Download presentation
Presentation is loading. Please wait.

1
第十章 细胞增殖 狭义的细胞生长:细胞大小的增加 广义的细胞生长:还包括细胞分裂和细胞分化
第十章 细胞增殖 狭义的细胞生长:细胞大小的增加 广义的细胞生长:还包括细胞分裂和细胞分化 细胞增殖:指细胞的增加或繁殖,是经过多次细胞分裂而使细胞数量快速膨胀的过程

2
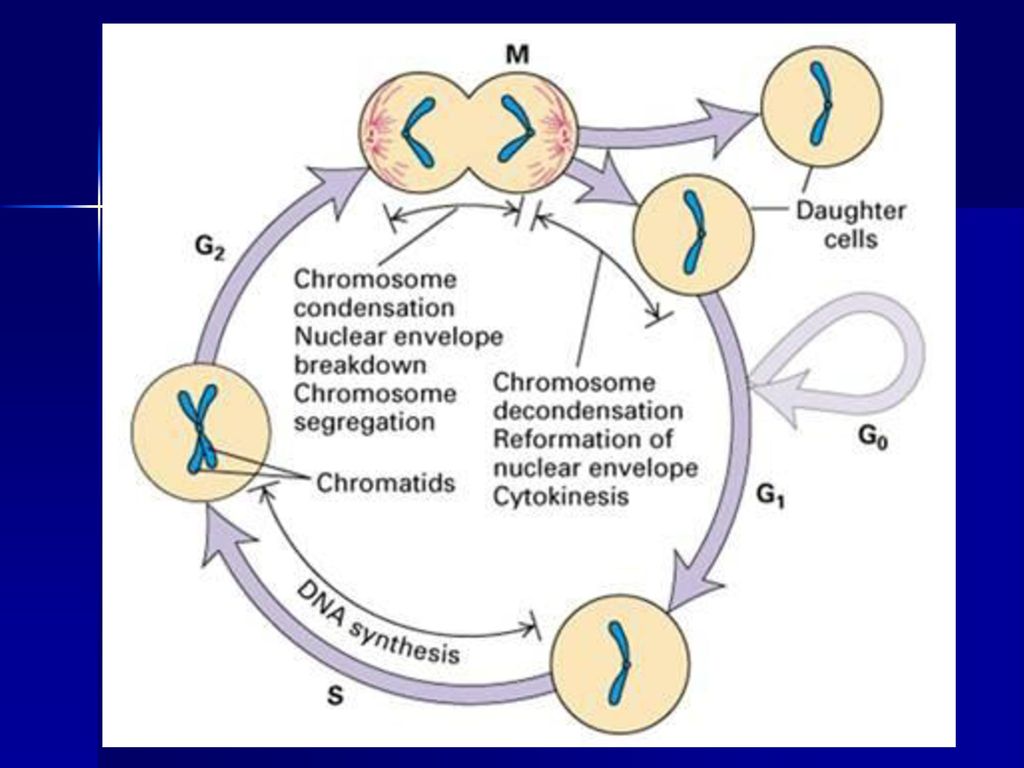
第一节 细胞周期的概念 一、什么是细胞周期 细胞周期指由细胞分裂结束到下一次细胞分裂结束所经历的过程,所需的时间叫细胞周期时间。
可分为四个阶段: ①G1期(gap1),指从有丝分裂完成到DNA复制之前的间隙时间; ②S期(synthesis phase),指DNA复制的时期,只有在这一时期H3-TdR才能掺入新合成的DNA中; ③G2期(gap2),指DNA复制完成到有丝分裂开始之前的一段时间; ④M期又称D期(mitosis or division),细胞分裂开始到结束。 分裂间期 分裂期
,指从有丝分裂完成到DNA复制之前的间隙时间; ②S期(synthesis phase),指DNA复制的时期,只有在这一时期H3-TdR才能掺入新合成的DNA中; ③G2期(gap2),指DNA复制完成到有丝分裂开始之前的一段时间; ④M期又称D期(mitosis or division),细胞分裂开始到结束。 分裂间期. 分裂期.")
4
① 周期中细胞(cycling cell):是指在细胞周期中连续运转
的细胞,有丝分裂活性高,又称为连续分裂细胞或可育细胞,如表皮生发层细胞、部分骨髓细胞、造血干细胞、雄性个体的生精细胞等。 ② 静止期细胞(quiescent cell):指的是暂时离开细胞周期,停止细胞分裂,去执行一定的生物学功能,但在适当的刺 激下可重新进入细胞周期的细胞,又称为G0期细胞或休眠细胞,如淋巴细胞、肝、肾细胞等。 ③ 终末分化细胞:指不可逆地脱离细胞周期,丧失分裂能力, 保持生理机能活动的细胞,又称终端细胞,如神经、肌肉、多形核细胞、红细胞等。
:指的是暂时离开细胞周期,停止细胞分裂,去执行一定的生物学功能,但在适当的刺. 激下可重新进入细胞周期的细胞,又称为G0期细胞或休眠细胞,如淋巴细胞、肝、肾细胞等。 ③ 终末分化细胞:指不可逆地脱离细胞周期,丧失分裂能力, 保持生理机能活动的细胞,又称终端细胞,如神经、肌肉、多形核细胞、红细胞等。")
5
细胞周期的时间长短与物种的细胞类型有关,如:小鼠十二指肠上皮细胞的周期为10小时,人类胃上皮细胞24小时,骨髓细胞18小时,培养的人成纤维细胞18小时,CHO细胞14小时,HeLa细胞21小时。不同类型细胞的G1长短不同,是造成细胞周期差异的主要原因。

6
细胞周期长短的测定 同种细胞间周期时间长短相似或相同;不同种类细胞间, 周期长短差别很大。
S+G2+M 的时间变化较小,细胞周期时间长短差别在G1期。 部分细胞的细胞周期没有G1、G2期。

7
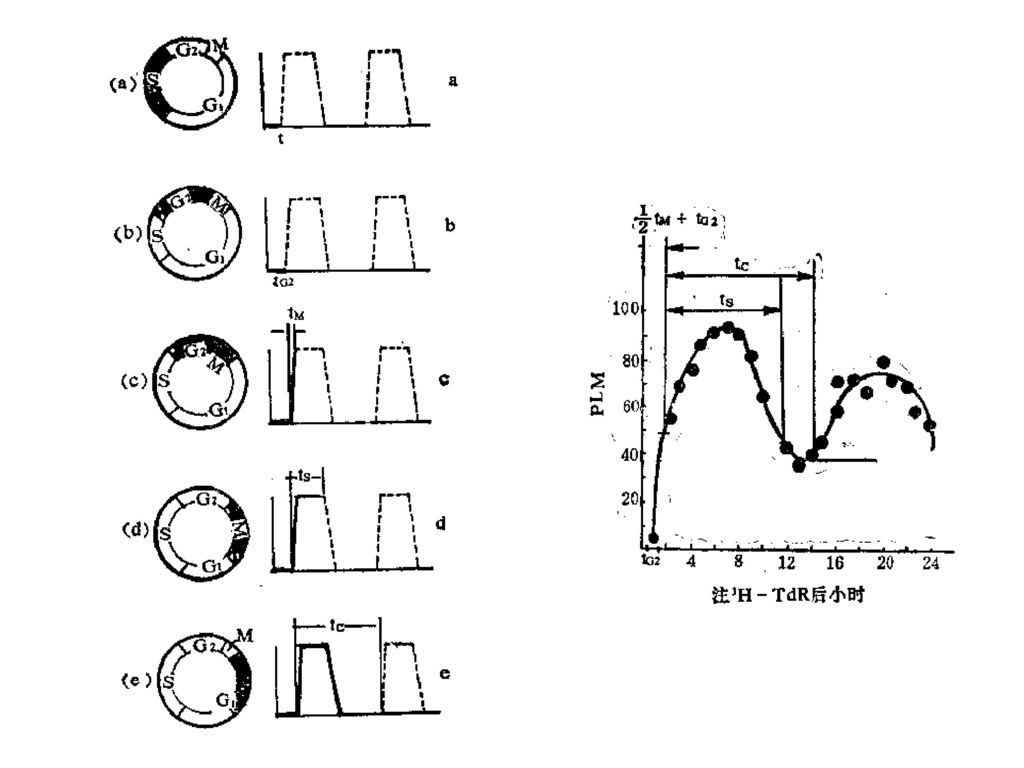
细胞周期长短的测定方法 mitoses,PLM) 流式细胞分选仪测定法(Flow Cytometry) 缩时摄像技术
标记有丝分裂百分数法(percentage labeled mitoses,PLM) 流式细胞分选仪测定法(Flow Cytometry) 缩时摄像技术
 流式细胞分选仪测定法(Flow Cytometry) 缩时摄像技术.")
8
标记有丝分裂百分数法(PLM) 原理:是一种常用的测定细胞周期时间的方法。其原理是对测定细胞进行脉冲标记、不同时间取样做细胞放射性自显影,找出正处于有丝分裂的分裂期细胞,计算其中带3H标记的细胞占有丝分裂细胞的百分数。 优点:可以测定细胞周期的总时间和各阶段的时间,结果明了易分析。 缺点:操作技术有一定的难度;具有一定的危险性;同位素的放射性逐渐衰减,误差较大。

9
测定原理: ① 待测细胞经3H-TdR标记后,所有S期细胞均被标记。 ② S期细胞经G2期才进入M期,所以一段时间内PLM=0。 ③开始出现标记M期细胞时,表示处于S期最后阶段的细胞,已渡过G2期,所以从PLM=0到出现PLM的时间间隔为TG2。 ④ S期细胞逐渐进入M期,PLM上升,到达到最高点的时候说明原先才进入M期的细胞,已完成M,进入G1期。所以从开始出现PLM到PLM达到最高点(≈100%)的时间间隔就是TM。 ⑤ 当PLM开始下降时,表明处于S期最初阶段的细胞也已进入M期,所以出现PLM到PLM又开始下降的一段时间等于TS。 ⑥ 从PLM出现到下一次PLM出现的时间间隔就等于TC,根据TC=TG1+TS+TG2+TM即可求出的TG1长度。
的时间间隔就是TM。 ⑤ 当PLM开始下降时,表明处于S期最初阶段的细胞也已进入M期,所以出现PLM到PLM又开始下降的一段时间等于TS。 ⑥ 从PLM出现到下一次PLM出现的时间间隔就等于TC,根据TC=TG1+TS+TG2+TM即可求出的TG1长度。")
11
细胞周期各阶段的时间与PLM的关系 Ts

12
二、细胞周期同步化 概念:细胞同步化是指在自然过程中发生的,或经人为处理 造成的细胞周期的同步化。 类型:自然同步化
人工同步化:选择同步化:有丝分裂选择法 细胞沉降分离法 诱导同步化:DNA合成阻断法 中期阻断法

13
(一)自然同步化 概念:自然界存在的细胞周期同步过程,称为自然同步化。 类型: 1.多核体
如粘菌只进行核分裂,而不发生胞质分裂,形成多核体。数量众多的核处于同一细胞质中,进行同步化分裂,使细胞核达108,体积达5~6cm。疟原虫也具有类似的情况。 2.某些水生动物的受精卵 如海胆卵可以同时授精,最初的3次细胞分裂是同步的,再如大量海参卵受精后,前9次细胞分裂都是同步化进行的。 3.增殖抑制解除后的同步分裂 如真菌的休眠孢子移入适宜环境后,它们一起发芽,同步分裂。

14
(二)人工同步化 1、选择同步化 1)有丝分裂选择法:使单层培养的细胞处于对数增殖期, 此时分裂活跃,MI高。有丝分裂细胞变圆隆起,与培养皿
培养液中,收集培养液,再加入新鲜培养液,依法继续收 集,则可获得一定数量的中期细胞。 — 优点:操作简单,同步化程度高,细胞不受药物的伤害。 — 缺点:获得的细胞数量少(分裂细胞约占1%~2%) 。
 。")
15
2)细胞沉降分离法:不同时期的细胞体积不同,而细胞在给定离心场中沉降的速度与其半径的平方成正比,因此可用离心的方法分离。
— 优点:可用于任何悬浮培养的细胞。 — 缺点:同步化程度低。

16
2、诱导同步化 1)DNA合成阻断法:选用DNA合成的抑制剂,可逆地抑制DNA合成,而不影响其他时期细胞的运转,最终可将细胞群阻断在S期或G/S交界处。5-氟脱氧尿嘧啶、羟基脲、高浓度ADR、GDR和TDR,均可抑制DNA合成使细胞同步化。其中高浓度TDR对S期细胞的毒性较小,因此常用TDR双阻断法诱导细胞同步化。 ?
DNA合成阻断法:选用DNA合成的抑制剂,可逆地抑制DNA合成,而不影响其他时期细胞的运转,最终可将细胞群阻断在S期或G/S交界处。5-氟脱氧尿嘧啶、羟基脲、高浓度ADR、GDR和TDR,均可抑制DNA合成使细胞同步化。其中高浓度TDR对S期细胞的毒性较小,因此常用TDR双阻断法诱导细胞同步化。")
17
TdR 阻断法阻滞细胞周期的原理是: TdR 是细胞DNA 合成不可缺少的前体, 但向培养基中加入过量的TdR, 能形成过量的三磷酸腺苷, 后者能反馈抑制其他核苷酸的磷酸化, 从而阻抑DNA合成。

18
DNA合成阻断法的操作过程 在细胞处于对数生长期的培养基中加入过量TDR,S期细胞被抑制,其它细胞继续运转,最后停在G1/S交界处。移去TDR,洗涤细胞并加入新鲜培养液、细胞又开始分裂。当释放时间大于TS时,所有细胞均脱离S期,再次加入过量TDR,细胞继续运转至G1/S交界处,被过量TDR抑制而停止。

19
关键:控制时间 第1次阻断时间相当于G2、M和G1期时间的总和或稍长,释放时间不短于S期时间,而小于G2+M+G1期时间,这样才能使所有位于G1/S期的细胞通过S期,而又不使沿周期前进最快的细胞进入下一个S期。第2次阻断时间同第1次,再释放。 HeLa细胞周期时间为21 h,其中G1期为10 h,S期为7 h,G2期为3 h,M期为1 h

20
优点:同步化程度高,适用于任何培养体系。可将几乎所
有的细胞同步化。 缺点:产生非均衡生长,个别细胞体积增大。 2)中期阻断法(M期抑制法):利用破坏微管的药物将细胞阻断在中期,常用的药物有秋水仙素和秋水仙酰胺,后者毒性较少。 优点:无非均衡生长现象;缺点:可逆性较差。
中期阻断法(M期抑制法):利用破坏微管的药物将细胞阻断在中期,常用的药物有秋水仙素和秋水仙酰胺,后者毒性较少。 优点:无非均衡生长现象;缺点:可逆性较差。")
21
细胞周期中各时期的特征 G1期:细胞表面有泡状物,染色质去凝集;蛋白质、糖 类、脂质等开始合成。
S期:细胞表面光滑,小泡和微绒毛明显减少,DNA复制, 中心体也在此期完成复制。 G2期:细胞表面许多微绒毛,合成一定数量的蛋白质和RNA。 M期:细胞分裂期,细胞呈球状,表面覆盖微绒毛。

22
植物细胞的细胞周期 植物细胞周期的时相和动物细胞的标准细胞周期相似,都含有G1、G2、S、M期。
植物细胞没有中心体,但细胞分裂时可以正常组装纺锤体; 植物细胞的形态不发生变化,以形成中间板的形式进行 胞质分裂。

23
植物细胞成膜体的形成

24
细菌的细胞周期 慢生长细菌细胞周期过程与真核细胞周期过程有一定相 似之处。其DNA复制之前的准备时间与G1期类似。分裂之
前的准备时间与G2期类似。再加上S期和M期,细菌的细 胞周期也基本具备四个时期。 细菌在快速生长情况下,如何协调快速分裂和最基本的 DNA复制速度之间的矛盾? 快速生长的细菌,在上一次细胞分裂结束时,细胞内的DNA经复制到一半进程,以保证迅速进行下一次分裂。

25
第二节 有丝分裂 一、细胞分裂的类型 细胞分裂(cell division)可分为无丝分裂(amitosis)、有丝分裂(mitosis)和减数分裂(meiosis)三种类型。 无丝分裂又称为直接分裂,由R. Remark(1841)首次发现于鸡胚血细胞。表现为细胞核伸长,从中部缢缩,然后细胞质分裂,其间不涉及纺锤体形成及染色体变化,故称为无丝分裂。无丝分裂不仅发现于原核生物,同时也发现于高等动植物,如植物的胚乳细胞、动物的胎膜,间充组织及肌肉细胞等等。
首次发现于鸡胚血细胞。表现为细胞核伸长,从中部缢缩,然后细胞质分裂,其间不涉及纺锤体形成及染色体变化,故称为无丝分裂。无丝分裂不仅发现于原核生物,同时也发现于高等动植物,如植物的胚乳细胞、动物的胎膜,间充组织及肌肉细胞等等。")
26
有丝分裂,又称为间接分裂,由W. Fleming (1882)年首次发现于动物及E
有丝分裂,又称为间接分裂,由W. Fleming (1882)年首次发现于动物及E. Strasburger(1880)年发现于植物。特点是有纺锤体染色体出现,子染色体被平均分配到子细胞,这种分裂方式普遍见于高等动植物。 减数分裂是指染色体复制一次而细胞连续分裂两次的分裂方式,是高等动植物配子体形成的分裂方式。
年首次发现于动物及E. Strasburger(1880)年发现于植物。特点是有纺锤体染色体出现,子染色体被平均分配到子细胞,这种分裂方式普遍见于高等动植物。 减数分裂是指染色体复制一次而细胞连续分裂两次的分裂方式,是高等动植物配子体形成的分裂方式。")
27
(二)有丝分裂(mitosis) 二、有丝分裂
有丝分裂过程是一个连续的过程,为了便于描述,人为的划分为六个时期:间期(interphase)、前期(prophase)、前中期(premetaphase)、中期(metaphase)、后期(anaphase)和末期(telophase)。其中间期包括G1期、S期和G2期,主要进行DNA复制等准备工作。
、前期(prophase)、前中期(premetaphase)、中期(metaphase)、后期(anaphase)和末期(telophase)。其中间期包括G1期、S期和G2期,主要进行DNA复制等准备工作。")
28
胞质分裂期 前中期

29
(一)前期 前期的主要事件是: ①染色质凝缩, ②分裂极确立与纺锤体开始形成, ③核仁解体, ④核膜消失。 前期最显著的特征是染色质通过螺旋化和折叠,变短变粗,形成光学显微镜下可以分辨的染色体,每条染色体包含2个染色单体。
前期 前期的主要事件是: ①染色质凝缩, ②分裂极确立与纺锤体开始形成, ③核仁解体, ④核膜消失。 前期最显著的特征是染色质通过螺旋化和折叠,变短变粗,形成光学显微镜下可以分辨的染色体,每条染色体包含2个染色单体。")
30
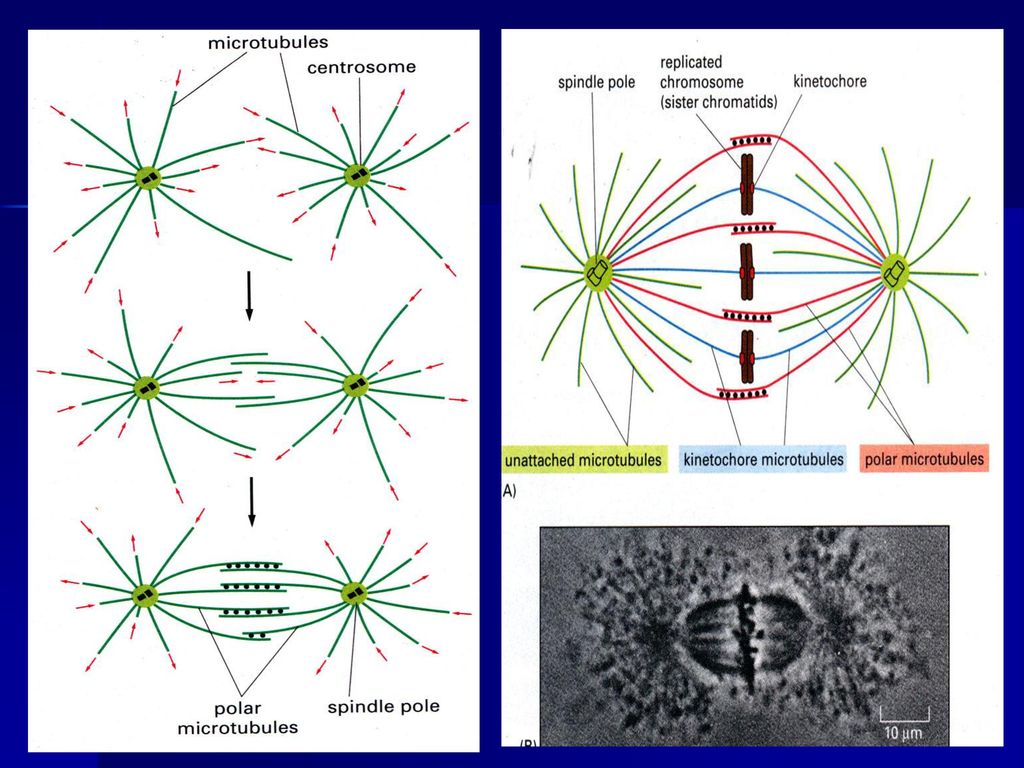
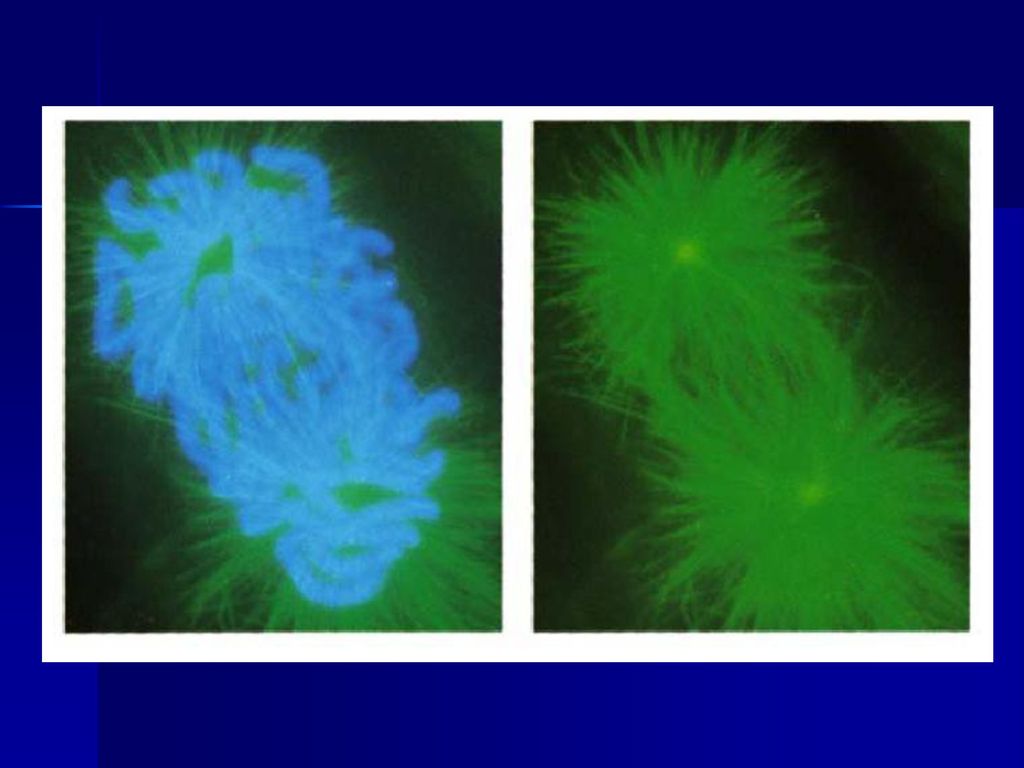
前期两个中心体向两极移动

31
早在S期两个中心粒已完成复制,在前期移向两极,两对中心粒之间形成纺锤体微管,当核膜解体时,两对中心粒已到达两极,并在两者之间形成纺锤体,纺锤体微管包括:
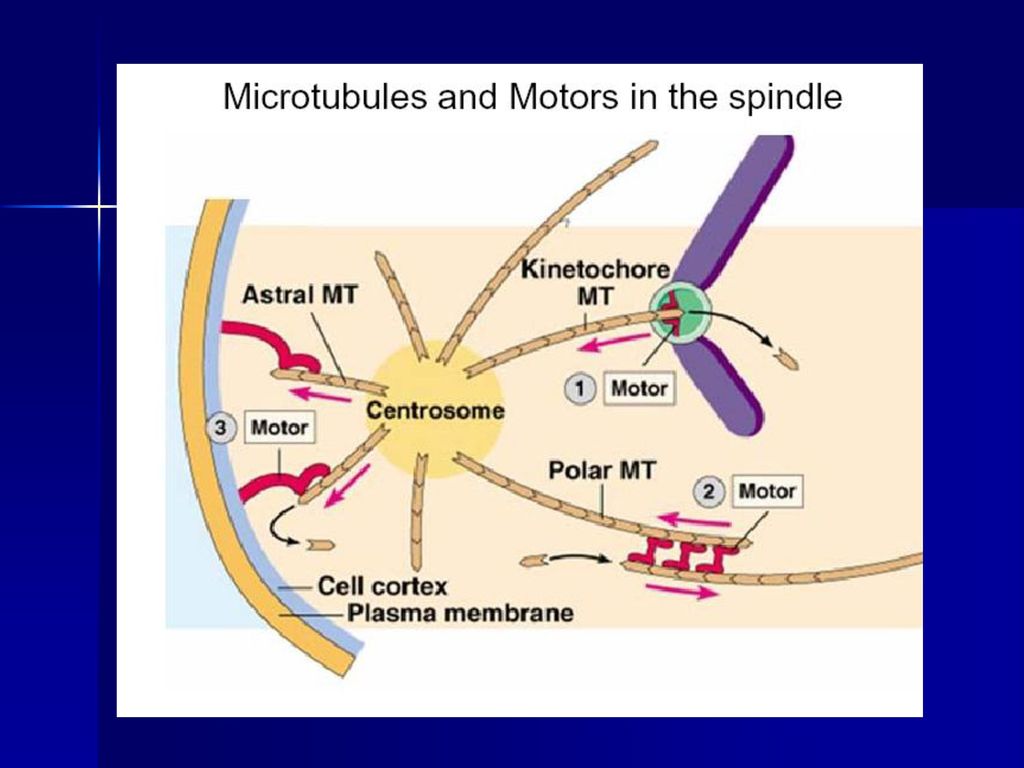
①着丝点微管(kinetochore mt):由中心体发出,连接在着丝点上,负责将染色体牵引到纺锤体上,着丝点上具有马达蛋白。 ②星体微管(astral mt):由中心体向外放射出,末端结合有分子马达,负责两极的分离,同时确定纺锤体纵轴的方向。 ③极体微管(polar mt或overlap mt):由中心体发出,在纺锤体中部重叠,重叠部位结合有分子马达,负责将两极推开。 有两类马达蛋白参与染色体、分裂极的分离,一类是dynein,另一类是kinesin。 植物没有中心粒和星体,其纺锤体叫作无星纺锤体,分裂极的确定机理尚不明确。
:由中心体发出,连接在着丝点上,负责将染色体牵引到纺锤体上,着丝点上具有马达蛋白。 ②星体微管(astral mt):由中心体向外放射出,末端结合有分子马达,负责两极的分离,同时确定纺锤体纵轴的方向。 ③极体微管(polar mt或overlap mt):由中心体发出,在纺锤体中部重叠,重叠部位结合有分子马达,负责将两极推开。 有两类马达蛋白参与染色体、分裂极的分离,一类是dynein,另一类是kinesin。 植物没有中心粒和星体,其纺锤体叫作无星纺锤体,分裂极的确定机理尚不明确。")
33
(二)前中期 指由核膜解体到染色体排列到赤道面(equatorial plane)这一阶段。纺锤体微管向细胞内部侵入,与染色体的着丝点结合。着丝点处的分子马达使染色体向微管的负端移动。在光镜下可以看到,此时染色体也就是既向一极移动也向另一极移动,是以振荡的方式移向纺锤体中部的。其原因是姊妹染色单体的着丝点都结合有微管和分子马达。
这一阶段。纺锤体微管向细胞内部侵入,与染色体的着丝点结合。着丝点处的分子马达使染色体向微管的负端移动。在光镜下可以看到,此时染色体也就是既向一极移动也向另一极移动,是以振荡的方式移向纺锤体中部的。其原因是姊妹染色单体的着丝点都结合有微管和分子马达。")
35
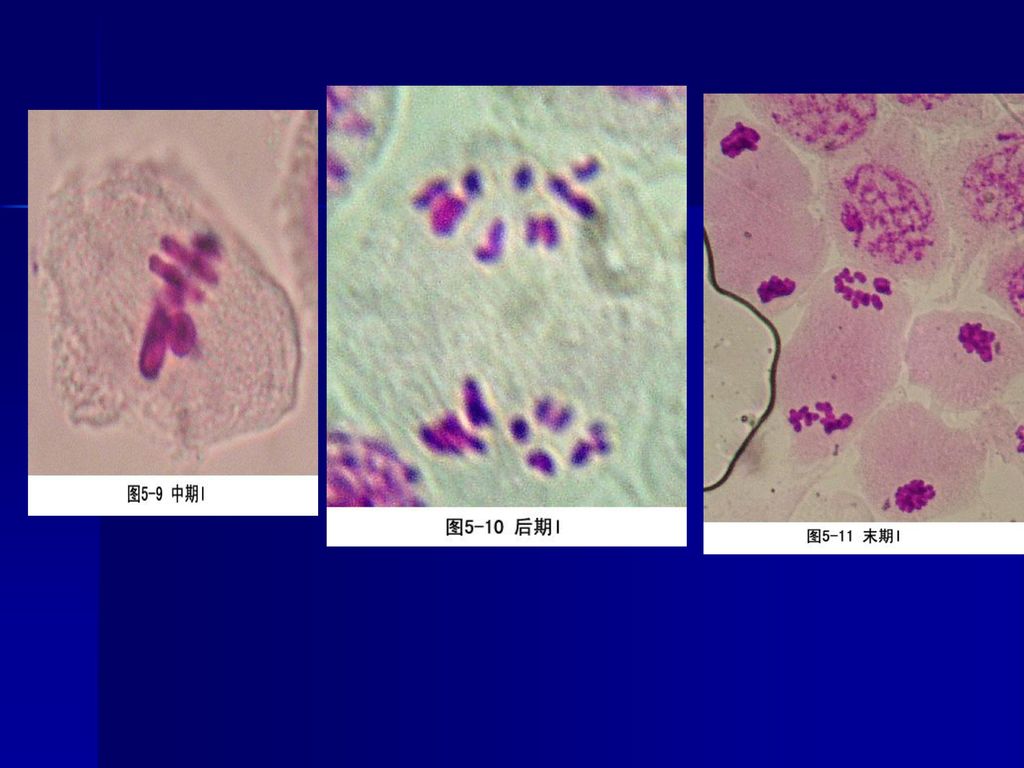
(三)中期 指从染色体排列到赤道面上,到姊妹染色单体开始分向两极的一段时间,纵向观动物染色体呈辐射状排列。染色体两边的牵引力就像拔河一样达到平衡。
中期 指从染色体排列到赤道面上,到姊妹染色单体开始分向两极的一段时间,纵向观动物染色体呈辐射状排列。染色体两边的牵引力就像拔河一样达到平衡。")
37
牵拉假说:染色体向赤道板方向运动,是由于动粒微管牵拉的结果。动粒微管越长,拉力越大,当来自两极的动粒微管的拉力相等时,染色体被稳定在赤道板上。
外推假说:染色体向赤道板方向的移动,是由于星体的排斥力将染色体外推的结果。染色体距离中心体越近,星体对染色体的外推力越强,当来自两极的推力达到平衡时,染色体被稳定在赤道板上。

38
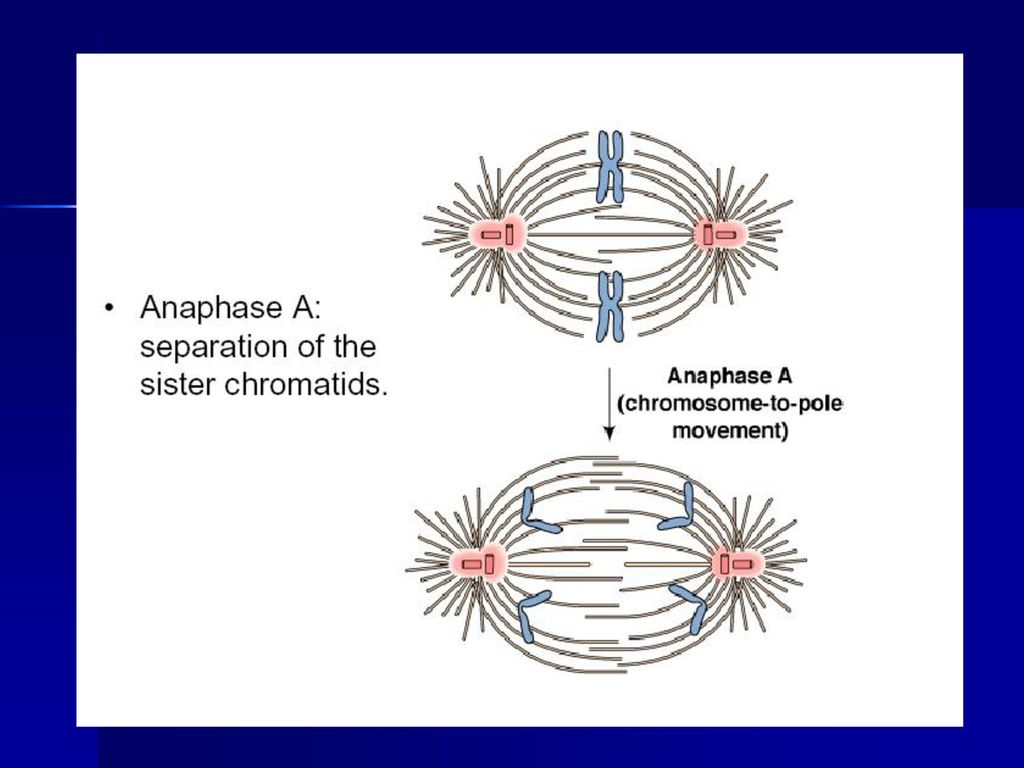
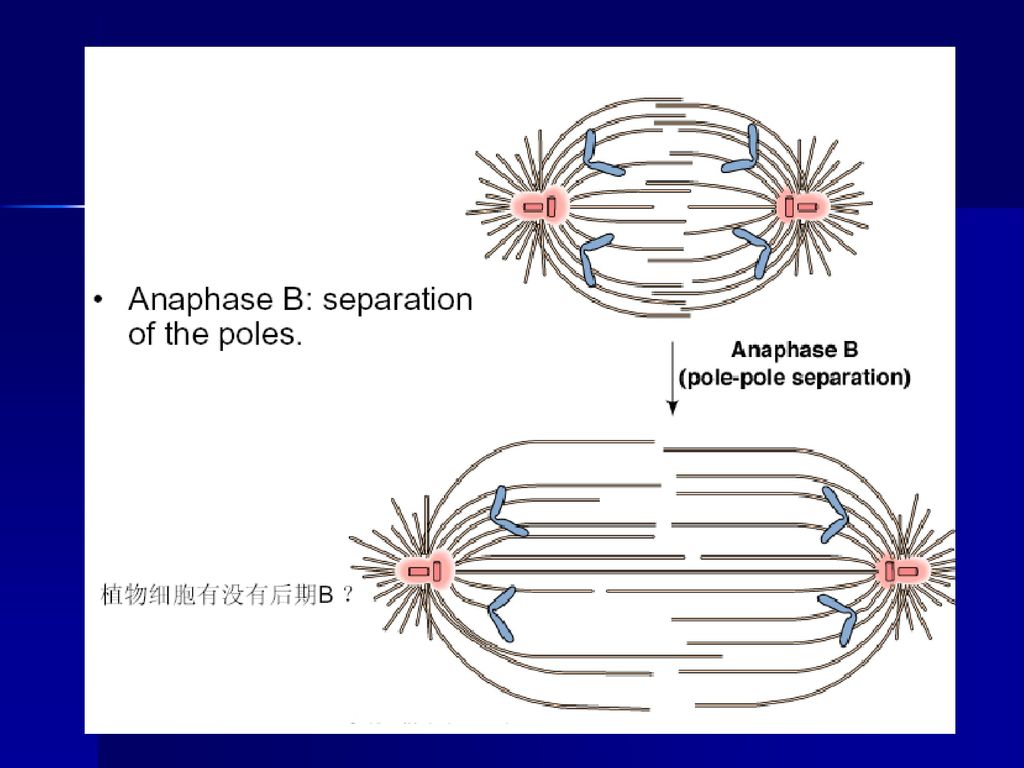
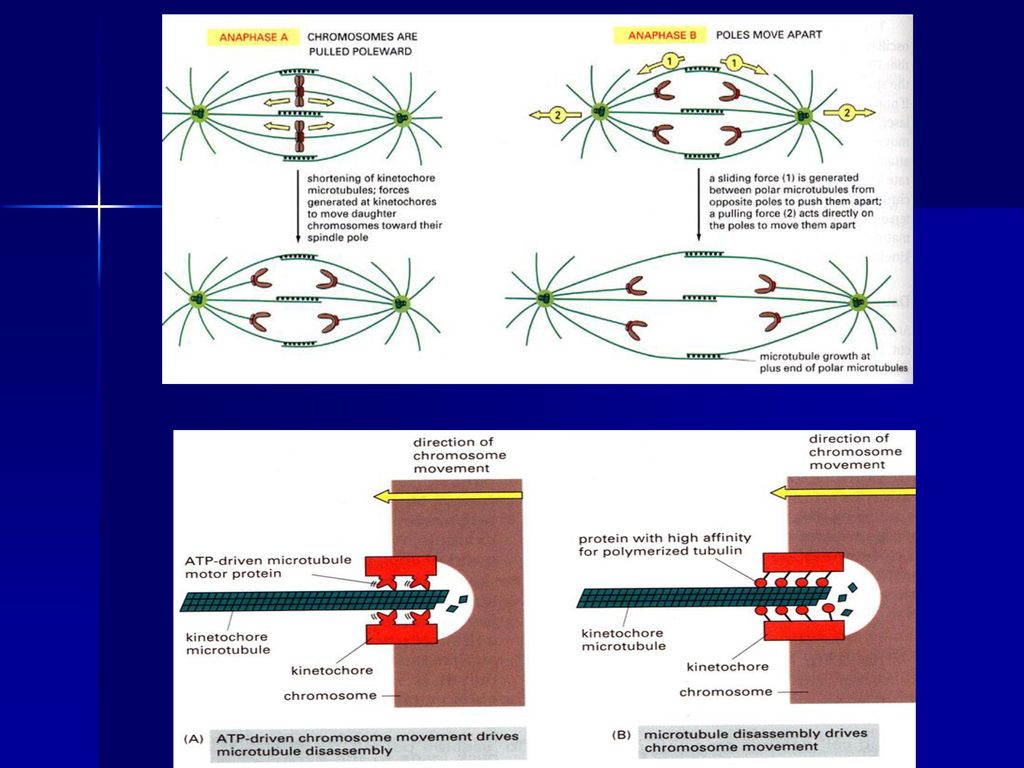
(四)后期 指姊妹染色单体分开并移向两极的时期,当子染色体到达两极后,标志这一时期结束。分为后期A和后期B。
后期 指姊妹染色单体分开并移向两极的时期,当子染色体到达两极后,标志这一时期结束。分为后期A和后期B。")
39
后期可以分为两个方面: ①后期A,指染色体向两极移动的过程。这是因为染色体着丝点微管在着丝点处去组装而缩短,在分子马达的作用下染色体向两极移动,体外实验证明即使在不存在ATP的情况下,染色体着丝点也有连接到正在去组装的微管上的能力,使染色体发生移动。 ②后期B,指两极间距离拉大的过程。这是因为一方面极体微管延长,结合在极体微管重叠部分的马达蛋白提供动力,推动两极分离,另一方面星体微管去组装而缩短,结合在星体微管正极的马达蛋白牵引两极距离加大。可见染色体的分离是在微管与分子马达的共同作用下实现的。

44
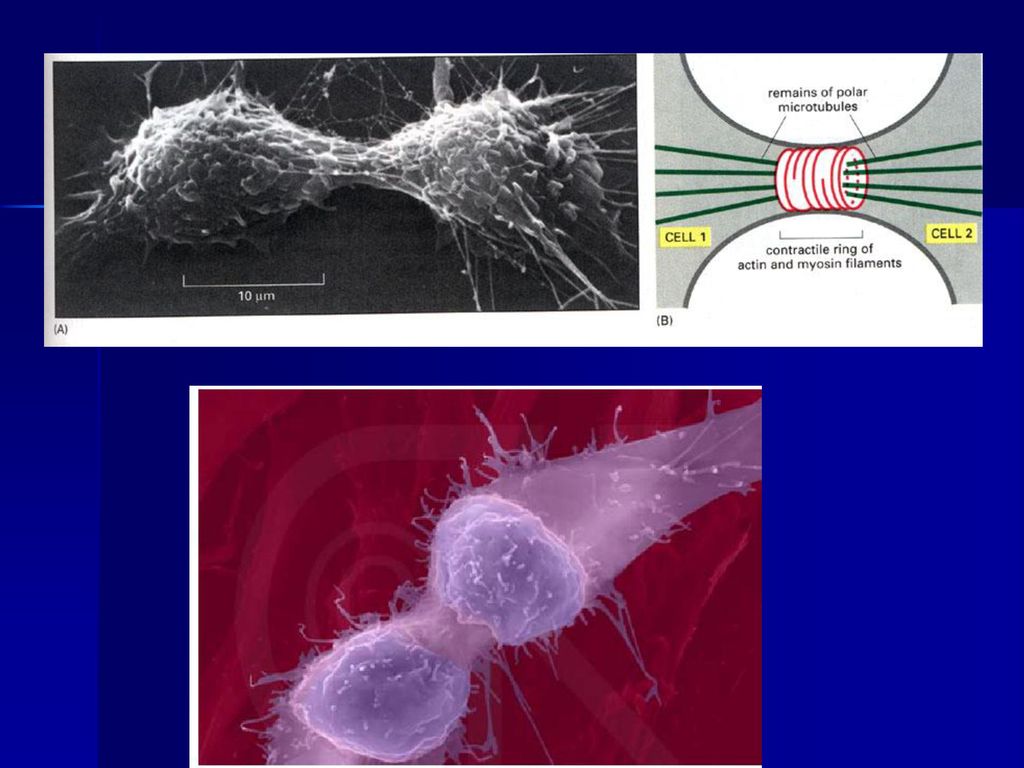
(五)末期 末期是从子染色体到达两极,至形成两个新细胞为 止的时期。末期涉及子核的形成和胞质分裂两个方面。
末期 末期是从子染色体到达两极,至形成两个新细胞为 止的时期。末期涉及子核的形成和胞质分裂两个方面。")
45
1、子核的形成 末期子核的形成,大体经历了与前期相反的过程,即染色体解聚缩,核仁出现和核膜重新形成。核仁由染色体上的核仁组织中心形成(NORs),几个NORS共同组成一个大的核仁,因此核仁的数目通常比NORs的数目要少。 前期核膜解体后,核纤层蛋白B与核膜残余小泡结合,末期核纤层蛋白B去磷酸化,介导核膜的重新装配。

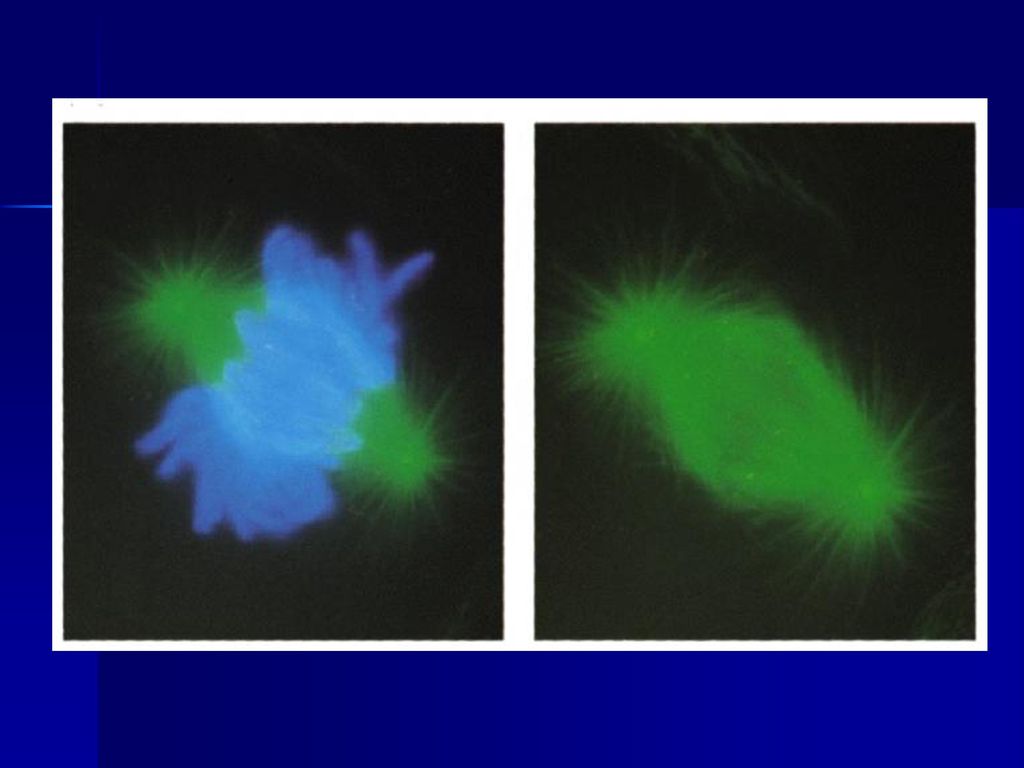
46
2、胞质分裂 虽然核分裂与胞质分裂(cytokinesis)是相继发生的,但属于两个分离的过程,例如大多数昆虫的卵,核可进行多次分裂而无胞质分裂,某些藻类的多核细胞可长达数尺,以后胞质才分裂形成单核细胞。 动物细胞的胞质分裂是以形成收缩环的方式完成的,收缩环在后期形成,由大量平行排列的肌动蛋白和结合在上面的myosin II等成分组成,用细胞松驰素及肌动蛋白和肌球蛋白抗体处理均能抑制收缩环的形成。不难想象胞质收缩环工作原理和肌肉收缩时一样的。

47
动细胞的胞质收缩环

48
胞质分裂(cytokinisis)
")
50
动物胞质分裂的另一特点是形成中体。末期纺锤体开始瓦解消失,但在纺锤体的中部微管数量增加,其中掺杂有高电子密度物质和囊状物,这一结构称为中体。在胞质分裂中的作用尚不清楚。
植物胞质分裂的机制不同于动物,后期或末期两极处微管消失,中间微管保留,并数量增加,形成桶状的成膜体(phragmoplast)。来自于高尔基体的囊泡沿微管转运到成膜体中间,融合形成细胞板。囊泡内的物质沉积为初生壁和中胶层,囊泡膜形成新的质膜,由于两侧质膜来源于共同的囊泡,因而膜间有许多连通的管道,形成胞间连丝。源源不断运送来的囊泡向细胞板融合,使细胞板扩展,形成完整的细胞壁,将子细胞一分为二。
。来自于高尔基体的囊泡沿微管转运到成膜体中间,融合形成细胞板。囊泡内的物质沉积为初生壁和中胶层,囊泡膜形成新的质膜,由于两侧质膜来源于共同的囊泡,因而膜间有许多连通的管道,形成胞间连丝。源源不断运送来的囊泡向细胞板融合,使细胞板扩展,形成完整的细胞壁,将子细胞一分为二。")
51
植物细胞成膜体的形成

52
第三节 减数分裂 减数分裂(Meiosis)的特点是DNA复制一次,而细胞连续分裂两次,形成单倍体的精子和卵子,通过受精作用又恢复二倍体,减数分裂过程中同源染色体间发生交换,使配子的遗传多样化,增加了后代的适应性,因此减数分裂不仅是保证生物种染色体数目稳定的机制,同且也是物种适应环境变化不断进化的机制。
的特点是DNA复制一次,而细胞连续分裂两次,形成单倍体的精子和卵子,通过受精作用又恢复二倍体,减数分裂过程中同源染色体间发生交换,使配子的遗传多样化,增加了后代的适应性,因此减数分裂不仅是保证生物种染色体数目稳定的机制,同且也是物种适应环境变化不断进化的机制。")
53
减数分裂可分为3种主要类型: 1、配子减数分裂(gametic meiosis),也叫终端减数分裂(terminal meiosis),其特点是减数分裂和配子的发生紧密联系在一起,在雄性脊椎动物中,一个精母细胞经过减数分裂形成4个精细胞,后者在经过一系列的变态发育,形成成熟的精子。在雌性脊椎动物中,一个卵母细胞经过减数分裂形成1个卵细胞和2-3个极体。 2、孢子减数分裂(sporic meiosis),也叫中间减数分裂(intermediate meiosis),见于植物和某些藻类。其特点是减数分裂和配子发生没有直接的关系,减数分裂的结果是形成单倍体的配子体(小孢子和大孢子)。小孢子再经过两次有次分裂形成包含一个营养核和两个雄配子(精子)的成熟花粉(雄配子体),大孢子经过三次有丝分裂形成胚囊(雌配子体),内含一个卵核、两个极核、3个反足细胞和两个助细胞。
,也叫终端减数分裂(terminal meiosis),其特点是减数分裂和配子的发生紧密联系在一起,在雄性脊椎动物中,一个精母细胞经过减数分裂形成4个精细胞,后者在经过一系列的变态发育,形成成熟的精子。在雌性脊椎动物中,一个卵母细胞经过减数分裂形成1个卵细胞和2-3个极体。 2、孢子减数分裂(sporic meiosis),也叫中间减数分裂(intermediate meiosis),见于植物和某些藻类。其特点是减数分裂和配子发生没有直接的关系,减数分裂的结果是形成单倍体的配子体(小孢子和大孢子)。小孢子再经过两次有次分裂形成包含一个营养核和两个雄配子(精子)的成熟花粉(雄配子体),大孢子经过三次有丝分裂形成胚囊(雌配子体),内含一个卵核、两个极核、3个反足细胞和两个助细胞。")
54
3、合子减数分裂(zygotic meiosis),也叫初始减数分裂(initial meiosis),仅见于真菌和某些原核生物,减数分裂发生于合子形成之后,形成单倍体的孢子,孢子通过有丝分裂产生新的单倍体后代。
,也叫初始减数分裂(initial meiosis),仅见于真菌和某些原核生物,减数分裂发生于合子形成之后,形成单倍体的孢子,孢子通过有丝分裂产生新的单倍体后代。")
55
此外某些生物还具有体细胞减数分裂(somatic meiosis)现象,如在蚊子幼虫的肠道中,有一些由核内有丝分裂形成的多倍体细胞(可高达32X),在蛹期又通过减数分裂降低了染色体倍性,增加了细胞数目。 减数分裂由紧密连接的两次分裂构成。通常减数分裂I分离的是同源染色体,所以称为异型分裂(heterotypic division)或减数分裂(reductional division)。减数分裂II分离的是姊妹染色体,类似于有丝分裂,所以称为同型分裂(homotypic division)或均等分裂(equational division)。和有丝分裂一样为了描述方便将减数分裂分为几个期和亚期
或减数分裂(reductional division)。减数分裂II分离的是姊妹染色体,类似于有丝分裂,所以称为同型分裂(homotypic division)或均等分裂(equational division)。和有丝分裂一样为了描述方便将减数分裂分为几个期和亚期.")
56
黑圆角蝉的精子发生

57
减数分裂模式图

58
一、间期 有丝分裂细胞在进入减数分裂之前要经过一个较长的间期,称前减数分裂间期(premeiotic interphase)或前减数分裂期(premeiosis)。 前减数分裂期也可分为G1期、S期和G2期,在G1期和S期把麝香百合的花粉每细胞在体外培养,则发现细胞进行有丝分裂,将G2晚期的细胞在体外培养则向减数分裂进行,说明G2期是有丝分裂向减数分裂转化的关键时期。 和有丝分裂不同的是,DNA不仅在S期合成,而且也在前期合成一小部分。D. E. Wimber和 W. Prensky(1963)认为合线期-粗线期合成大约2%的DNA。Y. Hotta等人(1966)在百合属(Lilium)和延龄草属(Trillium)中发现,粗线期合成大约0.3%的DNA。称为合线期DNA(zyg-DNA)或粗线期DNA(P-DNA)。这些DNA的合成可能与联会复合体的形成有关。
认为合线期-粗线期合成大约2%的DNA。Y. Hotta等人(1966)在百合属(Lilium)和延龄草属(Trillium)中发现,粗线期合成大约0.3%的DNA。称为合线期DNA(zyg-DNA)或粗线期DNA(P-DNA)。这些DNA的合成可能与联会复合体的形成有关。")
59
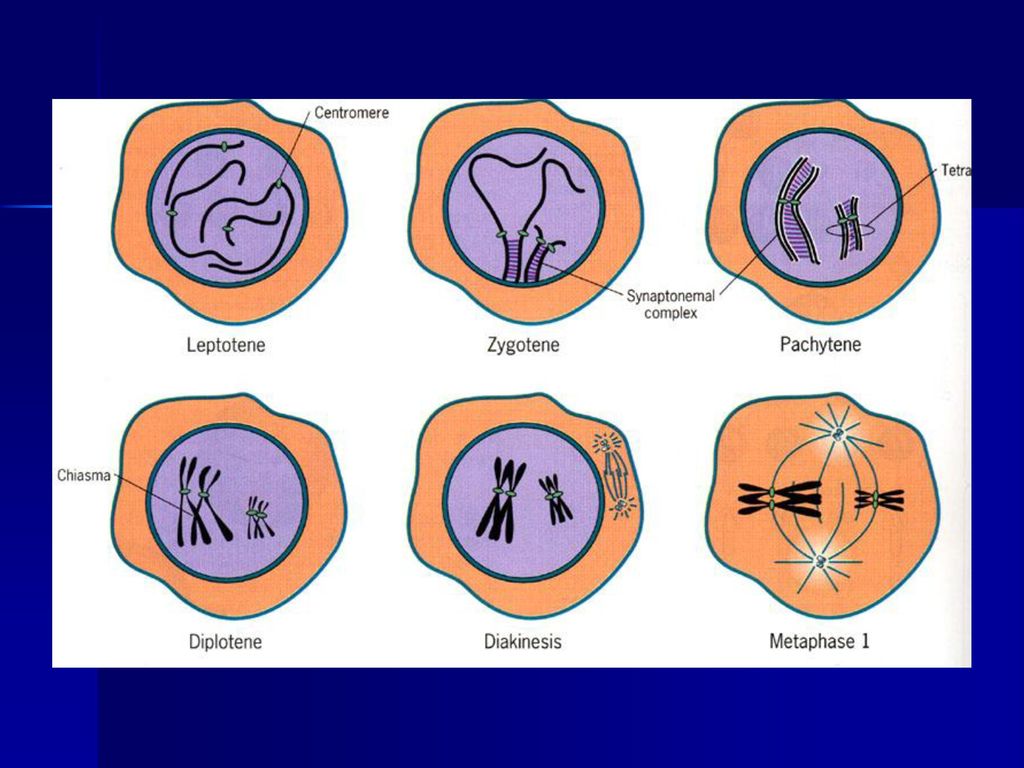
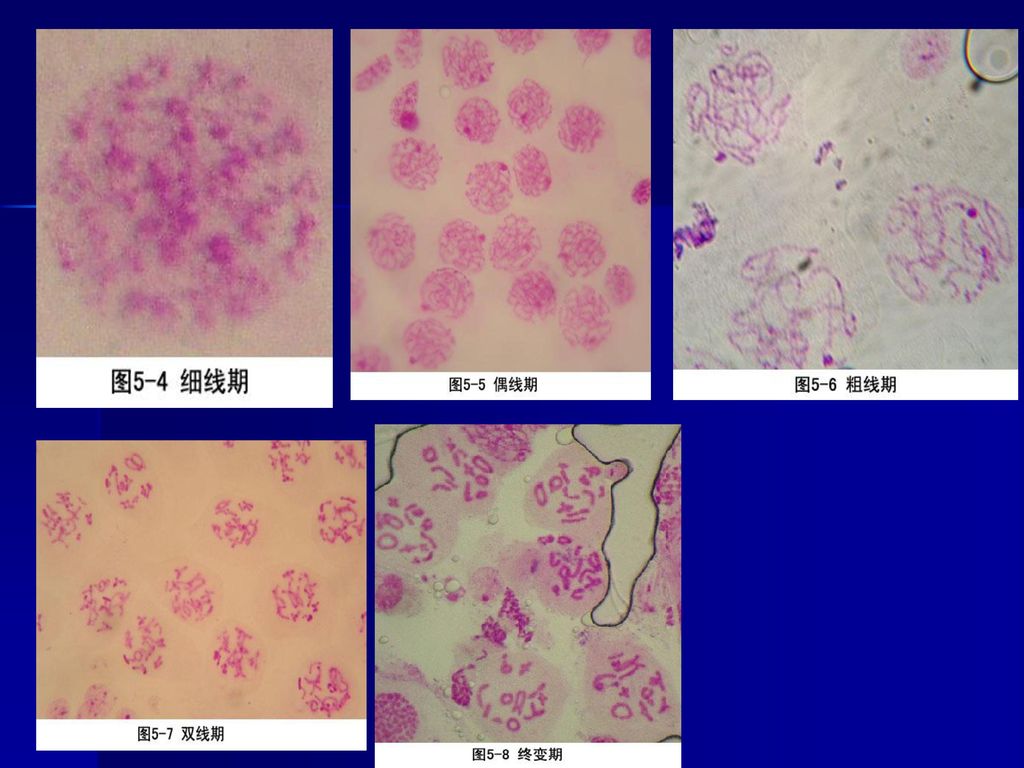
二、分裂期 (一)减数分裂I 1、前期I 减数分裂的特殊过程主要发生在前期I,通常人为划分为5个时期: ①细线期(leptotene) ②合线期(zygotene) ③粗线期(pachytene) ④双线期(diplotene) ⑤终变期(diakinesis)。
 ⑤终变期(diakinesis)。")
60
1)细线期: 染色体呈细线状,具有念珠状的染色粒。持续时间最长,占减数分裂周期的40%。细线期虽然染色体已经复制,但光镜下分辨不出两条染色单体。由于染色体细线交织在一起,偏向核的一方,所以又称为凝线期(synizesis),在有些物种中表现为染色体细线一端在核膜的一侧集中,另一端放射状伸出,形似花束,称为花束期(bouquet stage)。
细线期: 染色体呈细线状,具有念珠状的染色粒。持续时间最长,占减数分裂周期的40%。细线期虽然染色体已经复制,但光镜下分辨不出两条染色单体。由于染色体细线交织在一起,偏向核的一方,所以又称为凝线期(synizesis),在有些物种中表现为染色体细线一端在核膜的一侧集中,另一端放射状伸出,形似花束,称为花束期(bouquet stage)。")
61
2)合线期:持续时间较长,占有丝分裂周期的20%。亦称偶线期,是同源染色体配对的时期,这种配对称为联会(synapsis)。这一时期同源染色体间形成联会复合体(synaptonemal complex,SC)。在光镜下可以看到两条结合在一起的染色体,称为二价体(bivalent)。每一对同源染色体都经过复制,含四个染色单体,所以又称为四分体(tetrad)。
合线期:持续时间较长,占有丝分裂周期的20%。亦称偶线期,是同源染色体配对的时期,这种配对称为联会(synapsis)。这一时期同源染色体间形成联会复合体(synaptonemal complex,SC)。在光镜下可以看到两条结合在一起的染色体,称为二价体(bivalent)。每一对同源染色体都经过复制,含四个染色单体,所以又称为四分体(tetrad)。")
62
联会复合体 :联会复合体(synaptonemal complex, SC)是减数分裂合线期两条同源染色体之间形成的一种结构,它与染色体的配对,交换和分离密切相关。
SC是同源染色体间形成的梯子样的结构。在电镜下观察,两侧是约40nm的侧生组分(lateral element),电子密度很高,两侧之间为宽约100nm的中间区(intermediate space),在电镜下是明亮区,在中间区的中央为中央组分(central element),宽约30nm。侧生组分与中央组分之间有横向排列的粗约7~10nm的SC纤维,使SC外观呈梯子状
,电子密度很高,两侧之间为宽约100nm的中间区(intermediate space),在电镜下是明亮区,在中间区的中央为中央组分(central element),宽约30nm。侧生组分与中央组分之间有横向排列的粗约7~10nm的SC纤维,使SC外观呈梯子状.")
63
长期以来人们认为SC将同源染色体组织在一起,使伸入SC的DNA之间产生重组,但实验证明不仅SC的形成晚于基因重组的启动,而且基因突变不能形成SC的酵母中,同源染色体间照样可以发生交换。现在一般认为它与同源染色体间交换的完成有关。 在磷钨酸染色的SC中央,还可以看到呈圆形或椭圆形的重组节(recombination nodules,RNs),RNs是同源染色体发生交叉的部位,RNs上有基因交换所需要的酶。 从形态学来看,SC形成合线期,成熟于粗线期,并存在数天,消失于双线期。联会复合体的形成与合线期DNA(Zyg-DNA)有关,在细线期或合线期加入DNA合成抑制剂,则抑制SC的形成
,RNs是同源染色体发生交叉的部位,RNs上有基因交换所需要的酶。 从形态学来看,SC形成合线期,成熟于粗线期,并存在数天,消失于双线期。联会复合体的形成与合线期DNA(Zyg-DNA)有关,在细线期或合线期加入DNA合成抑制剂,则抑制SC的形成.")
64
3)粗线期:持续时间长达数天,此时染色体变短,结合紧密,在光镜下只在局部可以区分同源染色体,这一时期同源染色体的非姊妹染色单体之间发生交换的时期。在果蝇粗线期SC上具有与SC宽度相近的电子致密球状小体,称为重组节,与DNA的重组有关。
粗线期:持续时间长达数天,此时染色体变短,结合紧密,在光镜下只在局部可以区分同源染色体,这一时期同源染色体的非姊妹染色单体之间发生交换的时期。在果蝇粗线期SC上具有与SC宽度相近的电子致密球状小体,称为重组节,与DNA的重组有关。")
65
4)双线期:联会的同源染色体相互排斥、开始分离,但在交叉点(chiasma)上还保持着联系。双线期染色体进一步缩短,在电镜下已看不到联会复合体。
交叉的数目和位置在每个二价体上并非是固定的,而随着时间推移,向端部移动,这种移动现象称为端化(terminalization),端化过程一直进行到中期。 植物细胞双线期一般较短,但在许多动物中双线期停留的时间非常长,人的卵母细胞在五个月胎儿中已达双线期,而一直到排卵都停在双线期,排卵年龄大约在12-50岁之间。成熟的卵细胞直到受精后,才迅速完成两次分裂,形成单倍体的卵核。 在鱼类、两栖类、爬行类、鸟类以及无脊椎动物的昆虫中,双线期的二价体解螺旋而形成灯刷染色体,这一时期是卵黄积累的时期。
,端化过程一直进行到中期。 植物细胞双线期一般较短,但在许多动物中双线期停留的时间非常长,人的卵母细胞在五个月胎儿中已达双线期,而一直到排卵都停在双线期,排卵年龄大约在12-50岁之间。成熟的卵细胞直到受精后,才迅速完成两次分裂,形成单倍体的卵核。 在鱼类、两栖类、爬行类、鸟类以及无脊椎动物的昆虫中,双线期的二价体解螺旋而形成灯刷染色体,这一时期是卵黄积累的时期。")
66
5)终变期:二价体显著变短,并向核周边移动,在核内均匀散开。所以是观察染色体的良好时期。
由于交叉端化过程的进一步发展,故交叉数目减少,通常只有一至二个交叉。终变期二价体的形状表现出多样性,如V形、O形等。 核仁此时开始消失,核被膜解体,但有的植物,如玉米,在终变期核仁仍然很显著。

69
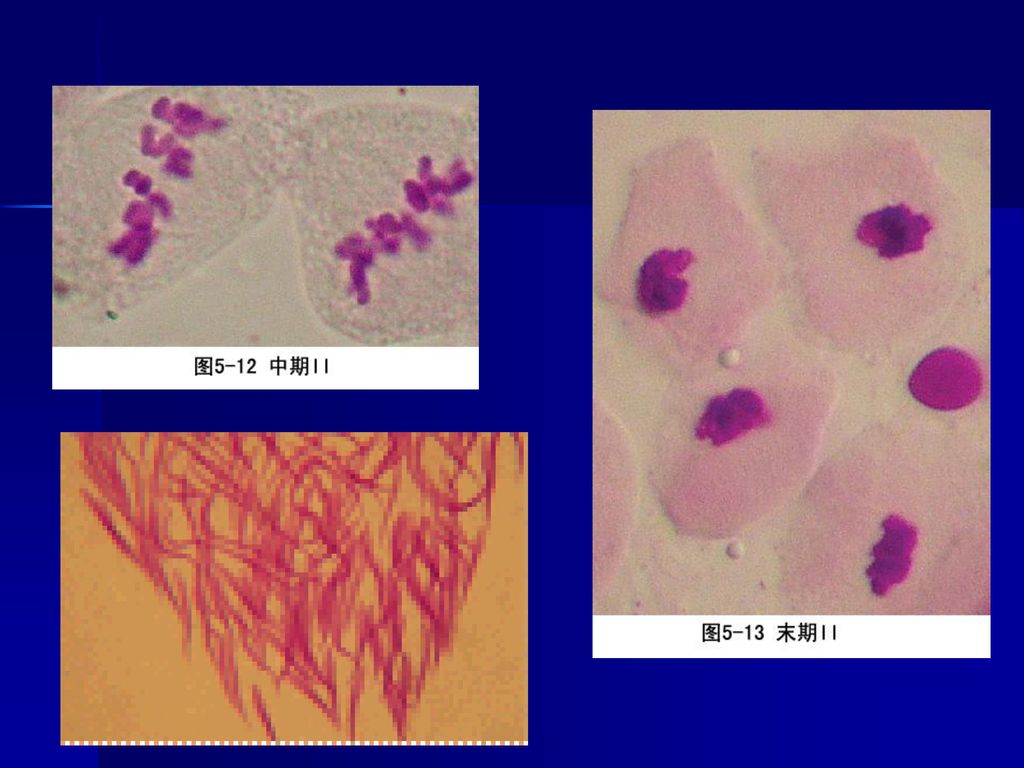
2、中期I 核仁消失,核被膜解体,标志进入中期I,中期I的主要特点是染色体排列在赤道面上。每个二价体有4个着丝粒、姊妹染色单位的着丝粒定向于纺锤体的同一极,故称联合定向(co-orientation)。
。")
70
3、后期I 二价体中的两条同源染色体分开,分别向两极移动。由于相互分离的是同源染色体,所以染色体数目减半。但每个子细胞的DNA含量仍为2C。同源染色体随机分向两极,使母本和父本染色体重所组合,产生基因组的变异。如人类染色体是23对,染色体组合的方式有223个(不包括交换),因此除同卵孪生外,几乎不可能得到遗传上等同的后代。
,因此除同卵孪生外,几乎不可能得到遗传上等同的后代。")
71
4、末期I 染色体到达两极后,解旋为细丝状、核膜重建、核仁形成,同时进行胞质分裂。

72
5、减数分裂间期 在减数分裂I和II之间的间期很短,不进行DNA的合成,有些生物没有间期,而由末期I直接转为前期II。

74
(二)减数分裂II 可分为前、中、后、末四个四期,与有丝分裂相似。 通过减数分裂一个精母细胞形成4个精子。 而一个卵母细胞形成一个卵子及2-3个极体。
减数分裂II 可分为前、中、后、末四个四期,与有丝分裂相似。 通过减数分裂一个精母细胞形成4个精子。 而一个卵母细胞形成一个卵子及2-3个极体。")
76
减数分裂的过程

77
二、减数分裂的意义 ◆ 有丝分裂确保世代间遗传的稳定性; ◆ 增加变异机会,确保生物的多样性,增强生物 适应环境变化的能力;
◆ 减数分裂是生物有性生殖的基础,是生物遗 传、生物进化和生物多样性的重要基础保证。

78
三、减数分裂的特点 ◆ 遗传物质只复制一次,细胞连续分裂两次,导致染 色体数目减半; ◆ S期持续时间较长,进行部分DNA的复制;
◆ 同源染色体在减数分裂期Ⅰ(MeiosisI)配对联会、基因重组; ◆ 减数分裂同源染色体配对排列在中期板上,第一次 分列时,同源染色体分开。
配对联会、基因重组; ◆ 减数分裂同源染色体配对排列在中期板上,第一次. 分列时,同源染色体分开。")
79
减数分裂前S期与有丝分裂前S期长度比较

80
第四节 细胞周期调控 一、研究背景 Rao和Johnson(1970、1972、1974)将Hela细胞同步于不同阶段,然后与M期细胞混合,在灭活仙台病毒介导下,诱导细胞融合,发现与M期细胞融合的间期细胞产生了形态各异的早熟凝集染色体(prematurely condensed chromosome,PCC),这种现象叫做早熟染色体凝集(premature chromosome condensation)。 G1期PCC为单线状,因DNA未复制。 S期PCC为粉末状,因DNA由多个部位开始复制。 G2期PCC为双线染色体,说明DNA复制已完成。
将Hela细胞同步于不同阶段,然后与M期细胞混合,在灭活仙台病毒介导下,诱导细胞融合,发现与M期细胞融合的间期细胞产生了形态各异的早熟凝集染色体(prematurely condensed chromosome,PCC),这种现象叫做早熟染色体凝集(premature chromosome condensation)。 G1期PCC为单线状,因DNA未复制。 S期PCC为粉末状,因DNA由多个部位开始复制。 G2期PCC为双线染色体,说明DNA复制已完成。")
81
不同形态的PCC

82
不仅同类M期细胞可以诱导PCC,不同类的M期细胞也可以诱导PCC产生,如人和蟾蜍的细胞融合时同样有这种效果,这就意味着M期细胞具有某种促进间期细胞进行分裂的因子,即成熟促进因子(maturation promoting factor,MPF)。 早在1960s,Yoshio Masui发现成熟蛙卵的提取物能促进未成熟卵的胚胞破裂(Germinal Vesicle Breakdown,GVBD),后来Sunkara将不同时期Hela细胞的提取液注射到蛙卵母细胞中,发现G1和S期的抽取物不能诱导GVBD,而G2和M期的则具有促进胚胞破裂的功能,它将这种诱导物质称为有丝分裂因子(MF)。后来在CHO细胞,酵母和粘菌中也提取出相同性质的MF。这类物质被统称为MPF。
,后来Sunkara将不同时期Hela细胞的提取液注射到蛙卵母细胞中,发现G1和S期的抽取物不能诱导GVBD,而G2和M期的则具有促进胚胞破裂的功能,它将这种诱导物质称为有丝分裂因子(MF)。后来在CHO细胞,酵母和粘菌中也提取出相同性质的MF。这类物质被统称为MPF。")
83
1960s Leland Hartwell以芽殖酵母为实验材料,利用阻断在不同细胞周期阶段的温度敏感突变株(在适宜的温度下和野生型一样),分离出了几十个与细胞分裂有关的基因(cell division cycle gene,CDC)。如芽殖酵母的cdc28基因,在G2/M转换点发挥重要的功能。Hartwell还通过研究酵母菌细胞对放射线的感受性,提出了checkpoint(细胞周期检验点)的概念,意指当DNA受到损伤时,细胞周期会停下来。
,分离出了几十个与细胞分裂有关的基因(cell division cycle gene,CDC)。如芽殖酵母的cdc28基因,在G2/M转换点发挥重要的功能。Hartwell还通过研究酵母菌细胞对放射线的感受性,提出了checkpoint(细胞周期检验点)的概念,意指当DNA受到损伤时,细胞周期会停下来。")
84
裂殖酵母细胞周期

85
1970s Paul Nurse等人以裂殖酵母为实验材料,同样发现了许多细胞周期调控基因,如:裂殖酵母cdc2、cdc25的突变型和在限制的温度下无法分裂;wee1突变型则提早分裂,而cdc25和wee1都发生突变的个体却会正常地分裂。进一步的研究发现cdc2和cdc28都编码一个34KD的蛋白激酶,促进细胞周期的进行。而weel和cdc25分别表现为抑制和促进CDC2的活性。这也解释了为何cdc25和wee1双重突变的个体可以恢复野生型的表型。

86
芽殖酵母细胞周期

87
Cdc25表达不足,细胞长得过长而不分裂; Wee1表达不足,细胞很小就开始分裂了

88
1983年Timothy Hunt首次发现海胆卵受精后,在其卵裂过程中两种蛋白质的含量随细胞周期剧烈振荡,在每一轮间期开始合成,G2/M时达到高峰,M结束后突然消失,下轮间期又重新合成,故命名为周期蛋白(cyclin)。后来在青蛙、爪蟾、海胆、果蝇和酵母中均发现类似的情况,各类动物来源的细胞周期蛋白mRNA均能诱导蛙卵的成熟。用海洋无脊椎动物和两栖类的卵为实验材料进行这类实验,好处在于卵的量比较大,而且在胚胎发育的早期,细胞分裂是同步化的。
。后来在青蛙、爪蟾、海胆、果蝇和酵母中均发现类似的情况,各类动物来源的细胞周期蛋白mRNA均能诱导蛙卵的成熟。用海洋无脊椎动物和两栖类的卵为实验材料进行这类实验,好处在于卵的量比较大,而且在胚胎发育的早期,细胞分裂是同步化的。")
89
MPF=CDC2+Cyclin B

90
1988年M. J. Lohka 纯化了爪蟾的MPF,经鉴定由32KD和45KD两种蛋白组成,二者结合可使多种蛋白质磷酸化。后来Paul Nurse(1990)进一步的实验证明P32实际上是CDC2的同源物,而P45是cyclinB的同源物,从而将细胞周期三个领域的研究联系在一起。2001年10月8日美国人Leland Hartwell、英国人Paul Nurse、Timothy Hunt因对细胞周期调控机理的研究而荣获诺贝尔生理医学奖
进一步的实验证明P32实际上是CDC2的同源物,而P45是cyclinB的同源物,从而将细胞周期三个领域的研究联系在一起。2001年10月8日美国人Leland Hartwell、英国人Paul Nurse、Timothy Hunt因对细胞周期调控机理的研究而荣获诺贝尔生理医学奖.")
91
2001年诺贝尔生理医学奖获得者

92
二、CDK CDC2与细胞周期蛋白结合才具有激酶的活性,称为细胞周期蛋白依赖性激酶(cyclin-dependent kinase,CDK),因此CDC2又被称为CDK1,激活的CDK1可将靶蛋白磷酸化而产生相应的生理效应,如将核纤层蛋白磷酸化导致核纤层解体、核膜消失,将H1磷酸化导致染色体的凝缩等等。这些效应的最终结果是细胞周期的不断运行。因此,CDK激酶和其调节因子又被称作细胞周期引擎。 目前发现的CDK在动物中有7种。各种CDK分子均含有一段相似的激酶结构域,这一区域有一段保守序列,即PSTAIRE,与周期蛋白的结合有关。
,因此CDC2又被称为CDK1,激活的CDK1可将靶蛋白磷酸化而产生相应的生理效应,如将核纤层蛋白磷酸化导致核纤层解体、核膜消失,将H1磷酸化导致染色体的凝缩等等。这些效应的最终结果是细胞周期的不断运行。因此,CDK激酶和其调节因子又被称作细胞周期引擎。 目前发现的CDK在动物中有7种。各种CDK分子均含有一段相似的激酶结构域,这一区域有一段保守序列,即PSTAIRE,与周期蛋白的结合有关。")
93
三、CKI 细胞中还具有细胞周期蛋白依赖性激酶抑制因子(CDK inhibitor,CKI)对细胞周期起负调控作用,目前发现的CKI分为两大家族: ①Ink4(Inhibitor of cdk 4),如P16ink4a、P15ink4b、P18ink4c、P19ink4d,特异性抑制cdk4·cyclin D1、cdk6·cyclin D1复合物。 ②Kip(Kinase inhibition protein):包括P21cip1 (cyclin inhibition protein 1)、P27kip1(kinase inhibition protein 1)、P57kip2等,能抑制大多数CDK的激酶活性,P21cip1还能与DNA聚合酶δ的辅助因子PCNA(proliferating cell nuclear antigen)结合,直接抑制DNA的合成
,如P16ink4a、P15ink4b、P18ink4c、P19ink4d,特异性抑制cdk4·cyclin D1、cdk6·cyclin D1复合物。 ②Kip(Kinase inhibition protein):包括P21cip1 (cyclin inhibition protein 1)、P27kip1(kinase inhibition protein 1)、P57kip2等,能抑制大多数CDK的激酶活性,P21cip1还能与DNA聚合酶δ的辅助因子PCNA(proliferating cell nuclear antigen)结合,直接抑制DNA的合成.")
94
P21cip1抑制CDK和PCNA

95
有丝分裂的变异 ① 不进行胞质分裂,形成二核或多核细胞; ② 染色体后期不开,或者进行核内有丝分裂,形成多倍体;
③ 姐妹染色单体不分离,形成双份染色体; ④ 细胞周期中缺少M期,核染色体反复加倍而不分开,形成 多线染色体; ⑤ 体细胞进行减数分裂,形成单倍体; ⑥ 多极分裂,由纺锤体极部纵裂并转向,引起多极分裂。

96
参与细胞有丝分裂的亚细胞结构 中心体(centrosome) 动粒(kinetochore)与着丝粒(centromere)
纺锤体(splindle)
")
97
中心体(centrosome) 中心体是动物细胞中的主要的微管组织中心。它由一对相互垂直的中心粒(centrioles)及其周围的基质构成。中心粒由微管构成,呈圆筒状结构,外围基质的主要成分是γ微管蛋白。中心体和外围的微管合成为星体,星体参与纺锤体的装配。 中心体周期(centrosome cycle):G1期末复制;G2期移向细胞两极,并组织星体和纺锤体;细胞分裂结束,分布在两个子细胞中。
:G1期末复制;G2期移向细胞两极,并组织星体和纺锤体;细胞分裂结束,分布在两个子细胞中。")
98
Centrosome cycle

99
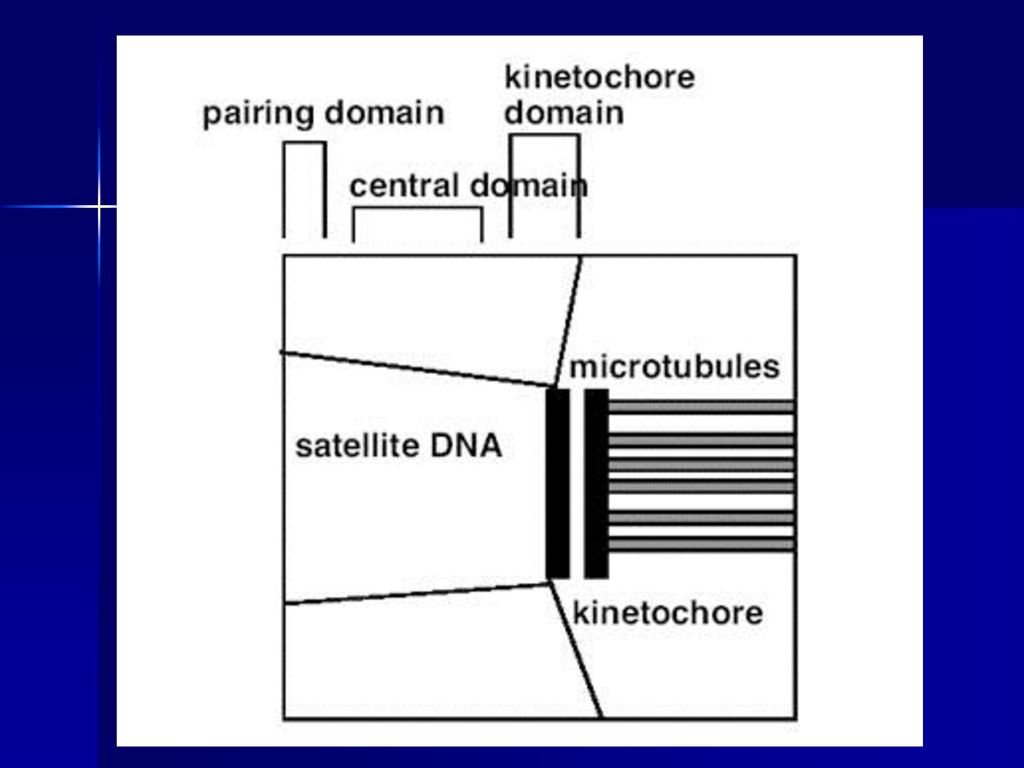
动粒与着丝粒 着丝粒:是指染色体主缢痕部位的染色质,它把姊妹染色体单体连接在一起,并把染色体分成两个臂。
动粒:是位于着丝粒两侧由蛋白质构成的三层盘状或球状结构。和纺锤体相连,与染色体的向极移动有关。 动粒的结构:内层(着丝粒染色质) 中层(细纤维横跨) 外层(微管)
 中层(细纤维横跨) 外层(微管)")
101
纺锤体(spindle) 概念(concept):由微管和微管蛋白组成的参与染色体向极移 动的纺锤式的结构。
结构组成:动粒微管:一端和中心体相连,另一端和动粒连。 极性微管:一端和中心体相连,另一端游离或者是 相互搭桥。 装配:微管在中心体周围的装配: γ微管蛋白 中心体的分离:移动素类蛋白(KRPs) 胞质动力蛋白(dynein)
 胞质动力蛋白(dynein)")
102
纺锤体的形态结构

103
纺锤体的装配

104
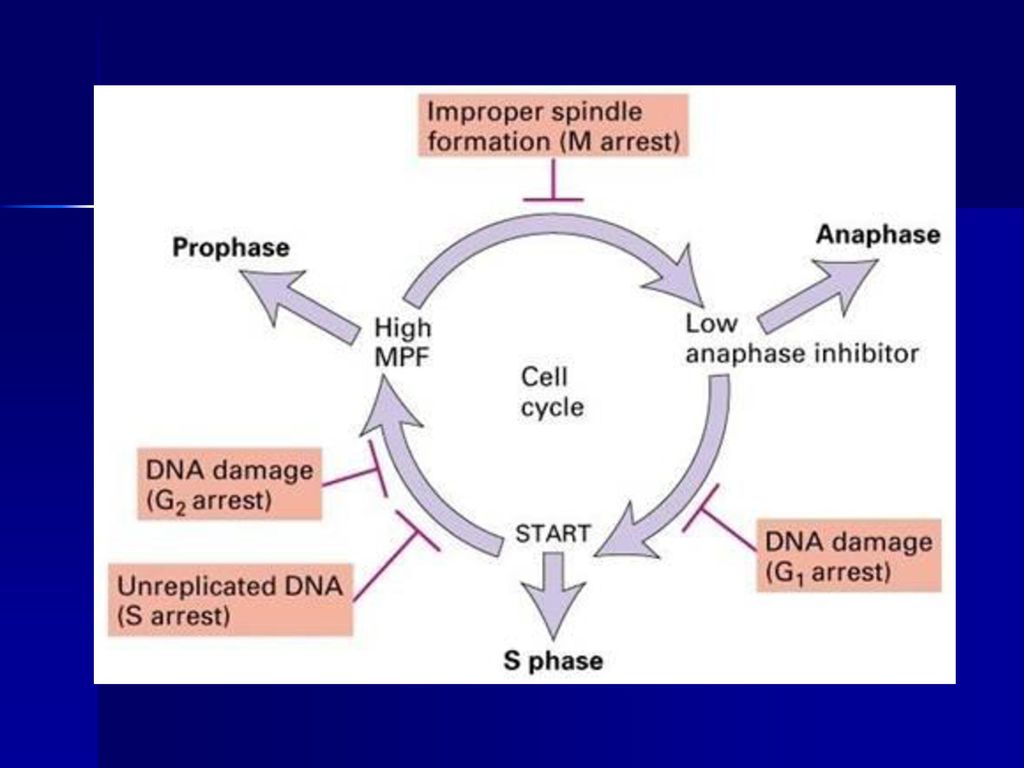
细胞周期中的检验点 G1/S检验点:start点或R点,控制细胞由静止状态的G1进入 DNA合成期,检查DNA是否损伤?细胞外环境是否适宜?
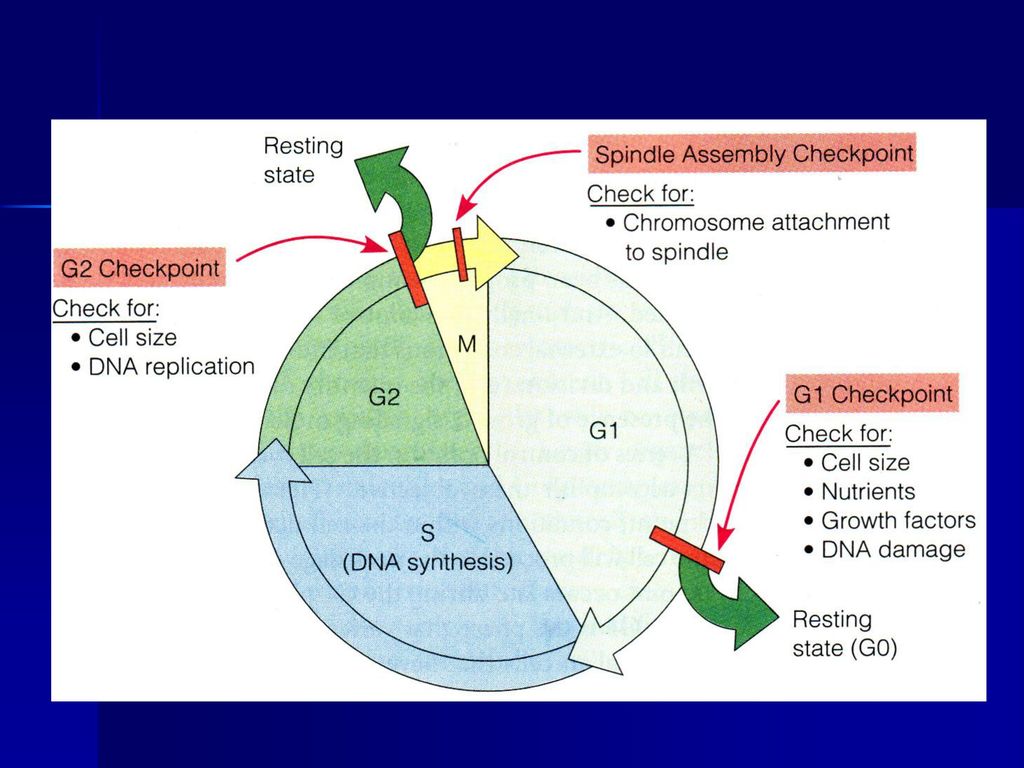
细胞体积是否足够大? S期检验点:DNA复制是否完成? G2/M检验点:是决定细胞一分为二的控制点,检查DNA是 否损伤?细胞体积是否足够大? 中-后期检验点(纺锤体组装检验点)
")
107
不同类型的周期蛋白 周期蛋白的结构 激酶复合体 脊椎动物 芽殖酵母 Cyclin CDK G1-CDK Cyclin D* CDK4 、6
Cln 3 CDK1(CDC28) G1/S-CDK Cyclin E CDK2 Cln 1、2 S-CDK Cyclin A Clb 5、6 M-CDK Cyclin B CDK1(CDC2) Clb 1-4 周期蛋白的结构
 G1/S-CDK. Cyclin E. CDK2. Cln 1、2. S-CDK. Cyclin A. Clb 5、6. M-CDK. Cyclin B. CDK1(CDC2) Clb 1-4. 周期蛋白的结构.")
108
End

Similar presentations





. 第一关:测基础 判正误 第二关:练规范 强素质 第一关:测基础 判正误 1. 病毒是一种生物,但它不是一个独立的生命系统 ( ) 2. 细胞学说揭示了细胞的统一性和多样性 ( ) 3. 原核细胞中只含有核糖体一种细胞器 ( ) 4. 蓝藻细胞不含有叶绿体,不能进行光合作用.>")

 考点二: 细胞膜与细胞壁(实验 “ 验证活细胞 吸收物质的选择性 ” :目的要求、材料用具、方 法步骤、实验现象和结果、讨论) 考点三:细胞质(实验 “ 观察叶绿体的形态和分 布.>")
 类型 :>")










 减数分裂(Meiosis)>")