Download presentation
Presentation is loading. Please wait.

1
第十二章 遗传与个体发育 第一节 细胞质在遗传中的作用 第二节 细胞分化的可逆性 第三节 基因表达的调控

2
第一节 细胞质在遗传中的作用 一、细胞质的不均一性和细胞分化 二、细胞质对染色体行为的影响 三、细胞质对性染色题的影响:X染色体失活

3
一、细胞质的不均一性和细胞分化 海胆受精卵发育 (1)海胆受精卵第一、第二次分裂,顺着对称轴方向进行。
(2)如果将四个卵裂细胞分开,每个卵裂细胞都能发育成小幼虫,各个卵裂细胞中细胞质相同。 (3)第三次卵裂方向与对称轴垂直,分裂的8个卵裂细胞分开后,就不能发育成幼虫。
如果将四个卵裂细胞分开,每个卵裂细胞都能发育成小幼虫,各个卵裂细胞中细胞质相同。 (3)第三次卵裂方向与对称轴垂直,分裂的8个卵裂细胞分开后,就不能发育成幼虫。")
4
一、细胞质的不均一性和细胞分化 1、海胆切割试验:
海胆受精卵最初两次分裂沿着对称轴进行,形成的4个细胞所含的物质基本上一致。沿对称轴分开,每个细胞可正常发育并长成小幼虫。说明各个卵裂细胞中细胞质是完全相同的。

5
一、细胞质的不均一性和细胞分化 1、海胆切割试验: 在卵裂开始时,顺着赤道面把卵切成两半,一半含有动物极,一半含有植物极。
上半部分发育成空心的球形胚,表面长着许多纤毛,但不能再往前发育,许多天后死亡; 下半部分发育成一个比较复杂但不完整的胚胎,它没有纤毛,能分化出内胚层,不久也死亡。

6
一、细胞质的不均一性和细胞分化 2、角贝的早期发育:角贝的细胞质可以分为三层:
动物极和植物极各有一层清澈的细胞质,在这两层之间是一层较宽的颗粒性细胞质,它组成卵子的主要部分。在卵裂开始的时候,植物极清澈的细胞质层形成圆球形的突出部分,称为极叶。

7
在卵裂开始的时候,植物极的细胞质层形成极叶和一个裂球相连,在第 1次卵裂结束的时候它又缩回,和这个裂球融合为一。因此这个裂球(CD裂球)包含所有3层细胞质,而另外一个裂球(AB裂球)只包含动物极清澈的细胞质层和颗粒层。第2次卵裂开始的时候,CD裂球再次形成极叶,在4细胞时期,极叶仍只和一个裂球(D裂球)相连。这样,4个裂球在细胞质的组成上是不均等的,只有D裂球包含了所有极叶的物质。
包含所有3层细胞质,而另外一个裂球(AB裂球)只包含动物极清澈的细胞质层和颗粒层。第2次卵裂开始的时候,CD裂球再次形成极叶,在4细胞时期,极叶仍只和一个裂球(D裂球)相连。这样,4个裂球在细胞质的组成上是不均等的,只有D裂球包含了所有极叶的物质。")
8
一、细胞质的不均一性和细胞分化 2、角贝的早期发育: 如果在4细胞时期,将裂球分离培养:
包含极叶的裂球可以发育为一个完整的、较小的担轮幼虫; 不含极叶的 3个则只能发育为缺少中胚层结构的、有缺陷的幼虫。 如果在第1或者第2次卵裂期将极叶切除, 虽然卵子包含动物极的和颗粒性的细胞质,而且也包含应有的细胞核,但因为缺少极叶的物质,也只能发育为一个缺少中胚层结构的、有缺陷的幼虫。 说明,极叶中包含的是细胞质而不是细胞核,决定裂球能独特地分化出中胚层结构。

9
一、细胞质的不均一性和细胞分化 2、角贝的早期发育:
使用角贝的近缘种,发现除去极叶后产生的畸形胚,与正常胚相比较,发现RNA合成的速率不同,进一步研究发现,是由于mRNA合成的降低。因此,极叶细胞质能调节核基因的活性,以某种形式影响基因的作用,从而诱导了细胞的分化。

10
二、细胞质对染色体行为的影响 小麦瘿蚊 受精卵卵裂时遗弃染色体,并非所有细胞都丢失,有选择性的丢失,哪些细胞丢失由细胞质决定。

11
小麦瘿蚊 二、细胞质对染色体行为的影响 受精卵卵裂最初时核分裂、胞质不分裂形成多核卵。核分裂时在细胞的一端出现极细胞质,当核分裂3~4次时,有两个核进入极细胞质中,然后卵裂,极细胞质中的核染色体不丢失(40条)之后发育成生殖细胞,其它核丢失32条,剩8条,之后仅能发育成体细胞。 结扎使核不能进入极细胞质区域,则不能形成生殖细胞,瘿蚊不育。

12
三、细胞质对性染色体的影响: X染色体失活 1、人的巴氏小体 2、三色猫(玳瑁猫):毛皮上有三种颜色,黄色、黑色和白色。
黄色: XO 黑色:Xo 在不是黄色和黑色的地方就是白色。 白色:由常染色体上的不完全显性基因S和s决定的。SS纯合体白色分布全身,Ss白色限于腹部和四肢。三色猫的基因型应为:XOXoS , X随机失活,细胞会产生黄色或黑色花斑。 和猫的颜色相关的基因机制比较复杂,涉及到很多条基因。 其中决定黑色的基因是B,控制合成一种酪氨酸酶,在代谢中能产生黑色素。而这个基因相对的隐性基因是b,能产生褐色或巧克力色。 决定白色的基因是W。和决定黑色的基因不同,W基因起到的是“捣乱”的作用,使得其他合成色素的细胞无法展现出自己原本的颜色。W是一个显性的基因,无论是WW还是Ww都会使猫变成白色而无视原本猫带有的成色基因。 好,回答你的问题吧。一只黑猫的基因型比较容易确定,即wwBB或wwBb。而白猫的基因型比较复杂,有WWBB,WWBb,WWbb,WwBB,WwBb或Wwbb 后代的可能性也会比较多变,比如说 wwBb(黑猫)和WwBb(白猫)的后代可能是WwBb(白猫),也有可能是wwBb(黑猫),甚至可以是wwbb(褐色猫) 结论1:白猫和黑猫的后代可以是全白,可以是全黑,也可以是褐色 杂花的毛色由另一个基因控制,即S基因。S基因的表达可以在猫的身上产生白色的斑纹,纯合的SS猫比杂合的Ss猫的斑纹来得更多。 如果上面这个例子中白猫的基因型是WwBbSS,而黑猫的基因型是wwBbss,那么它的后代可能是WwBbSs(白猫),也有可能是wwBbSs(黑白相间的猫),或者可以是wwbbSs(褐白相间的猫) 结论2:黑猫和白猫也可以生出黑白花的猫。 灰色猫的基因则需要涉及到一个“淡化”的概念,淡化是由D基因控制的,D基因的产物可以使色素在毛发里正常运输和沉积。当猫没有D基因时(即dd),那么它的毛发颜色就会变淡,比如黑色会变成灰色,橙色会变成淡黄色。 如果第一个例子中白猫的基因型是WwBbdd,而黑猫的基因型是wwBbDd,那么后代可能是WwBbdd(白猫),也有可能是wwBbdd(灰色的猫),或者可以是wwbbdd(淡褐色的猫)。当然,最初的黑猫(wwBbDd)和褐色猫(wwbbDd)也还是可能存在的 结论3:白猫和黑猫能够生出灰色的猫
和WwBb(白猫)的后代可能是WwBb(白猫),也有可能是wwBb(黑猫),甚至可以是wwbb(褐色猫) 结论1:白猫和黑猫的后代可以是全白,可以是全黑,也可以是褐色 杂花的毛色由另一个基因控制,即S基因。S基因的表达可以在猫的身上产生白色的斑纹,纯合的SS猫比杂合的Ss猫的斑纹来得更多。 如果上面这个例子中白猫的基因型是WwBbSS,而黑猫的基因型是wwBbss,那么它的后代可能是WwBbSs(白猫),也有可能是wwBbSs(黑白相间的猫),或者可以是wwbbSs(褐白相间的猫) 结论2:黑猫和白猫也可以生出黑白花的猫。 灰色猫的基因则需要涉及到一个 淡化 的概念,淡化是由D基因控制的,D基因的产物可以使色素在毛发里正常运输和沉积。当猫没有D基因时(即dd),那么它的毛发颜色就会变淡,比如黑色会变成灰色,橙色会变成淡黄色。 如果第一个例子中白猫的基因型是WwBbdd,而黑猫的基因型是wwBbDd,那么后代可能是WwBbdd(白猫),也有可能是wwBbdd(灰色的猫),或者可以是wwbbdd(淡褐色的猫)。当然,最初的黑猫(wwBbDd)和褐色猫(wwbbDd)也还是可能存在的 结论3:白猫和黑猫能够生出灰色的猫.")
14
第二节 细胞分化的可逆性 个体发育:细胞分裂分化的过程。 发育:高等生物从受精卵开始发育,经过一系列细胞分裂、分化,发育成新个体。

15
第二节 细胞分化的可逆性 ※一个受精卵是有全能性的,它可以产生各种类型的细胞。在个体发育的什么阶段,细胞的全能性受到了限制呢?一直认为植物细胞具有全能性(人为条件下完成的)动物细胞不具有全能性。 ※直到1997年2月,苏格兰学者Wilmut用体细胞核移植技术培养成“多莉”绵羊,这一动物细胞无全能性的观点才发生动摇。

16
第二节 细胞分化的可逆性 ※动物再生的例子:将海参横向环切为数段,经过3-5个月,每段又会生成一个完整的海参;海参排脏逃生,经30-50天再生新内脏;壁虎再生失去的尾巴,涡虫、蚯蚓可以再生失去的部分都说明动物细胞是具有全能性的,只不过是能够再生的组织和器官在数目和种类上是有限的,特别是高等动物。

17
一、植物的组织培养 植物的组织培养:在体外人工培养植物组织和细胞的技术。
※例:胡萝卜韧皮部的单个细胞重新发育成一个完整的个体。现在小麦玉米、水稻等很多植物都可以通过组织培养获得愈伤组织,产生新的个体。因此,分化的细胞,可以从高度的分化状态回复过来(脱分化),重新再分化,形成植株,即植物细胞具有全能性。 ※ 实践上的意义:有利于突变植株的获得,可缩短育种进程(诱变培养细胞,筛选需要的突变型,分化为成熟植株);快速无性繁殖(试管苗);单体培养快速获得纯合个体。
,重新再分化,形成植株,即植物细胞具有全能性。 ※ 实践上的意义:有利于突变植株的获得,可缩短育种进程(诱变培养细胞,筛选需要的突变型,分化为成熟植株);快速无性繁殖(试管苗);单体培养快速获得纯合个体。")
18
植物的组织培养 外植体 诱导产生愈伤组织 诱芽培养基 生根培养基 移植入土

19
马兰的组织培养

20
二、动物的核移植试验 核移植是将供体细胞核移入去核的卵母细胞中,使后者不经精子穿透等有性过程即可被激活、分裂并发育,让核供体的基因得到完全复制。培养一段时间后,在把发育中的卵母细胞移植到人或动物体内的方法。 核移植的细胞来源主要分为:供体细胞来源和受体细胞的来源两种。 核移植主要用于细胞移植和异种器官移植,细胞移植可以治疗由于细胞功能缺陷所引起的各种疾病。 汉斯·施佩曼,1869年生于德国。高中毕业后参军。退役后在家族产业中帮忙。后改变志向学医。毕业后在维尔茨堡大学动物系工作。历任罗斯托克大学、弗赖堡大学教授等职,因发现蝾螈胚胎体中的“组织中心”而获1935年诺贝尔生理学医学奖。1941年因希特勒迫害逝世。

21
二、动物的核移植试验 (一)移植开端 1938年德国汉斯.施佩曼提出他称之为“奇异的实验”的设想:从发育到后期的胚胎(成熟的或未成熟的胚胎均可)中取出细胞核,将其移植到一个卵子中。他被称为“克隆之父” 施佩曼毕生从事两栖类胚胎早期发育的研究,1902年,当蝾螈受精卵分裂为二细胞时,他用胎发在两个分裂球之间加以结扎,结果每个分裂球都发育成一个完整的胚胎,而如果在原肠期结扎,则两半各自发育为半个胚胎。这意味着胚胎在早期和晚期之间发生了某种变化,使胚胎各部分“决定”确定下来。1920年,又通过异位移植实验,把受精卵结扎为背腹两半,只有背部一半能产生正常胚胎,而把背唇移植到腹唇,可产生第二个胚胎。他把背唇区称为“组织者”。 汉斯·施佩曼,1869年生于德国。高中毕业后参军。退役后在家族产业中帮忙。后改变志向学医。毕业后在维尔茨堡大学动物系工作。历任罗斯托克大学、弗赖堡大学教授等职,因发现蝾螈胚胎体中的“组织中心”而获1935年诺贝尔生理学医学奖。1941年因希特勒迫害逝世。

22
二、动物的核移植试验 (一)移植开端 1952年,研究细胞全能性的第一个人美国科学家R.Briggs(罗伯特.布里格斯)豹纹蛙→ 囊胚期细胞核→去核的同种蛙卵→部分卵发育成个体(即克隆)。 胚胎后期到蝌蚪、成蛙的细胞核→移入去核的同种蛙卵中→失败。 认为胚胎早期细胞核具有全能性,而胚胎以后各个时期的细胞核难以体现全能性,即分化难以逆转,多数生物学家转向以未成熟的胚胎细胞克隆动物的领域。

23
二、动物的核移植试验 (二)胚胎细胞核移植—胚胎细胞克隆 胚胎早期(囊胚期)的细胞核移入去核的异种动物的受精卵中,使受精卵发育成个体的过程。
我国童第周和美籍华人牛满江教授在鱼类和两栖类中进行核移植,成功地得到了核质杂交品种。鲤鱼生长迅速、个体大、肉质粗厚、味道差。鲫鱼肉细味美,但个体小。(若二者杂交,子一代雄性不育,无法繁殖。) 童第周( ),字慰孙,浙江鄞县人。我国著名生物学家、教育家、中国试验胚胎学的创始人,是试验胚胎学、细胞生物学、发育生物学及海洋生物学等领域卓有建树的生物学家,被誉为“克隆先驱”。 1930年童第周在亲友们的资助下,远度重洋,来到北欧比利时的首都——布鲁塞尔比京大学留学。在欧洲著名生物学者勃朗歇尔教授的指导下,研究胚胎学。当时,他发现有的外国留学生对中国人抱着一种藐视的态度,说“中国人是弱国的国民”。和他同住的一个洋人学生公开说:“中国人太笨。”听到这些,童第周再也压抑不住满腔的怒火,对那个洋人说:“这样吧,我们来比一比,你代表你的国家,我代表我的国家,看谁先取得博士学位。” 童第周憋着一股气,在日记中写下了自己的誓言:“中国人不是笨人,应该拿出东西来,为我们的民族争光!” 研究胚胎学,经常要做卵细胞膜的剥除手术,有一次做实验,教授要求学生们设法把青蛙卵膜剥下来,这是一项难度很大的手术,青蛙卵只有小米粒大小,外面紧紧地包着三层象蛋白一样的软膜,因为卵小膜薄,手术只能在显微镜下进行。许多人都失败了,他们一剥开卵膜,就把青蛙卵也给撕破了。只有童第周一人不声不响地完成了这项实验任务。
 童第周( ),字慰孙,浙江鄞县人。我国著名生物学家、教育家、中国试验胚胎学的创始人,是试验胚胎学、细胞生物学、发育生物学及海洋生物学等领域卓有建树的生物学家,被誉为 克隆先驱 。 1930年童第周在亲友们的资助下,远度重洋,来到北欧比利时的首都——布鲁塞尔比京大学留学。在欧洲著名生物学者勃朗歇尔教授的指导下,研究胚胎学。当时,他发现有的外国留学生对中国人抱着一种藐视的态度,说 中国人是弱国的国民 。和他同住的一个洋人学生公开说: 中国人太笨。 听到这些,童第周再也压抑不住满腔的怒火,对那个洋人说: 这样吧,我们来比一比,你代表你的国家,我代表我的国家,看谁先取得博士学位。 童第周憋着一股气,在日记中写下了自己的誓言: 中国人不是笨人,应该拿出东西来,为我们的民族争光! 研究胚胎学,经常要做卵细胞膜的剥除手术,有一次做实验,教授要求学生们设法把青蛙卵膜剥下来,这是一项难度很大的手术,青蛙卵只有小米粒大小,外面紧紧地包着三层象蛋白一样的软膜,因为卵小膜薄,手术只能在显微镜下进行。许多人都失败了,他们一剥开卵膜,就把青蛙卵也给撕破了。只有童第周一人不声不响地完成了这项实验任务。")
24
二、动物的核移植试验 (二)胚胎细胞核移植—胚胎细胞克隆 具有生殖能力 肌肉蛋白含量比鲤鱼高3.78% 脂肪含量比鲤鱼高5.58%
胚胎细胞核移植—胚胎细胞克隆 具有生殖能力 肌肉蛋白含量比鲤鱼高3.78% 脂肪含量比鲤鱼高5.58%")
25
二、动物的核移植试验

26
(三)体细胞核移植—体细胞克隆 动物体细胞克隆技术又称体细胞核移植技术或无性繁殖技术,它是用动物特定发育阶段的核供体(体细胞核)及相应的核受体(去核的原核胚或成熟的卵母细胞、卵细胞)不经过有性繁殖过程,进行体外重构,并通过重构胚的胚胎移植,从而达到扩繁同基因型动物种群的目的。
体细胞核移植—体细胞克隆 动物体细胞克隆技术又称体细胞核移植技术或无性繁殖技术,它是用动物特定发育阶段的核供体(体细胞核)及相应的核受体(去核的原核胚或成熟的卵母细胞、卵细胞)不经过有性繁殖过程,进行体外重构,并通过重构胚的胚胎移植,从而达到扩繁同基因型动物种群的目的。")
27
(三)体细胞核移植—体细胞克隆 1、体细胞克隆的类型
(1)同种体细胞核移植:科学家用取自一只6岁成羊的乳腺细胞培育成功一只克隆羊,开创了成年哺乳动物克隆的先河。 (2)异种体细胞核移植:陈大元等将大熊猫的子宫上皮细胞、骨骼肌细胞和乳腺上皮细胞的细胞核移入去核的兔卵母细胞后,分别有9.9 %、6.8 % 和11.7 % 的重构卵发育到囊胚,染色体、线粒体DNA和核DNA分析均证实异种重构胚的核遗传物质来自大熊猫供体细胞。这不仅开创了大熊猫体细胞移核的先例,而且首次大熊猫体细胞核的异种移核。对大熊猫的快繁及保护提供了一线希望。
同种体细胞核移植:科学家用取自一只6岁成羊的乳腺细胞培育成功一只克隆羊,开创了成年哺乳动物克隆的先河。 (2)异种体细胞核移植:陈大元等将大熊猫的子宫上皮细胞、骨骼肌细胞和乳腺上皮细胞的细胞核移入去核的兔卵母细胞后,分别有9.9 %、6.8 % 和11.7 % 的重构卵发育到囊胚,染色体、线粒体DNA和核DNA分析均证实异种重构胚的核遗传物质来自大熊猫供体细胞。这不仅开创了大熊猫体细胞移核的先例,而且首次大熊猫体细胞核的异种移核。对大熊猫的快繁及保护提供了一线希望。")
28
(三)体细胞核移植—体细胞克隆 2、体细胞克隆的一般方法
胎儿细胞或成年动物已分化的细胞作为核供体,进行细胞核移植,得到与供体细胞具有同性质动物的过程。 克隆羊“多莉”的克隆方法如下:

29
体细胞克隆 A 多莉羊

30
克隆动物

31
克隆动物

32
(四)关于克隆人 1997年,克隆(即无性繁殖)羊“多莉”的问世,让人们听到了克隆人的脚步声。由于克隆人不单纯是科技问题,更是对人类社会及其未来命运有着非同寻常影响,所以国外各界人士纷纷从科学、哲学(包括科技哲学、价值哲学、伦理学、宗教哲学)、社会学、法学、心理学等不同角度,对克隆人问题展开了广泛的探讨。 克隆人“三疯子”的科学家是:美国“克隆基金”主任布瓦瑟利耶、肯塔基州列克星顿大学激素学院院长扎沃斯和意大利科学家安蒂诺里。由于此三人是克隆人的坚定推崇者,因此被学术界称为“科学疯子”。
羊 多莉 的问世,让人们听到了克隆人的脚步声。由于克隆人不单纯是科技问题,更是对人类社会及其未来命运有着非同寻常影响,所以国外各界人士纷纷从科学、哲学(包括科技哲学、价值哲学、伦理学、宗教哲学)、社会学、法学、心理学等不同角度,对克隆人问题展开了广泛的探讨。 克隆人 三疯子 的科学家是:美国 克隆基金 主任布瓦瑟利耶、肯塔基州列克星顿大学激素学院院长扎沃斯和意大利科学家安蒂诺里。由于此三人是克隆人的坚定推崇者,因此被学术界称为 科学疯子 。")
33
克隆三疯子 布丽吉特·布瓦瑟利耶 意大利科学家安蒂诺里 扎沃斯

34
(四)关于克隆人 1、克隆人就在我们身边 (1)体细胞克隆人的过程—人造人 (2)同卵双胞胎的产生过程—天然的克隆人
同卵双胞胎—由同一个受精卵一分二,而二个细胞各自发育形成二个独立个体,二个人不但细胞核、细胞质完全相同,而在完全相同的子宫环境发育成长,比由克隆技术产生的克隆人有更完美的吻合度。这个完美的天然的克隆人是一种更为完美的克隆人,真正意义上的克隆人—同卵双胞胎

35
(四)关于克隆人
关于克隆人")
36
天然的克隆人

37
(四)关于克隆人 2、治疗性克隆 治疗性克隆:体细胞核→卵细胞→新的细胞→“桑椹” →胚胎干细胞→克隆人体器官,供医学研究,解决器官移植供体不足问题等。 例如:治疗性克隆希望使用病人自己细胞中的遗传物质来产生胰岛以治疗糖尿病,或者产生神经细胞来修复受损的脊髓。

38
(四)关于克隆人 3、生殖性克隆 生殖性克隆:即通常所说的克隆人。体细胞核→卵细胞→新的细
→“桑椹”(胚胎干细胞) →胚胎植入到一个女性子宫→婴儿 如果干细胞保留时间超过14天,它就可能长出神经元,此时,这一 细胞就有了生命意义。 据报道,2001年11月美国科学家在马萨诸塞州的先进细胞技术公司,用人的皮肤细胞克隆出首个人类早期胚胎,12天后把它焚化烧毁。 目前国际上已对治疗性克隆规定了三条原则。一是取得的材料 卵子、体细胞,必须是自愿的,不能是骗来的,不能是买来的,提供 者有知情权。二是胚胎细胞保留时间不能超过14天,超过了,就被视 为动机不纯,有克隆人之嫌。三是不能将克隆的胚胎细胞植入人体子宫。
 →胚胎植入到一个女性子宫→婴儿. 如果干细胞保留时间超过14天,它就可能长出神经元,此时,这一. 细胞就有了生命意义。 据报道,2001年11月美国科学家在马萨诸塞州的先进细胞技术公司,用人的皮肤细胞克隆出首个人类早期胚胎,12天后把它焚化烧毁。 目前国际上已对治疗性克隆规定了三条原则。一是取得的材料. 卵子、体细胞,必须是自愿的,不能是骗来的,不能是买来的,提供. 者有知情权。二是胚胎细胞保留时间不能超过14天,超过了,就被视. 为动机不纯,有克隆人之嫌。三是不能将克隆的胚胎细胞植入人体子宫。")
39
我国首次人类胚胎体外培养 辽宁省计划生育研究院王彤博士等人在我国首次完成人类全胚胎体外培养取得成功。共培养成活28例人类胚胎,存活最长至今22天。左图是培养液中的约1cm长的人胚胎。受精后20-50天胚胎长度2mm-1cm。 2004年4月19日钱江晚报 患心脏病、红斑狼疮的母亲便可“生”下孩子。

40
(五)体细胞克隆目前存在的问题 体细胞克隆已在牛、绵羊、山羊、小鼠和猪等动物中取得了成功,但在其它动物上还有待进一步验证。其总体水平仍处于实验研究阶段,走向实践运用还面临许多问题,主要表现在以下几个方面: 1.费用高、效率低:通过体细胞克隆动物的费用非常高,如制作“多莉”的费用就超过了200万英镑。效率低,克隆绵羊只有3% 左右的重组胚能发育为正常后代。在克隆牛的研究中,重组胚的囊胚发育率最高能达49%,约有80% 的移植胚能发育为正常胎儿,但胎儿出生后的死亡率达50%。而Cibelli等在克隆牛的研究中,重组胚的囊胚率仅为17%,移植后只有14% 的囊胚发育为正常后代。克隆小鼠研究中,只有2%~3% 的重组胚能发育产仔,仔鼠死亡率也高达40%。以上结果说明,体细胞克隆技术仅处于初步研究阶段,还需要在其它动物上进一步充分研究,以提高生产效率。

41
(四)体细胞克隆目前存在的问题 2.克隆胎儿初生重增加,质量差:由于培养基中血清的存在,导致早期胚胎基因表达发生改变,致使许多克隆动物初生重增加,很难自然分娩,大多数需人工辅助。初生儿还伴有各种缺陷,死亡率增高。出现这一现象的原因还不清楚,目前只能从优化培养条件及操作程序入手以减少胎儿死亡率。 3.克隆动物生长发育不正常,非正常死亡率高:研究发现,有些用修饰过的细胞培育出的克隆动物存在缺陷,它们在生长过程中会出现难以预料的后果。“多莉”羊在生长过程中就出现了比同龄羊提前衰老的现象,而且现已死亡。亦有研究表明,克隆鼠成年后常患有肥胖症的原因与早期胚胎阶段的细胞缺陷有关。

42
(五)成功率低的原因 尽管也有少数成功率较高的报道,但目前公认的体细胞克隆成功率在1%~3%。克隆胚胎移植后的出生率平均不到10%。胎儿异常、流产和围产期死亡率高,出生后一周的死亡率最高可达100%。原因七方面: 1.不能高效率的去除受体卵母细胞的染色体:现有的各种去核技术都要损失部分卵母细胞的细胞质。 2.重组胚的融合和卵母细胞质的激活 3.供体核与受体胞质细胞周期同步的问题:供体核与受体胞质细胞周期同步与否是影响克隆胚胎发育的重要因素之一。目前通用的受体胞质是M 期的卵母细胞。关于血清饥饿G0/G1期细胞作为核供体的优越性曾存在争议。现在认为,成体细胞因对接触抑制敏感性高和细胞周期长,故无需饥饿也多处在G1期。而胎儿细胞则必须饥饿才能获得较高的克隆成功率。目前一般认为,除了S期以外,其它各时期细胞均可作为核供体。

43
(五)成功率低的原因 4.线粒体来源问题:无论直接注射还是细胞融合,供体核都带入一部分细胞质。于是,就出现了移核胚胎的线粒体来源问题。结论不一致:来源于受体;来源于供体与受体双方。 5.端粒的问题:端粒(telomere)是染色体端部特化部分,因无黏性而能防止染色体黏着。细胞每复制一次端粒核苷酸就减少50~100bp。端粒复制要靠端粒酶(telomerase)。正常细胞缺乏此酶,故端粒随细胞分裂而变短,细胞衰老。而生殖细胞和癌细胞则都有此酶,因此不衰老。那么,克隆动物是否会遗传供核细胞缩短的端粒而提前衰老呢?不一致:多莉短;克隆牛不短。
是染色体端部特化部分,因无黏性而能防止染色体黏着。细胞每复制一次端粒核苷酸就减少50~100bp。端粒复制要靠端粒酶(telomerase)。正常细胞缺乏此酶,故端粒随细胞分裂而变短,细胞衰老。而生殖细胞和癌细胞则都有此酶,因此不衰老。那么,克隆动物是否会遗传供核细胞缩短的端粒而提前衰老呢 不一致:多莉短;克隆牛不短。")
44
染 色 体

45
染 色 体

46
(五)成功率低的原因 6.X-染色体灭活问题:雌雄哺乳动物的基因剂量补偿是通过灭活雌性动物的一条X-染色体来实现的。正常情况下,着床前胚胎的两条X染色体都活动。后来,上胚层中X染色体随机灭活,而滋养层细胞则专门灭活父系X染色体。死亡克隆胎盘X染色体随机灭活,但活克隆为父亲X染色体灭活。
成功率低的原因 6.X-染色体灭活问题:雌雄哺乳动物的基因剂量补偿是通过灭活雌性动物的一条X-染色体来实现的。正常情况下,着床前胚胎的两条X染色体都活动。后来,上胚层中X染色体随机灭活,而滋养层细胞则专门灭活父系X染色体。死亡克隆胎盘X染色体随机灭活,但活克隆为父亲X染色体灭活。")
50
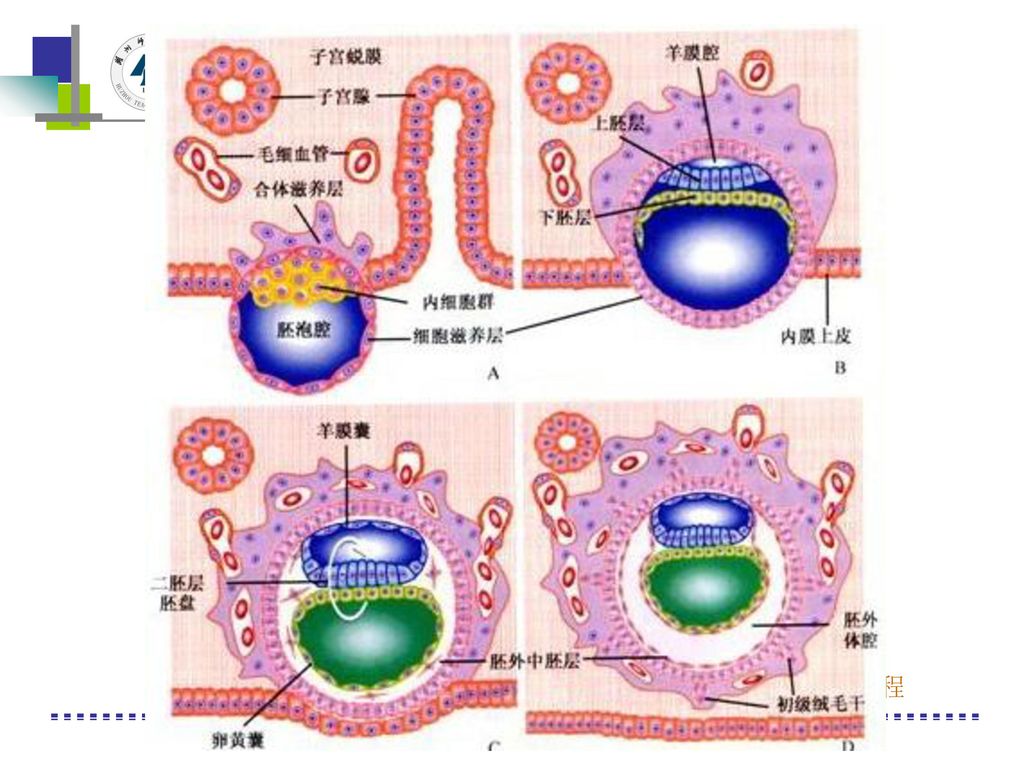
胎儿发育 卵子在受精后的2周内称孕卵或受精卵;受精后的第3~8周称为胚胎;第8周末起称为胎儿;妊娠各周胎儿发育的特征如下: 8周 胚胎长约3cm,初具人形。 头特别大、眼、耳、鼻、口已可辨认,早期心脏形成,有搏动,超声检查可以发现。 12周 胎儿身长7~9cm。 重约20g。 外生殖器已发生。 四肢有微弱活动,大多数骨骼中已出现骨化中心。 16周 胎儿身长约10~17cm。 重100~120g,皮肤色红,光滑透明。 有少量毳毛。骨骼入一步发育。 X线检查可见骨骼阴影。 外生殖器可辨男女。腹部检查可听到胎心音。 孕妇可感到胎动。 20周 胎儿身长18~27cm,重280~300g,皮肤暗红。 透明度减低,全身有胎脂孕33周胎儿发育情况,胎头占全身的1/3。 有头发生长。 开始出现吞咽活动。 (网址: .Com) 24周 胎儿身长28~34cm,重600~700g,皮下脂肪开始沉积,皮肤有皱纹。 28周胎儿身长35~38cm。 重100~1200g。全身细瘦,皮肤发红。 上有胎脂,指(趾)甲未达到指(趾)端。女性阴唇已发育。 大阴唇包藏小阴 唇及阴蒂,男性睾丸已降至阴囊。因皮下脂肪少,面部皱纹多。 形如老人。若出生。 则能啼哭。 会吞咽。 四肢能活动,但生活力弱。 需特殊护理方能生存。 32周 胎儿身长40cm,重1500~1700g,皮肤深红,面部胎毛已脱落,生后适当护理可存活。 36周 胎儿身长45~46cm。 约重2500g。皮下脂肪多,面部皱纹消失,指(趾)甲已达指(趾端)。出生后啼器与吸吮,成活机会很大。 40周 胎儿发育成熟,身长约50cm。 重约3000~3300g。皮肤粉红,皮下脂肪发育良好,胎儿大部分已消退,头发长2~3cm。指(趾)甲已过指(趾)端。四肢运动活泼,能大声啼器,有强烈吸吮反射。 胎儿身长、体重随妊娠月份逐渐增加。 为便于记忆,一般采用下列公式计算: 妊娠20周前 身怀孕前三个月的症状长=妊娠月数的平方(cm) 体重=妊娠月数的立方×2(g) 妊娠20周后 身长=妊娠月数×5(cm) 体重=妊娠月数的立方×3(g)
 24周 胎儿身长28~34cm,重600~700g,皮下脂肪开始沉积,皮肤有皱纹。 28周胎儿身长35~38cm。 重100~1200g。全身细瘦,皮肤发红。 上有胎脂,指(趾)甲未达到指(趾)端。女性阴唇已发育。 大阴唇包藏小阴 唇及阴蒂,男性睾丸已降至阴囊。因皮下脂肪少,面部皱纹多。 形如老人。若出生。 则能啼哭。 会吞咽。 四肢能活动,但生活力弱。 需特殊护理方能生存。 32周 胎儿身长40cm,重1500~1700g,皮肤深红,面部胎毛已脱落,生后适当护理可存活。 36周 胎儿身长45~46cm。 约重2500g。皮下脂肪多,面部皱纹消失,指(趾)甲已达指(趾端)。出生后啼器与吸吮,成活机会很大。 40周 胎儿发育成熟,身长约50cm。 重约3000~3300g。皮肤粉红,皮下脂肪发育良好,胎儿大部分已消退,头发长2~3cm。指(趾)甲已过指(趾)端。四肢运动活泼,能大声啼器,有强烈吸吮反射。 胎儿身长、体重随妊娠月份逐渐增加。 为便于记忆,一般采用下列公式计算: 妊娠20周前 身怀孕前三个月的症状长=妊娠月数的平方(cm) 体重=妊娠月数的立方×2(g) 妊娠20周后 身长=妊娠月数×5(cm) 体重=妊娠月数的立方×3(g)")
53
(五)成功率低的原因 7.克隆胚胎某些基因的再程序化异常:已经证明,受精后父系和母系基因组都发生广泛的去甲基化。有人证明,牛着床前移核胚胎中有些重复序列的甲基化异常,某些胚胎基因往往不能再激活。这说明,大多数克隆胚胎的后成性再程序化异常,不彻底的再程序化可能是造成克隆成功率低的原因。
成功率低的原因 7.克隆胚胎某些基因的再程序化异常:已经证明,受精后父系和母系基因组都发生广泛的去甲基化。有人证明,牛着床前移核胚胎中有些重复序列的甲基化异常,某些胚胎基因往往不能再激活。这说明,大多数克隆胚胎的后成性再程序化异常,不彻底的再程序化可能是造成克隆成功率低的原因。")
54
第三节 基因表达的调控 一、基因表达调控的例征 (一)不同条件下的基因激活
第三节 基因表达的调控 一、基因表达调控的例征 (一)不同条件下的基因激活 1、细菌和其他单细胞生物中,不同条件下可形成不同的酶是广泛存在的现象。许多单细胞生物生活在水溶液中,溶液的温度、pH、培养基成分经常变化,细胞必须随时改变酶的成分,才能适应周围环境的变化。 2、酵母菌小菌落在缺氧、有氧条件下,均酵解;大菌落无氧酵解,有氧进行有氧呼吸。 3、人类的乙醇脱氢酶、乙醛脱氢酶都是诱导酶。
不同条件下的基因激活. 1、细菌和其他单细胞生物中,不同条件下可形成不同的酶是广泛存在的现象。许多单细胞生物生活在水溶液中,溶液的温度、pH、培养基成分经常变化,细胞必须随时改变酶的成分,才能适应周围环境的变化。 2、酵母菌小菌落在缺氧、有氧条件下,均酵解;大菌落无氧酵解,有氧进行有氧呼吸。 3、人类的乙醇脱氢酶、乙醛脱氢酶都是诱导酶。")
55
一、基因表达调控的例征 (二)不同发育阶段的基因活性 多细胞生物的不同组织细胞的基因活性仍有很大差别。
1、脊椎动物细胞内的乳酸脱氢酶的变化:LDH有五类:LDH1—LDH5。 小鼠卵细胞中,只有LDH1,大约发育到第9天,主要是LDH5,再发育又出现LDH1,表明编码LDH1和 LDH5的基因在不同时期分 别处于打开和关闭的状态。 2、人的血红蛋白组成的变化:血红蛋白用于氧的运输,在人发育的各个阶段中,血红蛋白的组成各不相同。 早期胚胎(1-3个月):HbGower1、HbGower2、HbPotland三种血红 蛋白。 胎儿:HbF、HbA 新生儿:HbF70—80%、HbA30—20% 成人:HbA95%、HbA22.5%、HbF<1%
:HbGower1、HbGower2、HbPotland三种血红. 蛋白。 胎儿:HbF、HbA. 新生儿:HbF70—80%、HbA30—20% 成人:HbA95%、HbA22.5%、HbF<1%")
56
二、原核类基因表达的调控* 细菌和噬菌体的基因调控多数发生在转录水平上,操纵子是细菌主要的基因调控单位,即转录单位。
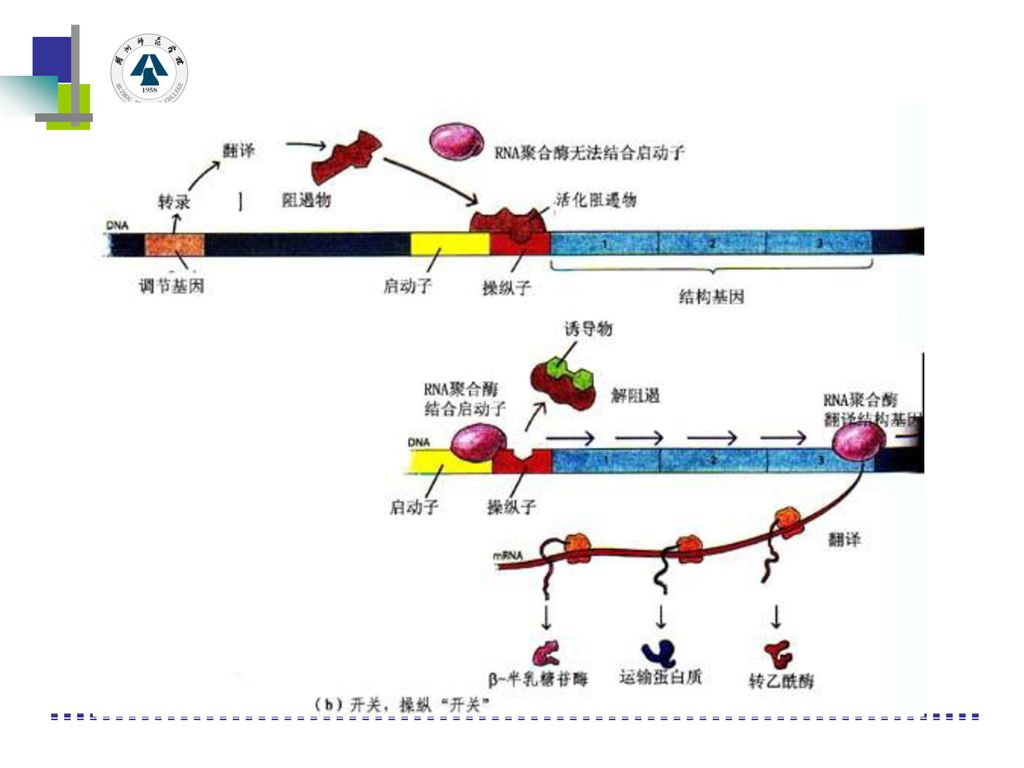
1960年雅各布和莫诺共同提出了乳糖操纵子模型。其调控机理: 乳糖操纵子=启动基因(P)+操纵基因(O)+结构基因(lacZ、lacY和lacA三个)组成,操纵子的前边有一个调节基因R。 乳糖操纵子的调节基因R能产生一种阻遏物(阻遏蛋白),当培养基中没有乳糖时,阻遏蛋白与操纵基因相结合,阻止RNA聚合酶的前进,mRNA转录就不能进行,乳糖代谢所需三种酶就不能合成。当培养基中有乳糖时,乳糖与阻遏蛋白结合,使阻遏蛋白构型发生变化,失去与操纵基因结合的能力,这时结构基因可转录出mRNA,进一步合成乳糖代谢所需三种酶利用乳糖。
+操纵基因(O)+结构基因(lacZ、lacY和lacA三个)组成,操纵子的前边有一个调节基因R。 乳糖操纵子的调节基因R能产生一种阻遏物(阻遏蛋白),当培养基中没有乳糖时,阻遏蛋白与操纵基因相结合,阻止RNA聚合酶的前进,mRNA转录就不能进行,乳糖代谢所需三种酶就不能合成。当培养基中有乳糖时,乳糖与阻遏蛋白结合,使阻遏蛋白构型发生变化,失去与操纵基因结合的能力,这时结构基因可转录出mRNA,进一步合成乳糖代谢所需三种酶利用乳糖。")
58
三、真核类基因表达的调控 (一)真核生物与原核生物基因表达调控的差异**
1、分子水平上的区别:真核细胞基因组的含量和基因数量远远高于原核细胞,并且DNA与组蛋白结合成核小体结构,同时与非组蛋白结合成染色体。 2、细胞水平上的区别:真核细胞的染色体是被包在核膜内,转录和翻译分别在细胞核和细胞质中进行。 3、个体水平上的区别:绝大多数真核生物都是复杂的多细胞有机体,个体发育过程复杂,不同组织细胞的基因,总是在不同的时空序列中被活化或阻遏。 因此,真核细胞基因表达的调控是一种多层次的调控系统,它涉及到DNA水平、转录水平、转录后水平、翻译水平等多个调控点。

59
三、真核类基因表达的调控 (二)真核类基因不同层次的表达和调控类型 1、染色体、DNA水平上的调控
(2)基因扩增:改变基因数量使之迅速增加而调节基因表达方式。即短期内细胞内大量产生某一基因拷贝的手段。如非洲爪蟾卵母细胞中rDNA的拷贝数可由平时的1500份急剧增加到200万份。它是适应胚胎发育时对核糖体的需求。 (3)转座因子:引起插入部位基因的开启和关闭。
基因扩增:改变基因数量使之迅速增加而调节基因表达方式。即短期内细胞内大量产生某一基因拷贝的手段。如非洲爪蟾卵母细胞中rDNA的拷贝数可由平时的1500份急剧增加到200万份。它是适应胚胎发育时对核糖体的需求。 (3)转座因子:引起插入部位基因的开启和关闭。")
60
三、真核类基因表达的调控 2、转录水平上的调控 (1)非组蛋白的调控作用:小鼠染色质重建实验说明非组蛋白对转录起着重要的作用。
骨髓细胞染色质→分离组分→DNA、组蛋白、非组蛋白 胸腺细胞染色质→分离组分→DNA、组蛋白、非组蛋白

61
三、真核类基因表达的调控 2、转录水平上的调控 (2)激素的调控作用:真核生物的细胞明显受激素的调控。在摇蚊、果蝇等双翅目昆虫幼虫的唾腺中的多线染色体上可看到一条条有特征的横纹。在幼虫、蛹期的各个发育阶段中,某些横纹变得疏松、膨大,这些疏松膨大区称为疏松区。疏松区出现一段时间后消失,它的出现有一定的时间程序。据测知,疏松区部位可以合成大量的RNA,而且具mRNA的性质。如果用蜕皮激素处理幼虫或离体的唾腺细胞,可诱发特定的基因转录。
激素的调控作用:真核生物的细胞明显受激素的调控。在摇蚊、果蝇等双翅目昆虫幼虫的唾腺中的多线染色体上可看到一条条有特征的横纹。在幼虫、蛹期的各个发育阶段中,某些横纹变得疏松、膨大,这些疏松膨大区称为疏松区。疏松区出现一段时间后消失,它的出现有一定的时间程序。据测知,疏松区部位可以合成大量的RNA,而且具mRNA的性质。如果用蜕皮激素处理幼虫或离体的唾腺细胞,可诱发特定的基因转录。")
62
三、真核类基因表达的调控

63
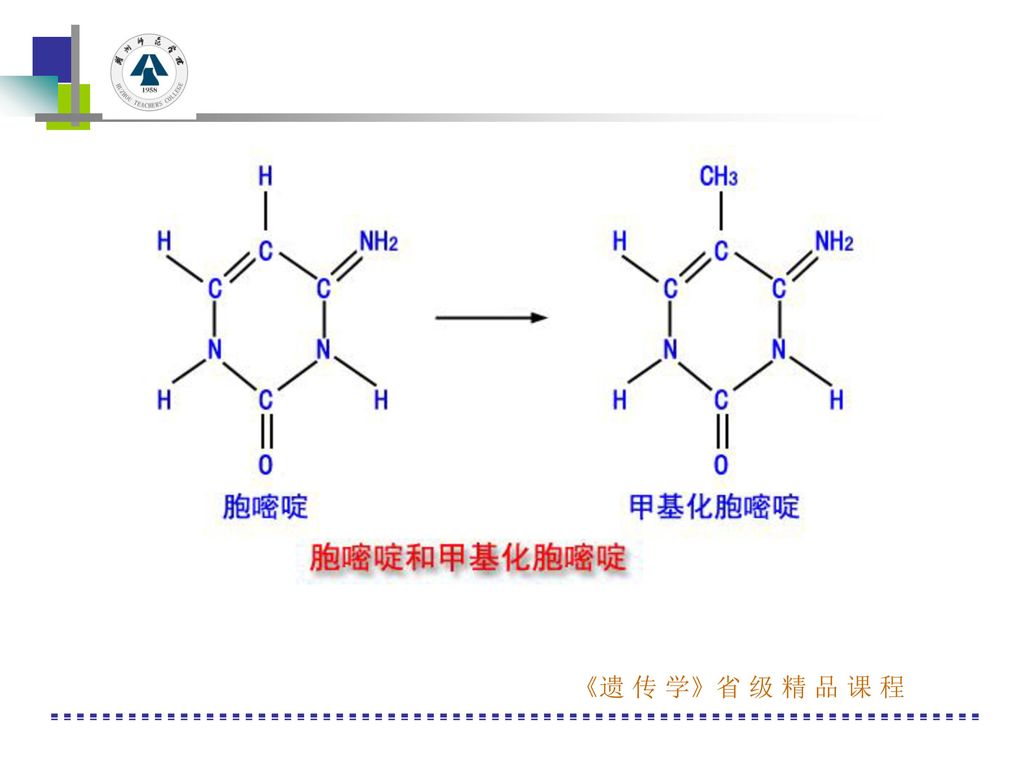
三、真核类基因表达的调控 2、转录水平上的调控 (3)异染色质化和修饰作用:真核生物可以通过改变染色体某些区域的异染色质化程度,从而调节基因的表达(巴氏小体)。真核细胞的修饰作用主要途径是通过胞嘧啶5′位上的甲基化反应来实现的。甲基化胞嘧啶即5′-甲基胞嘧啶不转录。 在不转录的DNA中GC有70%以上甲基化,而转录活性较高的DNA中,GC只有20-30%的甲基化。

65
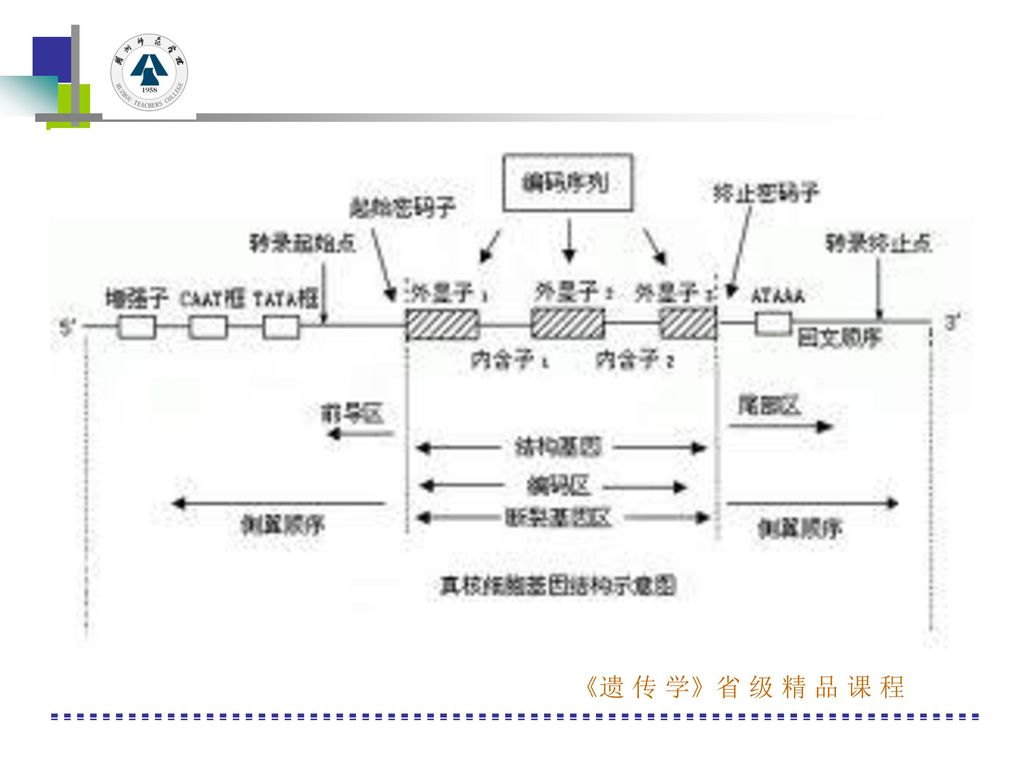
三、真核类基因表达的调控 (4)真核类基因转录调控的模式:从5′端开始,依次是增强子、CAAT框、TATA框、加帽点、起始密码子、外显子、内含子、终止密码子多聚A填加位点。 增强子:能够增强与之连锁的基因转录活性的调控序列。位于5′端启动子上-100bp。 CAAT框:在转录起始点的5′端-80—-70位置之间,共通顺序是GGTCAATCT,属于启动区域,作用不很肯定,一般认为只影响总的转录量,而不影响转录的位置。当此段顺序被改变,mRNA形成的量明显下降。 TATA框:在mRNA转录起始点的5′端上游20-30核苷酸地方的TATA框顺序。这是RNA聚合酶的重要接触点,可使酶定位在正确位置上而开始转录,当此框变化时,转录从不正常位置起始,转录水平下降。

67
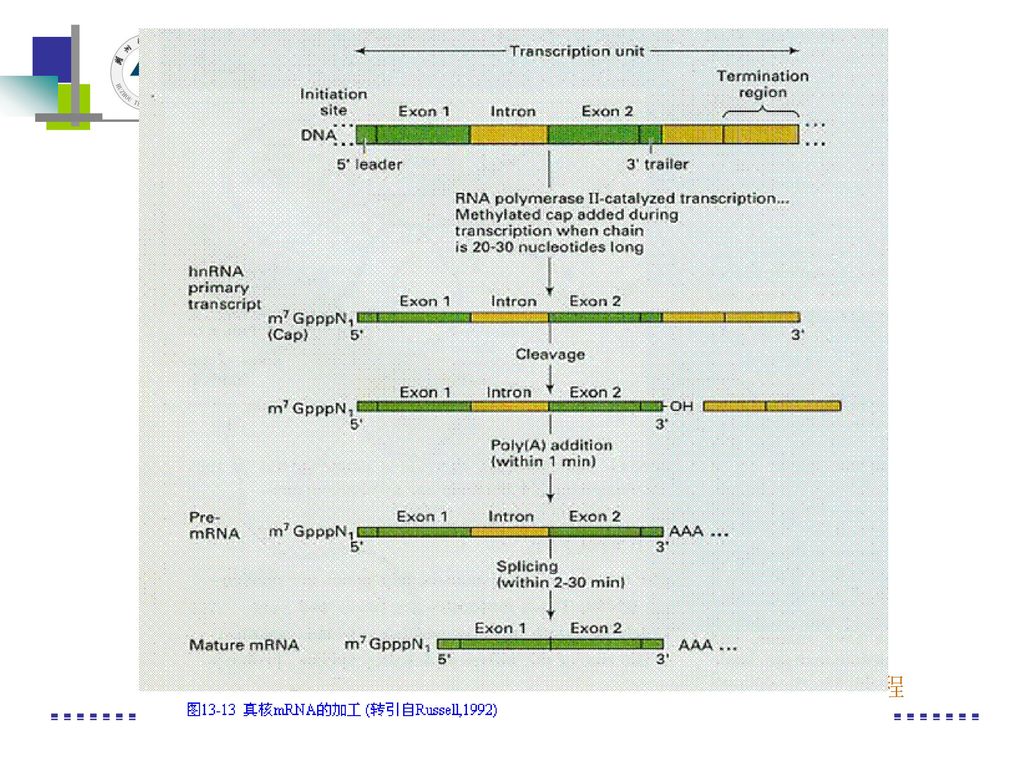
三、真核类基因表达的调控 3、RNA加工水平上的调控 (1)对mRNA前体(hnRNA)加工,主要在5′端加帽,即添加三磷酸甲基鸟苷(m7G-ppp-)。 (2)在3′端加多聚A尾巴,50—200bp的polyA。 (3)切除内含子,使hnRNA→mRNA。由于剪切的形式不同可产生功能不同的mRNA。
切除内含子,使hnRNA→mRNA。由于剪切的形式不同可产生功能不同的mRNA。")
69
真核生物翻译调节的主要形式是控制mRNA的稳定性(延长寿命)。
三、真核类基因表达的调控 4、转译水平上的调控** 真核生物翻译调节的主要形式是控制mRNA的稳定性(延长寿命)。 (1)mRNA 5′端的加帽作用,以及3′端加多聚A尾巴,都有助于提高mRNA分子的稳定性。 (2)某些真核生物中,mRNA进入细胞质后,不立即作为模板翻译蛋白质,而是与一些蛋白质结合成RNA-蛋白质颗粒(RNP),以失活的状态存在,从而延长寿命,提高其稳定性。例如种子萌发时,甚至没有RNA的合成,也能合成蛋白质,所以种子里一定贮存着mRNA。古莲子在地下埋藏1700多年之后,仍能出芽,可见其mRNA寿命之长。
。 (1)mRNA 5′端的加帽作用,以及3′端加多聚A尾巴,都有助于提高mRNA分子的稳定性。 (2)某些真核生物中,mRNA进入细胞质后,不立即作为模板翻译蛋白质,而是与一些蛋白质结合成RNA-蛋白质颗粒(RNP),以失活的状态存在,从而延长寿命,提高其稳定性。例如种子萌发时,甚至没有RNA的合成,也能合成蛋白质,所以种子里一定贮存着mRNA。古莲子在地下埋藏1700多年之后,仍能出芽,可见其mRNA寿命之长。")
70
作 业 1、名词 体细胞克隆 阻遏蛋白 2、乳糖操纵子的结构?乳糖操纵子模型的调 控机制? 3、真核生物与原核生物基因表达调控的差异?

Similar presentations

 导致胎儿缺氧的母体因素有 ①微小动脉供血不足:如妊高征等 ②红细胞携氧量不足:如重度贫血、一氧化碳中毒等;>")




 类型 :>")













