Download presentation
1
第五章 蛋白质(Protein)
")
2
第一节 引言 蛋白质是构成生物体的基本物质。 病毒,细菌,激素,植物和动物细胞原生质都是以蛋白质为基础的。
第一节 引言 蛋白质是构成生物体的基本物质。 病毒,细菌,激素,植物和动物细胞原生质都是以蛋白质为基础的。 酶是蛋白质 已经发现数以千计的酶。 蛋白质是由20种氨基酸构成的聚合物。

3
一、蛋白质的分类 (一)根据组成分类 简单蛋白质(homoproteins) 在细胞中未经酶催化改性的蛋白质。仅含有氨基酸。
结合蛋白质(conjugated proteins)或杂蛋白质(heteroproteins) 经过酶催化改性的或与非蛋白质组分复合的蛋白质。含有氨基酸和其它非蛋白质化合物。 辅基(prosthetic proteins) 结合蛋白中的非蛋白质组分。
或杂蛋白质(heteroproteins) 经过酶催化改性的或与非蛋白质组分复合的蛋白质。含有氨基酸和其它非蛋白质化合物。 辅基(prosthetic proteins) 结合蛋白中的非蛋白质组分。")
4
结合蛋白质包括: 核蛋白(核蛋白体) 糖蛋白(卵清蛋白、κ-酪蛋白) 磷蛋白(α-和β-酪蛋白、激酶、磷酸化酶)
核蛋白(核蛋白体) 糖蛋白(卵清蛋白、κ-酪蛋白) 磷蛋白(α-和β-酪蛋白、激酶、磷酸化酶) 脂蛋白(蛋黄蛋白质、几种肌浆蛋白质) 金属蛋白(血红蛋白、肌红蛋白和几种酶)。
 糖蛋白(卵清蛋白、κ-酪蛋白) 磷蛋白(α-和β-酪蛋白、激酶、磷酸化酶) 脂蛋白(蛋黄蛋白质、几种肌浆蛋白质) 金属蛋白(血红蛋白、肌红蛋白和几种酶)。")
5
(二)根据大体上的结构形式分类 球蛋白 以球状或椭圆状存在的蛋白质,由多肽链自身折叠而成;
球蛋白 以球状或椭圆状存在的蛋白质,由多肽链自身折叠而成; 纤维状蛋白 棒状分子,含有相互缠绕的线状多肽链(例如,原肌球蛋白、胶原蛋白、角蛋白和弹性蛋白)。纤维状蛋白也能由小的球状蛋白经线性聚集而成,例如肌动蛋白和血纤维蛋白。 大多数酶是球状蛋白,而纤维状蛋白总是起着结构蛋白的作用。
。纤维状蛋白也能由小的球状蛋白经线性聚集而成,例如肌动蛋白和血纤维蛋白。 大多数酶是球状蛋白,而纤维状蛋白总是起着结构蛋白的作用。")
6
二、蛋白质的生物功能 酶 结构蛋白 收缩蛋白(肌球蛋白、肌动蛋白、微管蛋白) 激素(胰岛素、生长激素)
传递蛋白(血清蛋白、铁传递蛋白、血红蛋白) 抗体(免疫球蛋白) 储藏蛋白(蛋清蛋白、种子蛋白) 储藏蛋白主要存在于蛋和植物种子中。 保护蛋白(毒素和过敏素)。
 抗体(免疫球蛋白) 储藏蛋白(蛋清蛋白、种子蛋白) 储藏蛋白主要存在于蛋和植物种子中。 保护蛋白(毒素和过敏素)。")
7
食品蛋白质 易于消化 无毒 富有营养 显示功能特性 来源丰富

8
表5-1 常见食物的蛋白质含量 ━━━━━━━━━━━━━━━━━━━━━━━━━ 食物 蛋白质 食物 蛋白质 食物 蛋白质 (%) (%) (%) ───────────────────────── 猪肉 鸡蛋 花生 牛肉 稻米 大豆 羊肉 小麦 菠菜 鸡肉 小米 油菜 肝 玉米 大白菜 1.1 脑 高梁 白萝卜 0.6 鲤 小麦粉 桔子 牛乳 红薯 苹果
 (%) (%) ───────────────────────── 猪肉 鸡蛋 13.4 花生 牛肉 稻米 8.5 大豆 羊肉 小麦 12.4 菠菜 1.4. 鸡肉 21.5 小米 9.4 油菜 1.4. 肝 玉米 8.6 大白菜 1.1. 脑 7-9 高梁 9.6 白萝卜 0.6. 鲤 18.1 小麦粉 11.0 桔子 0.9. 牛乳 3.5 红薯 1.3 苹果 0.2.")
9
第二节 氨基酸的物理化学性质 一、氨基酸的一般性质 (一)一般结构和分类 蛋白质含有碳、氢、氧和氮等元素,有些含有硫或磷元素,少数还含有锌、铁、铜、锰等元素。不同的蛋白质其组成和结构也不同。 各种蛋白质的含氮量很接近,其平均值为16%。

10
组成蛋白质的氨基酸除了脯氨酸和羟脯氨酸外都是α-氨基酸。
都是L-型的α-氨基酸。 决定物化性质

11
蛋白质中常见氨基酸根据其分子结构可归纳为六类
中性氨基酸 一氨基,一羧基 GLY(甘),ALA(丙),VAL(缬), LEU(亮), ILE(异亮) 酸性氨基酸 一氨基,二羧基 ASP(天冬),GLU(谷) 碱性氨基酸 二氨基,一羧基 ARG(精),LYS(赖) 含羟氨基酸 一氨基,一羧基,一羟基 SER(丝),THR(苏) 含硫氨基酸 一氨基,一羧基,一巯基 CYS( 胱氨酸),MET(蛋) 含环氨基酸 一氨基,一羧基,一环 PHE(苯丙),TRP(色), HIS(组),PRO(脯),TYR(酪)
,ALA(丙),VAL(缬), LEU(亮), ILE(异亮) 酸性氨基酸 一氨基,二羧基. ASP(天冬),GLU(谷) 碱性氨基酸 二氨基,一羧基. ARG(精),LYS(赖) 含羟氨基酸 一氨基,一羧基,一羟基. SER(丝),THR(苏) 含硫氨基酸 一氨基,一羧基,一巯基. CYS( 胱氨酸),MET(蛋) 含环氨基酸 一氨基,一羧基,一环. PHE(苯丙),TRP(色), HIS(组),PRO(脯),TYR(酪)")
12
按侧链基团的极性可分为四类 非极性氨基酸或疏水性氨基酸。含有脂肪族(Ala、Ile、Leu、Met、Pro和Val)和芳香族侧链的氨基酸(Phe,Trp)是疏水的。 不带电荷的极性氨基酸,侧链与水结合氢键如: SER,THR,TYR,CYS, ASP,GLY。 在pH7时(生理条件下)带正电荷的极性氨基酸(碱性氨基酸),如LYS,ARG,HIS。 在pH7时(生理条件下)带负电荷的极性氨基酸(酸性氨基酸), 如ASP,GLU。 碱性和酸性氨基酸具有很强的亲水性。
带正电荷的极性氨基酸(碱性氨基酸),如LYS,ARG,HIS。 在pH7时(生理条件下)带负电荷的极性氨基酸(酸性氨基酸), 如ASP,GLU。 碱性和酸性氨基酸具有很强的亲水性。")
13
甘丙缬亮异脂链 丝苏半蛋羟硫添 脯酪色苯杂芳环 天谷精赖组酸碱

14
必需氨基酸 有些氨基酸,机体合成不足,必需从食物或饲料中供给,如果食物或饲料中缺乏这些氨基酸,就会影响机体的正常生长和健康。
人体必需氨基酸有LYS,PHE, VAL, MET, TRP, LEU, ILE及THR八种,此外,HIS对于婴儿的营养也是必需的。

15
(二)氨基酸的立体化学 除甘氨酸外都具有旋光性。 在植物或动物组织的蛋白质水解物中,仅发现L-型异构体。
氨基酸的立体化学 除甘氨酸外都具有旋光性。 在植物或动物组织的蛋白质水解物中,仅发现L-型异构体。")
16
(三)氨基酸的酸碱性质:离子化 在中性pH范围,α-氨基和羧基都处在离子化状态,此时是偶极离子或两性离子。两性电解质。
偶极离子以电中性状态存在时的pH被称为等电点(pI)。
。")
17
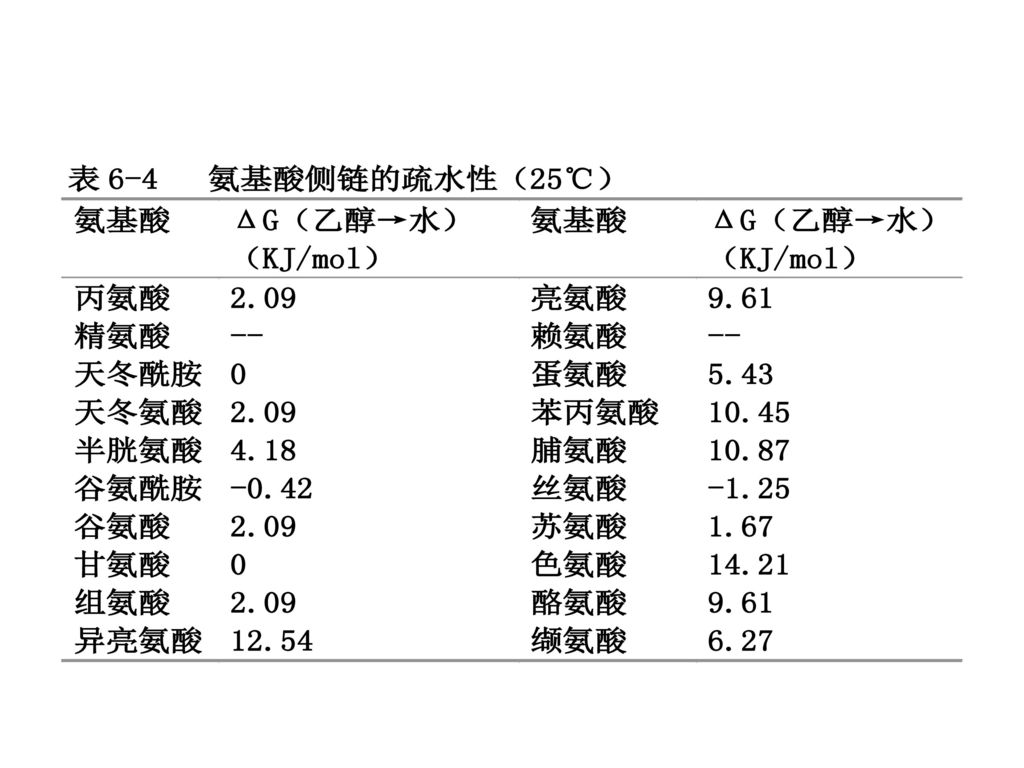
(四)氨基酸的疏水性 蛋白质在水中的溶解度主要取决于氨基酸组分侧链上极性(带电或不带电)和非极性(疏水)基团的分布。
氨基酸在水中和极性溶剂(例如乙醇)中的相对溶解度决定着氨基酸以及多肽和蛋白质的疏水性。 如果不考虑活度系数,那么1摩尔氨基酸从乙醇转移到水溶液的自由能变化可表示 氨基酸的疏水性。 ΔGt(Et→W)
中的相对溶解度决定着氨基酸以及多肽和蛋白质的疏水性。 如果不考虑活度系数,那么1摩尔氨基酸从乙醇转移到水溶液的自由能变化可表示 氨基酸的疏水性。 ΔGt(Et→W)")
18
缬氨酸(Val) ΔGt,Val=ΔGt,Gly+ΔGt,异丙基侧链 ΔGt,异丙基侧链=ΔGt,Val-ΔGt,Gly
 ΔGt,Val=ΔGt,Gly+ΔGt,异丙基侧链 ΔGt,异丙基侧链=ΔGt,Val-ΔGt,Gly")
20
具有大的正的ΔGt的AA侧链 疏水性强 优先选择处在有机相 倾向于配置在蛋白质分子的内部 具有负的ΔGt的AA侧链 亲水性的 配置在蛋白质分子的表面

21
Trp、Tyr和Phe在近紫外区(250-300 nm)吸收光。Trp和Tyr在紫外区还显示荧光。
(五)氨基酸的光学性质 Trp、Tyr和Phe在近紫外区( nm)吸收光。Trp和Tyr在紫外区还显示荧光。
氨基酸的光学性质. Trp、Tyr和Phe在近紫外区( nm)吸收光。Trp和Tyr在紫外区还显示荧光。")
22
二、氨基酸的化学反应性 存在于游离氨基酸和蛋白质分子中的反应基团 氨基、羧基、巯基、酚羟基、羟基、硫醚基(Met)、咪唑基和胍基
、咪唑基和胍基")
23
与茚三酮反应 常用来定量游离氨基酸。 1摩尔的氨基酸,产生1摩尔的氨、醛、CO2和二氢化茚亭。释放出的氨随即与1摩尔茚三酮和1摩尔二氢化茚亭反应生成一种被称为Ruhemann’s紫的紫色物质,后者在570 nm显示最高吸收。 脯氨酸和羟基脯氨酸产生一种黄色物质,它在440 nm显示最高吸收。

24
与邻-苯二甲醛反应 当存在2-巯基乙醇时氨基酸与邻-苯二甲醛反应(1,2-苯二甲醛)反应生成高荧光的衍生物,它在380 nm激发时在450 nm具有最高荧光发射。
反应生成高荧光的衍生物,它在380 nm激发时在450 nm具有最高荧光发射。")
25
与荧光胺反应 含有伯胺的氨基酸、肽和蛋白质与荧光胺反应成高荧光的衍生物,它在390 nm激发时,在475 nm具有最高荧光发射。此法能被用于定量氨基酸以及蛋白质和肽。

26
第三节 蛋白质的结构 一、蛋白质的结构水平 (一)一级结构 一级结构 指氨基酸在蛋白质分子中按一定顺序以肽键连接形成多肽链。
第三节 蛋白质的结构 一、蛋白质的结构水平 (一)一级结构 一级结构 指氨基酸在蛋白质分子中按一定顺序以肽键连接形成多肽链。 构成蛋白质的主要氨基酸有20种。
一级结构. 一级结构 指氨基酸在蛋白质分子中按一定顺序以肽键连接形成多肽链。 构成蛋白质的主要氨基酸有20种。")
27
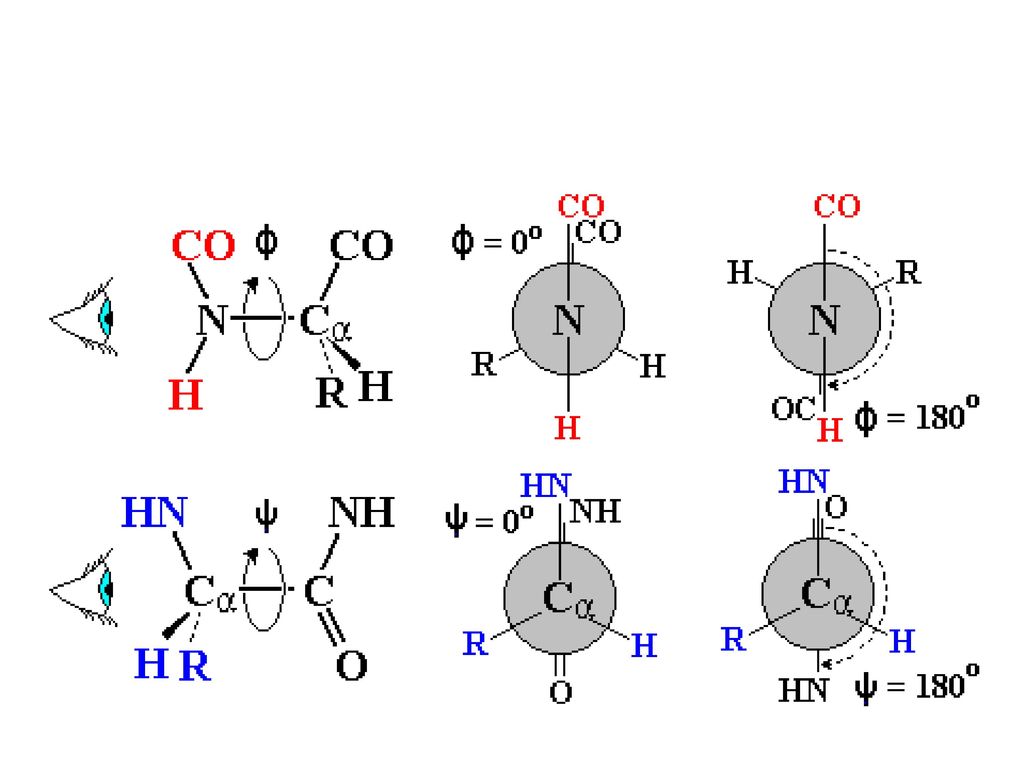
C—N键具有部分双键的性质 C—O键仅有60%的双键性质而C—N键亦有40%的双键性质.
N-Cα和Cα-C键具有转动自由度,分别被定义为φ和ψ两面角,也称为主链扭转角。 理论上φ和ψ具有360度转动自由度,但实际转动自由度由于Cα原子上侧链原子的立体位阻而被限制.

28
多肽链主链基本上可被描述为通过Cα原子连结的一系列 -Cα-CO-NH-Cα-平面

30
几乎所有的蛋白质肽键都以反式构型存在在热力学上反式比顺式稳定

31
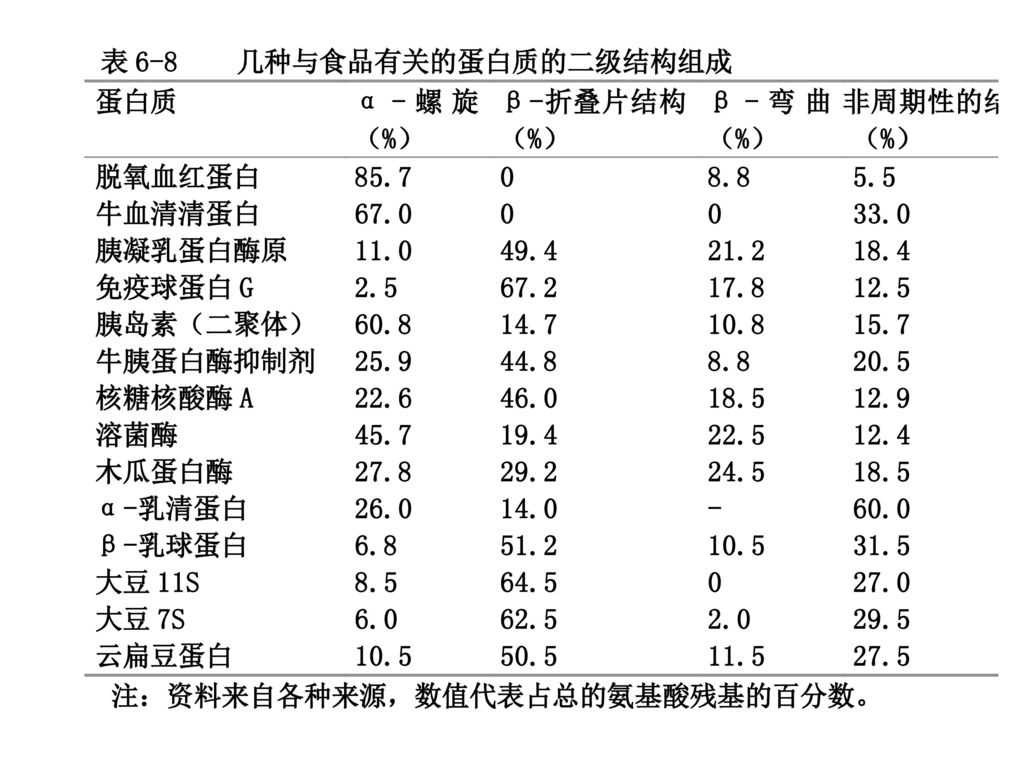
(二)二级结构 蛋白质的二级结构 多肽链的某些部分的氨基酸残基周期性的(有规则的)空间排列,即沿着肽链轴所采取的特有的空间结构。
蛋白质的二级结构 多肽链的某些部分的氨基酸残基周期性的(有规则的)空间排列,即沿着肽链轴所采取的特有的空间结构。 连续的氨基酸残基采取同一套φ和ψ扭转角时就形成周期性的结构。氨基酸残基侧链之间近邻或短距离的非共价相互作用导致局部自由能下降,这就驱动了φ和ψ角的扭转。 非周期性或随机结构是指连续的氨基酸残基具有不同套的φ和ψ扭转角所形成的区域。
空间排列,即沿着肽链轴所采取的特有的空间结构。 连续的氨基酸残基采取同一套φ和ψ扭转角时就形成周期性的结构。氨基酸残基侧链之间近邻或短距离的非共价相互作用导致局部自由能下降,这就驱动了φ和ψ角的扭转。 非周期性或随机结构是指连续的氨基酸残基具有不同套的φ和ψ扭转角所形成的区域。")
32
1. 螺旋结构 连续的氨基酸残基的φ和ψ角按同一套值扭转。 通过选择不同的φ角和ψ角组合,理论上可能产生几种几何形状的螺旋结构。
在蛋白质分子中仅存在三种螺旋结构,即α-螺旋、310-螺旋和π-螺旋。

33
α-螺旋 每圈包含3.6个AA残基。每一个AA转角为100˚。侧链按垂直于螺旋的轴的方向定向。
依靠氢键稳定。每一个残基的N-H基与前面第四个残基的C=O基形成氢键,在此氢键圈中包含13个主链原子;于是α-螺旋有时也被称为3.613螺旋。氢键平行于螺旋轴而定向。氢键的N、H和O原子几乎处在一条直线上。 α-螺旋能以右手和左手螺旋两种方式存在,右手螺旋较稳定。 天然蛋白质中只存在右手α-螺旋。

34
2. β-折叠片结构 一种据齿形结构。它比α螺旋较为伸展

35
主链上的氨基酸残基的侧链按垂直于片状结构的平面定 向。
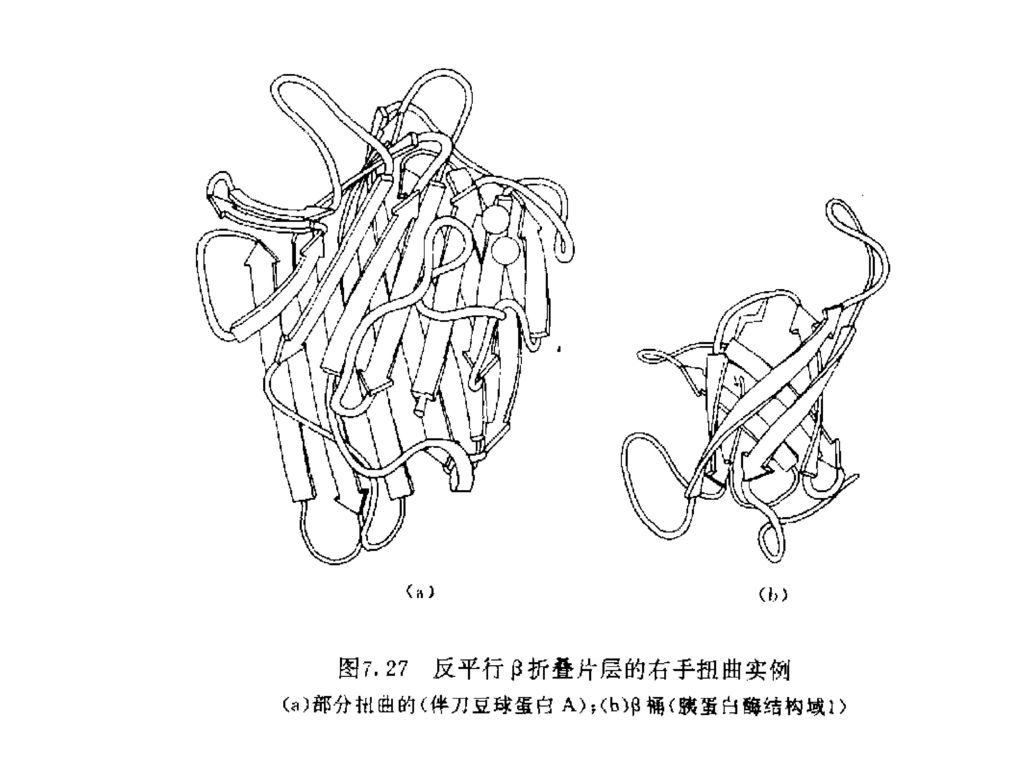
氢键只可能在多肽链的两个片断之间形成。 在反平行β-折叠片结构中,N-H…O原子处在一条直线 上(0氢键角),增加了氢键的稳定性。反平行β-折叠 片结构比平行β-折叠片结构更为稳定。
,增加了氢键的稳定性。反平行β-折叠 片结构比平行β-折叠片结构更为稳定。")
36
含有高比例的β-折叠片结构的蛋白质一般呈现高变性温度。
当α-螺旋类型的蛋白质溶液经加热和冷却时,α-螺旋结构通常转变成β-折叠片结构。β-折叠片结构不会转变成α-螺旋结构。

37
β-弯曲(β-bend)或β-旋转(β-turn)
β-折叠片结构中多肽链反转180°就形成β-旋转。 一个β-旋转结构包含4个氨基酸残基,此结构由氢键所稳定。最常见于β-旋转结构中的氨基酸残基是Asp、Cys、Asn、Gly、Tyr和Pro。

38
Beta turn

40
(三)三级结构
三级结构")
41
三级结构的形成包括在蛋白质中各种不同的基团之间相互作用(疏水、静电和范德华尔)和氢键的优化,使得蛋白质分子的自由能尽可能地降至最低。
一级结构决定着蛋白质分子的形状。含有大量的并均匀地分布在氨基酸序列中的亲水性氨基酸残基的蛋白质分子将呈拉长或棒状。含有大量的疏水性氨基酸残基的蛋白质分子将呈球状。

44
(四)四级结构 蛋白质的四级结构 蛋白质亚基非共价结合的结果。 稳定四级结构的键或力除了二硫键外和稳定三级结构的是相同的。
四级结构 蛋白质的四级结构 蛋白质亚基非共价结合的结果。 稳定四级结构的键或力除了二硫键外和稳定三级结构的是相同的。")
46
(五)蛋白质结构中的相互作用和键 ---稳定蛋白质结构的作用力
(五)蛋白质结构中的相互作用和键 ---稳定蛋白质结构的作用力 蛋白质的天然构象是一种热力学状态,在此状态有利的相互作用达到最大,不利的相互作用降到最小。 蛋白质的稳定性 使蛋白质分子能达到和保持它们的天然构象的一种状态,但这并不排除生理功能所必需的构象的调整。 影响蛋白质折叠的作用力包括两类: (a)蛋白质分子固有的作用力所形成的分子内相互作用。范德华相互作用(Van der Waals interaction)和空间相互作用(steric interaction) (b)受周围溶剂影响的分子内相互作用。氢键、静电相互作用和疏水相互作用。
蛋白质结构中的相互作用和键 ---稳定蛋白质结构的作用力. 蛋白质的天然构象是一种热力学状态,在此状态有利的相互作用达到最大,不利的相互作用降到最小。 蛋白质的稳定性 使蛋白质分子能达到和保持它们的天然构象的一种状态,但这并不排除生理功能所必需的构象的调整。 影响蛋白质折叠的作用力包括两类: (a)蛋白质分子固有的作用力所形成的分子内相互作用。范德华相互作用(Van der Waals interaction)和空间相互作用(steric interaction) (b)受周围溶剂影响的分子内相互作用。氢键、静电相互作用和疏水相互作用。")
47
1. 空间相互作用 大多数氨基酸残基含有庞大的侧链,某些扭角是不可能存在的。
理论上φ和ψ角在具有360°的转动自由度,实际上由于侧链原子的空间位阻而使它们的转动受到很大的限制。 2. Van der Waals 相互作用 存在于原子之间。原子间距离较大,相互作用小;距离减小,吸引力逐渐增大;如果距离进一步减小,斥力将增大。 中性原子之间偶极-诱导偶极和诱导偶极-诱导偶极相互作用。吸引和排斥 吸引能6次方反比于相互作用的原子间的距离,而排斥相互作用7次方反比于该距离。 范德华尔相互作用对于蛋白质的折叠和稳定性的贡献是很显著的。

48
3. 静电相互作用 蛋白质含有一些带有可离解基团的氨基酸残基。 在中性pH,Asp和Glu残基带负电荷,而Lys、Arg和His带正电荷;在碱性pH,Cys和Tyr残基带负电荷。在中性pH,蛋白质分子带净的负电荷或净的正电荷,这取决于分子中负电荷和正电荷残基的相对数目。 蛋白质分子净电荷为0时的pH被定义为蛋白质的等电点(pI)。 等离子点是指不存在电解质时蛋白质溶液的pH。
。 等离子点是指不存在电解质时蛋白质溶液的pH。")
49
除少数例外,蛋白质中几乎所有的带电基团都分布在分子的表面。
排斥作用或许会导致蛋白质结构不稳定。吸引作用有助于蛋白质结构的稳定。 处在蛋白质分子表面的带电基团对蛋白质结构的稳定性没有重要的贡献。 水溶液中水的介电常数高,引力和排斥力均小。 处在蛋白质分子内部的带电基团对蛋白质结构的稳定性有重要的影响 环境的介电常数比水的小,引力和排斥力均大。

50
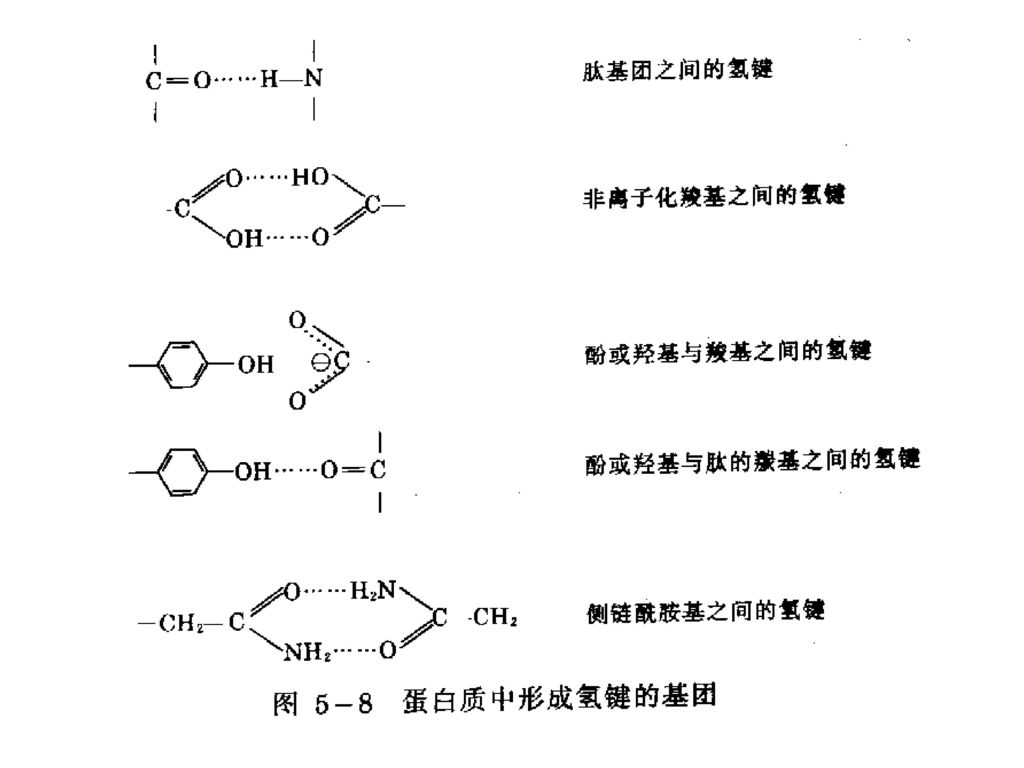
4. 氢键 主要通过羰基及亚氨基通过氢氧而形成的一种键。
蛋白质含有一些能形成氢键的基团。可在两肽链间,或一条肽链的不同部位形成氢键。 在α-螺旋和β-折叠片结构中肽键的N-H和C=O之间形成了最大数目的氢键。 氢键对于维持蛋白质二级结构,保持蛋白质稳定性,起着重要的作用。

52
5.疏水相互作用 在水溶液中,非极性基团之间的疏水相互作用是水与非极性基团之间热力学上不利的相互作用的结果。 在水溶液中水结构诱导的非极性基团之间的相互作用被称为疏水相互作用。 在蛋白质中,氨基酸残基非极性侧链之间的疏水相互作用是蛋白质折叠成独特的三维结构的主要原因,

53
6. 二硫交联 二硫键(-S-S-) 两个硫原子间的化学键 由二个半胱氨酸分子通过脱氢氧化而结合成的一种化学键。键能很大( 千卡/ 摩尔),对稳定蛋白质空间结构起着重要作用。二硫键数目多、稳定,如毛、发。 天然存在于蛋白质中的唯一的共价侧链交联。既能存在于分子内,也能存在于分子间。 在单体蛋白质中二硫键的形成是蛋白质折叠的结果。二硫键一旦形成就能帮助稳定蛋白质的折叠结构。

54
7. 金属离子 一些蛋白质结合特定的金属离子,如钙,镁和钠离子等。这种结合能稳定蛋白质的结构 稳定的机制或许是通过中和电荷的效应和促进其他相互作用(疏水相互作用使蛋白质的分子构型更牢固)。 α-淀粉酶蛋白 钙稳定酶蛋白的构象。 牛乳清蛋白天然结构稳定的必需条件是结合钙离子。 总之,一个独特的三维蛋白质结构的形成是各种排斥和吸引的非共价相互作用以及几个共价二硫键的净结果。

55
蛋白质变性的一般概念 第四节 蛋白质的变性 一、概述
第四节 蛋白质的变性 一、概述 蛋白质变性的一般概念 外界因素的作用使构成空间结构的氢键等副键遭受破坏,导致蛋白质的二级,三级和四级结构的变化,天然蛋白质的空间构型则解体,有秩序的螺旋型,球状构型变为无秩序的伸展肽链,使天然蛋白质的理化性质改变并失去原来的生理活性,这种作用称为蛋白质的变性作用。 在某些情况下,变性过程是可逆的,例如,有的蛋白质在加热时发生变性,冷却后,又可复原。 可逆变性一般只涉及蛋白质分子的三级和四级结构变化, 不可逆变性能使二级结构也发生变化。 如果二硫键起着稳定蛋白质构象的作用,那么它的断裂往往导致不可逆的变性。

56
(1)因疏水性基团的暴露而导致溶解度的下降
蛋白质变性后其物理化学性质变化表现为: (1)因疏水性基团的暴露而导致溶解度的下降 (2)结合水能力的改变 (3)失去生物活力(酶活力或免疫活力) (4)对蛋白酶的敏感性(肽键暴露) (5)蛋白质固有粘度增加 (6)没有结晶能力
因疏水性基团的暴露而导致溶解度的下降. (2)结合水能力的改变. (3)失去生物活力(酶活力或免疫活力) (4)对蛋白酶的敏感性(肽键暴露) (5)蛋白质固有粘度增加. (6)没有结晶能力.")
57
引起蛋白质变性的因素 物理因素: 温度、紫外线、超声波、高压等。 化学因素: 酸、碱、有机溶剂、重金属盐类、脲、胍、表面活性剂等。

58
二、引起蛋白质变性的因素 (一)物理因素 1. 热 最常见的变性现象,热变性。 热变性所需温度与蛋白质本质、纯度和pH值有关。 蛋白质受热变性的机理 在较高温度下,保持蛋白质空间构象的那些副键断裂,破坏了肽链的特定排列,原来在分子内部的一些非极性基团暴露到了分子的表面,因而降低了蛋白质的溶解度,促进了蛋白质分子之间相互结合而凝集,形成不可逆的凝胶而凝固。

59
氢键、静电和范德华尔相互作用具有放热的性质。它们在高温下去稳定而在低温下稳定。
疏水相互作用是吸热的。它们在高温下稳定,而在低温下去稳定。然而,疏水相互作用的稳定性也不会随温度的提高而无限制地增强疏水相互作用的强度在60-70℃时达到最高。 构象熵:-TSconf

60
一般认为,温度愈低蛋白质的热稳定性愈高,但并非总是如此。

61
极性和非极性相互作用对蛋白质稳定性的影响
蛋白质的分子中极性相互作用超过非极性相互作用,蛋白质在冻结温度和低于冻结温度时比较高温度时更稳定。 蛋白质的稳定主要依靠疏水相互作用时,蛋白质在室温时比在冻结温度时更稳定。

62
含有较高比例的疏水性氨基酸残基(尤其是Val、Ile、Leu和Phe)的蛋白质比亲水性较强的蛋白质一般更为稳定。
蛋白质的平均疏水性和热变性温度之间的正相关只是一个近似,二硫键、盐桥等对蛋白质的稳定性也有贡献。

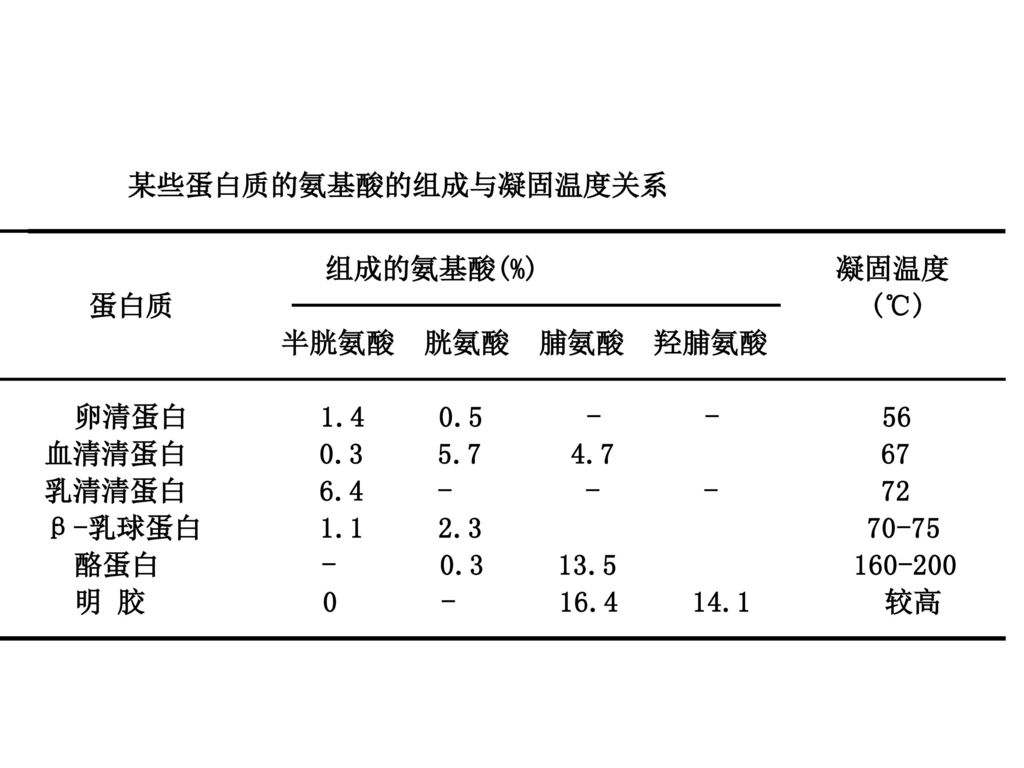
63
影响热变性的因素 (1)组成蛋白质的氨基酸的种类 蛋白质分子中-SH 的含量与变性蛋白质在水中凝固作用成正比。如大豆蛋白质(含硫氨基酸较少) 脯氨酸或羟脯氨酸能阻碍蛋白质分子彼此形成交联,使蛋白质不易凝固。如醇溶谷蛋白(含脯及羟脯氨酸较多) 。
 脯氨酸或羟脯氨酸能阻碍蛋白质分子彼此形成交联,使蛋白质不易凝固。如醇溶谷蛋白(含脯及羟脯氨酸较多) 。 .")
64
(2)温度 一般是在50-70℃之间 蛋白质凝固的温度因其种类而不同, 容易凝固的蛋白质有可溶性清蛋白和球蛋白。如牛乳中含有酪蛋白,有少许乳清蛋白,所以在一般情况下,牛乳不易凝。
温度 一般是在50-70℃之间 蛋白质凝固的温度因其种类而不同, 容易凝固的蛋白质有可溶性清蛋白和球蛋白。如牛乳中含有酪蛋白,有少许乳清蛋白,所以在一般情况下,牛乳不易凝。")
66
(3)水 水分愈少所需变性温度愈高。 水能显著地促进蛋白质的热变性。干蛋白质粉对热变性是非常稳定的。 在干燥状态,蛋白质具有静止的结构,多肽链段的移动受到限制。水分含量的增加,水合作用和水部分地穿透至蛋白质结构的空洞的表面导致蛋白质的肿胀。蛋白质的肿胀提高了多肽链的移动性和柔性,使蛋白质分子可以采取动力学上更为熔融的结构,当加热时,此动力学上柔性结构比起干燥状态能提供给水更多的机会接近盐桥和肽的氢键,于是造成较低的Td值。

68
(4)电解质 蛋白质凝固温度因电解质的存在而降低,其反应速度则增加,化合价大的离子易使蛋白质凝固。例如制造豆腐时,豆浆中的球蛋白仅加热是不会凝固的,但在70℃以上添加氯化镁或硫酸钙即可凝固。 (5)氢离子浓度 蛋白质凝固温度,亦受氢离子浓度的影响,一般在等电点范围内,最易凝固(分子间容易靠近)。加酸过多至pH4.8以下,则凝固温度上升,甚至不会凝固。
。加酸过多至pH4.8以下,则凝固温度上升,甚至不会凝固。")
69
低温能导致一些蛋白质的变性。 2. 冷 一些在室温下稳定的酶在0℃时变得较不稳定。 一些蛋白质在低温时聚集或沉淀。
高疏水性-极性氨基酸比例的组成及结构决定疏水相互作用,易低温变性。

70
3. 机械处理 揉搓、振动、打擦等操作产生的机械剪切可能使蛋白质变性。揉搓或滚压产生剪切力可打破α螺旋。 当一个转动的叶片产生高剪切时,造成亚音速的脉冲,在叶片的尾随边缘也出现空化,导致蛋白质变性。剪切速度愈高,蛋白质变性程度愈高。 高温和高剪切力相结合能导致蛋白质不可逆的变性。

71
4. 压力 压力诱导蛋白质变性主要是因为蛋白质是柔性的和可压缩的。球状蛋白质分子结构的内部有一些空穴仍然存在,这使得蛋白质分子结构具有可压缩性。 压力诱导的蛋白质变性是高度可逆的。 大多数纤维蛋白无空穴,故其对静水压的稳定性较高。 大多数蛋白质在100~1200 Mpa压力下诱导变性。 压力诱导转变的中点出现在400~800 Mpa。 高压处理:灭菌和蛋白质的凝胶。

72
5. 辐照 芳香族氨基酸残基吸收紫外线,若能量高,能打断二硫键。其它射线能起氧化或打破共价键作用。 6. 界面 当分子吸附在水和空气、水和非水液体或水和固体的界面时,通常会导致不可逆变性。

73
(二)引起蛋白质变性的化学因素 1. pH 常温下,大多数蛋白质仅在pH4-10的范围内是稳定的。 酸碱引起蛋白质变性的机理 蛋白质溶液pH值的改变导致多肽链中某些基团发生解离,从而破坏了静电作用形成的键。 蛋白质在等电点时比在任何其他pH值时对变性作用更加稳定(自身不容易展开) 在中性pH附近,净负电推斥能量小于其他稳定蛋白质的相互作用的能量,因此大多数蛋白质是稳定的。 在极端pH值,高静电荷引起的强烈的分子内电推斥力导致蛋白质分子的肿胀和展开。
 在中性pH附近,净负电推斥能量小于其他稳定蛋白质的相互作用的能量,因此大多数蛋白质是稳定的。 在极端pH值,高静电荷引起的强烈的分子内电推斥力导致蛋白质分子的肿胀和展开。")
74
pH诱导的蛋白质变性多数是可逆的 然而,如果肽键的部分水解,Asn和Gln的脱酰胺,巯基的破坏,或者聚集作用能导致蛋白质的不可逆变性。
酸和碱可以加速热变性。 一般水果罐头钉杀菌的温度较蔬菜低。

75
2. 金属 碱金属作用有限,碱土金属具有较高的能力,过渡金属容易反应,其中许多能形成络合物。用螯合剂除去这些金属离子,降低蛋白质对热和蛋白酶的稳定性。 3. 有机溶剂 有机溶剂以不同的方式影响蛋白质的疏水相互作用、氢键和静电相互作用。 在蛋白质水溶液中加入大量的有机溶剂,能引起蛋白质的变性作用主要是影响蛋白质的疏水相互作用 非极性侧链在有机溶剂中比在水中更易溶解,因此会削弱疏水相互作用。

76
4. 有机化合物的水溶液 (1)一些有机化合物,如脲素和盐酸胍(GuHCl)的水溶液,能打断氢键导致蛋白质不同程度的变性。这些物质通过提高疏水氨基酸残基在水相的浓度,也降低了疏水相互作用。 (2)表面活性剂是强有力的变性剂。其作用如同蛋白质疏水区和亲水环境的媒介物,打断了疏水相互作用。
一些有机化合物,如脲素和盐酸胍(GuHCl)的水溶液,能打断氢键导致蛋白质不同程度的变性。这些物质通过提高疏水氨基酸残基在水相的浓度,也降低了疏水相互作用。 (2)表面活性剂是强有力的变性剂。其作用如同蛋白质疏水区和亲水环境的媒介物,打断了疏水相互作用。 .")
77
5. 促溶盐(chaotropic salts)和变性
盐以两种不同的方式影响蛋白质的稳定性。 在低浓度时,离子通过非特异性的静电相互作用与蛋白质作用,此类蛋白质电荷的静电中和一般稳定了蛋白质的结构。完全的电荷中和出现在离子强度等于或低于0.2,与盐的性质无关。 在较高的浓度(>1M),盐具有影响蛋白质结构稳定性的离子特异效应,像Na2SO4和NaF这样的盐能促进蛋白质结构的稳定性,而NaSCN和NaClO4的作用相反。
,盐具有影响蛋白质结构稳定性的离子特异效应,像Na2SO4和NaF这样的盐能促进蛋白质结构的稳定性,而NaSCN和NaClO4的作用相反。")
78
阴离子对蛋白质结构的影响甚于阳离子 图 6-17 各种钠盐对处在pH7.0下β-乳球蛋白变性温度的影响。 Δ-NaCl,□-NaBr, ●-NaClO4,▲-NaSCN,■-脲素, o –Na2SO4

79
在等离子强度下各种阴离子影响蛋白质结构稳定性的能力一般遵循下面顺序(Hofmeister系列或促溶)系列。
F-<SO42-<Cl-<Br-<I-<ClO4-<SCN-<Cl3CCOO-。 氟化物、氯化物和硫酸盐是结构稳定剂,而其它阴离子盐是结构去稳定剂。NaSCN和NaClO4是强变性剂。 高浓度的盐总是对蛋白质的结构稳定性产生不利的影响。

80
稳定蛋白质的盐能促进蛋白质的水合作用并与蛋白质微弱地结合。
使蛋白质不稳定的盐能降低蛋白质的水合作用并与蛋白质强烈地结合。

81
(六)还原剂 还原剂能还原蛋白质分子中的二硫键交联,因而改变了蛋白质的构象。
还原剂 还原剂能还原蛋白质分子中的二硫键交联,因而改变了蛋白质的构象。")
82
第五节 蛋白质的功能性质 蛋白质对食品的感官品质具有重要的影响 蛋白质的功能性质指的是能使蛋白质对人们所期望的食品特征作出贡献的那些物理化学性质。 食品蛋白质的“功能性质”(“Functionality”)的定义:在食品加工、保藏、制备和消费期间影响蛋白质在食品体系中的性能的那些蛋白质的物理和化学性质。
的定义:在食品加工、保藏、制备和消费期间影响蛋白质在食品体系中的性能的那些蛋白质的物理和化学性质。")
83
表5-12 食品蛋白质在食品体系中的功能作用 功能 机制 食品 蛋白质种类 溶解性 亲水性 饮料 乳清蛋白 粘度
水结合、流体动力学分子大小和形状 汤、肉汁、色拉调味料和甜食 明胶 水结合 氢键、离子水合 肉、香肠、蛋糕和面包 肌肉蛋白质、鸡蛋蛋白质 凝胶作用 水截留和固定、网状结构形成 肉、凝胶、蛋糕、焙烤食品和奶酪 肌肉蛋白质、鸡蛋和乳蛋白质 粘结—粘合 疏水结合、离子结合和氢键 肉、香肠、面条和焙烤食品 肌肉蛋白质、鸡蛋蛋白质和乳清蛋白质 弹性 疏水结合和二硫交联 肉和焙烤食品 肌肉蛋白质和谷物蛋白质 乳化 在界面上吸附和形成膜 香肠、大红肠、汤、蛋糕和调味料 肌肉蛋白质、鸡蛋蛋白质和乳蛋白质 起泡 界面吸附和形成膜 搅打起泡的浇头、冰淇淋、蛋糕和甜食 鸡蛋蛋白质和乳蛋白质 脂肪和风味物的结合 疏水结合或截留 低脂焙烤食品和油炸面包圈 乳蛋白质、鸡蛋蛋白质和谷物蛋白质

84
食品蛋白质的功能性质可以分成三个主要类别:
(1) 水化性质(取决于蛋白质-水的相互作用) 包括水吸收及保留、湿润性、肿胀、粘着性、分散性、溶解度和粘度。 (2)与蛋白质-蛋白质相互作用有关的性质 包括沉淀作用、凝胶作用和形成各种其它结构(蛋白质面团和纤维)。 (3)表面性质 主要关系到蛋白质的表面张力、乳化作用和泡沫特征。 上述这几类性质不是完全独立的。
 水化性质(取决于蛋白质-水的相互作用) 包括水吸收及保留、湿润性、肿胀、粘着性、分散性、溶解度和粘度。 (2)与蛋白质-蛋白质相互作用有关的性质. 包括沉淀作用、凝胶作用和形成各种其它结构(蛋白质面团和纤维)。 (3)表面性质. 主要关系到蛋白质的表面张力、乳化作用和泡沫特征。 上述这几类性质不是完全独立的。")
85
食品的流变和质构性质取决于水与其他食品组分,尤其像蛋白质和多糖那样的大分子的相互作用。
一、蛋白质的水合 食品的流变和质构性质取决于水与其他食品组分,尤其像蛋白质和多糖那样的大分子的相互作用。 1. 取决于水-蛋白质相互作用的蛋白质的功能性质: 分散性、湿润性、肿胀、溶解性、增稠、粘度、持水能力、胶凝作用、凝结、乳化和起泡。

86
1 水与蛋白质分子一些基团的结合,通过: 离子(蛋白质分子中的带电基团)-偶极相互作用 偶极(蛋白质分子中的极性基团)-偶极相互作用
偶极-诱导偶极(蛋白质分子中的非极性基团)相互作用 疏水(蛋白质分子中的非极性基团)相互作用 2. 蛋白质结合水的能力(水合能力) 定义 当干蛋白质粉与相对湿度为90-95%的水蒸汽达到平衡时,每克蛋白质所结合的水的克数被定义为蛋白质结合水的能力。
相互作用. 疏水(蛋白质分子中的非极性基团)相互作用 2. 蛋白质结合水的能力(水合能力) 定义 当干蛋白质粉与相对湿度为90-95%的水蒸汽达到平衡时,每克蛋白质所结合的水的克数被定义为蛋白质结合水的能力。")
87
蛋白质的水合能力部分地与它的氨基酸组成有关,带电的氨基酸残基数目愈大,水合能力愈大。
带电基团:6 mol水/mol残基 不带电的极性残基:2 mol /mol残基。 非极性残基:1 mol /mol残基。 a:水合能力,g水/g蛋白质; fc, fp, fN:分别代表蛋白质分子中带电的、极性的和非极性残基所占的分数。

88
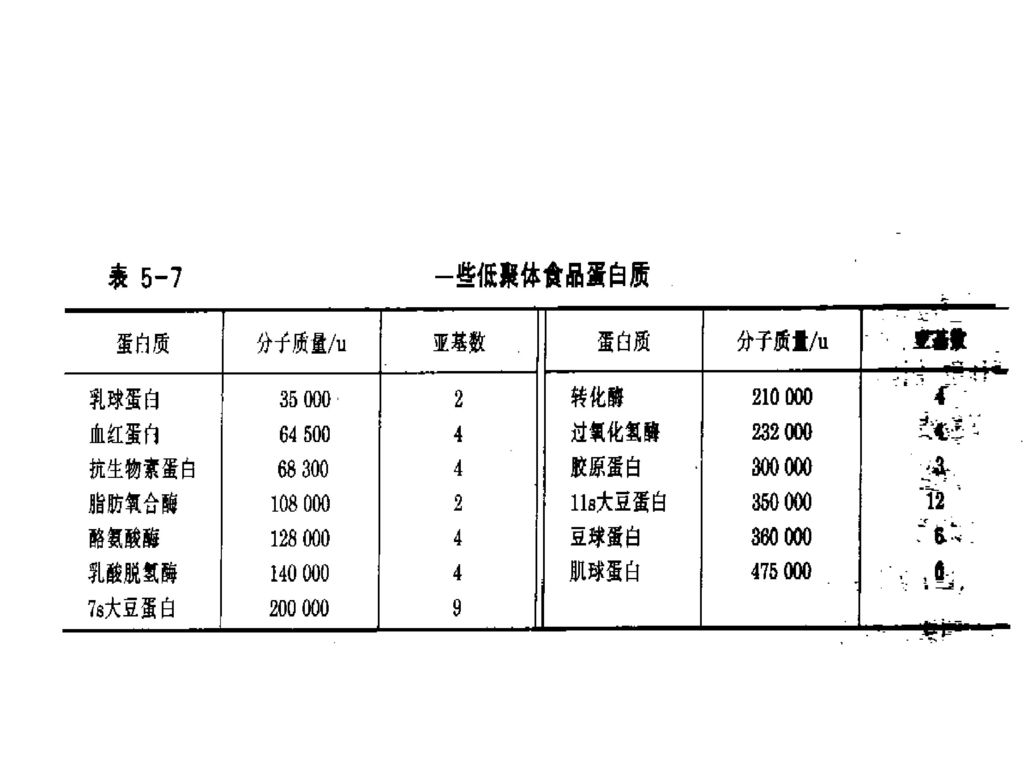
表5-14 各种蛋白质的水合能力 蛋白质 水合能力(g H2O/g 蛋白质) 纯蛋白质 商业蛋白质产品 核糖核酸酶 0.53 乳清浓缩蛋白
表5-14 各种蛋白质的水合能力 蛋白质 水合能力(g H2O/g 蛋白质) 纯蛋白质 商业蛋白质产品 核糖核酸酶 0.53 乳清浓缩蛋白 溶菌酶 0.34 酪蛋白酸钠 肌红蛋白 0.44 大豆蛋白 0.33 β-乳球蛋白 0.54 胰凝乳蛋白酶原 0.23 血清清蛋白 血红蛋白 0.62 胶原蛋白 0.45 酪蛋白 0.40 卵清蛋白 0.30
 纯蛋白质. 商业蛋白质产品. 核糖核酸酶 乳清浓缩蛋白 溶菌酶 酪蛋白酸钠 肌红蛋白 大豆蛋白 β-乳球蛋白 胰凝乳蛋白酶原 血清清蛋白. 血红蛋白 胶原蛋白 酪蛋白 卵清蛋白")
89
结合而被吸附 3. 蛋白质的水化过程 (1) (2) 干蛋白质 水分子通过与极性部位 多层水吸附 液态水凝聚(3) 肿胀(4)
3. 蛋白质的水化过程 (1) (2) 干蛋白质 水分子通过与极性部位 多层水吸附 结合而被吸附 液态水凝聚(3) 肿胀(4) 溶剂分散(5) 肿胀的不溶性粒子或块 溶液
 (2) 干蛋白质 水分子通过与极性部位 多层水吸附. 结合而被吸附. 液态水凝聚(3) 肿胀(4) 溶剂分散(5) 肿胀的不溶性粒子或块. 溶液.")
91
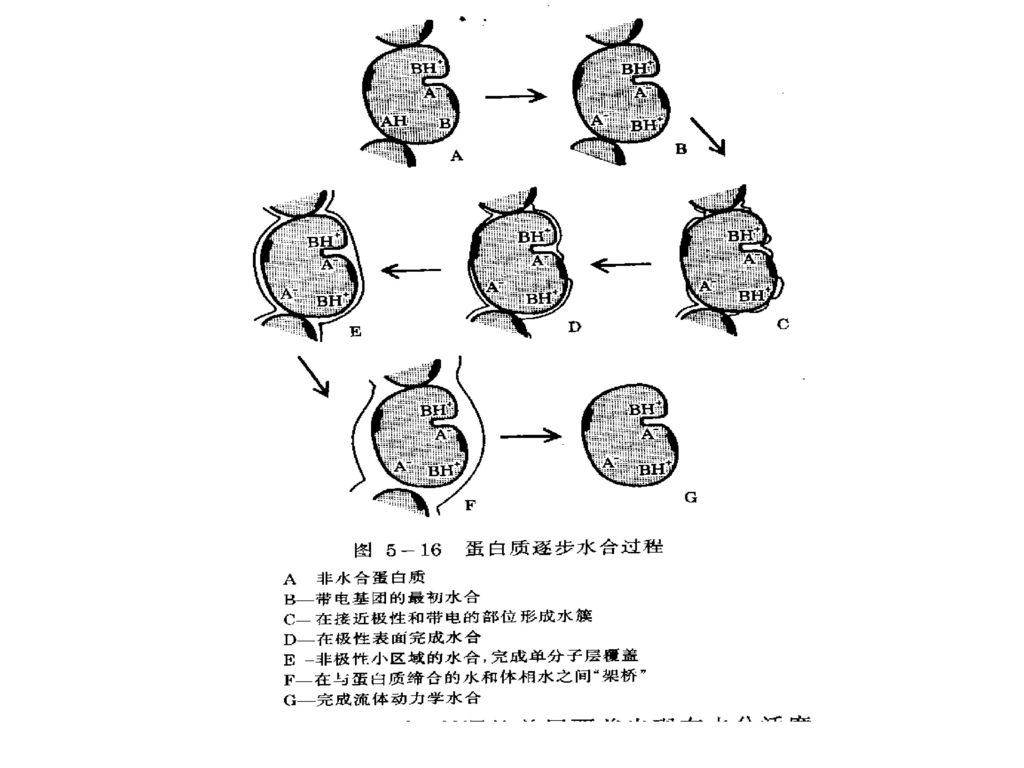
4. 蛋白质的吸着等温线 蛋白质的吸附等温线为S形曲线。 单层覆盖出现在水分活度(aW) 。 在水分活度 范围形成多层水。 在aw=0.9时,蛋白质结合约 g H2O/g 蛋白质。多数在0℃时不能冻结。 当aw>0.9时,液态(大量)水凝聚在蛋白质分子结构的裂缝中或不溶性蛋白质的毛细管中。这部分水的性质类似于体相水,被称为流体动力学水,随蛋白质分子一起运动。
水凝聚在蛋白质分子结构的裂缝中或不溶性蛋白质的毛细管中。这部分水的性质类似于体相水,被称为流体动力学水,随蛋白质分子一起运动。")
92
5. 影响蛋白质结合水能力的环境因素 (1) pH pH会影响蛋白质分子的离子化作用和净电荷数值,从而改变蛋白质分子间的吸引力和推斥力以及蛋白质分子同水结合的能力。 在等电点pH时,蛋白质显示最低的水合作用。 高于或低于等电点pH,由于净电荷和推斥力的增加使蛋白质肿胀和结合较多的水。 大多数蛋白质结合水的能力在pH 9-10时比任何其它pH时来得大,这是由于巯基和酪氨酸残基的离子化;当pH超过10时,赖氨酸残基的ε-氨基上正电荷的失去使蛋白质结合水的能力下降。

93
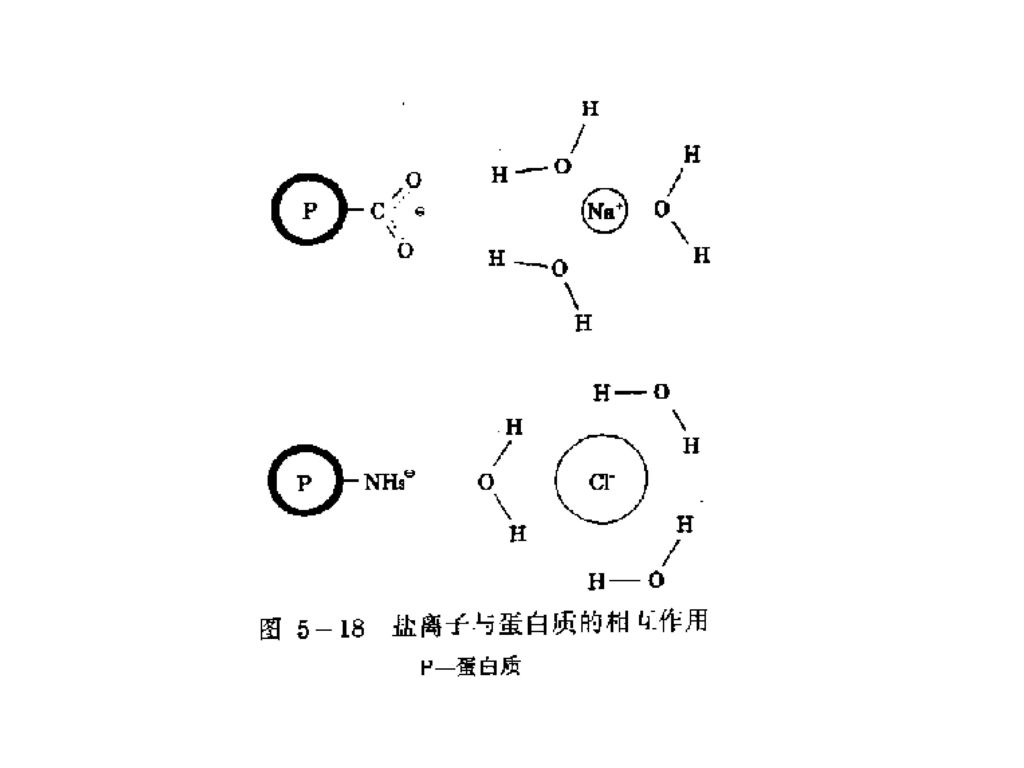
(2) 离子的种类和浓度 离子的种类和浓度对蛋白质的吸收水,肿胀和溶解度具有显著的影响。 低浓度(<0.2M)的盐能提高蛋白质结合水的能力,这是由水合盐离子与蛋白质分子上带电基团微弱地结合所造成的。 高浓度的盐会导致蛋白质的脱水,因为更多的水与盐离子结合。

94
(3) 温度 温度升高后,氢键作用和离子基团的水合作用减弱,蛋白质结合水的能力一般随之下降。 A.加热时,变性和凝聚作用,降低蛋白质的表面积及极性氨基酸的有效性。另一方面,加热时,内部的疏水性氨基酸转向表面。 B. 一些蛋白质,如乳清蛋白,加热时形成凝胶。如将凝胶干燥,蛋白质网中毛细管作用力使蛋白质吸收的能力显著地增加。

95
(4)蛋白质变性 变性蛋白质结合水的能力一般比天然蛋白质约高10%。因为蛋白质变性时,随着一些原来埋藏的疏水基团的暴露,表面积与体积之比增加。 然而,如果变性导致蛋白质聚集,那么蛋白质结合水的能力由于蛋白质—蛋白质相互作用而下降。 注意:蛋白质的溶解性不仅取决于结合水的能力,还取决于其他热力学因素。

96
二、蛋白质的溶解度 最受蛋白质溶解度影响的功能性质 增稠、起泡、乳化和胶凝作用。 高的起始溶解度是其它功能性质的先决条件,不溶性蛋白质在食品中的应用非常有限。 初始溶解的主要优点是它能使蛋白质分子或颗粒快速和广泛地分散,从而形成一个良好分散的胶体体系。 只有具有高溶解度乳清蛋白质和一些其它蛋白质才能在乳状液,泡沫和凝胶中表现出良好的功能性质。

97
蛋白质—蛋白质 + 溶剂—溶剂 蛋白质—溶剂 影响蛋白质溶解性质的主要的相互作用: 疏水相互作用能促进蛋白质—蛋白质相互作用,使蛋白质溶解度降低; 离子相互作用能促进蛋白质—水相互作用,使蛋白质溶解度增加。

98
疏水与离子作用 第一推斥力:带相同电荷离子化残基间; 第二推斥力:蛋白质分子的离子基团的水合壳间的排斥力 Bigelow公式 平均疏水性
电荷频率 平均疏水性小且电荷频率大,则溶解度高

99
描述食品蛋白质的一些术语 l WSP:水溶性蛋白质 l WDP:水可分散蛋白质 l PDI:蛋白质分散性指标 l NSI:氮溶解性指标 PDI和NSI已被确定为美国油脂化学家协会的法定方法:

100
氮溶解度指标(NSI)、溶解度-pH或溶解度-离子强度或溶解度-热处理曲线是食品加工中最进行的试验。
热处理时,大多数蛋白质的溶解度显著地和不可逆地降低。

101
根据溶解度性质的蛋白质分类 (1) 清蛋白,能溶于pH 6.6的水,例如,血清清蛋白、卵清蛋白和α-乳清蛋白都是属于这类蛋白质; (2) 球蛋白,能溶于pH 7.0的稀盐溶液,例如,大豆球蛋白、菜豆球蛋白和β-乳球蛋白都是属于这类蛋白质; (3) 谷蛋白,仅能溶于酸(pH 2)和碱(pH 12)溶液,例如,小麦谷蛋白是属于这类蛋白质; (4) 醇溶谷蛋白,能溶于70%乙醇,玉米醇溶蛋白和麦醇溶蛋白是属于这类蛋白质。 谷蛋白和醇溶谷蛋白是高疏水性蛋白质。
 谷蛋白,仅能溶于酸(pH 2)和碱(pH 12)溶液,例如,小麦谷蛋白是属于这类蛋白质; (4) 醇溶谷蛋白,能溶于70%乙醇,玉米醇溶蛋白和麦醇溶蛋白是属于这类蛋白质。 谷蛋白和醇溶谷蛋白是高疏水性蛋白质。")
102
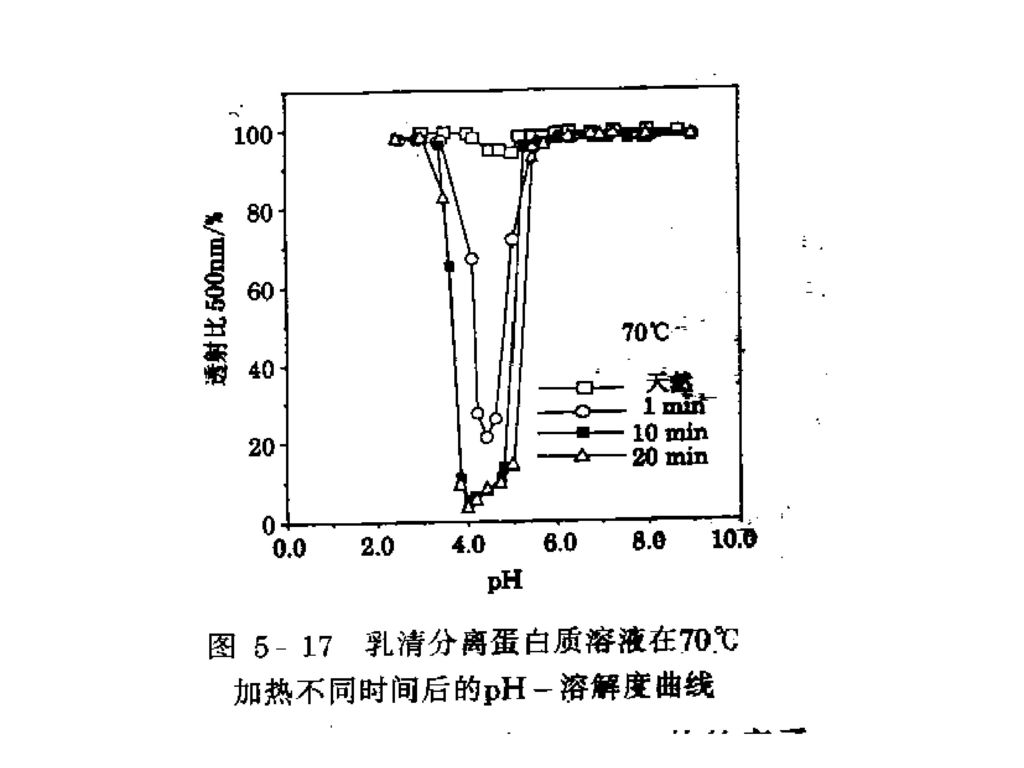
其它影响溶解度的因素: 1.pH 的影响 当pH高于或低于等电点时,蛋白质带净的负电荷或净的正电荷,水分子能同这些电荷相互作用并起着稳定作用。含有相同符号电荷的蛋白质链具有相互排斥、离解和展开的倾向。 大多数食品蛋白质的溶解度-pH是一条U-形曲线,最低溶解度出现在蛋白质的等电点附近 热变性会改变蛋白质的pH—溶解度曲线的形状。

104
2.离子强度的影响 “盐溶” 中性盐的离子在0.1-1M能提高蛋白质的溶解度。 “盐析” 中性盐的离子大于1M,蛋白质的溶解度降低, 并可能导致蛋白质沉淀。

106
蛋白质在盐溶液中的溶解度一般遵循以下关系
S和S0:蛋白质在盐溶液和水中的溶解度 Ks:盐析常数; Cs:盐的摩尔浓度;:常数 Ks:正值 盐析 Ks:负值 盐溶

107
在等电点pH离子强度和离子类型对羧基血红蛋白溶解度的影响

108
3.非水溶剂的影响 能与水互溶的有机溶剂(如乙醇和丙酮)会降低水介质的介电常数,从而提高蛋白质分子内和分子间的静电作用力(推斥和吸引)。 分子内的静电推斥相互作用导致蛋白质分子结构的展开。在此展开状态,介电常数的降低能促进暴露的肽基团之间的分子间氢键的形成和带相反电荷的基团之间的分子间静电相互吸引作用。 这些分子间的极性相互作用导致蛋白质在有机溶剂—水体系中溶解度下降或沉淀。

109
4.温度的影响 0-40 ℃蛋白质的溶解度随温度的提高而提高。 例外。高疏水性蛋白质,如β-酪蛋白和一些谷类蛋白质,溶解度与温度副相关。温度超过40℃时,由于热动能的增加导致蛋白质结构的展开(变性),蛋白质内部的疏水基团暴露,促进蛋白质聚集和沉淀。
,蛋白质内部的疏水基团暴露,促进蛋白质聚集和沉淀。")
110
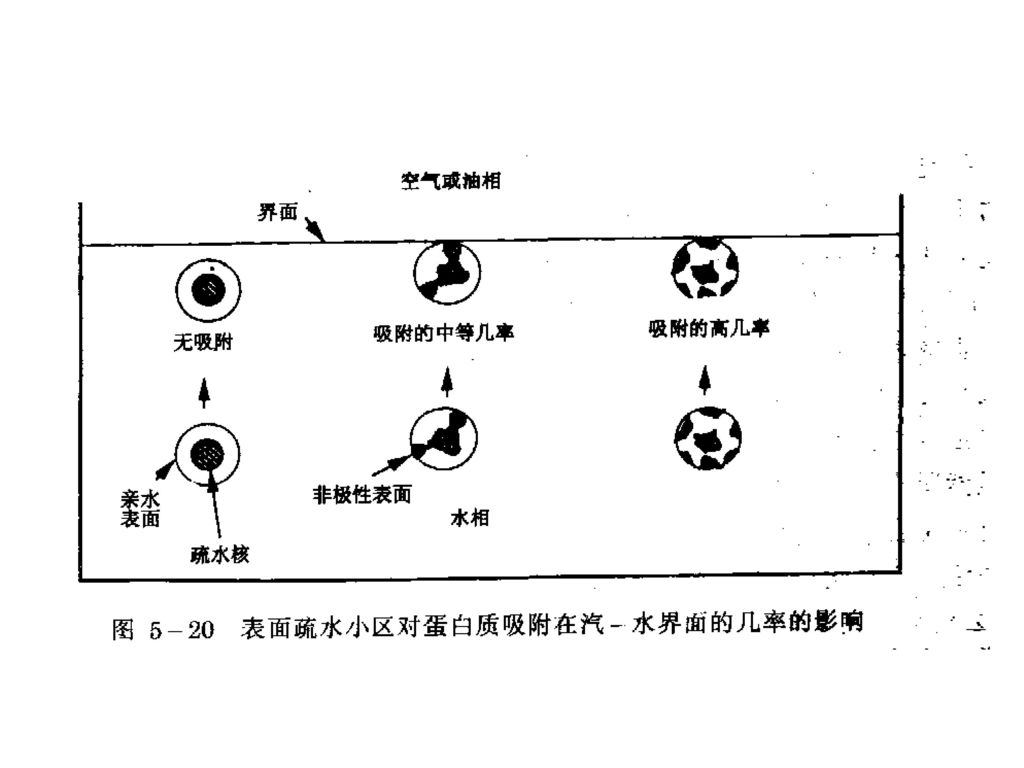
三、蛋白质的界面性质 蛋白质是两亲物质,能自发地迁移至气—水界面或油—水界面。在界面区域的浓度总是高于在体相水相中的浓度。 蛋白质不同于低相对分子质量表面活性剂,它们能在界面形成高粘弹性薄膜(能承受保藏和处理中的机械冲击)。 不同蛋白质在表面活性性质上存在着显著差别。差别主要与它们在构象上的差别有关。 重要的构象因素包括多肽链的稳定性/柔性、对环境改变适应的难易程度和亲水与疏水基团在蛋白质表面的分布模式。

111
最为主要的是蛋白质分子的构象

112
理想的表面活性蛋白质具有三个性能: (1) 能快速地吸附至界面 (2) 能快速地展开并在界面上再定向 (3) 一旦达到界面,能与邻近分子相互作用,形成具有强的粘合和粘弹性质并能忍受热和机械运动的膜。
 能快速地吸附至界面 (2) 能快速地展开并在界面上再定向 (3) 一旦达到界面,能与邻近分子相互作用,形成具有强的粘合和粘弹性质并能忍受热和机械运动的膜。")
114
多肽链在界面上采取的构型: 列车状 当多肽片段直接与界面接触时呈列车状; 圈状 当多肽片段悬浮在水相时呈圈状;
列车状 当多肽片段直接与界面接触时呈列车状; 圈状 当多肽片段悬浮在水相时呈圈状; 尾状 蛋白质分子的N-和C-末端片段通常处在水相呈尾状。

115
三种构象的相对分布取决于蛋白质的构象特征。列车状构象的比例愈大,蛋白质愈是强烈地与界面相结合,并且表面张力愈是低。

116
界面上蛋白质膜的机械强度取决于粘合的分子间的相互作用
静电相互作用 氢键 疏水相互作用 吸附的蛋白质通过-S-S- === 2-SH相互交换形成的界面聚合也能提高膜的粘弹性质。 吸引、推斥和水合作用力之间的适当平衡是形成稳定的粘弹膜的必要条件。

117
(一)乳化性质 蛋白质起着乳化剂的作用。 天然牛乳乳状液是由脂肪球“膜”稳定的,此膜是由依次相继吸附着的三酰基甘油,磷脂,不溶解的脂蛋白和可溶解的蛋白质所构成。 当牛奶被均质时,脂蛋白膜被酪蛋白胶束和乳清蛋白质组成的膜所取代。在抵抗乳状液分层方面均质牛奶比天然牛奶较为稳定,这是因为酪蛋白胶束——乳清蛋白质膜比天然蛋白质膜更强。

118
1. 评价食品乳化性质的方法 l 油滴大小分布 l 乳化活力(emulsifying activity)
1. 评价食品乳化性质的方法 l 油滴大小分布 l 乳化活力(emulsifying activity) l 乳化能力(emulsion capacity) l 乳化稳定性(emulsion stability)
 l 乳化能力(emulsion capacity) l 乳化稳定性(emulsion stability)")
119
(1)油滴大小分布 由蛋白质稳定的乳状液的物理和感官性质取决于所形成的液滴的大小和总界面面积。
(1)油滴大小分布 由蛋白质稳定的乳状液的物理和感官性质取决于所形成的液滴的大小和总界面面积。 测定乳状液平均液滴的大小的方法有: 光学显微镜法(不是非常可靠) 电子显微镜法 光散射 Coulter计数器(液滴通过已知大小的小孔)。 界面面积 φ:分散相的体积分数; R:乳状液粒子的平均半径。
油滴大小分布 由蛋白质稳定的乳状液的物理和感官性质取决于所形成的液滴的大小和总界面面积。 测定乳状液平均液滴的大小的方法有: 光学显微镜法(不是非常可靠) 电子显微镜法. 光散射. Coulter计数器(液滴通过已知大小的小孔)。 界面面积. φ:分散相的体积分数; R:乳状液粒子的平均半径。")
120
(2) 乳化活力指标(Emulsifying Activity Index)
乳化活力指标(EAI) 单位质量蛋白质所产生的界面面积 m:蛋白质质量
 单位质量蛋白质所产生的界面面积. m:蛋白质质量.")
121
浊度法测定蛋白质的EAI (简便且更实际)
乳状液的浊度是由下式所确定 A:吸光度(500nm);ι:光路长度。 根据光散射的Mie 理论,乳状液的界面积是浊度的2倍。 φ:油的体积分数;(1-φ)ρ:单位体积乳状液中总的蛋白质量。
;ι:光路长度。 根据光散射的Mie 理论,乳状液的界面积是浊度的2倍。 φ:油的体积分数;(1-φ)ρ:单位体积乳状液中总的蛋白质量。")
122
(3) 蛋白质的载量(Protein Load )
蛋白质的载量 单位界面面积上吸附的蛋白质量。 将乳状液离心,使水相分离出来,然后重复地洗油相和离心以除去任何松散的被吸附的蛋白质。最初乳状液中总蛋白质量和从油相洗出的液体中蛋白质量之差即为吸附在乳化粒子上的蛋白质的量。 一般情况下,蛋白质的载量在1-3 mg/m2界面面积范围内。 在蛋白质含量保持不变的条件下,蛋白质的载量随乳状液中油相体积分数增加而降低。对于高脂肪乳状液和小尺寸液滴,需要有更多的蛋白质才足以涂布在界面上和稳定乳状液。

123
(4)乳化能力 乳化能力(EC)是指在乳状液相转变前(从O/W乳状液转变成W/O乳状液)每克蛋白质所能乳化的油的体积。 测定蛋白质乳化能力的方法如下: 在不变的温度和速度下,将油或熔化的脂肪加至在食品捣碎器中被连续搅拌的蛋白质水溶液,根据后者粘度和颜色(通常将染料加入油中)的突然变化或电阻的增加检测相的转变。 对于一个由蛋白质稳定的乳状液,相转变通常会发生在φ为 范围。 相转变并非是一个瞬时过程,相转变出现之前先形成W/O/W双重乳状液。
的突然变化或电阻的增加检测相的转变。 对于一个由蛋白质稳定的乳状液,相转变通常会发生在φ为 范围。 相转变并非是一个瞬时过程,相转变出现之前先形成W/O/W双重乳状液。")
124
乳化能力随相转变达到时蛋白质浓度的增加而减少。未吸附的蛋白质累积在水相。
为了比较不同蛋白质的乳化能力,应采用EC—蛋白质浓度曲线,取代在特定蛋白质浓度下的EC。

125
(5)乳状液稳定性 保藏在高温或在离心力下分离这样的剧烈条件来评价乳状液的稳定性。 若采用离心的方法,可用乳状液界面面积(即浊度)减少的百分数、或者分出的乳油的百分数、或者乳油层的脂肪含量表示乳状液的稳定性。常用下式表示乳状液稳定性: 乳油层体积是在乳状液经受标准化的离心处理后测得的。 乳状液,刻度离心管,1300×g离心5min;有时为了避免油滴聚结,在180×g下离心15min。 乳状液稳定性指标(ESI)的定义是乳状液的浊度达到起始值的一半所需要的时间。
的定义是乳状液的浊度达到起始值的一半所需要的时间。")
126
乳状液的破裂能或不能导致一个水层和(或)一个油层的分离。油层向上移动,水相向下移动。
许多食品乳状液是以油水分离方式失去它的稳定性。 通常,值为 时,O/W乳状液具有最高的稳定性。当值较低时发生排水;而在较高时倾向于产生聚结和油层分离。

127
2. 影响蛋白质乳化作用的因素 内在因素 pH、离子强度、温度、存在的低相对分子质量表面活性剂、糖、油相体积、蛋白质类型和使用的油的熔点等。 外在因素 如制备乳状液的设备的类型、能量输入的速度和剪切速度。 目前没有一致认可的系统地评价蛋白质乳化性质的标准方法。

128
影响蛋白质乳化作用的因素 (1)溶解度 蛋白质的溶解度在它的乳化性质方面起着重要的作用。 高度不溶性的蛋白质不是良好的乳化剂。 在25-80%溶解度范围内不存在蛋白质溶解度和乳化性质之间确实的关系。 良好的乳化性质所必需的最低溶解度取决于蛋白质的品种。在香肠那样的肉乳状液中,0.5M NaCl对肌纤维蛋白质的增溶作用促进了它的乳化性质。商业大豆分离蛋白质由于在加工中经受热处理而使它们的溶解度很低,并导致它们的乳化性质很差。

129
(2)pH pH以多种不同的方式影响着蛋白质的乳化性质 等电点时溶解度较低,影响其乳化能力。但是,在等电点时有利于蛋白质与脂肪的相互作用。 在等电点时缺乏净电荷和静电推斥相互作用,这有助于在界面达到最高蛋白质载量和促使高粘弹膜的形成。 在等电点具有高溶解度的蛋白质(例如血清清蛋白、明胶和蛋清蛋白)在此pH具有最高乳化活力和乳化能力。 大多数食品蛋白质(酪蛋白、商品乳清蛋白、肉蛋白、大豆蛋白、花生蛋白、牛血清蛋白和肌纤维蛋白)在它们的等电点pH时是微溶和缺乏静电推斥力的,因此在此pH它们一般不是良好的乳化剂。然而,这些蛋白质在远离它们的等电点pH时可能是有效的乳化剂。
在此pH具有最高乳化活力和乳化能力。 大多数食品蛋白质(酪蛋白、商品乳清蛋白、肉蛋白、大豆蛋白、花生蛋白、牛血清蛋白和肌纤维蛋白)在它们的等电点pH时是微溶和缺乏静电推斥力的,因此在此pH它们一般不是良好的乳化剂。然而,这些蛋白质在远离它们的等电点pH时可能是有效的乳化剂。")
130
(3) 疏水性 蛋白质的乳化性质与它的表面疏水性存在着一个弱正相关联,然而与平均疏水性无关。 一些蛋白质,像β-乳球蛋白、α-乳清蛋白和大豆蛋白,它们的乳化性质与表面疏水性之间不存在紧密的关联。
 疏水性 蛋白质的乳化性质与它的表面疏水性存在着一个弱正相关联,然而与平均疏水性无关。 一些蛋白质,像β-乳球蛋白、α-乳清蛋白和大豆蛋白,它们的乳化性质与表面疏水性之间不存在紧密的关联。")
131
(4)蛋白质分子的柔性 在油—水界面的蛋白质分子的柔性可能是决定蛋白质乳化性质最重要的因素。 蛋白质在乳化作用前的部分变性(展开),如果没有造成不溶解,通常能改进它们的乳化性质。因为提高了分子的柔性和表面疏水性。 蛋白质在界面上展开的速度取决于原始分子的柔性。 在展开状态,含有游离巯基和二硫键的蛋白质通过 -SH和-S-S- 交换反应经受缓慢的聚合作用,这会导致在油—水界面形成高粘弹性的膜。

132
(5)加热作用 加热处理通常能降低吸附在界面上的蛋白膜的粘度和硬度,因而降低了乳状液的稳定性。然而,高度水化的界面蛋白质膜的凝胶作用提高了表面粘度和硬度,从而稳定了乳状液。 足以造成蛋白质不溶解的热变性会损害蛋白质的乳化性质。 (6)小分子的表面活性剂 小分子的表面活性剂,降低了蛋白质膜的硬度和减弱了使保留在界面上的作用力。损害由蛋白质稳定的乳状液的稳定性。
小分子的表面活性剂. 小分子的表面活性剂,降低了蛋白质膜的硬度和减弱了使保留在界面上的作用力。损害由蛋白质稳定的乳状液的稳定性。")
133
(二) 起泡性质 1. 食品泡沫 气泡分散在含有可溶性表面活性剂的连续液体或半固体相中的分散体系。 糖蛋白、蛋糕、冰淇淋、面包、搅打奶油、蛋白甜饼、蛋奶酥、奶油冻和果汁软糖。气体则是空气或二氧化碳,连续相为水溶液或悬浊液。 气/液界面能达到1m2/ml液体的水平。需要加入表面活性剂降低界面张力和在截留的气泡之间形成一个弹性的保护壁垒。 由蛋白质稳定的泡沫一般是蛋白质溶液经吹气泡、搅打和摇振而形成的。

134
2. 蛋白质起泡能力的评价指标 在汽-液界面形成坚韧的薄膜使大量气泡并入和稳定的能力。
2. 蛋白质起泡能力的评价指标 在汽-液界面形成坚韧的薄膜使大量气泡并入和稳定的能力。 起泡能力(Foamability or Foaming Capacity):蛋白质能产生的界面面,有几种表示和评价方式积的量,有如下表示和评价方式 : 膨胀率(Overrun) 稳定状态泡沫值(Steady-state Foam Value) 起泡力(Foaming Power) 泡沫膨胀(Foam Expansion)
:蛋白质能产生的界面面,有几种表示和评价方式积的量,有如下表示和评价方式 : 膨胀率(Overrun) 稳定状态泡沫值(Steady-state Foam Value) 起泡力(Foaming Power) 泡沫膨胀(Foam Expansion)")
136
3. 形成泡沫的方法 气体通过一个多孔的喷洒器进入一个低蛋白质浓度的水溶液(0.01-2%)。 如果在体系中导入大量的气体,液体能完全转变成泡沫。 10倍的膨胀(1000%)是常见的。有时100倍的膨胀也可能达到。
。 如果在体系中导入大量的气体,液体能完全转变成泡沫。 10倍的膨胀(1000%)是常见的。有时100倍的膨胀也可能达到。 .")
137
(2) 在有整体的气相存在的条件下,通过搅打或振荡一个蛋白质水溶液也能形成泡沫。
打擦是将空气引入大多数充气食品时所优先采用的方法。打擦能产生较激烈的机械应力和剪切作用以及一个较为均一的气体分散体系。样品的体积增加一般在 %。 (3) 将一个预先被加压的溶液突然减压。当物料从气溶胶容器分离出来时形成了打擦奶油。
 将一个预先被加压的溶液突然减压。当物料从气溶胶容器分离出来时形成了打擦奶油。")
138
蛋白质稳定处在重力和机械力下的泡沫的能力。
泡沫稳定性(Foam Stability) 蛋白质稳定处在重力和机械力下的泡沫的能力。 表示为:50%液体从泡沫中排出所需要的时间或者泡沫体积减少50%所需要的时间。 作为有效的起泡剂,蛋白质必须满足以下要求: 快速吸附至气-水界面 易于在界面上展开和重排 通过分子间相互作用形成粘合性膜
 蛋白质稳定处在重力和机械力下的泡沫的能力。 表示为:50%液体从泡沫中排出所需要的时间或者泡沫体积减少50%所需要的时间。 作为有效的起泡剂,蛋白质必须满足以下要求: 快速吸附至气-水界面. 易于在界面上展开和重排. 通过分子间相互作用形成粘合性膜.")
139
影响蛋白质起泡性质的分子性质有: 溶解度 分子柔性 疏水性(两亲性) 带电基团和极性基团
 带电基团和极性基团")
140
4. 影响泡沫形成和稳定性的环境因素 (1)pH 在等电点时,泡沫不是很大,但稳定性较好。如球蛋白,面筋蛋白,乳清蛋白。 在等电点时,缺乏静电斥力,有利于形成粘合膜 不溶解的蛋白质对起泡能力没有贡献。因此,若等电点时溶解度太低,数量少,但稳定性仍好。 (2)盐 盐能影响蛋白质的溶解度、粘度、展开和聚集,因而能改变起泡性质。 钙离子能通过在蛋白质的羧基之间形成桥接而改进泡沫的稳定性。
盐. 盐能影响蛋白质的溶解度、粘度、展开和聚集,因而能改变起泡性质。 钙离子能通过在蛋白质的羧基之间形成桥接而改进泡沫的稳定性。")
141
乳清蛋白的起泡性质和泡沫稳定性随NaCl浓度的提高而降低,这是NaCl对乳清蛋白(特别是其中的ß-乳球蛋白)的盐溶效应引起的。
一般来说,蛋白质被盐析则显示较好的起泡性质。

142
(3)糖 由于蔗糖能增加粘度,抑制了泡沫的膨胀,但却改进了泡沫的稳定性。加糖蛋白泡沫打擦时,在后阶段加入糖,泡沫膨胀已经发生。 (4)脂 磷脂,比蛋白质更好的表面活性 污染的脂类会严重损害蛋白质的起泡性质。一般不含脂类的蛋白质有较好的起泡性质。具有表面活性的极性脂类能吸附在气/水界面从而妨碍了吸附的蛋白质膜形成最理想的构象。 (5)蛋白质浓度 蛋白质浓度增加,稳定性提高。蛋白质浓度的增加提高了黏度,产生了较小的气泡和坚硬的膜。有最大值,一般为2-8%W/V。
蛋白质浓度. 蛋白质浓度增加,稳定性提高。蛋白质浓度的增加提高了黏度,产生了较小的气泡和坚硬的膜。有最大值,一般为2-8%W/V。")
143
(6)温度 降低温度,疏水作用下降; 部分热变性,改进起泡性质;过高,形成-S-S-,不能吸附在表面。
温度 降低温度,疏水作用下降; 部分热变性,改进起泡性质;过高,形成-S-S-,不能吸附在表面。")
144
可以利用蛋白质作为结合期望的风味物的载体。
四、风味结合 可以利用蛋白质作为结合期望的风味物的载体。 (一)挥发性物质同蛋白质之间的相互作用 风味物结合包括吸附到食品的表面或通过扩散穿透到食品的内部(吸收)。 (1)通过van der Waals相互作用的物理吸附 (2)通过共价或静电键的化学吸附。 (3)通过疏水相互作用。
挥发性物质同蛋白质之间的相互作用. 风味物结合包括吸附到食品的表面或通过扩散穿透到食品的内部(吸收)。 (1)通过van der Waals相互作用的物理吸附. (2)通过共价或静电键的化学吸附。 (3)通过疏水相互作用。")
145
挥发性性风味化合物主要通过疏水相互作用与水合蛋白质结合。任何能改变蛋白质构象的因素都能影响挥发性化合物的结合。
(二)影响风味结合的因素 挥发性性风味化合物主要通过疏水相互作用与水合蛋白质结合。任何能改变蛋白质构象的因素都能影响挥发性化合物的结合。 热变性的蛋白质显示较高的结合风味物的能力。
影响风味结合的因素. 挥发性性风味化合物主要通过疏水相互作用与水合蛋白质结合。任何能改变蛋白质构象的因素都能影响挥发性化合物的结合。 热变性的蛋白质显示较高的结合风味物的能力。")
146
五、凝胶作用 (1)蛋白质凝胶的一般性质 缔合:在亚基或分子水平上的变化,聚合或聚集反应一般包括形成的复合物。
沉淀:导致完全或部分地丧失溶解性质的所有聚集反应 絮凝:不存在蛋白质变性作用时的随机聚集反应,这种现象往往是由于多肽链之间的静电排斥作用受到抑制而产生的. 凝结:变性引起的随机聚集反应和蛋白质-蛋白质相互作用超过蛋白质-溶剂相互作用而引起的聚集反应,凝结作用会引起在的结块形成。 凝胶:变性的分子聚集以形成一个有规则的蛋白质网

147
(2)凝胶形成和凝胶结构的特征 形成 三维蛋白质网的形成是蛋白质-蛋白质和蛋白质-溶剂相互作用之间以及相邻多肽链吸引力和排斥力之间一个平衡的结果。 吸引力 疏水相互作用(高温能提高此类作用),静电相互作用(钙),氢键(冷却)和(或)二硫交联。 排斥力 静电排斥和蛋白质-水相互作用能将多肽链保持在分离的状态。 蛋白质分子的相互吸引在高浓度下较易发生。

148
主要依靠非共价相互作用维持的凝胶网状结构是可逆的,加热时它们熔化成预凝胶状态。
主要依靠氢键形成的凝胶网状结构更是如此。明胶凝胶属这种情况。 依靠疏水相互作用形成的凝胶网状结构是不可逆的,疏水相互作用随温度升高而增强。 蛋清凝胶就属于这种情况。 含有半胱氨酸和胱氨酸的蛋白质在加热时通过-SH和-S-S相互交换反应产生聚合和在冷却时形成的连续的共价的网状结构(凝胶)通常是不可逆的。 卵清蛋白、β-乳球蛋白和乳清蛋白凝胶属于此种类型。
通常是不可逆的。 卵清蛋白、β-乳球蛋白和乳清蛋白凝胶属于此种类型。")
149
※ 不同种类的蛋白质在一起加热时能形成凝胶(共凝胶 作用)。
※ 蛋白质也能通过和多糖胶凝剂的相互作用形成凝胶。 带正电荷的明胶和带负电荷的海藻酸酸,酪蛋白带正 电荷区域和多硫化卡拉胶能产生特异性的离子相互作 用。

150
蛋白质能形成两类凝胶 凝结块(不透明)凝胶 含有大量非极性氨基酸残基的蛋白质在变性时产生疏水性聚集,随后这些不溶性的聚集体随机缔合而凝结成不可逆的凝结块类型的凝胶。 当Val、Pro、Leu、Ile、Phe和Trp残基的总和超过31.5摩尔百分数时,倾向于形成凝结块凝胶。 透明凝胶 仅含有少量非极性氨基酸残基的蛋白质在变性时形成可溶性复合物。可溶性复合物的缔合速度低于变性速度,凝胶网状结构主要是通过氢键相互作用而形成的,因此蛋白质溶液在加热后冷却时才能凝结成凝胶。冷却时可溶性复合物缓慢的缔合速度有助于形成有序的透明的凝胶网状结构。 当****低于31.5%,通常形成半透明类型的凝胶。

151
4. 影响蛋白质胶凝性质的因素 A.具有高摩尔量和高百分比的疏水氨基酸倾向于建立坚固的网。 B.蛋白质分子的解离和(或)展开一般会增加反应基团特别是球状蛋白质的疏水基团的暴露。这有利于蛋白质-蛋白质相互作用。 有限制的水解有时能促进蛋白质凝胶的形成。 在牛乳酪蛋白胶束中加入凝乳酶导致凝结块类凝胶的形成。

152
C.高温有利于疏水相互作用,而低温条件有利于氢键的形成。
加热处理能暴露内部的SH基团和促进二硫键的形成或交换。高百分数的SH和-S-S-强化了分子间的网状结构和倾向于形成热不可逆凝胶。 D.钙桥能提高许多凝胶的硬度和稳定性。 采用像Ca2+和Mg2+这样的二价阳离子在蛋白质分子的带负电荷基团之间形成交联。从大豆蛋白制备豆腐是此类凝胶中的一个很好的例子。

153
E.pH对胶凝性质的影响: 在或近等电点pH,蛋白质通常形成凝结块类凝胶。在极端pH,由于强烈的静电推斥作用,蛋白质形成弱凝胶。对于大多数蛋白质,形成凝胶的最适pH约7-8。 凝胶作用pH范围随蛋白质浓度的增加而增大,这说明高浓度疏水基团及二硫键补偿远离等电点pH下由于高的净电荷而产生的电荷斥力。在等电点,由于缺乏推斥力,因而形成较少的膨胀,较少的水化和硬度较低的凝胶。

154
F. 蛋白质浓度 为了形成一个静止后自动凝结的凝胶网状结构,一个最低蛋白质浓度,即最小浓度终点(least concentration endpoint LCE)是必需的。 大豆蛋白质、鸡蛋清蛋白和明胶的LCE值分别为8%、3%和0.6%。 G. 凝胶网状结构对热和机械力的稳定性取决于每单体链所形成的交联数目。 蛋白质凝胶硬度的平方根与相对分子质量呈线性关系。 相对分子质量小于23,000的球状蛋白质除非含有一个游离的巯基和一个二硫键,否则在任何合理的蛋白质浓度下不能形成热诱导凝胶。

155
测验题 1. 稳定蛋白质二级结构的主要作用力是什么? 稳 定蛋白质三级结构的主要作用力是什么? 2. 影响蛋白质变性的物理因素有哪些?化学因素有 哪些? 3. 蛋白质在食品中的主要功能性质有哪些? 4. 影响蛋白质起泡性质的因素有哪些?






 血液中转氨酶活力的测定 一. 目的 : 了解转氨酶在代谢过程中的重要作用及其在临 床诊断中的意义, 学习转氨酶活力测定的原理和方 法。 二. 原理 : 生物体内广泛存在的氨基转换酶也称转氨酶, 能 催化 α – 氨基酸的 α – 氨基与 α – 酮基互换, 在氨基酸 的合成和分解尿素和嘌呤的合成等中间代谢过程中.>")






 蔡哲瑜(4BS ) 詹君瑞(4BS ) 指導老師 : 潘文彬 老師>")







蛋白质的降解: 外源蛋白的消化 内源性蛋白的选择性降解 (2)氨基酸的分解代谢:>")
>")
