Download presentation
Presentation is loading. Please wait.

1
第九章 脂 类 代 谢 Metabolism of Lipids

2
第一节 脂类的概念及生理作用 第二节 脂肪的水解 第三节 甘油的代谢 第四节 脂肪酸的β-氧化作用

3
第一节 脂类的概念及生理作用 脂类(lipids)是生物体维持正常生命活动不可缺少的一大类有机化合物,是与糖类、蛋白质、核酸并列为四大类重要基本物质之一。脂类的一个共性就是难溶于水,易溶于有机溶剂。脂类的生物学功能主要包括:
是生物体维持正常生命活动不可缺少的一大类有机化合物,是与糖类、蛋白质、核酸并列为四大类重要基本物质之一。脂类的一个共性就是难溶于水,易溶于有机溶剂。脂类的生物学功能主要包括:")
4
同等重量计算,脂肪的代谢能量实际高达糖原的6倍
一、 供能、贮能 1 g 脂肪:38 kJ (9kcal) 1 g 葡萄糖:16.9kJ (4kcal) 脂肪是非极性化合物,以无水状态贮存 糖原是极性化合物,以水合形式贮存 同等重量计算,脂肪的代谢能量实际高达糖原的6倍
 1 g 葡萄糖:16.9kJ (4kcal) 脂肪是非极性化合物,以无水状态贮存. 糖原是极性化合物,以水合形式贮存. 同等重量计算,脂肪的代谢能量实际高达糖原的6倍.")
5
二、是生物细胞的结构物质 类脂:胆固醇,磷脂 其中的磷脂是构成细胞生物膜(Biomembrane)的重要结构物质。 三、生物活性物质 前列腺素,VD,性激素 四、缓冲作用
的重要结构物质。 三、生物活性物质 前列腺素,VD,性激素 四、缓冲作用")
6
脂的分类 (1) 单纯脂——是脂肪酸和醇类所形成的酯,其中典型的为甘油三酯。 (2) 复合脂——除醇类、脂肪酸外还含有其它物质,如磷酸、含氮化合物、糖基及其衍生物、鞘氨醇及其衍生物等。磷脂 (3) 其它脂——为一类不含有脂肪酸、非皂化的脂,包括萜类、前列腺素类和甾类化合物等。
 其它脂——为一类不含有脂肪酸、非皂化的脂,包括萜类、前列腺素类和甾类化合物等。 .")
7
单纯脂

8
复合脂 磷酸甘油脂,又称甘油磷脂,是最具有代表性的复合脂,广泛存在于动物、植物和微生物。磷脂甘油脂是细胞膜结构重要的组分之一,在动物的脑、心、肾、肝、骨髓、卵以及植物的种子和果实中含量较为丰富。最简单的磷酸甘油脂结构如图

9
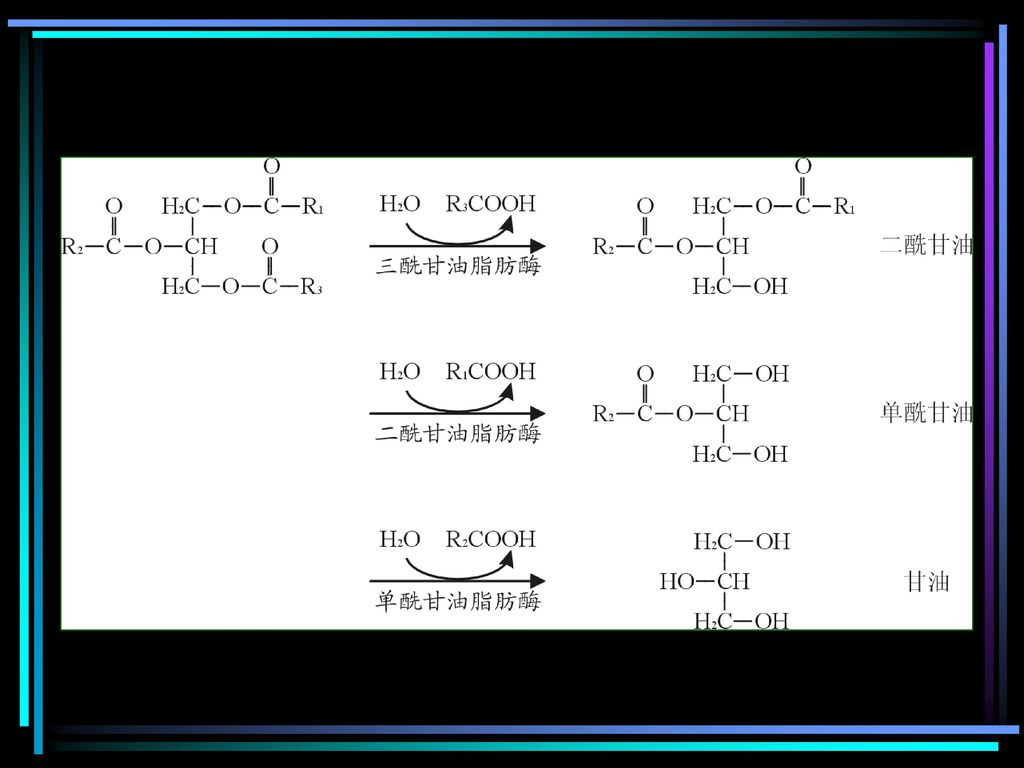
第二节 脂肪的水解

11
第三节 甘油的水解 甘油在甘油激酶的催化下,被磷酸化成3-磷酸甘油,然后氧化脱氢生成磷酸二羟丙酮。 其反应如下:

12
其中第一步反应需要消耗ATP,而第二步反应可生成还原辅酶Ⅰ。
磷酸二羟丙酮为磷酸丙糖,是糖酵解途径的中间产物,因此既可以继续氧化,经丙酮酸进入三羧酸循环彻底氧化成CO2和水,又可经糖异生作用合成葡萄糖,乃至合成多糖。

13
第四节 脂肪酸的水解 步骤 一、脂肪酸的活化(线粒体外膜) 二、脂肪酸的转运 三、脂肪酸的β-氧化过程(线粒体基质) 四、氧化磷酸化(线粒体基质)
 二、脂肪酸的转运 三、脂肪酸的β-氧化过程(线粒体基质) 四、氧化磷酸化(线粒体基质)")
14
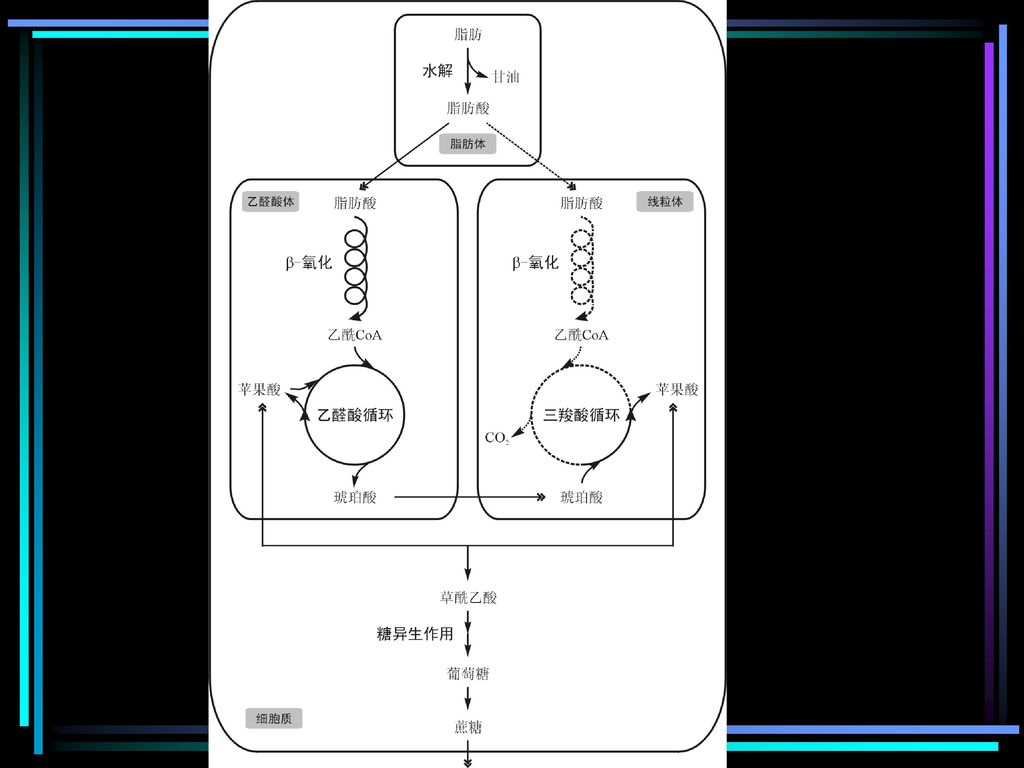
乙醛酸循环体(glyoxysome,简称乙醛酸体)
b-氧化作用的部位: 线粒体基质 乙醛酸循环体(glyoxysome,简称乙醛酸体) 油料作物种子萌发时
 油料作物种子萌发时.")
15
一、 脂肪酸的活化 脂肪酸的活化是指脂肪酸的羧基与CoA酯化成脂酰CoA的过程。反应如下:

16
脂肪酸的活化需要ATP的参与。每活化1分子脂肪酸,需要1分子ATP转化为AMP,即要消耗2个高能磷酸键。这可以折算成需要2分子ATP水解成ADP。
在体内,焦磷酸很快被磷酸酶水解,使得反应不可逆。

17
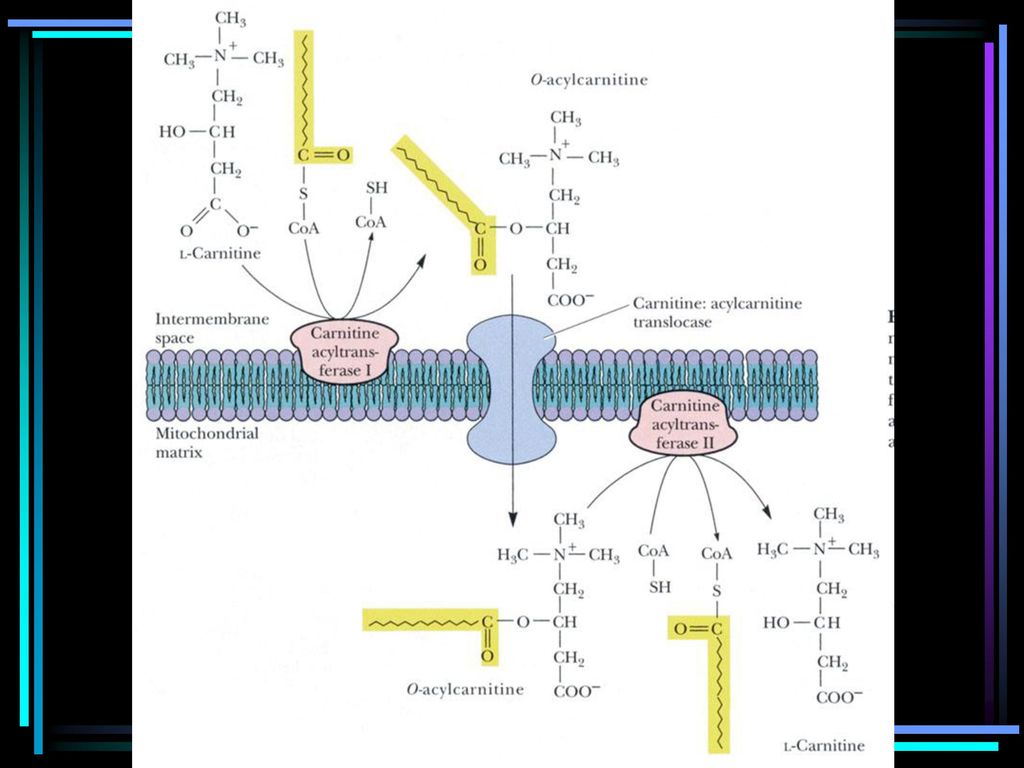
二、脂肪酸的转运 脂肪酸的 -氧化作用通常是在线粒体的基质中进行的,中、短链脂肪酸可直接穿过线粒体内膜,而长链脂肪酸需依靠肉碱(也叫肉毒碱,Carnitine)携带,以脂酰肉碱的形式跨越内膜而进入基质,故称肉碱转运。
携带,以脂酰肉碱的形式跨越内膜而进入基质,故称肉碱转运。")
18
肉碱(也叫肉毒碱,Carnitine)的结构如下:
的结构如下:")
19
肉毒碱是季胺类化合物,是一种人体必需的营养素,有着重要的生物学功能和临床应用价值。近年来肉毒碱在心脑血管疾病、消化疾病、儿童疾病的预防和治疗,以及血液透析病人的营养支持和运动医学等领域已得到广泛的研究和应用。

20
其中的肉碱脂酰转移酶Ⅰ和Ⅱ是一组同工酶。前者在线粒体外催化脂酰CoA上的脂酰基转移给肉碱,生成脂酰肉碱;后者则在线粒体内将运入的脂酰肉碱上的脂酰基重新转移至CoA,游离的肉碱被运回内膜外侧循环使用。

22
三、脂肪酸的 β-氧化 脂酰CoA进入线粒体后,经历多次b-氧化作用而逐步降解成多个二碳单位 —— 乙酰CoA。
1. 偶数碳链脂肪酸的氧化 脂酰CoA进入线粒体后,经历多次b-氧化作用而逐步降解成多个二碳单位 —— 乙酰CoA。 每次b-氧化作用包括四个步骤:

23
a. 氧化( 脂酰辅酶A脱氢酶)
")
24
b. 水合 (烯脂酰CoA水合酶)
")
25
c. 氧化 (羟脂酰CoA脱氢酶)
")
26
d. 断裂/硫解 (-酮脂酰CoA硫解酶)
")
27
乙酰CoA

28
对于长链脂肪酸,需要经过多次b-氧化作用,每次降解下一个二碳单位,直至成为二碳(当脂肪酸含偶数碳时)或三碳(当脂肪酸含奇数碳时)的脂酰CoA。
或三碳(当脂肪酸含奇数碳时)的脂酰CoA。")
29
下图是软脂酸(棕榈酸 C15H31COOH)的b-氧化过程,它需经历七轮b-氧化作用而生成8分子乙酰CoA。
的b-氧化过程,它需经历七轮b-氧化作用而生成8分子乙酰CoA。")
30
对于偶数碳饱和脂肪酸,b-氧化过程的化学计量:
脂肪酸在b-氧化作用前的活化作用需消耗能量,即1分子ATP转变成了AMP,消耗了2个高能磷酸键,相当于2分子ATP。 在b-氧化过程中,每进行一轮,使1分子FAD还原成FADH2、1分子NAD+还原成NADH,两者经呼吸链可分别生成2分子和3分子ATP,因此每轮b-氧化作用可生成5分子ATP。 b-氧化作用的产物乙酰CoA可通过三羧酸循环而彻底氧化成CO2和水,同时每分子乙酰CoA可生成12分子ATP。

31
1分子软脂酸彻底氧化 生成ATP的分子数 一次活化作用 -2 7轮b-氧化作用 +5×7 = +35 8分子乙酰CoA的氧化 +12×8 = +96 总 计 +129 在油料种子萌发时乙醛酸体中通过b-氧化产生的乙酰CoA一般不用作产能形成ATP,而是通过乙醛酸循环(见后)转变成琥珀酸,再经糖的异生作用转化成糖。
转变成琥珀酸,再经糖的异生作用转化成糖。")
32
丙酰CoA的代谢在动物体内依照如下图所示的途径进行,先进行羧化,然后经过两次异构化,形成琥珀酰CoA。
2. 奇数碳链脂肪酸的氧化 大多数脂肪酸含偶数碳原子,它们通过b-氧化可全部转变成乙酰CoA,但一些植物和海洋生物能合成奇数碳脂肪酸,它们在最后一轮b-氧化作用后,产生丙酰CoA。 丙酰CoA的代谢在动物体内依照如下图所示的途径进行,先进行羧化,然后经过两次异构化,形成琥珀酰CoA。

33
油酰CoA 的氧化

34
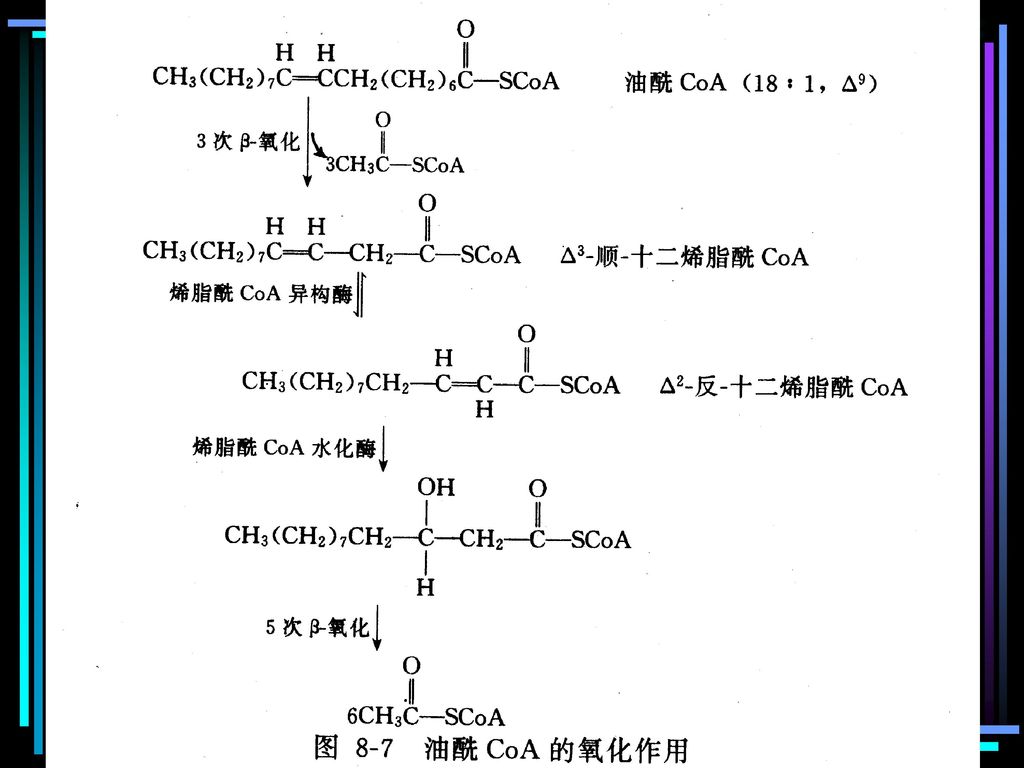
3.不饱和脂肪酸的氧化 生物体中的不饱和脂肪酸的双键都是顺式构型,而且位置也相当有规律 —— 第一个双键都是在C9和C10之间(写作D9),以后每隔三个碳原子出现一个。例如,亚油酸18:2D9,12;-亚油酸18:3D9,12,15。 不饱和脂肪酸的氧化与饱和脂肪酸基本相同,只是某些步骤还需其它酶的参与,现以油酸为例加以说明。

36
它经历了三轮b-氧化作用后,产物在b,g位有一顺式双键,因此接下来的反应不是脱氢,而是双键的异构化,生成反式的a,b双键,然后b-氧化作用继续正常进行。因此油酸的氧化与相同碳的饱和脂肪酸(硬脂酸)相比,只是以一次双键异构化反应取代了一次脱氢反应,所以少产生一分子FADH2。 不仅是单不饱和脂肪酸,所有的多不饱和脂肪酸的前四轮b-氧化作用都与油酸相类同,都在第四轮时需要一种异构酶的参与。

37
四、脂肪酸的α-氧化途径 脂肪酸在一些酶的催化下,在a-碳原子上发生氧化作用,分解出一个一碳单位CO2,生成缩短了一个碳原子的脂肪酸。这种氧化作用称为脂肪酸的a-氧化作用。 a-氧化作用是1956年由P.K.Stumpf首先在植物种子和叶片中发现的,后来在动物脑和肝细胞中也发现了脂肪酸的这种氧化作用。

38
该途径以游离脂肪酸作为底物,在a-碳原子上发生羟化(-OH)或氢过氧化(-OOH),然后进一步氧化脱羧,其可能的机理下图所示。
或氢过氧化(-OOH),然后进一步氧化脱羧,其可能的机理下图所示。")
39
a-氧化作用对于生物体内 奇数碳脂肪酸的形成; 含甲基的支链脂肪酸的降解; 过长的脂肪酸(如C22、C24)的降解 起着重要的作用 哺乳动物将绿色蔬菜中植醇降解就是通过这种途径而实现的
的降解 起着重要的作用 哺乳动物将绿色蔬菜中植醇降解就是通过这种途径而实现的")
40
五、脂肪酸的-氧化途径 脂肪酸的-氧化是指脂肪酸的末端(-端)甲基发生氧化,先转变成羟甲基,继而再氧化成羧基,从而形成a, -二羧酸的过程。
甲基发生氧化,先转变成羟甲基,继而再氧化成羧基,从而形成a, -二羧酸的过程。")
41
生成的,-二羧酸可从两端进行β-氧化作用而降解。

42
动物体内的十二碳以下的脂肪酸常常通过w-氧化途径进行降解。
植物体内的在w-端具有含氧基团(羟基、醛基或羧基)的脂肪酸大多也是通过w-氧化作用生成的,这些脂肪酸常常是角质层或细胞壁的组成成分。 一些需氧微生物能将烃或脂肪酸迅速降解成水溶性产物,这种降解过程首先要进行w-氧化作用,生成二羧基脂肪酸后再通过b-氧化作用降解,如海洋中的某些浮游细菌可降解海面上的浮油,其氧化速率可高达0.5克/天/平方米。
的脂肪酸大多也是通过w-氧化作用生成的,这些脂肪酸常常是角质层或细胞壁的组成成分。 一些需氧微生物能将烃或脂肪酸迅速降解成水溶性产物,这种降解过程首先要进行w-氧化作用,生成二羧基脂肪酸后再通过b-氧化作用降解,如海洋中的某些浮游细菌可降解海面上的浮油,其氧化速率可高达0.5克/天/平方米。")
43
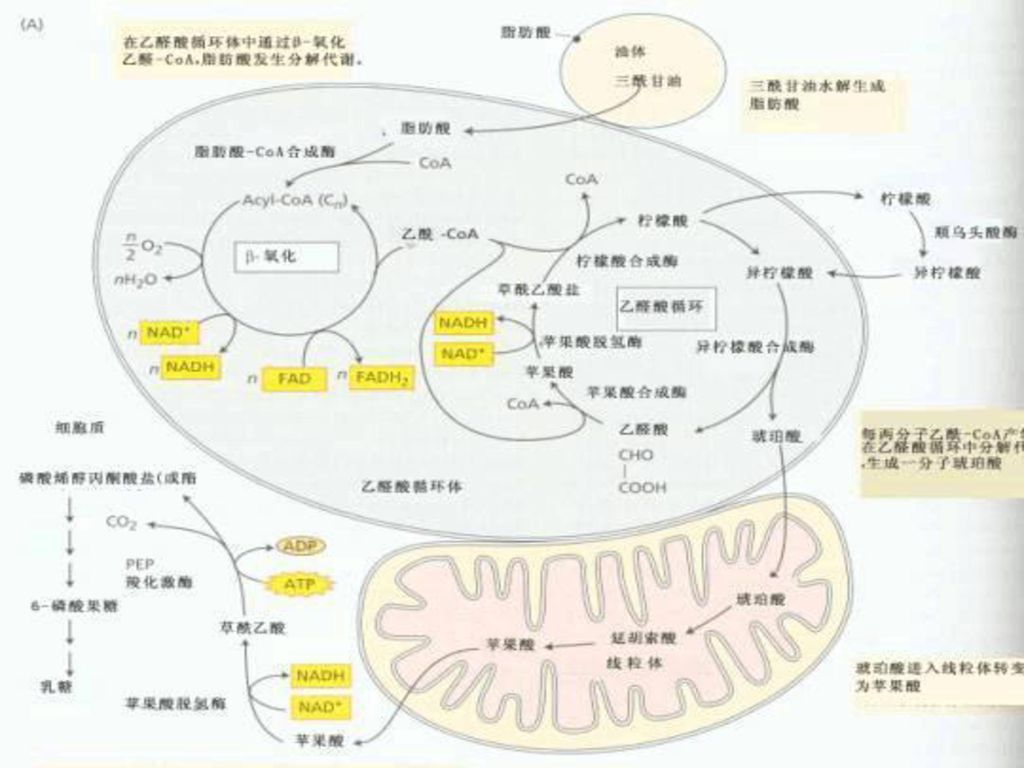
六. 乙醛酸循环(Glyoxylate cycle)
乙醛酸循环不存在于动物及高等植物的营养器官内,它存在于一些细菌、藻类和油料植物的种子的乙醛酸体中。 油料植物的种子中主要的贮藏物质是脂肪,在种子萌发时乙醛酸体大量出现,由于它含有脂肪分解和乙醛酸循环的整套酶系,因此可以将脂肪分解,并将2分子乙酰CoA合成1分子四碳化合物琥珀酸。

44
微生物、植物细胞内脂肪酸氧化分解为乙酰CoA之后,在乙醛酸体(glyoxysome)内生成琥珀酸、乙醛酸和苹果酸;琥珀酸可用于糖的合成,该过程称为乙醛酸循环(glyoxylic acid cycle,GAC)。
内生成琥珀酸、乙醛酸和苹果酸;琥珀酸可用于糖的合成,该过程称为乙醛酸循环(glyoxylic acid cycle,GAC)。")
45
动物和人类细胞中没有乙醛酸体,无法将脂肪酸转变为糖。
植物和微生物有乙醛酸体。油料植物种子(花生、油菜、棉籽等)萌发时存在着能够将脂肪酸转化为糖的乙醛酸循环。 水稻叶片中也分离出了乙醛酸循环中的两个关键酶——异柠檬酸裂解酶和苹果酸合成酶。
萌发时存在着能够将脂肪酸转化为糖的乙醛酸循环。 水稻叶片中也分离出了乙醛酸循环中的两个关键酶——异柠檬酸裂解酶和苹果酸合成酶。")
46
琥珀酸可异生成糖并以蔗糖的形式运至种苗的其它组织供给它们生长所需的能源和碳源;而当种子萌发终止、贮脂耗尽,同时叶片能进行光合作用时,植物的能源和碳源可以由太阳光和CO2获得时,乙醛酸体的数量迅速下降以至完全消失。 对于一些细菌和藻类,乙醛酸循环使它们能以乙酸盐为能源和碳源生长。

47
1. 乙醛酸循环的过程

48
乙醛酸循环的净结果是把两分子乙酰CoA转变成一分子琥珀酸。其总反应为:
1. 乙醛酸循环的过程 乙醛酸循环的净结果是把两分子乙酰CoA转变成一分子琥珀酸。其总反应为:

50
乙醛酸循环与三羧酸循环相比,可以看成是三羧酸循环的一个支路,它在异柠檬酸处分支,绕过了三羧酸循环的两步脱羧反应,因而不发生氧化降解。
1. 乙醛酸循环的过程 乙醛酸循环与三羧酸循环相比,可以看成是三羧酸循环的一个支路,它在异柠檬酸处分支,绕过了三羧酸循环的两步脱羧反应,因而不发生氧化降解。 参与乙醛酸循环的酶除了异柠檬酸裂解酶和苹果酸合酶外,其余的酶都与三羧酸循环的酶相同。 异柠檬酸裂解酶和苹果酸合酶是乙醛酸循环的关键酶。

52
由乙醛酸循环转变成的琥珀酸,需要在线粒体中通过三羧酸循环的部分反应转化为苹果酸,然后进入细胞质,沿糖异生途径转变成糖类。
乙醛酸循环中有苹果酸中间体,它也可以到细胞质中异生成糖,但它需要及时回补,以保证循环的正常进行,这仍来自循环的产物琥珀酸在线粒体中的转变。

53
八. 酮体代谢 由脂肪酸的-氧化及其它代谢所产生的乙酰CoA,在一般的细胞中可进入三羧酸循环进行氧化分解;但在动物的肝脏、肾脏、脑等组织中,尤其在饥饿、禁食、糖尿病等情形下,乙酰CoA还有另一条代谢去路,最终生成乙酰乙酸、 -羟丁酸和丙酮,这三种产物统称为酮体(ketone bodies)。
。 .")
54
(1) 两分子乙酰CoA缩合成乙酰乙酰CoA,反应由硫解酶催化。此外,脂肪酸β-氧化作用的最后一轮也能产生乙酰乙酰CoA。
1. 酮体的合成 八. 酮体代谢 (1) 两分子乙酰CoA缩合成乙酰乙酰CoA,反应由硫解酶催化。此外,脂肪酸β-氧化作用的最后一轮也能产生乙酰乙酰CoA。
 两分子乙酰CoA缩合成乙酰乙酰CoA,反应由硫解酶催化。此外,脂肪酸β-氧化作用的最后一轮也能产生乙酰乙酰CoA。")
55
(2) 又一分子乙酰CoA与乙酰乙酰CoA缩合,生成-羟--甲基戊二酸单酰CoA(HMG-CoA),反应由HMG-CoA合成酶催化。
1. 酮体的合成 八. 酮体代谢 (2) 又一分子乙酰CoA与乙酰乙酰CoA缩合,生成-羟--甲基戊二酸单酰CoA(HMG-CoA),反应由HMG-CoA合成酶催化。 3-Hydroxy-3methylglutaryl-CoA (HMG-CoA)
 又一分子乙酰CoA与乙酰乙酰CoA缩合,生成-羟--甲基戊二酸单酰CoA(HMG-CoA),反应由HMG-CoA合成酶催化。 3-Hydroxy-3methylglutaryl-CoA (HMG-CoA)")
56
(3) MG-CoA分解成乙酰乙酸和乙酰CoA,反应由HMG-CoA裂解酶催化。
1. 酮体的合成 八. 酮体代谢 (3) MG-CoA分解成乙酰乙酸和乙酰CoA,反应由HMG-CoA裂解酶催化。
 MG-CoA分解成乙酰乙酸和乙酰CoA,反应由HMG-CoA裂解酶催化。")
57
1. 酮体的合成 八. 酮体代谢 (4) 生成的乙酰乙酸一部分可还原成-羟丁酸,反应由-羟丁酸脱氢酶催化;也有极少一部分可脱羧形成丙酮,反应可自发进行,也可由乙酰乙酸脱羧酶催化。
 生成的乙酰乙酸一部分可还原成-羟丁酸,反应由-羟丁酸脱氢酶催化;也有极少一部分可脱羧形成丙酮,反应可自发进行,也可由乙酰乙酸脱羧酶催化。")
58
八. 酮体代谢 2. 酮体的分解

59
八. 酮体代谢 2. 酮体的分解

60
第四节 脂肪酸的β-氧化作用 1904年德国Knoop: 1-5C 苯脂酸 喂狗
第四节 脂肪酸的β-氧化作用 1904年德国Knoop: 1-5C 苯脂酸 喂狗 马尿酸 苯乙尿酸 1941年德国Schoeuheimer: 氘标记硬脂酸 喂小鼠

61
(一)饱和脂肪酸的氧化 1. 脂肪酸的活化 内质网,线粒体外膜 2. 脂酰CoA的转移 肉碱:β羟γ三甲基氨基丁酸 carnitine
1. 脂肪酸的活化 内质网,线粒体外膜 Mg2+ 脂酰CoA合成酶 RCOOH + HS-CoA + ATP RCO SCoA + AMP + PPi 2. 脂酰CoA的转移 肉碱:β羟γ三甲基氨基丁酸 carnitine CH3 | CH3-N+-CH2-CH-CH2-COOH | | CH OH

63
脂酰CoA + 肉碱 CoASH + 脂酰肉碱 肉碱脂酰转移酶I: 限速酶 肉碱脂酰转移酶II: 脂肪酸 脂酰 SCoA 肉碱
线粒体外 线粒体内膜 线粒体内 脂肪酸 脂酰 SCoA 肉碱 脂酰 SCoA 酶I 酶II 脂酰 肉碱 CoASH CoASH 肉碱脂酰转移酶I: 限速酶 肉碱脂酰转移酶II: 肉碱:肝 赖氨酸; 肾

64
3. 脂肪酸的β 氧化 β-oxidation 线粒体 继续β氧化 脂酰CoA 脱氢酶 烯酰CoA β脂酰CoA 水合酶 硫解酶
RCH2-CH2-CH2-CO SCoA 脂酰CoA 脱氢酶 FAD FADH2 RCH2CH=CH-CO SCoA RCH2-CO ~ SCoA + CH3-CO ~ SCoA β脂酰CoA 硫解酶 CoASH H2O 烯酰CoA 水合酶 β羟脂酰 CoA脱氢酶 NAD+ NADH + H+ RCH2-CH-CH2-CO SCoA | OH RCH2-C-CH2-CO SCoA || O

66
4.乙酰CoA的彻底氧化 5. 能量的生成 软脂酸 CH3-(CH2)14-COOH + 23O2 16CO2 + 16H2O kcal/mol 活化: ATP 7次β氧化: 5 X 7 = 35 ATP 129 ATP 8个乙酰CoA:12 X 8 = 96 ATP

67
1 mol 高能磷酸键: 7.3 kcal 7.3 kcal × 129 = 940 kcal 940
能量利用率 = ×100 % = 40.2 % 2340 软脂酸 葡萄糖 1 mol 129 mol ATP mol ATP 100 g 50.4 mol ATP mol ATP 能量利用率 40.2 % % 调节:脂酰肉碱抑制丙酮酸脱氢酶系的活性

68
丙酰CoA CH3-CH2-CO SCoA 琥珀酰CoA
(二)奇数碳原子脂肪酸的氧化: 丙酰CoA CH3-CH2-CO SCoA 琥珀酰CoA (三)不饱和脂肪酸的氧化 天然不饱和脂肪酸多为顺式 异构酶
奇数碳原子脂肪酸的氧化: 丙酰CoA CH3-CH2-CO SCoA 琥珀酰CoA. (三)不饱和脂肪酸的氧化. 天然不饱和脂肪酸多为顺式. 异构酶.")
69
三、酮体的生成和利用 酮体 ketone bodies: 肝合成 血: < 3 mg % ( 0.3 mmol/L)
乙酰乙酸 CH3-C-CH2-COOH % || O β羟丁酸 CH3-CH-CH2-COOH % | OH 丙酮 CH3-C-CH 少量 || O

70
(一)酮体的生成 肝 线粒体 HMG辅酶A合酶:限速酶 β酮硫解酶 CoASH β羟丁 HMG辅酶 酸脱氢酶 A合酶 HMG辅酶A 裂解酶
(一)酮体的生成 肝 线粒体 2 CH3-CO SCoA HOOC-CH2-CH-CH3 | OH CH3-C-CH3 || O β酮硫解酶 CoASH β羟丁 酸脱氢酶 NADH +H+ NAD+ CH3-CO-CH2-CO SCoA CO2 CH3-CO ~ SCoA HMG辅酶 A合酶 CoASH HMG辅酶A 裂解酶 CH3-CO ~ SCoA CH3 | HOOC-CH2-C-CH2-CO SCoA OH HOOC-CH2-C-CH3 || O HMG辅酶A合酶:限速酶
酮体的生成 肝 线粒体. 2 CH3-CO SCoA. HOOC-CH2-CH-CH3. | OH. CH3-C-CH3. || O. β酮硫解酶. CoASH. β羟丁. 酸脱氢酶. NADH +H+ NAD+ CH3-CO-CH2-CO SCoA. CO2. CH3-CO ~ SCoA. HMG辅酶. A合酶. CoASH. HMG辅酶A. 裂解酶. CH3-CO ~ SCoA. CH3. | HOOC-CH2-C-CH2-CO SCoA. OH. HOOC-CH2-C-CH3. || O. HMG辅酶A合酶:限速酶.")
71
(二)酮体的利用 心肌,骨骼肌,大脑等 琥珀酰CoA 乙酰乙酰 β酮脂酰 CoA合成酶 CoA转移酶 琥珀酸
(二)酮体的利用 心肌,骨骼肌,大脑等 CH3-CH(OH)-CH2-COOH ATP + CoASH CH3-CO-CH2-COOH 琥珀酰CoA 乙酰乙酰 CoA合成酶 β酮脂酰 CoA转移酶 AMP + PPi 琥珀酸 CH3-CO-CH2-CO SCoA CoASH 2 CH3-CO SCoA TCA
酮体的利用 心肌,骨骼肌,大脑等. CH3-CH(OH)-CH2-COOH. ATP + CoASH. CH3-CO-CH2-COOH. 琥珀酰CoA. 乙酰乙酰. CoA合成酶. β酮脂酰. CoA转移酶. AMP + PPi. 琥珀酸. CH3-CO-CH2-CO SCoA. CoASH. 2 CH3-CO SCoA. TCA.")
72
(三)酮体生成的意义 能源物质:肝外组织 大脑、肌肉 (四)影响因素 1。短期饥饿: 生理: 2。长期饥饿: 糖尿病 酮症 ketosis: 酮血症,酮尿症(> 70 mg %),酮症酸中毒
酮体生成的意义 能源物质:肝外组织 大脑、肌肉 (四)影响因素 1。短期饥饿: 生理: 2。长期饥饿: 糖尿病 酮症 ketosis: 酮血症,酮尿症(> 70 mg %),酮症酸中毒")
73
四、甘油代谢 glycerol 肝、肾、肠 CH2-OH | CH-OH 1. CH2-OH | CH-OH CH2-O- P 甘油磷酸激酶
ATP ADP 2. CH2-OH | CH-OH CH2-O- P + NAD+ CH2-OH | C=O CH2-O- P + NADH + H+ 3磷酸甘油脱氢酶 TCA 糖

74
五. 脂肪酸的合成 部位: 肝, 脑, 肺, 乳腺, 脂肪组织 胞液 (一)饱和脂肪酸的合成 原料: CH3CO~ SCoA 1. 运出线粒体: CH3CO~SCoA 柠檬酸----丙酮酸穿梭 citrate pyruvate cycle

75
丙酮酸 苹果酸 线粒体 胞液 柠檬酸 柠檬酸 草酰乙酸 草酰乙酸 苹果酸 苹果酸 丙酮酸 丙酮酸 苹果酸酶 CoASH CoASH
载体 CoASH ATP ADP + Pi 草酰乙酸 草酰乙酸 CH3CO~SCoA CH3CO~SCoA NADH + H+ NAD+ 苹果酸脱氢酶 NADH + H+ NAD+ 丙酮酸羧化酶 ATP ADP + Pi CO2 苹果酸 载体 苹果酸 NADP+ NADPH + H+ 苹果酸酶 CO2 丙酮酸 载体 丙酮酸 丙酮酸 苹果酸 NADP+ NADPH + H+ 苹果酸酶 CO2

76
2. 丙二酸单酰CoA的合成 乙酰CoA羧化酶 生物素 解聚: 无活性 乙酰CoA羧化酶: 限速酶 多聚体: 有活性 柠檬酸: 促进
CH3-CO SCoA + HCO3- + ATP HOOC-CH2-CO SCoA + ADP + Pi 解聚: 无活性 多聚体: 有活性 乙酰CoA羧化酶: 限速酶 柠檬酸: 促进 长链脂肪酰CoA: 抑制

77
3. 软脂酸的合成 大肠杆菌 脂肪酸合成酶复合体: 脂肪酸合成酶复合体 脂酰基载体蛋白 acyl carrier protein, ACP
3. 软脂酸的合成 大肠杆菌 脂肪酸合成酶复合体 14NADPH + H+ 14NADP+ CH3CO SCoA + 7 HOOC-CH2-CO SCoA CH3-(CH2)14-COOH + 7CO2 + 8CoASH + 6H2O 脂肪酸合成酶复合体: 脂酰基载体蛋白 acyl carrier protein, ACP 辅基: 4`-磷酸泛酰氨基乙硫醇 O CH O O || | || || Ser36—O-P-O-CH2-C-CHOH-C-NH-CH2-CH2-C-NHCH2-CH2-SH | | OH CH 泛酸 氨基乙硫醇
14-COOH + 7CO2 + 8CoASH + 6H2O. 脂肪酸合成酶复合体: 脂酰基载体蛋白 acyl carrier protein, ACP. 辅基: 4`-磷酸泛酰氨基乙硫醇. O CH3 O O. || | || || Ser36—O-P-O-CH2-C-CHOH-C-NH-CH2-CH2-C-NHCH2-CH2-SH. | | OH CH3 泛酸 氨基乙硫醇.")
78
1. 乙酰基转移酶 2. 丙二酰基转移酶 3. β酮脂酰合酶 4. β酮脂酰还原酶 5. β羟脂酰脱水酶 6. 烯酰还原酶 7. 硫脂酶

79
合成过程: 转移 缩合 还原 硫解 加氢 脱水 再加氢

80
(二)脂肪酸碳链的延长 1. 内质网酶体系 缩合 加氢 加氢 脱水 O O || ||
1. 内质网酶体系 O O || || R-CH2-C~SCoA HOOC-CH2-C~SCoA 缩合 O || R-CH2-CH2-CH2-C~SCoA O O || || R-CH2-C-CH2-C~SCoA 加氢 NADPH + H+ NADP+ 加氢 NADPH + H+ NADP+ 脱水 H2O O || R-CH2-CH=CH-C~SCoA OH O | || R-CH2-CH-CH2-C~SCoA

81
2. 线粒体酶体系 乙酰CoA供给二碳单位 (三) 不饱和脂肪酸的合成 内质网: 去饱和酶 硬脂酸 油酸( 18:1, ∆9 ) 软脂酸 软油酸 (16:1, ∆9 ) 亚油酸(18:2, ∆9,12) 亚麻酸(18:3, ∆9,12,15) 花生四烯酸(20:4, ∆5,8,11,14) 必需脂肪酸 Essential fatty acid
 亚麻酸(18:3, ∆9,12,15) 花生四烯酸(20:4, ∆5,8,11,14) 必需脂肪酸. Essential. fatty acid.")
82
(四) 脂肪酸合成的调节 限速酶: 乙酰CoA羧化酶 1. 代谢物的调节 饥饿,禁食 脂肪动员 高脂膳食 脂酰CoA 乙酰CoA羧化酶 高糖膳食 ATP 异柠檬酸脱氢酶 柠檬酸堆积 乙酰CoA羧化酶

83
2. 激素的调节 ①胰岛素: 诱导乙酰CoA羧化酶 , 脂肪酸合酶, 柠檬酸裂解酶 脂肪酸合成 ②胰高血糖素, 生长素, 肾上腺素 乙酰CoA羧化酶 P , 活性 脂肪酸合成

84
3. 脂肪酸的合成与分解 合成: 丙二酰CoA 抑制分解: 肉碱脂酰转移酶

85
六、甘油三脂的合成 triglyceride
1. 肝细胞 O ‖ O CH2-O-C-R1 ‖ | R2-C-O-CH | CH2- P CH2-OH | CH-OH ① CH2-OH | CH-OH CH2-O- P 甘油磷酸激酶 ATP ADP 2RCO~SCoA 2CoASH 脂酰转移酶 O ‖ O CH2-O-C-R1 ‖ | R2-C-O-CH | CH2- OH O ‖ O CH2-O-C-R1 ‖ | R2-C-O-CH O | || CH2-O-C-R3 磷酸酶 Pi RCO~SCoA CoASH 脂酰转移酶

86
2. 脂肪组织 ② 甘油三脂 甘油三脂 CH2-OH | C=O CH2-O- P CH2-OH | CH-OH CH2-O- P

87
3. 肠粘膜细胞 脂酰转移酶 O ‖ O CH2-O-C-R1 ‖ | R2-C-O-CH O | || CH2-O-C-R3
‖ | R2-C-O-CH O | || CH2-O-C-R3 O CH2-OH ‖ | R2-C-O-CH | CH2-OH 2RCO~SCoA 2CoASH 脂酰转移酶

88
O ‖ O CH2-O-C-R1 ‖ | R2-C-O-CH | CH2- P -X 甘油磷脂

89
第四节 磷脂的代谢 phospholipid 构成细胞膜 脂类运输 卵磷脂 (ecithin)
第四节 磷脂的代谢 phospholipid 构成细胞膜 脂类运输 卵磷脂 (ecithin) 磷脂酰胆碱 (phosphstidyl choline, PC) 脑磷脂 (cephalin) 磷脂酰乙醇胺 (phosphatidyl ethanolamine, PE) 甘油磷脂 磷脂酰丝氨酸 (phosphatidyl serine, PS) 磷脂酰肌醇 (phosphatidyl inositol, PI) 磷脂酰甘油 (phosphatidyl glycerol, PG) 心磷脂 (cardiolipin)
 磷脂酰胆碱 (phosphstidyl choline, PC) 脑磷脂 (cephalin) 磷脂酰乙醇胺 (phosphatidyl ethanolamine, PE) 甘油磷脂. 磷脂酰丝氨酸 (phosphatidyl serine, PS) 磷脂酰肌醇 (phosphatidyl inositol, PI) 磷脂酰甘油 (phosphatidyl glycerol, PG) 心磷脂 (cardiolipin)")
90
甘油磷脂 O ‖ O CH2-O-C-R1 ‖ | R2-C-O-CH | CH2- P -X
‖ | R2-C-O-CH | CH2- P -X 甘油磷脂 PC: X= HO-CH2-CH2-N+-(CH3)3
3.")
91
O ‖ O CH2-O-C-R1 ‖ | R2-C-O-CH | CH2- P -X 甘油磷脂 PE: X=HO-CH2-CH2-NH2

92
甘油磷脂 O ‖ O CH2-O-C-R1 ‖ | R2-C-O-CH | CH2- P -X PS: X= HO-CH2-CH-COOH
‖ | R2-C-O-CH | CH2- P -X 甘油磷脂 PS: X= HO-CH2-CH-COOH | NH2

93
O ‖ O CH2-O-C-R1 ‖ | R2-C-O-CH | CH2- P -X 甘油磷脂 6 5 PI: X = 4 1 2 3

94
O ‖ O CH2-O-C-R1 ‖ | R2-C-O-CH | CH2- P -X 甘油磷脂 CH2-OH | PG: X = CH-OH

95
甘油磷脂 心磷脂:X = PG O ‖ O CH2-O-C-R1 ‖ | R2-C-O-CH | CH2- P -X O O ‖ ||
‖ | R2-C-O-CH | CH2- P -X 甘油磷脂 心磷脂:X = PG O O ‖ || O CH2-O-C-R CH2 – O-C - R3 ‖ | | R2-C-O-CH O OH O CH – O-C - R4 | || | || | || CH2-O-P-O-CH2-CH-CH2-O-P-O- CH O | | OH OH

96
一、甘油磷脂的合成 (一)卵磷脂 脑磷脂 磷脂酰丝氨酸 3 (S腺苷蛋氨酸) COOH | CHNH2 CH2OH +
(一)卵磷脂 脑磷脂 磷脂酰丝氨酸 COOH | CHNH2 CH2OH + CH2-N(CH3)3 | CH2OH CH2NH2 | CH2OH 3 (S腺苷蛋氨酸) CO2 ATP ADP CH3-S-AR | CH2 CH-NH2 COOH + CH2-N(CH3)3 | CH2O- P CH2NH2 | CH2O- P CTP PPi CDP-乙醇胺 CDP-胆碱
卵磷脂 脑磷脂 磷脂酰丝氨酸. COOH. | CHNH2. CH2OH. + CH2-N(CH3)3. | CH2OH. CH2NH2. | CH2OH. 3 (S腺苷蛋氨酸) CO2. ATP. ADP. CH3-S-AR. | CH2. CH-NH2. COOH. + CH2-N(CH3)3. | CH2O- P. CH2NH2. | CH2O- P. CTP. PPi. CDP-乙醇胺. CDP-胆碱.")
97
CDP-乙醇胺 CMP 磷酸乙醇胺 转移酶 CDP-胆碱 CMP 磷酸胆碱 转移酶 3 (S腺苷蛋氨酸) O ‖ O CH2-O-C-R1
‖ | R2-C-O-CH | CH2- P O ‖ O CH2-O-C-R1 ‖ | R2-C-O-CH | CH2- OH 磷酸酶 Pi CDP-乙醇胺 CMP 磷酸乙醇胺 转移酶 CDP-胆碱 CMP 磷酸胆碱 转移酶 O ‖ O CH2-O-C-R1 ‖ | R2-C-O-CH | CH2- P - 胆碱 O ‖ O CH2-O-C-R1 ‖ | R2-C-O-CH | CH2- P - 乙醇胺 3 (S腺苷蛋氨酸)
")
98
丝氨酸 乙醇胺 (二)磷脂酰肌醇 心磷脂 CTP PPi CTP – 磷脂酸 胞苷酸转移酶 CDP-甘油二脂 O ‖
O CH2-O-C-R1 ‖ | R2-C-O-CH | CH2- P - 乙醇胺 O ‖ O CH2-O-C-R1 ‖ | R2-C-O-CH | CH2- P - 丝氨酸 丝氨酸 乙醇胺 (二)磷脂酰肌醇 心磷脂 O ‖ O CH2-O-C-R1 ‖ | R2-C-O-CH | CH2- P CTP PPi CTP – 磷脂酸 胞苷酸转移酶 CDP-甘油二脂
磷脂酰肌醇 心磷脂. O. ‖ O CH2-O-C-R1. ‖ | R2-C-O-CH. | CH2- P. CTP. PPi. CTP – 磷脂酸. 胞苷酸转移酶. CDP-甘油二脂.")
99
CDP-甘油二脂 3 P 甘油 CMP 磷脂酰甘油3磷酸 H2O Pi 磷脂酰甘油(PG)
")
100
CDP-甘油二脂 磷脂酰肌醇(PI) 心磷脂 磷脂酰甘油3磷酸 (二磷脂酰甘油) 磷脂酰甘油(PG) 肌醇 CMP 磷脂酰甘油 CMP
H2O Pi 磷脂酰甘油(PG)
")
101
PIP2 磷脂酰 肌醇激酶 PI 磷脂酶C DAG IP3 O ‖ ‖ O CH2-O-C-R1 O CH2-O-C-R1 ‖ | ‖ |
‖ | R2-C-O-CH O | || CH2-O-P-OH | PIP2 O ‖ O CH2-O-C-R1 ‖ | R2-C-O-CH O | || CH2-O-P-OH | 磷脂酰 肌醇激酶 2ATP 2ADP PI 磷脂酶C O ‖ O CH2-O-C-R1 ‖ | R2-C-O-CH | CH2- OH DAG P O IP3

102
二、甘油磷脂的分解 ① 小肠: 水解磷脂,帮助吸收 ② 细胞: 降解磷脂, 新陈代谢 ③产生第二信使: IP3, DAG 等
O ‖ O CH O C R1 ‖ | R C O CH O | || CH O P O X | OH A2 C D 磷脂酶A2: 溶血磷脂 ① 小肠: 水解磷脂,帮助吸收 ② 细胞: 降解磷脂, 新陈代谢 ③产生第二信使: IP3, DAG 等

103
第五节 胆固醇的代谢 cholesterol 甾体 胆固醇

104
一、胆固醇的合成 1. 合成原料 1940年 Bloch 同位素标记: CH3COOH 喂大鼠 胆固醇被标记 乙酰CoA 2. 合成部位
主要: 肝 ~ 80 % 其它: 小肠,肾上腺皮质, 卵巢, 睾丸等 胞液及内质网

105
3. 合成过程 胆固醇 C27 羊毛固醇 C30(环) HMGCoA 合酶 鲨烯 C30 HMGCoA 还原酶 2 CH3-CO SCoA
CoASH CH3-CO-CH2-CO SCoA 羊毛固醇 C30(环) CH3-CO~ SCoA HMGCoA 合酶 CoASH 鲨烯 C30 CH3 | HOOC-CH2-C-CH2-CO~SCoA OH 6X CH3 | CH3-C=CH-CH2-O- P - P HMGCoA 还原酶 2 NADPH + H+ 2 NADP+ CH3 | HOOC-CH2-C-CH2-CH2-OH OH
 CH3-CO~ SCoA. HMGCoA. 合酶. CoASH. 鲨烯 C30. CH3. | HOOC-CH2-C-CH2-CO~SCoA. OH. 6X. CH3. | CH3-C=CH-CH2-O- P - P. HMGCoA. 还原酶. 2 NADPH + H+ 2 NADP+ CH3. | HOOC-CH2-C-CH2-CH2-OH. OH.")
106
HMG CoA还原酶: 限速酶 ①代谢物 外源性胆固醇↑ 内源性胆固醇合成↓ 外源性胆固醇↓ 内源性胆固醇合成↑ ② 激素 胰岛素 胆固醇合成↑ 胰高血糖素, 皮质醇 胆固醇合成↓ 甲状腺素 胆固醇合成↑ 胆固醇转变为胆汁酸↑ 血胆固醇↓

107
lecithin-cholesterol acyl transferase, LCAT
二、胆固醇的转变 1. 血浆中: 卵磷脂胆固醇脂肪酰基转移酶 lecithin-cholesterol acyl transferase, LCAT 胆固醇 胆固醇酯 LCAT O ‖ O CH2-C-O-R1 ‖ | R2-C-O-CH | CH2- P - 胆碱 溶血卵磷脂

108
2. 组织中 脂酰CoA胆固醇脂酰转移酶 acyl CoA : choesterol acyl transferase, ACAT 3. 生物活性物质 肾上腺皮质: 肾上腺皮质激素 性腺:雌激素,雄激素 皮肤:VD3

109
第六节 血浆脂蛋白的代谢 lipoprotein 一、血脂 含量不很稳定 包括:甘油三脂,磷脂,总胆固醇,游离脂肪酸 二、血浆脂蛋白分类
第六节 血浆脂蛋白的代谢 lipoprotein 一、血脂 含量不很稳定 包括:甘油三脂,磷脂,总胆固醇,游离脂肪酸 二、血浆脂蛋白分类 1. 密度法: 乳糜微粒 chylomicron, CM 极低密度脂蛋白 very low density lipoprotein, VLDL

110
低密度脂蛋白 low density lipoprotein, LDL
高密度脂蛋白 high density lipoprotein, HDL 2.电泳法: CM β 脂蛋白 前β 脂蛋白 α 脂蛋白 LDL VLDL HDL CM

111
三、组成 载脂蛋白 甘油三脂 磷脂 总胆固醇 CM VLDL LDL HDL 2 % 10 % 20 % 45 % 88 55 8 10
2 % % % % 甘油三脂 磷脂 总胆固醇

112
载脂蛋白 apolipoprotein AI: 激活LCAT CII: 激活脂蛋白脂肪酶 B100:促进脂蛋白与细胞膜表面受体结合
VLDL LDL HDL CM 载脂蛋白 apolipoprotein AI,CII CII B AI, D AI: 激活LCAT CII: 激活脂蛋白脂肪酶 B100:促进脂蛋白与细胞膜表面受体结合 D: 促进HDL的胆固醇酯转移到LDL上

113
四、脂蛋白的结构 胆固醇酯 甘油三脂 载脂蛋白 胆固醇 磷脂

114
五、脂蛋白的代谢 1. 乳 糜微粒(CM) 肠粘膜细胞合成 淋巴,血液 全身组织: 肝 毛细血管壁脂蛋白脂肪酶 作用:运输外源性甘油三脂到全身各组织 饱食脂肪餐后 血CM :半衰期 10分钟 2小时内清除

115
2. 极低密度脂蛋白 VLDL 肝脏合成:磷脂,胆固醇,载脂蛋白 VLDL 运输到肝外组织 甘油三脂水解 肝外组织摄取
毛细血管壁脂蛋白脂肪酶 VLDL 运输到肝外组织 甘油三脂水解 肝外组织摄取 作用:运输内源性甘油三脂到机体各组织中

116
3. 低密度脂蛋白 LDL 合成场所:血液 胆固醇占48 % apoB100 作用:运输肝脏合成的胆固醇到肝外组织 4. 高密度脂蛋白 HDL 合成场所:肝脏 作用:收集血液中胆固醇及apo运回肝脏 *血液中游离脂肪酸的运输:与清蛋白结合

117
六、脂蛋白代谢异常 高脂蛋白血症

118
肝糖原 1磷酸葡萄糖 葡萄糖 6磷酸葡萄糖 戊糖旁路 甘油 3磷酸甘油 磷酸丙糖 NADPH + H+ 脂肪酸 丙酮酸 胆固醇 乙酰CoA β 羟丁酸 乙酰乙酸 草酰乙酸 TCA 丙酮

Similar presentations

生物生长发育和代谢所必需 的一类微量有机物质;( 2 )需量少,每日仅需 mg 或 g 级;( 3 )在体内不能合成或合成不足, 必需由食物供给( 4 )机体缺乏会导致物质谢障 碍,引起缺乏症。 分类:维生素一般习惯分为脂溶性和水溶性两大 类。其中脂溶性维生素在体内可直接参与代谢的.>")
 和二十二碳六 烯酸 (DHA) 等。>")

>")
 磷脂質 游離脂肪酸>")






蛋白质的降解: 外源蛋白的消化 内源性蛋白的选择性降解 (2)氨基酸的分解代谢:>")

是参与生物生长发育和代谢所必需的一类微量有机物质。这类物质由于体内不能合成或者合成量不足,所以必需由食物供给。已知绝大多数维生素作为酶的辅酶或辅基的组成成分,在物质代谢中起重要作用。机体缺乏维生素时,物质代谢发生障碍,引起维生素缺乏症。 脂溶性维生素和水溶性维生素.>")
.>")
脂类及脂蛋白测定 ——董雷鸣.>")


 電子傳遞鏈 傳導概論>")
