关,属非代谢性的交换吸附。 2.离子进入根部内部 (1)质外体途径 (2)共质体途径
根部吸收矿物质的过程 1.把离子吸附在根部表面 交换吸附(exchange absorption): 不需能,与温度无关,属非代谢性的交换吸附。 2.离子进入根部内部 (1)质外体途径 (2)共质体途径")
66
3.离子进入导管 机理:两种不同观点 (1)被动进入 证据:裸露玉米种根微管组织吸收、保持钾、氯离子的能力很低。 (2)主动运输 证据:运输受阻时,中柱鞘及导管周围薄壁细胞中离子多。
被动进入 证据:裸露玉米种根微管组织吸收、保持钾、氯离子的能力很低。 (2)主动运输 证据:运输受阻时,中柱鞘及导管周围薄壁细胞中离子多。")
67
三、根部对土壤中非溶解状态矿质元素的吸收
(一)根部对吸附在土壤胶体上的矿质元素的吸收 主要通过两种方式: 1.通过土壤溶液而得到 ,具体过程如下: (1)根部呼吸放出co2和土壤溶液中的H2O形成H2CO3; (2) H2CO3从细胞质表面逐渐接近土粒表面; (3)土粒表面的K+和H2CO3的H+进行离子交换; (4)K+HCO 3 -返回根表面; (5)K+和H+进行离子交换,K+便进入根部,也可连HCO3-一起进入根部。
根部对吸附在土壤胶体上的矿质元素的吸收 主要通过两种方式: 1.通过土壤溶液而得到 ,具体过程如下: (1)根部呼吸放出co2和土壤溶液中的H2O形成H2CO3; (2) H2CO3从细胞质表面逐渐接近土粒表面; (3)土粒表面的K+和H2CO3的H+进行离子交换; (4)K+HCO 3 -返回根表面; (5)K+和H+进行离子交换,K+便进入根部,也可连HCO3-一起进入根部。")
68
2.直接交换得到(接触交换,contact exchange): 在根部和土壤微粒表面上的离子不同地振动,如果根部和土壤微粒的距离小于离子振动的空间,即可发生直接交换。
: 在根部和土壤微粒表面上的离子不同地振动,如果根部和土壤微粒的距离小于离子振动的空间,即可发生直接交换。")
69
(二)根部对难溶解矿物质的利用 1.呼吸产生二氧化碳溶于水成碳酸; 2.分泌有机酸; 3.生理酸性盐。 等方式先溶解在按前述方式吸收。
根部对难溶解矿物质的利用 1.呼吸产生二氧化碳溶于水成碳酸; 2.分泌有机酸; 3.生理酸性盐。 等方式先溶解在按前述方式吸收。")
70
四、影响根部吸收矿物质的条件 根部对矿物质的吸收主要有主动吸收和交换吸附,凡能影响这两个方面任何一方面的条件均可影响。 (一)温度 一定范围内根部吸收矿质的速率随土温的升高而加快。 1.温度过高不利吸收: (1)酶钝化,速率下降; (2)细胞透性增加,原生质外流。 2.温度过低: 代谢弱,主动吸收慢; 细胞质粘性增大,离子进入困难。
温度 一定范围内根部吸收矿质的速率随土温的升高而加快。 1.温度过高不利吸收: (1)酶钝化,速率下降; (2)细胞透性增加,原生质外流。 2.温度过低: 代谢弱,主动吸收慢; 细胞质粘性增大,离子进入困难。")
71
(二)通气状况 在一定范围内,氧气供应越好,根系对矿质元素的吸收越多。
通气状况 在一定范围内,氧气供应越好,根系对矿质元素的吸收越多。")
72
(三)溶液浓度 在浓度较稀时,随着溶液浓度的升高,根部对离子的吸收数量也增加;当浓度进一步增加时,并无此关系,原因何在?离子载体的饱和效应。况且,当外界溶液浓度过大时,会使植物组织脱水,出现烧苗现象。 (四)氢离子浓度 1.直接影响: 由于组成的主要成分是蛋白质,而蛋白质是两性电解质,在不同的pH条件下带电情况不同,对外界离子的吸附情况也就不一样
氢离子浓度. 1.直接影响: 由于组成的主要成分是蛋白质,而蛋白质是两性电解质,在不同的pH条件下带电情况不同,对外界离子的吸附情况也就不一样.")
73
2.间接影响: (1)土壤溶液反应改变,可以引起溶液中养分的溶解或沉淀; (2)土壤溶液反应也影响土壤微生物的活动。 (五)离子间的相互作用 一种离子的存在会影响对另一种离子的吸收。 竞争结合位点——阻碍; 激活结合位点——促进。

74
五、植物地上部分对矿质元素的吸收 根外营养,主要指叶片营养(foliar nutrition)。
1.要保证吸收,必须保证溶液能很好地被吸附在叶片上。措施:用表面活性剂、喷雾液滴要细 2.达到细胞质的途径: (1)气孔进入 (2)角质层进入 3.影响营养元素进入叶片的内外因素: (1)叶片的生理状态,嫩叶快; (2)温度,影响代谢; (3)液面保湿时间。
气孔进入. (2)角质层进入. 3.影响营养元素进入叶片的内外因素: (1)叶片的生理状态,嫩叶快; (2)温度,影响代谢; (3)液面保湿时间。")
75
4.浓度:1.5%-2.0%以下 5.根外施肥的优点: (1)生育后期或临界营养期补充营养; (2)克服易被土壤固定肥料利用率地的不足; (3)补充微量元素。 思考:如何充分发挥根外施肥的作用?
生育后期或临界营养期补充营养; (2)克服易被土壤固定肥料利用率地的不足; (3)补充微量元素。 思考:如何充分发挥根外施肥的作用?")
76
第四节 无机养料的同化 高等植物与动物的显著区别之一就在于能将无机养料同化为有机养料。 一、硝酸盐的代谢还原 1.植物所需的氮素主要是通过从土壤中获得铵盐和硝态盐,再同化为自身组成物。 植物吸收铵盐后可以直接合成氨基酸,而硝态盐必须通过代谢还原(metabolic reduction)才能利用因为蛋白质的氮为高度还原态的氮,而硝态氮为高度氧化态氮。
才能利用因为蛋白质的氮为高度还原态的氮,而硝态氮为高度氧化态氮。")
77
2. 硝态氮的还原过程: HNO3 +2e HNO2 +2e [H2N2O2] +2e [NH2OH] +2e NH3 3
2.硝态氮的还原过程: HNO3 +2e HNO2 +2e [H2N2O2] +2e [NH2OH] +2e NH3 3.硝态氮还原为亚硝酸盐过程的部位、酶及电子传递过程: (1)硝酸盐还原为亚硝酸盐在细胞质内进行; (2)硝酸还原酶(含有钼和黄素辅酶FAD)催化; 硝酸还原酶(nitrate reductase)是一种诱导酶。 诱导酶(induced enzyme):又叫适应酶(adaptive enzyme),指植物体内本来不含有某酶,但在特定的外来物质影响下,可生成这种酶。 硝酸还原酶为诱导酶的证据: (3)电子传递过程 NADH FAD MO5+ NO3- NAD+ NADH2 MO6+ NO2-
![2. 硝态氮的还原过程: HNO3 +2e HNO2 +2e [H2N2O2] +2e [NH2OH] +2e NH3 3](http://slidesplayer.com/slide/11408995/61/images/77/2.+%E7%A1%9D%E6%80%81%E6%B0%AE%E7%9A%84%E8%BF%98%E5%8E%9F%E8%BF%87%E7%A8%8B%EF%BC%9A+HNO3+%2B2e+HNO2+%2B2e+%5BH2N2O2%5D+%2B2e+%5BNH2OH%5D+%2B2e+NH3+3.jpg "2.硝态氮的还原过程: HNO3 +2e HNO2 +2e [H2N2O2] +2e [NH2OH] +2e NH3 3.硝态氮还原为亚硝酸盐过程的部位、酶及电子传递过程: (1)硝酸盐还原为亚硝酸盐在细胞质内进行; (2)硝酸还原酶(含有钼和黄素辅酶FAD)催化; 硝酸还原酶(nitrate reductase)是一种诱导酶。 诱导酶(induced enzyme):又叫适应酶(adaptive enzyme),指植物体内本来不含有某酶,但在特定的外来物质影响下,可生成这种酶。 硝酸还原酶为诱导酶的证据: (3)电子传递过程 NADH FAD MO5+ NO3- NAD+ NADH2 MO6+ NO2-")
78
4.亚硝酸盐的还原 (1)部位:叶绿体内进行 (2)酶:亚硝酸还原酶(nitrite reductase) (3)反应过程: NO2-+6e-+8H+ NH4++2H2O (4)电子传递: 铁氧还蛋白(还原型) NO2- e- 光合作用的光反应 铁氧还蛋白(氧化型) NH4+
部位:叶绿体内进行 (2)酶:亚硝酸还原酶(nitrite reductase) (3)反应过程: NO2-+6e-+8H+ NH4++2H2O (4)电子传递: 铁氧还蛋白(还原型) NO2- e- 光合作用的光反应 铁氧还蛋白(氧化型) NH4+")
79
二、氨的同化 植物吸收铵盐以后,或当植物所吸收的硝酸盐被还原成氨后,氨就立即被同化,否则就会毒害植物。因为氨可能抑制呼吸过程中的电子传递系统。 氨的同化方式有以下几种: 1.还原氨基化:还原氨直接使酮酸氨基化形成相应氨基酸的过程,即氨与a-酮酸结合形成氨基酸的过程叫还原氨基化(reduced amination)。如: a-酮戊二酸与氨结合在谷氨酸去氢酶作用下,以NADH+H+为氢供体,还原为谷氨酸。
。如: a-酮戊二酸与氨结合在谷氨酸去氢酶作用下,以NADH+H+为氢供体,还原为谷氨酸。")
80
整个反应中必须有转氨酶和磷酸吡哆醛的参与。 如: 谷氨酸 磷酸吡哆醛 天冬氨酸
2.氨基交换作用(transamination):一种氨基酸的氨基被转移到另一种酮酸的酮基上,而使之氨基化反应,接受体便变成一种新的氨基酸,而供体则变成另一种酮酸。 整个反应中必须有转氨酶和磷酸吡哆醛的参与。 如: 谷氨酸 磷酸吡哆醛 天冬氨酸 转氨酶 a-酮戊二酸 磷酸吡哆胺 草酰乙酸 磷酸吡哆醛的作用如何?
:一种氨基酸的氨基被转移到另一种酮酸的酮基上,而使之氨基化反应,接受体便变成一种新的氨基酸,而供体则变成另一种酮酸。 整个反应中必须有转氨酶和磷酸吡哆醛的参与。 如: 谷氨酸 磷酸吡哆醛 天冬氨酸. 转氨酶. a-酮戊二酸 磷酸吡哆胺 草酰乙酸. 磷酸吡哆醛的作用如何?")
81
3. 与二氧化碳形成氨甲酰磷酸 氨与二氧化碳结合,形成氨甲酰磷酸: NH3+CO2+ATP NH2COOP+ADP 4

82
四、硫酸盐的同化 1.植物获得硫主要有两种途径:从土壤中获取硫酸根离子和叶片从空气中吸收二氧化硫,最后二氧化硫也转变为硫酸根离子,然后再进行同化。 2.同化部位:既可以在根部同化,也可以在地上部同化。 3.同化过程: (1)SO42-活化: SO42-+ATP ATP-硫酸化酶 APS+Ppi-焦磷酸 APS+ATP APS激酶 PAPS+ADP APS与PAPS可相互转化。 (2)活化硫酸盐的还原 活化硫酸根+载体蛋白 还原态Fd………半胱氨酸
SO42-活化: SO42-+ATP ATP-硫酸化酶 APS+Ppi-焦磷酸 APS+ATP APS激酶 PAPS+ADP APS与PAPS可相互转化。 (2)活化硫酸盐的还原 活化硫酸根+载体蛋白 还原态Fd………半胱氨酸")
83
五、磷酸盐的同化 1.同化部位:同化部位不限 2.同化方式: (1)氧化磷酸化作用 ADP+Pi ATP+H2O (2)光合磷酸化作用 (3)转磷酸作用(底物水平磷酸化) 1,3-二磷酸甘油酸+ADP 3-磷酸甘油酸+ATP
氧化磷酸化作用 ADP+Pi ATP+H2O (2)光合磷酸化作用 (3)转磷酸作用(底物水平磷酸化) 1,3-二磷酸甘油酸+ADP 3-磷酸甘油酸+ATP")
84
第五节 矿质元素在植物体内的运输 一、矿物质运输的形式、途径和速度
1.运输形式 (1)氮的运输形式:主要有氨基酸、酰氨,还有少量以硝酸盐形式向上运输; (2)磷酸运输形式:主要以正磷酸形态运输,但也有在根部转变为有机磷化物然后才向上运输; (3)硫的运输形式:主要以硫酸根离子形式运输,但有少数以蛋氨酸和谷光甘肽之类形式运输; (4)金属离子:以离子状态运输
氮的运输形式:主要有氨基酸、酰氨,还有少量以硝酸盐形式向上运输; (2)磷酸运输形式:主要以正磷酸形态运输,但也有在根部转变为有机磷化物然后才向上运输; (3)硫的运输形式:主要以硫酸根离子形式运输,但有少数以蛋氨酸和谷光甘肽之类形式运输; (4)金属离子:以离子状态运输")
85
2.运输途径: (1)研究方法:放射性同位素与蜡纸阻隔相结合; (2)根部吸收无机离子:木质部上升 从木质部扩散到韧皮部; (3)叶片吸收的无机离子: 向下运输:以韧皮部为主,并横向运输到木质部; 向上运输:也是通过韧皮部,但有些矿质能从韧皮部扩散到木质部而向上运输。 3.运输速度:矿质元素运输速率约为30-100cm/h.
研究方法:放射性同位素与蜡纸阻隔相结合; (2)根部吸收无机离子:木质部上升 从木质部扩散到韧皮部; (3)叶片吸收的无机离子: 向下运输:以韧皮部为主,并横向运输到木质部; 向上运输:也是通过韧皮部,但有些矿质能从韧皮部扩散到木质部而向上运输。 3.运输速度:矿质元素运输速率约为30-100cm/h.")
86
二、矿物质在植物体内的分布 (一)可参与循环的元素与不参与循环的元素 1.可参与循环的元素 某些元素进入地上部后仍成离子状态,如钾;某些元素形成不稳定化合物,如氮、磷、镁; 2.不参与循环的元素 一些在体内形成稳定化合物,不能被再利用,如硫、钙、铁、锰、硼,尤其是钙、铁、锰。 再利用的元素以磷、氮最典型,不能再利用的元素以钙最典型。
可参与循环的元素与不参与循环的元素 1.可参与循环的元素 某些元素进入地上部后仍成离子状态,如钾;某些元素形成不稳定化合物,如氮、磷、镁; 2.不参与循环的元素 一些在体内形成稳定化合物,不能被再利用,如硫、钙、铁、锰、硼,尤其是钙、铁、锰。 再利用的元素以磷、氮最典型,不能再利用的元素以钙最典型。")
87
(二)元素在体内的分布 1.参与循环的元素大多分布在生长点和嫩叶等代谢旺盛的部分; 2.不参与代谢的元素分布在老叶。 因而缺素时能参与循环的元素表现在老叶,缺不参与循环的元素,病症表现在嫩叶。 3.可移动元素在体内可重新分布,同时可以被排除体外,参与生态循环。 植株被雨淋时洗出的主要物质是钾、氮、糖、有机酸和植物激素。

88
第六节 合理施肥的生理基础 合理施肥的目的:根据矿质元素对植物的生理功能和作用,结合作物的需水规律,适时地、适量地施肥,做到少肥高效。 一、作物的需肥规律 1.不同作物对三要素所要求的绝对量和相对比例不一样; 2.同一作物不同生育期对矿质元素的吸收不一样; 3.作物在不同生育期各有明显的生育中心。 肥料优先分配到生长中心 最高生产效率期(植物营养最大效率期):不同生育期施肥,对植物生长影响不同,其中施肥营养效果最好的时期为最高生产 效率期。
:不同生育期施肥,对植物生长影响不同,其中施肥营养效果最好的时期为最高生产 效率期。")
89
二、合理施肥的指标 (一)形态指标 1.叶貌 2.叶色 (1)叶色是反映作物体内营养状况的最灵敏指标; (2)叶色是反映 植物体内代谢类型的良好指标; (3)叶色反映快、敏感。 3.茎的颜色也可以作为施肥的指标
形态指标 1.叶貌 2.叶色 (1)叶色是反映作物体内营养状况的最灵敏指标; (2)叶色是反映 植物体内代谢类型的良好指标; (3)叶色反映快、敏感。 3.茎的颜色也可以作为施肥的指标")
90
(二)生理指标: 一般以功能叶作为测定对象 1
(二)生理指标: 一般以功能叶作为测定对象 1.营养元素 结合不同施肥水平,不同产量,通过化学分析,找出不同生育期、不同组织、不同营养浓度与产量的关系。 作物养分浓度在严重缺乏与适量两个浓度之间有一个获得最高产量的最低浓度,叫临界浓度(critical concentration)。 2.酰氨 监测氮素含量 3.酶活性 某些离子与酶活性密切相关,甚至是其组分,通过对酶活性的检测,从而推测元素含量。
生理指标: 一般以功能叶作为测定对象 1.营养元素 结合不同施肥水平,不同产量,通过化学分析,找出不同生育期、不同组织、不同营养浓度与产量的关系。 作物养分浓度在严重缺乏与适量两个浓度之间有一个获得最高产量的最低浓度,叫临界浓度(critical concentration)。 2.酰氨 监测氮素含量 3.酶活性 某些离子与酶活性密切相关,甚至是其组分,通过对酶活性的检测,从而推测元素含量。")
91
三、施肥增产的原因 1.植物施肥与给动物喂饲料有本质区别:植物是从无机物到有机物,增产效果是间接的;动物是从有机物到有机物。
2.施肥可改善光合性能: 增大光合面积 提高光合能力 延长光合时间 有利于产品分配利用 施肥增产的实质在于改善光合性能,形成更多的有机物,获得增产。 3.施肥还可以改善栽培条件,特别是土壤条件。

92
四、发挥肥效的措施 1.适当灌溉 以水促肥,水是肥的开关 2.适当深耕 土壤容纳更多的水、肥,促进根系发达,增加吸收面积 3.改善光照条件 适当密植 4.改善施肥方式 (1)根外施肥 (2)深层施肥
根外施肥 (2)深层施肥")
93
第三章 植物的光合作用 碳素营养是植物的生命基础: (1)植物体的干物质有90%左右是有机化合物,有机化合物中都含有碳素;
(2)碳原子是组成所有有机化合物的主要骨架。 按碳素营养方式的不同把植物分为两类: (1)异养植物(heterophyte):只能利用现成的有机物作营养的植物; (2)自养植物(autophyte):可以利用无机碳化合物作营养的植物。 碳素同化(carbon assimilation):自养植物吸收二氧化碳转变成有机物的过程。包括:细菌光合作用、绿色植物光合作用和化能合成作用三种类型。
碳原子是组成所有有机化合物的主要骨架。 按碳素营养方式的不同把植物分为两类: (1)异养植物(heterophyte):只能利用现成的有机物作营养的植物; (2)自养植物(autophyte):可以利用无机碳化合物作营养的植物。 碳素同化(carbon assimilation):自养植物吸收二氧化碳转变成有机物的过程。包括:细菌光合作用、绿色植物光合作用和化能合成作用三种类型。")
94
第一节 光合作用的重要性 1.什么是光合作用(photosynthesis)? 绿色植物吸收光能,同化二氧化碳和水,制造有机物并释放氧的过程。 2.光合作用的重要意义: (1)把无机物变成有机物; 每秒钟地球上同化碳素超过6000吨,约40%由浮游植物同化,60%由陆生植物同化。食物、化工原料、药材 (2)蓄积太阳的能量; 每年固定3X1021J,主要能源 (3)环境保护。 如果不释放氧气,地球上的氧3000年就会用完,同时形成臭氧,防紫外辐射。 研究光合作用的意义:
? 绿色植物吸收光能,同化二氧化碳和水,制造有机物并释放氧的过程。 2.光合作用的重要意义: (1)把无机物变成有机物; 每秒钟地球上同化碳素超过6000吨,约40%由浮游植物同化,60%由陆生植物同化。食物、化工原料、药材 (2)蓄积太阳的能量; 每年固定3X1021J,主要能源 (3)环境保护。 如果不释放氧气,地球上的氧3000年就会用完,同时形成臭氧,防紫外辐射。 研究光合作用的意义:")
95
第二节 叶绿体及叶绿体色素 叶片是光合作用的主要器官,叶绿体是光合作用的重要细胞器; 离体叶绿体光合速率可达完整叶片的80-90%;
叶绿体是光合作用的形态单位,但单独一个叶绿体不一定是光合作用的完整单位。

96
一、叶绿体的结构和成分 目前采用细胞匀浆法和分级离心技术将各种细胞器分开进行分析 (一)叶绿体的结构
1.外观:椭圆形,直径3-6um,厚2-3um。叶片中叶绿体含量高每平方毫米含量为7个数量级,有利于表面积的扩大,扩大吸收利用光能和二氧化碳。 2.结构:两层选择透性膜,膜以内的物质为构成片层的底物——间质(stroma),间质中有将光能转变为化学能的场所——基粒(grana),同时,间质中还有一些具有脂类贮藏功能的由亲酯性醌类物质组成的嗜锇滴。
,间质中有将光能转变为化学能的场所——基粒(grana),同时,间质中还有一些具有脂类贮藏功能的由亲酯性醌类物质组成的嗜锇滴。")
97
外被:即叶绿体膜 间质:叶绿体以内的基础物质。是构成片层的底物,主要为可溶性蛋白质(酶)和其他活跃物质。 基粒:光合色素集中之地,光能转变为化学能的场所。 类囊体:构成叶绿体的片层系统中的每个片层都是闭喝囊状,内为水溶液。 基粒类囊体: 间质类囊体: 嗜锇滴:叶绿体间质中的容易与锇酸结合的颗粒。其主要成分为亲酯性醌类物质。生理功能是贮藏脂类物质。 不同植物或同一植物的不同部位的叶绿体内的基粒类囊体数目不同。 凡光合细胞都有类囊体。 3.原核细胞与真核细胞类囊体膜在细胞内分布不同: 4.类囊体垛叠的生理意义: (1)有效收集光能,加速光反应; (2)酶的有序排列,有利于代谢的顺利进行。
有效收集光能,加速光反应; (2)酶的有序排列,有利于代谢的顺利进行。")
98
(二)叶绿体的成分 1.水分:约占75% 2.蛋白质:叶绿体的结构基础,占干重的30-45%,作为酶催化各类反应。 3.脂类:膜成分 4.色素:光合色素 5.无机盐: 6.核苷酸(NAD+、NADP+)和醌:传递电子或氢原子。
叶绿体的成分 1.水分:约占75% 2.蛋白质:叶绿体的结构基础,占干重的30-45%,作为酶催化各类反应。 3.脂类:膜成分 4.色素:光合色素 5.无机盐: 6.核苷酸(NAD+、NADP+)和醌:传递电子或氢原子。")
99
二、光合色素的化学特性 光合色素主要分为3类:叶绿素、类胡萝卜素和藻胆素

100
(一)光合色素的化学结构 1.叶绿素(chlorophyll):在高等植物中主要含有叶绿素a和叶绿素b。 (1)物理性质: 溶解性:叶绿素a、b都不溶于水,但能溶于酒精、丙酮和石油醚等有机溶剂; 颜色:叶绿素a呈蓝绿色,叶绿素b呈黄绿色。 (2)化学性质: 分子式:
化学性质: 分子式:")
101
结构特征: 为叶绿酸的酯 分子中有四个吡咯环和一个羰基与羧基组成的副环 四个吡咯环与四个甲稀基连接成一个大环叫卟啉 镁原子位于卟啉环的中央 羰基与甲醇结合,叶绿醇第四环的丙酸结合 具有双亲媒性 (3)功能
功能.")
102
2.类胡萝卜素(carotenoid): 包括胡萝卜素(carotene)和叶黄素(xanthophyll)。 (1)物理性质 溶解性:不溶于水,但能溶于有机溶剂; 颜色:类胡萝卜素呈橙黄色,叶黄素呈黄色。 (2)结构特征:不饱和碳氢化合物,有三种同分异构体 两者区别:在于紫罗兰酮环上一个位置的羟基代替氢 (3)功能 收集光能; 防护光照伤害叶绿素。
结构特征:不饱和碳氢化合物,有三种同分异构体. 两者区别:在于紫罗兰酮环上一个位置的羟基代替氢. (3)功能. 收集光能; 防护光照伤害叶绿素。")
103
3.藻胆素(phycobilin): 藻类进行光合作用的主要色素。 常与藻类中的蛋白质结合为藻胆蛋白。结合紧密只有用强酸煮沸才能分开。 可分为藻红蛋白和藻蓝蛋白。 结构与叶绿素有相似之处。
: 藻类进行光合作用的主要色素。 常与藻类中的蛋白质结合为藻胆蛋白。结合紧密只有用强酸煮沸才能分开。 可分为藻红蛋白和藻蓝蛋白。 结构与叶绿素有相似之处。")
104
(二)色素在叶绿体内的排列 两种不同看法: 1.叶绿体的片层是由含水蛋白质层和脂类层相间排列而成。色素头尾分别插入其中。 2.叶绿体片层是脂类双层膜,球形蛋白镶嵌在脂类双层膜之内,直达膜的里外,叶绿素分子整个排列在蛋白质中。
色素在叶绿体内的排列 两种不同看法: 1.叶绿体的片层是由含水蛋白质层和脂类层相间排列而成。色素头尾分别插入其中。 2.叶绿体片层是脂类双层膜,球形蛋白镶嵌在脂类双层膜之内,直达膜的里外,叶绿素分子整个排列在蛋白质中。")
105
三、光合色素的光学特性 (一)辐射能量 光是一种电磁波,对光合作用有效的是波长为 nm之间。 根据光的波粒二相性,光同时又是离子流,这种离子称为光子。光子携带能量与波长的关系为: E=N h v/入 由此可见,光子的能量与波长成反比。
辐射能量 光是一种电磁波,对光合作用有效的是波长为 nm之间。 根据光的波粒二相性,光同时又是离子流,这种离子称为光子。光子携带能量与波长的关系为: E=N h v/入 由此可见,光子的能量与波长成反比。")
106
(二)吸收光谱 1. 连续光谱: 2. 吸收光谱(absorption spectrum): 3
(二)吸收光谱 1.连续光谱: 2.吸收光谱(absorption spectrum): 3.叶绿素的吸收光谱特征 (1)有两个最强区:640nm-660nm的红光部分和 nm的蓝紫光部分; (2)对绿光吸收最少。 4.叶绿素a、b吸收光谱比较: (1)叶绿素a在红光部分吸收带宽些,在蓝紫光部分窄些,叶绿素b相反; (2)叶绿素a在红光部分吸收带偏向长波长,在蓝紫光部分吸收带偏向短光波。 5.胡萝卜素与叶黄素的吸收光谱:最大吸收在蓝紫光部分,不吸收红光 6.藻胆素的吸收光谱:主要吸收绿光、橙光
吸收光谱 1.连续光谱: 2.吸收光谱(absorption spectrum): 3.叶绿素的吸收光谱特征 (1)有两个最强区:640nm-660nm的红光部分和 nm的蓝紫光部分; (2)对绿光吸收最少。 4.叶绿素a、b吸收光谱比较: (1)叶绿素a在红光部分吸收带宽些,在蓝紫光部分窄些,叶绿素b相反; (2)叶绿素a在红光部分吸收带偏向长波长,在蓝紫光部分吸收带偏向短光波。 5.胡萝卜素与叶黄素的吸收光谱:最大吸收在蓝紫光部分,不吸收红光 6.藻胆素的吸收光谱:主要吸收绿光、橙光")
107
(三)荧光现象与磷光现象 1.荧光现象: (1)定义:叶绿素溶液在透射光下呈绿色,而在反射光下呈红色(叶绿素a为 血红光,叶绿素b为棕红光)的现象。 (2)发生荧光现象的机理(见图): 2.磷光现象: (1)定义: (2)产生磷光现象的机理: 3.荧光磷光的寿命:荧光寿命很短为10-8—10-9S;磷光较长为10-2S。 4.叶绿素荧光现象和磷光现象的意义: 思考:荧光与磷光有何区别?
荧光现象与磷光现象 1.荧光现象: (1)定义:叶绿素溶液在透射光下呈绿色,而在反射光下呈红色(叶绿素a为 血红光,叶绿素b为棕红光)的现象。 (2)发生荧光现象的机理(见图): 2.磷光现象: (1)定义: (2)产生磷光现象的机理: 3.荧光磷光的寿命:荧光寿命很短为10-8—10-9S;磷光较长为10-2S。 4.叶绿素荧光现象和磷光现象的意义: 思考:荧光与磷光有何区别?")
108
四、叶绿素的形成 (一)叶绿素的生物合成 谷氨酸 a-酮戊二酸 脱植基叶绿素a 叶绿素a 二氧戊酸 原脱植基叶绿素a ALA Mg-原卟啉 胆色素原 原卟啉IX 尿卟啉原III 粪卟啉原III
叶绿素的生物合成 谷氨酸 a-酮戊二酸 脱植基叶绿素a 叶绿素a 二氧戊酸 原脱植基叶绿素a ALA Mg-原卟啉 胆色素原 原卟啉IX 尿卟啉原III 粪卟啉原III")
109
(二)植物的叶色 1.叶色是植物叶子各种色素的综合表现。但主要为叶绿素和类胡萝卜素两类色素的比例。 2.正常叶片中主要色素的比例 叶绿素/类胡萝卜素约为:3:1 叶绿素a/叶绿素b约为:3:1 叶黄素/胡萝卜素约为:2:1 3.关于叶色的几个问题 (1)为什么树叶一般都为绿色? (2)为什么叶片衰老时会呈黄色? (3)红叶又是如何形成的?
为什么树叶一般都为绿色? (2)为什么叶片衰老时会呈黄色? (3)红叶又是如何形成的?")
110
4.影响叶绿素形成的因素: (1)光照:为其主要影响因素 (2)温度:通过影响酶活力而影响; (3)矿质元素缺乏:主要是氮、镁、铁、锰、铜、锌 5.几种现象: 黄化现象(etiolation): 早春寒潮过后水道秧苗变白现象 6.叶绿素的开发利用:牙膏、美容、药用等

111
第三节 光合作用的机理 概 述 光合作用的三大步骤: 1.光能的吸收、传递和转换(通过原初反应完成);
概 述 光合作用的三大步骤: 1.光能的吸收、传递和转换(通过原初反应完成); 2.电能转变为活跃的化学能(通过电子传递和光合磷酸化完成); 3.活跃的化学能转变为稳定的化学能(通过碳素同化完成); 以上三个步骤又可根据反应中是否需光分为光反应和暗反应两个阶段;上述1、2步基本上属于光反应,第3步属于暗反应。 1.光反应(light reaction):必须在光下进行,由光所引起的光化反应,它主要在基粒类囊体膜(光合膜)上进行; 2.暗反应(dark reaction):可以在暗处进行的由若干酶所催化的化学反应,暗反应是在间质中进行的。
; 2.电能转变为活跃的化学能(通过电子传递和光合磷酸化完成); 3.活跃的化学能转变为稳定的化学能(通过碳素同化完成); 以上三个步骤又可根据反应中是否需光分为光反应和暗反应两个阶段;上述1、2步基本上属于光反应,第3步属于暗反应。 1.光反应(light reaction):必须在光下进行,由光所引起的光化反应,它主要在基粒类囊体膜(光合膜)上进行; 2.暗反应(dark reaction):可以在暗处进行的由若干酶所催化的化学反应,暗反应是在间质中进行的。")
112
一、原初反应 1.光合单位(photosynthetic unit):是指结合在类囊体膜上能进行光合作用的最小结构单位。
2.光合单位的组成:聚光色素系统(light-harvesting pigment system)+作用中心(reaction centre)。 3.按功能光合色素可分为以下两类: (1)作用中心色素(reaction centre pigment)又叫陷阱:少数特殊状态的叶绿素a分子属于此类,它具有光化学活性。既是光的捕捉器,又是光能的转换器。
+作用中心(reaction centre)。 3.按功能光合色素可分为以下两类: (1)作用中心色素(reaction centre pigment)又叫陷阱:少数特殊状态的叶绿素a分子属于此类,它具有光化学活性。既是光的捕捉器,又是光能的转换器。")
113
(2)聚光色素(light-harvesting pigment)又叫天线色素:没有光化学活性,只具有收集光能的作用,除作用中心色素以外的色素均具有聚光作用4. 光量子的传递: 5.光合作用中心 (1)定义: (2)组成: (3)作用:光能转化为化学能 思考:原初反应分为能量吸收、传递、转化是如何完成的?
定义: (2)组成: (3)作用:光能转化为化学能 思考:原初反应分为能量吸收、传递、转化是如何完成的? .")
114
二、电子传递和光合磷酸化 (一)光系统 1.光系统的发现 : 红降(red drop): 量子产额(quantum yield):
(一)光系统 光系统的发现 : 红降(red drop): 量子产额(quantum yield): 爱默生效应(Emerson effect): 2.两个光系统: (1)颗粒大小 (2)分布部位 (3)反应波长
光系统 1.光系统的发现 : 红降(red drop): 量子产额(quantum yield): 爱默生效应(Emerson effect): 2.两个光系统: (1)颗粒大小. (2)分布部位. (3)反应波长.")
115
(4)主要特征 3. 光合链: (二)氧的释放 1. 希尔反应(Hill reaction): 2. 放氧系统的五种状态: 3
主要特征 3. 光合链: (二)氧的释放 1. 希尔反应(Hill reaction): 2. 放氧系统的五种状态: 3")
116
(三)光合磷酸化 1. 定义: 2. 分类: (1)非循环式光合磷酸化 (2)循环式光合磷酸化 3. 光合磷酸化的机理:化学渗透学说 4
(三)光合磷酸化 1.定义: 2.分类: (1)非循环式光合磷酸化 (2)循环式光合磷酸化 3.光合磷酸化的机理:化学渗透学说 4.同化能力(assimilatory power):
光合磷酸化 1.定义: 2.分类: (1)非循环式光合磷酸化 (2)循环式光合磷酸化 3.光合磷酸化的机理:化学渗透学说 4.同化能力(assimilatory power):")
117
三、碳同化 碳素同化是光合作用的一个重要方面。从能量转换角度看,碳同化是将ATP和NADP中的活跃化学能转换为储存在糖类中稳定的化学能,较长时间供给生命活动的需要;从物质生产角度看,占植物体干重90%以上的有机物基本上都是通过碳同化形成的。 碳同化在叶绿体间质中进行,需要多种酶协同作用。 碳同化的途径有三条:卡尔文循环、C4途径、景天科酸代谢途径,但只有卡尔文循环才具有合成淀粉的能力,其他两条途径只能固定转运二氧化碳,仍需通过卡尔文循环才能完成。

118
(一)卡尔文循环 1. 卡尔文循环的别名: 光合环、还原磷酸戊糖途径(RPPP)、C3途径等。 2
(一)卡尔文循环 1.卡尔文循环的别名: 光合环、还原磷酸戊糖途径(RPPP)、C3途径等。 2.卡尔文循环分为四个阶段: (1)羧化(固定二氧化碳) 固定二氧化碳,形成3-磷酸甘油酸(PGA)
卡尔文循环 1.卡尔文循环的别名: 光合环、还原磷酸戊糖途径(RPPP)、C3途径等。 2.卡尔文循环分为四个阶段: (1)羧化(固定二氧化碳) 固定二氧化碳,形成3-磷酸甘油酸(PGA)")
119
(2)还原(贮能完成) 由PGA到DPGA(1,3-二磷酸甘油酸),再到GAPld(3-磷酸甘油醛)。
还原(贮能完成) 由PGA到DPGA(1,3-二磷酸甘油酸),再到GAPld(3-磷酸甘油醛)。")
120
(3)更新: 有GAPld经过一系列变化,再形成RuBP的过程。
更新: 有GAPld经过一系列变化,再形成RuBP的过程。")
124
(4)合成: 还原过程中形成的3-磷酸甘油醛在叶绿体内合成淀粉,也可透出叶绿体在细胞质中合成蔗糖。 C3循环变化简图:
合成: 还原过程中形成的3-磷酸甘油醛在叶绿体内合成淀粉,也可透出叶绿体在细胞质中合成蔗糖。 C3循环变化简图:")
125
3. 卡尔文循环的贮能情况: 固定3分子二氧化碳,要6个NADPH分子和9个ATP分子,形成1个PGAld分子。 4

126
(二)C4途径 1. C4途径,又叫 C4-二羧酸途径、 C4光合碳同化(PCA)环、Hatch-Slack途径。它是在卡尔文循环前附加的固定二氧化碳途径。 2. C4途径的过程: 叶肉细胞胞质中的PEP为二氧化碳受体,在PEPC作用下,生成OAA,OAA被还原为苹果酸或天冬氨酸,被运输到维管束鞘细胞中去。再脱羧后运回到叶肉细胞。 3. C4途径的调节: (1)光调节 (2)效应剂调节 (3)二价金属离子调节
光调节 (2)效应剂调节 (3)二价金属离子调节 .")
127
(三)景天科植物酸代谢 1.景天科植物酸代谢过程 2.CAM的调节 (1)短期调节 指昼夜调节方式。要完成这种调节CAMPEPC有两种形式: 晚上型: 白天型: (2)长期调节
景天科植物酸代谢 1.景天科植物酸代谢过程 2.CAM的调节 (1)短期调节 指昼夜调节方式。要完成这种调节CAMPEPC有两种形式: 晚上型: 白天型: (2)长期调节")
128
四、光合作用的产物 1. 光合作用产物(photOOyythytic prodllCt)主要是糖类,包括单糖、双糖和多糖,其中以蔗糖和淀粉最为普遍。 2.不同植物的主要光合产物不同。大多数高等植物的光合产物是淀粉,有些植物(如洋葱、大蒜)的光合产物是葡萄糖和果糖,不形成淀粉。 3.长期以来,糖类曾被认为是光合作用的唯一产物,而其他物质(如蛋白质、脂肪和有机酸)是植物利用糖类再度合成的。的确,这些物质一部分是再度合成的,但也有一部分却是光合作用直接产物,特别是在藻类和高等植物正在发育的叶片中。 4.磷酸丙糖是形成光合产物的重要中间产物。
是植物利用糖类再度合成的。的确,这些物质一部分是再度合成的,但也有一部分却是光合作用直接产物,特别是在藻类和高等植物正在发育的叶片中。 4.磷酸丙糖是形成光合产物的重要中间产物。")
129
5.淀粉是在叶绿体内合成的。当卡尔文循环形成F6P时通过G6P、G1P转变为AD P-葡萄糖(ADPG),在引子(麦芽糖)帮助下,就形成淀粉。
6.蔗糖是在细胞质内形成的。叶绿体中形成的DHAP(二羟丙酮磷酸),外运到细胞质,经过FBP、 F6P、G6P、G1P形成UDP-葡萄糖(UDPG),最后转变为蔗糖。 7.DHAP从叶绿体流出是通过专一载体“磷酸运转器”的。 每运出一个磷酸酶就有一个正磷酸离子从细胞质运回来。 8.ATP是不易透过叶绿体膜的。它通过3-磷酸甘油酸/二羟丙酮磷酸穿梭(PGA/DHAP shuttle)而到达细胞质的。 9.在叶绿体里的淀粉合成和在细胞质里的蔗糖合成会竞争底物——磷酸丙糖。具体方式见图。
,外运到细胞质,经过FBP、 F6P、G6P、G1P形成UDP-葡萄糖(UDPG),最后转变为蔗糖。 7.DHAP从叶绿体流出是通过专一载体 磷酸运转器 的。 每运出一个磷酸酶就有一个正磷酸离子从细胞质运回来。 8.ATP是不易透过叶绿体膜的。它通过3-磷酸甘油酸/二羟丙酮磷酸穿梭(PGA/DHAP shuttle)而到达细胞质的。 9.在叶绿体里的淀粉合成和在细胞质里的蔗糖合成会竞争底物——磷酸丙糖。具体方式见图。")
130
第四节光呼吸 植物的绿色细胞依赖光照,放出CO2和吸收O2的过程,被称为光呼吸(photorespiration)。
一般生活细胞的呼吸在光照或黑暗中都可以进行,对光照没有特殊要求,这种呼吸相对地称为暗呼吸(dark respiration),通常所说的呼吸就是指暗呼吸。
,通常所说的呼吸就是指暗呼吸。")
131
一、光呼吸的生物化学及功能 1.进行部位:叶绿体、线粒体、过氧化体 2.底物:乙醇酸 3.生化过程:
(1)叶绿体内:RuBP+氧+水——2-磷酸乙醇酸 2-磷酸乙醇酸+水——乙醇酸+磷酸 (2)过氧化体内:乙醇酸+氧——乙醛酸+过氧化氢 乙醛酸+谷氨酸——甘氨酸+a-酮戊二酸 (3)线粒体:甘氨酸——丝氨酸+二氧化碳 (4)过氧化体:丝氨酸——羟基丙酮酸——甘油酸 (5)叶绿体:甘油酸——3-磷酸甘油酸,进入卡尔文循环
叶绿体内:RuBP+氧+水——2-磷酸乙醇酸. 2-磷酸乙醇酸+水——乙醇酸+磷酸. (2)过氧化体内:乙醇酸+氧——乙醛酸+过氧化氢. 乙醛酸+谷氨酸——甘氨酸+a-酮戊二酸. (3)线粒体:甘氨酸——丝氨酸+二氧化碳. (4)过氧化体:丝氨酸——羟基丙酮酸——甘油酸. (5)叶绿体:甘油酸——3-磷酸甘油酸,进入卡尔文循环.")
133
3.在整个循环中氧的吸收发生于哪些细胞器?二氧化碳的释放又发生于哪些细胞器?
4.光呼吸的调节与外界条件密切有关 (1)首先是氧及二氧化碳的浓度,二氧化碳抑制光呼吸而促进光合作用, 氧则抑制光合作用而促进光呼吸; (2)随着光强、温度和 PH的增高,光呼吸也加强,其实质也是二氧化碳和氧对RuBP的竞争。 5.光呼吸的生理功能 光呼吸释放的CO2量占光合作用CO2固定量的20~27%,也就是把光合作用固定四分之一左右的碳又变成 CO2释放出去。光呼吸是一个消费过程。 为什么植物在进化过程中会演化出这样一个过程呢?它对植物有什么好处呢? (1)近年来不少人认为光呼吸可保护光合器免受伤害。在干旱和高辐射期间,气孔关闭光合组织合成乙醇酸,并使光呼吸的CO2重新固定,消耗过剩的光能,保护叶绿体免受伤害。 (2)防止氧对光合碳同化的抑制作用 (3)磷酸丙糖和氨基酸合成的补充途径
首先是氧及二氧化碳的浓度,二氧化碳抑制光呼吸而促进光合作用, 氧则抑制光合作用而促进光呼吸; (2)随着光强、温度和 PH的增高,光呼吸也加强,其实质也是二氧化碳和氧对RuBP的竞争。 5.光呼吸的生理功能. 光呼吸释放的CO2量占光合作用CO2固定量的20~27%,也就是把光合作用固定四分之一左右的碳又变成 CO2释放出去。光呼吸是一个消费过程。 为什么植物在进化过程中会演化出这样一个过程呢?它对植物有什么好处呢? (1)近年来不少人认为光呼吸可保护光合器免受伤害。在干旱和高辐射期间,气孔关闭光合组织合成乙醇酸,并使光呼吸的CO2重新固定,消耗过剩的光能,保护叶绿体免受伤害。 (2)防止氧对光合碳同化的抑制作用. (3)磷酸丙糖和氨基酸合成的补充途径.")
136
二、C3植物和C4植物的光合特征 1.概念:C3植物(最初产物为3-磷酸甘油酸)、C4植物(最初产物为草酰乙酸) 2.C4植物比C3植物具有较强的光合作用,其原因主要从两个方面来探讨: (1)从结构上看: C4植物叶片的维管束薄壁细胞较大,其中含有许多较大的叶绿体,叶绿体没有基粒或基拉发育不良;维管束鞘的外侧密接一层成环状或近于环状排列的叶肉细胞,组成了“花环型”结构。 C3植物的维管束鞘薄壁细胞较小,不含或很少叶绿体,没有“花环型”结构,维管束鞘周围的叶肉细胞排列松散。

137
C4植物进行光合作用时,只有维管束鞘薄壁细胞形成淀粉,在叶肉细胞中没有淀粉。而水稻等 C3植物由于仅有叶肉细胞含有叶绿体,整个光合过程都是在对肉细胞里进行,淀粉亦只是积累在叶肉细胞中,维管束鞘薄壁细胞不积存淀粉。 (2)在生理上,C4植物一般比C3植物具有较强的光合作用,这与PEP羧化酶活性强(PEP羧化酶的活性约为RuBPC的60多倍,对二氧化碳的亲合力远远大于,RuBPC,故C4植物能利用低浓度的二氧化碳,C3植物则不能),光呼吸弱(由于提高了CO2/O2的比率)有关。 3.几个问题 (1)为什么C4植物被称作低补偿植物? (2)为什么C4植物的耐旱性强于C3植物?
在生理上,C4植物一般比C3植物具有较强的光合作用,这与PEP羧化酶活性强(PEP羧化酶的活性约为RuBPC的60多倍,对二氧化碳的亲合力远远大于,RuBPC,故C4植物能利用低浓度的二氧化碳,C3植物则不能),光呼吸弱(由于提高了CO2/O2的比率)有关。 3.几个问题. (1)为什么C4植物被称作低补偿植物? (2)为什么C4植物的耐旱性强于C3植物?")
139
第五节光合作用的进化 1.细菌光合作用 含有色素的光合细菌,在光照下利用硫化氢、异丙醇等无机的或有机的还原剂,把二氧化碳还原为有机物的过程,称为细菌光合作用。 主要代表:红硫细菌科和红色无硫细菌科(多为嫌气性细菌)。 光合细菌的细胞不存在叶绿体,但具双层膜的球状膜粒,类似叶绿体中的类囊体,另称为载色体。载色体含有细菌叶绿素和类胡萝卜素,能吸收光能和传递光能,进行光合作用。 2. 化能合成作用 不含色素的化能合成细菌,在暗处利用硫化氢、氢、氨等氧化时,释放的化学能来同化二氧化碳为有机物的过程,称为化能合成作用。 主要代表:化能合成细菌都是好气性细菌。硝化细菌是属于化能合成细菌。硝他细菌包括两群微生物,即亚硝酸细菌和硝酸细菌。

140
3.植物碳素同化作用三类过程的异同: 过 程 碳源(氢受体) 能量来源 氢来源 绿色植物光合作用 二氧化碳 光 水 细菌光合作用 二氧化碳 光 硫化氢、有机物等 化能合成作用 二氧化碳 化学能 水、氨等 4.光合作用的进化 细菌光合作用——绿色植物光合作用——化能合成作用 思考:光合作用都要放出氧气?

141
第六节影响光合作用的因素 一、外界条件对光合速率的影响 光合速率 净光合速率(表观光合速率) 真正光合速率 (一)光照 1.光强 光饱和现象
光补偿点 阳生植物 阴生植物 阳生植物与阴生植物比较: 2.光质

142
(二)二氧化碳 植物对二氧化碳的利用与光照强度有关,在弱光情况下,只能利用较低的二氧化碳浓度,光合慢,随着光照的加强,植物就能吸收利用较高的二氧化碳浓度,光合加快。 二氧化碳补偿点: (三)温度 主要通过影响酶的活性来影响光合速率,同时还通过影响体内物质运输和分子扩散速率来影响。 低温光和速率不高和高温光合下降的原因分别是什么? 思考:对一种植物来说,二氧化碳补偿点是定值。

143
(四)矿质元素 矿质元素直接或间接影响光合作用。 1.氮、镁、铁、锰等是叶绿素生物合成所必需的矿质元素; 2.钾、磷等参与糖类代谢,缺乏时便影响糖类的转变和运输,这样也就间接影响了光合作用;同时,磷也参与光合作用中间产物的转变和能量传递,所以对光合作用影响很大; 3.在一定范围内,营养元素越多,光合速率就越快。三要素中以氮肥对光合作用的效果最明显; 4.追施氮肥促使光合速率的原因有两方面:一方面是促进叶片面积增大,叶片数目增多,增加光合面积,这是间接的影响。另一方面是直接的影响,即影响光合能力。

144
(五)水分 水分缺乏主要是间接的影响光合作用下降: 1.缺水——气孔关闭——二氧化碳吸收受阻——光合速率下降 2.缺水——叶片淀粉水解加强,糖类堆积——光合产物输出缓慢——光合速率下降 (六)氧 1.氧胁迫 2.氧抑制光合作用的原因: (1) 加强氧与二氧化碳对RuBP结合的竞争,提高光呼吸速率; (2)氧能与NADP+竞争接受电子, NADPH合成量就少,碳同化所需的还原能力减少; (3)氧接受电子后形成的超氧自由基,会破坏光合膜; (4)在强光下,氧参与光合色素的光氧化,破坏光合色素,等等。
 加强氧与二氧化碳对RuBP结合的竞争,提高光呼吸速率; (2)氧能与NADP+竞争接受电子, NADPH合成量就少,碳同化所需的还原能力减少; (3)氧接受电子后形成的超氧自由基,会破坏光合膜; (4)在强光下,氧参与光合色素的光氧化,破坏光合色素,等等。")
145
(七)光合速率的日变化 1.在温暖的日子里,如水分供应充足,太阳光照成为主要矛盾,光合过程为单峰曲线。
2.当晴天无云而太阳光照强烈时,光合进程便形成双峰曲线,中午前后光合速率下降,呈现“午休”现象。 3.为什么会出现午休现象? ( 1)水分在中午供给不上,气孔关闭; ( 2)二氧化碳供应不足; (3)光合产物淀粉等来不及分解运走,积累在叶肉细胞的细胞质中,阻碍细胞内二氧化碳的运输。
水分在中午供给不上,气孔关闭; ( 2)二氧化碳供应不足; (3)光合产物淀粉等来不及分解运走,积累在叶肉细胞的细胞质中,阻碍细胞内二氧化碳的运输。")
146
二、内部因素对光合速率的影响 (一)不同部位
由于叶绿素具有接受和转换能量的作用,所以,植株中凡是绿色的、具有叶绿素的部位都进行光合作用,在一定范围内,叶绿素含量越多,光合越强。同一植物不同部位的叶片光合速率,因叶子发育状况不同而呈规律性的变化。 (二)不同生育期 一株作物不同生育期的光合速率,一般都以营养生长中期为最强,到生长末期就下降。
不同生育期. 一株作物不同生育期的光合速率,一般都以营养生长中期为最强,到生长末期就下降。")
147
第七节植物对光能的利用 一、植物的光能利用率
1.太阳常数(solar constant):地球外层垂直于太阳光的平面上,收到的太阳能量称为太阳常数。 2.光能利用率(efficiency for solar energy utilization):是指植物光合作用所累积的有机物所含的能量,占照射在单位地面上的日光能量的比率。 3.计算光能利用率的基准及优缺点 (1)从全生育期出发,计算全生育期的光能利用率,优点是比较全面,着眼于最大限度利用全生育期的光能(如通过间作、套种以增大全田的光能利用率),但计算比较笼统。 (2)从经济器官形成期出发,计算形成经济器官时期这一段时间的光能利用率,范围小,计算较准确,但是未能考虑生育前、中期的光能利用情况,不够全面。
:地球外层垂直于太阳光的平面上,收到的太阳能量称为太阳常数。 2.光能利用率(efficiency for solar energy utilization):是指植物光合作用所累积的有机物所含的能量,占照射在单位地面上的日光能量的比率。 3.计算光能利用率的基准及优缺点. (1)从全生育期出发,计算全生育期的光能利用率,优点是比较全面,着眼于最大限度利用全生育期的光能(如通过间作、套种以增大全田的光能利用率),但计算比较笼统。 (2)从经济器官形成期出发,计算形成经济器官时期这一段时间的光能利用率,范围小,计算较准确,但是未能考虑生育前、中期的光能利用情况,不够全面。")
148
二、提高光能利用率的途径 (一)延长光合时间 延长光合时间就是最大限度地利用光照时间,提高光能利用率。延长光合时间的措施有:
1.提高复种指数 复种指数:就是全年内农作物的收获面积对耕地面积之比。提高复种指数就是增加收获面积,延长单位土地面积上作物的光合时间。 2.延长生育期 在不影响耕作制度的前提下,适当延长作物的生育期。 3.补充人工光照 在小面积的栽培中,当阳光不足或日照时间过短时,还可用人工光照补充。

149
(二)增加光合面积 光合面积即植物的绿色面积,主要是叶面积。但叶面积过大,又会影响群体中的通风透光而引起一系列矛盾。所以,光合面积要适当。
1.合理密植 合理密植是提高光能利用率的主要措施之一。不可太稀,不可太密。种得过稀,个体发展较好,但群体得不到充分发展,光能利用率低。种得过密,下层叶子受到光照少,在光补偿点以下,变成消费器官,光合生产率减弱,也会减产。 2.改变株型 优良株型即秆矮,叶直而小、厚,分蘖密集。株型改善,就能增加密植程度,增大光合面积,耐肥不倒伏,充分利用光能,提高光能利用率。

150
(三)加强光合效率 1.增加二氧化碳浓度 有三个措施值得试行:(1)控制栽值规格和肥水,因地制宜选好行向,使后期通风良好。(2)增施有机肥料,放出二氧化碳。(3)深施碳酸氢铵肥料。 温室效应(greenhouse effect):指在地球周围的大气层中,人类无限制地向地球大气层中排放二氧化碳,使二氧化碳浓度不断增长。本来太阳辐射下来的热,地球以红外线形式重新辐射到空间。由于大气层中的二氧化碳能强烈地吸收红外线,太阳辐射的能量在大气层中就“易入难出”,温度上升,似温室一样。 2.降低光呼吸 措施主要有两种:(1)利用光呼吸抑制剂去抑制光呼吸,提高光合效率。( 2 )改变环境成分,尤其增加二氧化碳浓度,使核酮糖二磷酸羧化酶/氧化酶的羧化反应占优势,减少其氧化反应的比例(减少光呼吸),光能利用率就能大大提高。
:指在地球周围的大气层中,人类无限制地向地球大气层中排放二氧化碳,使二氧化碳浓度不断增长。本来太阳辐射下来的热,地球以红外线形式重新辐射到空间。由于大气层中的二氧化碳能强烈地吸收红外线,太阳辐射的能量在大气层中就 易入难出 ,温度上升,似温室一样。 2.降低光呼吸. 措施主要有两种:(1)利用光呼吸抑制剂去抑制光呼吸,提高光合效率。( 2 )改变环境成分,尤其增加二氧化碳浓度,使核酮糖二磷酸羧化酶/氧化酶的羧化反应占优势,减少其氧化反应的比例(减少光呼吸),光能利用率就能大大提高。")
151
第四章 植物的呼吸作用 前面各章都是说明植物把外界物质改造为自身物质的过程,是新陈代谢的同化作用方面;呼吸作用,是将植物体内的物质不断分解的过程,是新陈代谢的异化作用方面。呼吸作用释放的能量供给各种生理活动的需要,它的中间产物在植物体各主要物质之间的转变起着枢纽作用,所以,呼吸作用是植物代谢中心,十分重要。

152
第一节 呼吸作用的概念及其生理意义 一、呼吸作用的概念 呼吸作用(respiration)包括有氧呼吸和无氧呼吸。
第一节 呼吸作用的概念及其生理意义 一、呼吸作用的概念 呼吸作用(respiration)包括有氧呼吸和无氧呼吸。 1.有氧呼吸(aerobic respiration)指生活细胞在氧气的参与下,把某些有机物质彻底氧化分解,放出二氧化碳并形成水,同时释放能量的过程。 2.无氧呼吸(anaerobic respiration)一般指在无氧条件下,细胞把某些有机物分解成为不彻底的氧化产物,同时释放能量的过程。这个过程用于高等植物,习惯上称为无氧呼吸,如应用于微生物,则惯称为发酵(fermentation)。 3.有氧呼吸是由无氧呼吸进化而来的。
包括有氧呼吸和无氧呼吸。 1.有氧呼吸(aerobic respiration)指生活细胞在氧气的参与下,把某些有机物质彻底氧化分解,放出二氧化碳并形成水,同时释放能量的过程。 2.无氧呼吸(anaerobic respiration)一般指在无氧条件下,细胞把某些有机物分解成为不彻底的氧化产物,同时释放能量的过程。这个过程用于高等植物,习惯上称为无氧呼吸,如应用于微生物,则惯称为发酵(fermentation)。 3.有氧呼吸是由无氧呼吸进化而来的。")
153
二、呼吸作用的生理意义 1.呼吸作用提供植物生命活动所需要的大部分能量
呼吸作用释放能量的速度较慢,而且逐步释放,适合于细胞和用。释放出来的能量,一部分转变为热能而散失掉,一部分以ATP形式贮存着。 2.呼吸过程为其他化合物合成提供原料 呼吸过程产生一系列的中间产物,这些中间产物很不稳定,成为进一步合成植物体内各种重要化合物的原料,也就是在植物体内有机物转变方面起着枢纽作用。

154
第二节 植物的呼吸代谢途径 一、糖酵解 1.定义:淀粉在无氧状态下分解成丙酮酸的过程,通称为糖酵解(glycolysis)。
第二节 植物的呼吸代谢途径 一、糖酵解 1.定义:淀粉在无氧状态下分解成丙酮酸的过程,通称为糖酵解(glycolysis)。 发酵是指微生物分解糖类产生酒精或乳酸的过程。 2.底物:一般为淀粉 3.发生部位:细胞质 4.过程: 5.能量状况: 6意义:供能、中间产物、为糖异生提供基本途径
。 发酵是指微生物分解糖类产生酒精或乳酸的过程。 2.底物:一般为淀粉. 3.发生部位:细胞质. 4.过程: 5.能量状况: 6意义:供能、中间产物、为糖异生提供基本途径.")
155
二、三羧酸循环 1.别名:TCA、Krebs环 2.底物:丙酮酸 3.过程 4.能量与关键物 5.值得注意的几个问题: 第一、三羧酸循环中一系列的脱羧反应是呼吸作用释放二氧化碳的来源。三羧酸循环过程中释放的二氧化碳,不是靠大气中的氧直接把碳氧化,而是靠被氧化底物中的氧和水分子中的氧来实现的。 第二、在三羧酸循环中有五次脱氢过程,氢经过一系列呼吸传递体的传递,释放出能量,最后与氧结合成水。因此,氢的氧化过程实际是放能过程。 第三、三羧酸循环是糖、脂肪、蛋白质和核酸及其他物质的共同代谢过程。

156
三、戊糖磷酸途径 1. 不经过无氧呼吸生成丙酮酸而进行有氧呼吸的途径,葡糖-6-磷酸可直接氧化脱氢,故亦称已糖磷酸途径(HMP途径),又称磷酸葡糖酸途径、戊糖磷酸途径(简称PPP)。 2. 进行部位:细胞质内 3过程:氧化阶段、非氧化阶段 4能量 5.氧化-还原辅酶不同是PPP与EMP-TCA的重要区别之一 6两种途径在葡萄糖降解中所占的比例,随植物的种类、器官、年龄和环境而异。

157
第三节生物氧化 有机物质在生物体内进行氧化(伴随着还原),包括消耗氧生成二氧化碳、水和放出能量的过程,称为生物氧化(biological oxidation)。 一、呼吸链 1.呼吸链(respiratory chain):就是呼吸代谢中间产物的电子和质子,沿着一系列有顺序的电子传递体组成的电子传递途径,传递到分子氧的总轨道。呼吸链是由许多氧化还原迅速而可逆的电子传递体所组成的,所以又称电子传递链(electron transport chain)。 2.组成:氢传递体和电子传递体 氢传递体传递氢(包括质子和电子),这一类氢传递体是作为脱氢酶的辅酶或辅基。植物细胞的主要酶辅助因素有下列三种: NAD即辅酶 1、NADP即辅酶11)和F(黄素蛋白),它们都能进行氧化还原。
:就是呼吸代谢中间产物的电子和质子,沿着一系列有顺序的电子传递体组成的电子传递途径,传递到分子氧的总轨道。呼吸链是由许多氧化还原迅速而可逆的电子传递体所组成的,所以又称电子传递链(electron transport chain)。 2.组成:氢传递体和电子传递体. 氢传递体传递氢(包括质子和电子),这一类氢传递体是作为脱氢酶的辅酶或辅基。植物细胞的主要酶辅助因素有下列三种: NAD即辅酶 1、NADP即辅酶11)和F(黄素蛋白),它们都能进行氧化还原。")
158
呼吸链中的电子传递体是指细胞色素体系,它们只传递电子。细胞色素是一类以铁卟啉为辅基(色素辅基)的结合蛋白质,根据吸收光谱的不同可分为a、b和c三类,每类又再分为若干种。细胞色素传递电子的机理,主要是通过铁卟啉辅基中的铁离子完成的。 3.高等植物呼吸链的构成及传递 呼吸链各电子载体的顺序是固定不变的,而且电子只能从底物传递到氧分子。其原因,一方面是各个酶系统有专一性;另一方面是各电子载体的氧化还原电位不同,电子总是从低电位向高电位传递。底物脱氢反应时电位最低(丢失电子的倾向大),顺次下来,分子氧最高。所以底物电子总是流向氧分子。 4.末端氧化酶(terminal oxidase):
,顺次下来,分子氧最高。所以底物电子总是流向氧分子。 4.末端氧化酶(terminal oxidase):")
159
二、氧化磷酸化 1.氧化磷酸化(oxidative phosphorylation):氧化过程伴随着ATP的合成,即氧化作用与磷酸化作用同时进行,这一过程称为氧化磷酸化。 2. P/O比:每吸收一个氧原子所酯化无机磷酸(Pi)分子数的比,称为P/O比(P/O ratio)。也可以说, P/O比是指每消耗一个氧原子有几个ADP变成ATP,因此P/O比亦称ADP/O比。 3.解偶联剂(uncoupling agent):象2,4-二硝基苯酚(DNP)等药剂可阻碍磷酸化(形成高能磷酸键)而不影响氧化(电子传递),就使偶联反应遭到破坏一类物质为解偶联剂。 4.呼吸抑制剂:能使电子传递链打断,磷酸化作用因得不到氧化作用所放出的能量,则氧化磷酸化也不能进行,甚至生命将因此告终的药剂。
分子数的比,称为P/O比(P/O ratio)。也可以说, P/O比是指每消耗一个氧原子有几个ADP变成ATP,因此P/O比亦称ADP/O比。 3.解偶联剂(uncoupling agent):象2,4-二硝基苯酚(DNP)等药剂可阻碍磷酸化(形成高能磷酸键)而不影响氧化(电子传递),就使偶联反应遭到破坏一类物质为解偶联剂。 4.呼吸抑制剂:能使电子传递链打断,磷酸化作用因得不到氧化作用所放出的能量,则氧化磷酸化也不能进行,甚至生命将因此告终的药剂。")
160
三、呼吸酸在细胞中的位置 1.研究方法:细胞匀浆法和分级离心技术将细胞器分离,然后分别测定细胞各部分(亚颗粒)的生化反应。 2.酶的存在位置
(1)EMP和PPP的酶是在细胞质的可溶部分(即衬质); (2)TCA和生物氧化的酶都在线粒体内。 (3)细胞质是糖酵解和PPP进行的场所,线粒体是TCA和生物氧化进行的位置。 3.线粒体 成分 结构、功能
EMP和PPP的酶是在细胞质的可溶部分(即衬质); (2)TCA和生物氧化的酶都在线粒体内。 (3)细胞质是糖酵解和PPP进行的场所,线粒体是TCA和生物氧化进行的位置。 3.线粒体. 成分. 结构、功能.")
161
四、呼吸过程中的氧化酶 (一)氧化酶 1.细胞色素氧化酶(cytochrome oxidase)
2.酚氧化酶(phenol oxidase) 单酚氧化酶(monoPhenol oxidase)(亦称酪氨酸酶)和多酚氧化酶(polyphenol oxidase)(亦称儿茶酚氧化酶)。 红茶、绿茶制法 3.抗坏血酸氧化酶(ascorbic acid oxidase) 与植物的受精过程有密切关系,并且有利于胚珠的发育。 4.黄素氧化酶(flavin oxidase,亦称黄酶) 存在于乙醛酸循环体中,能把脂肪酸氧化分 解,最后形成过氧化氢。 5.交替氧化酶(alternate oxidase)
 单酚氧化酶(monoPhenol oxidase)(亦称酪氨酸酶)和多酚氧化酶(polyphenol oxidase)(亦称儿茶酚氧化酶)。 红茶、绿茶制法. 3.抗坏血酸氧化酶(ascorbic acid oxidase) 与植物的受精过程有密切关系,并且有利于胚珠的发育。 4.黄素氧化酶(flavin oxidase,亦称黄酶) 存在于乙醛酸循环体中,能把脂肪酸氧化分 解,最后形成过氧化氢。 5.交替氧化酶(alternate oxidase)")
162
(三)呼吸酶的适应性 1.以对温度来说,黄酶对温度变化不敏感,温度降低时黄酶活性降低不多,故在低温下生长的植物及其器官以这种酶为主,而细胞色素氧化酶对温度变化最为敏感,在果实成熟过程中酶系统的更替正好反映了酶系统对温度的适应。例如,柑桔的果实有细胞色素氧化酶、多酚氧化酶和黄酶,在果实未成熟时,气温尚高,呼吸氧化是以细胞色素氧化酶为主,到果实成熟时,气温渐低,则以黄酶为主,这就保证了成熟后期呼吸活动的水平,同时也反映了植物对低温的适应。 2.以对氧浓度的要求来说,细胞色素氧化酶对氧的亲和力量强,所以在低氧浓度的情况下,仍能发挥良好的作用,而酚氧化酶和黄酶对氧的亲和力弱、只有较高氧浓度下才能顺利地发挥作用。苹果果向中酶分布也正好反映了酶对氧供应的适应,内层以细胞色素氧化线为主,表层以黄酶和酚氧化酶为主。水稻幼苗之所以能够适应淹水低氧条件,是因为在低氧时细胞色素氧化酶活性加强而黄酶活性降低之故。

163
(四)植物呼吸代谢的多样性 1.它表现在呼吸途径多样性(EMP、TCA和PPP等); 2.呼吸链电子传递系统的多样性(电子传递主路、几条支路和抗氰途径); 3.末端氧化系统的多样性(细胞色素氧化酶、酚氧化酶、抗坏血酸氧化酶、乙醇酸氧化酶和交替氧化酶)。 这些多样性,是植物在长期进化过程中对不断变化的环境的适应表现。

164
第四节呼吸过程中能量的贮存和利用 一、贮存能量 1. 贮能的主要形式:高能键,主要是高能磷酸键,特别是三磷酸腺苷; 2.能量贮存过程
1. 贮能的主要形式:高能键,主要是高能磷酸键,特别是三磷酸腺苷; 2.能量贮存过程 (1)氧化磷酸化(oxidative phosphorylation): (2)转磷酸化作用(transphosphorylation): (3)非氧化作用
氧化磷酸化(oxidative phosphorylation): (2)转磷酸化作用(transphosphorylation): (3)非氧化作用.")
165
二、利用能量 1.1mol葡萄糖完全氧化得多少ATP,如何计算? 2.植物有氧呼吸的能量利用率:

166
三、光合作用和呼吸作用的关系 1.光合作用与呼吸作用比较 2.光合与呼吸的辨证关系:
1.光合作用所需的ADP(供光合磷酸化产生ATP之用)和辅酶NADP(供产生NADPH之用),与呼吸作用所需的 ADP和 NADP是相同的。这两种物质在光合和呼吸中共用。 2.光合作用的碳循环与呼吸作用的成糖磷酸途径基本上是正反反应的关系。光合作用和呼吸作用之间有许多糖类(中间产物)是可以交替使用的。 3.光合释放的氧可供呼吸利用,而呼吸作用释放的二氧化碳亦能为光合作用所同化。
和辅酶NADP(供产生NADPH之用),与呼吸作用所需的 ADP和 NADP是相同的。这两种物质在光合和呼吸中共用。 2.光合作用的碳循环与呼吸作用的成糖磷酸途径基本上是正反反应的关系。光合作用和呼吸作用之间有许多糖类(中间产物)是可以交替使用的。 3.光合释放的氧可供呼吸利用,而呼吸作用释放的二氧化碳亦能为光合作用所同化。")
167
第五节 呼吸作用的调节和控制 一、巴斯德效应和糖酵解的调节
第五节 呼吸作用的调节和控制 一、巴斯德效应和糖酵解的调节 1.巴斯德效应(Pasteur effect):氧可以降低糖类的分解代谢和减少糖醉解产物的积累,这种现象被称为巴斯德效应。 2.糖酵解调节 (1)过程 (2)调节酶:磷酸果糖激酶和丙酮酸激酶 (3)关键酶:磷酸果糖激酶 (4)效应物:负效应物:ATP和柠檬酸 正效应物:ADP、Pi
:氧可以降低糖类的分解代谢和减少糖醉解产物的积累,这种现象被称为巴斯德效应。 2.糖酵解调节. (1)过程. (2)调节酶:磷酸果糖激酶和丙酮酸激酶. (3)关键酶:磷酸果糖激酶. (4)效应物:负效应物:ATP和柠檬酸. 正效应物:ADP、Pi.")
168
二、戊糖磷酸途径和三羧酸循环的调节 1.戌糖磷酸途径主要是受NADPH的调节。 2. 三羧酸循环的调节是多方面的。
NADH是主要负效应物,NADH水平过高,会抑制丙酮酸脱氢酶(多酶复合体)、异柠檬酸脱氨酶、苹果酸脱氢酶和苹果酸酶等的活性。 ATP对柠檬酸合成酶和苹果酸脱氢酶起抑制作用。 AMP对酮戊二酸脱氢酶活性、COA对苹果酸酶活性都有促进作用。 根据质量作用原理,产物(如乙酸CoA税用酸CoA和草酰乙酸)的浓度过高时也会抑制各自有关酶的活性。
、异柠檬酸脱氨酶、苹果酸脱氢酶和苹果酸酶等的活性。 ATP对柠檬酸合成酶和苹果酸脱氢酶起抑制作用。 AMP对酮戊二酸脱氢酶活性、COA对苹果酸酶活性都有促进作用。 根据质量作用原理,产物(如乙酸CoA税用酸CoA和草酰乙酸)的浓度过高时也会抑制各自有关酶的活性。")
169
三、腺苷酸 荷的调节 1. 腺苷酸(adenylic acid)对呼吸的影响是多方面的,它能调整细胞的代谢。
2.能荷(energy charge)={[ATP]+1/2[ADP]}/{[ATP]+[ ADP]+[ AMP]} 3.如果细胞的腺青酸全部为ATP,则能荷为1。如果全部为ADP,则为0.5;活细胞的能行一般稳定在0.75~0.95。 4.当ATP/AMP比率很低时,表示能量贮存很少,能行低,ATP合成反应加快,能行变大时,合成反应就减慢。因此,能荷是细胞中ATP合成反应和利用反应的调节因素。
={[ATP]+1/2[ADP]}/{[ATP]+[ ADP]+[ AMP]} 3.如果细胞的腺青酸全部为ATP,则能荷为1。如果全部为ADP,则为0.5;活细胞的能行一般稳定在0.75~0.95。 4.当ATP/AMP比率很低时,表示能量贮存很少,能行低,ATP合成反应加快,能行变大时,合成反应就减慢。因此,能荷是细胞中ATP合成反应和利用反应的调节因素。")
170
第六节影响呼吸作用的因素 一、呼吸速率和呼吸商 呼吸作用的指标有两个,即呼吸速率和呼吸商。
1.呼吸速率(respiratory rate):可以用植物的单位鲜重、干重或原生质(以含氮量)表示,在一定时间内所放出的二氧化碳的体积,或所吸收的氧的体积来表示。 2.呼吸商又称呼吸系数,是表示呼吸底物的性质和氧气供应状态的一种指标。植物组织在一定时间内,放出二氧化碳的mol数与吸收氧气的mol数的比率叫做呼吸商(respiratory quotient简称R.Q.)或呼吸系数(respiratory coefficient)。
:可以用植物的单位鲜重、干重或原生质(以含氮量)表示,在一定时间内所放出的二氧化碳的体积,或所吸收的氧的体积来表示。 2.呼吸商又称呼吸系数,是表示呼吸底物的性质和氧气供应状态的一种指标。植物组织在一定时间内,放出二氧化碳的mol数与吸收氧气的mol数的比率叫做呼吸商(respiratory quotient简称R.Q.)或呼吸系数(respiratory coefficient)。")
171
二、内部因素对呼吸速率的影响 1.不同植物具有不同的呼吸速率。一般可以说,凡是生长快的植物呼吸速率就快,生长慢的植物呼吸速率也慢;
2.同一植株不同防器官,因为代谢不同、非代谢(结构)组成的相对比重不同,以及与氧气接触程度不同,所以呼吸速率有很大的差异; 3.同一器官的个别组织,在呼吸速率上彼此也很不相同; 4.同一器官在不同的生长过程中,呼吸速率亦有较大的变化。
组成的相对比重不同,以及与氧气接触程度不同,所以呼吸速率有很大的差异; 3.同一器官的个别组织,在呼吸速率上彼此也很不相同; 4.同一器官在不同的生长过程中,呼吸速率亦有较大的变化。")
172
三、外界条件对呼吸速率的影响 外界条件对呼吸速率的影响也和对所有生理进程的影响一样。可以分为三个基点,即最低、最适和最高三点。某环境因素使某生理过程持续地最快地进行,此点就是最适点,而使该生理过程能够进行的最低或最高的限度,分别称为最低点和最高点。 (一)温度 1.温度之所以能影响呼吸速率主要是影响呼吸酶的活性。 2.超过最适点,呼吸速率则会随着温度的增高而下降 3.呼吸作用的最低温度依植物体的生理状况而有差异。 4.呼吸作用的最适温度总是比光合作用的最适温度高,
温度. 1.温度之所以能影响呼吸速率主要是影响呼吸酶的活性。 2.超过最适点,呼吸速率则会随着温度的增高而下降. 3.呼吸作用的最低温度依植物体的生理状况而有差异。 4.呼吸作用的最适温度总是比光合作用的最适温度高,")
173
3.呼吸商的特点: (1)呼吸底物性质与呼吸商有密切关系 A、当呼吸底物是糖类(如葡萄糖)而又完全氧化时,呼吸商是1。 B、如果呼吸底物是一些富于氢的物质,如脂肪或蛋白质,则呼吸商小于1。 C、如果呼吸底收只是一些比糖类含氧较多的物质,如已局部氧化的有机酸。则呼吸商大干1。 (2)氧气供应状态时呼吸商影响也很大。
氧气供应状态时呼吸商影响也很大。")
174
5.较长时间维持最快呼吸速率的温度才是呼吸最适温度;
6.温度系数(temperature coefficient):由于温度升高10℃而引起反应速度的增加的倍数 ,通常称为温度系数。 7.应用:根据温度对呼吸速率影响的原理,实践上贮藏果实和蔬菜时应该使温度降低,以减少呼吸消耗,但是温度又不能低到破坏植物组织的程度,否则,细胞受损害,对病原微生物的抵抗力大减,也易腐烂损坏。
:由于温度升高10℃而引起反应速度的增加的倍数 ,通常称为温度系数。 7.应用:根据温度对呼吸速率影响的原理,实践上贮藏果实和蔬菜时应该使温度降低,以减少呼吸消耗,但是温度又不能低到破坏植物组织的程度,否则,细胞受损害,对病原微生物的抵抗力大减,也易腐烂损坏。")
175
(二)氧气 1.氧气影响呼吸速率和呼吸性质; 2.为什么植物不能长时间忍受无氧呼吸? (1)无氧呼吸产生酒精。酒精使细胞质的蛋白质变性; (2)能量消耗过多。因为无氧呼吸利用每mol葡萄糖产生的能量很少,相当于有氧呼吸的百分之几,植物要维持正常生理需要,就要消耗更多的有机物,这样,植物体内养料耗损过多; (3)没有丙酮酸氧化过程,需要中间产物形成的物质就无法继续合成。
能量消耗过多。因为无氧呼吸利用每mol葡萄糖产生的能量很少,相当于有氧呼吸的百分之几,植物要维持正常生理需要,就要消耗更多的有机物,这样,植物体内养料耗损过多; (3)没有丙酮酸氧化过程,需要中间产物形成的物质就无法继续合成。")
176
(三)二氧化碳 1.浓度高抑制呼吸。 2.高浓度二氧化碳抑制呼吸这个原理可利用于贮藏果蔬。 (四)机械损伤 机械损伤会显著加快组织的呼吸速率,理由有两个; (1)是原来氧化酶与其底物在结构上是隔开的,机械损伤使原来的间隔破坏,酚类化合物就迅速地被氧化;(2)是机械损伤使某些细胞转变为分生组织状态,以形成愈伤组织去修补伤处,这些生长旺盛的生长细胞的呼吸速率,当然比原来休眠或成熟组织的呼吸速率快得多。
是原来氧化酶与其底物在结构上是隔开的,机械损伤使原来的间隔破坏,酚类化合物就迅速地被氧化;(2)是机械损伤使某些细胞转变为分生组织状态,以形成愈伤组织去修补伤处,这些生长旺盛的生长细胞的呼吸速率,当然比原来休眠或成熟组织的呼吸速率快得多。")
177
第七节 呼吸作用与农业生产 一、呼吸作用与作物栽培 1.呼吸是代谢的中心
第七节 呼吸作用与农业生产 一、呼吸作用与作物栽培 1.呼吸是代谢的中心 既能释放能量以供作物各种生理过程的需要,它的中间产物又在作物体各主要有机物之间的转变起着枢纽作用,所以呼吸不仅影响作物的无机营养和有机营养,亦影响物质的运输和转变,最后就导致新细胞和器官的形成,植株长大。 2.栽培措施 3.常见的生理障碍

178
二、呼吸作用与粮食贮藏 1.种子呼吸加快的危害:引起有机物的大量消耗;呼吸放出的水分,又会使粮堆湿度增大;呼吸放出的热量,又使粮温增高,反过来又促使呼吸增强,最后导致发热霉变,使粮食变质变量。 2.贮藏条件: (1)晒干; (2)净(无微生物和害虫); (3)低温(通风和密闭); (4)低氧。 3.保管方法:化学保管法、脱氧保管法。
晒干; (2)净(无微生物和害虫); (3)低温(通风和密闭); (4)低氧。 3.保管方法:化学保管法、脱氧保管法。")
179
三、呼吸作用与果蔬贮藏 1.果树贮藏的特点:不能干燥。
2.与粮食贮藏相同,果蔬贮藏亦可以应用降低氧浓度或降低温度的原理。同时注意避免机械损伤。 3.贮藏方法——自体保藏法: 由于果实蔬菜本身不断呼吸,放出二氧化碳,在密闭环境里,二氧化碳浓度逐渐增高,抑制呼吸作用(但容器中二氧化碳浓度不能超过10%,否则,果实中毒变坏),可以稍为延长贮藏期间。如能密封加”低温(1~5℃),贮藏时间更长。
,可以稍为延长贮藏期间。如能密封加 低温(1~5℃),贮藏时间更长。")
180
第五章 光合产物运输分配和植物信息传递系统

181
第一节植物体内的信息传递 物理信号 化学信号 细胞信号传导 细胞信号传导的分子途径

182
一、胞间信号及其传递 二、跨膜信号转换 三、细胞内信号 (一)磷酸肌醇信号系统
磷酸肌醇信号系统")
183
第二节 高等植物的运输 叶片是进行光合作用合成有机物的主要基地,植株各器官、各组织所需要的有机物,主要由叶片供应。
第二节 高等植物的运输 叶片是进行光合作用合成有机物的主要基地,植株各器官、各组织所需要的有机物,主要由叶片供应。 从有机物生产发源地到消耗地或贮藏场所之间,必然有一个运输过程。 细胞组织之间之所以能互通有无,制造或吸收器官与消费或积累器官之间之所以能同时共存,作物体之所以能保持一个统一的整体,都完全依赖着有效的运输机构。 有机物运输是决定产量高低和品质好坏的一个重要因素;光合作用形成大量有机物,生物产量高,但人类所需要的是较有经济价值的部分。从较高的生物产量变成较高经济产量,这就存在一个有机物的运输问题,即同化产物的分配问题。

184
一、有机物的短距离运输 (一)细胞内的有机物运输
1.方式:细胞内有机物的运输,主要通过扩散和布朗运动等在细胞器和细胞溶质之间移动,也可通过原生质运动使细胞器移位。 2.速度:各种物质在细胞内部运输速度不同。 (二)细胞间的有机物运输 (1)质外体运输; (2)共质体运输。
细胞间的有机物运输. (1)质外体运输; (2)共质体运输。")
185
二、有机物的长距离运输 (一)运输的途径 1.有机物的运输途径:由韧皮部 2.研究方法: 环割试验 同位素示踪法 (二)运输方向
韧皮部内的物质可同时作双向运输。同化物也可以横向运输。但正常状态下其量甚微,只有当纵向运输受阻时,横向运输才加强。

186
(三)运输的速度和形式 1.速度:一般是约100cm/h 不同植物的有机物运输速度是多少有点差异; 同一作物,由于生育期不同,有机物运输的速度也有所不同。 2.研究方法:利用蚜虫的吻刺法结合同位素示踪进行测定。 3.筛管汁液成分: 干重90%以上是蔗糖,蔗糖是糖类运输的主要形式; 某些植物的筛管汁液中还有少量棉子糖,水苏糖和毛蕊花糖; 还含有微量的氨基酸、酰氨、生长物质、有机酸和糖醇。

187
三、运输特点 1.运输的速率和速度很高 2.韧皮部汁液有溢泌现象 3.筛管中的汁液很复杂 4.韧皮部中物质可以同时向两个方向运输
5.韧皮部中的物质的运输只有当细胞活着的时候才能发生,并且与呼吸作用有关。

188
第三节韧皮部运输的机制 一、装载和卸出 1.韧皮部运输的关键:同化物怎样从“源”细胞装载入筛管分子,以及怎样从筛管分子卸出到“库”细胞。
2.同化物装载到韧皮部细胞的途径:由叶肉细胞,经过自由空间再进入韧皮部筛管分子,也就是“共质体一质外体一共质体”途径。糖-质子协同运输机理。 3.韧皮部卸出有两种途径: (1)共质体途径; (2)质外体途径:有两种情况 A.蔗糖在质外体部分代谢,被转化酶分裂为葡萄糖或果糖; B.蔗糖通过质外体并无变化,直接进入贮藏空间。
共质体途径; (2)质外体途径:有两种情况. A.蔗糖在质外体部分代谢,被转化酶分裂为葡萄糖或果糖; B.蔗糖通过质外体并无变化,直接进入贮藏空间。")
189
二、有机物运输的机理 (一)压力流动学说(Pressure flow theory)
基本论点:有机物在筛管中随着液流的流动而移动。这种液流的流动,是由于输导系统两端的压力势的差异而引起的,所以称为压力流动学说。 物理原理: 3.证据: 4.难题: (1)筛板充满韧皮蛋白纤丝和胼胝质,对糖溶液流动有相当大的阻力。要使糖液保持那么快的速度,其所需的压力势差异比筛管的大得多。(2)这个学说对一个筛管细胞同时进行双向运输的事实难以解释。
筛板充满韧皮蛋白纤丝和胼胝质,对糖溶液流动有相当大的阻力。要使糖液保持那么快的速度,其所需的压力势差异比筛管的大得多。(2)这个学说对一个筛管细胞同时进行双向运输的事实难以解释。")
190
(二)细胞质泵动学说(cytoplasmic pumping theory)
可以解释双向运输现象。同一筛管中不同胞纵连束,在相同时候可进行相反方向的移动,糖分也就向相反方向运输。 (三)收缩蛋白学说(contractile protein theory) 筛管分子的内腔有一种由微纤丝相连的网状结构。微纤丝一端固定,一端游离于筛管细胞质内,微纤丝似鞭毛一样颤动。 活的筛管分子内有两种运动:脉膊状流动,集体流动。 微纤丝之所以运动因为它是由韧皮蛋白(P-蛋白)收缩丝所组成的。 总的来看,同化物运输的学说很多,但从运输动力来说,可分为两大方面:渗透动力和代谢动力。
收缩蛋白学说(contractile protein theory) 筛管分子的内腔有一种由微纤丝相连的网状结构。微纤丝一端固定,一端游离于筛管细胞质内,微纤丝似鞭毛一样颤动。 活的筛管分子内有两种运动:脉膊状流动,集体流动。 微纤丝之所以运动因为它是由韧皮蛋白(P-蛋白)收缩丝所组成的。 总的来看,同化物运输的学说很多,但从运输动力来说,可分为两大方面:渗透动力和代谢动力。")
191
第四节同化物的分配及影响因素 一、同化物的分配规律 (一)分配方向 以不同生育期来说,光合产物优先分配到生长中心。
以不同叶位的叶片来说,它的光合产物分配有“就近供应,同侧运输”的特点。 以不同叶龄的叶片来说,叶片生长尚未完成时,本身产生较少光合产物,不向外运出,反而从别处输入光合产物,供助叶生长用。一俟叶片长成,形成大量光合产物,就向外运送光合产物。

192
(二)分配规律 1.供应能力 供应能力就是指该器官或部位的同化产物能否输出以及输出多少的能力。 “代谢源” 2.竞争能力 “代谢库”(metabolic sink)是指消耗或贮藏同化物养料的器官。同化物之所以能够向“代谢库”输送,主要是“代谢库”有一种“拉力”。 3.运输能力 运输能力包括输出和输入部分之间输导系统联系、畅通程度和距离远近。 总之,有机物究竟分配到哪里,分配多少,以供应能力、竞争能力和运输能力三个因素的综合而定。

193
二、影响和调节同化物运输的因素 一、内部因素 (一)源器官的代谢调节 叶内蔗糖的浓度与下列因素有关
1.绿色细胞内蔗糖合成酶与蔗糖-6-磷酸合成酶的活性与蔗糖浓度有关 2.叶细胞内K/Na 的比值 3.叶细胞内无机磷含量与蔗糖浓度有关 (二)库器官的代谢调节 1.库器官的特点 2.库强度 3.激素对库活力的调节
库器官的代谢调节. 1.库器官的特点. 2.库强度. 3.激素对库活力的调节.")
194
二、外界条件对有机物运输的影响 凡影响输导组织两端有机物浓度和代谢强度的因素,皆影响物质的运输速度。 (一)温度 (二)水分 (三)光
(四)矿质元素 影响同化物运输的矿质元素,主要是硼、磷、钾等元素。 (五)植物激素
矿质元素. 影响同化物运输的矿质元素,主要是硼、磷、钾等元素。 (五)植物激素.")
195
第三篇植物的生长和发育 新陈代谢是植物生长和发育的基础。 生长(growth)是植物体积的增大,它是通过细胞分裂和伸长来完成的;
发育(development)则指在整个生活史中,植物体的构造和机能从简单到复杂的变化过程,它的表现就是细胞、组织和器官的分化(differentiation)。 在植物体的发育过程中,由于部分细胞逐渐丧失了分裂和伸长的能力,向不同方向分化,从而形成了具有各种特殊构造和机能的细胞、组织和器官。
则指在整个生活史中,植物体的构造和机能从简单到复杂的变化过程,它的表现就是细胞、组织和器官的分化(differentiation)。 在植物体的发育过程中,由于部分细胞逐渐丧失了分裂和伸长的能力,向不同方向分化,从而形成了具有各种特殊构造和机能的细胞、组织和器官。")
196
第六章植物生长物质 1.植物生长物质(plant growth substances):是一些调节植物生长发育的物质。 2.分类:
(1)植物激素(plant hormones或phytohormones); (2)植物生长调节剂(plant growth regulators)。 3.植物激素是指一些在植物体内合成,并从产生处运送到别处,对生长发育产生显著作用的微量有机物;
植物激素(plant hormones或phytohormones); (2)植物生长调节剂(plant growth regulators)。 3.植物激素是指一些在植物体内合成,并从产生处运送到别处,对生长发育产生显著作用的微量有机物;")
197
4.植物生长调节剂是指一些具有植物激素活性的人工合成的物质。
5.目前,大家公认的植物激素有五类,即生长素类、赤霉素类、细胞分裂素类、乙烯和脱落酸。前三类都是促进生长发育的物质,脱落酸是一种抑制生长发育的物质,而乙烯则主要是一种促进想官成熟的物质。 6.有些生长调节剂的生理效能比植物激素的还好

198
第一节生长素类 一、生长素的发现 1.生长素(auxin)是最早发现的一种植物激素。 2.生长素发现的一些关键性试验:
(1)英国的Charles Darwin和他的儿子Francis Darwin:胚芽鞘产生向光弯曲是由于幼苗在单侧光照下,产生某种影响,从上部传到下都,造成背光面和向光面生长快慢不同。 (2)丹麦的Boysen-Jensen(1913)的实验证明,Darwin所谓“影响”是可以透过凝胶片的。
英国的Charles Darwin和他的儿子Francis Darwin:胚芽鞘产生向光弯曲是由于幼苗在单侧光照下,产生某种影响,从上部传到下都,造成背光面和向光面生长快慢不同。 (2)丹麦的Boysen-Jensen(1913)的实验证明,Darwin所谓 影响 是可以透过凝胶片的。")
199
(3)匈牙利的A.Paal(1914,1919)的实验证明胚芽鞘顶端的扩散性化合物具促进生长的效应。
(4)荷兰的F.W.Went(1928)证明促进生长的影响可从路尖传到琼胶,再传到去顶胚芽鞘,这种影响确是化学本质,Went称之为生长素。创造燕麦试法,定量测定生长素含量,推动了植物激素的研究。 3.生长素的分离: 荷兰的F.Kogl(1934)等从玉米油、根霉、麦芽等分离和纯化刺激生长的物质,经鉴定是IAA。这个工作大大推动了植物激素研究向前发展。 4.植物体内的生长素类物质以吲哚乙酸最普遍。
荷兰的F.W.Went(1928)证明促进生长的影响可从路尖传到琼胶,再传到去顶胚芽鞘,这种影响确是化学本质,Went称之为生长素。创造燕麦试法,定量测定生长素含量,推动了植物激素的研究。 3.生长素的分离: 荷兰的F.Kogl(1934)等从玉米油、根霉、麦芽等分离和纯化刺激生长的物质,经鉴定是IAA。这个工作大大推动了植物激素研究向前发展。 4.植物体内的生长素类物质以吲哚乙酸最普遍。")
200
二、生长素在植物体内的分布和传导 1.分布: 生长素在高等植物中分布很广,根、茎、叶、花、果实、种子及胚芽鞘中都有。但大多集中在生长旺盛的部分,而在趋向衰老的组织和器官中则甚少。 2.含量:每克鲜重植物材料,一般含10~100ng生长素。 3.生长素运输: (1)包括两个系统: 一是需能的、单方向的极性运输(即生长素只能从植物体的形态学上端向下端运输,而不能倒转过来运输)系统;
包括两个系统: 一是需能的、单方向的极性运输(即生长素只能从植物体的形态学上端向下端运输,而不能倒转过来运输)系统;")
201
二是被动的、通过韧皮部的、无极性运输系统。
(2)生长素是通过韧皮部运输的,运输的速度大约为l-2.4cm。 (3)生长素的极性运输是主动的运输过程: A.其运输速度比物理的扩散约大10倍; B.在缺氧的条件下会严重地阻碍生长素的运输; C.生长素可以逆浓度陡度运输。 (4)极性运输机理尚不完全清楚,这里介绍化学渗透学说(chemiosmotic theory)。 主要内容是:细胞大部质膜比下部质膜易于透过生长素。
生长素是通过韧皮部运输的,运输的速度大约为l-2.4cm。 (3)生长素的极性运输是主动的运输过程: A.其运输速度比物理的扩散约大10倍; B.在缺氧的条件下会严重地阻碍生长素的运输; C.生长素可以逆浓度陡度运输。 (4)极性运输机理尚不完全清楚,这里介绍化学渗透学说(chemiosmotic theory)。 主要内容是:细胞大部质膜比下部质膜易于透过生长素。")
202
三、生长素生物合成 1.合成部位: 叶原基、嫩叶和发育中的种子。成熟叶片和根尖也产生生长素,但数量很微。 2.合成前体:色氨酸。 3.合成途径: (1)吲哚丙酮酸途径:本途径在高等植物中占优势,对一些植物来说是唯一的生长素合成途径。 (2)色胺途径:植物中占少数,而大麦、燕麦、烟草和番茄枝条中则同时进行这两条途径。 (3)吲哚乙醇途径:
色胺途径:植物中占少数,而大麦、燕麦、烟草和番茄枝条中则同时进行这两条途径。 (3)吲哚乙醇途径:")
203
四、自由生长素和束缚生长素 1.分类: 生长素在植物组织内呈不同化学状态。 易于提取的生长素称为自由生长素(free auxin), 受酶解、水解或自溶作用从束缚物释放出来的那部分生长素,称为束缚生长素(bound auxin)。 2.活性: 自由生长素具有活性,而束缚生长素则没有活性。自由生长素和束缚生长素可相互转变。束缚生长素是生长素与不同化合物(糖、氨基酸)结合而形成的,类型不同,生理作用也有差异。
结合而形成的,类型不同,生理作用也有差异。")
204
3.束缚生长素在植物体内的作用: (1)作为贮藏形式; (2)作为运输形式; (3)解毒作用; (4)防止氧化; (5)调节自由生长素含量。
作为贮藏形式; (2)作为运输形式; (3)解毒作用; (4)防止氧化; (5)调节自由生长素含量。")
205
(一)酶促降解 分为脱羧降解和不脱羧降解。 1.脱羧降解 2.不脱羧降解: (二)光氧化
五、生长素的降解 (一)酶促降解 分为脱羧降解和不脱羧降解。 1.脱羧降解 在IAA氧化酶作用下氧化时脱羧产生二氧化碳。 2.不脱羧降解: IAA降解物仍然保留IAA侧链的两个碳原子,不脱羧。 (二)光氧化 体外的吲哚乙酸在植物色素核黄素催化下,可被光氧化,产物也是吲哚醛和亚甲基羟吲哚。 植物体内自由生长素水平是随着生长发育而变化的。它是通过生物合成、生物降解、运输、结合和区域化(贮存在液泡)等途径,调节体内生长素水平,以适应生长发育的需要。
酶促降解 分为脱羧降解和不脱羧降解。 1.脱羧降解. 在IAA氧化酶作用下氧化时脱羧产生二氧化碳。 2.不脱羧降解: IAA降解物仍然保留IAA侧链的两个碳原子,不脱羧。 (二)光氧化. 体外的吲哚乙酸在植物色素核黄素催化下,可被光氧化,产物也是吲哚醛和亚甲基羟吲哚。 植物体内自由生长素水平是随着生长发育而变化的。它是通过生物合成、生物降解、运输、结合和区域化(贮存在液泡)等途径,调节体内生长素水平,以适应生长发育的需要。")
206
六、生长素的作用机理 生长素作用于细胞时,首先与受体结合,经过一系列过程,使细胞壁介质酸化和蛋白质形成。最终表现出细胞长大。
(一)生长素受体 1.生长素与细胞中的生长素受体结合,是生长素在细胞中作用的开始。生长素受体是激素受体的一种。 2.激素受体(hormone receptor):具有与激素特异地结合的物质,能识别激素信号,并能将信号转化为一系列细胞内的生物化学变化,最终表现出生物效应。 3.生长素受体在细胞中的存在位置有多种说法。主要有两种:一种在质膜;另一种是在细胞质(或细胞核),前者促进细胞,壁松弛,后者促进核酸和蛋白质的合成。生长素受体是不耐热的蛋白质。
生长素受体. 1.生长素与细胞中的生长素受体结合,是生长素在细胞中作用的开始。生长素受体是激素受体的一种。 2.激素受体(hormone receptor):具有与激素特异地结合的物质,能识别激素信号,并能将信号转化为一系列细胞内的生物化学变化,最终表现出生物效应。 3.生长素受体在细胞中的存在位置有多种说法。主要有两种:一种在质膜;另一种是在细胞质(或细胞核),前者促进细胞,壁松弛,后者促进核酸和蛋白质的合成。生长素受体是不耐热的蛋白质。")
207
(二)细胞壁酸化作用 1.生长素和质膜上的受体质子泵(ATP酶)结合,作为活化质子泵的效应物,使质子泵活化,把细胞质内的质子(H+)分沁到细胞壁去。当细胞壁环境酸化后,一些在细胞壁中的酶被激活。此外,在酸性环境中,对酸不稳定键的键(H)易断裂。因此,细胞壁多糖分子间结构交织点破裂,联系松弛,细胞壁可塑性增加。 2.酸-生长学说: 由于生长素和酸性溶液都可同样促进细胞伸长,生长素促使H+分泌速度和细胞伸长速度一致,故此,把生长素诱导细胞壁酸化并使其可塑性增大而导致细胞伸长的理论,称为酸-生长学说(acid growth theory)。 3.生长素作用于质子泵而引起细胞壁酸化、松弛软化的过程,是一种快反应,一般仅维持十多分钟。然而生长素促进生长可以稳定几个小时,这就暗示生长素除了促进H+分泌外,必定还有其他作用。
结合,作为活化质子泵的效应物,使质子泵活化,把细胞质内的质子(H+)分沁到细胞壁去。当细胞壁环境酸化后,一些在细胞壁中的酶被激活。此外,在酸性环境中,对酸不稳定键的键(H)易断裂。因此,细胞壁多糖分子间结构交织点破裂,联系松弛,细胞壁可塑性增加。 2.酸-生长学说: 由于生长素和酸性溶液都可同样促进细胞伸长,生长素促使H+分泌速度和细胞伸长速度一致,故此,把生长素诱导细胞壁酸化并使其可塑性增大而导致细胞伸长的理论,称为酸-生长学说(acid growth theory)。 3.生长素作用于质子泵而引起细胞壁酸化、松弛软化的过程,是一种快反应,一般仅维持十多分钟。然而生长素促进生长可以稳定几个小时,这就暗示生长素除了促进H+分泌外,必定还有其他作用。")
208
(三)核酸和蛋白质的合成 1.生长素促进细胞生长除了使细胞壁松弛外,还促进RNA和蛋白质的合成。
证据:用IAA处理豌豆上胚轴DNA、RNA含量增多。且RNA合成抑制剂放线菌素D可减少RNA合成速率,如用蛋白质合成抑制剂亚胺环己酮处理,则抑制蛋白质的合成。 2.生长素对双子叶植物生长的作用模式: 3.综上所述,生长素一方面活化质膜上的ATP酶,促使细胞壁环境酸化,增加可塑性,从而增强细胞渗透吸水的能力,液抱不断增大,细胞体积也加大;另一方面,生长素促进RNA和蛋白质的合成,为原生质体和细胞壁的合成提供原料,保持持久性生长。

209
七、生长素的生理作用 1.促进:增加雌花,单性结实,子房壁生长,细胞分裂,维管束分化,光合产物分配,叶片扩大,茎伸长,偏上性,乙烯产生,叶片脱落,形成层活性,伤口愈合,不定根的形成。种子发芽,侧根形成,根瘤形成,种子和果实生长,座果,顶端优势。 2. 抑制:花朵脱落,侧枝生长,块根形成,叶片衰老。 3.生长素对细胞伸长的促进作用,与生长素浓度、细胞年龄和植物器官种类有关。 (1)一般在低浓度时可促进生长,浓度较高则会抑制生长,如果浓度更高则会使植物受伤; (2)细胞年龄不同对生长素的敏感程度不同。一般来说,幼嫩细胞对生长素反应非常敏感,老细胞则比较迟钝。 (3)不同器官对生长素的反应敏感也不一样,根最敏感,茎最不敏感,芽居中。
一般在低浓度时可促进生长,浓度较高则会抑制生长,如果浓度更高则会使植物受伤; (2)细胞年龄不同对生长素的敏感程度不同。一般来说,幼嫩细胞对生长素反应非常敏感,老细胞则比较迟钝。 (3)不同器官对生长素的反应敏感也不一样,根最敏感,茎最不敏感,芽居中。")
210
八、人工合成的生长素类及其应用 1.人工合成生长素类: (1)有吲哚环的:最早发现的是吲哚丙酸(简称IPA)和吲哚丁酸(简称IBA)。
(2)没有吲哚环而具有萘环的化合物:a-萘乙酸(简称NAA) (3)具有苯环的化合物,如2,4-二氯苯氧乙酸(简称2,4-D),也都有吲哚乙酸的生理活性。 (4)还发现和这些化合物有关的化合物,如萘氧乙酸(简称NOA)、 2, 4, 5-三氯苯氧乙酸。 (5)还有一类合成的生长素衍生物,如a-(对氯苯氧基)异丁酸(简称PCIB),它本身不具或具很少生长素活性,但对生长素有专一的抑制效应,故称为抗生长素(antiauxin)。
没有吲哚环而具有萘环的化合物:a-萘乙酸(简称NAA) (3)具有苯环的化合物,如2,4-二氯苯氧乙酸(简称2,4-D),也都有吲哚乙酸的生理活性。 (4)还发现和这些化合物有关的化合物,如萘氧乙酸(简称NOA)、 2, 4, 5-三氯苯氧乙酸。 (5)还有一类合成的生长素衍生物,如a-(对氯苯氧基)异丁酸(简称PCIB),它本身不具或具很少生长素活性,但对生长素有专一的抑制效应,故称为抗生长素(antiauxin)。")
211
PCIB在植物体内与生长素竞争受体,所以抑制正常的生长素作用。如果施用过量的生长素。可以克服PCIB的抑制作用。
2.人工合成生长素类应用于农业生产,主要有下列几个方面: (1)促使插枝生根 (2)阻止器官脱落 防止离层激形成。 (3)促进结实 可形成无籽果实。 (4)促进菠萝开花
促使插枝生根. (2)阻止器官脱落. 防止离层激形成。 (3)促进结实. 可形成无籽果实。 (4)促进菠萝开花.")
212
第二节 赤霉素类 赤霉素(gibberellins)是日本人黑泽英一从水稻恶苗病的研究中发现的。 一、赤霉素的结构 1.结构:赤霉素是一种双萜,由4个异成二烯单位组成。其基本结构是赤霉素烷。 2.分类: (1)根据分子中碳原子总数的不同分为C19和C20两类赤霉素。前者包含的种类大大多于后者,生理活性高;而后者生理活性低。各类赤霉素都含有羧酸,所以赤霉素是酸性。 (2)根据结合情况分为:自由赤霉素和结合赤霉素。
根据分子中碳原子总数的不同分为C19和C20两类赤霉素。前者包含的种类大大多于后者,生理活性高;而后者生理活性低。各类赤霉素都含有羧酸,所以赤霉素是酸性。 (2)根据结合情况分为:自由赤霉素和结合赤霉素。")
213
二、赤霉素的分布与运输 1.分布: 广泛分布于被子植物、裸子植物、蕨类植物、褐藻、绿藻、真菌和细菌中。赤霉素的种类很多,至1991年底已报道有86种赤霉素。 赤霉素和生长素一样,较多存在干生长旺盛的部分。 果实和种子(尤其是未成熟种子)的赤霉素含量比营养器官的多两个数量级。每个器官或组织都含有两种以上的赤霉素,而且赤霉素的种类、数量和状态都因植物发育时期而异。 2.运输: 赤霉素在植物体内的运输没有极性。根尖合成的赤霉素沿导管向上运输,而嫩叶产生的赤霉素则沿筛管向下运输。 运输速度,不同植物差异很大。
的赤霉素含量比营养器官的多两个数量级。每个器官或组织都含有两种以上的赤霉素,而且赤霉素的种类、数量和状态都因植物发育时期而异。 2.运输: 赤霉素在植物体内的运输没有极性。根尖合成的赤霉素沿导管向上运输,而嫩叶产生的赤霉素则沿筛管向下运输。 运输速度,不同植物差异很大。")
214
三、赤霉素的生物合成 赤霉素在高等植物中生物合成的位置至少有三处:发育着的果实(或种子),伸长着的茎端和根部。 细胞内合成的部位:质体 GA12-7-醛是各种赤霉素相互转变的分支点。 甲瓦龙酸(又名甲羟戊酸)—牛儿牛儿焦磷酸(GGPP)—古巴焦磷酸(CPP)——内根-贝壳杉烯—内根-贝壳杉烯醇—内根-贝壳杉烯醛—内根-贝壳杉烯酸—内根-7a-羟基贝壳杉烯酸—GA12-7-醛—…..赤霉素
—牛儿牛儿焦磷酸(GGPP)—古巴焦磷酸(CPP)——内根-贝壳杉烯—内根-贝壳杉烯醇—内根-贝壳杉烯醛—内根-贝壳杉烯酸—内根-7a-羟基贝壳杉烯酸—GA12-7-醛—…..赤霉素.")
215
四、赤霉素的作用机理 (一)促进生长 1.赤霉素显著地促进植物生长,包括细胞分裂和细胞扩大两个方面。 2.GA显著促进燕麦节间初段伸长的同时,细胞壁可塑性也增加。 3.赤霉素促进细胞生长和生长素促进细胞生长的作用机理不完全相同,表现在: (1)生长素促进细胞延长与渗透吸水有关,而赤霉素对高直下胚轴的延长与细胞液渗透压浓度无关,但增加细胞壁的伸展性;
生长素促进细胞延长与渗透吸水有关,而赤霉素对高直下胚轴的延长与细胞液渗透压浓度无关,但增加细胞壁的伸展性;")
216
(2)生长素引起细胞壁酸化而疏松,而赤霉素不引起细胞壁酸化;
(3)生长素对细胞廷长的影响有较短的停滞期,而赤霉素的停滞期长达45min。 4.赤霉素促进细胞延长的原因 (1)有人用赤霉素消除细胞壁中 Ca的作用来解释赤霉素促进细胞延长的原因。 (2)有人认为赤霉素阻止细胞壁的硬化过程。赤霉素能抑制细胞壁过氧化物酶的活性,所以细胞壁不硬化,有延展性,细胞就延长。在诱发细胞延长的同时,赤霉素也加强细胞壁聚合物的生物合成。 (二)促进RNA和蛋白质合成 赤霉素对a-淀粉酶合成的影响是控制DNA转录为mRNA, 能一定程度地增强翻译水平,产生a-淀粉酶。
生长素对细胞廷长的影响有较短的停滞期,而赤霉素的停滞期长达45min。 4.赤霉素促进细胞延长的原因. (1)有人用赤霉素消除细胞壁中 Ca的作用来解释赤霉素促进细胞延长的原因。 (2)有人认为赤霉素阻止细胞壁的硬化过程。赤霉素能抑制细胞壁过氧化物酶的活性,所以细胞壁不硬化,有延展性,细胞就延长。在诱发细胞延长的同时,赤霉素也加强细胞壁聚合物的生物合成。 (二)促进RNA和蛋白质合成. 赤霉素对a-淀粉酶合成的影响是控制DNA转录为mRNA, 能一定程度地增强翻译水平,产生a-淀粉酶。")
217
五、赤霉素的生理作用及应用 1.生理作用 (1)促进:两性花的雄花形成,单性结实,某些植物开花,细胞分裂,叶片扩大,抽首,茎延长,侧枝生长,胚轴弯钩变直,种子发芽,果实生长,某些植物座果。 (2)抑制:成熟,侧芽休眠,衰老,块茎形成。
促进:两性花的雄花形成,单性结实,某些植物开花,细胞分裂,叶片扩大,抽首,茎延长,侧枝生长,胚轴弯钩变直,种子发芽,果实生长,某些植物座果。 (2)抑制:成熟,侧芽休眠,衰老,块茎形成。 .")
218
2.生产上的应用: (1)促进麦芽糖化 赤霉素诱发a-淀粉酶的形成用于啤酒生产。 (2)促进营养生长 赤霉素对根的伸长无促进作用,但显著促进茎叶生长。 (3)防止脱落 可阻止花柄和果辆离层的形成,防止花果脱落,提高座果率。 (4)打破休眠
打破休眠.")
219
第三节细胞分裂素类 1.细胞分裂素类是一类促进细胞分裂的植物激素。此类物质中最早被发现的是激动素。 2.发现过程: 1955年F.Skoog等培养烟草髓部组织时,偶然在培养基中加入放置很久的鲜鱼精子DNA,髓部细胞分裂就加快;如加入新鲜的DNA,则完全无效;可是当新鲜的DNA与培养基一起高压灭菌后,又能促进细胞分裂。 从DNA降解物中分离出一种物质—6-呋喃氨基嘌呤,它能促进细胞分裂,被命名为激动素。 3.把具有和激动素相同生理活性的天然的和人工合成的化合物,都称为细胞分裂素(cytokinin)。
。")
220
一、细胞分裂素的种类及化学结构 细胞分裂素是腺嘌呤的衍生物。当6位氨基、2位碳原子和9位氮原子上的氢原子被取代时,则形成各种不同的细胞分裂素。

221
(一)天然的细胞分裂素 天然存在的细胞分裂素又可分为游离的细胞分裂素和在tRNA中的细胞分裂素。 1.游离的细胞分裂素 植物和微生物中都含有游离的细胞分裂素,共20多种。其中玉米素的生理活性比激动素强得多。玉米素核苷。二氢玉米素、异戊烯基腺苷。 2.在tRNA中的细胞分裂素 细胞分裂素本身就是tRNA的组成部分。植物tRNA中的细胞分裂素主要有:异戊烯基腺苷、反式玉米素核苷、甲硫基异戊烯基腺苷、甲硫基玉米素核苷。

222
(二)人工合成的细胞分裂素 根据激动素的结构,人们合成了大量的衍生物,它们都具促进细胞分裂能力。 常用的有激动素、6-苄基腺嘌呤(简称6-BA)和四氢吡喃苄基腺嘌呤(又称多氯苯甲酸,简称PBA)。 二苯脲的结构则特殊,它不具腺嘌呤的结构,但却有细胞分裂素的生理功能。 此外,一些人工会成的化合物能堵塞细胞分裂素的作用(可能与细胞分裂素竞争受体),起着细胞分裂素拮抗剂作用。 如果多施细胞分裂素,则可克服这种抑制效应。最有效的细胞分裂素拮抗剂是3-甲基-7-(3-甲基丁氨基)吡唑啉[4,3-右旋]嘧啶。
,起着细胞分裂素拮抗剂作用。 如果多施细胞分裂素,则可克服这种抑制效应。最有效的细胞分裂素拮抗剂是3-甲基-7-(3-甲基丁氨基)吡唑啉[4,3-右旋]嘧啶。")
223
二、细胞分裂素的生物合成及运输 1.细胞分裂素分布于细菌、真菌、藻类和高等植物中,高等植物的细胞分裂素主要存在于进行细胞分裂的部位,如茎尖、根尖、未成熟的种子、萌发的种子和生长着的果实等。 2.一般来说,细胞分裂素含量为1~1000ng/g干重。从高等植物中发现的细胞分裂素,大多数是玉米素或玉米素核苷。 3.一般认为,细胞分裂素在根尖合成,经木质部运到地上部分。细胞分裂素生物合成是在细胞里的微粒体中进行的。

224
4.植物内游离细胞分裂素的来源,有一部分是tRNA降解所致;但有一部分是不依赖tRNA的细胞分裂素。
5.细胞分裂素的前体是甲瓦龙酸,经过异戊烯基焦磷酸(iPP)和AMP,在细胞分裂素合成酶催化下,形成异戊烯基腺苷-5’-磷酸盐。这是游离细胞分裂素生物合成的关键步骤。 6.细胞分裂素在植物体中运输是无极性的。 7.细胞分裂素氧化酶可能对细胞分裂素起钝化作用,防止细胞分裂素积累过多,产生毒害。
和AMP,在细胞分裂素合成酶催化下,形成异戊烯基腺苷-5’-磷酸盐。这是游离细胞分裂素生物合成的关键步骤。 6.细胞分裂素在植物体中运输是无极性的。 7.细胞分裂素氧化酶可能对细胞分裂素起钝化作用,防止细胞分裂素积累过多,产生毒害。")
225
三、细胞分裂素的作用机理 (一)细胞分裂素的结合位点 专一性、高亲和性的CTK结合蛋白 (二)细胞分裂素对转录和翻译的控制 1.细胞分裂素促进转录作用。 激动素能与豌豆芽染色质结合,调节基因活性,促进 RNA合成。 2.促进翻译 细胞分裂素可以促进蛋白质的生物合成。因为细胞分裂素存在于核糖体上促进核糖体与mRNA结合,形成多核糖体,加速翻译速度,形成新的蛋白质。

226
四、细胞分裂素的生理作用和应用 (一)细胞分裂素的生理作用如下: 1.促进:细胞分裂,地上部分化,侧芽生长,叶片扩大,气孔张开,偏上性,伤口愈合,种子发芽,形成层活动,根瘤形成,果实生长,某些植物座果。 抑制:不定根形成,侧根形成,叶片衰老(延缓)。 (二)几种生理作用与应用 1.促进细胞分裂和扩大 细胞分裂素的生理作用也是多方面的,它主要的作用是促进细胞分裂; 细胞分裂素也可以使细胞体积加大,但和生长素不同的是它的作用主要是使细胞扩大,而不是伸长。
几种生理作用与应用. 1.促进细胞分裂和扩大. 细胞分裂素的生理作用也是多方面的,它主要的作用是促进细胞分裂; 细胞分裂素也可以使细胞体积加大,但和生长素不同的是它的作用主要是使细胞扩大,而不是伸长。")
227
2.诱导芽的分化 愈伤组织产生根或产生芽,取决于生长素和激动素依次的比值。当激动素/生长素的比值低时,诱导根的分化;两者比值处于中间水平时,愈伤组织只生长而不分化;两者比值较高时,则诱导芽的形成。 3.延缓叶对衰老 延缓叶片衰老是细胞分裂素特有的作用。 细胞分裂素可以显著延长保绿时间,推迟离休叶片衰老。原因有两个: (1)细胞分裂素抑制核糖核酸酶、脱氧核糖核酸酶和蛋白酶等的活性,延缓了核酸、蛋白质和叶绿素等的降解。 ( 2)细胞分裂素促使营养物质向其应用部位移动。 激动素诱发定向运输
细胞分裂素抑制核糖核酸酶、脱氧核糖核酸酶和蛋白酶等的活性,延缓了核酸、蛋白质和叶绿素等的降解。 ( 2)细胞分裂素促使营养物质向其应用部位移动。 激动素诱发定向运输.")
228
第四节脱落酸 植物在它的生活周期中,如果生活条件不适宜,部分器官(如果实、叶片等)就会脱落:或者到了生长季节终了,叶子就会脱落,生长停止,进人休眠。在这些过程中,植物体内就会产生一类抑制生长发育的植物激素,即脱落酸(abscisic acid,简称ABA)。 一、脱落酸的化学结构 脱落酸是一种类倍半萜化合物。由于分子中有一个不对称碳原子,所以有两个旋光异构体。天然的脱落酸是右旋的(+)ABA表示;它的对映作左旋,以(-)ABA表示,两者都有生物活性,但后者不能促进气孔关闭。
ABA表示;它的对映作左旋,以(-)ABA表示,两者都有生物活性,但后者不能促进气孔关闭。")
229
二、脱落酸的分布和运输 (一)分布 1.存在植物类群: 存在于全部维营植物中,包括被子植物、裸子植物和蕨类植物。苔类和藻类植物中含有半月苔酸。细菌和真菌无脱落酸,藓类有无脱落酸尚未确定。 2.分布的组织器官: 高等植物各器官和组织中都有脱落酸,其中以将要脱落或进入休眠的器官和组织中较多,在逆境条件下含量会迅速增多。

230
(二)运输 1.脱落酸运输不存在极性。 2.运输形式:主要以游离型的形式运输,也有部分以脱落酸糖着形式运输。 3.运输速度:在植物体的运输速度很快,在茎或叶柄中的运输速率大约是20mm/h。
运输 1.脱落酸运输不存在极性。 2.运输形式:主要以游离型的形式运输,也有部分以脱落酸糖着形式运输。 3.运输速度:在植物体的运输速度很快,在茎或叶柄中的运输速率大约是20mm/h。")
231
MVA——类胡萝卜素——黄质醛——ABA
三、脱落酸的生物合成和代谢 脱落酸在根冠和萎蔫的叶片中合成得较多,茎和种子等各处都有合成脱落酸的能力。 脱落酸以离子化状态大量积累在叶绿体中。 (一)脱落酸的生物合成 脱落酸合成途径有两条: 1.类萜途径(terpenoid pathway)脱落酸的合成是由甲瓦龙酸(MVA)经过法尼焦磷酸(FPP),再经过一些未明的过程而形成脱落酸。 2.类胡萝卜途径(carotenoid Pathway) MVA——类胡萝卜素——黄质醛——ABA
脱落酸的生物合成. 脱落酸合成途径有两条: 1.类萜途径(terpenoid pathway)脱落酸的合成是由甲瓦龙酸(MVA)经过法尼焦磷酸(FPP),再经过一些未明的过程而形成脱落酸。 2.类胡萝卜途径(carotenoid Pathway) MVA——类胡萝卜素——黄质醛——ABA.")
232
(二)脱落酸的代谢 1.脱落酸会氧化降解为活性很低的中间产物(红花菜豆酸,简称PA); 2.同时也会形成没有活性的脱落酸结合物。 3.脱落酸的合成和代谢总结
脱落酸的代谢 1.脱落酸会氧化降解为活性很低的中间产物(红花菜豆酸,简称PA); 2.同时也会形成没有活性的脱落酸结合物。 3.脱落酸的合成和代谢总结")
233
四、脱落酸的作用机理 (一)脱落酸的结合位点 两种不同的观点: 1.以蚕豆叶片保卫细胞原生质体为材料证明,脱落酸专一地与质膜受体结合。 2.用棉花子房细胞的质膜和核做材料,发现脱落酸只与细胞核专一结合,而与质膜结合不专一。 (二)脱落酸抑制核酸和蛋白质合成 1.脱落酸只在转录水平上起作用,而不在翻译水平上起作用。 2.然而,许多试验也证明脱落酸抑制蛋白质的合成。
脱落酸抑制核酸和蛋白质合成. 1.脱落酸只在转录水平上起作用,而不在翻译水平上起作用。 2.然而,许多试验也证明脱落酸抑制蛋白质的合成。")
234
五、脱落酸的生理作用和应用 (一)生理作用
1.促进:叶、花、果脱落,气孔关闭,侧芽、块茎休眠,叶片衰老,光合产物运向发育着的种子,果实产生乙烯,果实成熟。 2.抑制:种子发芽,IAA运输,植株生长。 (二)几种主要生理作用的应用 1.促进脱落 2.促进休眠 3.提高抗逆性 脱落酸在逆境条件下迅速形成,使植物的生理发生变化,以适应环境,所以脱落酸又称为“应激激素”。
几种主要生理作用的应用. 1.促进脱落. 2.促进休眠. 3.提高抗逆性. 脱落酸在逆境条件下迅速形成,使植物的生理发生变化,以适应环境,所以脱落酸又称为 应激激素 。")
235
第五节 乙烯 1901年,俄国植物生理学家Neljubow报道,照明气中的乙烯会引起黑暗中生长的豌豆幼苗,产生“三重反应”,他认为乙烯是生长调节剂。 英国Gane(1934)首先证明乙烯是植物的天然产物。 美国Crocker等认为乙烯是一种果实催熟激素,同时也有调节营养器官的作用。 Burg(1965)提出,乙烯是一种植物激素,以后得到公认。
提出,乙烯是一种植物激素,以后得到公认。")
236
一、乙烯的分布与生物合成 (一)乙烯的分布 高等植物各器官都能产生乙烯,但不同组织、器官和发育时期,乙烯的释放量是不同的。 成熟组织释放乙烯较少;分生组织、种子萌发、花刚凋谢和果实成熟时产生乙烯最多。 (二)乙烯的生物合成 1.前体:蛋氨酸 2.合成过程
乙烯的生物合成. 1.前体:蛋氨酸. 2.合成过程.")
237
(三)乙烯生物合成的酶调节 1.ACC合成酶 ACC合成酶催化SAM转变为ACC。 2.乙烯形成酶 ACC转变为乙烯是由一种氧化酶(或酶系统)所催化的,一般称它为乙烯形成酶(EFE)与其底物ACC有很高的亲和力。 3.ACC丙二酰基转移酶 ACC丙二酸基转移酶的作用就是促使ACC起丙二酸化反应,形成 MACC。这种酶活性强时,形成MACC多,ACC就少,乙烯释放量就少;否则乙烯增多。

238
(四)乙烯的代谢 乙烯在植物体内形成以后也会转变为二氧化碳和乙烯氧化物等气体代谢物,也会形成可溶性代谢物,如乙烯乙二醇(ethylene glycol)和乙烯葡萄糖结合体等。 乙烯代谢的功能是除去乙烯或使乙烯钝化,使乙烯水平达到适合植物生长发育需要。
乙烯的代谢 乙烯在植物体内形成以后也会转变为二氧化碳和乙烯氧化物等气体代谢物,也会形成可溶性代谢物,如乙烯乙二醇(ethylene glycol)和乙烯葡萄糖结合体等。 乙烯代谢的功能是除去乙烯或使乙烯钝化,使乙烯水平达到适合植物生长发育需要。")
239
二、乙烯的作用机理 乙烯形成以后,还需要与金属(可能是一价铜)蛋白质结合,进一步通过代谢后才起生理作用。 银离子抑制乙烯作用,其原因可能是银离子取代金属蛋白质中的金属,而使金属蛋白质无法与乙烯结合。 EDTA是一种与金属结合的螫合物,所以Fe-EDTA也抑制乙烯的作用; 二氧化碳也抑制乙烯的作用,因为二氧化碳与乙烯竞争一个作用部位。 一般来说,与乙烯结合的膜蛋白质对热敏感,乙烯可促进核酸和蛋白质的合成。

240
三、乙烯的生理作用和应用 (一)生理作用 1.促进:解除休眠,地上部和根的生长和分化,不定根形成,叶片和果实脱落,某些植物的花诱导形成,两性花中雌花形成,开花,花和果实衰老,果实成熟,茎增粗,萎蔫。 2.抑制:某些植物开花,生长素的转运,茎和根的伸长生长。 乙烯是气体,在合成部位起作用,不被转运,但是乙烯的前体ACC在植物体内是能被运输的。所以有人认为ACC才是激素,因为它符合植物激素的定义。不过,这仅是一些人的观点。

241
(二)乙烯的生理功能的主要表现 1.促进细胞扩大 黄化豌豆幼苗对乙烯的生长反应是“三重反应”。即矮化、加粗、偏上生长。 2.促进果实成熟 乙烯促进果实成熟的原因可能是:增强质膜的透性,加速呼吸,引起果肉有机物的强烈转化。 3.促进器官脱落 在叶片脱落过程中,乙烯能促进离层中纤维素酶的合成,并促使该酶由原生质体释放到细胞壁中,引起细胞壁分解,同时也刺激寓居区近侧细胞膨胀,叶柄便分离开。

242
(三)乙烯利在农业生产上的应用 1.果实催熟和改善品质 2.促进次生物质排出 乙烯也能增加漆树、松树和印度紫檀等重要木本经济植物的次生物质的产量。 3.促进开花 用乙烯利对菠萝灌心催花,开花提早,花期一致。 4.化学杀雄

243
第六节 油菜素内酯 1.发现:美国的Mitchell研究小组在70年代初期研究花粉激素,发现油菜等花粉中有一种提取物,对菜豆幼苗生长具极强烈的促进作用。 2.提取:Grove等(1979年)用227kg油菜花粉,得到10mg的高活性结晶,它是固醇内酯化合物,命名为油菜素内酯(brassinolide)。 3.特点:油菜素内酯具有许多与上述五大激素不同的独特生理效应,大部分植物激素的含量和用量以mm/L为单位,而油菜素内酯则以ug/L为单位,所以油菜素内酯应是一类新型的激素。这种提法目前尚未得到公认。 4.油菜素内酯是一种固体物质,结构与动物激素非常相似。说明动、植物是同源的。
用227kg油菜花粉,得到10mg的高活性结晶,它是固醇内酯化合物,命名为油菜素内酯(brassinolide)。 3.特点:油菜素内酯具有许多与上述五大激素不同的独特生理效应,大部分植物激素的含量和用量以mm/L为单位,而油菜素内酯则以ug/L为单位,所以油菜素内酯应是一类新型的激素。这种提法目前尚未得到公认。 4.油菜素内酯是一种固体物质,结构与动物激素非常相似。说明动、植物是同源的。")
244
一、油菜素内酯的种类及生物合成 1.定义:高等植物中普遍存在着一大类以甾类化合物为骨架的具生理活性的天然甾体类,总称为油菜素内酯(brassinosteroid)。 2.油菜素内酯的含量和生理活性最高,一般以油菜素内酯代表油菜素甾体类。 3.分布:裸子植物和被子植物中存在油菜素甾体类,藻类也有。在高等植物中枝、叶、花各器官都有,尤其是花粉最多。 4.人工合成:表油菜素内酯等,且油菜素内酯已可人工合成。

245
二、油菜素内酯的生理作用 1.促进细胞伸长和分裂 原因在于油菜素内酯可使绿豆和斑豆的DNA聚合酶和RNA聚合酶的活性增大, DNA和RNA含量增多,蛋白质合成也多。油菜素内酯又会刺激质膜上的ATP酶活性,促使质膜分泌H+到细胞壁。所以油菜素内酯可能同样遵循“酸生长”学说,促使细胞伸长。 2.促进光合作用 提高RuBPC活性、叶绿素含量。 3.抵抗低温伤害 油莱索内酯提高抗寒的原因,可能是处理后恢复内源生长素水平。

246
三、油菜素内酯的应用 1.提高产量 在开花期施用油菜素内酯,可提高玉米、小麦、番茄、黄瓜和茄子的产量。 2.提高抗逆性 油莱素内酯具有明显的抗冷性、抗旱性和抗盐性,在自然条件不良时更是突出,所以油菜素内酯又称逆境条件的缓和剂。 3.抗病 油菜素内酯可提高作物对病害抵抗能力,但对稻瘟病和黄瓜白粉病无效。

247
第七节 生长抑制物质 1.生长抑制剂(growth inhibitor) (1)特点:它们抑制顶端分生组织生长,丧失顶端优势,使植株形态发生很大的变化,外施赤霉素不能逆转这种抑制效应。生长抑制剂也抑制种子和芽的萌发。 (2)代表:天然生长抑制剂有ABA、肉桂酸、香豆素、水杨酸、绿原酸、咖啡酸和茉莉酸等。人工合成的生长抑制剂有三腆苯甲酸、整形素等。
代表:天然生长抑制剂有ABA、肉桂酸、香豆素、水杨酸、绿原酸、咖啡酸和茉莉酸等。人工合成的生长抑制剂有三腆苯甲酸、整形素等。")
248
2.生长延缓剂(growth retardant)
(1)特点:它们抑制茎部近顶端分生组织的细胞延长,节间缩短,叶数和节数不变,株型紧凑,矮小,生殖器官不受影响或影响不大。全部为人工合成,外施赤霉素可以逆转其抑制效应。 (2)代表:CCC、B9、PP333等,它们都能抑制赤霉素的生物合成,所以是抗赤霉素。
特点:它们抑制茎部近顶端分生组织的细胞延长,节间缩短,叶数和节数不变,株型紧凑,矮小,生殖器官不受影响或影响不大。全部为人工合成,外施赤霉素可以逆转其抑制效应。 (2)代表:CCC、B9、PP333等,它们都能抑制赤霉素的生物合成,所以是抗赤霉素。")
249
一、生长抑制剂 (一)茉莉酸 1.茉莉酸(简称JA)是从茉莉属等植物的茎、叶中分离出来的,故名茉莉酸。
2.分布:被子植物和裸子植物里有茉莉酸或茉莉酸甲酯。现已人工合成。 3.生理作用:茉莉酸甲酯抑制水稻、花生幼苗生长,诱导气孔关闭,促进燕麦和豇豆的叶片衰老和脱落,提高水稻幼苗的抗寒性和花生幼苗的抗旱性,其生理功能类似ABA。 (二)三碘苯甲酸 三碘苯甲酸(简称TIBA)是一种阻碍生长素运输的物质,它能抑制顶端分生组织细胞分裂,使植株矮化,消除顶端优势,使分枝增加。 TIBA多用于大豆。
三碘苯甲酸. 三碘苯甲酸(简称TIBA)是一种阻碍生长素运输的物质,它能抑制顶端分生组织细胞分裂,使植株矮化,消除顶端优势,使分枝增加。 TIBA多用于大豆。")
250
(三)马来酰肼 马来酰肼(简称MH)在我国又称青鲜素,是第一种人工合成的生长抑制剂。 其作用正好同生长素相反,因为它的结构与RNA的组成部分尿嘧啶非常相似,MH进入植物体内可代替尿嘧啶的位置,但不能起代谢中的作用,因而阻止正常代谢的进行,从而抑制了生长。 MH是一种危险的化合物,可能致癌和使动物染色体畸变。 (四)整形素 整形素常用于木本植物。它能抑制顶端分生组织细胞分裂及伸长,抑制植株茎伸长,腋芽滋生,使植株发育成矮小灌木的形状。整形素还能刺激花及果实脱落。
整形素. 整形素常用于木本植物。它能抑制顶端分生组织细胞分裂及伸长,抑制植株茎伸长,腋芽滋生,使植株发育成矮小灌木的形状。整形素还能刺激花及果实脱落。")
251
二、生长延缓剂 (一) CCC CCC我国俗称矮壮素,是常用的一种生长延缓剂。能抑制赤霉素生物合成过程中的GGPP环化为CPP的过程,也可以抑制 CPP转变为贝壳杉烯过程,是一种抗赤霉素剂。喷施矮壮素可使节间缩短,植株变矮,茎粗,叶色加深。有利于改善透光条件和光合作用,并抗倒伏。 (二) Pix Pix国内俗称缩节安、助壮素等。与CCC相似,在棉株始花期施用,能有效地控制棉铃营养生长,节间短,叶面积小,增加叶绿素含量,提高光合速率,可使光合产物较多运到幼铃,减少蕾铃脱落,提高棉花产量。
 Pix. Pix国内俗称缩节安、助壮素等。与CCC相似,在棉株始花期施用,能有效地控制棉铃营养生长,节间短,叶面积小,增加叶绿素含量,提高光合速率,可使光合产物较多运到幼铃,减少蕾铃脱落,提高棉花产量。")
252
(三)B9 B9,又名Afar,SADH。其作用机理是抑制贝壳杉烯醛的合成,所以也是抑制赤霉素的生物合成。可抑制果树顶端分生组织的细胞分裂,使枝条生长缓慢,可代替人工整枝,增加次年开花座果的数量。B9有防止采前落果及促进果实着色的作用。溶液喷施叶面,抑制茎枝生长,减少地上部分营养生长的物质消耗,使光合产物较多地运到地下部,尤其是结实器官,提高产量。 (四)PP333 PP333(氯丁唑,我国俗称多效唑)。这种延缓剂破坏微粒体中细胞色素P450催化作用,抑制贝壳杉烯、贝壳杉烯醇、贝壳杉烯醛的合成,所以赤霉素含量大减。PP333对营养生长的抑制能力比B9或CCC更大。
PP333. PP333(氯丁唑,我国俗称多效唑)。这种延缓剂破坏微粒体中细胞色素P450催化作用,抑制贝壳杉烯、贝壳杉烯醇、贝壳杉烯醛的合成,所以赤霉素含量大减。PP333对营养生长的抑制能力比B9或CCC更大。")
253
(五)其 他 在生长延缓剂中还有一些药剂是常见的。 例如,福斯方-D;又如阿莫-1618。这两种延缓剂抑制CPP的合成,所以赤霉素含量低。它们可使茎杆矮化,外色浓绿。 醇草定(又名嘧啶醇)抑制贝壳杉烯至贝壳杉烯酸的氧化,影响赤霉素的合成,故此施用醇草定可抑制植株生长。
抑制贝壳杉烯至贝壳杉烯酸的氧化,影响赤霉素的合成,故此施用醇草定可抑制植株生长。")
254
第八节多胺和钙调素 多胺和钙调素不是植物生长物质,但它们与植物生长发育的关系密切。 一、多胺 1.分布:不同器官的多胺含量是不同的,一般来说,细胞分裂旺盛的地方,多胺含量较多。 2.生物合成:多胺是由三种氨基酸转变来的:(1)精氨酸和赖氨酸构成多胺氨基部分的碳架;(2)蛋氨酸向腐胺提供丙氨基而逐步形成亚精胺和精胺;(3)赖氨酸脱羧形成尸胺。 3.运输形式:多胺本身是不能在植物体内运输,而它们的前身—氨基酸可以运输到生长活跃部位转变为多胺。
精氨酸和赖氨酸构成多胺氨基部分的碳架;(2)蛋氨酸向腐胺提供丙氨基而逐步形成亚精胺和精胺;(3)赖氨酸脱羧形成尸胺。 3.运输形式:多胺本身是不能在植物体内运输,而它们的前身—氨基酸可以运输到生长活跃部位转变为多胺。")
255
多胺的生理功能 1.促进生长 加快DNA的转录,RNA聚合酶活性和氨基酸掺入蛋白质速率加快,通过影响核酸代谢,促进蛋白质的合成。 2.调节与光敏色素有关的生长和形态建成 光照影响光敏色素,接着影响精氨酸脱羧酶活性,多胺生物合成,最后影响生长和形态建成。

256
3.延迟衰老 (1)多胺能保存叶绿体类囊体膜的完整性,减慢蛋白质丧失和RN-case活性,阻止叶绿素破坏,衰老较慢。 (2)亚精胺和精胺与乙烯的前体都是S-腺苷蛋氨酸,多胺和乙烯就会竞争S-腺苷蛋氨酸,所以多胺可抑制乙烯的生成,也会延缓衰老。 4.适应逆境条件 植物在缺钾和缺镁时,精氨酸脱羧酶活性提高几倍到几十倍,积累腐胺,以代替钾等主要无机阳离子,影响细胞pH。 此外,外施IAA、GA、CTK等植物激素可促进多胺生物合成。
多胺能保存叶绿体类囊体膜的完整性,减慢蛋白质丧失和RN-case活性,阻止叶绿素破坏,衰老较慢。 (2)亚精胺和精胺与乙烯的前体都是S-腺苷蛋氨酸,多胺和乙烯就会竞争S-腺苷蛋氨酸,所以多胺可抑制乙烯的生成,也会延缓衰老。 4.适应逆境条件 植物在缺钾和缺镁时,精氨酸脱羧酶活性提高几倍到几十倍,积累腐胺,以代替钾等主要无机阳离子,影响细胞pH。 此外,外施IAA、GA、CTK等植物激素可促进多胺生物合成。")
257
二、钙调素 1.钙离子的受体蛋白叫钙调素(calmodulin,简称CaM) 2.CaM位于植物细胞的细胞溶质和细胞器(质体、线粒体和细胞核)中,是一种耐热的球状蛋白。CaM和Ca。十有很高的亲和力。一个CaM分子可与四个Ca2+结合。CaM本身没有活性,但和Ca2+结合后,就引起构象变化,使原来埋藏在CaM分子内部的流水区域翻转出来,暴露于表面,CaM就是通过这个疏水区域与酶结合,形成酶一ca2+-CaM复合体,出现活性。 3.活性的酶- Ca2+-CaM复合体参与 Ca2+·CaM为调节细胞功能的第二信使。
中,是一种耐热的球状蛋白。CaM和Ca。十有很高的亲和力。一个CaM分子可与四个Ca2+结合。CaM本身没有活性,但和Ca2+结合后,就引起构象变化,使原来埋藏在CaM分子内部的流水区域翻转出来,暴露于表面,CaM就是通过这个疏水区域与酶结合,形成酶一ca2+-CaM复合体,出现活性。 3.活性的酶- Ca2+-CaM复合体参与 Ca2+·CaM为调节细胞功能的第二信使。")
258
第七章光形态建成 1. 光对植物的影响主要有两个方面: (1)光是绿色植物光合作用所必需的; (2)光调节植物整个生长和发育。
2.光形态建成(photomorphogenesis):依赖光控制植物生长、发育和分化的过程。 3.光合作用是高能反应,它将光能转变为化学能;而光形态建成是低能反应,光只作为一个信号去激发光受体,推动细胞内一系列反应,最终表现为形态结构的变化,光形态建成所需红闪光的能量和一般光合作用光补偿点总能量相差10个数量级,甚为微弱。
:依赖光控制植物生长、发育和分化的过程。 3.光合作用是高能反应,它将光能转变为化学能;而光形态建成是低能反应,光只作为一个信号去激发光受体,推动细胞内一系列反应,最终表现为形态结构的变化,光形态建成所需红闪光的能量和一般光合作用光补偿点总能量相差10个数量级,甚为微弱。")
259
第一节光敏色素的发现和分布 一、光敏色素的发现 二、光敏色素的分布 光敏色素分布在植物的各个器官中。
一般来说,蛋白质丰富的分生组织中含有较多的光敏色素。 一般认为,光敏色素与膜系统结合,分布在质膜、线粒体膜、核膜。叶绿体膜和内质网膜上。

260
第二节光敏色素的化学性质及光化学转换 一、光敏色素的化学性质 光敏色素是一种易溶于水的色素蛋白质。
有两个组成部分:生色团和蛋白质。光敏色素生色团是一长链状的 4个吡咯环,与胆色素的胆绿素结构相似。 生色团有两种类型:红光吸收型(Pr)和远红光吸收型(Pfr),两者的光学特性不同。 光敏色素的蛋白质单体分子量是124kD,Pfr型才有生理活性,那些蛋白质降解后的大光敏色素(分子量为114~118kD)和小光敏色素(分子量为60kD)是没有生理活性的。 Pr型色素的生物合成是在黑暗条件下进行的,其合成过程可能类似脱植基叶绿素的合成过程,因为两者都具四吡咯环。
和远红光吸收型(Pfr),两者的光学特性不同。 光敏色素的蛋白质单体分子量是124kD,Pfr型才有生理活性,那些蛋白质降解后的大光敏色素(分子量为114~118kD)和小光敏色素(分子量为60kD)是没有生理活性的。 Pr型色素的生物合成是在黑暗条件下进行的,其合成过程可能类似脱植基叶绿素的合成过程,因为两者都具四吡咯环。")
261
二、光敏色素系统的光化学转换 1.总光敏色素:
2.光稳定平衡(photostationary equilibrium):在一定波长下具有生理活性的Pfr型浓度与总光敏色素浓度的比例。 3.光敏色素的两种反应: (1)慢反应 (2)快反应 4.光敏色素的光转变过程:
:在一定波长下具有生理活性的Pfr型浓度与总光敏色素浓度的比例。 3.光敏色素的两种反应: (1)慢反应. (2)快反应. 4.光敏色素的光转变过程:")
262
第三节光敏色素的生理作用 一、光敏色素和酶 许多酶的光调节是通过光敏色素为媒介的。 现在已知有多种酶是受光敏色素控制的。 二、光敏色素与植物激素 植物的光形态建成是由体外和体内的多因子决定的。激素可能是最重要的体内调节因子。 植物激素在光形态建成中的作用,有时是中间调节物质,有时与光协同起调节作用。

263
第四节 光敏色素的作用机理 两种假说;膜假说和基因调节假说。 一、膜假说
第四节 光敏色素的作用机理 两种假说;膜假说和基因调节假说。 一、膜假说 1.内容:光敏色素位子膜系统上,当发生光转换时,跨膜的离子流动和膜上酶的分布都会改变,影响代谢,经过一系列的生理生化变化,最终表现出形态建成的改变。 2.证据:如含羞草、合欢叶片运动;转板藻的叶绿体运动等。 3.棚田效应(Tanada effect) :离体绿豆根尖在红光下诱导膜产生少量正电荷,所以能粘附在带负电行的玻璃表面,而远红光照射则逆转这种粘附现象,这就是棚田效应。
 :离体绿豆根尖在红光下诱导膜产生少量正电荷,所以能粘附在带负电行的玻璃表面,而远红光照射则逆转这种粘附现象,这就是棚田效应。")
264
二、基因调节假说 1.内容:在接受红光后,Pfr型经过一系列过程,将信号转移到基因,活化或抑制某些特定基因,使转录出单股mRNA的速度发生改变,mRNA翻译成特殊蛋白质(酶),最后表现出形态建成。 2.这个假说能解释前述光敏色素的慢反应,需要较长的停滞期,形成各种蛋白质和酶。 3.光活化膜上光敏色素经过Ca和 CaM调节蛋白质磷酸化的模式。

265
第五节蓝光反应 1.植物界存在的第二类光形态建成是受蓝光调节的反应。 2.藻类、真菌、旅类和种子植物都有蓝光效应。
蓝光可使褐藻中的萱藻平面地生长,而红光则诱导丝状生长,在蓝光下正常发育,形成心形原叶体,而红光下则纵向生长成绿色的丝状体。真菌的光形态建成反应主要受蓝光和紫外光诱导。 3.隐花色素的作用光谱的特征是:在近紫外光350~380nm波段有一尖峰,在蓝光部分有三个峰组成的宽阔的峰,其最大值在440~460nm,在480nm有一“陡肩”,420nm有一可分辨的“小肩”。

266
第八章 植物的生长生理 第一节种子的萌发 一、影响种子萌发的外界条件 (一)水分 1.种皮软化, 2.氧易透过,使胚易突破种皮;
3.使代谢加强; 4.促进物质运输。 (二)氧气 (三)温度 (四)光 需光种子(light seed)、需暗种子(dark seed)。
氧气. (三)温度. (四)光. 需光种子(light seed)、需暗种子(dark seed)。")
267
二、种子萌发的生理生化变化

268
(一)种子的吸水 种子的吸水可分为 3个阶段,即急剧 的吸水、吸水的停 止和胚根长出后的 重新迅速吸水。
种子的吸水 种子的吸水可分为 3个阶段,即急剧 的吸水、吸水的停 止和胚根长出后的 重新迅速吸水。")
269
(二)呼吸作用的变化和酶的形成 1.初期的呼吸主要是无氧呼吸,而随后是有氧呼吸。
2.在吸水的第二阶段,种子中各种酶亦在形成着。萌发种子酶的形成有两种来源: (1)从已存在的束缚态酶释放或活化而来; (2)通过核酸诱导下合成的蛋白质形成新的酶。
从已存在的束缚态酶释放或活化而来; (2)通过核酸诱导下合成的蛋白质形成新的酶。")
270
(三)有机物的转变 淀粉种子、油料种子和豆类种子 1.淀粉的转变
被淀粉酶水解为麦芽糖,再由麦芽糖酶分解为葡萄糖,供细胞代谢所利用,或转化为蔗糖。 2.脂肪的转变 脂肪酶的作用下水解为甘油和脂肪酸。甘油在酶的催化下变成磷酸甘油,再转变成磷酸二羟丙酮参加糖酵解反应,或转变为葡萄糖等。脂肪酸氧化分解为乙酰辅酶A,通过乙酸酸循环变为蔗糖。 3.蛋白质的转变 种子萌发经历从异养到自养的过程。

271
三、种子的寿命 (1)利用组织还原力 活的种子一定有呼吸作用,而呼吸作用会使物质还原呈现特定的颜色,根据颜色反应,即可知种子的生活力。
1. 种子寿命(seed longevity)是指种子从采收至失去发芽力的时间。 2.影响种子寿命的因素:湿度、温度、生物污染 3.检查种子生活力的方法有三类: (1)利用组织还原力 活的种子一定有呼吸作用,而呼吸作用会使物质还原呈现特定的颜色,根据颜色反应,即可知种子的生活力。 (2)利用原生质的着色能力 活种子的细胞质不易着色,死种子的细胞质易着色。 (3)利用细胞中的萤光物质 蛋白质、核酸和核着酸等重要有机物,都有荧光性质。
是指种子从采收至失去发芽力的时间。 2.影响种子寿命的因素:湿度、温度、生物污染. 3.检查种子生活力的方法有三类: (1)利用组织还原力. 活的种子一定有呼吸作用,而呼吸作用会使物质还原呈现特定的颜色,根据颜色反应,即可知种子的生活力。 (2)利用原生质的着色能力. 活种子的细胞质不易着色,死种子的细胞质易着色。 (3)利用细胞中的萤光物质. 蛋白质、核酸和核着酸等重要有机物,都有荧光性质。")
272
第二节细胞的生长和分化(略讲) 植物的组织和器官,以至整体的生长,是以细胞的生长为基础的,即通过细胞分裂增加细胞数目和通过细胞伸长增加体积来实现的。 一、细胞分裂的生理 分期及各期的主要事件。
 植物的组织和器官,以至整体的生长,是以细胞的生长为基础的,即通过细胞分裂增加细胞数目和通过细胞伸长增加体积来实现的。 一、细胞分裂的生理 分期及各期的主要事件。")
273
二、细胞伸长的生理 细胞壁疏松加大的两种方式: ( 1)化学滑行(chemical creep); (2)水解。 赤霉素和生长素促进细胞伸长,脱落酸抑制细胞伸长,细胞分裂素和乙烯则促进细胞扩大。
化学滑行(chemical creep); (2)水解。 赤霉素和生长素促进细胞伸长,脱落酸抑制细胞伸长,细胞分裂素和乙烯则促进细胞扩大。")
274
三、细胞分化的生理 细胞分化是指形成不同形态和不同功能细胞的过程。 (一)细胞全能性
1. 细胞全能性(totipotency):是指植物体的每个细胞携带着一套完整的基因组,并具有发育成完整植株的潜在能力。 2.细胞全能性是细胞分化的理论基础,而细胞分化是细胞全能性的具体表现。
:是指植物体的每个细胞携带着一套完整的基因组,并具有发育成完整植株的潜在能力。 2.细胞全能性是细胞分化的理论基础,而细胞分化是细胞全能性的具体表现。")
275
(二)极性 1.极性(Polarity):指在器官、组织甚至细胞中在不同轴向上存在某种形态结构和生理生化上的梯度差异。 2.极性造成了细胞内物质(如代谢物、蛋白质、激素等)分布不均匀,建立起轴向,两极分化,因此细胞不均等分裂(不是指染色体,而是指细胞质的构造和物质)。 3.木质部和韧皮部的分化与糖浓度有关。低木高韧,中均有。 4.光对植物组织的分化也有影响。 5.植物激素在细胞分化中也起着重要的作用。

276
四、组织培养 1.组织培养(Tissue culture):指在无菌条件下,分离并在培养基中培养离体植物组织(器官或细胞)的技术。
2.组织培养的理论依据:植物细胞具有全能性。 3.分类: (1)根据培养对象,分为器官培养、组织培养、胚胎培养、细胞培养和原生质体培养等。 (2)根据培养过程,分为初代培养、继代培养(subculture)等。 (3)根据培养基物理状态,分为固体培养、液体培养。
根据培养对象,分为器官培养、组织培养、胚胎培养、细胞培养和原生质体培养等。 (2)根据培养过程,分为初代培养、继代培养(subculture)等。 (3)根据培养基物理状态,分为固体培养、液体培养。")
277
4.组织培养的优点:可以研究被培养部分在不受植物体其他部分干扰下的生长和分化的规律,并且可以用各种培养条件影响它们的生长和分化,以解决理论上和生产上的问题。 5.组织培养的特点是: (1)取材少,培养材料经济; (2)人为控制培养条件,不受自然条件影响; (3)生长周期短,繁殖率高; (4)管理方便,利于自动化控制。 6.常用消毒剂:次氯酸钙、过氧化氢、氯化汞 7.培养基的组成:无机营养物、碳源、维生素、生长调节物质和有机附加物等
取材少,培养材料经济; (2)人为控制培养条件,不受自然条件影响; (3)生长周期短,繁殖率高; (4)管理方便,利于自动化控制。 6.常用消毒剂:次氯酸钙、过氧化氢、氯化汞 7.培养基的组成:无机营养物、碳源、维生素、生长调节物质和有机附加物等")
278
8.组织培养所需的条件: (1)温度:一般是25~27℃;最好有昼夜温差。 (2)光照:也因组织不同而异。 茎尖、叶片组织培养要光照,以便进行光合作用,.花果培养要避免直射光,以散射光或暗中培养较宜,根组织培养通常在暗处进行。 9.在组织培养中的几个阶段: 植物体—外植体—愈伤组织—生长点—植株 名词:脱分化(dedifferentiation)、愈伤组织、再分化(rededifferentiation)
、愈伤组织、再分化(rededifferentiation)")
279
第三节 植物的生长 1.植物的生长实际上就是细胞数目的增多和体积的增大,因此,植物的生长是一个体积或重量的不可逆的增加过程。
2.植物生长的性质和动物的有本质上的区别。脊椎动物在出生后已具备了成年动物的一切主要器官,它的生长不过是各部分(头、躯干和四肢)体积的同时增大而已,并且动物的生长迟早总会达到一定的限度。种子植物的生长则不同。植物出生后,在整个生活过程中,都在继续不断地产生新的器官,而且,由于茎和根尖端的组织始终保持胚胎状态,茎和报中又有形成层,所以,可以不断的生长。
体积的同时增大而已,并且动物的生长迟早总会达到一定的限度。种子植物的生长则不同。植物出生后,在整个生活过程中,都在继续不断地产生新的器官,而且,由于茎和根尖端的组织始终保持胚胎状态,茎和报中又有形成层,所以,可以不断的生长。")
280
一、茎的生长 (一)茎生长特性 (l)停滞期(lag phase); (2)对数生长期(logarithmic growth phase);
控制茎生长最重要的组织是顶端分生组织和近顶端分生组织。茎的生长速率都表现出“慢—快—慢”的基本规律,即开始时生长缓慢,以后逐渐加快,达到最高点,然后生长速率又减慢以至停止,这三个阶段总合起来叫做生长大周期(grand Period of growth)。可细分为四个时期: (l)停滞期(lag phase); (2)对数生长期(logarithmic growth phase); (3)直线生长期(linear growth phase); (4)衰老期(senescence Phase)。
。可细分为四个时期: (l)停滞期(lag phase); (2)对数生长期(logarithmic growth phase); (3)直线生长期(linear growth phase); (4)衰老期(senescence Phase)。")
281
(二)影响茎生长的条件 1.温度 生长最适温度与协调的最适温度 温周期现象(thermoperiodicity of growth):在自然条件下,也具有日温较高和夜温较低的周期性变化。植物这种对昼夜温度周期性变化的反应,称为生长的温周期现象。 2.光 光对茎生长有抑制作用。

282
光线抑制生长的原因有: (1)光照使自由IAA转变为无活性的结合态IAA; (2)光照提高IAA氧化酶活性,IAA含量就下降。然而,光照也会促进虽菜黄质分解民生长抑制物。光照既会减少生长素含量,又会增加生长抑制物的含量,所以抑制茎的生长; (3)红光增加细胞质钙离子浓度,活化CaM,分泌离子到细胞壁,细胞延长减慢。 3.水分 4.植物激素 5.机械刺激
红光增加细胞质钙离子浓度,活化CaM,分泌离子到细胞壁,细胞延长减慢。 3.水分. 4.植物激素. 5.机械刺激.")
283
(三)顶端优势 1.植物体是各个部分的统一整体。因此,植物各部分间的生长互相有着极密切的关系。植物各部分间的相互制约与协调的现象,称为相关性(correlation)。 2.顶端在生长上占有优势的现象,称为顶端优势(apical dominance)。 3.为什么会发生顶端优势现象呢? 和生长素有关。

284
二、根的生长 (一)根生长特性 根的生长部位也有顶端分生组织,根的生长也具生长大周期。根也有顶端优势。 (二)影响根生长的条件 1.光;
2.土壤阻力; 根在土壤中生长受阻力后会促进内源植物激素的合成。 3.土壤水分; 4.土壤温度。

285
(三)根和地上部的相关 1.为什么必须根生长得好,地上部分才能很好地生长呢?(1)首先,地上部分生长所需要的水分和矿物质,主要是由根系供应的。 (2)根系对地上部分的生长还有其他重要作用。 根的伤流液中还有细胞分裂素,这说明根系可产生细胞分裂素供应地上部分; 根系还能合成植物碱等其他各种复杂的含氮化合物,烟草叶中的烟碱也是在根部合成后,才运送到叶子去的; 2.植物地上部分对根的生长也有促进作用。 3.在某些条件下,根和地上部分的生长也会相互抑制。

286
三、叶的生长 (一)叶的发生 叶原基—叶轴—叶片—叶柄 (二)影响叶生长的条件
叶片的大小主要决定于细胞数目,而细胞大小是次要因子。影响叶生长的条件如下: 1.矿质营养氮肥 2.水分 3.光照 4.温度 5.植物激素

287
四、营养生长和生殖生长的相关 生殖器官生长所需的养料,大部分是由营养器官供应的。营养器官生长不好,生殖器官的生长自然也不会好。 营养器官和生殖器官生长之间也是有矛盾的,它表现在营养器官生长对生殖器官生长的抑制,和生殖器官生长对营养器官生长的抑制两个方面。 当营养器官生长过旺,消耗较多养分,便会影响到生殖器官的生长。反过来,生殖器官生长同样也影响营养器官生长。

288
第四节植物的运动 植物体的器官在空间可以产生位置移动,这就是植物的运动(movement)。
高等植物的运动可分为向性运动(tropic movement)和感性运动(nastic movement)。 向性运动是由光、重力等外界刺激而产生的,它的运动方向取决于外界的刺激方向。 感性运动是由外界刺激(如光暗转变、触摸等)或内部时间机制而引起的,外界刺激方向不能决定运动方向。
和感性运动(nastic movement)。 向性运动是由光、重力等外界刺激而产生的,它的运动方向取决于外界的刺激方向。 感性运动是由外界刺激(如光暗转变、触摸等)或内部时间机制而引起的,外界刺激方向不能决定运动方向。")
289
一、向性运动 1.向性运动包括三个步骤: (1)感受(感受到外界刺激); (2)传导(将感受到的信息传导到向性发生的细胞), (3)反应(接受信息后,弯曲生长)。 2.向性运动是生长引起的、不可逆的运动。 3.分类:向光性、向重力性、向化性、向水性等。

290
(-)向光性 1.向光性(phototropism) 植物随光的方向而弯曲的能力,称为向光性。
2.分类:正向光性、负向光性及横向光性之分。一般来说,地上部器官具正向光性,根部为负向光性。 3.向光性在植物生活中具有重要的意义。 4.植物感受光的部位:茎尖、芽鞘尖端、根尖、某些叶片或生长中的茎。 5.向光性与光能有关,而光能又与光照强度和时间有关。 6.向光性反应的光受体是什么?向光性的光受体是存在于质膜上的核黄素(riboflavin)。 7.向光性的原因: 一是生长素分布不均匀; 一是抑制物质分布不均匀。
。 7.向光性的原因: 一是生长素分布不均匀; 一是抑制物质分布不均匀。")
291
(二)向重力性 1.向重力性就是植物在重力影响下,保持一定方向生长的特性。根顺着重力方向向下生长,称为正向重力性;茎背离重力方向向上生长,称为负向重力性;地下茎则水平方向生长,称为横向重力性。 2.感受重力的细胞器是平衡石。植物的平衡石是指淀粉体,一个细胞内有4~12个淀粉体,每个淀份体外有一层膜,内有1~8个淀粉粒。 3.向重力性的机理——平衡石假说 4.意义:

292
(三)向化性 1.向化性(chemotropism):是由某些化学物质在植物周围分布不平均引起的生长。 2.植物根部生长的方向就有向化现象,它们是朝向肥料较多的土壤生长的。 3.应用:水稻深层施肥的目的之一,就是深施肥料,使稻根向深处生长,分布广,吸收更多养分。 此外还有向水性(hydrotropism)。向水性是当土壤中水分分布不均匀时,根趋向较湿的地方的特性。
。向水性是当土壤中水分分布不均匀时,根趋向较湿的地方的特性。")
293
二、感性运动 1.感性运动方向和外界刺激方向无关。 2.感性运动是由生长着器官两侧或上下面生长不等引起的。 3.感性运动有两类: (1)生长运动(growth movement),不可逆的细胞伸长,如偏上性运动等; (2)紧张性运动(turgor movement),由叶枕膨压变化产生,是可逆性变化,如叶片感夜运动等。
紧张性运动(turgor movement),由叶枕膨压变化产生,是可逆性变化,如叶片感夜运动等。")
294
(一)偏上性和偏下性 1.偏上性(epinasty):植物器官向下弯曲生长的现象。 2.偏下性(hyponasty):植物器官向上弯曲生长的现象。 3.原因:叶柄两侧生长素浓度的不同。
偏上性和偏下性 1.偏上性(epinasty):植物器官向下弯曲生长的现象。 2.偏下性(hyponasty):植物器官向上弯曲生长的现象。 3.原因:叶柄两侧生长素浓度的不同。")
295
(二)感夜性 1.定义:许多植物的叶子(或小叶)白天高挺张开、晚上会合拢或下垂,这就是感夜性。 2.感夜运动的产生可能原因是: 叶片在白天合成许多生长素,主要运到叶柄下半侧,K和CI也运输到生长素浓度高的地方,水分就进入叶枕,细胞膨胀,导致叶片高挺。到晚上,生长素运输量减少,进行相反反应,叶片就下垂。 3.植物之所以对光暗有反应,是因为光敏色素起作用,这种昼夜有内在节奏的变化是由生物钟控制的。

296
(三)感热性 1.植物对温度变化起反应的感性运动,称为感热性(thermonasty)。 2. 意义: 使植物在适宜的温度下进行授粉,并且保护花的内部免受不良条件的影响。 (四)感震性 1.由震动引起植物器官运动称之为感震性。 2.含羞草叶子下垂的机理 3.震动刺激在含羞草中的传递

297
三、生理钟 1.定义:在没有昼夜变化和温度变化的稳恒条件下,叶子的升起和下降运动的每一周期接近27h。 由于这个周期不正好是24h,而只接近这个数值,因此,这样的周期叫做近似昼夜节律(circadian rhythm)。 2.生物节奏的几个特性: 首先,节奏的引起必须有一个信号,而一旦节奏开始,在稳恒的条件下仍然继续显示。 其次,一旦节奏开始,就以大约24h的节奏自由的运行。 3.植物借助干生理钟准确地进行测时过程。

298
第九章植物的生殖生理 在营养器官生长的基础上,在适宜的外界条件下,植物就分化出生殖器官(花),最后结出果实。植物要在适宜开花的季节才能分化出花,而季节变化的主要特征是温度高低和日照长短,植物开花就与温度及日照长度有密切的关系。研究表明,春化作用和光周期是控制植物开花的两种主要机制。春化作用是指植物需要经过低温后才能开花.光周期是指植物在适合的日照长度才能开花。然而。要使植物从营养生长转变为生殖生长,还需要经过幼年期,植株有一定营养体,才能开花。所以,幼年期、春化作用和光周期是控制植物开花的三个重要阶段。
,最后结出果实。植物要在适宜开花的季节才能分化出花,而季节变化的主要特征是温度高低和日照长短,植物开花就与温度及日照长度有密切的关系。研究表明,春化作用和光周期是控制植物开花的两种主要机制。春化作用是指植物需要经过低温后才能开花.光周期是指植物在适合的日照长度才能开花。然而。要使植物从营养生长转变为生殖生长,还需要经过幼年期,植株有一定营养体,才能开花。所以,幼年期、春化作用和光周期是控制植物开花的三个重要阶段。")
299
第一节 幼年期 1.幼年期(juvenility)是植物早期生长的阶段。在此期间,任何处理都不能诱导开花。 2.植物必须达到一定年龄或经过一定时期的生长后,才能开花。许多植物完成幼年期后即转入成熟开花期,在适宜环境中诱导开花。 3.幼年期时间长短因植物种类而异。 木本植物长于草本植物; 有的植物根本没有幼年期,因为种子中已具备花原基。

300
一、幼年期的特征 1.幼年期和成熟期的区别 (1)形态特征: (2)生理特征 幼年期生长快,呼吸强,核酸代谢和蛋白质合成都快。 幼年期茎的切段易发根,可能与幼年期切条内合较多生长素有关。 2.植株从幼年期转变为成年期是由茎基向顶端转变,所以植株不同部位的成熟度不一样。 3.植株一旦成熟非常稳定,除非经过有性生殖,重新进入幼年期。 4.在生产中的应用:扦插、嫁接

301
二、提早成熟 植株处于幼年期不能开花,所以要设法加速生长,迅速通过幼年期,或者设法减慢生长以提早开花。 提早开花,可能是长日照促进植物生长的缘故。 树的大小决定幼年期长短。 内源赤霉素在幼年期转变为成年期中起作用。 靠近地面的根对维持幼年期很重要。

302
第二节春化作用 一、春化作用的条件 1.春化作用(vernalization):低温促使植物开花的作用,称为春化作用。
2.措施:闷麦法、七九小麦。 3.各种植物春化所要求的温度有所不同,有效温度界于零度以下和10度之间。最适温度是1-7度。 4.一般说来,冬性愈强,要求的春化温度愈低,春化的天数也愈长。 5.一些长日植物的春化作用可被高温(约30℃)或短日处理而消失。春化作用一旦完成,就非常稳定,高温处理就不起作用。这种由另一条件消除春化的现象,称为脱春化作用(devernalization)。 6影响春化的因素:糖、氧、水分
或短日处理而消失。春化作用一旦完成,就非常稳定,高温处理就不起作用。这种由另一条件消除春化的现象,称为脱春化作用(devernalization)。 6影响春化的因素:糖、氧、水分.")
303
二、春化作用的时期、部位和刺激传导 1.时期: (1)可在种子萌发或在植株生长的任何时期中进行; (2)少数植物只有在绿色幼苗长到一定大小,才能通过春化。 2.感受部位: 接受低温影响的部位是茎尖生长点。只有正在分裂的细胞才具有春化能力。 证据:芹菜试验 3.春化作用的刺激传导 以春化素(vernalin)形式传导。
形式传导。")
304
三、春化作用的生理生化变化 1.春化处理的小麦种子呼吸速率升高; 2.在春化过程中核酸(特别是RNA)在体内含量增加,代谢加速,而且RNA性质也有所变化; 3.低温处理的冬小麦种子中可溶性蛋白及游离氨基酸含量增加,其中脯氨酸增加较多; 4.春化处理作物体内赤霉素含量增加。

305
1.MeIchers-Lang假说:认为春化作用至少有两个阶段
四、春化作用的机理 1.MeIchers-Lang假说:认为春化作用至少有两个阶段 第一个阶段是春化作用的前作物在低温下转变成不稳定的中间产物(中间产物如在高温下可被破坏或被钝化); 第二个阶段是在 20度下,中间产物转变为热稳定的物质,即春化作用的最终产物. 2.谭克辉将春化过程依次分为5个分过程: (1)初期以旺盛的氧化及氧化磷酸化为主的过程; (2)以脱氢酶活性逐渐占优势的代谢过程; (3)与核酸代谢密切相关的过程; (4)与蛋白质代谢密切相关的过程; (5)低温诱导末期及其稳定时期。
; 第二个阶段是在 20度下,中间产物转变为热稳定的物质,即春化作用的最终产物. 2.谭克辉将春化过程依次分为5个分过程: (1)初期以旺盛的氧化及氧化磷酸化为主的过程; (2)以脱氢酶活性逐渐占优势的代谢过程; (3)与核酸代谢密切相关的过程; (4)与蛋白质代谢密切相关的过程; (5)低温诱导末期及其稳定时期。")
306
第三节光周期 1.光周期(photoperiod): 在一天之中,白天和黑夜的相对长度,称为光周期。
2.对多数植物来说,特别是一年生和二年生植物,当同一种植物生长在特定的纬度的时候,每年都大约在相同的日子开花。 3.植物对于白天和黑夜的相对长度的反应,称为光周期现象(photoperiodism)。
。")
307
一、光周期反应类型 1.光周期反应类型可分为三种 短日植物(short-day plant): 长日植物(long-day plant): 日中性植物(intermediate-day plant): 2.临界日长(critical daylenth):指诱导短日植物开花所需的最长日照时数,或诱导长日植物开花所需的最短日照时数。不同植物临界日长不一。 3.临界暗期(critical dark period):在昼夜周期中短日植物能够开花所必需的最短暗期长度,或长日植物能够开花所必需的最长暗期长度。
:指诱导短日植物开花所需的最长日照时数,或诱导长日植物开花所需的最短日照时数。不同植物临界日长不一。 3.临界暗期(critical dark period):在昼夜周期中短日植物能够开花所必需的最短暗期长度,或长日植物能够开花所必需的最长暗期长度。")
308
4.长短日植物(LSDP):经短日照完成花诱导后,再在长日照条件下形成花器官的植物;
5.短长日植物(SLDP):经长日照完成花诱导后,再在短日照条件下形成花器官的植物。 6.植物开花究竟决定于日长或夜长? 7.不同纬度地球表面一年中光周期的变化与植物的适应性。 8.春化作用与光周期有相互作用。
:经长日照完成花诱导后,再在短日照条件下形成花器官的植物。 6.植物开花究竟决定于日长或夜长? 7.不同纬度地球表面一年中光周期的变化与植物的适应性。 8.春化作用与光周期有相互作用。")
309
二、光周期刺激的感受和传导 1.感受光周期刺激的部位是 叶子。 通常植物生长到一定程度后,才有可能接受光周期的诱导。 不同植物开始对光周期敏感的年龄不同,年龄越大,光周期诱导的时间也越短。 2.诱导开花部位是茎尖端的生长点 因此,由叶中产生的开花刺激物必定有一个传导问题。嫁接试验即可证明。 3.两种光周期反应的植物所产生的开花刺激物,没有什么区别。 4.运输途径主要是韧皮部。

310
三、光周期诱导 1.光周期诱导(photoperiodic induction):植物只需要一定时间适宜的光周期处理,以后即使处于不适宜的光周期下,仍然可以长期保持刺激的效果,即花的分化不出现在适宜的光周期处理的当时,而是在处理后若干天,这种现象称为光周期诱导。 2.光周期诱导所需的光周期处理天数,因植物种类而异。 3.每种植物光周期诱导需要的天数随植物的年龄以及环境条件,特别是温度、光强及日照的长度而定。

311
四、光对暗期中断 1.在自然条件下,光周期诱导所要求的光照强度是很微弱的,远远低于光合作用所需要的光照强度。 2. 一般认为光周期诱导的光照强度约在50~100lX,但不同植物甚至品种的反应亦不同。 3.短日植物应该称为长夜植物,它的开花决定于暗期的长度,而不决定于光期时间的长度。 4.无论是抑制短日植物开花,还是诱导长日植物开花,都是红光最有效。 5.如果在红光照过之后立即再照以远红光,就不能发生夜间断(night break)的作用,也就是被远红光的作用所抵消。在植物的花诱导上,也有着光敏色素的参与。
的作用,也就是被远红光的作用所抵消。在植物的花诱导上,也有着光敏色素的参与。")
312
五、光敏色素与花诱导 (一)花诱导的作用光谱 作用光谱曲线:在暗期间断试验中,如果在暗期中间用各种单色光对植物进行闪光处理。几天之后,开始观察花原基的发生。根据达到同样开花水平(如和对照相比抑制50%)所需的不同波长光的能量,绘出的光谱曲线。 阻止短日植物(大豆和苍耳)开花的作用光谱和促进长日植物(各大麦)开花的作用光谱很相似。 红光对植物开花最有效,蓝光效果很差。
开花的作用光谱和促进长日植物(各大麦)开花的作用光谱很相似。 红光对植物开花最有效,蓝光效果很差。")
313
(二)光敏色素与诱导开花 1.短日植物的开花诱导要求是暗期的前期“高Pfr反应”,后期是“低Pfr反应”。 2.长日植物在暗期前期是“低Pfr反应”,后期是“高Pfr反应”。 综上所述,短日植物和长日植物对日长的本质区别,似乎在于暗期的前期。短日植物在暗期的前期是高Pfr水平,然后转变为低Pfr水平,长日植物暗期的前期是低P fr水平,后期是高Pfr水平。

314
六、光周期诱导开花的机理 (-)成花素假说 1.适宜的光周期下,植物的叶中形成开花的刺激物,可以从一株植物传递到另一株植物。这种物质被认为是一种激素,即成花素。 2.近年来,业已确定赤霉素对某些长日植物(如天仙子、金光菊、黑眼菊)可代替光照条件,而在非诱导的短日条件下开花。对某些冬性长田植物(如胡萝卜、甘蓝)又可代替低温,即不经春化即可开花。 3.也证明这些植物经过诱导之后,内源的赤霉素含量都有所增加。因此,使人很容易联想到赤霉素和诱导植物开花有关。
可代替光照条件,而在非诱导的短日条件下开花。对某些冬性长田植物(如胡萝卜、甘蓝)又可代替低温,即不经春化即可开花。 3.也证明这些植物经过诱导之后,内源的赤霉素含量都有所增加。因此,使人很容易联想到赤霉素和诱导植物开花有关。")
315
4.赤霉素还不是开花激素: 首先,在长日照的作用下,花原基的分化和茎的伸长是同时发生的,而在赤霉素诱导的作用下,茎的伸长在花原基分化之前; 其次,赤霉素不能促进短日植物在非诱导条件下开花。 5.激素假说:假定成花素(即诱导开花的激素)是由形成茎所必需的赤霉素和形成花所必需的开花素两组具有活力的物质组成。植物体内存在赤霉素和开花素,才能开花。 6.中性植物本身具有赤霉素和开花素,所以,不论在长、短日照条件下都能开花。 7.长日植物在长日条件下,短日植物在短日条件下,都具有赤霉素和开花素,所以,都可以开花。 8.赤霉素限制长日植物开花,而开花素限制短日植物开花。
是由形成茎所必需的赤霉素和形成花所必需的开花素两组具有活力的物质组成。植物体内存在赤霉素和开花素,才能开花。 6.中性植物本身具有赤霉素和开花素,所以,不论在长、短日照条件下都能开花。 7.长日植物在长日条件下,短日植物在短日条件下,都具有赤霉素和开花素,所以,都可以开花。 8.赤霉素限制长日植物开花,而开花素限制短日植物开花。")
316
(二)光敏色素假说 1.内容:光敏色素会活化某些特定的基因,促进参与新的蛋白质或酶。 2.证据: (1)许多试验证实,在光周期诱导下,叶片中的核酸(主要是RNA)和蛋白质都发生了不同程度的变化,说明这些物质有可能参与开花的诱导过程。 (2)试验表明,红光照射后提高子叶的NAD P含量,远红光则降低红光反应,减少NAD P含量;同时证实从Pfr促进NAD激酶活性 可见 Pfr作用与NAD P合成或分解有关。 (3)也有实验证明, Pfr很可能使膜蛋白质结构发生变化,导致膜透性改变,从而引起一系列变化。
试验表明,红光照射后提高子叶的NAD P含量,远红光则降低红光反应,减少NAD P含量;同时证实从Pfr促进NAD激酶活性. 可见 Pfr作用与NAD P合成或分解有关。 (3)也有实验证明, Pfr很可能使膜蛋白质结构发生变化,导致膜透性改变,从而引起一系列变化。")
317
(三)碳氮比理论 1.内容:凡是C/N较大时,则开花,反之,C/N较小,则延迟开花或不开花。 2.证据: 果树栽培上,利用刀砍伤或用绞缢树皮以使C/N加大,促使早开花结果。 作物栽培上施用大量氮肥,会重C/N变小,延迟作物的开花,使作物晚熟。 3.应注意的是,Klebs进行研究时所用的植物,都是长日植物或者是中性植物,而没有短日植物。 4.可以认为只有适当的C/N比,才能保证花器官正常形成的需要,使能顺利地开花。

318
七、春化和光周期理论在农业上的应用 (一)春化处理 提早成熟 冬作春播 加速育种。 (二)控制开花 鲜花适时供应 (三)引种 “短南北移迟”,其余可推导
春化处理 提早成熟 冬作春播 加速育种。 (二)控制开花 鲜花适时供应 (三)引种 短南北移迟 ,其余可推导")
319
第四节 花器官形成的生理 一、花分化初期茎生长点的形态及生理变化 1.花分化时生长锥的表面积都变大。 2.主要形态和生理变化:
第四节 花器官形成的生理 一、花分化初期茎生长点的形态及生理变化 1.花分化时生长锥的表面积都变大。 2.主要形态和生理变化: (1)生长锥的表面一层或数层细胞分裂加速,细胞小而细胞质浓,而中部的一些细胞则分裂减慢,细胞变大,细胞质稀薄,有的出现了液泡。 (2)生长锥表面的细胞具有较高的蛋白质和RNA含量。 说明花的分化是与DNA-RNA-蛋白质系统活化有关。 (3)由于表层的分生细胞的迅速分裂,使生长锥表面出现皱折,在原来形成叶原基的地方形成花原基,在花原基上再分化出花的各部分的原基。
生长锥的表面一层或数层细胞分裂加速,细胞小而细胞质浓,而中部的一些细胞则分裂减慢,细胞变大,细胞质稀薄,有的出现了液泡。 (2)生长锥表面的细胞具有较高的蛋白质和RNA含量。 说明花的分化是与DNA-RNA-蛋白质系统活化有关。 (3)由于表层的分生细胞的迅速分裂,使生长锥表面出现皱折,在原来形成叶原基的地方形成花原基,在花原基上再分化出花的各部分的原基。")
320
二、花器官形成所需的条件 在植物的成花过程中,花的诱导和花器官形成是两个过程。 影响花器官形成的内外条件: (-)气象条件
1.光:照光时间越长,光强度越大,对花形成愈有利。 2.温度。 (二)栽培条件 1.水分; 2.肥料; 3.栽培密度。 (三)生理条件 1.体内有机物养分充足与否,是决定花器官形成好坏的主要条件, 2.在营养生长和生殖生长之间有养分分配问题,在同一花序的小花之间也有这个问题。 强势花、弱势花。
栽培条件. 1.水分; 2.肥料; 3.栽培密度。 (三)生理条件. 1.体内有机物养分充足与否,是决定花器官形成好坏的主要条件, 2.在营养生长和生殖生长之间有养分分配问题,在同一花序的小花之间也有这个问题。 强势花、弱势花。")
321
三、植物性别的分化 (一)雌雄个体的代谢差异 1.雌株处于较还原状态,雄株处于相对氧化状态; 2.多数雄株组织的呼吸速率大干雌株的;
3.多数雄株的过氧化氢酶活性也比雌株高。 (二)外界条件对植物性别形成的影响 1.光周期:短日照促使短日植物多开雌花,长日植物多开雄花。 2.糖氮比值低时,将提高雌花分化的百分数。 3.土壤条件对植物性别分化比较明显。 4.植物激素对花的性别分化也有影响。 5.伤害也可使雄株转变为雌株。
外界条件对植物性别形成的影响. 1.光周期:短日照促使短日植物多开雌花,长日植物多开雄花。 2.糖氮比值低时,将提高雌花分化的百分数。 3.土壤条件对植物性别分化比较明显。 4.植物激素对花的性别分化也有影响。 5.伤害也可使雄株转变为雌株。")
322
第五节 受精生理 一、花粉寿命与贮藏 1.花粉寿命:花粉离开母体后,维持能授粉能力的时间长短。不同种类植物的花粉生活力有很大的差异。
第五节 受精生理 一、花粉寿命与贮藏 1.花粉寿命:花粉离开母体后,维持能授粉能力的时间长短。不同种类植物的花粉生活力有很大的差异。 2.影响花粉寿命的条件: (1)湿度 (2)温度 (3)空气中二氧化碳和氧的含量 (4)光线对花粉的贮存也有影响,一般以遮荫或在黑暗处贮存较好。 3.粉贮存期生活力逐渐降低的原因 贮藏物质消耗过多、酶活性下降和水分过度缺乏。泛酸减少。
湿度. (2)温度. (3)空气中二氧化碳和氧的含量. (4)光线对花粉的贮存也有影响,一般以遮荫或在黑暗处贮存较好。 3.粉贮存期生活力逐渐降低的原因. 贮藏物质消耗过多、酶活性下降和水分过度缺乏。泛酸减少。")
323
二、柱头的生活能力 雌蕊柱头承受花粉能力持续时间长短,主要与柱头的生活能力有关。 柱头的生活能力一般都能持续一个时期,具体时间长短则因植物的种类而异。 在作物杂交育种过程中,需要了解柱头的生活能力,什么时候开始,什么时候最强,什么时候丧失能力,为授粉工作提供科学根据,以便提高制种的产量和质量。 一个雄穗上住头丧失生活能力的顺序,和花柱在穗轴上发生的先后是一致的,即穗中、下部先丧失,顶端后丧失。

324
三、外界条件对授粉的影响 (一)温度 水稻抽穗开花期的最适温度是30~35℃。 (二)湿度 水稻开花的最适湿度是70一80%,否则也影响授粉。 (三)风 对风媒花的授粉也有较大影响,无风或大风都不利于作物授粉。 (四)土壤中的肥料
土壤中的肥料.")
325
四、花粉和柱头的相互“识别” 1.识别反应决定于花粉壁中蛋白质和柱头乳突细胞表面的蛋白质表膜之间的相互关系。 2.识别过程:当花粉受到潮湿时,外壁蛋白迅速释放。因此,花粉的识别物质是外壁蛋白。而雌蕊的识别感受器是柱头表面的亲水的蛋白质表膜(pellicle)具粘性,易捕捉花粉。 (1)当种内进行杂交时,亲和性花粉释放出外壁蛋白并扩散入柱头表面,与柱头表层感受器——蛋白质表膜相互作用,相互“认识”。如认可,花粉管便萌发穿过柱头,直至受精。
当种内进行杂交时,亲和性花粉释放出外壁蛋白并扩散入柱头表面,与柱头表层感受器——蛋白质表膜相互作用,相互 认识 。如认可,花粉管便萌发穿过柱头,直至受精。")
326
(2)如果是远缘杂交,不亲和性花粉的外壁蛋白和柱头表面的蛋白质表膜不相认识,相互拒绝,花粉管生长受阻,不能穿入柱头;而柱头乳突细胞也产生胼胝质,阻碍花粉管穿入,而且花粉管尖端也被胼胝质封闭,使受精失败. 3.意义:花粉与柱头的相互识别的机能,是植物在长期进化过程中形成的,保证物种稳定与繁衍。 4.克服不亲和性的方法之一—混合花粉授粉法。 方法是接生活的不亲和性花粉的同时,混合一些杀死的亲和的花粉。亲和的花粉可使柱头不能识别不亲和的花粉,被称为蒙导花粉。

327
五、花粉的萌发和花粉管的伸长 1.花粉落在雌蕊的柱头上,受到柱头分泌物的刺激,开始吸水萌发。花粉中含有淀粉和脂肪等后含物,水势较低,于是便从柱头吸水,花粉内壁便通过萌发孔向外突出,形成花粉管。 2.生理生化变化:花粉萌发时,酶的活性加强,其中磷酸化酶、淀粉酶、转化酶等活性剧烈增强,有时甚至比原来高6倍之多。花粉萌发时,呼吸速率剧增,蛋白质合成也加快。 3.花粉管的生长是由它的尖端延伸,这种延伸是由花粉管内膨压推动的。 4.花粉的萌发有群体效应 5.花粉向胚囊定向生长的原因:花粉管的向化性运动。

328
六、受精前后雌蕊的代谢变化 1.花粉中不断向花柱分泌各种酶类 雌蕊组织中的糖类和蛋白质的代谢作用都在加强 呼吸作用也在加强 授粉后雌蕊组织的吸收水分和无机盐的能力也都在增强。 2.授粉后雌蕊中的生长素含量大大增加 (1)花粉含有生长素有关。 (2)主要是花粉中含有使色氨酸转变成吲哚乙酸酶体系,花粉管在生长过程中,能将这些酶分泌到雌蕊组织,因此,引起花柱和子房形成大量生长素。
花粉含有生长素有关。 (2)主要是花粉中含有使色氨酸转变成吲哚乙酸酶体系,花粉管在生长过程中,能将这些酶分泌到雌蕊组织,因此,引起花柱和子房形成大量生长素。")
329
随着花粉管的伸长,雌蕊各部分生长素不同时间雌蕊组织各部分量高峰按花枝顶端、花柱基部和子房的顺序出的生长素含量变化现。
3.卵细胞受精前处于休眠状态,受精后受到激活,呼吸速率也伴随着发生变化。 4.受精引起子房代谢剧烈变化的原因之一,是子房的生长素含量迅速增加。大量生长素“吸引”营养器官的养料集中运到生殖器官。

330
第十章植物的成熟和衰老生理 植物受精后各部分发育目标: 受精卵发育成胚; 胚珠发育成种子; 子房壁发育成果皮; 子房发育形成果实。

331
第一节种子成熟时的生理生化变化 种子的成熟过程,实质上就是胚从小长大,以及营养物质在种子中变化和积累的过程。
种子成熟期间的物质变化,大体上和种子萌发时的变化相反,植株营养器官的养料,以可溶性的低分子化合物状态(如葡萄糖、蔗糖、氨基酸等形式)运往种子,在种子中逐渐转化为不溶性的高分子化合物(如淀粉、蛋白质和脂肪等),并且积累起来。
运往种子,在种子中逐渐转化为不溶性的高分子化合物(如淀粉、蛋白质和脂肪等),并且积累起来。")
332
一、主要有机物的变化 1.淀粉的合成: 淀粉磷酸化酶在淀粉合成中起主导作用。 增强淀粉磷酸化酶活性的条件如下: (1)淀粉磷酸化酶的最适PH值是6.5; ( 2)该酶活性最适温度是26~46度; (3)磷酸含量要适当。 磷酸供给过多会影响淀粉合成速度,过少不利于糖的运输。
磷酸含量要适当。 磷酸供给过多会影响淀粉合成速度,过少不利于糖的运输。")
333
2.蛋白质的合成:总氮变化不大,非蛋白质态氮不断下降,蛋白质氮的含量不断增加。
3.脂肪的形成:脂肪是由糖类转化而来的。 油脂形成有两个特点: (1)成熟期所形成的大量游离脂肪酸,随着种子的成熟,游离脂肪酸逐渐合成复杂的油脂。 (2)种子成熟时先形成饱和脂肪酸,然后,由饱和脂肪酸变为不饱和脂肪酸, 碘值 总之,在种子成熟过程中,可溶性糖类转化为不溶性糖类,非蛋白质氮转变为蛋白质,而脂肪则是由糖类转化而来的。
成熟期所形成的大量游离脂肪酸,随着种子的成熟,游离脂肪酸逐渐合成复杂的油脂。 (2)种子成熟时先形成饱和脂肪酸,然后,由饱和脂肪酸变为不饱和脂肪酸, 碘值. 总之,在种子成熟过程中,可溶性糖类转化为不溶性糖类,非蛋白质氮转变为蛋白质,而脂肪则是由糖类转化而来的。")
334
二、其他生理变化 1.呼吸速率: 有机物积累迅速时,呼吸作用也旺盛,种子接近成熟时,呼吸作用逐渐降低。 2.激素变化: 首先出现的是玉米素,可能调节建成籽粒的细胞分裂过程,然后是赤霉素和生长素可能调节有机物向籽粒的运输和积。此外,籽粒成熟期脱落酸大量增加,可能和籽粒的休眠有关。 3种子含水量: 与有机物的积累恰好相反,它是随着种子的成熟而逐渐减少的。

335
三、外界条件对种子成熟和化学成分的影响 1.干燥与热风 (1)造成籽粒瘦小,产量大减。 (2)干旱也可使籽粒的化学成分发生变化。风旱不实的种子中蛋白质的相对含量较高。 2.温度 种子成熟期间,适当的低温有利于油脂的累积。在油脂品质上,种子成熟时温度较低而昼夜温差大时,利于不饱和脂肪酸的形成;在相反的情形下,利于饱和脂肪酸的形成。 3.营养条件 氮肥、钾肥、磷肥

336
第二节 果实成熟时的生理生化变化 肉质果实成熟时的生理生化变化。 一、果实的生长 1.肉质果实的生长也具有生长大周期,呈S型生长曲线; 2.但也有一些核果及某些非核果的生长曲线,则呈双S型,即在生长的中期有一个缓慢期。这个时期正好是珠心和珠被生长停止,幼胚生长强烈的时期,这时核也正在变硬。

337
3.单性结实: (1)定义:不经受精作用而形成不含种子的果实的,称为单性结实(parthenocarpy)。 (2)分类:单性结实有天然的单性结实和刺激性单性结实之分。 天然的单性结实:指不需要经过受精作用就产生无子果实的。同一种植物,无子种的子房中生长素含量较有子种的为高。 刺激性单性结实:指必需给以某种刺激才能产生无子果实。 (3)原因: 果实生长与受精后子房生长素含量增多有关。
原因: 果实生长与受精后子房生长素含量增多有关。")
338
二、呼吸骤变 1.当果实成熟到一定程度时,呼吸速率首先是降低,然后突然增高,最后又下降,此时果实使进入完全成熟。这个呼吸高峰,便称为呼吸骤变(respiratory climacteric)。 2.具呼吸骤变的骤变型果实有:苹果、香蕉、梨、桃、番木瓜、忙果和鳄梨等;不具呼吸骤变的非骤变型集实有:橙、凤梨、葡萄、草毒和柠檬等。

339
3.骤变型果实和非骤变型果实的主要区别 (1)前者含有复杂的贮藏物质(淀粉或脂肪),在摘果后达到完全可食状态前,贮藏物质强烈水解,呼吸加强,而后者并不如此。 (2)骤变型果实成熟比较迅速,非骤变型果实成熟比较缓慢。 4.果实发生呼吸骤变的原因: 由于果实中产生乙烯的结果。乙烯可增加果皮细胞的透性,加强内部氧化过程,促进果实的呼吸作用,加速果实成熟。 5.后熟作用:出现呼吸骤变期间果实内部的变化称为果实的后熟作用。 6.应用:以推迟或提早果实成熟。 控制气体法延熟及人工催熟

340
三、肉质果实成熟色香味变化 1.果实变甜 呼吸骤变出现后,淀粉转变为可溶性糖。 2.酸味减少 在成熟过程中,多数果实有机酸含量下降,因为有些有机酸转变为糖,有些则由呼吸作用氧化成二氧化碳和水,有些则被K,Ca等所中和。 3.涩味消失 果实成熟时,单宁被过氧化物酶氧化成无涩味的过氧化物,凝结成不溶于水的胶状物质。

341
4.香味产生 果实成熟时产生一些具有香味的物质,主要是酯类,还有一些特殊的醛类等。 5.由硬变软 果肉细胞壁中层的果胶质变为可溶性的果胶。 果肉细胞中的淀粉粒的消失(淀粉转变为可溶性糖)。 6.色泽变艳 果皮中叶绿素被破坏丧失绿色,类胡萝卜素仍较多存在,呈现黄色,或者由于形成花色素而呈现红色。 肉质果实果肉的有机物的变化,明显受温度和湿度的影响。在夏凉多雨的条件下,果实中含酸量较多,而糖分则相对减少;而在阳光充足、气温较高及昼夜温差较大的条件下,果实中含酸少而糖分较多。

342
四、果实成熟时蛋白质和激素的变化 1.果实成熟与蛋白质合成有关。 证据:蛋白质合成抑制剂抑制果实成熟。 2.在果实成熟过程中,生长素、赤霉素、细胞分裂素、脱落酸和乙烯五类植物激素,都是有规律地参加到代谢反应中。

343
第三节 种子和延存器官的休眠 成熟种子或器官在合适的萌发条件下仍不萌发的现象,称为休眠(dormancy)。 一、种子休眠的原因和破除
第三节 种子和延存器官的休眠 成熟种子或器官在合适的萌发条件下仍不萌发的现象,称为休眠(dormancy)。 一、种子休眠的原因和破除 (一)种皮限制 1.原因:种皮不能透水或透水性弱;种皮不透气;种皮大坚硬,胚不能突破种皮,也难以萌发。 2.破除:现在一般采用物理、化学方法来破坏种皮,使种皮透水透气。 (二)种子未完成后熟 1.种子的后熟作用:有些种子的胚已经发育完全,但在适宜条件下也不能萌发它们一定要经过休眠,在胚内部发生某些生理生化变化,才能萌发。这些种子在休眠期内发生的生理生化过程,称为后熟作用(after ripening)。
。 一、种子休眠的原因和破除. (一)种皮限制. 1.原因:种皮不能透水或透水性弱;种皮不透气;种皮大坚硬,胚不能突破种皮,也难以萌发。 2.破除:现在一般采用物理、化学方法来破坏种皮,使种皮透水透气。 (二)种子未完成后熟. 1.种子的后熟作用:有些种子的胚已经发育完全,但在适宜条件下也不能萌发它们一定要经过休眠,在胚内部发生某些生理生化变化,才能萌发。这些种子在休眠期内发生的生理生化过程,称为后熟作用(after ripening)。")
344
在后熟过程中,种子内的淀粉、蛋白质、脂肪等有机物的合成作用加强,呼吸减弱,酸度降低。经过后熟作用后,种皮透性增加,呼吸增强,有机物开始水解。
2.破除:这类种子必需经低温处理,即用湿砂将种子分层堆积在低温(5度左右)的地方1~3个月,经过后熟才能萌发。这种催芽的技术称为层积处理(stratification)。 (三)胚未完全发育 (四)抑制物质的存在 有些植物的果实或种子存在抑制种子萌发的物质,以防止种子的萌发。 生长抑制剂抑制种子萌发有重要的生物学意义。
的地方1~3个月,经过后熟才能萌发。这种催芽的技术称为层积处理(stratification)。 (三)胚未完全发育. (四)抑制物质的存在. 有些植物的果实或种子存在抑制种子萌发的物质,以防止种子的萌发。 生长抑制剂抑制种子萌发有重要的生物学意义。")
345
二、延存器官休眠的打破和延长 1.破除休眠: 用赤霉素破除休眠是当前最有效的方法。 晒种法效果也很好。 硫脲来破除马铃薯块茎的休眠。
2.延长休眠: 萘乙酸甲酯粉剂处理

346
第四节植物的衰老 植物衰老(senescence)是指一个器官或整个植株生命功能逐渐衰退过程。
人根据植物生长习性,开花植物有两类不同的衰老方式:一类是一生中能多次开花的植物;另一类是一生中只开一次花的植物。 一、衰老时的生理生化变化 (一)蛋白质显著下降 原因有两种可能:一是蛋白质合成能力减弱,一是蛋白质分解加快,两者可能同时进行。
蛋白质显著下降. 原因有两种可能:一是蛋白质合成能力减弱,一是蛋白质分解加快,两者可能同时进行。")
347
(二)核酸含量降低 衰老时RNA含量下降,是与RNA合成能力降低有关。也有认为是降解加快所致。 (三)光合速率下降 1.叶片衰老时叶绿体被破坏。 2.叶绿素含量迅速下降 3.蛋白水解酶活性增强的过程,核酮糖1,5-二磷酸羧化酶减少。 因此,认为在衰老的过程中,光合电子传递和光合磷酸化受作用受阻,所以光合速率下降。

348
(四)呼吸速率下降 在叶子衰老过程中,线粒体的变化不如叶绿体的那么大。 在衰老早期,线粒体体积变小,摺皱膨胀,数目减少,然而功能线粒体一直到衰老末期还是保留着。 叶片衰老时,呼吸速率迅速下降,后来又急剧上升,再迅速下降,似果实一样,有呼吸骤变,这种现象和乙烯出现高峰有关,因为乙烯加速透性,呼吸加强。 衰老过程的呼吸商与正常呼吸的不同,这说明它利用的不是糖而是氨基酸。 此外,衰老时呼吸过程的氧化磷酸化逐步解偶联,产生的ATP量也减少了,细胞中合成过程所需的能量不足,更促进衰老的发展。

349
二、影响衰老的外界条件 1.光 延缓叶片衰老是通过环式光合磷酸化而供给ATP,用于聚合物的再合成,或降低蛋白质、叶绿素和RNA的降解。 光敏色素在衰老过程中也起作用。 2.温度 低温和高温都会加速叶片衰老。 3.水分 干旱促进蛋白质降解和提高呼吸速率,叶绿体片层结构破坏,光合磷酸化受抑制,光合速率下降。 4.营养

350
三、植物衰老的原因 (一)营养亏缺理论 1.生殖器官从其他器官获得大量营养物质,致使其他器官缺乏营养而死亡,称营养亏缺理论。 2.不能说明下列问题: (1)即使供给已开花结实植株充分养料,也无法使植株免于衰老; (2)雌雄异株的大麻和菠菜,在雄株开雄花后,不能给实,谈不上积集营养体养分,但维株仍然衰老死亡。
雌雄异株的大麻和菠菜,在雄株开雄花后,不能给实,谈不上积集营养体养分,但维株仍然衰老死亡。")
351
(二)植物激素调控理论 1.理论:植物激素调控理论认为,单捻植物的衰老是由一种或多种激素综合控制的。 2.两种解释: (1)植株营养生长时,根系合成的细胞分裂素运到叶片,促使叶片蛋白质合成,推迟植株衰老。但是植株开花结实时,一方面是根系合成细胞分裂素数量减少,叶片得不到足够的细胞分裂素;
植物激素调控理论 1.理论:植物激素调控理论认为,单捻植物的衰老是由一种或多种激素综合控制的。 2.两种解释: (1)植株营养生长时,根系合成的细胞分裂素运到叶片,促使叶片蛋白质合成,推迟植株衰老。但是植株开花结实时,一方面是根系合成细胞分裂素数量减少,叶片得不到足够的细胞分裂素;")
352
另一方面是花和果实内细胞分裂素含量增大,成为植株代谢旺盛的生长中心,促使叶片的养料运问果实,这就是叶片缺乏细胞分裂素导致叶片衰老的原因。
(2)花或种子中形成促进衰老的激素(脱落酸和乙烯),运到植株营养器官所致。衰老来源于籽粒本身,而不是由根部造成的。 一般来说,细胞分裂素和赤霉素等可延缓叶片衰老,而脱落酸和乙烯等则促进叶片衰老。 生产上用苄基腺瞟吟(BA)延缓果蔬和食用菌的衰老,用乙烯利催熟香蕉、番茄、棉铃等。
花或种子中形成促进衰老的激素(脱落酸和乙烯),运到植株营养器官所致。衰老来源于籽粒本身,而不是由根部造成的。 一般来说,细胞分裂素和赤霉素等可延缓叶片衰老,而脱落酸和乙烯等则促进叶片衰老。 生产上用苄基腺瞟吟(BA)延缓果蔬和食用菌的衰老,用乙烯利催熟香蕉、番茄、棉铃等。")
353
第五节植物器官的脱落 脱落(abscission)是指植物细胞组织或器官与植物体分离的过程。 植物器官脱落是植物自我调节的手段。 是植物对外界环境的一种适应。
是指植物细胞组织或器官与植物体分离的过程。 植物器官脱落是植物自我调节的手段。 是植物对外界环境的一种适应。")
354
一、环境因子对脱落的影响 (一)温度 温度过高和过低都会加速器官脱落。 (二)水分 通常季节性的干旱会使树木落叶。 (三)光照 光照不足,器官容易脱落。 (四)氧 (五)矿质营养 氮素含量高时,生长素也较多,叶片不易脱落。缺钙的植株的营养器官很容易脱落。

355
二、脱落时细胞及生化变化 (一)离区的解剖 离区分为离层和保护层两部分。 (二)脱落时细胞的变化 1.RNA含量增加,内质网增多,高尔基体和小泡都增多。 2.小泡聚积在质膜,释放出酶到细胞壁和中胶层。 3.细胞壁和中胶层分解并膨大,以中胶层最为明显。

356
(三)脱落的生物化学 主要是水解离层的细胞壁和中胶层,使细胞分离,成为离层;促使细胞壁物质的合成和沉积,保护分离的断面,形成保护层。 在脱落发生之前,植物叶片或果实内植物激素含量发生变化,在激素信号的作用下,离区内合成RNA,翻译成蛋白质(酶),呼吸加强,提供上述变化的能量,因此脱落是一个需氧过程。
,呼吸加强,提供上述变化的能量,因此脱落是一个需氧过程。")
357
脱落主要酶类: 1.纤维素酶: 纤维素酶定位在离层,该酶在脱落过程中可能扮演主要角色。PI酸性和PI碱性纤维素酶,前者与细胞壁木质化关系密切,受IAA控制,后者与细胞壁分解关系密切,受乙烯控制。 2.果胶酶(pecLinase) 果胶甲酯酶(PME)或称果胶酶,主要是催化水解果胶酸的甲酯键。其活性与脱落成负相关。 ( 2)聚半乳糖醛酸酶(PG),主要催化多聚半乳糖醛酸的糖醛键,生成半乳糖醛酸,其活性与脱落呈正相关。
或称果胶酶,主要是催化水解果胶酸的甲酯键。其活性与脱落成负相关。 ( 2)聚半乳糖醛酸酶(PG),主要催化多聚半乳糖醛酸的糖醛键,生成半乳糖醛酸,其活性与脱落呈正相关。")
358
三、脱落与植物激素 (-)生长素 “生长素梯度”学说(auxin gradient theory) 决定脱落的不是生长素绝对含量,而是相对浓度,即离层两侧生长素浓度梯度起着调节脱落的作用。当远基端浓度高于近基端时,器官不脱落;当两端浓度差异小或不存在时,器官脱落;当远基端浓度低于近基端时,加速脱落。 (二)脱落酸 脱落酸能促进分解细胞壁酶的分泌,也能抑制叶柄内生长素的传导,所以促进器官脱落。短日照有利于脱落酸的合成,这正是说明短日照成为叶片脱落的环境信号的原因。
脱落酸. 脱落酸能促进分解细胞壁酶的分泌,也能抑制叶柄内生长素的传导,所以促进器官脱落。短日照有利于脱落酸的合成,这正是说明短日照成为叶片脱落的环境信号的原因。")
359
(三)乙烯 1.乙烯含量与脱落有密切关系。 乙烯引起脱落的机理,可能有两方面的作用: (1)诱导离区果胶酶和纤维素酶的合成,增加膜透性; (2)促使生长素钝化和抑制生长素向离区输导,使离区生长素含量少。 2.赤霉素和细胞分裂素也影响脱落,不过都不是直接的。 3.脱落的关键是果胶和胞壁等物质和可溶性糖之间的平衡。 4.保花保果的措施: (1)改善营养条件,使花果得到足够的光合产物。 (2)应用植物生长调节剂
改善营养条件,使花果得到足够的光合产物。 (2)应用植物生长调节剂.")
Similar presentations
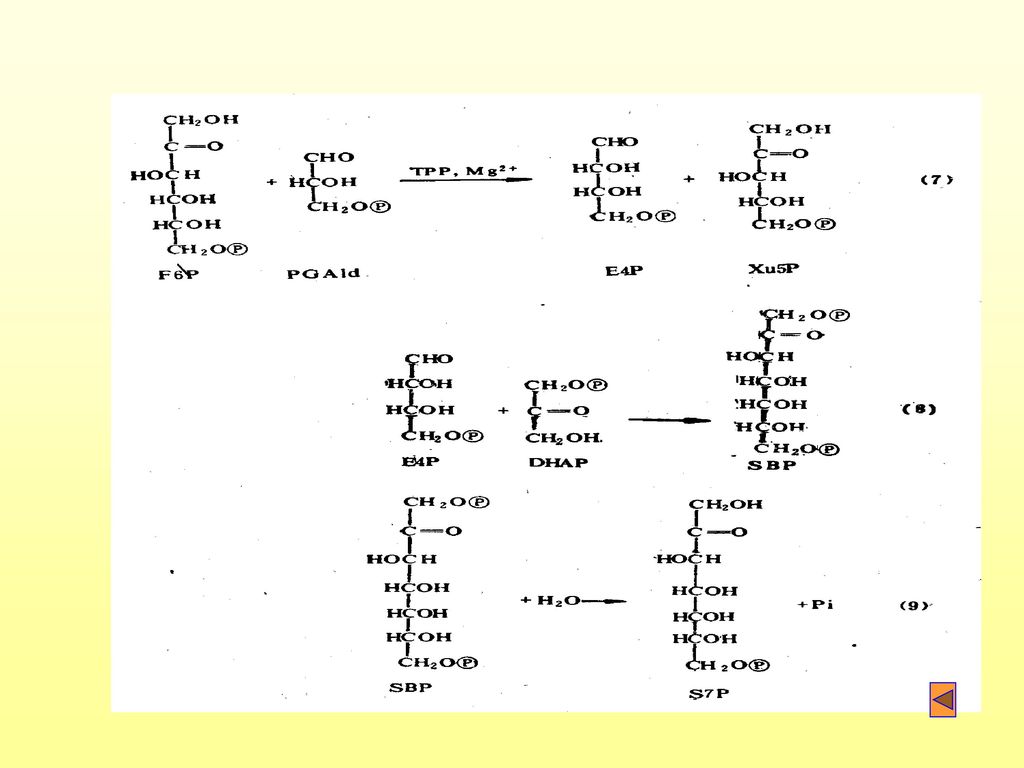
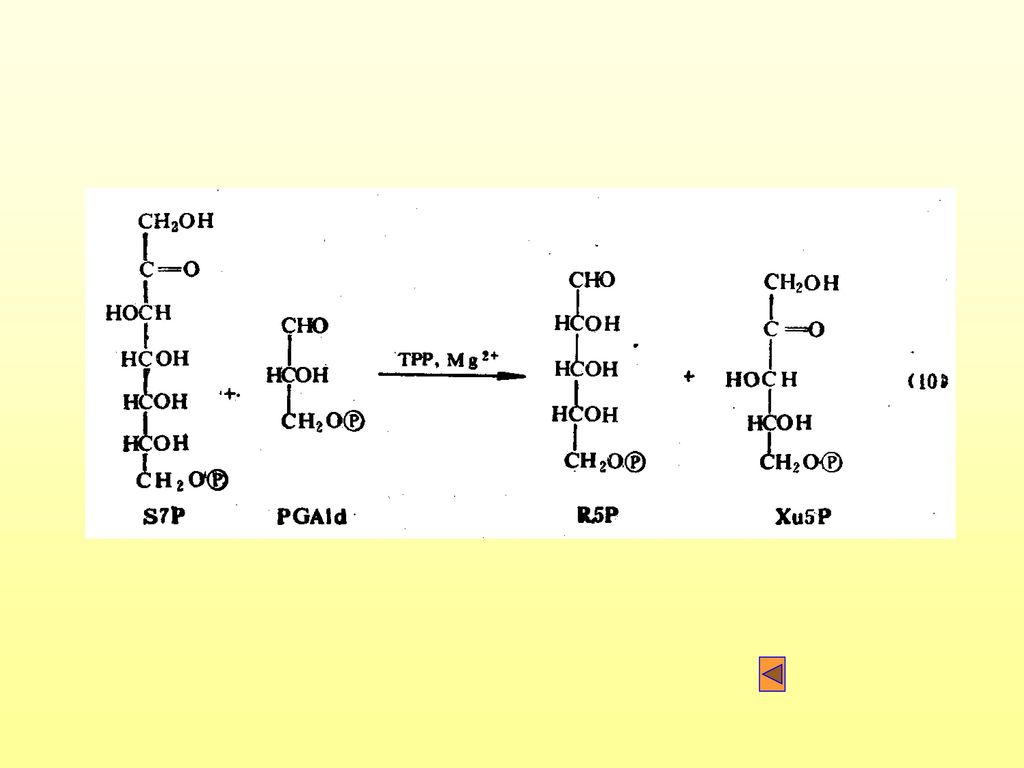
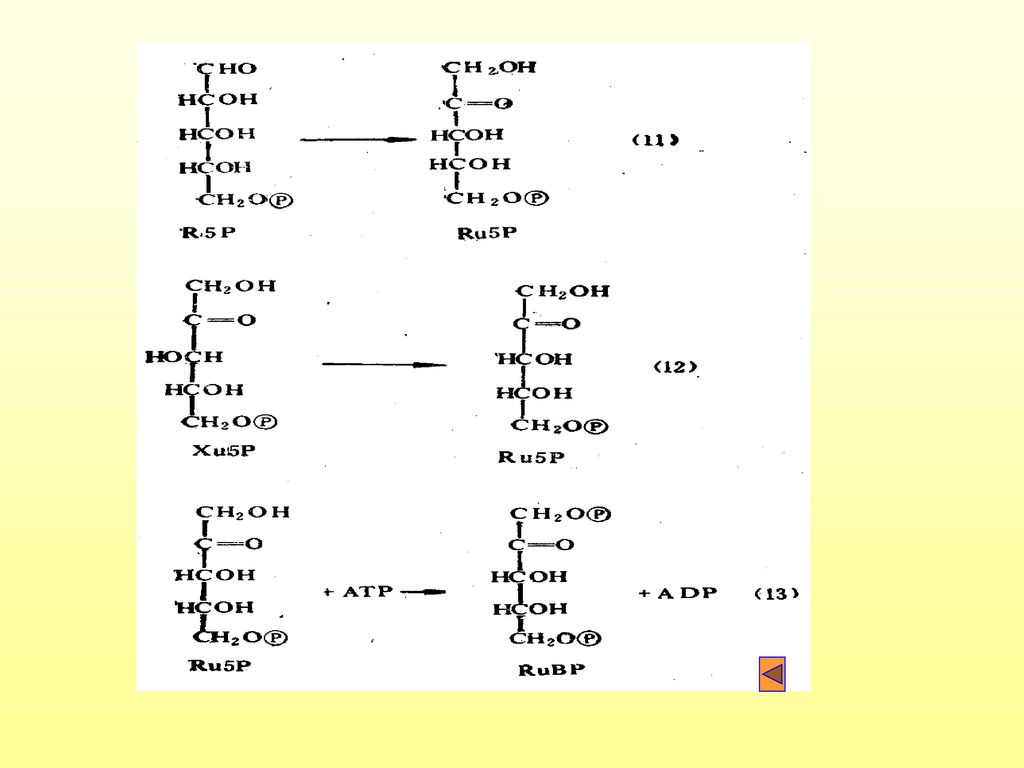
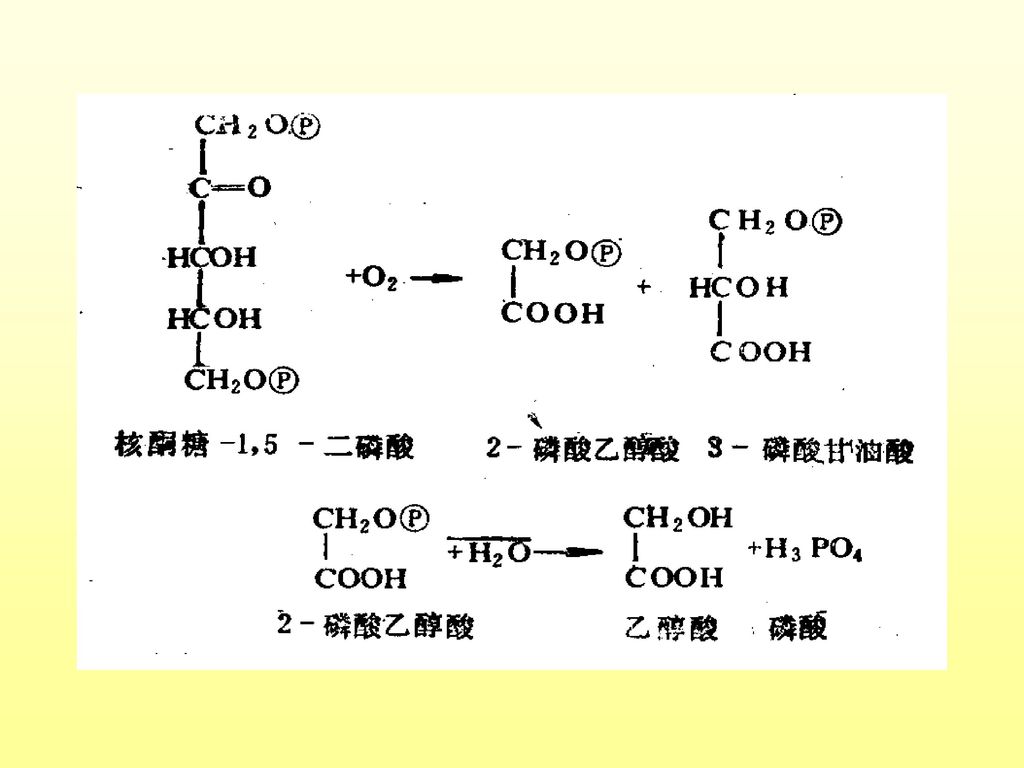
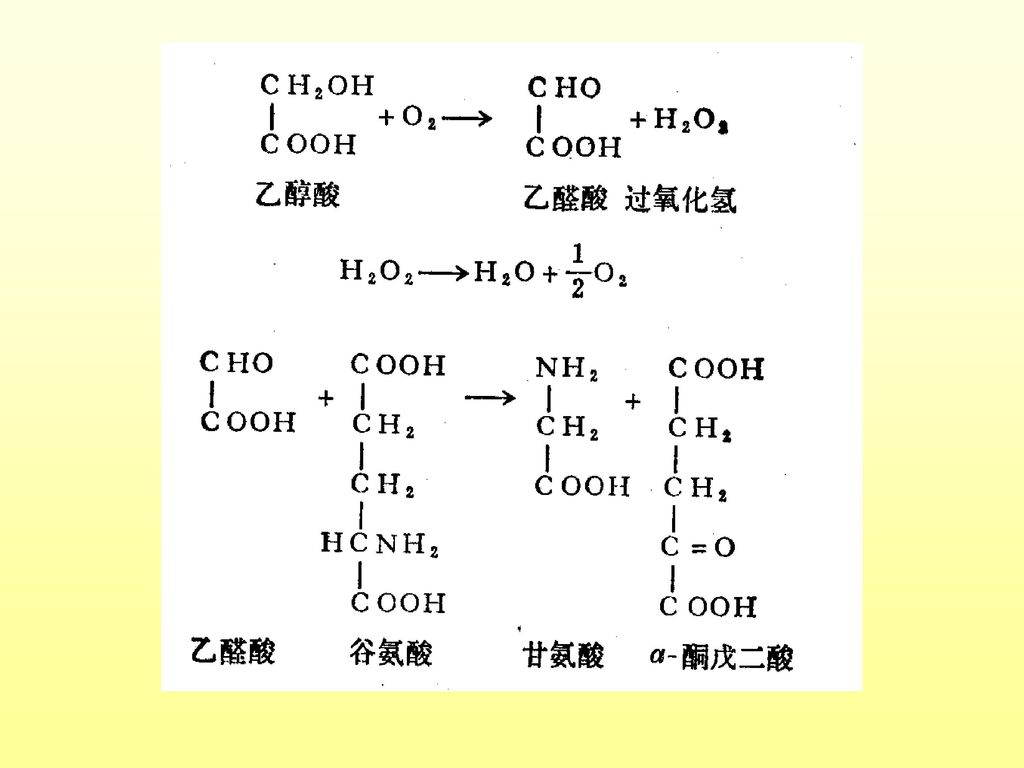

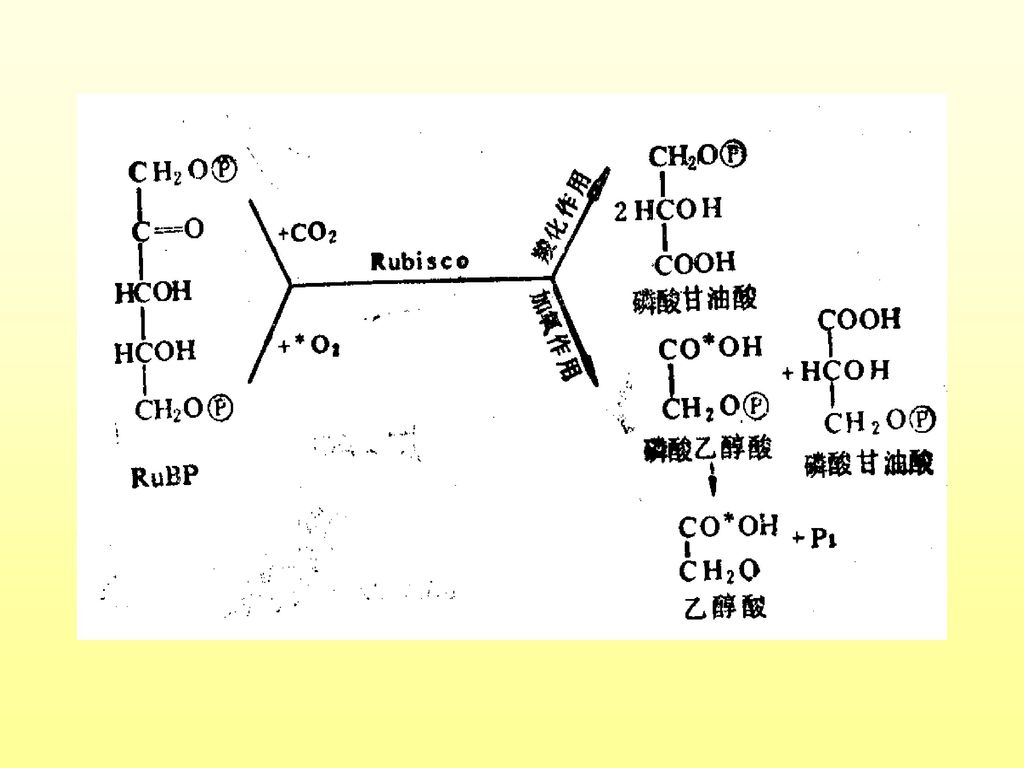








>")
蛋白质的降解: 外源蛋白的消化 内源性蛋白的选择性降解 (2)氨基酸的分解代谢:>")



.>")

 電子傳遞鏈 傳導概論>")



