Download presentation
2
A11 Regulation of gene expression in eukaryotes

3
2 真核生物基因表达调控的特点 真核生物表达调控与原核生物的不同: ( 1 )染色体结构不同; ( 2 )原核生物具有正调控和负调控并重的特点,真核 生物目前已知的主要是正调控; ( 3 )原核生物的转录和翻译是相偶联的,真核生物的 转录和翻译在时空上是分开的; ( 4 )真核生物是多细胞的,在生物的个体发育过程中 其基因表达在时间和空间上具有特异性,即细胞特异性 或组织特异性表达。
染色体结构不同; ( 2 )原核生物具有正调控和负调控并重的特点,真核 生物目前已知的主要是正调控; ( 3 )原核生物的转录和翻译是相偶联的,真核生物的 转录和翻译在时空上是分开的; ( 4 )真核生物是多细胞的,在生物的个体发育过程中 其基因表达在时间和空间上具有特异性,即细胞特异性 或组织特异性表达。")
4
Contents Gene expression in eukaryotes Regulation on DNA level Regulation of transcription Transcription factors Regulation of gene expression by hormones and cytokines Post-transcriptional regulation Translational regulation Gene expression and development

5
1. Gene expression in eukaryotes Cells express about 15% of their genes Cell-specific expression: e.g. In lymphocytes ['l ɪ mfə,sa ɪ t], antibody genes are expressed at a high level.

6
Expression pattern can change during the lifetime of a cell: e.g. from primitive progenitor [prə ʊ 'd ʒ en ɪ tə] cells to blood cells Importance in understanding gene expression

7
Various levels of regulation that are possible during the expression of genetic material

9
2. Regulation on DNA level 基因的丢失:不可逆 某些真核生物随细胞的分化丢失了染色体上的某些 DNA 片段的现 象称为基因丢失. 如小麦瘿蚊、马蛔虫体细胞 基因扩增( copy number increased ) 爪蟾卵母细胞的灯刷染色体 : 非洲爪蟾卵母细胞 rRNA 基因卵裂 时,扩增 2000 倍,达 1012 个核糖体。 药物:诱导抗药性基因的扩增; 肿瘤细胞:原癌基因拷贝数异常增加。 基因重排( rearrangement ) 酵母交配型的转变 抗体多样性 如免疫球蛋白基因能发生重排,形成多样性。
 爪蟾卵母细胞的灯刷染色体 : 非洲爪蟾卵母细胞 rRNA 基因卵裂 时,扩增 2000 倍,达 1012 个核糖体。 药物:诱导抗药性基因的扩增; 肿瘤细胞:原癌基因拷贝数异常增加。 基因重排( rearrangement ) 酵母交配型的转变 抗体多样性 如免疫球蛋白基因能发生重排,形成多样性。.")
10
酵母交配型的转变: 当活性暗盒信息被沉默暗盒信息所取代时,新 “ 装 进 ” 活性暗盒的信息就可以表达,就可以发生交配型转换。可见,细胞通过 交替改变 MAT 座位中的等位基因,控制细胞的交配型

11
The structure of antibody 轻链 L 重链 H 轻链 L 重链 H V 区的高变异 性决定了抗体 的多样性

12
1976 年, Hozumi 和利根川进 (Tonegawa) 发现在胎鼠细 胞中编码V区和C区的 DNA 序列相隔较远,但在成熟的 抗体细胞中(淋巴瘤细胞)这两个区域却紧密相连,表明 : 在淋巴细胞分化过程中,免疫球蛋白基因曾发生过体细胞重组。 λ 轻链 V 基因只编码 V 区 108 个氨基酸中的 95 个,最后的 39bp ( 13 个密码子)消失了,移到了 C 基因附近,即连接 V 和 C 基因的 DNA 序列 J ( Joining )区附近。 1982 年 Hood 和利根川进等又在 H 链的编码区中发现了编 码高变区氨基酸的多样性基因( diversity ),即 D 基因, 其位置在 V 、 J 基因之间。 证明抗体的多样性由体细胞水平上的基因重排产生。
 发现在胎鼠细 胞中编码V区和C区的 DNA 序列相隔较远,但在成熟的 抗体细胞中(淋巴瘤细胞)这两个区域却紧密相连,表明 : 在淋巴细胞分化过程中,免疫球蛋白基因曾发生过体细胞重组。 λ 轻链 V 基因只编码 V 区 108 个氨基酸中的 95 个,最后的 39bp ( 13 个密码子)消失了,移到了 C 基因附近,即连接 V 和 C 基因的 DNA 序列 J ( Joining )区附近。 1982 年 Hood 和利根川进等又在 H 链的编码区中发现了编 码高变区氨基酸的多样性基因( diversity ),即 D 基因, 其位置在 V 、 J 基因之间。 证明抗体的多样性由体细胞水平上的基因重排产生。")
13
小鼠在发育过程中重链通 过基因重组发生 DNA 重排 小鼠在发育过程中轻链通 过基因重组发生 DNA 重排

14
3. Regulation of transcription 顺式作用元件 ( cis-acting element ) 反式作用因子 (trans-acting Factor)
 反式作用因子 (trans-acting Factor)")
15
顺式作用元件是指具有调节功能的特定 DNA 序列或影响自身基 因表达活性的 DNA 序列,在基因的同一条 DNA 分子上; 顺式作用元件类型: ( 1 )启动子 (promoter) :核心启动子和上游启动子 ( 2 )增强子 (enhancer) :可以改变染色体的构象;为转录因子进入启动子区 提供位点 加强相连基因从正确起始位点的转录活性; 增强子无论是在下游或在上游均可激活转录; 无论是在下游或在上游,可在远离起始位点 1Kb 以上发挥作用; 两个启动子串联在一起时,增强子优先激活距离最近的那一个 ( 3 )沉默子 (silencer ) :负性调节元件,起阻遏作用。 ( 4 )绝缘子 (insulator, boundary element): 在真核基因及其调控区的一段 DNA 序列 。
启动子 (promoter) :核心启动子和上游启动子 ( 2 )增强子 (enhancer) :可以改变染色体的构象;为转录因子进入启动子区 提供位点 加强相连基因从正确起始位点的转录活性; 增强子无论是在下游或在上游均可激活转录; 无论是在下游或在上游,可在远离起始位点 1Kb 以上发挥作用; 两个启动子串联在一起时,增强子优先激活距离最近的那一个 ( 3 )沉默子 (silencer ) :负性调节元件,起阻遏作用。 ( 4 )绝缘子 (insulator, boundary element): 在真核基因及其调控区的一段 DNA 序列 。")
17
反式作用元件的结构和功能: ( 1 )概念: 反式作用元件是 DNA 结合蛋白(转录因子),可 使邻近基因开放(正调控)或关闭(负调控)。 The transcription factors are synthesized in the cytoplasm of the cell but exert their effects in the nucleus, so are called trans-acting factors. 核蛋白 ( 2 )通用或基本转录因子 —RNA 聚合酶结合启动子所必需的 一组蛋白因子。 如: TF Ⅱ A 、 TF Ⅱ B 、 TF Ⅱ D 、 TF Ⅱ E 等。 ( 3 )特异转录因子 ( special transcription factors)— 个别基 因转录所必需的转录因子. 如: OCT-2 :在淋巴细胞中特异性表达,识别 Ig 基因的启动 子和增强子。
通用或基本转录因子 —RNA 聚合酶结合启动子所必需的 一组蛋白因子。 如: TF Ⅱ A 、 TF Ⅱ B 、 TF Ⅱ D 、 TF Ⅱ E 等。 ( 3 )特异转录因子 ( special transcription factors)— 个别基 因转录所必需的转录因子. 如: OCT-2 :在淋巴细胞中特异性表达,识别 Ig 基因的启动 子和增强子。.")
18
4. Transcription factors Transcription factors are a large family of proteins that regulate the expression of protein coding genes. The overall effect on transcription depends on the complement of transcription factors bound.

19
Transcription factors consist of discrete protein domains with specific functions. Common domains DNA binding domains DNA 结合结构域 Dimerization domains 二聚化结构域 Transactivation domains 转录激活结构域

21
DNA 结合域: 1.Zinc finger 2.Helix-Turn-Helix 3.Basic domains

22
21 锌指结构域 The zinc finger domain 锌指结构有 2 种形式: C 2 H 2 zinc finger 和 C4 zinc finger C 2 H 2 zinc finger :由 12 个氨基酸组成的环,通过 2 个半胱氨酸( C , Cys )和 2 个组氨酸( H , His )残基固定,这 4 个残基与 Zn 2+ 在空间 上形成一个四面体结构。 一般情况下需要 3 个或更多的 C 2 H 2 型锌指 才能与 DNA 结合,如在 TFIIA 有 9 个这样的锌指结构,转录因子 SP1 有 3 个。 C4 zinc finger : Zn 2+ 与 4 个半胱氨酸( C , Cys )结合,存在于类 固醇激素受体转录因子中。
和 2 个组氨酸( H , His )残基固定,这 4 个残基与 Zn 2+ 在空间 上形成一个四面体结构。 一般情况下需要 3 个或更多的 C 2 H 2 型锌指 才能与 DNA 结合,如在 TFIIA 有 9 个这样的锌指结构,转录因子 SP1 有 3 个。 C4 zinc finger : Zn 2+ 与 4 个半胱氨酸( C , Cys )结合,存在于类 固醇激素受体转录因子中。")
23
β- 折叠 α- 螺旋 半胱氨酸 Cys 碱性保守氨 基酸 组氨酸 His C 2 H 2 zinc finger C4 zinc finger 立体结构 半胱氨酸 Cys DNA binding

24
23 The zinc finger domain 锌指结构域 半胱氨酸 组氨酸 β- 折叠 α- 螺旋

26
25 The helix-turn-helix domain 螺旋 - 转角 - 螺旋结构域 特点: DNA 结合蛋白包含有 60 个氨基酸的 同源域( Homeodomain ),由同源 框序列编码( homeobox )。 DNA 结合结构域由 4 个 -helices (螺 旋)组成,螺旋Ⅱ和 Ⅲ保持了适当的 角度,并被一个特异的 β-turn (转角) 所分隔。 α 识别螺旋处于 DNA 的大沟中,并与 DNA 发生作用
,由同源 框序列编码( homeobox )。 DNA 结合结构域由 4 个 -helices (螺 旋)组成,螺旋Ⅱ和 Ⅲ保持了适当的 角度,并被一个特异的 β-turn (转角) 所分隔。 α 识别螺旋处于 DNA 的大沟中,并与 DNA 发生作用")
28
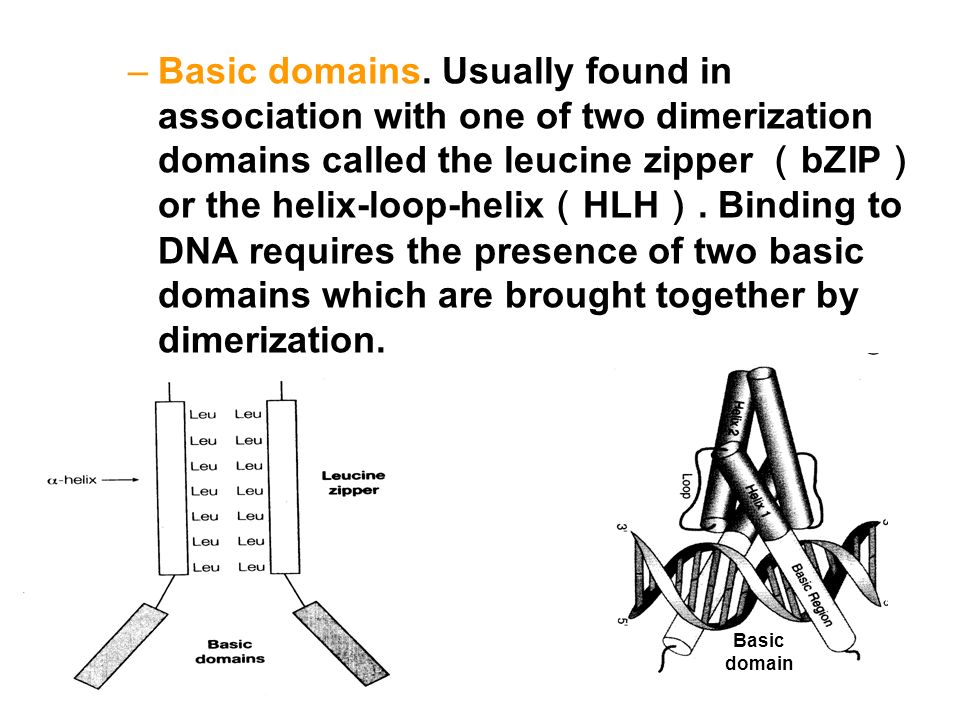
–Basic domains. Usually found in association with one of two dimerization domains called the leucine zipper ( bZIP ) or the helix-loop-helix ( HLH ). Binding to DNA requires the presence of two basic domains which are brought together by dimerization. Basic domain
 or the helix-loop-helix ( HLH ). Binding to DNA requires the presence of two basic domains which are brought together by dimerization. Basic domain.")
29
Dimerization domains –Basic Leucine zipper ( bZIP ) –Helix-Loop-Helix ( HLH )
 –Helix-Loop-Helix ( HLH )")
30
29 亮氨酸拉链 (leucine zipper) 结构域 ( 1 )亮氨酸拉链( Leucine zipper )的肽链上每隔 7 个残基 就有一个疏水的亮氨酸残基,这 些残基位于 DNA 结合域的 C 端 α- 螺旋上,形成一个疏水表面,疏 水表面的相互作用,形成了二聚 体 —— 卷曲的卷曲结构 ( coiled-coil structure )如 bZIP 转录因子。 ( 3 )蛋白的二聚作用使 2 个碱性 结构域相邻,进而可与 DNA 发生 作用。
 结构域 ( 1 )亮氨酸拉链( Leucine zipper )的肽链上每隔 7 个残基 就有一个疏水的亮氨酸残基,这 些残基位于 DNA 结合域的 C 端 α- 螺旋上,形成一个疏水表面,疏 水表面的相互作用,形成了二聚 体 —— 卷曲的卷曲结构 ( coiled-coil structure )如 bZIP 转录因子。 ( 3 )蛋白的二聚作用使 2 个碱性 结构域相邻,进而可与 DNA 发生 作用。")
31
30 亮氨酸拉链 (leucine zipper)
")
32
螺旋 - 环 - 螺旋结构域 The helix-loop-helix domain , HLH 结构域。 Two helices separated by a nonhelical loop. Dimerization by hydrophobic (疏水的) amino acids present on one side of the carboxyl terminal helix Homodimers and heterodimers: e.g. MyoD family in regulating gene expression in developing muscle cells
 amino acids present on one side of the carboxyl terminal helix Homodimers and heterodimers: e.g. MyoD family in regulating gene expression in developing muscle cells.")
33
可变连接区 DNA 特异位点

34
螺旋 - 环 - 螺旋 DNA 特异位点

35
Activation domains ( Rich in certain amino acids ) 酸性激活结构域 Acidic activition domain: 酵母 Gcn4 和 Gal4 、哺乳动物糖皮质激素受体、疱疹病毒激活子的 转录激活结构域均含有高比例的酸性氨基酸,称为 酸性激活结构域 富含谷氨酰胺结构域 Glutamine-rich domains 在 SP1 转录因子的 2 个激活区域上发现富含谷氨酰胺结构域,这 些结构域与果蝇触角蛋白质的结构域可以互换。 富含脯氨酸结构域 Prolin-rich domain: 在 c-Jun 、 AP2 和 OCT-2 等转录因子中发现富含脯氨酸结构域 ,这 个结构域可以激活转录。
 酸性激活结构域 Acidic activition domain: 酵母 Gcn4 和 Gal4 、哺乳动物糖皮质激素受体、疱疹病毒激活子的 转录激活结构域均含有高比例的酸性氨基酸,称为 酸性激活结构域 富含谷氨酰胺结构域 Glutamine-rich domains 在 SP1 转录因子的 2 个激活区域上发现富含谷氨酰胺结构域,这 些结构域与果蝇触角蛋白质的结构域可以互换。 富含脯氨酸结构域 Prolin-rich domain: 在 c-Jun 、 AP2 和 OCT-2 等转录因子中发现富含脯氨酸结构域 ,这 个结构域可以激活转录。")
36
Transcription factors regulate the expression of target genes by binding to the transcription initiation complex and changing its stability. Some transcription factors can repress transcription either directly or indirectly

37
5. Regulation of gene expression by hormones and cytokines Steroid hormone (类固醇激素) : lipid- soluble, can pass through the cell membrane into the cytoplasm and bind to steroid hormone receptor, release and dimerize and translocate to the nucleus, bind promoter and activate transcription
 : lipid- soluble, can pass through the cell membrane into the cytoplasm and bind to steroid hormone receptor, release and dimerize and translocate to the nucleus, bind promoter and activate transcription.")
38
Polypeptide hormone (多肽类激素 ) and cytokines (细胞因子) : bind receptor, signal transduction, stimulate transcription
 and cytokines (细胞因子) : bind receptor, signal transduction, stimulate transcription")
39
5. Post-transcriptional regulation 5′ capping and 3′ polyA tailing Alternative splicing 外显子选择 (optional exon) 、内含子选择 (optional intron) 、互斥外显 子、内部剪接位点 RNA editing micro RNA (miRNA)/RNA interference(RNAi)
 、内含子选择 (optional intron) 、互斥外显 子、内部剪接位点 RNA editing micro RNA (miRNA)/RNA interference(RNAi).")
40
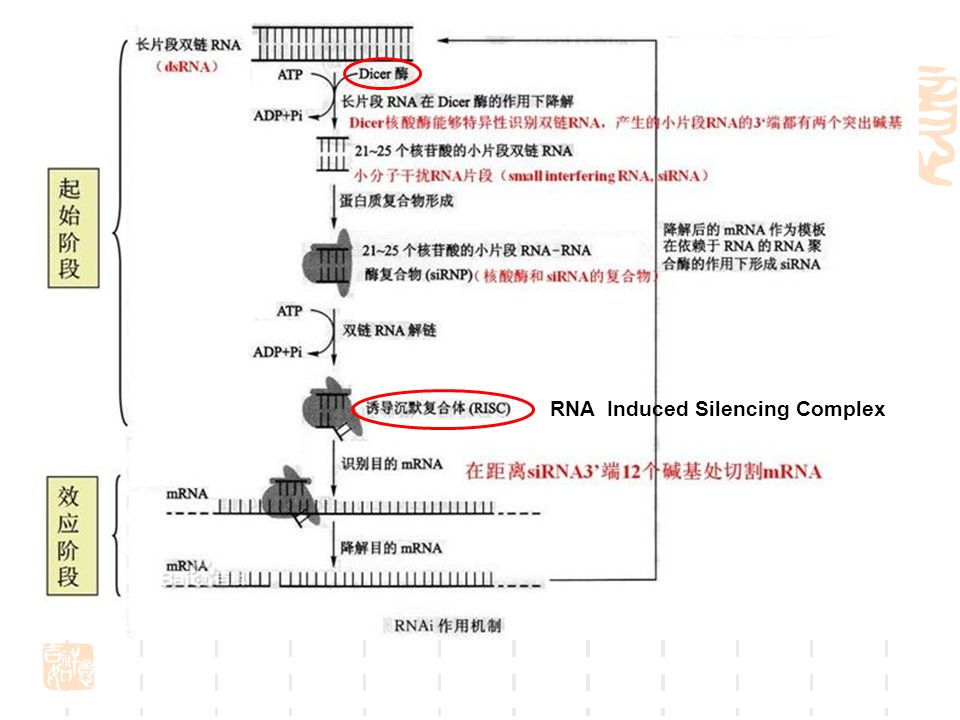
miRNA/ RNAi miRNA 长度为 22nt 左右 5′ 端带磷酸基团、 3′ 端带羟基的非蛋白编码的调控小 RNA 家 族, 参与发育、衰老、死亡的时序控制、神经 分化、细胞凋亡、新陈代谢、癌症、脂肪 细胞分化的基因表达调控。

42
RNA Induced Silencing Complex

43
42 5’ 端 UTR( 非翻译区)结构与翻译起始的调节 5‘ 帽子结构的甲基化: 1 )保护 mRNA 免遭 5’ 外切酶 的降解。 2 )为 mRNA 的从核中输出提供转运信号。 3 ) 提高翻译模板的稳定性和翻译效率。 翻译起始因子的调控: eIF-2-4F 的磷酸化能提高翻译速度 eIF-2α 的磷酸化能抑制翻译起始 6. Translational regulation

44
43 翻译起始因子 (eIF 2 ) 的调控
 的调控")
45
44 3‘UTR (非翻译区)结构与 mRNA 稳定性 多聚腺苷酸尾的调节作用 poly A 尾巴的功能: 1 )稳定 mRNA 分子,抵抗 3’- 端外切酶攻击的作用。 2 )有助于细胞质中成熟 mRNA 的翻译。 3 ) 3‘ UTR 区域具有保护 5‘ 端帽子结构的作用。
结构与 mRNA 稳定性 多聚腺苷酸尾的调节作用 poly A 尾巴的功能: 1 )稳定 mRNA 分子,抵抗 3’- 端外切酶攻击的作用。 2 )有助于细胞质中成熟 mRNA 的翻译。 3 ) 3‘ UTR 区域具有保护 5‘ 端帽子结构的作用。")
46
45 3‘UTR 序列及结构对 mRNA 稳定性的调节 球蛋白 mRNA 的 3’URT 序列含有许多 CCUCC 重复蛋 白序列,这些序列发生突变将降低 mRNA 的稳定性。 某些不稳定 mRNA 的 3’UTR 含有 50nt 的共有序列 AUUUA ,称为 ARE 。 ARE 结合蛋白可以聚集外切多 聚腺苷酸酶和内切酶,加速 mRNA 的降解。

47
7.Gene expression and development 不同发育阶段基因表达的调控 个体发育受多个基因群相互作用的多层次网络控制, 这些基因的表达在时空上相互联系和配合,控制发育 的遗传程序, 如果蝇 不同组织器官基因表达的调控 基因表达的空间特异性

48
细胞凋亡 多细胞有机体为调控机体发育,维持内环境稳定,由 基因调控的细胞主动死亡过程。 凋亡基因: bcl-2

49
Questions The levels of regulation of gene expression in eukaryotes. The mechanism that transcription is regulated in eukaryotes. List cis-elements and transcription factors in eukaryotes. What does post-transcriptional regulation mean?







 谢喜秀 1040800114.>")



>")
 第10章 生命為何偏愛螺旋狀 (生物大分子螺旋)>")









主要由多细胞组成,每个细胞基因组中蕴藏的遗传信息量及基因数量都大大高于原核生物。人类细胞单倍体基因组有3×109bp,为大肠杆菌总DNA的800倍,噬菌体的10万倍左右!>")



>")