Download presentation
Presentation is loading. Please wait.

1
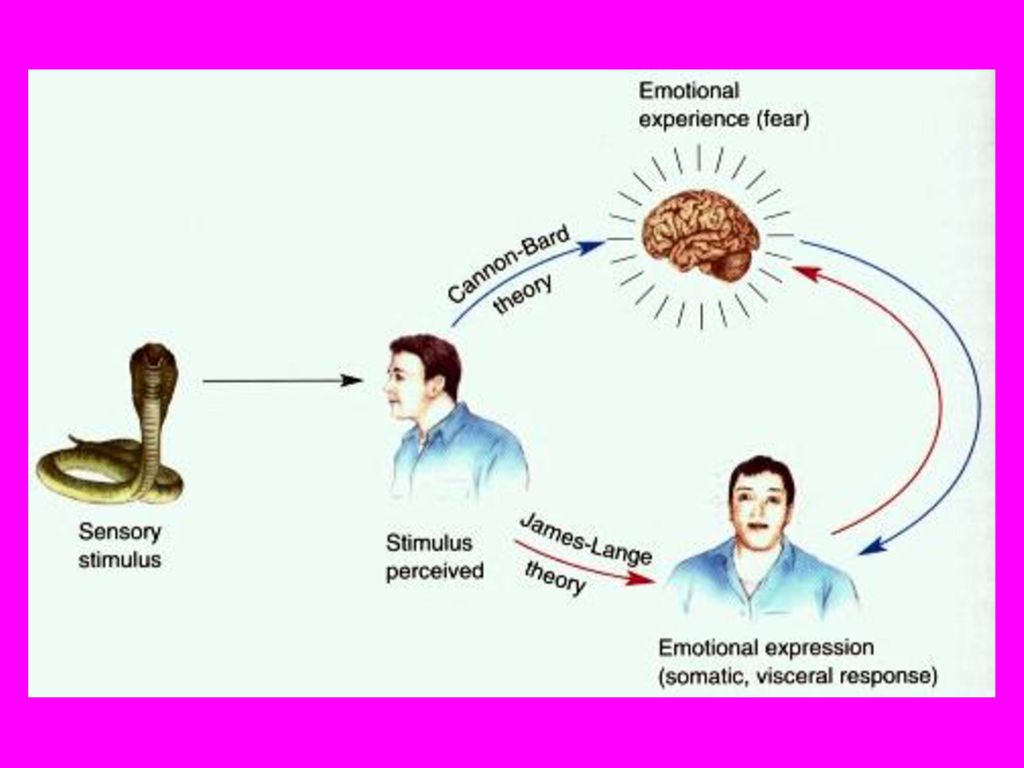
第十七章 情绪的脑机制 情绪活动对人类有重要意义,其表达占据人类生活的很大一部分。 情绪机制的研究有很大难度,必须把情绪体验与情绪表达加以区别. 情绪反应通常来自身体对外界内外环境的感觉,并由运动系统、自 主神经系统和下丘脑内分泌系统所控制。而情绪体验我们了解甚少. 现在对情绪神经机制研究主要来自:有关动物情绪的表达和临床病 例研究这两个方面。情绪研究可简化为一个关于输入-输出的问题。 问题是输入的感觉刺激如何引起代表情绪表达的行为和生理反应。 第一节 什么是情绪 情绪,例如爱、恨、厌恶、喜欢、嫉妒、内疚、恐惧和焦虑等我们 可经常体验到。但我们很难精确地给它们下定义。有两种不同学说. 一. 情绪学说 19世纪著名科学家达尔文和弗洛伊德研究过脑在情绪表达中的作用. 1. James-Lange学说 认为情绪体验是对身体生理变化的反应。例如因为我们哭了,所以 我们感到悲伤,而不是因为感到悲伤才哭。感觉系统把环境信息传 给大脑,大脑把加工后的信号传到身体并作出一定反应,感觉系统

2
再对这些变化作出相应的反应。认为生理变化产生情绪。如果这些
生理变化被解除,情绪也随之消失。这种今天看来不可思议的想法, 被同时代的许多人所接受。James的思维实验说明,生理变化与情 绪的关系。消除愤怒时的生理变化很难保持愤怒的情绪。但这并不 意味没有明显的生理变化信号就不可能有情绪的产生。 2. Cannon-Bard学说 James-Lange学说流行不久就受到攻击。产生了Cannon-Bard情绪 学说,认为在没有情绪表达的情况下情绪体验也可以发生。 Cannon-Bard学说与James-Lange学说的的第一个对立之处是, 认为 即使没有感觉到生理变化, 情绪也可以被体验到。脊髓横断的病人, 丧失断面以下的躯体感觉,却仍有体验情绪的表现。另一个对立处 是认为情绪体验和身体的生理状态之间没有必然的相关。如恐惧产 生心跳加快等现象,但在愤怒或无情绪生病 发烧时也出现这些生理变化。Cannon-Bard 学说强调在情绪体验中丘脑所起的特定作用。 认为大脑皮层的刺激-反应神经回路与情绪无 关。右图示两学说间的不同。

4
二. 情绪机制的实验研究 实验研究证明, 每种学说既有优点也有不足。例如恐惧和愤怒都激 活自主神经系统的交感支,但它们分别与不同的生理反应相关联。 虽然还未能证明这两种情绪是不同生理反应的产物,但至少二者的 生理反应是不同的。这与Cannon-Bard学说不一致。另一个被实验 研究证实与Cannon-Bard学说不一致的观点是,情绪有时受到脊髓 损伤的影响,但结果不一致。另外,如果强迫微笑的情绪行为表达, 使人感到比较愉快。有的情绪体验依赖情绪行为的表达,有的不依 赖。因此,对情绪产生的机制有待更多的研究积累。 第二节 边缘系统 特定的感觉信息沿特定的解剖学通路传递到新皮层,形成特定的感 觉系统。研究发现脑内存在与情绪体验有关的系统,称为边缘系统. 一. Broca 边缘叶 1878年法国神经学家Broca发现所有哺乳动物小脑内侧表面有一组 与周围皮层截然不同的结构, 它围绕脑干形成一个环或边界, 将它 命名为边缘叶(limbic lobe)。它由胼胝体周围的皮层,主要是扣带 回和颞叶内表面皮层(包括海马)组成, 起初一直认为它与嗅觉有关,
。它由胼胝体周围的皮层,主要是扣带. 回和颞叶内表面皮层(包括海马)组成, 起初一直认为它与嗅觉有关,")
5
后来的研究证明, 这些结构与情绪有密切的关系。
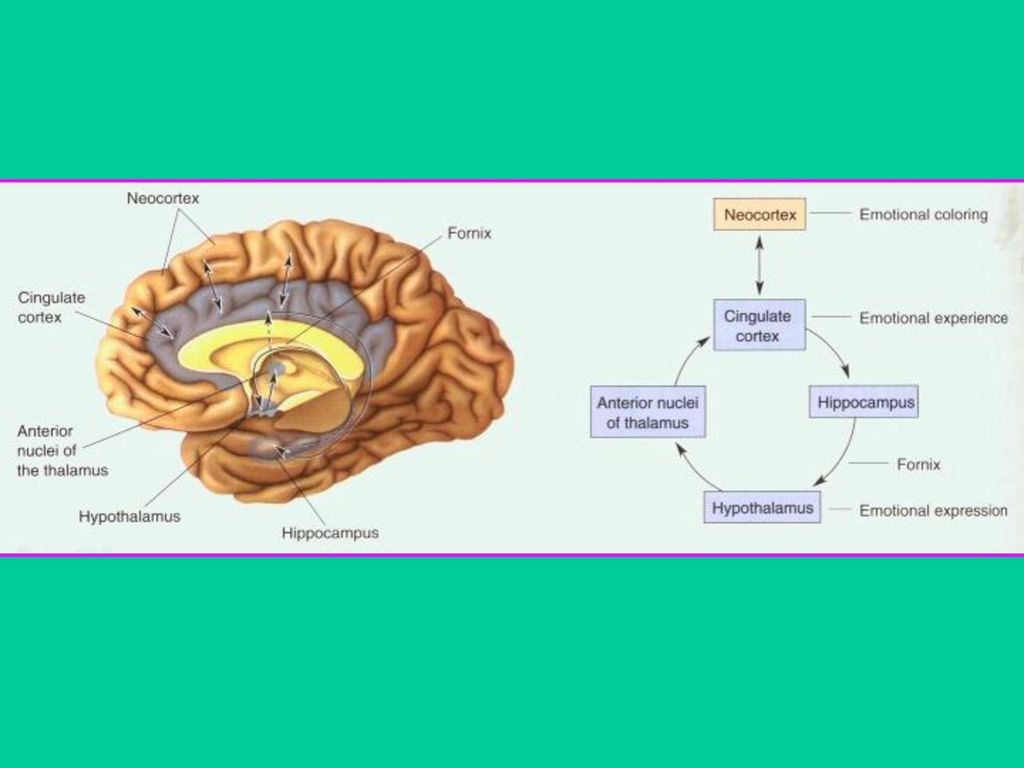
二. Papez 回路 美国神经学家Papez认为在脑内侧壁有一个情绪系统, 它把新皮层 和下丘脑互相连接起来, 它在情绪体验和情绪表达中起关键作用, 被称为Papez回路(下图)。当肿瘤使扣带回受损时,引起恐惧、激 怒和压抑等情绪失调。证明下丘脑整合自主神经系统的活动并控 制情绪的行为表达,把下丘脑和新皮层连接起来,使二者能彼此 影响。这种双向交流说明Papez回路既符合James-Lange学说也符 合Cannon-Bard学说。解剖学研究证明 Papez 回路的存在, 也证明 其结构与情绪有关。 Papez回路与Broca边缘叶之间具有相似性,它们通常被称为边缘系 统(limbic system)。认为边缘系统是脑的三大基本功能系统之一, 即爬行脑、旧哺乳脑和新哺乳脑中的旧哺乳脑。边缘系统的出现使 动物具有体验和表达情绪的能力,使动 物从由脑干(爬行脑)所支配的刻板行为 中解放出来。同样,新皮层(新哺乳脑) 的出现使动物具有更高级、更复杂功能。
。当肿瘤使扣带回受损时,引起恐惧、激. 怒和压抑等情绪失调。证明下丘脑整合自主神经系统的活动并控. 制情绪的行为表达,把下丘脑和新皮层连接起来,使二者能彼此. 影响。这种双向交流说明Papez回路既符合James-Lange学说也符. 合Cannon-Bard学说。解剖学研究证明 Papez 回路的存在, 也证明. 其结构与情绪有关。 Papez回路与Broca边缘叶之间具有相似性,它们通常被称为边缘系. 统(limbic system)。认为边缘系统是脑的三大基本功能系统之一, 即爬行脑、旧哺乳脑和新哺乳脑中的旧哺乳脑。边缘系统的出现使. 动物具有体验和表达情绪的能力,使动. 物从由脑干(爬行脑)所支配的刻板行为. 中解放出来。同样,新皮层(新哺乳脑) 的出现使动物具有更高级、更复杂功能。")
7
第三节 恐惧与焦虑 感觉信息如何引起焦虑和恐惧相关的生理和行为反应?有证据表明, 位于颞叶内的杏仁核在其中起关键作用。 一. Klüver-Bucy 综合症 研究发现切除或损伤猴的双侧颞叶严重影响其恐惧反应, 表现出许 多异乎寻常的变态行为反应。并归纳为精神盲、口倾向、复变态、 性行为改变和情绪改变五类。总称Klüver-Bucy 综合症。 切除颞叶的猴子能看见物体,却不能识别,称精神盲;它们捡起视 野内每件物体察看,然后放入口中一一鉴别,无论其是否可食,称 口倾向;它们有难以抗拒的检查物体的倾向,情不自禁地靠近并触 摸每一件物体,然后放入口中,称复变态;它们对性兴趣明显增大, 包括异性性行为、同性性行为和手淫。最典型的情绪改变是对恐惧 反应的明显降低。对实验者不但不逃避,而且主动靠近和触摸人。 对猴的天敌蛇也无恐惧反应,在受到攻击后仍再次靠近蛇并试图观 察它。这并不是由于愚蠢和记忆丧失,而确实是情绪降低造成的。 这在切除颞叶的病人身上也都可以见到所有这些症状,包括视觉识 别有问题,口倾向和性欲亢进,情绪行为非常呆板。

8
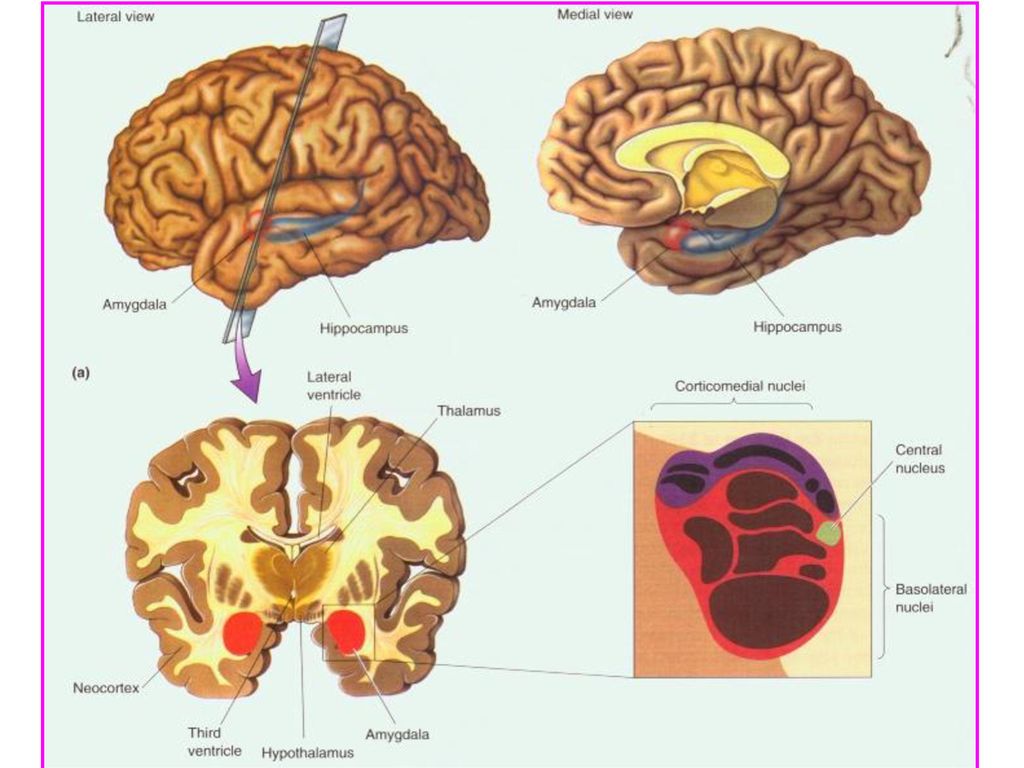
二. 杏仁核 上述颞叶切除中包括杏仁核、海马等皮层下结构。 Klüver-Bucy 综 合症中的情绪障碍可能是由于杏仁核损毁导致的。杏仁核除了与恐 惧反应有关外,还与许多情绪反应有关。 1. 杏仁核的解剖学 杏仁核位于颞叶端部、内侧颞皮层下方, 形似杏仁, 是一个核团复合 体,由三部分组成:基底外侧核群(basolateral nuclei)、皮层内侧核 群(corticomedial nuclei)和中央核(central nucleus)(下图)。杏仁核接 受来自海马、扣带回及新皮层各脑区的输入。特别注意:所有感觉 系统的信息都传至杏仁核,特别是其基底外侧核。不同感觉系统在 杏仁核有不同的投射模式。杏仁核各部分之间的相互联系使来自不 同感觉系统的信息在此整合。杏仁核和下丘 脑之间有两条主要通路:腹侧离杏仁核通路 (ventral amygdalofugal pathway)和终纹(stria terminalis)。 2. 损毁和刺激杏仁核产生的影响 动物实验证明, 双侧损毁杏仁核导致与Klü-
、皮层内侧核. 群(corticomedial nuclei)和中央核(central nucleus)(下图)。杏仁核接. 受来自海马、扣带回及新皮层各脑区的输入。特别注意:所有感觉. 系统的信息都传至杏仁核,特别是其基底外侧核。不同感觉系统在. 杏仁核有不同的投射模式。杏仁核各部分之间的相互联系使来自不. 同感觉系统的信息在此整合。杏仁核和下丘. 脑之间有两条主要通路:腹侧离杏仁核通路. (ventral amygdalofugal pathway)和终纹(stria. terminalis)。 2. 损毁和刺激杏仁核产生的影响. 动物实验证明, 双侧损毁杏仁核导致与Klü-")
10
ver-Bucy 综合症相似的呆板情绪反应。除了对攻击行为和记忆有影
响外,还严重降低动物的恐惧行为。例如,切除杏仁核的大鼠主动 接近猫并咬它的耳朵。现在认为这是由于基底外侧核损毁所致。损 毁杏仁核的Urbach-Wiethe病人能根据照片辨认人物,并能根据面 部表情分辨出高兴、悲伤和厌恶三种表情;但对愤怒和害怕不能辨 认。杏仁核损毁似乎选择性地降低了病人对恐惧行为的识别。反过 来,电刺激杏仁核将引起动物警惕性和注意力增加及猛烈的攻击行 为;在人则引起焦虑和恐惧。 三. 习得性恐惧 与恐惧相关的记忆能够很快形成,而且长久地保存。孩提时受到某 种痛苦伤害,以后再不会重复遭受同类伤害。杏仁核虽然不是记忆 储存的主要位点,但它似乎参与把情绪活动有关的内容传送到记忆 系统中。实验证明,动物经过学习,有关痛苦的刺激能引起恐惧性 的条件反射,如心率加快。训练后杏仁 中央核对这种痛苦刺激起反应。损毁杏 仁核将消除这种习得性的反应。习得性 恐惧的回路如右图示。

11
第四节 愤怒与攻击 攻击可分为多种类型。攻击的目的可能是捕食、护幼、争夺配偶和 恐吓潜在的敌人等等。一些研究表明不同类型的攻击行为具有不同 的神经基础。雄激素(androgen)水平是影响攻击的一个因素。动物 雄激素季节性变化和攻击行为之间相互关联。这在人类不明显。 捕食性攻击(pedatory aggression)和情感性攻击(affective aggression) 是不同性质的攻击。捕食性攻击时动物不出现交感神经系统活动的 增强,不发出声音,直接瞄准捕向猎物;而情感性攻击伴有交感神 经系统活动的增强,一般都发出叫声摆出威胁性或防御性的姿势。 这两种类型的攻击行为及其生理表现都由躯体运动系统和自主神经 系统介导,但二者的神经传导通路上有某些不同,从而造成在行为 反应上的显著差异。 一. 下丘脑与攻击行为 下丘脑是与攻击行为相关的重要结构之一。 1. 假怒 实验显示切除双侧大脑皮层的动物在行为上出现显著变化。术前不 易激怒的动物,术后小小的刺激则使之处于狂怒状态。这种状态称
和情感性攻击(affective aggression) 是不同性质的攻击。捕食性攻击时动物不出现交感神经系统活动的. 增强,不发出声音,直接瞄准捕向猎物;而情感性攻击伴有交感神. 经系统活动的增强,一般都发出叫声摆出威胁性或防御性的姿势。 这两种类型的攻击行为及其生理表现都由躯体运动系统和自主神经. 系统介导,但二者的神经传导通路上有某些不同,从而造成在行为. 反应上的显著差异。 一. 下丘脑与攻击行为. 下丘脑是与攻击行为相关的重要结构之一。 1. 假怒. 实验显示切除双侧大脑皮层的动物在行为上出现显著变化。术前不. 易激怒的动物,术后小小的刺激则使之处于狂怒状态。这种状态称.")
12
为假怒,因为通常在这种情况下不至于引起愤怒,而现在却有愤怒
反应的种种表现;或虽有愤怒反应的行为表现,而无实际攻击行为. 损毁下丘脑可以反转双侧大脑皮层切除引起的假怒行为效应。只损 毁下丘脑前部不能反转假怒,而只切除后半部下丘脑就能使假怒现 象不再出现。提示下丘脑后半部可能对于愤怒和攻击行为的表达特 别重要。通常这种功能被大脑皮层抑制。 2. 下丘脑电刺激实验 在研究电刺激间脑造成的行为影响时发现, 电刺激下丘脑可引起动 物打喷嚏、喘气、进食、恐惧和愤怒等行为反应以及心率变化、瞳 孔放大及肠胃蠕动等现象。这提示下丘脑有两个基本功能:内环境 稳定和情绪表达。认为下丘脑是情绪表达系统中一个重要组成部分. 电刺激下丘脑引起的动物愤怒表达与大脑皮层损毁后动物表现出的 假怒现象很相似。低强度电刺激下丘脑如猫受到敌人威胁时一样, 表现出流口水、嘶叫、耳朵向后、毛发竖立等复杂行为。有时突然 逃跑,如同躲避想象中的攻击者。当刺激强度增加,猫会做出一个 真正的攻击反应,用爪子猛烈敲打或扑向假想敌人。刺激停止,猫 的愤怒行为立刻消失,甚至蜷成一团睡觉。

13
后来的研究发现,刺激下丘脑内侧部引起情感性攻击,动物弓背、
嘶叫、流口水,但并不发动真正攻击。刺激下丘脑外侧部引起捕食 性攻击,动物稍稍弓起背,毛发轻微竖起,迅速扑向大鼠并狠狠咬 住其颈部。没有情感性攻击时那种明显的威胁姿势。 二. 中脑与攻击行为 下丘脑通过两条通路把信号传送至脑干: 内侧前脑束(medial forebrain bundle)和背侧纵束(dorsal longitudinal fasciculus)。下丘 脑外侧部的传出纤维组成部分内侧前脑束,投射到中脑被盖腹侧区 (ventral tegmental area, VTA)。如同刺激下丘脑外侧部那样,电刺 激VTA使动物表现出捕食性攻击的行为特征。相反,损毁VTA则减 少动物的捕食性攻击。内侧前脑束被切断后,电刺激下丘脑则不引 起动物的攻击行为。提示下丘脑是通过影响VTA而参与攻击行为的. 下丘脑内侧部传出纤维经背侧纵束至中脑导水管周围灰质(periaqu- eductal gray mater)。如同刺激下丘脑内侧部那样,电刺激中脑导 水管周围灰质使动物发动情感性攻击,而损毁这部分结构则减少情 感性攻击行为。
和背侧纵束(dorsal longitudinal fasciculus)。下丘. 脑外侧部的传出纤维组成部分内侧前脑束,投射到中脑被盖腹侧区. (ventral tegmental area, VTA)。如同刺激下丘脑外侧部那样,电刺. 激VTA使动物表现出捕食性攻击的行为特征。相反,损毁VTA则减. 少动物的捕食性攻击。内侧前脑束被切断后,电刺激下丘脑则不引. 起动物的攻击行为。提示下丘脑是通过影响VTA而参与攻击行为的. 下丘脑内侧部传出纤维经背侧纵束至中脑导水管周围灰质(periaqu- eductal gray mater)。如同刺激下丘脑内侧部那样,电刺激中脑导. 水管周围灰质使动物发动情感性攻击,而损毁这部分结构则减少情. 感性攻击行为。")
14
三. 杏仁核与攻击行为 杏仁核也与攻击行为有关。杏仁核双侧损毁的猴王将降为最低的位 置,原来排名第二的猕猴成为新猴王;然后再损伤新猴王双侧杏仁 核,同样这只猴的地位降至最低。提示杏仁核对攻击行为是非常重 要的,而攻击能力的强弱直接决定猕猴在社会群体中的地位。据此 一些神经外科医生认为切除杏仁核有可能降低那些具有暴力倾向人 的攻击行为。临床报告显示,这类手术在降低攻击行为、增强注意 力、降低过多的活动、减少癫痫发作等方面取得相当大的成功。这 类手术称为精神手术(psychosurgery)。20世纪早期常用于治疗与焦 虑、攻击等行为有关的严重疾病或神经官能症。但这种手术损毁相 当大一部分脑组织,是一种很危险的方法,只有等手术恢复后才能 了解是否损害了病人的认知能力或行为功能。 杏仁核对攻击行为起多方面的影响。电刺激基底外侧核导致感情性 攻击,损毁基底外侧核降低动物的感情性攻击。皮层内侧核对攻击 行为有抑制作用。损毁皮层内侧核或终纹显著地增强动物的捕食性 攻击行为。因此通常情况下,皮层内侧核可能对下丘脑起抑制作用, 从而抑制捕食性攻击行为。
。20世纪早期常用于治疗与焦. 虑、攻击等行为有关的严重疾病或神经官能症。但这种手术损毁相. 当大一部分脑组织,是一种很危险的方法,只有等手术恢复后才能. 了解是否损害了病人的认知能力或行为功能。 杏仁核对攻击行为起多方面的影响。电刺激基底外侧核导致感情性. 攻击,损毁基底外侧核降低动物的感情性攻击。皮层内侧核对攻击. 行为有抑制作用。损毁皮层内侧核或终纹显著地增强动物的捕食性. 攻击行为。因此通常情况下,皮层内侧核可能对下丘脑起抑制作用, 从而抑制捕食性攻击行为。")
15
四. 5-羟色胺与攻击行为 5-羟色胺能神经元位于脑干的中缝核群。这些神经元内侧前脑束上 行投射至下丘脑及与情绪有关的边缘系统。研究证明,5-羟色胺可 能参与抑制攻击行为。 在饲养笼隔离4周后,鼠通常变得活动过多,对其它鼠发动猛烈攻 击。隔离对5-羟色胺总体水平没有影响,但降低其合成、释放和重 新合成的速率。用阻断5-HT合成的药物PCPA处理后攻击行为X显 著增多。应用分子生物学技术可把小鼠编码5-HT1B受体的基因敲除 (gene knockout)。缺少5-HT1B受体的变异小鼠行为表现正常,群居 时没有相互攻击行为。但在外界环境压力下,它们的攻击行为明显 增多。解剖学研究表明,杏仁核、中脑导水管周围灰质、中缝核群 和基底神经节等与攻击行为有关的脑结构中,都有5-HT1B受体的分 布,这就解释了为什么5-HT1B受体激活后小鼠攻击性降低,缺乏这 一受体的小鼠攻击性增强。通常5-HT可能通过5-HT1B受体抑制杏 仁核、中脑导水管周围灰质和中缝核群神经元的活动,从而抑制攻 击行为。
。缺少5-HT1B受体的变异小鼠行为表现正常,群居. 时没有相互攻击行为。但在外界环境压力下,它们的攻击行为明显. 增多。解剖学研究表明,杏仁核、中脑导水管周围灰质、中缝核群. 和基底神经节等与攻击行为有关的脑结构中,都有5-HT1B受体的分. 布,这就解释了为什么5-HT1B受体激活后小鼠攻击性降低,缺乏这. 一受体的小鼠攻击性增强。通常5-HT可能通过5-HT1B受体抑制杏. 仁核、中脑导水管周围灰质和中缝核群神经元的活动,从而抑制攻. 击行为。")
16
第五节 强化和奖赏 在大鼠脑内埋植一个电极可随时进行电刺激。将大鼠置于一个有踏 板的盒子内,当大鼠踩到踏板时将受到一个短暂的电刺激(下图)。 起初,大鼠在盒子内随意活动,只偶尔踩到踏板时受到电刺激。不 久后,大鼠学会不断踩踏板。这种行为称为自我电刺激(electrical self-stimulation)。大鼠如此热衷于才踏板,以至有时它们忘记饮食, 吃水,只有当体力耗尽后才会停下来。由此发现了强化脑刺激这一 现象。这引发了随后许多的研究工作。 一. 自我电刺激与强化 上述实验中大鼠不断踩踏板的原因还不很清楚, 可能是大鼠从电刺 激中获得一种正向感觉, 希望获得更多刺激。引起强化脑刺激的脑 位点称为愉快中枢(pleasure center)。但这一术语可能在两方面引起 误解:一是我们并不知道大鼠是否体验到愉快的感觉, 它只是希望得到更多刺激,但刺激并不一定引起愉快 感觉。如有时一个人不由自主地一个劲喝酒,但喝酒 并不给人带来愉快感觉。第二即使刺激是愉快的,也 不一定存在特异的中心负责这种强化行为,可能有许
。大鼠如此热衷于才踏板,以至有时它们忘记饮食, 吃水,只有当体力耗尽后才会停下来。由此发现了强化脑刺激这一. 现象。这引发了随后许多的研究工作。 一. 自我电刺激与强化. 上述实验中大鼠不断踩踏板的原因还不很清楚, 可能是大鼠从电刺. 激中获得一种正向感觉, 希望获得更多刺激。引起强化脑刺激的脑. 位点称为愉快中枢(pleasure center)。但这一术语可能在两方面引起. 误解:一是我们并不知道大鼠是否体验到愉快的感觉, 它只是希望得到更多刺激,但刺激并不一定引起愉快. 感觉。如有时一个人不由自主地一个劲喝酒,但喝酒. 并不给人带来愉快感觉。第二即使刺激是愉快的,也. 不一定存在特异的中心负责这种强化行为,可能有许.")
18
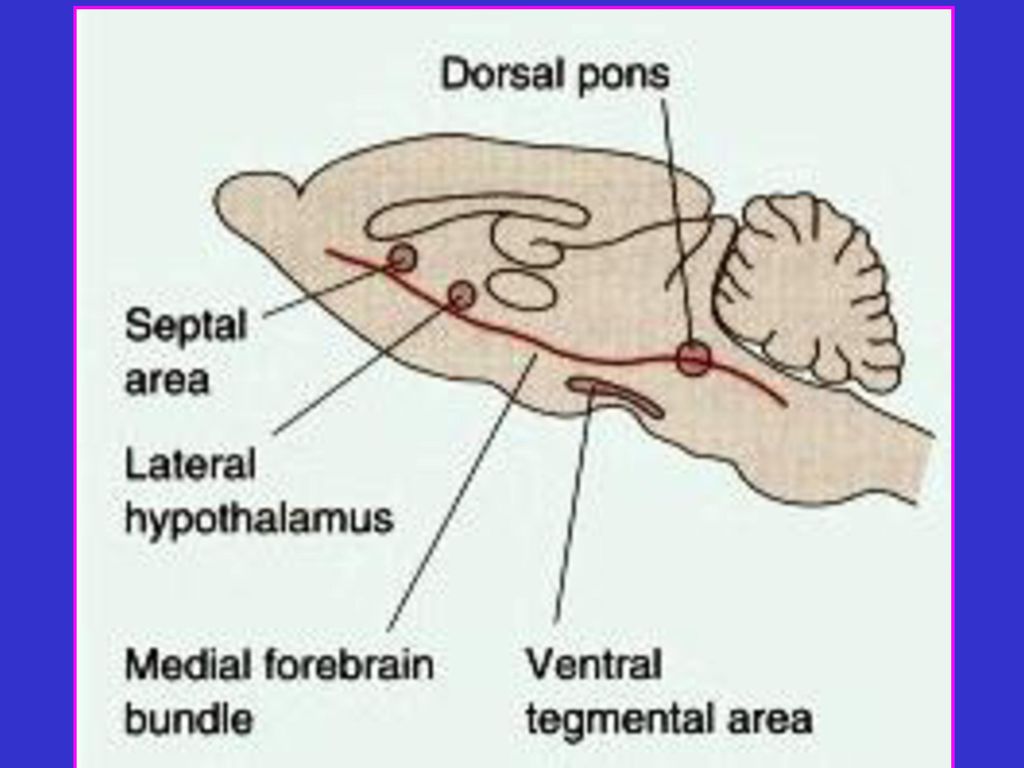
多散在的位点的刺激也可能引起自我电刺激行为;还电刺激可能只
影响纤维束,而不是核团。 在随后的数十年中,在大鼠脑的边缘结构和其他地方发现了许多自 我电刺激位点,包括隔区、下丘脑外侧部、内侧前脑束、中脑被盖 腹侧区和脑桥背侧部(下图)。其中,刺激前脑内侧束产生很强的强化 效应。另外,对某些脑区位点的电刺激产生回避行为,动物还能学 习某种操作以终止这些部位的电刺激。这些不愉快中枢(displeasure center)或负向强化位点位于下丘脑的内侧部和中脑被盖腹侧区的外 侧部。刺激这些部位引起负向感觉(恐惧)或产生负向强化行为。 二. 人的脑刺激 为了确定电刺激引起的感觉, 刺激人脑然后问其感觉。但这只能在某 些手术时要求病人在手术过程中处于清醒状态,对脑施加电刺激进 行。最常见的是严重癫痫的手术治疗。对脑电刺 激可产生愉快感觉,愉快中枢并非完全不适。但 病人选择的不断刺激位点并非引起愉快感觉的某 一位点,其原因可根据下列两例病例了解。 一个病人患有严重嗜睡病(narcolepsy), 可由清醒
。其中,刺激前脑内侧束产生很强的强化. 效应。另外,对某些脑区位点的电刺激产生回避行为,动物还能学. 习某种操作以终止这些部位的电刺激。这些不愉快中枢(displeasure. center)或负向强化位点位于下丘脑的内侧部和中脑被盖腹侧区的外. 侧部。刺激这些部位引起负向感觉(恐惧)或产生负向强化行为。 二. 人的脑刺激. 为了确定电刺激引起的感觉, 刺激人脑然后问其感觉。但这只能在某. 些手术时要求病人在手术过程中处于清醒状态,对脑施加电刺激进. 行。最常见的是严重癫痫的手术治疗。对脑电刺. 激可产生愉快感觉,愉快中枢并非完全不适。但. 病人选择的不断刺激位点并非引起愉快感觉的某. 一位点,其原因可根据下列两例病例了解。 一个病人患有严重嗜睡病(narcolepsy), 可由清醒.")
20
状态直接进入深睡状态。为了寻找使他保持清醒状态的自我刺激位
点,在其脑的不同部位埋植14根电极。刺激海马感受中等程度的愉 快。刺激中脑被盖区感到很清醒,但不愉快。他经常选择的刺激位 点是前脑隔区,感到很清醒和很舒服的感觉。 另一个癫痫病患者情况更复杂。刺激隔区和中脑被盖区引起愉快感 觉。刺激隔区产生性感觉,刺激中脑被盖区产生开怀畅饮的感觉。 刺激杏仁核或尾状核产生中等程度舒服感觉。病人经常自我刺激的 位点是内侧丘脑,引起愤怒的不愉快感觉。病人选择的理由是可引 起一种快要回忆起一件往事的感觉,想通过反复刺激回忆起来。 从以上可得出这样结论:人脑自我刺激的位点并不总引起愉快感觉. 三. 多巴胺和强化 大量散在位点能导致自我电刺激是因为这些散在位点可通过一条神 经通路与正常奖励行为的共同神经通路彼此联系。而且中脑腹侧被 盖区是多巴胺神经元细胞体所在部位,它们的纤维经内侧前脑束到 达脑的许多区域。多巴胺与强化和奖赏行为有联系,但对其本质还 不清楚。实际上有些实验并不支持内侧前脑束多巴胺传出纤维在强 化和奖赏行为中关键作用。它只在某些情况下对强化行为是重要的.

21
That is great!

22
第十八章 睡眠与觉醒的脑机制 睡眠是机体对环境的反应降低、与环境相互作用减弱的状态,很容 易逆转。昏迷和全身麻醉不易逆转,不属于睡眠。人大约1/3的时 间在睡眠中度过,其中1/4时间处于活跃的做梦状态。睡眠和摄食, 呼吸一样对生命极其重要。所有哺乳动物、鸟类和爬行类都睡眠, 如此普遍的行为必然有至关重要的生物学意义。有些生活在深水或 急流中的动物似乎有理由不睡眠,如水豚大约一分钟呼吸一次空气, 但它们的睡眠时间几乎与人一样多。 目前还没有一种公认的关于睡眠功能理论。最合适的设想有两种: 恢复理论和适应理论。但遗憾的是尚未鉴定出那些生理过程明确地 经睡眠而恢复,或在睡眠中生成哪些重要物质或降解毒素。而 睡眠的适应理论可表述为多种形式:睡眠可躲避危险,节约体能等. 第一节 REM睡眠和非REM睡眠 人在正常的一天中经历两种迥异的行为:觉醒与睡眠。睡眠又包含 分明的时相。一夜中有数次进入快动眼睡眠(rapid eye movement sleep, REM睡眠)状态。REM睡眠期间,脑电图看起来更像觉醒, 脑内呈现出梦的生动、详细的情景。REM睡眠相以外的睡眠时间
状态。REM睡眠期间,脑电图看起来更像觉醒, 脑内呈现出梦的生动、详细的情景。REM睡眠相以外的睡眠时间.")
23
称为非REM睡眠,期间脑内不生成复杂的梦。又称为慢波睡眠,
因其脑电图以大而慢的节律为主。觉醒、REM睡眠和非REM睡眠 是由脑功能三种不同状态造成的。每种状态伴有机体功能显著变化. 一. 非REM睡眠的特征 全身的肌张力下降, 运动减至最小。期间机体能运动,但脑很少指 挥 其进行运动,只是稍微调整体位。机体体温和能耗降低。由于 自主神经的副交感活性增高,心率、呼吸和泌尿均减慢,消化活动 增强。脑的能耗和神经元总体放电及精神活动都达日最低。被唤醒 后通常回忆不起有任何思维活动和梦境。为休闲的脑和可动的身体. 二. REM睡眠的特征 它是做梦的睡眠。被唤醒后可报告清晰、详细而生动的梦境,经常 有奇异的情节。期间脑电图呈现迅速、低电压的波动,看起来与活 跃而觉醒的脑无异。脑氧耗比清醒状态下高。骨骼肌张力几乎完全 丧失,躯体实际上动弹不了。呼吸肌继续工作,但很微弱。耳肌和 眼肌例外地惊人的活跃。眼睑闭合,眼球时而迅速地来回运动,准 确地预示着生动的梦,在眼动中和眼动后被唤醒者90-95%报告做 了梦。期间自主神经系统的交感活性占主导地位。心率加速而不规

24
则。体温控制系统基本不控制,深部体温开始下降。REM睡眠中
脑似乎不在休息。是一种活跃的脑和瘫痪的身体。 第二节 睡眠周期 睡眠过程中,脑反复循环于不同活动状态。健康成人睡眠总时间的 75%为非REM睡眠,25%为REM睡眠。非REM睡眠又分为4个不 同的时期。整个睡眠过程中,非REM睡眠与REM睡眠周期性的交 替,平均每90min重复一个周期(p347)。随睡眠进程非REM睡眠的 时程普遍缩短, 而REM睡眠延长。一夜的REM睡眠有一半发生在 最后1/3睡眠时间。最长可持续30-50min。当在两个REM睡眠期之 间似乎有30min的强制不应期,即在每个REM睡眠期后必须至少经 过30min的非REM睡眠后,下一个REM睡眠期才有可能出现。 非REM睡眠1期是过渡性睡眠,通常持续几分钟,是睡眠最浅,最 易被唤醒的阶段。在此期觉醒的脑电 α 节律变得不规则,并逐渐消 失.2期睡眠稍深,持续5-15min,脑电图有偶发的8-14Hz振荡及大 幅度的K-复合体(K-Complex; 即δ节律和下丘脑产生的睡眠梭形波 的复合波), 眼动几乎停止。 3 期睡眠期间脑电图出现大幅度的、慢 速的 δ 节律,眼和躯体停止运动。4 期睡眠最深,脑电图呈大幅的
。随睡眠进程非REM睡眠的. 时程普遍缩短, 而REM睡眠延长。一夜的REM睡眠有一半发生在. 最后1/3睡眠时间。最长可持续30-50min。当在两个REM睡眠期之. 间似乎有30min的强制不应期,即在每个REM睡眠期后必须至少经. 过30min的非REM睡眠后,下一个REM睡眠期才有可能出现。 非REM睡眠1期是过渡性睡眠,通常持续几分钟,是睡眠最浅,最. 易被唤醒的阶段。在此期觉醒的脑电 α 节律变得不规则,并逐渐消. 失.2期睡眠稍深,持续5-15min,脑电图有偶发的8-14Hz振荡及大. 幅度的K-复合体(K-Complex; 即δ节律和下丘脑产生的睡眠梭形波. 的复合波), 眼动几乎停止。 3 期睡眠期间脑电图出现大幅度的、慢. 速的 δ 节律,眼和躯体停止运动。4 期睡眠最深,脑电图呈大幅的.")
25
2Hz或小于2Hz的δ节律,持续20-40min。之后,又回到3期、2期,
随后又突然进入REM睡眠。 REM睡眠期间,脑电图出现快速的 β 节律,眼球运动迅速而频繁。下图为睡眠各期的脑电图。 研究提示,成人对睡眠的正常需要差异很大,从每晚5h至10h。平 均7. 5h,约68%青年在 h 之间。衡量睡眠充足与否的指标是 在觉醒时的状态。每个人需要一定量的睡眠以维持必要的觉醒。 第三节 睡眠与觉醒的机制 1940年前人们普遍认为睡眠是被动的过程:只要消除对脑的感觉输 入脑就会入睡。然而阻断脑的感觉输入,动物仍有觉醒-睡眠周期. 现在知道睡眠是一个主动过程,且要求多个脑区的同时参加。已知: ①对睡眠和觉醒控制最关键的是弥散调 制性神经递质系统。②脑干去甲肾上腺 素能和5-HT能神经元在觉醒期间放电, 增强觉醒状态;部分胆碱能神经元增强 关键的REM事件,而其他胆碱能神经 元在觉醒时活动。③弥散调制性神经递 质系统控制丘脑的节律活动,丘脑又控

26
制大脑皮层的脑电节律;丘脑缓慢的与睡眠相关的节律阻断感觉信
息输入皮层。④睡眠还涉及下行调制系统的活动。如做梦时脑通过 下行调制系统有效地抑制运动神经元。 一. 上行网状激活系统与觉醒 脑干损伤导致睡眠和昏迷, 提示该处神经元的活动是保持觉醒所必 须的。实验研究发现,损毁脑干中线结构导致类似于非REM睡眠 状态,但损毁外侧被盖阻断上行感觉输入不引起非REM睡眠状态. 反之,电刺激网状结构内的中脑中线被盖可使皮层从非REM睡眠 的慢速、节律性脑电活动转变为更警觉、更唤醒的状态,其脑电图 与觉醒相似。被刺激的这个区域被称为上行网状激活系统。 一些神经元在觉醒时放电频率增高,包括去甲肾上腺素能的蓝斑细 胞、5-HT能的中缝核细胞、基底前脑的胆碱能神经元。它们都直 接与整个丘脑、大脑皮层以及其他脑区形成突触。它们释放递质的 总效应是使神经元去极化,兴奋性增高,抑制节律性放电。这在丘 脑中际神经元表现最明显。 二. 入睡与非REM睡眠 入睡过程包含短时间内的一系列进行性改变, 然后发展为非REM睡

27
眠状态。期间大多数脑干调制神经元放电频率普遍降低。早期脑电
图有丘脑神经元固有节律产生的睡眠梭形波。随非REM睡眠进展, 梭形波消失,为缓慢的δ节律(小于4Hz)取代.δ节律似乎也是丘脑 细胞产生的;当其膜电位比产生梭形波时更负(比觉醒时负得更多) , 就产生δ节律.梭形波或δ节律期间神经元活动的同步化是由于丘脑内的(也许还有丘脑和劈柴之间的)神经互联的活动而产生。由于丘脑和大脑皮层之间强的双向兴奋性联系,彼此产生显著相互影响. 三. REM睡眠 其与非REM睡眠不同, 神经机制也截然不同。大脑皮层在REM睡 眠期间的活跃程度至少不低于觉醒时。如在REM睡眠期间视皮层 神经元自发放电,运动皮层神经元也高频放电。复杂的梦也需要大 脑皮层的活动。但皮层并非REM睡眠所必须。 REM睡眠的控制和其他脑功能状态的控制一样, 来自脑干深部, 特 别是脑桥的弥散调制神经递质系统。上位脑干的两大系统:即蓝斑 和中缝核群的放电频率随着REM睡眠的起始几乎降到零。而脑桥 胆碱能神经元的放电频率急剧上升。有证据表明,胆碱能神经元的 活动诱发REM睡眠。REM睡眠期间,可能是由于乙酰胆碱的作用,
取代.δ节律似乎也是丘脑. 细胞产生的;当其膜电位比产生梭形波时更负(比觉醒时负得更多) , 就产生δ节律.梭形波或δ节律期间神经元活动的同步化是由于丘脑内的(也许还有丘脑和劈柴之间的)神经互联的活动而产生。由于丘脑和大脑皮层之间强的双向兴奋性联系,彼此产生显著相互影响. 三. REM睡眠. 其与非REM睡眠不同, 神经机制也截然不同。大脑皮层在REM睡. 眠期间的活跃程度至少不低于觉醒时。如在REM睡眠期间视皮层. 神经元自发放电,运动皮层神经元也高频放电。复杂的梦也需要大. 脑皮层的活动。但皮层并非REM睡眠所必须。 REM睡眠的控制和其他脑功能状态的控制一样, 来自脑干深部, 特. 别是脑桥的弥散调制神经递质系统。上位脑干的两大系统:即蓝斑. 和中缝核群的放电频率随着REM睡眠的起始几乎降到零。而脑桥. 胆碱能神经元的放电频率急剧上升。有证据表明,胆碱能神经元的. 活动诱发REM睡眠。REM睡眠期间,可能是由于乙酰胆碱的作用,")
28
使丘脑和皮层的活动与它们在觉醒状态下非常相似。当蓝斑的去甲
肾上腺素能和中缝核群的5-HT神经元重新开始放电时,REM睡眠 期终止。下图示REM睡眠起始和终止时刻,脑桥胆碱能神经元及 蓝斑的去甲肾上腺素能和中缝核群的5-HT神经元的放电活动。 梦多发生在REM睡眠期。做梦时为什么很少有真正的活动呢?因 为控制睡眠过程的深部脑干系统有效地抑制脊髓运动神经元,阻止 下行的运动指令变成实际的运动。这种适应性机制防止我们在梦中 因为运动而伤害自己。有少数患REM睡眠行为疾病的老年人做梦 时经常有行为活动而反复受伤,原因是介导REM无张力的脑干系 统发生故障。损伤猫脑桥的某些部位能引起相似的症状。 四. 促睡因子 瞌睡是感冒、流感等感染性疾病的常见症状之一。机体对感染的免 疫反应与睡眠调控之间可能有直接联系。在剥夺睡眠动物中鉴定出 多种促睡物质,如山羊脑脊液中的胞壁 酰二肽,以及白介素-1等,它们能激活 免疫系统。它们促睡的机制还不清楚。 但可能睡眠有助于抵御感染。

29
第四节 REM睡眠和梦 如何解释梦的意义目前尚无一致认识。对梦的解释极大的依赖于对 REM睡眠而非梦本身的研究。因为REM睡眠可以客观地测量,而 梦则很难研究。因为看不到梦,和难以对其进行研究。 我们需要做梦吗?不知道,但机体似乎渴求REM睡眠。每次当受试者进入REM睡眠状态被唤醒时,就特异性地剥夺其REM睡眠。1-2分钟后他再次入睡时,必进入非REM睡眠状态。这样受试者整夜睡眠是比较纯粹的非REM睡眠。经过数天这种不愉快的处理后,受试者试图进入REM睡眠状态的次数大大增多。当最后被允许不受干扰地入睡时,其REM睡眠的时程延长,与被剥夺REM的时程成比例. Freud曾提出梦的多种功能,认为梦是对伪装起来的愿望的一种满 足,是表达性和攻击幻想的一种无意识的方式,而这种幻想在觉醒 时是被禁止的。近年对梦的解释发现更多的生物学基础。认为梦, 至少梦的某些奇异特征,代表大脑皮层的记忆或联系的形成过程, 这是REM睡眠期间脑桥随机放电激活的。但对随机放电如何触发许多梦的怪异性及其复杂而流畅的故事情节和一夜一夜反复出现的梦,尚无法解释。有证据表明,经历高强度学习后REM睡眠时程增加。

30
What a lovely day today!

Similar presentations

 装配 后调整检测.>")




声音的产生:声音是由于物体的振动产生的。 凡是发声的物体都在振动。振动停止,发声也停止。 ( 2 )声源:正在发声的物体叫声源。固体、液体、气体 都可以作为声源,有声音一定有声源。 ( 3 )声音的传播:声音的传播必须有介质,声音可以在.>")






.>")






