Download presentation
Presentation is loading. Please wait.

1
基础分子生物学

3
第十二章 遗传信息的传递与表达

4
本章重点介绍 DNA、RNA、蛋白质的生物合成与调控
RNA转录的定义、特点(方式、方向、酶、蛋白因子)、过程(起始、延长、终止);RNA转录后修饰。 翻译的定义、特点(合成体系、几种RNA的作用、方向、酶、蛋白因子)、过程(起始、延长、终止);新生蛋白质的转运及加工。 基因表达的调节控制。 教材第16、17、18、19章
、过程(起始、延长、终止);RNA转录后修饰。 翻译的定义、特点(合成体系、几种RNA的作用、方向、酶、蛋白因子)、过程(起始、延长、终止);新生蛋白质的转运及加工。 基因表达的调节控制。 教材第16、17、18、19章.")
5
中心法则: DNA dependent DNA polymerase——DDDP
DNA dependent RNA polymerase——DDRP RNA dependent RNA polymerase——RDRP RNA dependent DNA polymerase——RDDP

6
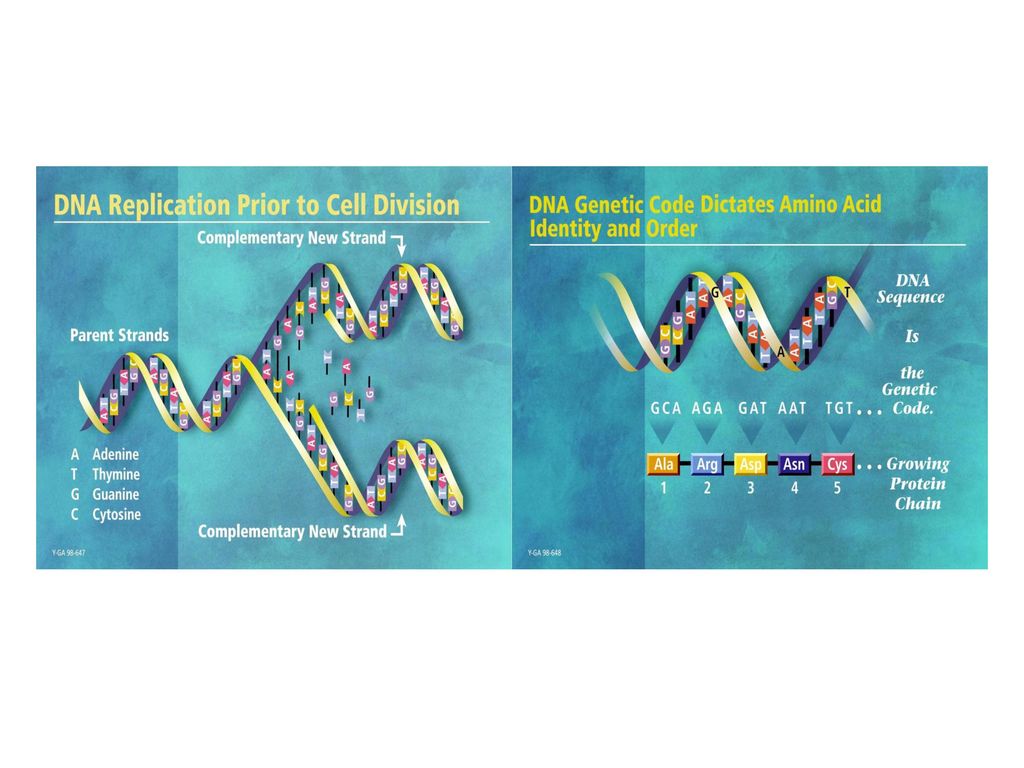
第一节 DNA复制的特点 一、定义: 二、基本特点: 以亲代DNA为模板,按碱基互补配对原则倍增两个完全
相同的子代DNA的过程称为复制(replication)。 二、基本特点: 1、合成体系 (1)模 板(template):母链DNA,并解链解旋。 (2)原 料:游离的dNTP、即dATP、dGTP、dCTP、dTTP (3)酶:DNA聚合酶、引物酶、解链酶、解旋酶、DNA连接酶。 (4)引 物(primer) :RNA
。 二、基本特点: 1、合成体系. (1)模 板(template):母链DNA,并解链解旋。 (2)原 料:游离的dNTP、即dATP、dGTP、dCTP、dTTP (3)酶:DNA聚合酶、引物酶、解链酶、解旋酶、DNA连接酶。 (4)引 物(primer) :RNA.")
7
2、复制方式 : P534图16-2 半保留复制,1958年Meselson 和 Stahl 实验证明

8
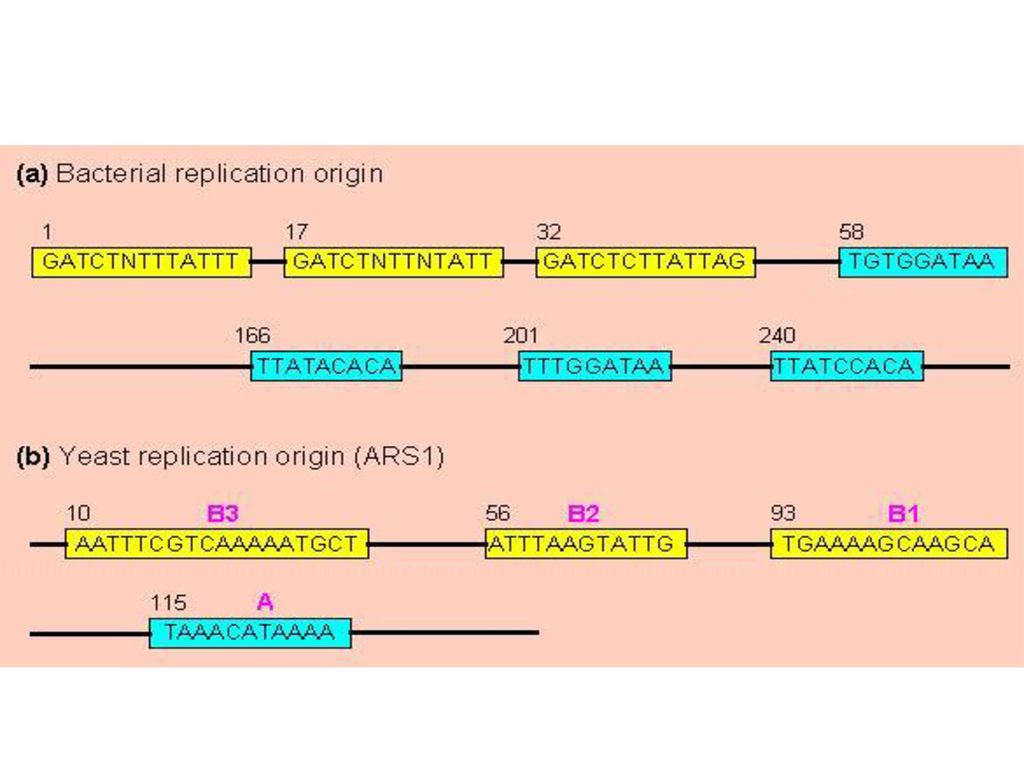
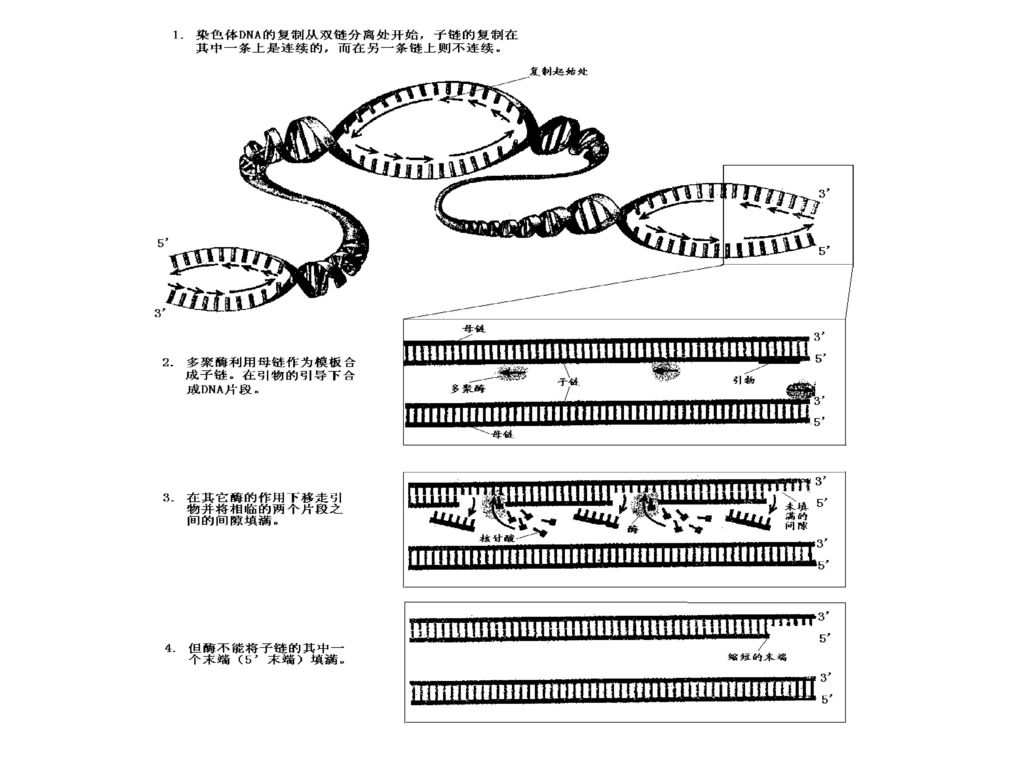
DNA复制时,以亲代DNA的每一条链作模板,合成完全相同的两个双链子代DNA,每个子代DNA中都含有一条亲代DNA链,另一条链是新合成的,这种现象称为DNA的半保留复制(semiconservative replication)。 3、复制起始点: 能独立进行复制的单位称为复制子(replicon)。 DNA复制是从特定位点起始,是一些具有特定核苷酸排列顺序的片段,即复制起始点(origin)。在原核生物中,复制起始点通常为一个,而在真核生物中则为多个。(p541图16-3)
。 DNA复制是从特定位点起始,是一些具有特定核苷酸排列顺序的片段,即复制起始点(origin)。在原核生物中,复制起始点通常为一个,而在真核生物中则为多个。(p541图16-3)")
11
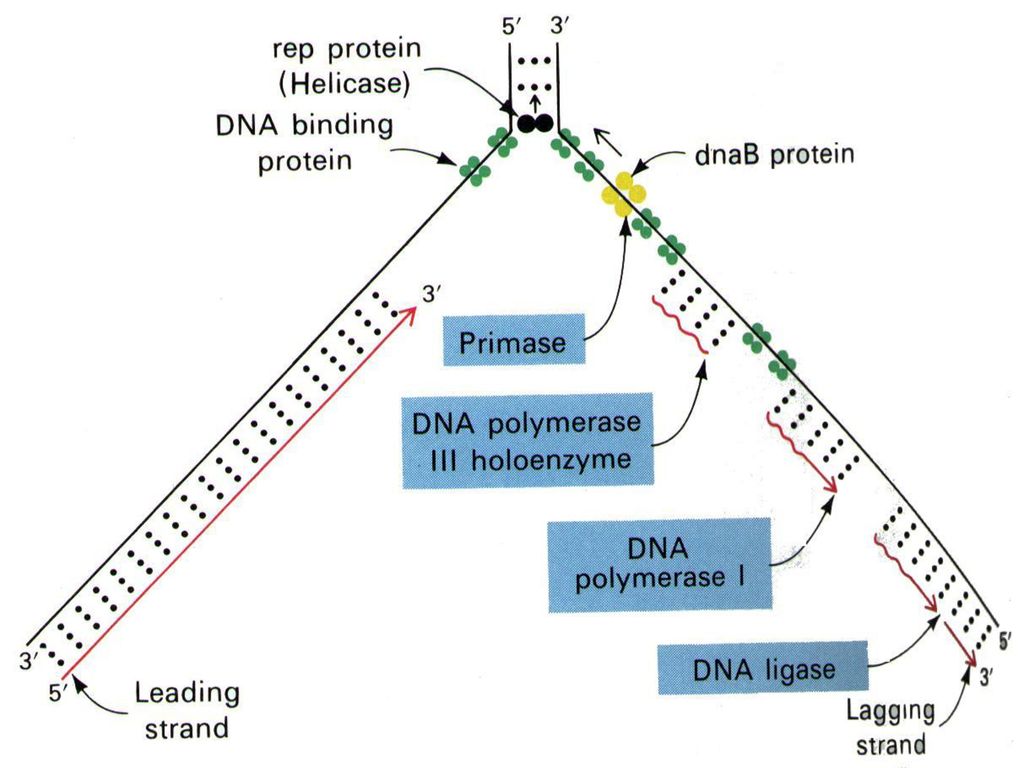
4、双向复制: P540 DNA复制时,以复制起始点为中心,向两个方向进行复制。 复制叉(replication fork) :亲代DNA复制起始时出现双链局部解链解旋而使DNA新链合成的部位象一个“叉子”,所以这个部位称为复制叉。
 :亲代DNA复制起始时出现双链局部解链解旋而使DNA新链合成的部位象一个 叉子 ,所以这个部位称为复制叉。")
12
在低等生物中,单链环状DNA主要采用单向复制(滚环式复制),如:噬菌体φX174DNA.(P544图16-16)
3′ 5′ 5′ 3′ 5′

13
5、复制方向: 5’→3’ 1968年,冈崎等人用噬菌体感染的大肠杆菌为实验材料,使用含有3H标记的胸苷的培养基进行培养,然后分离出DNA,测定新合成的DNA。

14
半不连续复制(semi-discontinuous replication):
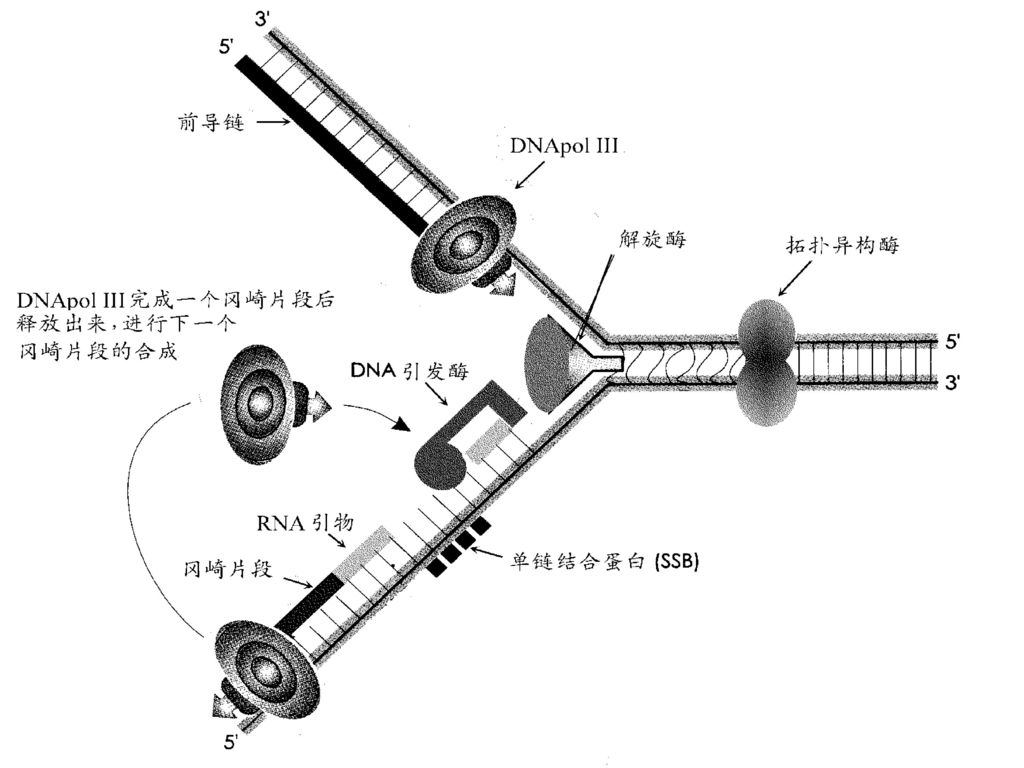
以3’→5’方向的亲代DNA链作模板的子代链在复制时是连续进行的,其子代链的聚合方向为5’ →3’,这一条链被称为前导链(leading strand)。 以5’→3’方向的亲代DNA链为模板的子代链在复制时则是不连续的,其链的聚合方向也是5’ →3’,这条链被称为滞后链(lagging strand)。 由滞后链所形成的一些子代DNA短链称为冈崎片段(Okazaki fragment),原核生物中约为1000~2000个核苷酸,真核生物中约为100~200个核苷酸。
。 以5’→3’方向的亲代DNA链为模板的子代链在复制时则是不连续的,其链的聚合方向也是5’ →3’,这条链被称为滞后链(lagging strand)。 由滞后链所形成的一些子代DNA短链称为冈崎片段(Okazaki fragment),原核生物中约为1000~2000个核苷酸,真核生物中约为100~200个核苷酸。")
15
6、复制起始于RNA引物的合成 DNA聚合酶不能从头开始进行聚合反应,只能将核苷酸加到已有的多核苷酸链上,因此DNA复制时需要在复制叉处合成一段短的与模板部分互补的RNA引物,包括每个冈崎片段的合成都是开始于一个RNA引物,在DNA聚合酶催化下从引物3’端延伸形成冈崎片段。 RNA引物是通过引发酶合成的,引物RNA(几个核苷酸)在复制过程中暂时存在,最后被外切酶水解。
在复制过程中暂时存在,最后被外切酶水解。")
16
三、参与DNA复制的酶及蛋白因子 复制是在酶催化下进行的核苷酸聚合过程。 1.解链、解旋酶
(1)解螺旋酶(helicase)/解链酶:是个需要ATP的酶,在移动的复制叉前面沿着单链DNA滑动而使双链解旋,形成适于DNA聚合酶和引发酶作用的单链模板DNA。
解螺旋酶(helicase)/解链酶:是个需要ATP的酶,在移动的复制叉前面沿着单链DNA滑动而使双链解旋,形成适于DNA聚合酶和引发酶作用的单链模板DNA。")
17
(2)拓扑异构酶(topoisomerase):
是用来将解链酶作用后产生的扭转张力释放掉而快速解除复制叉移动产生的超螺旋。 酶Ⅰ:断开单链,使 DNA链下游沿轴松解方向旋转成松驰状态,然后封闭切口,不耗能。 酶Ⅱ:断开双链,链呈松驰状态后封闭切口,耗能时能使DNA分子转变为负超螺旋,更好地起模板作用。 (3)单链结合蛋白(single-strand binding protein,SSB) 防止解开的螺旋恢复原来状态及被核酸酶水解,对单链DNA有非常高的亲和性。
单链结合蛋白(single-strand binding protein,SSB) 防止解开的螺旋恢复原来状态及被核酸酶水解,对单链DNA有非常高的亲和性。")
18
(4)DNA引发酶(primase) P541表16-3
特殊的RNA聚合酶,以DNA为模板催化一小段引物RNA合成,带有3’-OH 端,合成后以氢键与模板结合。每秒钟催化10个核苷酸的RNA引物的合成。(依赖ATP在模板链上移动) 引发体成分
 引发体成分.")
20
催化dNTP聚合到核酸链上,是依赖DNA的DNA聚合酶。 (DNA聚合酶的反应特点,p538)
2.DNA聚合酶( polymerase) 催化dNTP聚合到核酸链上,是依赖DNA的DNA聚合酶。 (DNA聚合酶的反应特点,p538) PPi H+
 催化dNTP聚合到核酸链上,是依赖DNA的DNA聚合酶。 (DNA聚合酶的反应特点,p538) PPi. H+")
21
种类: 原核生物:DNA聚合酶Ⅰ、Ⅱ、Ⅲ(PolⅠ、Ⅱ、Ⅲ),比活最大起真正复制作用的是PolⅢ ,含量最多的是 PolⅠ,这三种酶都属于具有多种酶活性的多功能酶。 (p538表16-1) 修复损伤
,比活最大起真正复制作用的是PolⅢ ,含量最多的是 PolⅠ,这三种酶都属于具有多种酶活性的多功能酶。 (p538表16-1) 修复损伤")
22
polⅠ是单一肽链的大分子蛋白质,可被蛋白酶水解为两个片段,其中的大片段称为Klenow fragment (Mr68000),具有5’→3’聚合酶活性和3’→5’外切酶的活性。小片段(Mr35000)具有 5’→3’外切酶的活性。
,具有5’→3’聚合酶活性和3’→5’外切酶的活性。小片段(Mr35000)具有 5’→3’外切酶的活性。")
23
DNA pol I降解RNA引物的同时合成DNA填补空隙,这种同时合成和降解的过程称为切口平移(nick translation),其效果是合成方向中切口的移动。
,其效果是合成方向中切口的移动。")
24
polⅢ由十种亚基组成,其中α亚基具有5’→3’聚合DNA的酶活性,具有复制DNA的功能;而ε亚基具有3’→5’外切酶的活性,与DNA复制的校正功能有关

25
DNA聚合酶校正反应:

26
真核生物:DNA聚合酶α、β、γ、δ、ε。 α、δ在复制中起主要作用。(p539表16-9)
引物的合成 前导链和滞后链的合成 DNA复制的保真性: 1.遵守严格的碱基配对规律; 2.DNA聚合酶在复制时对碱基的正确选择; 3.对复制过程中出现的错误及时进行校正。

27
3.DNA连接酶(DNA ligase) 连接DNA链3’-OH末端和另一DNA链5’-P 末端,使二者生成磷酸二酯键。 DNA连接酶催化的条件是:① 需一段DNA片段具有3'-OH,而另一段DNA片段具有5'-Pi基;② 未封闭的缺口位于双链DNA中,即其中有一条链是完整的;③ 需要消耗能量,在原核生物中由NAD+供能,在真核生物中由ATP供能。

28
作用:冈崎片段的连接;引物水解留下的空缺由PolⅠ催化填补,连接酶缝合缺口;DNA修复、重组、剪接。

30
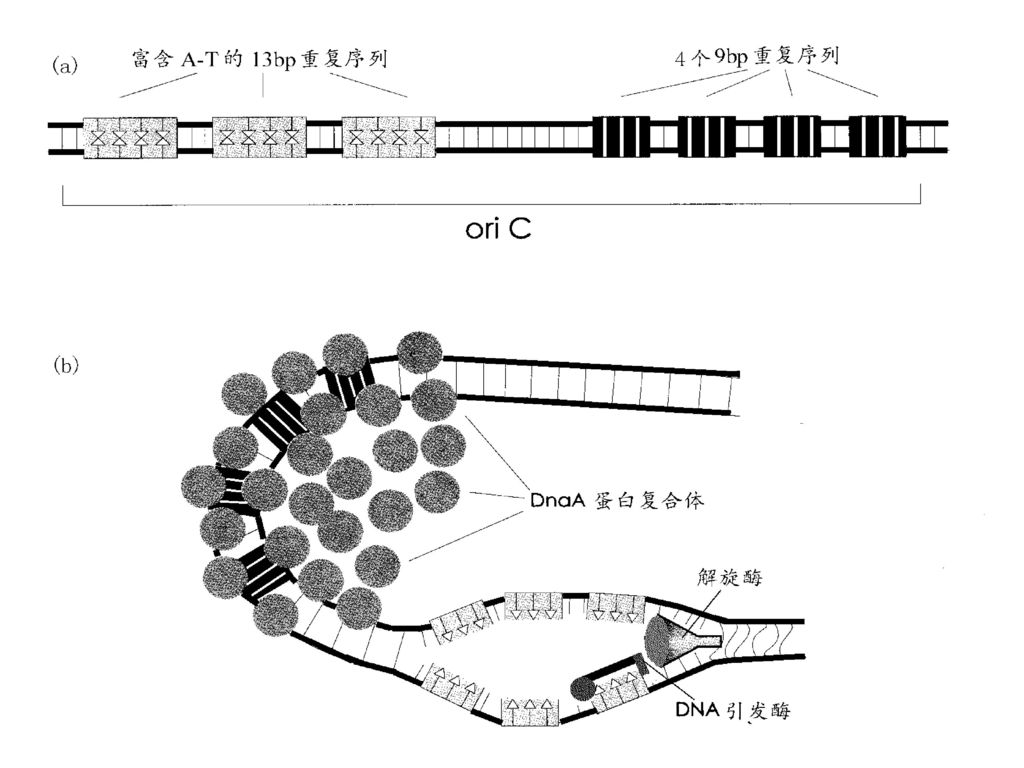
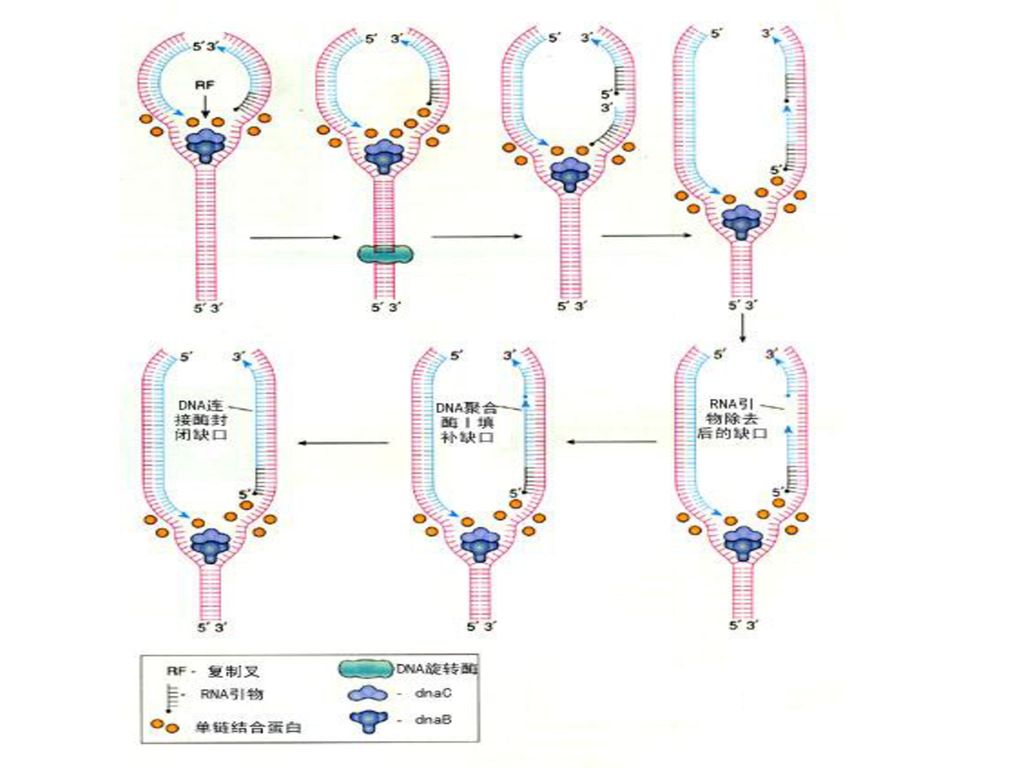
第二节 DNA复制过程 一、复制的起始 —— 形成复制叉 1、预引发
(1)引发体组装:由蛋白因子(如Dna A等)识别复制起始点,并与其他蛋白因子以及引物酶一起组装形成引发体。 (2)解旋解链,形成复制叉:由拓扑异构酶和解链酶作用,使DNA的超螺旋及双螺旋结构解开,碱基间氢键断裂,形成两条单链DNA。单链DNA结合蛋白(SSB)结合在两条单链DNA上,形成复制叉。 2、引发:在引物酶的催化下,以DNA为模板,合成一段短的RNA片段(10多个核苷酸),从而获得3'端自由羟基(3'-OH)。
引发体组装:由蛋白因子(如Dna A等)识别复制起始点,并与其他蛋白因子以及引物酶一起组装形成引发体。 (2)解旋解链,形成复制叉:由拓扑异构酶和解链酶作用,使DNA的超螺旋及双螺旋结构解开,碱基间氢键断裂,形成两条单链DNA。单链DNA结合蛋白(SSB)结合在两条单链DNA上,形成复制叉。 2、引发:在引物酶的催化下,以DNA为模板,合成一段短的RNA片段(10多个核苷酸),从而获得3 端自由羟基(3 -OH)。")
31
× * NI N1 RNA引物 5' 3' 拓扑异构酶 解链酶 单链结合蛋白 引物酶 聚合酶Ⅲ

32
二、复制的延长 1、聚合子代DNA: 由DNA聚合酶催化,从5'→3'方向聚合子代DNA链。 2、引发体移动:
引发体向前移动,解开新的局部双螺旋,形成新的复制叉,随从链重新合成RNA引物,继续进行链的延长。

33
* 5' 3'

34
三、复制的终止 1、去除引物,填补缺口: 在原核生物中,由DNA聚合酶Ⅰ水解去除RNA引物,并催化延长引物缺口处的DNA,而在真核生物中,RNA引物的去除,由一种特殊的核酸酶来水解,而冈崎片段由DNA聚合酶ε来延长。 2、连接冈崎片段: 在DNA连接酶的催化下,形成最后一个磷酸酯键,将冈崎片段连接起来,形成完整的DNA长链。

35
. 聚合酶Ⅰ RNA酶 DNA连接酶 5' 3'

36
E.Coli中的ter(terminus)区组织,含有5个非对称的ter部位,与Tus (terminus utilization substance,终止子利用物质)结合成Tus-ter复合物,通过阻止解旋酶解旋DNA来封闭复制叉的通路。
区组织,含有5个非对称的ter部位,与Tus (terminus utilization substance,终止子利用物质)结合成Tus-ter复合物,通过阻止解旋酶解旋DNA来封闭复制叉的通路。")
38
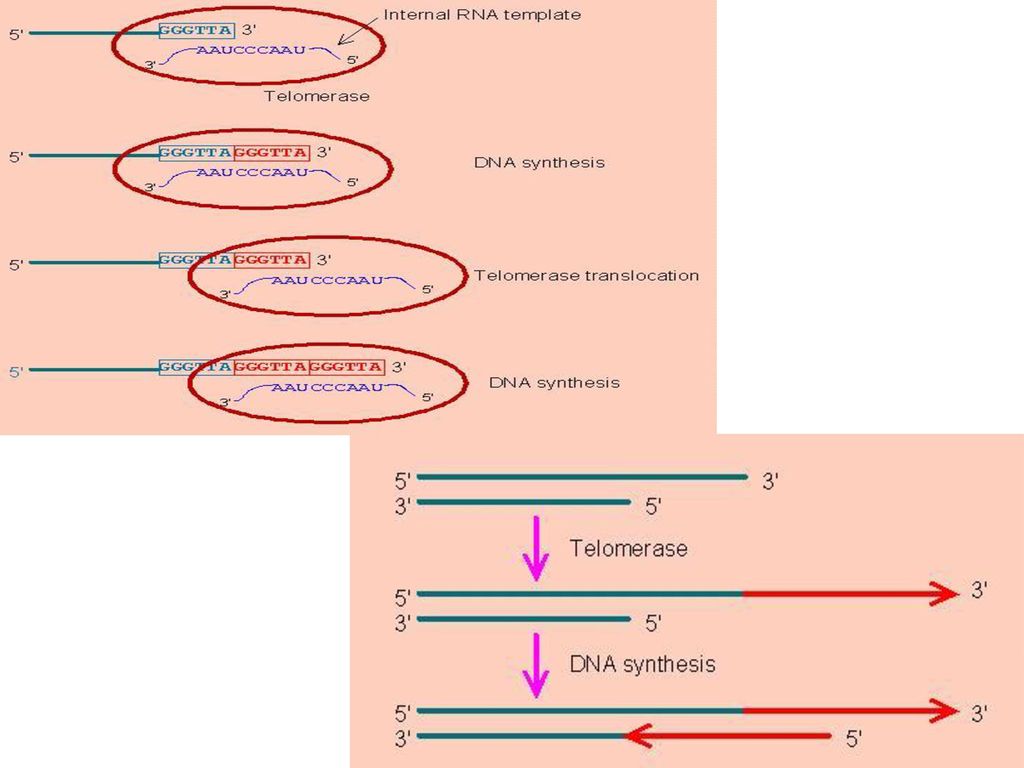
四、真核生物的端粒 真核细胞线性DNA在复制完成后,其末端由于引物RNA的水解而可能出现缩短,故通过形成端粒解决。
端粒(telomere)是指真核生物染色体线性DNA分子末端的结构部分,通常膨大成粒状。其共同的结构特征是由一些富含G、C的短重复序列构成,可重复数十次至数百次。如:人的端粒为TTAGGG。 端粒的作用:填补5’末端消除RNA引物后形成的空隙;稳定染色体末端结构,防止染色体间末端连接;决定细胞的寿命。 端粒酶(telomerase)是一种RNA-蛋白质复合体,以其RNA为模板,对末端DNA链进行延长,主要存在于生殖细胞中。
是指真核生物染色体线性DNA分子末端的结构部分,通常膨大成粒状。其共同的结构特征是由一些富含G、C的短重复序列构成,可重复数十次至数百次。如:人的端粒为TTAGGG。 端粒的作用:填补5’末端消除RNA引物后形成的空隙;稳定染色体末端结构,防止染色体间末端连接;决定细胞的寿命。 端粒酶(telomerase)是一种RNA-蛋白质复合体,以其RNA为模板,对末端DNA链进行延长,主要存在于生殖细胞中。")
40
端粒酶的结构

42
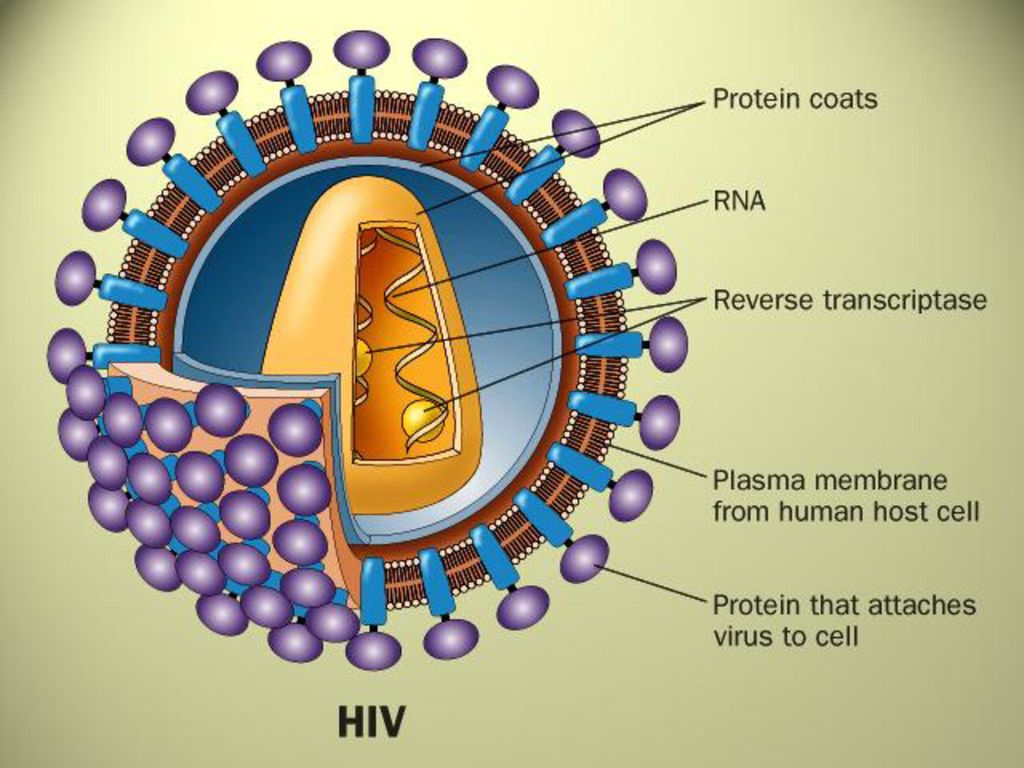
五、逆转录现象 (p545) 在RNA病毒(逆转录病毒,如致癌RNA病毒)中,以RNA为模板,按碱基配对确定核苷酸顺序,合成DNA的过程称为逆转录作用(reverse transcription)。由逆转录酶催化合成的DNA为cDNA(complementary DNA),即互补DNA。 DNA双链 逆转录酶 DNA聚合酶 DNA单链 n1dATP n2dCTP n3dGTP n4dTTP + RNA (模板) RNA-DNA
 RNA-DNA.")
43
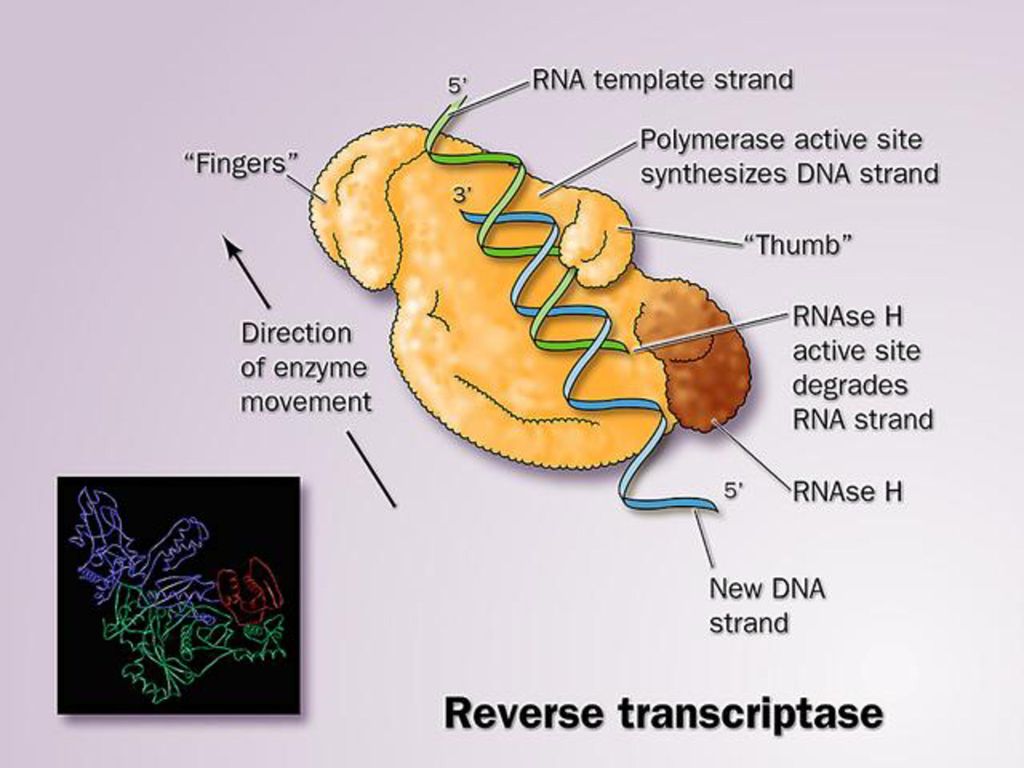
将逆转录病毒RNA转换成DNA的最关键酶是逆转录酶(reverse transcriptase)。该酶具有Rnase H活性(降解RNA活性)和DNA聚合酶活性(RDDP、DDDP)。
逆转录酶没有3’-5’外切酶活性或校正活性,它的错误率比任何DNA聚合酶都高,例如人免疫缺陷病毒I(human immune deficiency virus I, HIV I)逆转录酶每合成2000到4000核苷酸就会掺入一个不正确的碱基,说明HIV I的高突变率。 意 义: 丰富中心法则内容;解释致癌病毒产生癌细胞的机制; 用于基因工程作为载体而获得目的基因。
逆转录酶每合成2000到4000核苷酸就会掺入一个不正确的碱基,说明HIV I的高突变率。 意 义: 丰富中心法则内容;解释致癌病毒产生癌细胞的机制; 用于基因工程作为载体而获得目的基因。")
45
逆转录病毒的生活循环

47
六、聚合酶链式反应(polymerase chain reaction,PCR)
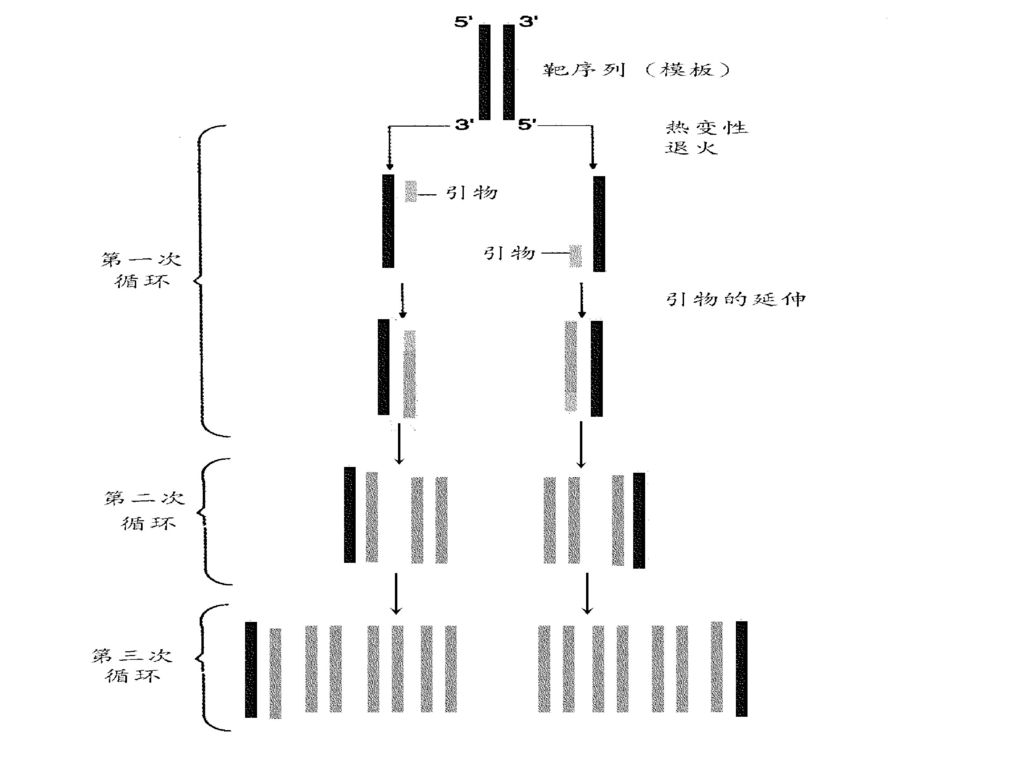
Kary Mullis1985年发明,获得1993年的诺贝尔化学奖 嗜热水生菌Thermus aquaticus的DNA聚合酶称之Taq聚合酶(Taq polymerase),是单一多肽链,具有良好的热稳定性,在70~75℃时具有最高生物学活性。 PCR技术是DNA的体外酶促扩增,又称为无细胞分子克隆法。是在试管内使用Taq聚合酶进行DNA的合成反应,并用合成的DNA引物代替RNA引物(20~24个核苷酸) ,通过加热(变性)、冷却(退火)、保温(延伸)等改变温度的方法使DNA得以复制。在重复20至30个循环后,复制出的DNA产量可达230拷贝,大约为109拷贝。
,是单一多肽链,具有良好的热稳定性,在70~75℃时具有最高生物学活性。 PCR技术是DNA的体外酶促扩增,又称为无细胞分子克隆法。是在试管内使用Taq聚合酶进行DNA的合成反应,并用合成的DNA引物代替RNA引物(20~24个核苷酸) ,通过加热(变性)、冷却(退火)、保温(延伸)等改变温度的方法使DNA得以复制。在重复20至30个循环后,复制出的DNA产量可达230拷贝,大约为109拷贝。")
49
PCR技术的应用 用于合成特异探针 用于DNA测序 产生和分析基因突变 重组DNA 扩增未知的DNA 比较基因组序列 应用于临床医学与法医学

50
七、原核生物与真核生物DNA复制比较: 相同点:前导链合成是连续的,随后链合成是不连续的,引物合成,冈崎片段合成,引物水解后填充与模板链互补的核苷酸等过程。 区别: (1)复制形式: 原核:θ模型(E.coli)、滚环模型( φX174) 真核:超螺旋、双链闭环、线状、双链开环 (2) 真核生物复制叉处需要的辅助蛋白不同。 真核生物所含的DNA聚合酶种类更多些。 真核生物复制是双向的,但E.coli染色体含有唯一的复制起点,而真核生物染色体存在许多复制起点。
 真核生物复制叉处需要的辅助蛋白不同。 真核生物所含的DNA聚合酶种类更多些。 真核生物复制是双向的,但E.coli染色体含有唯一的复制起点,而真核生物染色体存在许多复制起点。")
51
第三节 DNA的损伤与修复 一、DNA的损伤(突变)
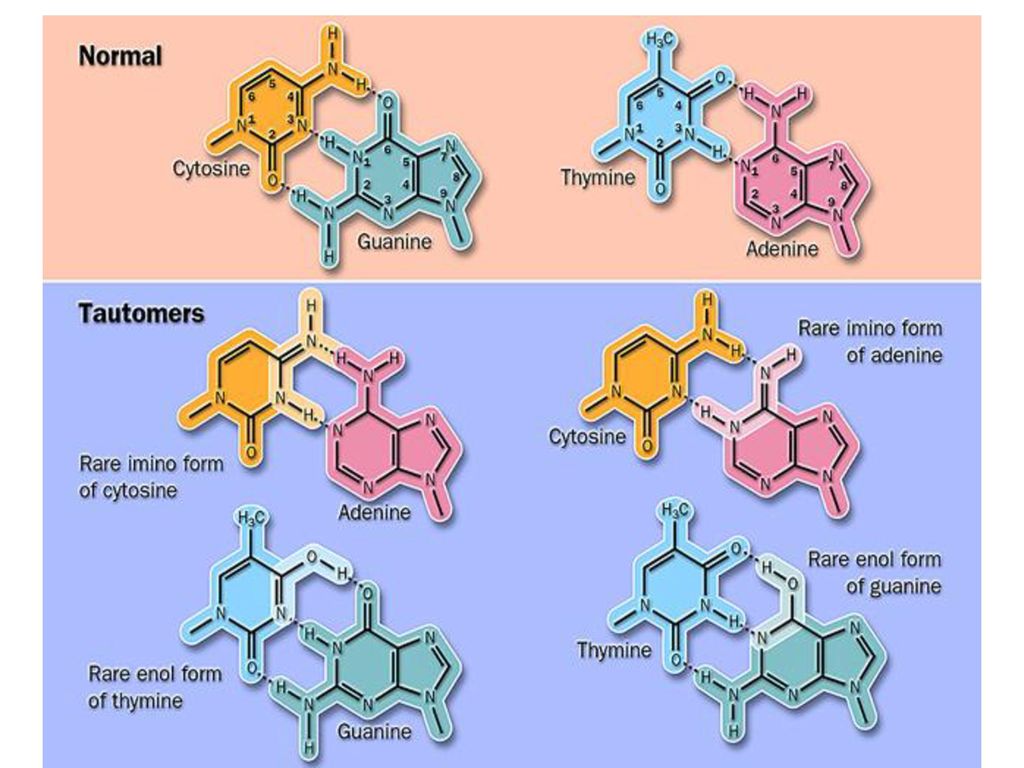
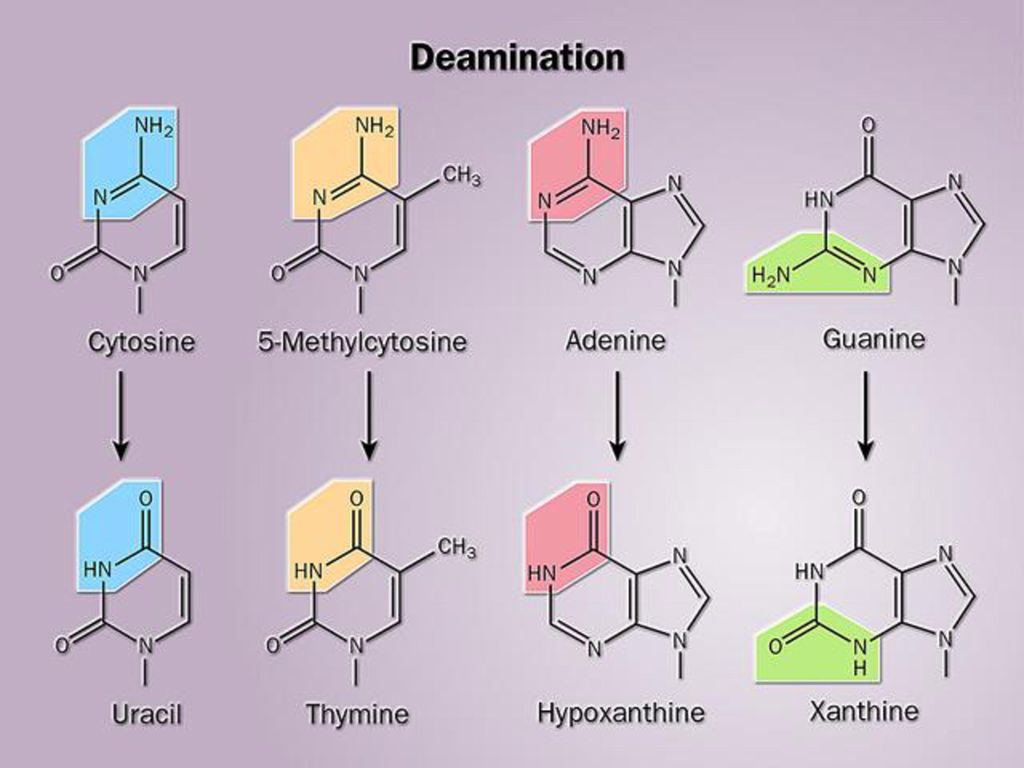
由自发的或环境因素引起DNA一级结构的任何异常改变称为DNA的损伤,也称为突变(mutation)。 DNA的损伤包括碱基脱落、碱基修饰、交联、链的断裂,重组等。 1、引起突变的因素: (1)自发因素: 自发脱碱基:由于N-糖苷键的自发断裂,引起嘌呤或嘧啶碱基的脱落。每日可达近万个核苷酸残基。 自发脱氨基:胞嘧啶自发脱氨基可生成尿嘧啶,腺嘌呤自发脱氨基可生成次黄嘌呤。每日可达几十到几百个核苷酸残基。 复制错配:由于复制时碱基配对错误引起的损伤,发生频率较低。
。 DNA的损伤包括碱基脱落、碱基修饰、交联、链的断裂,重组等。 1、引起突变的因素: (1)自发因素: 自发脱碱基:由于N-糖苷键的自发断裂,引起嘌呤或嘧啶碱基的脱落。每日可达近万个核苷酸残基。 自发脱氨基:胞嘧啶自发脱氨基可生成尿嘧啶,腺嘌呤自发脱氨基可生成次黄嘌呤。每日可达几十到几百个核苷酸残基。 复制错配:由于复制时碱基配对错误引起的损伤,发生频率较低。")
53
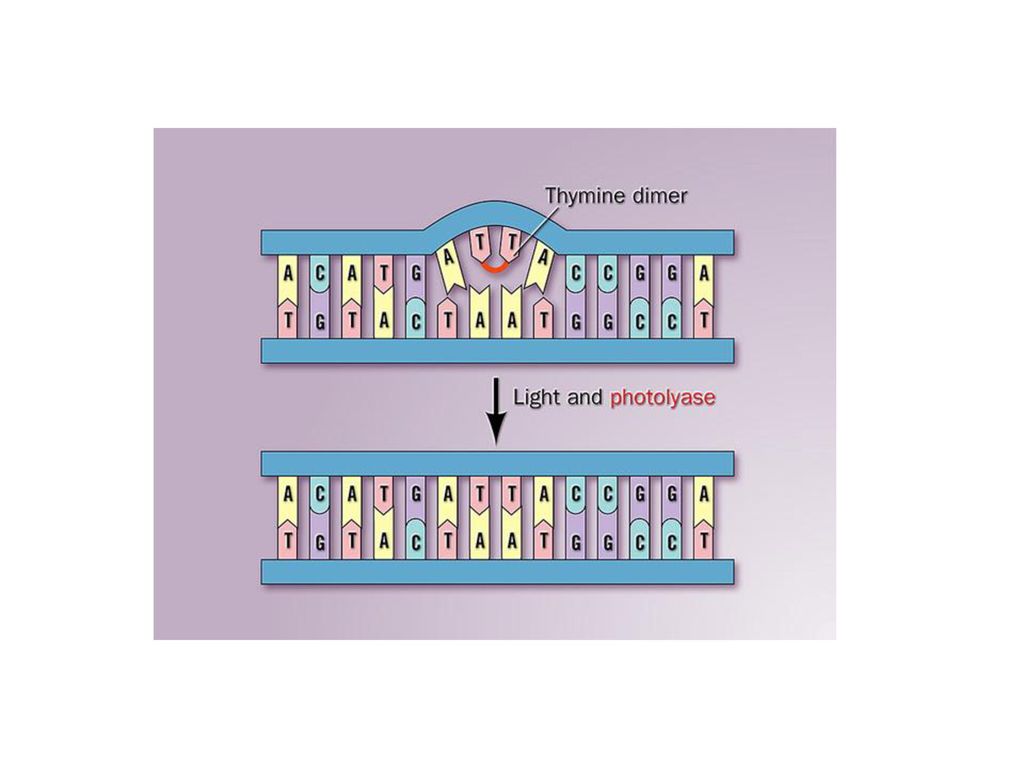
(2)物理因素: 由紫外线、电离辐射、X射线等引起的DNA损伤。其中,X射线和电离辐射常常引起DNA链的断裂,而紫外线常常引起嘧啶二聚体的形成,如TT,TC,CC等二聚体。这些嘧啶二聚体由于形成了共价键连接的环丁烷结构,因而会引起复制障碍。
物理因素: 由紫外线、电离辐射、X射线等引起的DNA损伤。其中,X射线和电离辐射常常引起DNA链的断裂,而紫外线常常引起嘧啶二聚体的形成,如TT,TC,CC等二聚体。这些嘧啶二聚体由于形成了共价键连接的环丁烷结构,因而会引起复制障碍。")
54
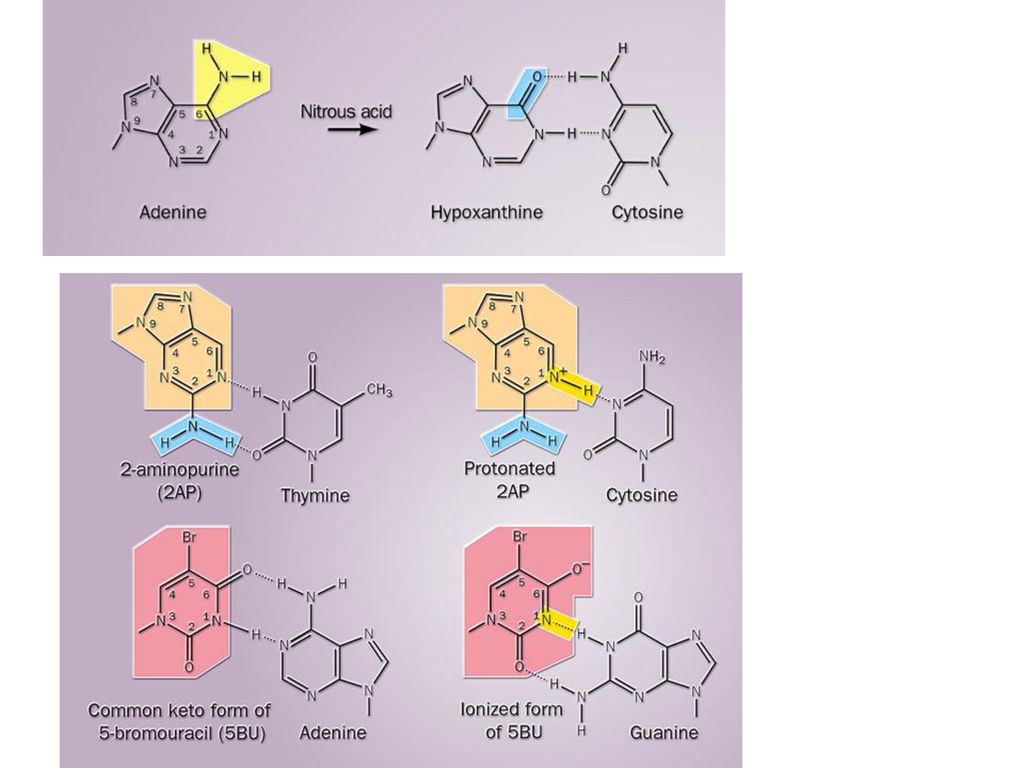
(3)化学因素: 脱氨剂:如亚硝酸与亚硝酸盐使C脱氨基生成U,A脱氨基生成I。 烷基化剂:如氮芥、乙基乙烷磺酸等是一类带有活性烷基的化合物,可提供甲基或其他烷基,引起碱基或磷酸基的烷基化,甚至可引起邻近碱基的交联。 DNA加合剂:如苯并芘,在体内代谢后生成四羟苯并芘,与嘌呤共价结合引起损伤。 碱基类似物:如5-FU,6-MP等,可掺入到DNA分子中引起损伤或突变。 断链剂:如过氧化物,含巯基化合物等,可引起DNA链的断裂。

57
2、DNA突变的类型:

58
3、DNA突变的效应: 同义突变:基因突变导致mRNA密码子第三位碱基的改变但不引起密码子意义的改变,其翻译产物中的氨基酸残基顺序不变,但有时可引起翻译效率降低。 误义突变:基因突变导致mRNA密码子碱基被置换,其意义发生改变,翻译产物中的氨基酸残基顺序发生改变。 无义突变:基因突变导致mRNA密码子碱基被置换而改变成终止密码子,引起多肽链合成的终止。 移码突变:基因突变导致mRNA密码子碱基被置换,引起突变点之后的氨基酸残基顺序全部发生改变。

59
二、DNA损伤的修复 分为直接修复和取代修复两大类。

60
1、直接修复: (1)光复活: 修复任何嘧啶二聚体的损伤,修复过程为:光复活酶识别嘧啶二聚体并与之结合形成复合物→在300~600nm可见光照射下,酶获得能量,将嘧啶二聚体的丁酰环打开,使之完全修复→光复活酶从DNA上解离。 (2)转甲基作用: 在转甲基酶的催化下,将DNA上的被修饰的甲基去除,转甲基酶自身被甲基化而失活。 (3)直接连接: DNA断裂形成的缺口,可以在DNA连接酶的催化下,直接进行连接而封闭缺口。
转甲基作用: 在转甲基酶的催化下,将DNA上的被修饰的甲基去除,转甲基酶自身被甲基化而失活。 (3)直接连接: DNA断裂形成的缺口,可以在DNA连接酶的催化下,直接进行连接而封闭缺口。")
62
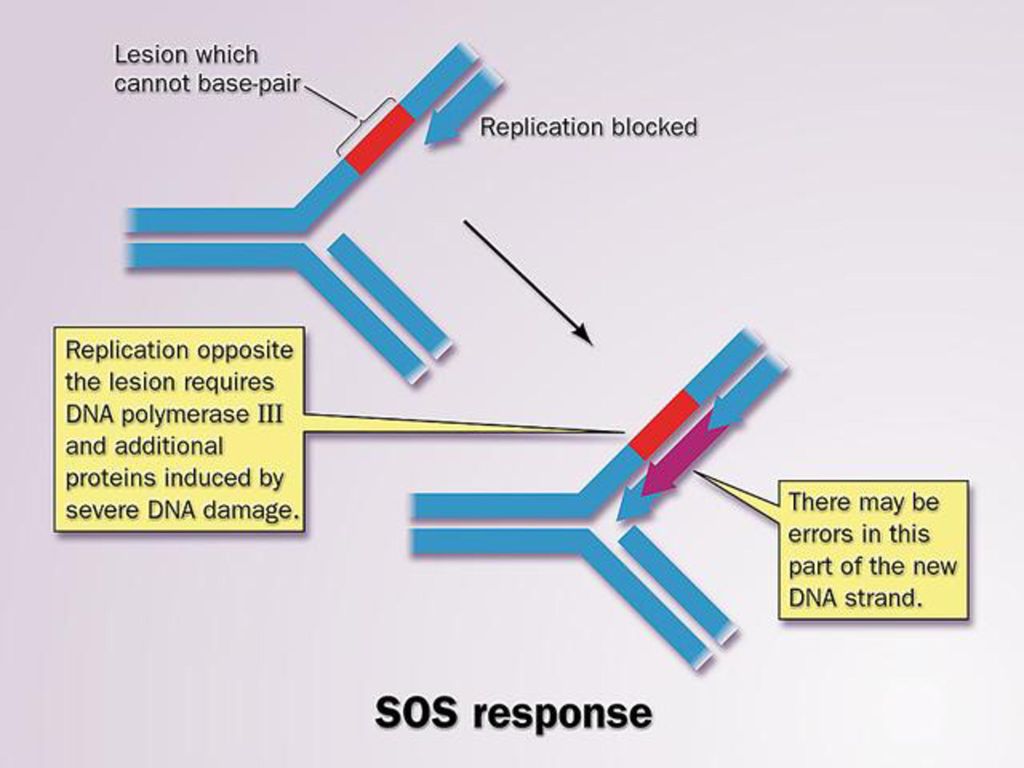
2、取代修复: (1)切除修复: 适用于多种DNA损伤的修复。该修复机制可以分别由两种不同的酶来发动,一种是核酸内切酶,另一种是DNA糖苷酶。 (2)重组修复: 是DNA的复制过程中所采用的一种有差错的修复方式。 (3)SOS修复: 一种在DNA分子受到较大范围损伤并且使复制受到抑制时出现的修复机制。 损伤诱导一种特异性较低的新的DNA聚合酶,以及重组酶等的产生。 由这些特异性较低的酶继续催化损伤部位DNA的复制,复制完成后,保留许多错误的碱基,从而造成突变。
SOS修复: 一种在DNA分子受到较大范围损伤并且使复制受到抑制时出现的修复机制。 损伤诱导一种特异性较低的新的DNA聚合酶,以及重组酶等的产生。 由这些特异性较低的酶继续催化损伤部位DNA的复制,复制完成后,保留许多错误的碱基,从而造成突变。")
65
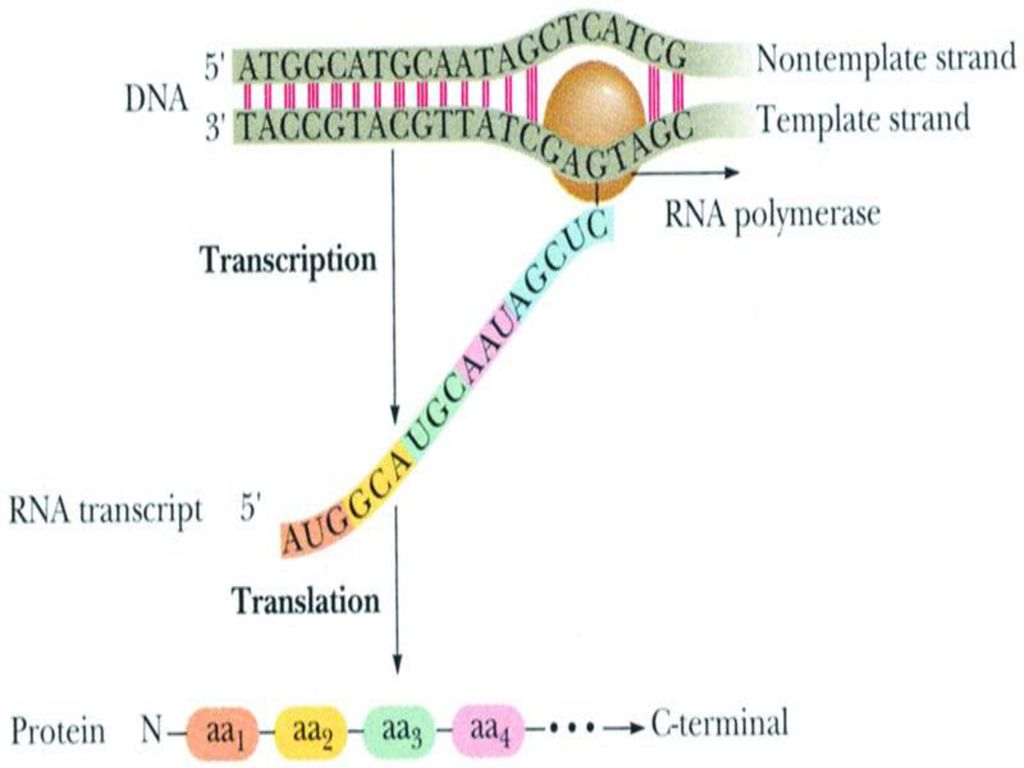
第四节 RNA的转录特点 一、转录(transcription)的定义 二、特点:
在RNA聚合酶催化下,以一段DNA链为模板合成RNA并将DNA所携带的遗传信息传递给RNA的过程称为转录。 经转录生成的RNA包括rRNA,tRNA,mRNA,snRNA等。 二、特点: 1.合成体系: (1)模板:以一段单链DNA作为模板(转录单位)。 (2)原料:游离的NTP,即ATP、UTP、CTP、GTP。 (3)酶:转录酶即RNA聚合酶,无校对功能。 (4)蛋白因子:终止因子ρ蛋白、激活因子CAP、转录因子TF等。 (5)无机离子:Mg2+、Mn2+。
模板:以一段单链DNA作为模板(转录单位)。 (2)原料:游离的NTP,即ATP、UTP、CTP、GTP。 (3)酶:转录酶即RNA聚合酶,无校对功能。 (4)蛋白因子:终止因子ρ蛋白、激活因子CAP、转录因子TF等。 (5)无机离子:Mg2+、Mn2+。")
66
2.方式 — 不对称转录 转录的不对称性是指以双链DNA中的一条链作为模板进行转录,从而将遗传信息由DNA传递给RNA。 对于不同的基因,转录信息可以存在于两条不同的DNA链上。能够转录RNA的那条DNA链称为模板链 (非信息链,反意义链),而与之互补的另一条DNA链称为编码链(信息链、有意义链)。
,而与之互补的另一条DNA链称为编码链(信息链、有意义链)。")
67
3.转录方向:5’-3’ RNA转录合成时,所依赖的模板DNA链的方向为3'→5',而RNA链的合成方向为5'→3'。 4、转录的连续性 RNA转录合成时,以DNA作为模板,在RNA聚合酶的催化下,连续合成一段RNA链,各条RNA链之间无需再进行连接。 合成的RNA中,如只含一个基因的遗传信息,称为单顺反子; 如含有几个基因的遗传信息,则称为多顺反子。 顺反子(cistron):是表示基因功能的单位,是一种结构基因,是为多肽链编码的一段染色体。
:是表示基因功能的单位,是一种结构基因,是为多肽链编码的一段染色体。")
68
特定起始点和特定终止点之间的DNA链构成一个转录单位,通常由转录区和有关的调节顺序构成。
5、特定的起始和终止位点 特定起始点和特定终止点之间的DNA链构成一个转录单位,通常由转录区和有关的调节顺序构成。 1

69
位于基因上游,与RNA聚合酶识别、结合并形成转录起始复合物的相关DNA顺序称为启动子(promoter),本身不被转录。通常还包括促进这一过程的调节物的结合点。(约70~80bp,p555图17-2)
包 括: a、转录起始部位,指导转录第一个核苷酸的碱基对,常为嘌呤,+1bp。 b、识别部位:RNA聚合酶识别应转录的DNA模板所必需,位于转录起始点-35区的5’- TTGACA序列-3’ 。 c、结合部位:是与RNA聚合酶结合的部位,位于-10区的TATAAT序列(Pribnow盒)。真核生物的转录起始区上游也存在一段富含TA的顺序,被称为Hogness盒或TATA盒(-25区)。另外在-75区存在CAAT盒,为5’- GCCAATCT序列-3’ 。在-100区存在增强子等。
。真核生物的转录起始区上游也存在一段富含TA的顺序,被称为Hogness盒或TATA盒(-25区)。另外在-75区存在CAAT盒,为5’- GCCAATCT序列-3’ 。在-100区存在增强子等。")
70
原核生物中转录起始区的共同序列

71
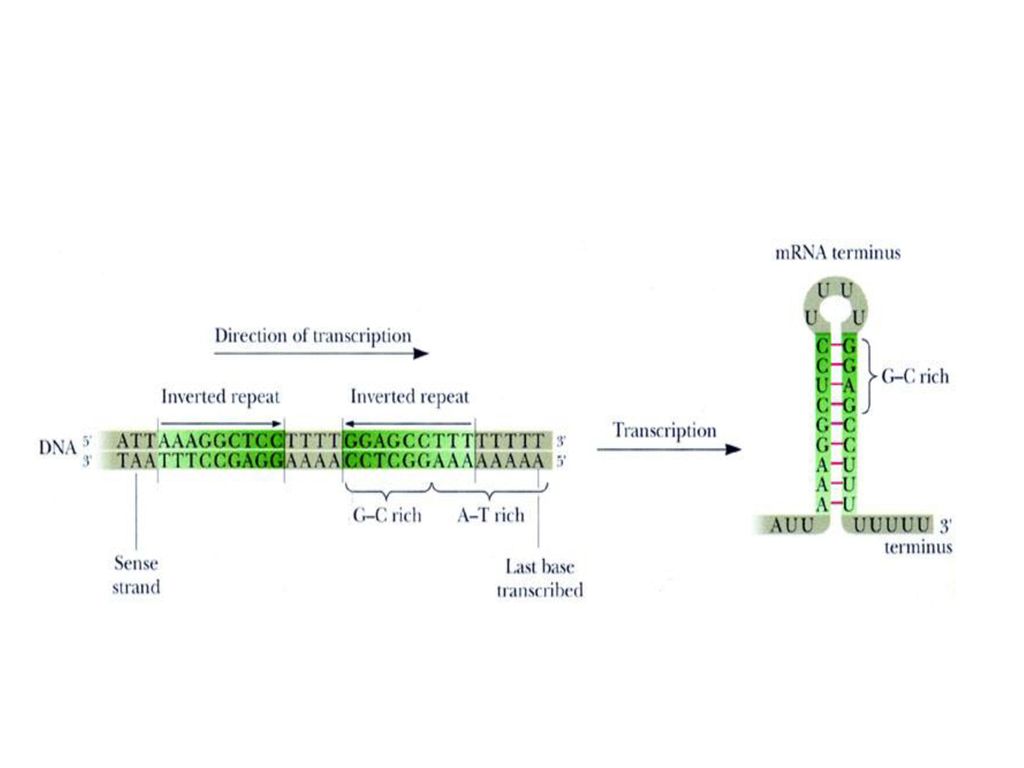
转录终止信号: 终止子(terminator):即终止信号,指DNA上促进转录终止的一段核苷酸序列,给RNA聚合酶提供终止信号,并发出RNA链延伸终止的信号。 (1)以蛋白因子ρ识别终止信号。 (2)DNA终止信号:含倒转重复顺序的GC丰富区,后随富含AT节段。生成的RNA可形成茎环结构/回文结构,有抗水解和保护作用,同时与DNA的AT节段作用弱,便于转录终了释放产物RNA。P556图17-3。
DNA终止信号:含倒转重复顺序的GC丰富区,后随富含AT节段。生成的RNA可形成茎环结构/回文结构,有抗水解和保护作用,同时与DNA的AT节段作用弱,便于转录终了释放产物RNA。P556图17-3。")
74
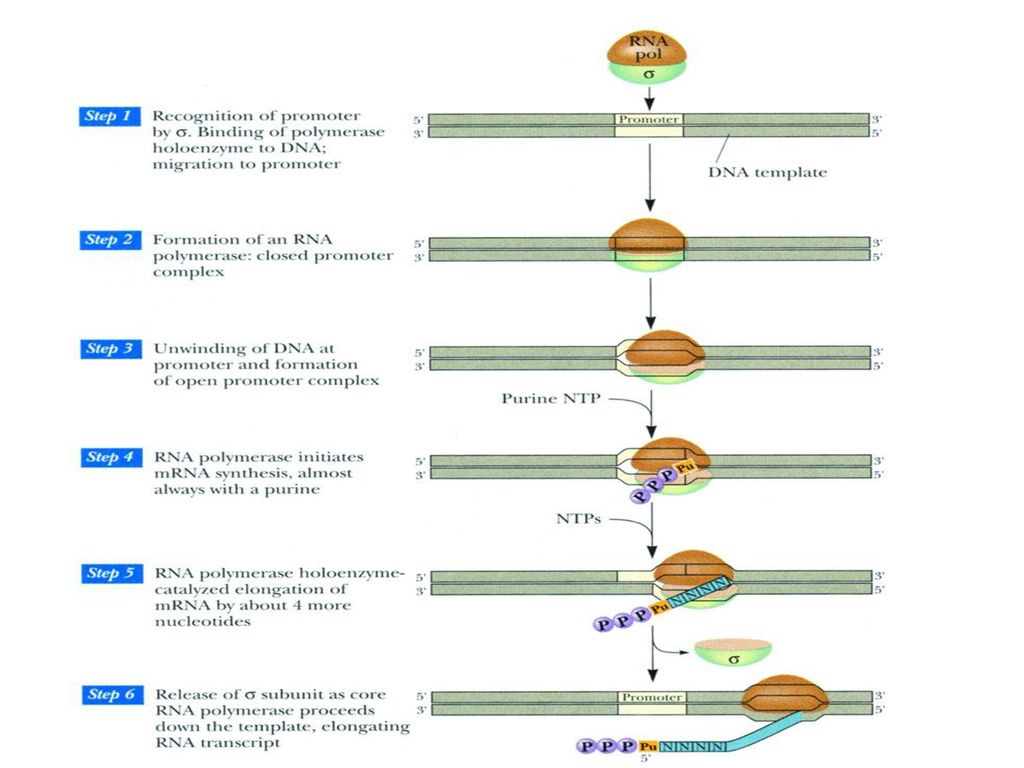
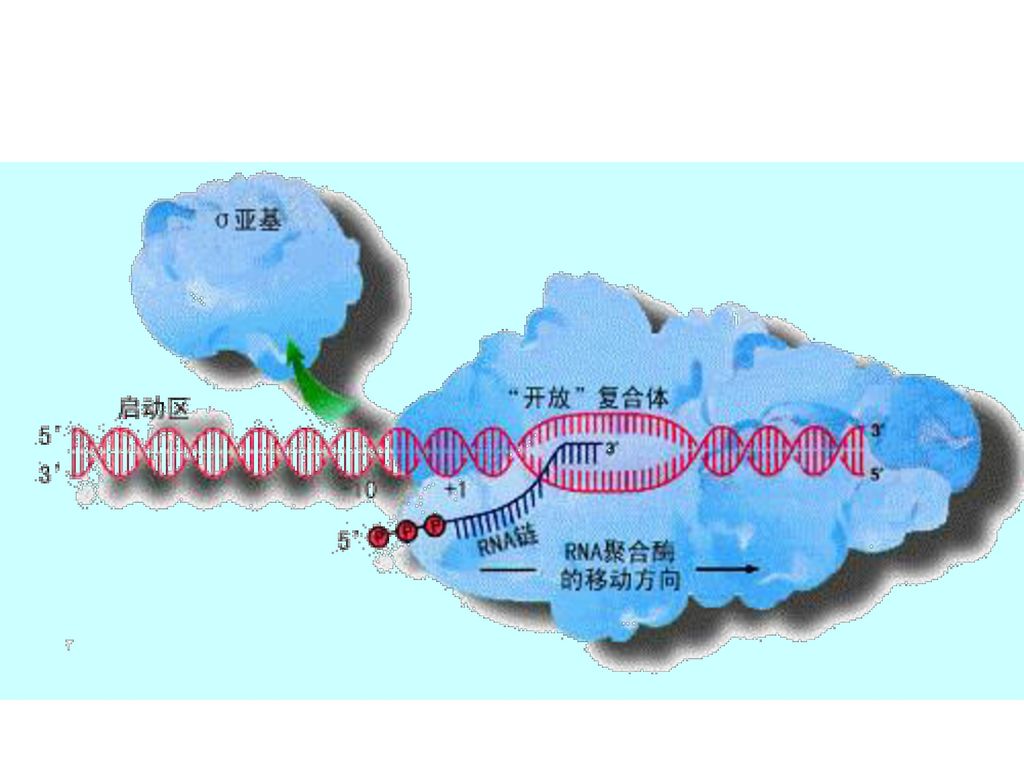
三、催化RNA合成的酶 1、RNA聚合酶(DDRP)
在单链DNA模板以及四种核糖核苷酸存在的条件下,不需要引物,即可从5’→3’聚合RNA,催化各种RNA的合成。 原核生物中的RNA聚合酶全酶(分子量约为465000)由六个亚基构成,即α2ββ’ωσ,含两个Zn2+。 σ亚基与转录起始点的识别(分子量大约是32000~92000,p460表36-3)有关,而在转录合成开始后被释放,余下的部分(α2ββ’ )被称为核心酶(分子量大约是 ),与RNA链的聚合有关。 (p554表17-1)
由六个亚基构成,即α2ββ’ωσ,含两个Zn2+。 σ亚基与转录起始点的识别(分子量大约是32000~92000,p460表36-3)有关,而在转录合成开始后被释放,余下的部分(α2ββ’ )被称为核心酶(分子量大约是 ),与RNA链的聚合有关。 (p554表17-1)")
75
大肠杆菌RNA聚合酶全酶

76
α-鹅膏蕈碱是真核细胞RNA聚合酶的抑制剂。按其对α-鹅膏蕈碱敏感性分为三种,均由10~12个大小不同的亚基所组成,结构复杂,功能不同,分子量约为50万,含锌。(p557表17-2)
")
77
酵母RNA聚合酶Ⅰ和Ⅱ

78
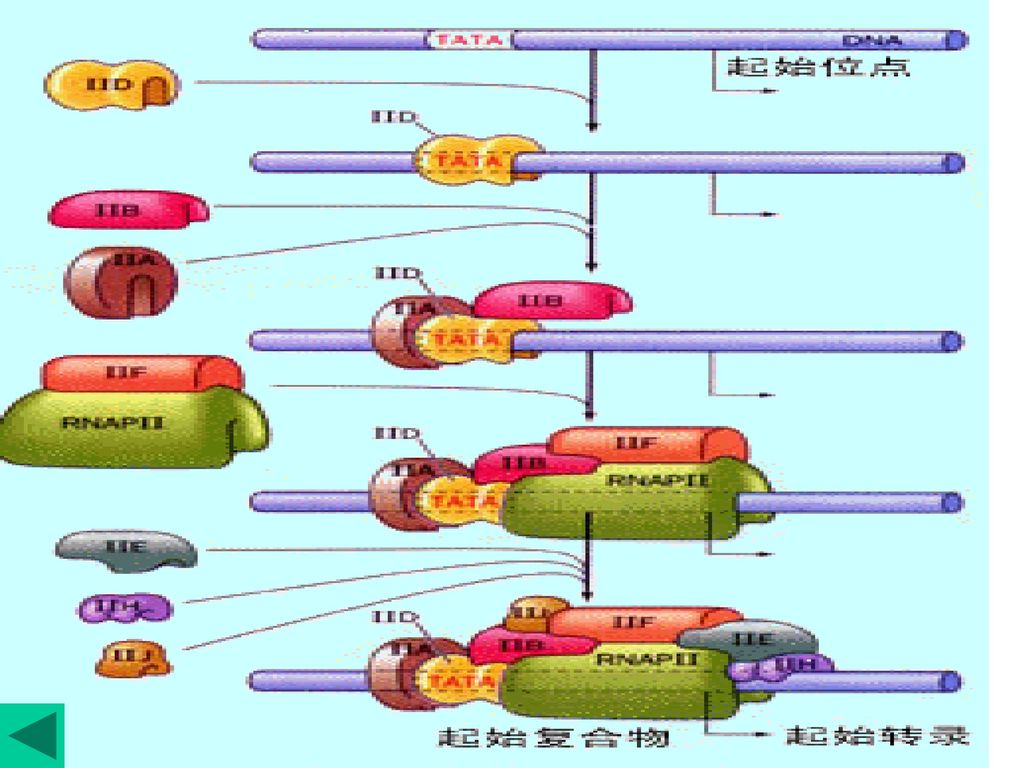
反式作用因子,是直接或间接辨认、结合启动子及聚合酶的调控基因转录效率的DNA结合蛋白质,如TF。
真核生物中RNA聚合酶需要蛋白因子(称为转录因子,transcriptional factor, TF)装配形成参与转录复合体,才能起始转录。如酶Ⅱ至少有六种不同的TF。 转录因子 功 能 TFⅡA 稳定TFⅡD结合 TFⅡB 促进polⅡ结合在模板 TFⅡD 辨认TATA盒 TFⅡE ATPase TFⅡF 解链酶,结合polⅡ TFⅡH ATP酶、解螺旋酶、激酶 反式作用因子,是直接或间接辨认、结合启动子及聚合酶的调控基因转录效率的DNA结合蛋白质,如TF。 顺式作用元件:对基因表达有调节活性的DNA序列,只 影响与其同处一个DNA分子上的基因,不编码蛋白质,如内含子。
装配形成参与转录复合体,才能起始转录。如酶Ⅱ至少有六种不同的TF。 转录因子 功 能. TFⅡA 稳定TFⅡD结合. TFⅡB 促进polⅡ结合在模板. TFⅡD 辨认TATA盒. TFⅡE ATPase. TFⅡF 解链酶,结合polⅡ. TFⅡH ATP酶、解螺旋酶、激酶. 反式作用因子,是直接或间接辨认、结合启动子及聚合酶的调控基因转录效率的DNA结合蛋白质,如TF。 顺式作用元件:对基因表达有调节活性的DNA序列,只. 影响与其同处一个DNA分子上的基因,不编码蛋白质,如内含子。")
79
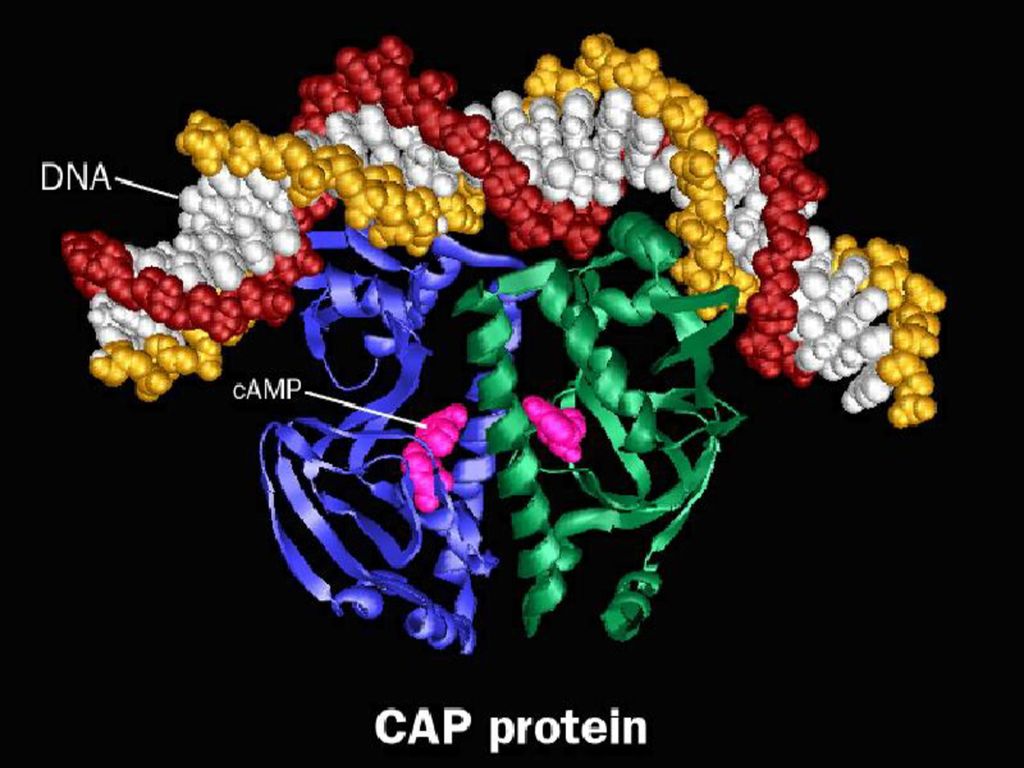
2、蛋白因子: (1)终止因子 ρ蛋白:分子量为50kd的蛋白质,多以六聚体形式存在。有ATP酶和解链酶的活性,与RNA结合后其构象发生变化,使RNA-DNA杂化双链变性转录产物释放。 Nus A蛋白:分子量69kd的酸性蛋白,在延长过程中与核心酶结合,并在终止部位使RNA聚合酶停顿及释放RNA。 (2)激活因子 降解产物基因激活蛋白(catabolite gene activator protein,CAP,又称cAMP受体蛋白,cyclic AMP receptor protein,CRP):二聚体蛋白质,亚基分子量为23kd,含DNA结合区和转录活化区。该蛋白与cAMP结合后,可结合到受调节的启动子,刺激RNA聚合酶与起始部位结合,从而起始转录过程。
激活因子. 降解产物基因激活蛋白(catabolite gene activator protein,CAP,又称cAMP受体蛋白,cyclic AMP receptor protein,CRP):二聚体蛋白质,亚基分子量为23kd,含DNA结合区和转录活化区。该蛋白与cAMP结合后,可结合到受调节的启动子,刺激RNA聚合酶与起始部位结合,从而起始转录过程。")
81
第五节 转录的过程 一、起始 识别起始位点:原核生物RNA聚合酶中的σ因子识别转录起始点,并促使核心酶结合形成全酶复合物。
RNA聚合酶全酶促使局部双链解开,并催化ATP或GTP与另外一个三磷酸核苷聚合,形成第一个3’,5’-磷酸二酯键。 利福霉素通过直接与细菌中的RNA聚合酶的亚基结合来抑制RNA合成,特异地抑制第一个磷酸二酯键的形成,抑制转录起始。利福平已用作为临床的抗结核菌药。

84
二、延长 σ因子从全酶上脱离,余下的核心酶继续沿DNA链移动,按照碱基互补原则,不断聚合RNA。 E.Coli RNA聚合酶催化的延伸速率大约是每秒钟50个核苷酸。 放线菌素D可插入到相邻的G-C碱基对之间,与鸟嘌呤形成特殊的氢键,结构中的环状多肽占据了DNA双螺旋的空间,抑制DNA模板的功能,有效地抑制原核生物和真核生物转录的延伸过程。

85
RNA聚合酶催化的延伸反应

87
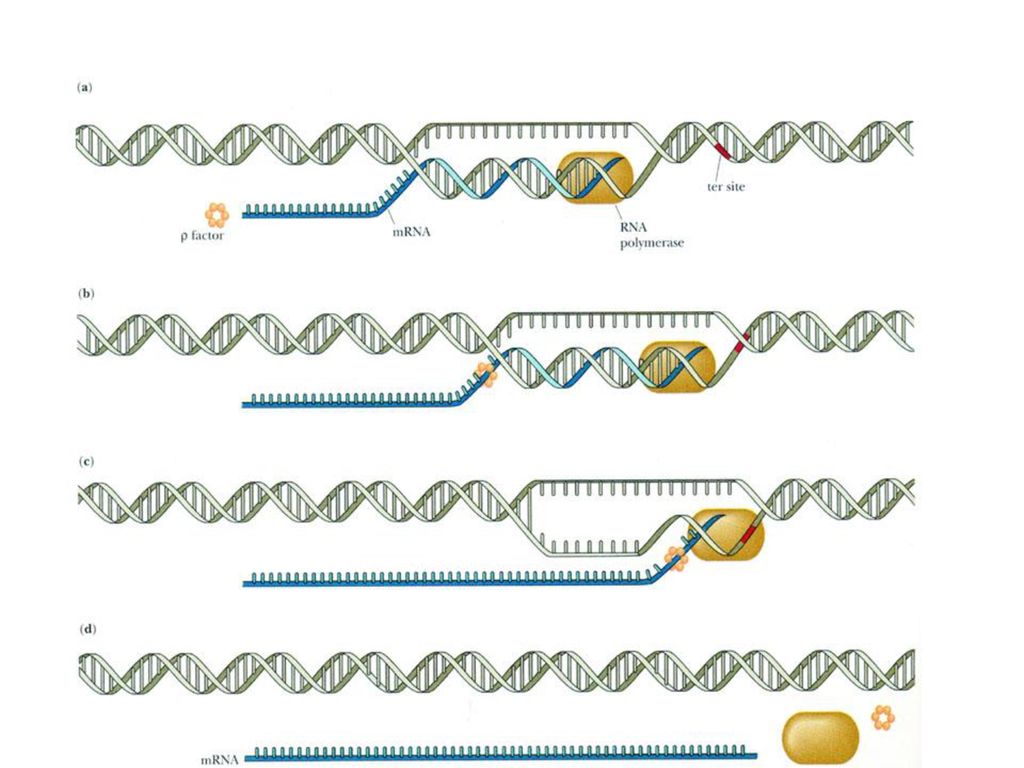
三、终止 RNA转录合成的终止机制有两种:
自动终止:模板DNA链在接近转录终止点处存在相连的富含GC和AT的区域,使RNA转录产物形成寡聚U及发夹形的二级结构,引起RNA聚合酶变构及移动停止,导致RNA转录的终止。

88
依赖辅助因子的终止:终止蛋白与RNA聚合酶复合体相互作用恰好终止在一个发卡环处,发卡环是在新合成的转录链上形成的,由ρ因子识别特异的终止信号,并促使RNA的释放。

89
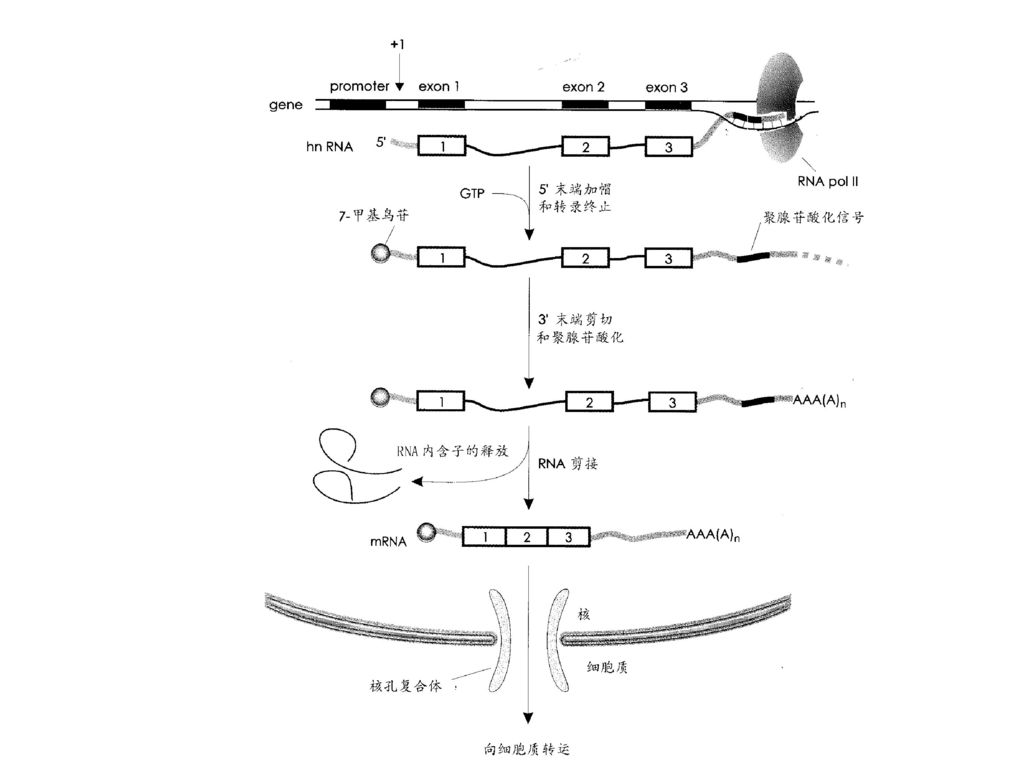
第六节 真核生物RNA转录后的修饰 一、mRNA的转录后加工(细胞核内) 1.加帽(adding cap):
在hnRNA的5’-端加上m7G5’ppp的结构。5’ 帽子对于蛋白质合成的起始是很重要的,同时保护mRNA转录后不被核酸外切酶降解。 2.加尾(adding tail): 由核酸外切酶切去3’- 端一些过剩的核苷酸,再由多聚腺苷酸聚合酶(PAP)催化加入polyA。聚腺苷酸化反应是一个重要的调节步骤,因为聚腺苷酸尾巴的长度能调节mRNA的稳定性和翻译效率。
: 由核酸外切酶切去3’- 端一些过剩的核苷酸,再由多聚腺苷酸聚合酶(PAP)催化加入polyA。聚腺苷酸化反应是一个重要的调节步骤,因为聚腺苷酸尾巴的长度能调节mRNA的稳定性和翻译效率。")
90
3.剪接(splicing): 真核生物结构基因是断裂基因,包含外显子,内含子。 内含子(编码链)的左端(5’端)均为GT,右端(3’端)均为AG,对应于RNA为GU-AG。 真核生物HnRNA的剪接一般需snRNA(100~300个核苷酸)参与构成的核蛋白体(snRNP,核内小核糖核蛋白)参加,通过形成套索状结构而将内含子切除掉。目前存在5种snRNP:U1 snRNP,U2 snRNP,U5 snRNP和U4-U6 snRNP,它们与内含子形成剪接复合体。U3snRNP与rRNA前体的加工有关。
参与构成的核蛋白体(snRNP,核内小核糖核蛋白)参加,通过形成套索状结构而将内含子切除掉。目前存在5种snRNP:U1 snRNP,U2 snRNP,U5 snRNP和U4-U6 snRNP,它们与内含子形成剪接复合体。U3snRNP与rRNA前体的加工有关。")
92
聚腺苷酸化作用断裂特异因子(CPSF) 断裂刺激因子(CSTF)
 断裂刺激因子(CSTF)")
93
通过RNA-DNA杂化检测出DNA模板链中的内含子

95
转酯基反应 P562图17-10

97
二、tRNA的转录后加工 (1)剪 接: 核酸内切酶,外切酶作用,切去5’-端,3’-端一定核苷酸及相当于反密码环的部分核苷酸(14-60)插入序列。 (2)加成: 3’-端加-CCA (3)修 饰: tRNA中稀有碱基的生成,在蛋白质合成过程中影响tRNA对密码的识别。 甲基化:A Am 还原加氢:U DHU 碱基转位:U 脱氨:A I
修 饰: tRNA中稀有碱基的生成,在蛋白质合成过程中影响tRNA对密码的识别。 甲基化:A Am 还原加氢:U DHU 碱基转位:U 脱氨:A I.")
99
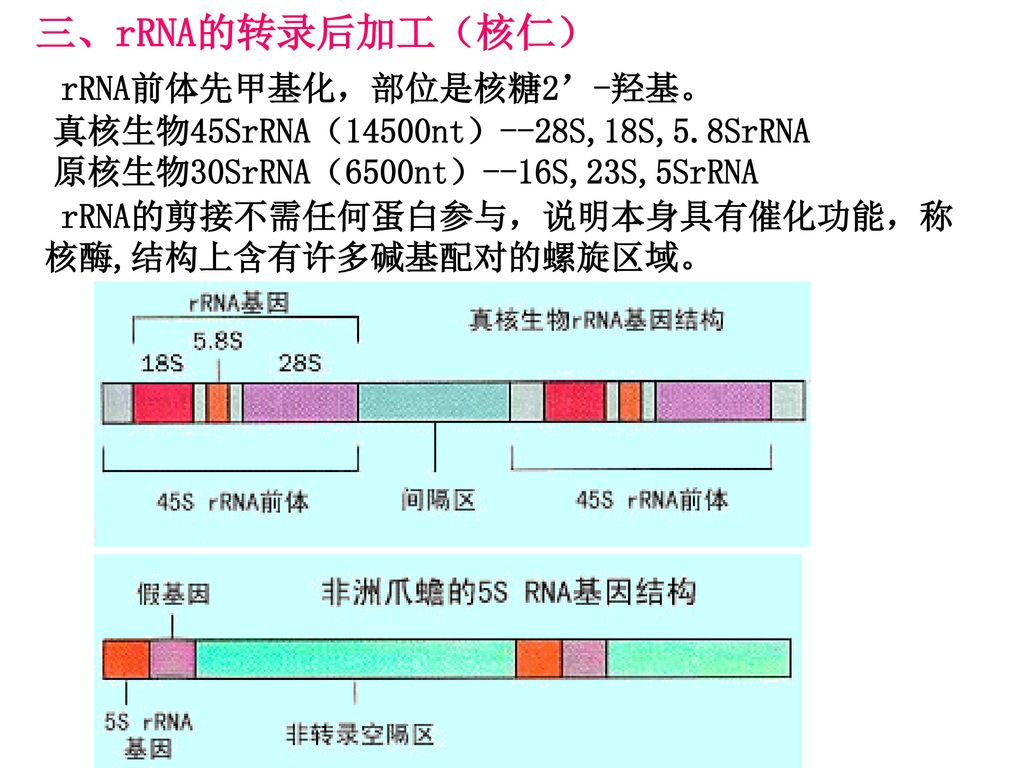
三、rRNA的转录后加工(核仁) rRNA前体先甲基化,部位是核糖2’-羟基。
真核生物45SrRNA(14500nt)--28S,18S,5.8SrRNA 原核生物30SrRNA(6500nt)--16S,23S,5SrRNA rRNA的剪接不需任何蛋白参与,说明本身具有催化功能,称核酶,结构上含有许多碱基配对的螺旋区域。
--28S,18S,5.8SrRNA. 原核生物30SrRNA(6500nt)--16S,23S,5SrRNA. rRNA的剪接不需任何蛋白参与,说明本身具有催化功能,称核酶,结构上含有许多碱基配对的螺旋区域。")
100
人45S rRNA加工成28S.5.8S和18S 有功能的rRNA的过程

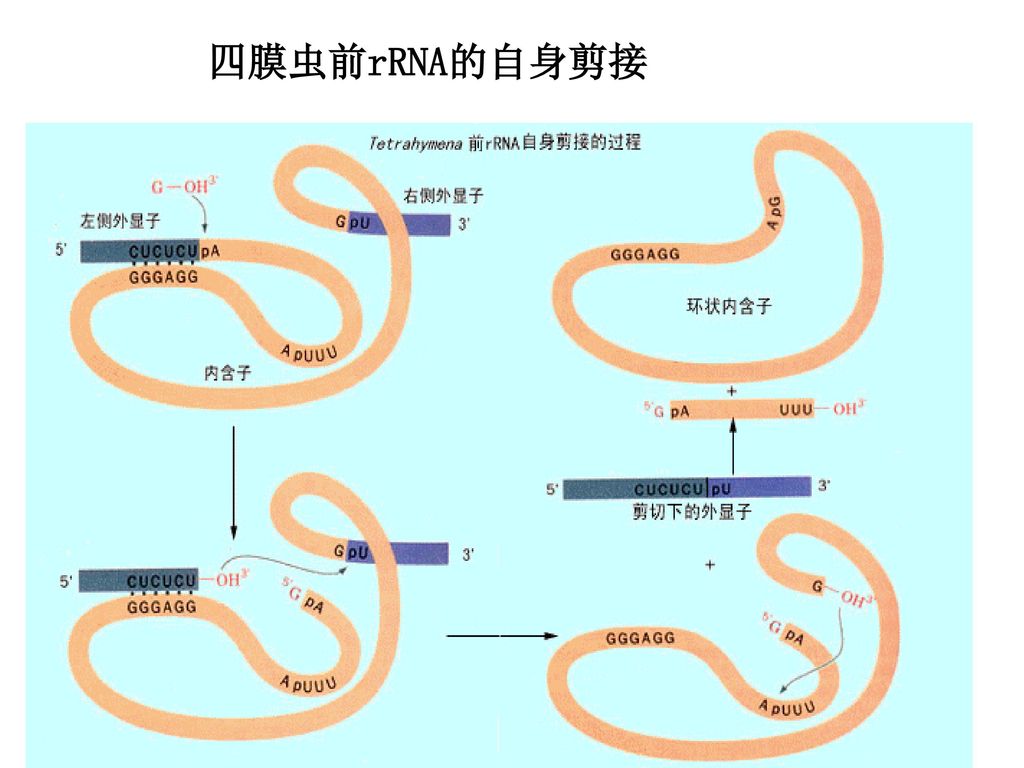
101
四膜虫前rRNA的自身剪接

102
基因工程ftz锤头核酶和其作用的靶序列:fushi taruzu(ftz)mRNA的相关序列
mRNA的相关序列")
103
第七节 蛋白质合成的特点 一、翻译(translation)的定义 二、蛋白质合成体系
即蛋白质的生物合成过程,就是将DNA传递给mRNA的遗传信息即核苷酸顺序,再具体的解译为蛋白质中氨基酸排列顺序的过程,是两种分子语言的转换。 二、蛋白质合成体系 1、原料:20种氨基酸 2、各种RNA:mRNA、tRNA、rRNA 3、酶及蛋白因子:氨基酰-tRNA合成酶、转肽酶、IF、EF、RF 等 4、供能物质及无机离子: ATP、GTP、Mg2+、K+

104
三、几种RNA的功能 1、mRNA 遗传信息的携带者,是蛋白质合成的直接模板。
mRNA分子上按5’→3’方向每三个相邻的核苷酸组成三联体,代表一个氨基酸的信息,此三联体就称为密码子( coden ),是蛋白质合成中某一特定氨基酸的编码单位。 遗传密码的特点:①方向性,即解读方向为5′→ 3′;②连续性;③简并性;④摆动性;⑤通用性(线粒体或叶绿体例外);⑥ 起始密码:AUG(原核:GUG、UUG)终止密码:UAA、UAG、UGA。
,是蛋白质合成中某一特定氨基酸的编码单位。 遗传密码的特点:①方向性,即解读方向为5′→ 3′;②连续性;③简并性;④摆动性;⑤通用性(线粒体或叶绿体例外);⑥ 起始密码:AUG(原核:GUG、UUG)终止密码:UAA、UAG、UGA。")
105
M. Nirenberg Jones,Khorana 证明三联体密码的三个著名实验的示意图 p569

106
1966年,全部密码都被破译。共有64个密码,其中61个密码是编码氨基酸的,其余3种是终止密码(无义密码)。
。")
108
遗传密码的连续性:密码之间无任何标点隔开,必须连续三个一组读码翻译。
例: 5’…GCUAUAUGAA…3’ 丙 异亮 半胱 ’…GCUAAUGUA…3’ 丙 天冬 缬 酰胺 移码

109
遗传密码阅读时一般不重叠: 开放阅读框:指DNA或RNA分子中一组连续的不重叠的编码序列(不包括终止密码)。
。")
111
遗传密码的简并性(degenerate):一种氨基酸可由两种或更多密码子编码。
例如:Ser有6个密码,Gly有4个密码,Lys有2个密码。 只有一个密码子的氨基酸是Trp和Met。 同义密码:编码同一种氨基酸的密码子。 一个密码的头两个核苷酸残基确定一个给定的氨基酸。 例:Gly的密码有4个,都开始于GG:GGU,GGC,GGA和GGG。 意义:1)减少碱基突变造成的有害效应。2)具有类似序列的密码往往指定化学性质类似的氨基酸。 例:Asp和Glu的密码开始于GA,仅第3位核苷酸不同。
减少碱基突变造成的有害效应。2)具有类似序列的密码往往指定化学性质类似的氨基酸。 例:Asp和Glu的密码开始于GA,仅第3位核苷酸不同。")
112
按5’ → 3’方向,mRNA 1 → tRNA 3,mRNA 2 → tRNA 2, mRNA 3 → tRNA 1.
遗传密码的摆动性(变偶性,wobble):翻译过程中氨基酸正确加入需靠mRNA上的密码与tRNA上的反密码互相辨认,密码的第三位碱基与反密码的第一位碱基不完全按碱基互补规律配对,具有较大的灵活性,又称密码的变偶性。减少密码阅读时的误差,增加翻译的准确性。 按5’ → 3’方向,mRNA 1 → tRNA 3,mRNA 2 → tRNA 2, mRNA 3 → tRNA 1. 反密码5’端碱基 G C A U I 密码3’端碱基 U , C G, C, U U A, G A, U, C
:翻译过程中氨基酸正确加入需靠mRNA上的密码与tRNA上的反密码互相辨认,密码的第三位碱基与反密码的第一位碱基不完全按碱基互补规律配对,具有较大的灵活性,又称密码的变偶性。减少密码阅读时的误差,增加翻译的准确性。 按5’ → 3’方向,mRNA 1 → tRNA 3,mRNA 2 → tRNA 2, mRNA 3 → tRNA 1. 反密码5’端碱基. G. C. A. U. I. 密码3’端碱基. U , C. G, C, U. U. A, G. A, U, C.")
114
遗传密码的通用性:所有的生物包括真核的与原核的,均共用一套密码。只是动物细胞线粒体,植物细胞叶绿体中密码并不完全相同。
例: 通用密码 线粒体 UGA 终止密码 Trp AUA Ile 起始密码(Met/Ile) AUG 起始密码 起始密码(Met) AUU Ile 起始密码(Ile) CUA Leu Thr AGA Arg 终止密码 AGG Arg 终止密码
 AUG 起始密码 起始密码(Met) AUU Ile 起始密码(Ile) CUA Leu Thr. AGA Arg 终止密码. AGG Arg 终止密码.")
115
2、tRNA 起接合器作用,借助于tRNA将氨基酸线性排列在mRNA模板上,将其碱基序列转换为氨基酸序列。
氨基酸结合位点:3’-末端CCA-OH。一种氨基酸可以和2--6种tRNA特异结合.结合相同氨基酸而反密码子不同的一组tRNA分子称为同工tRNA(isoacceptor tRNA)。 识别模板的部位:tRNA反密码环中部的三个核苷酸构成三联体,识别mRNA上相应的密码,此三联体就称为反密码(anticoden)。密码子与反密码子是反向平行配对的。 核糖体识别位点:tRNA分子链中-GTC-(TC环)核苷酸序列,起着将多肽链连接在核糖体的作用。
。 识别模板的部位:tRNA反密码环中部的三个核苷酸构成三联体,识别mRNA上相应的密码,此三联体就称为反密码(anticoden)。密码子与反密码子是反向平行配对的。 核糖体识别位点:tRNA分子链中-GTC-(TC环)核苷酸序列,起着将多肽链连接在核糖体的作用。")
116
在原核生物和真核生物中,均存在另一种携带蛋氨酸的tRNA,识别非起始部位的蛋氨酸密码AUG,即tRNAeMet。
能够识别mRNA中5′端起始密码AUG的tRNA是一种特殊的tRNA,称为起始tRNA。在原核生物中,起始tRNA是一种携带甲酰蛋氨酸的tRNA,即tRNAfMet;而在真核生物中,起始tRNA是一种携带蛋氨酸的tRNA,即tRNAiMet。 在原核生物和真核生物中,均存在另一种携带蛋氨酸的tRNA,识别非起始部位的蛋氨酸密码AUG,即tRNAeMet。 fMet-tRNAfMet

117
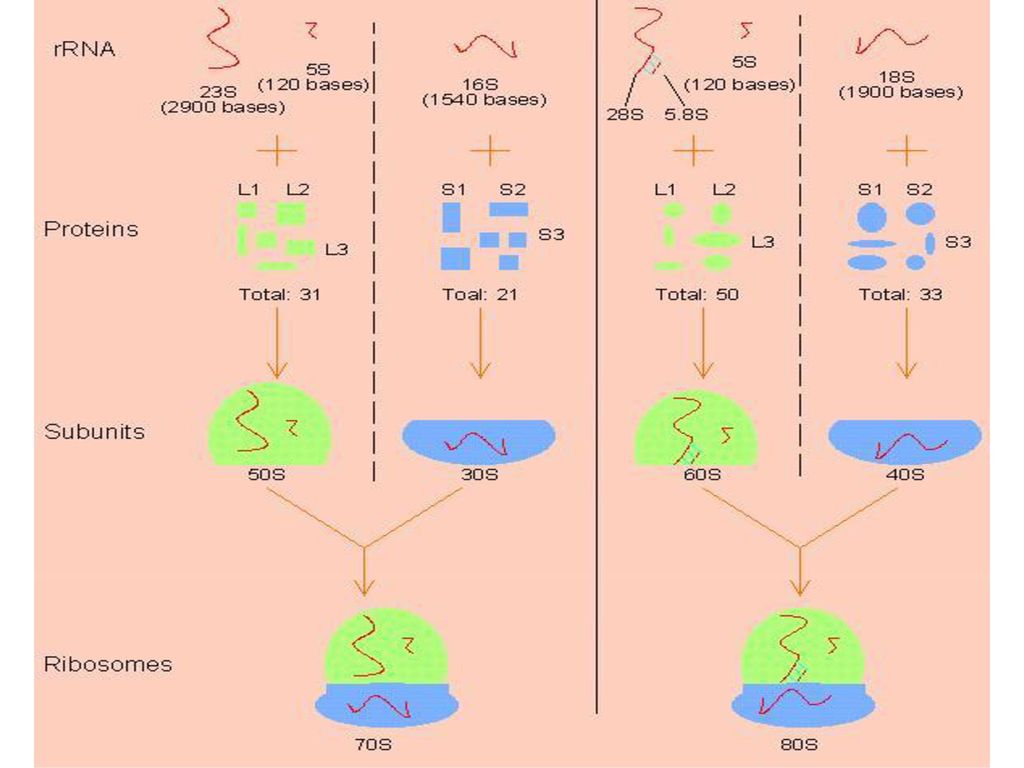
3、rRNA和核蛋白体 核蛋白体是蛋白质生物合成的场所,是一种无膜的 细胞器。 E.coli核糖体(70S) 高等动物核糖体(80S)
30S亚基 S亚基 S亚基 60S亚基 16SRNA SRNA SRNA SRNA 5SRNA SRNA 21种蛋白质 种蛋白质 SRNA 33种蛋白质 种蛋白质

119
大肠杆菌核蛋白体

120
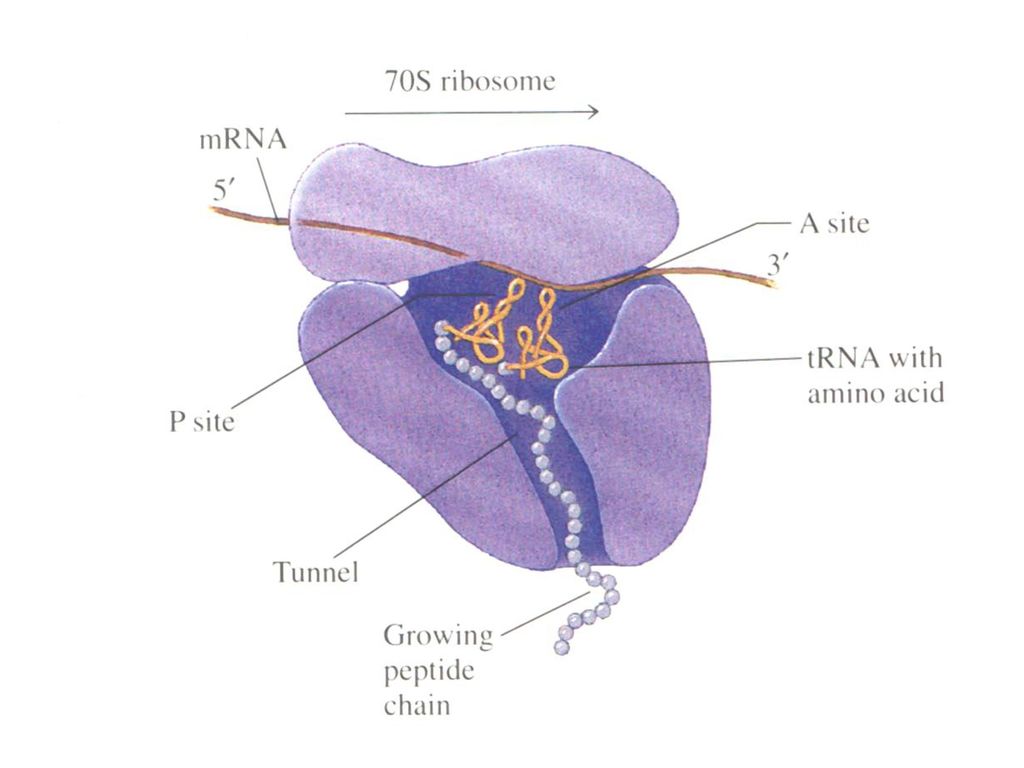
核蛋白体的大、小亚基分别有不同的功能: 1.小亚基:与mRNA、GTP和起始tRNA结合。 16srRNA3’末端有一富含嘧啶特异序列-UCCU-,与mRNA起始密码上游含嘌呤碱基的区域-AGGA-序列(SD序列, Shine-Dalgarno siquence )配对识别。(p575图18-2) 2.大亚基: 1)具有两个不同的tRNA结合点:A位(右),即受位或氨酰基位,与新进入的氨基酰tRNA结合;P位(左),即给位或肽酰基位,与延伸中的肽酰基tRNA结合。 2)具有起始因子、延长因子及释放因子的出入部位。 3)具有转肽酶活性:将给位上的肽酰基转移给受位上的氨基酰tRNA,形成肽键。 4)具有GTPase活性,水解GTP,获得能量。
配对识别。(p575图18-2) 2.大亚基: 1)具有两个不同的tRNA结合点:A位(右),即受位或氨酰基位,与新进入的氨基酰tRNA结合;P位(左),即给位或肽酰基位,与延伸中的肽酰基tRNA结合。 2)具有起始因子、延长因子及释放因子的出入部位。 3)具有转肽酶活性:将给位上的肽酰基转移给受位上的氨基酰tRNA,形成肽键。 4)具有GTPase活性,水解GTP,获得能量。")
121
E.coli mRNA的SD序列

124
四、参与蛋白质合成的酶及蛋白因子 1、氨基酰tRNA合成酶
每种氨基酰tRNA合成酶对相应氨基酸以及携带氨基酸的tRNA具有高度特异性。 该酶对tRNA的识别,是因为在tRNA的氨基酸臂上存在特定的识别密码,即第二套遗传密码。 在催化反应中,特异的tRNA3’端CCA上的2’或3’位自由羟基与相应的活化氨基酸以酯键相连接,形成氨基酸tRNA。 具有纠错功能。

125
氨基酸+ATP+E 氨基酰-AMP-E+PPi 氨基酰-AMP-E+tRNA 氨酰~tRNA+AMP+E
氨基酸+tRNA +ATP →氨酰~tRNA+AMP+PPi

126
2、转肽酶 (p578图18-6)
")
127
3、起始因子(IF) 是与多肽链合成起始有关的蛋白因子。原核生物中存在3种起始因子,分别称为IF1-3。在真核生物中存在11种起始因子(eIF)。其作用主要是促进核蛋白体大小亚基解离以及小亚基与起始tRNA、模板mRNA结合。 4、延长因子(EF) 原核生物中存在3种延长因子(EFTU,EFTS,EFG),真核生物中存在2种(EF1,EF2)。其作用主要促使氨基酰tRNA进入核蛋白体的受位,并可促进移位过程。
 原核生物中存在3种延长因子(EFTU,EFTS,EFG),真核生物中存在2种(EF1,EF2)。其作用主要促使氨基酰tRNA进入核蛋白体的受位,并可促进移位过程。")
128
5、释放因子(RF) 原核生物中有3种,在真核生物中只有1种。其主要作用是识别终止密码,协助多肽链的释放。
 原核生物中有3种,在真核生物中只有1种。其主要作用是识别终止密码,协助多肽链的释放。")
129
第八节 蛋白质生物合成过程 一、氨基酸的活化与搬运 蛋白质生物合成过程包括三大步骤: ①氨基酸的活化与搬运 ②活化氨基酸在核蛋白体上的缩合
第八节 蛋白质生物合成过程 蛋白质生物合成过程包括三大步骤: ①氨基酸的活化与搬运 ②活化氨基酸在核蛋白体上的缩合 ③多肽链合成后的加工修饰 一、氨基酸的活化与搬运 由氨基酰tRNA合成酶催化氨基酰-tRNA的生成,活化1分子氨基酸,消耗2ATP。 氨基酰tRNA的合成,可使氨基酸活化、搬运、定位。

130
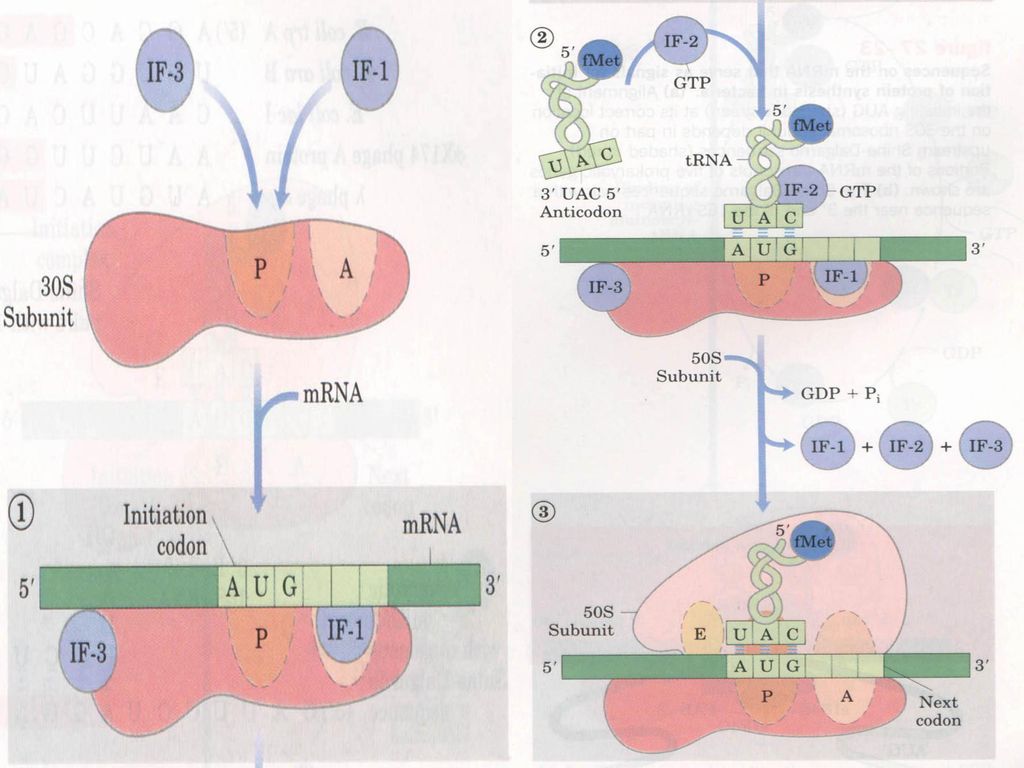
二、活化氨基酸的缩合——核蛋白体循环 即蛋白质的生物合成,以mRNA为模板,tRNA为氨基酸载体,在核蛋白体上通过进位,转肽,移位等过程反复翻译mRNA上的密码子并缩合生成多肽链的循环反应,称为核蛋白体循环。 核蛋白体循环分为起始、延长和终止三个阶段。 (一)起始阶段:起始复合体 由核糖体大亚基,小亚基,模板mRNA,起始蛋氨酰-tRNA(原核fMet-tRNAfmet,真核Met-tRNAimet)构成,此过程耗能,IF参与。 (p576图18-4)
起始阶段:起始复合体. 由核糖体大亚基,小亚基,模板mRNA,起始蛋氨酰-tRNA(原核fMet-tRNAfmet,真核Met-tRNAimet)构成,此过程耗能,IF参与。 (p576图18-4)")
132
1.30S起始复合物的形成:在IF1、IF3的促进下,30S小亚基与mRNA的起始部位结合。
原核mRNA起始部位的SD序列被核蛋白体小亚基辨认结合。 真核生物中的mRNA具有帽子结构,需一种特殊的帽子结合蛋白(CBP)识别。 2.70S前起始复合体的形成:IF-2-GTP结合在30S亚基上,形成了一个由fMet-tRNAfMet、IF-2-GTP及mRNA组成的前起始复合体。 3.70S起始复合体的形成:GTP被水解,IF1、IF3、IF2从复合物上脱落,50S大亚基与复合体结合,tRNAfmet的反密码CAU与mRNA上的起始密码AUG互补结合,tRNAfmet结合在核蛋白体的给位(P位)。
识别。 2.70S前起始复合体的形成:IF-2-GTP结合在30S亚基上,形成了一个由fMet-tRNAfMet、IF-2-GTP及mRNA组成的前起始复合体。 3.70S起始复合体的形成:GTP被水解,IF1、IF3、IF2从复合物上脱落,50S大亚基与复合体结合,tRNAfmet的反密码CAU与mRNA上的起始密码AUG互补结合,tRNAfmet结合在核蛋白体的给位(P位)。")
133
(二)肽链延长阶段: 1.进位: 与mRNA下一个密码相对应的氨基酰-tRNA进入核蛋白体的受位。此步骤需GTP,Mg2+,和EF参与。
肽链延长阶段: 1.进位: 与mRNA下一个密码相对应的氨基酰-tRNA进入核蛋白体的受位。此步骤需GTP,Mg2+,和EF参与。")
134
2.转肽: 在转肽酶的催化下,将给位上的tRNA所携带的甲酰蛋氨酰基或肽酰基转移到受位上的氨基酰tRNA上,与其α-氨基缩合形成肽键。此步骤需Mg2+,K+。给位上已失去蛋氨酰基或肽酰基的tRNA从核蛋白上脱落。

135
3.移位: 核蛋白体向mRNA的3’端滑动相当于一个密码的距离,同时使肽酰tRNA从受位移到给位。此步骤需EF(EFG)、GTP和Mg2+参与。此时,核蛋白体的受位留空,与下一个密码相对应的氨基酰tRNA即可再进入,重复以上循环过程,使多肽链不断延长。 在肽链延伸过程中,核糖体和有关的辅助因子沿着模板mRNA的5’→3’方向移动,而蛋白质合成是从N端向C端。

136
(三)肽链终止阶段: 核蛋白体沿mRNA链滑动,不断使多肽链延长,直到终止信号进入受位。 1、识别:RF识别终止密码,进入核蛋白体的受位。
2、水解:RF使转肽酶变为水解酶,多肽链与tRNA之间的酯键被水解,多肽链释放。 3、解离:通过水解GTP,使核蛋白体与mRNA分离,tRNA、RF脱落,核蛋白体解离为大、小亚基。

137
蛋白质合成是个耗能过程,氨基酸活化时需消耗2分子高能磷酸键,肽键形成时又消耗2分子高能磷酸键,故缩合一分子氨基酸残基需消耗4分子高能磷酸键。
在蛋白质生物合成过程中,常常由若干核蛋白体结合在同一mRNA分子上,同时进行翻译,但每两个相邻核蛋白之间存在一定的间隔,形成念球状结构。 由若干核蛋白体结合在一条mRNA上同时进行多肽链的翻译所形成的念球状结构称为多核蛋白体。

139
常见的蛋白质生物合成抑制剂: 嘌呤霉素的结构类似于氨酰-tRNA的3’末端的AMP结构,可以进入核糖体的A位。肽酰转移酶催化新生成的多肽转移至嘌呤霉素的游离的氨基上。由于肽酰-嘌呤霉素在A位处的结合弱,很快就从核糖体上解离,可终止蛋白质的合成。 四环素和原核生物核糖体的30S亚基结合,封住了A位点,阻止氨酰-tRNA进入A位。 氯霉素与50S的核糖体亚基相互作用,抑制肽酰转移酶,广泛用作治疗细菌感染的药物。 链霉素抑制蛋白质合成的起始,也能引起核糖体构象改变,使肽链延伸阶段的mRNA错读,引起翻译错误。 白喉毒素是已知的最有潜力的毒素,只要一分子的白喉毒素就足可以使真核细胞内的延伸因子eEF-2失活,导致死亡。 蓖麻毒蛋白是从蓖麻中分离出来的,其作用模式是通过切断N苷键除去腺嘌呤碱基使真核生物的28SrRNA失活,导致翻译终止。

140
真核生物蛋白质合成特点 核糖体更大 起始结构不同:起始密码子、帽子结构、单顺反子、起始tRNA。 辅助因子更多
合成过程:形成80S复合物、肽链的延伸、终止与释放。 转录与翻译过程分离

142
第九节 蛋白质合成后的加工修饰 一、一级结构的加工修饰: 1.N端甲酰蛋氨酸或蛋氨酸的切除:
① 去甲酰化: 甲酰化酶 甲酰蛋氨酸-肽 甲酸 + 蛋氨酸-肽 ② 去蛋氨酰基: 蛋氨酸氨基肽酶 蛋氨酰-肽 蛋氨酸 + 肽

143
2、切除信号肽:分泌型蛋白质末端有一段15~30氨基酸残基序列,可引导多肽到细胞的相应部分,称为信号肽。完成任务后可被特异的信号肽酶水解。
3、氨基酸的修饰: 由专一性的酶催化进行修饰,包括糖基化、羟基化、磷酸化、甲酰化等。 4、二硫键的形成: 由专一性的氧化酶催化,将-SH氧化为-S-S-。 5、肽段的切除: 由专一性的蛋白酶催化,将部分肽段切除。

144
二、高级结构的形成: 1.构象的形成:在分子内伴侣、辅助酶及分子伴侣的协助下,形成特定的空间构象。
分子伴侣:是指细胞中普遍存在的一组蛋白质,在与细胞中靶蛋白短暂结合的过程中改变蛋白质的空间构象从而调节蛋白质的功能。 2.亚基的聚合。 3.辅基的连接:如细胞色素C与血红素的结合、乙酰CoA羧化酶与生物素的结合。

145
第十节 基因表达的调控 基因表达(gene expression):是指在一定调节因素作用下,DNA分子上特定的基因被激活并转录生成特定的RNA或引起特异性蛋白质合成的过程。 通过基因表达合成特异性蛋白质,从而赋予细胞特定的生理功能或形态,以适应环境、维持发育和分化。 一般来说,处于转录激活状态的基因仅占5%。 基因表达的时间特异性:是指特定基因的表达严格按照特定的时间顺序发生,以适应细胞或个体特定分化、发育阶段的需要。故又称为阶段特异性。 基因表达的空间特异性:是指多细胞生物个体在某一特定生长发育阶段,同一基因的表达在不同的细胞或组织器官不同,从而导致特异性的蛋白质分布于不同的细胞或组织器官。又称为细胞特异性或组织特异性。

146
一、基因表达的方式 组成性基因表达:是指在个体发育的任一阶段都能在大多数细胞中持续进行的基因表达。其基因表达产物是对生命过程必需的,且较少受环境因素影响。这类基因被称为管家基因。 诱导表达(induction):是指在特定环境因素刺激下,基因被激活而使基因表达产物增加。这类基因称为可诱导基因。 阻遏表达(repression):是指在特定环境因素刺激下,基因被抑制而使基因表达产物减少。这类基因称为可阻遏基因。
:是指在特定环境因素刺激下,基因被抑制而使基因表达产物减少。这类基因称为可阻遏基因。")
147
二、基因表达调控的基本原理 1、基因表达的多级调控:
基因表达的调控可分为转录水平(基因激活及转录起始),转录后水平(加工及转运),翻译水平及翻译后水平。 2、基因转录激活调节基本要素: (1)特异DNA序列:具有调节功能的DNA序列。 顺式作用元件(cis-acting element)又称分子内作用元件,指存在于DNA分子上的一些与基因转录调控有关的特殊顺序。 原核生物中,其顺式作用元件主要由启动基因、操纵基因和调节基因组成。真核生物中,顺式作用元件主要有启动子(promoter)、增强子(enhancer)和沉默子(silencer)。
,转录后水平(加工及转运),翻译水平及翻译后水平。 2、基因转录激活调节基本要素: (1)特异DNA序列:具有调节功能的DNA序列。 顺式作用元件(cis-acting element)又称分子内作用元件,指存在于DNA分子上的一些与基因转录调控有关的特殊顺序。 原核生物中,其顺式作用元件主要由启动基因、操纵基因和调节基因组成。真核生物中,顺式作用元件主要有启动子(promoter)、增强子(enhancer)和沉默子(silencer)。")
148
(2)调节蛋白: 反式作用因子(trans-acting factor):又称为分子间作用因子,指一些与基因表达调控有关的蛋白质因子。 原核生物中的反式作用因子主要分为特异因子、激活蛋白和阻遏蛋白;而真核生物中的反式作用因子通常称为转录因子。 (3)DNA-蛋白质的相互作用: 反式作用因子与顺式作用元件之间的共同作用,才能够达到对特定基因进行调控的目的,其结合是非共价结合。
DNA-蛋白质的相互作用: 反式作用因子与顺式作用元件之间的共同作用,才能够达到对特定基因进行调控的目的,其结合是非共价结合。")
149
三、原核基因转录调节 1961年,Francis Jacob和Jacques Monod提出著名的操纵子学说,开创了基因表达调控的新领域。
存在于原核生物中的一种主要的调控模式是操纵子(operon)调控模式,该模式也见于低等真核生物中。 操纵子:是原核生物的调控系统,是控制若干相关结构基因表达的DNA功能区。
调控模式,该模式也见于低等真核生物中。 操纵子:是原核生物的调控系统,是控制若干相关结构基因表达的DNA功能区。")
150
1、操纵子的结构与功能 典型的操纵子可分为控制区和信息区两部分。控制区由各种调控基因所组成,而信息区则由若干结构基因串联在一起构成。(p562图39-21 酶的诱导与酶的阻遏) 调节基因(I):为阻抑蛋白编码基因 控制区 启动基因(P):为cAMP受体蛋白、RNA聚合酶等结合区 操纵基因(O):为阻抑蛋白结合位点,不编码蛋白质 信息区:由一个或数个结构基因串联在一起组成
:为cAMP受体蛋白、RNA聚合酶等结合区. 操纵基因(O):为阻抑蛋白结合位点,不编码蛋白质. 信息区:由一个或数个结构基因串联在一起组成.")
151
2、乳糖操纵子的调控机制 控制区 信息区 乳糖操纵子(Lac operon)控制区包括调节基因(阻遏基因),启动基因(其CRP结合位点位于RNA聚合酶结合位点上游)和操纵基因(lacO);信息区由β-半乳糖苷酶基因(lacZ),β-半乳糖苷通透酶基因(lacY)和乙酰基转移酶基因(lacA)串联在一起构成。(p521图15-2)
控制区包括调节基因(阻遏基因),启动基因(其CRP结合位点位于RNA聚合酶结合位点上游)和操纵基因(lacO);信息区由β-半乳糖苷酶基因(lacZ),β-半乳糖苷通透酶基因(lacY)和乙酰基转移酶基因(lacA)串联在一起构成。(p521图15-2)")
152
乳糖操纵子结构基因及其表达产物

153
当培养基中乳糖浓度升高而葡萄糖浓度降低时
(1)乳糖操纵子的诱导: 当培养基中乳糖浓度升高而葡萄糖浓度降低时 细胞中cAMP浓度升高 乳糖作为诱导剂与阻抑蛋白结合 cAMP与CRP结合并使之激活 促使阻抑蛋白与操纵基因分离 CRP与启动基因结合并促使 RNA聚合酶与启动基因结合 基因转录激活
乳糖操纵子的诱导: 当培养基中乳糖浓度升高而葡萄糖浓度降低时. 细胞中cAMP浓度升高 乳糖作为诱导剂与阻抑蛋白结合. cAMP与CRP结合并使之激活. 促使阻抑蛋白与操纵基因分离. CRP与启动基因结合并促使. RNA聚合酶与启动基因结合. 基因转录激活.")
155
(2)乳糖操纵子的阻遏: 当培养基中乳糖浓度降低而葡萄糖浓度升高时 细胞中cAMP浓度降低 缺乏乳糖与阻抑蛋白结合 CRP失活
CRP失活 阻抑蛋白与操纵基因结合 CRP及RNA聚合酶不能与 启动基因结合 基因转录被阻遏

157
3、色氨酸操纵子的调控机制 细菌E.coli具备合成色氨酸所需要的酶。这些酶编码基因串成一个操纵子。
色氨酸操纵子(trp operon)属于阻遏型操纵子,主要参与调控一系列用于色氨酸合成代谢的酶蛋白的转录合成。当细胞内缺乏色氨酸时,此操纵子开放,而当细胞内合成的色氨酸过多时,此操纵子被关闭。 色氨酸操纵子中含有五个结构基因,即trpE、trpD、trpC、trpB、trpA,编码三个合成色氨酸的酶。另外还存在一个前导肽编码基因(trpL)。(p523图15-5)
属于阻遏型操纵子,主要参与调控一系列用于色氨酸合成代谢的酶蛋白的转录合成。当细胞内缺乏色氨酸时,此操纵子开放,而当细胞内合成的色氨酸过多时,此操纵子被关闭。 色氨酸操纵子中含有五个结构基因,即trpE、trpD、trpC、trpB、trpA,编码三个合成色氨酸的酶。另外还存在一个前导肽编码基因(trpL)。(p523图15-5)")
159
色氨酸操纵子通常情况下处于开放状态,其辅阻遏蛋白不能与操纵基因结合而阻遏转录。而当色氨酸合成过多时,色氨酸作为辅阻遏物与辅阻遏蛋白结合而形成阻遏蛋白,后者与操纵基因结合而使基因转录关闭。
色氨酸操纵子的调控还涉及转录衰减(attenuation)机制。因为在色氨酸操纵子序列中有2个色氨酸密码子,且第一个结构基因与启动基因之间存在有一衰减区域,同时可形成特殊的发夹结构。
机制。因为在色氨酸操纵子序列中有2个色氨酸密码子,且第一个结构基因与启动基因之间存在有一衰减区域,同时可形成特殊的发夹结构。")
161
当细胞内色氨酸浓度很高时,通过与转录相偶联的翻译过程,形成一个衰减子结构,使RNA聚合酶从DNA上脱落,导致转录终止。
当色氨酸缺乏时,核蛋白体翻译停止在密码子前,阻止衰减子的形成,转录继续进行。

163
本 章 小 结 基因表达的内容,即中心法则。 DNA的合成(复制):复制的定义、体系(模板、酶、原料、辅助因子)、方向、方式、特点、过程(前导链、随后链合成的区别,起始、延长。终止);逆转录及意义;DNA的损伤与修复。 RNA的合成(转录):转录的定义、体系(模板、酶、原料、辅助因子)、方向、特点、过程(起始、延长、终止)、加工修饰。 蛋白质的合成(翻译):翻译的定义、体系(模板、酶、原料、辅助因子)、场所、方向、特点(各种RNA所起的作用与结构特点)、过程(起始、延长、终止)、加工修饰。 基因表达的调控及意义。转录水平的调控、操纵子模式(概念、结构、调控方式)。 名词解释:中心法则、半保留复制、冈崎片段、端粒、端粒酶、拓扑异构酶、逆转录、启动子、不对称转录、聚合酶链式反应、核酶、转录、翻译、核蛋白体循环、密码子、反密码子、连续性、简并性、摆动配对、操纵子、乳糖操纵子
:转录的定义、体系(模板、酶、原料、辅助因子)、方向、特点、过程(起始、延长、终止)、加工修饰。 蛋白质的合成(翻译):翻译的定义、体系(模板、酶、原料、辅助因子)、场所、方向、特点(各种RNA所起的作用与结构特点)、过程(起始、延长、终止)、加工修饰。 基因表达的调控及意义。转录水平的调控、操纵子模式(概念、结构、调控方式)。 名词解释:中心法则、半保留复制、冈崎片段、端粒、端粒酶、拓扑异构酶、逆转录、启动子、不对称转录、聚合酶链式反应、核酶、转录、翻译、核蛋白体循环、密码子、反密码子、连续性、简并性、摆动配对、操纵子、乳糖操纵子.")
164
思 考 题 1、试述生物机体是如何保证DNA复制的准确性。 2、以乳糖操纵子为例说明原核生物是如何调控基因表达的。
3、遗传密码是如何编码的?有何特点。 4、以各种氨基酸、tRNA等为基本原料,用mRNA作为模板,生物合成含有300个氨基酸残基的多肽,至少需要消耗多少摩尔高能磷酸键? 5、简述RNA的加工修饰。

165
结 束

Similar presentations















>")

蛋白质的降解: 外源蛋白的消化 内源性蛋白的选择性降解 (2)氨基酸的分解代谢:>")


>")


 Chapter 12 Protein Biosynthesis,Translation>")



 一、DNA的复制方式~半保留复制>")



