第36章、RNA合成 36.1 基因转录需要DNA依赖性的RNA聚合酶 36.2 RNA合成涉及三个过程:起始,延伸和终止 36.5 核酶是个位置特异的核酸内切酶 36.6 基因转录调控-操纵子模型
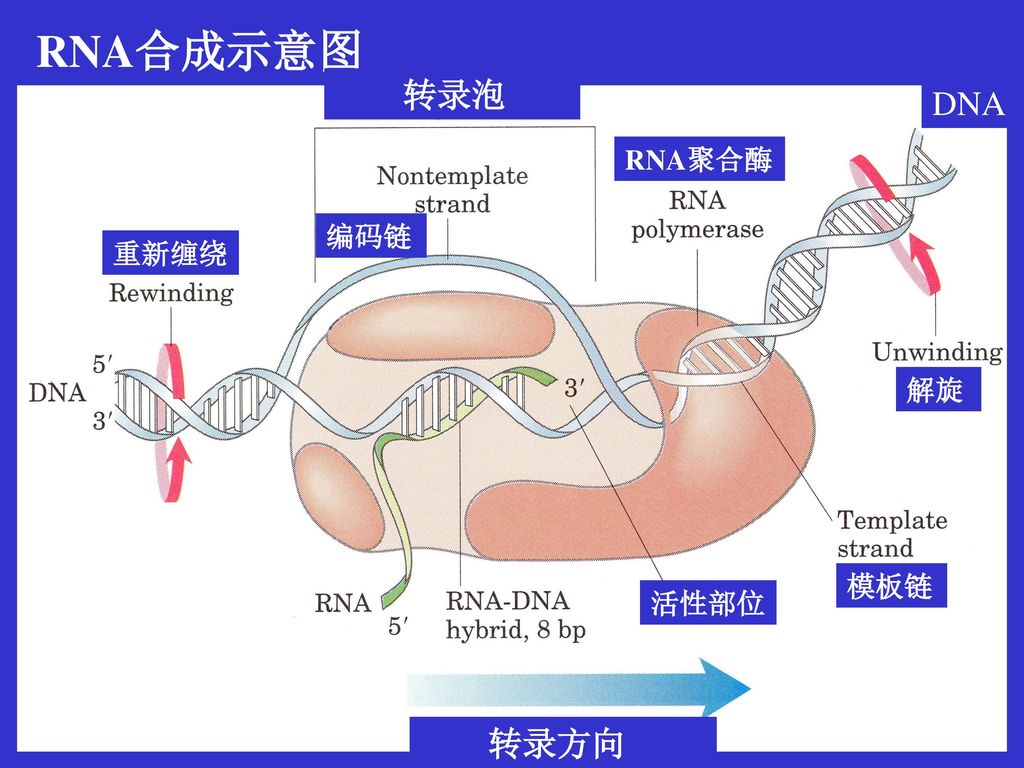
RNA合成示意图 转录泡 DNA RNA聚合酶 编码链 重新缠绕 解旋 模板链 活性部位 转录方向
36.1 基因转录需要DNA依赖性的RNA聚合酶 大肠杆菌RNA聚合酶的核心酶是由5个蛋白亚基组成的,分别被命名为,ˊ,(2个),和亚基。其中亚基是催化亚基,核心酶的分子量大约是350000。 大肠杆菌RNA聚合酶全酶还含有第六个亚基,称之亚基(分子量为70000),与核心的RNA聚合酶瞬时结合,其功能是识别模板上的启动子,使RNA聚合酶与启动子结合。一旦延伸开始亚基就脱离聚合酶。
真核生物存在着三种不同的RNA聚合酶:RNA聚合酶I(RNA pol I)、RNA聚合酶II(RNA pol II)和RNA聚合酶III(RNA pol III)。每一种聚合酶都含有很多亚基,它们承担的任务各不相同。
36.2 RNA合成包括三个阶段:起始、延伸和终止
(a)转录在启动子调控下起始 当E.coli RNA聚合酶结合到模板上的启动子后,就开始了RNA的合成。细菌启动子要行使其功能需要两个高度保守DNA序列,一个序列区是处于开始转录的第一个核苷酸的5ˊ端之前(习惯称之上游)的-35区,(上游核苷酸编号为“-”),提供RNA聚合酶识别信号。另一个保守区称为-10区,即转录起始点上游的10个核苷酸,提供DNA双链解旋信号。 RNA聚合酶首先沿着DNA滑动直至遇到-35区,形成一个起始的蛋白-DNA复合体(包括亚基),称为封闭性复合体。这个复合体向3ˊ方向移动到-10区,启动DNA双链解旋,将模板上的转录起始点暴露出来,此时RNA聚合酶和DNA形成的复合体称为开放性复合体,它构成了活性的转录单元。-10区是个富含A-T的区,由于A-T碱基对之间的氢键弱,所以容易使螺旋解旋。
298个大肠杆菌启动子共有序列 来自噬菌体和细菌的10个基因的启动子序列来自噬菌体和细菌的10个基因的启动子序列,这些启动子都可被RNA聚合酶中的70亚基(E.coli中最常见的一种亚基)识别。
(b)链延伸是一个核苷酰基转移反应 RNA聚合酶催化链沿着5ˊ至3ˊ方向延伸。链延伸所需的下一个核苷三磷酸经聚合酶验证与模板上相应的未配对的核苷酸正确地形成氢键,然后RNA聚合酶催化延伸着的RNA链的3ˊ-羟基对新进来的核苷三磷酸的α-磷进行亲核攻击,这一核苷酸基转移反应的结果是一个新的磷酸二酯键的形成和释放出焦磷酸,焦磷酸水解为无机磷酸释放能量保证了反应的进行。 当开始延伸后,亚基立刻从 RNA聚合酶上脱落。随着RNA的延伸,已合成的RNA链离开模板,原来解旋的DNA又恢复双螺旋。DNA双链解旋时会使延伸泡前面产生扭曲应力,拓扑异构酶会防止扭曲应力的净增加。
RNA聚合酶 催化的延伸反应
转录延伸 编码链 模板链 转录 转录的RNA 翻译
(c)转录终止在特殊的终止子序列 在大肠杆菌中存在着两种终止机制,一种是蛋白依赖性的链终止,即一个终止蛋白与RNA聚合酶复合体相互作用恰好终止在一个发卡环处,发卡环是在新合成的转录链上形成的。 蛋白是一个ATP依赖性的RNA-DNA解旋酶,其功能是破坏了RNA-DNA杂化体,导致延伸复合体的解离,使合成的RNA链释放出来。
蛋白依赖性的链终止 终止位置 蛋白 蛋白沿着mRNA移动 当聚合酶停止在终止位置时,蛋白与酶结合 蛋白与酶作用,新合成mRNA释放
另一种是蛋白非依赖性的终止作用,其特征是也有一个类似的发卡结构,发卡下游处的模板链上恰好存在一串腺苷酸,对应的转录链应是一串尿苷酸。 所以转录终止也许是当延伸复合体停止在发卡结构处时,使得A-U配对碱基的RNA-DNA杂化体不稳定,导致复合体的解体而终止转录。 转录方向 反向重复序列 反向重复序列 转录 被转录的最后一个碱基 DNA中模板链 转录形成发卡环
蛋白依赖性转录终止 蛋白非依赖性的转录终止
36.3 一些抗生素是RNA合成的抑制剂 放线菌素D来自链霉菌,可以嵌入双链DNA中。放线菌素D中的吩恶嗪酮环插入到相邻的G-C碱基对之间,结构中的环状多肽占据了DNA双螺旋的空间,阻塞了转录的延伸。即使在很低的浓度下,放线菌素D都能有效地抑制原核生物和真核生物转录的延伸过程。 利福霉素是另一个非常有用的抗生素,也是从链霉菌分离出来的。利福霉素通过直接与细菌中的RNA聚合酶的亚基结合来抑制RNA合成,特异地抑制第一个磷酸二酯键的形成。由于利福霉素不抑制真核生物的RNA聚合酶,所以一个合成的利福霉素衍生物利福平已用作为临床的抗结核菌药。
放线菌素 D
36.4 初始 RNA 转录产物都需经转录后加工 1、初始mRNA转录产物的加工 初始mRNA转录产物要经过在其5ˊ端加一个甲基化的“帽子”和在3ˊ端加上一个多聚腺苷酸的尾巴,并且要切去内含子,将外显子拼接在一起构成一个功能性的mRNA。 大多数真核生物中编码蛋白质的基因是由编码蛋白质氨基酸序列的外显子和散布在外显子中非编码序列的内含子构成的,即外显子被非编码序列内含子隔开,所以真核生物基因又称之为断裂基因。刚转录出的RNA产物,也称为非均一RNA(hnRNA)必须通过一种称之RNA剪接机制进行加工。 细菌基因的编码序列是连续的,转录后的mRNA不需加工就可直接作为模板翻译蛋白质。
通过成熟RNA-DNA杂交给出了鸡卵清蛋白的非编码序列(内含子) 灰色表示非编码序列(内含子) 蓝色表示编码序列(外显子) 绿色表示成熟mRNA
真核生物mRNA的5ˊ端有一个“帽子”结构,是由稀有的7-甲基鸟嘌呤通过5ˊ,5ˊ-三磷酸键与初始转录物的起始( 5ˊ )核苷酸连接形成的。这个帽子是在转录到20个核苷酸时在转鸟嘌呤核苷酸酶催化下加到5ˊ端的。 mRNA上的5ˊ帽子对于蛋白质合成的起始是很重要的,同时它可保护mRNA转录本不被核酸外切酶降解。
真核生物的mRNA的3ˊ端是通过涉及一个大的蛋白复合物的两步反应进行转录后修饰的。在第一步反应中,一个聚腺苷酸化作用断裂特异因子(CPSF)识别并结合出现于大多数mRNA的3ˊ端的AAUAA序列。另一个称为断裂刺激因子(CSTF)的蛋白复合物与CPSF相互作用激活mRNA在AAUAA下游大约20个核苷酸的部位断裂。 在第二步3ˊ加工反应中,聚腺苷酸聚合酶(PAP)结合于转录产物的游离的3ˊ端,加上大约20个腺苷酸的3ˊ端的poly(A)尾巴,这个短的尾巴通过寡聚腺苷酸结合蛋白刺激PAP活性再进一步延伸到大约200个腺苷酸。聚腺苷酸化反应是一个重要的调节步骤,因为聚腺苷酸尾巴的长度能调节mRNA的稳定性和翻译效率。
真核生物mRNA 3ˊ端的聚腺苷酸化
hnRNA加工成 mRNA的过程 向细胞质转运 外显子1 外显子2 外显子3 5’末端加帽和转录终止 3’末端剪切和聚腺苷酸化 内含子 剪去内含子并拼接 向细胞质转运
大多数初始mRNA剪接涉及到两步转酯基反应 一些小核RNA和辅助蛋白形成核内核糖核蛋白(snRNP)识别剪接位点和参与剪接。 在第一次转酯基反应中,内含子中的一个腺苷酸2 ˊ羟基和同一内含子中5ˊ核苷酸之间形成一个不常见的2ˊ,5ˊ-磷酸二酯键。 在第二次转酯基反应中,上游外显子的3ˊ羟基攻击处于下游的外显子-内含子边界的5ˊ磷酸。这一拼接反应涉及称之套索的一个中间体结构的形成,它是第二次转酯基反应之后释放出来的。 套索中的2ˊ,5ˊ-磷酸二酯键位于称之分支点的部位,这个部位形成了一个带有3个磷酸二酯键的结构,即腺苷酸残基与内含子的其余部分是通过标准的3ˊ,5ˊ-磷酸二酯键连接的。
内含子 外显子 1 外显子 2 5剪接部位 3剪接部位 内含子 外显子 1 外显子 2 剪接点 外显子 1 外显子 2
2、初始 rRNA加工 大肠杆菌中在单一的转录本内就含有7个rRNA基因的拷贝。初级转录物大约含有6500个核苷酸,编码23S和16S rRNA,同时还编码着几种tRNA和小的5 S rRNA。 分子克隆和核酸测序表明,23 S rRNA大约含有2900核苷酸,而16S rRNA大约含有1600核苷酸。将初始转录物加工成23 S rRNA和16S rRNA需要特异的核酸酶(RNases)催化。 下图给出了把人的45S rRNA加工成28S 和18S 两个有功能的rRNA的加工过程的示意图。
将人45S rRNA 加工成 28S 和18S 两个有功能的rRNA的过程
3、 tRNA的加工 2.对某些tRNA通过核苷酸基转移酶加上末端CCA序列。 3.进行特异碱基修饰。 1. RNase通过tRNA剪接反应除去内含子。 2.对某些tRNA通过核苷酸基转移酶加上末端CCA序列。 3.进行特异碱基修饰。
加氨基酸部位 加工 反密码子
36.5 核酶是个位置特异的核酸内切酶 外显子2 外显子1 1981年Cech发现四膜虫rRNA内含子的切除和外显子的拼接不需要酶催化,而是一个RNA自我拼接的过程。具有催化功能的RNA分子称为核酶。 内含子 剪接过程: 1、外部的鸟苷与内含子的5ˊ核苷酸形成新的磷酸二酯键,释放出外显子1的3 ˊOH。 2、出外显子1的3 ˊOH攻击外显子2的5ˊ磷酸,形成磷酸二酯健,将两个外显子连接在一起。 3、内含子去除5 ˊ端片段后形成环状。
36.6 基因转录调控-操纵子学说 一个基因指的是编码蛋白质多肽链和功能RNA的DNA(某些病毒基因为RNA),基因表达就是遗传信息杜转录和翻译过程。 基因表达的调控可以在不同水平,例如转录水平,或翻译水平上进行。 原核生物的基因调控主要是在转录水平上进行,因为他们的转录和翻译过程可同时进行。 真核生物的转录和翻译过程在时间和空间上都被分隔开,而且转录和翻译后都要经过复杂的加工过程,所以其基因调控在不同水平都需要进行调控。
Jacob和Monod在对大肠杆菌乳糖发酵过程中酶的生物合成研究时,探讨了基因表达调控机制,并提出了乳糖操纵子模型。 操纵子:由一个启动子共转录的几个不同基因组成的转录单位,即由启动子序列、操纵基因和受操纵基因调控的一个或多个相关基因(结构基因)组成的基因表达单位。操纵子模型可以很好说明原核生物基因表达的调控机制。 控制部位 几个相关基因(结构基因) 调节物基因 操纵子的一般结构
大肠杆菌乳糖操纵子 大肠杆菌乳糖操纵子包括依次排列的启动子、操纵基因和三个结构基因。结构基因 Z 编码分解乳糖的-半乳糖苷酶、Y 编码吸收乳糖的-半乳糖苷透性酶、 a 编码-半乳糖苷乙酰基转移酶。O代表操纵基因,i 代表调节基因,P 代表启动子。如果进行转录,转录产物是一条多顺反子mRNA。 大肠杆菌能够利用乳糖作为它的唯一碳源,但需要合成代谢乳糖的三个酶: -半乳糖苷酶(降解乳糖生成半乳糖和葡萄糖)、-半乳糖苷透性酶(使乳糖进入细胞内)、 -半乳糖苷乙酰基转移酶(半乳糖代谢时需要的酶)。这三个酶都是乳糖存在时诱导生成的酶。 乳糖操纵子
当细胞中没有乳糖或其他诱导物时,调节物基因经转录、表达生成阻遏物,然后阻遏物特异与操纵基因结合,阻止 z、y、a 基因的转录。
当细胞中存在乳糖或其他诱导物时,诱导物与阻遏物结合,使阻遏物构象发生变化,从而使阻遏物不能与操纵基因结合。这种状况下,在RNA聚合酶催化下可以进行 z、y、a 基因的转录。
要点归纳 1. RNA合成需要DNA依赖性的RNA聚合酶全酶,E.coli RNA聚合酶全酶亚基组成为αβ2βˊσ,其中的σ亚基在转录起始时被用来识别启动子,该酶的核心酶亚基组成为αβ2βˊ。真核生物的细胞核中存在3种不同的RNA聚合酶:RNA pol Ⅰ负责合成rRNA前体,而RNA pol Ⅱ负责合成mRNA前体,RNA pol Ⅲ负责合成tRNA前体。 2. RNA聚合酶催化以DNA模板链为模板转录生成互补的、反平行的RNA,用的底物是核糖核苷三磷酸,不需要引物。RNA聚合酶催化3′羟基与核苷三磷酸的5′-α-磷酸基团形成磷酸二酯键,使核苷酸整合到延伸的聚核苷酸链中,同时释放出焦磷酸,焦磷酸水解释放的能量驱动聚合反应。RNA合成方向是5′→3′方向。另外RNA聚合酶不具有校正核酸酶活性。
3. 转录包括3个阶段:起始、延伸和终止。在原核生物中(如E 3. 转录包括3个阶段:起始、延伸和终止。在原核生物中(如E.coli),当σ亚基与RNA聚合酶的核心酶结合形成全酶后附着在DNA的启动子区时,转录起始。然后通过自启作用结合ATP(GTP),ATP(GTP)提供3′-羟基用于第二个核苷酸的聚合,即RNA合成不需要预先存在的引物。启动转录开始合成新链后,σ亚基从全酶上解离。 4. 当转录进行时,DNA局部解旋,形成所谓的转录泡,转录泡以每秒50个核苷酸的速度沿着DNA模板移动。新合成的RNA链含有结束转录的终止信号。一种终止信号是一个RNA发卡,该发卡位于几个U残基后面。不同的终止信号可以被rho蛋白(一种ATPase)识别。
5. 无论是原核生物还是真核生物的rRNA和tRNA的大的初始转录物都要经过加工(包括碱基修饰)形成小的、成熟的RNA。原核生物的mRNA不用加工,直接作为模板翻译成蛋白质。 真核生物的mRNA初级转录物需要切去内含子,并将被内含子隔开的各个外显子剪接起来。进一步加工涉及到在5′端形成甲基化的帽子,以及在3′端形成聚腺苷酸(pol(A))尾部RNA。 6. 某些RNA分子具有催化活性,例如来自四膜虫的26S核糖体RNA在没有蛋白质参与下可以进行自我剪接。该rRNA内含子的自我剪接表现出了真正的催化活性,所以现在把这样的RNA称之为核酶。
7. 在原核细胞中,酶的合成会受到阻遏物抑制。酶的诱导和抑制可以用操纵子假说解释,依据该假说,转录的功能单位是一个由控制元件(启动子)和编码特定蛋白的结构基因组成的操纵子。操纵子模型可以很好说明原核生物基因表达的调控机制。但真核生物不组成操纵子。 8. 大肠杆菌乳糖操纵子包括依次排列的启动子、操纵基因和三个结构基因。在没有诱导物存在时,阻遏物与操纵基因结合,阻止结构基因的转录;而当有诱导物存在时,诱导物与阻遏物结合,使得该阻遏物不能与操纵基因结合。结果在乳糖存在下,乳糖操纵子中结构基因被转录为一条多顺反子mRNA,然后表达出乳糖代谢所需要的三个酶。