Download presentation
Presentation is loading. Please wait.

1
杀虫杀螨剂 第二章 (1)杀虫剂的推广应用,对控制农作物害虫的危害起到了良好的防治效果,推动了农药的发展和病虫草害的化学防治;
(2)推进了化学防治与其他防治措施相协调的综合技术的发展; (3)推进了有害生物抗药性的研究与治理。;
推进了化学防治与其他防治措施相协调的综合技术的发展; (3)推进了有害生物抗药性的研究与治理。;")
2
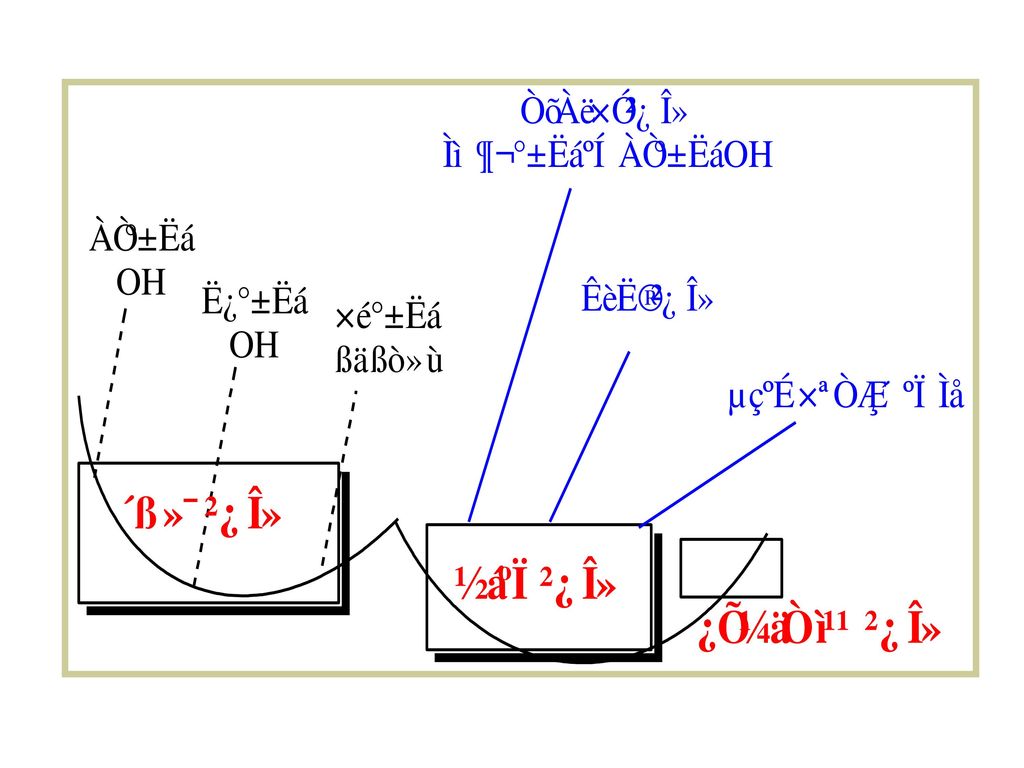
第一节 杀虫剂进入昆虫体内的途径 杀虫剂处理昆虫,不管是什么方式或是什么杀虫剂,必须通过昆虫的吸收。 经口摄入 肠吸收
第一节 杀虫剂进入昆虫体内的途径 杀虫剂处理昆虫,不管是什么方式或是什么杀虫剂,必须通过昆虫的吸收。 经口摄入 肠吸收 穿透肠道进入体内,进而扩散 生物转换(贮存、降解、活化) 排泄 到达作用靶标,引起昆虫死亡
 排泄. 到达作用靶标,引起昆虫死亡.")
3
一、杀虫剂进入昆虫体内的途径 1.从口腔进入 杀虫剂主要从昆虫的口腔、体壁及气门侵入体内而起作用。 昆虫中肠是杀虫剂吸收和穿透的主要场所。
一是杀虫剂能否在中肠溶解及溶解的速度, 二是对围食膜和肠壁细胞质膜的穿透能力。 a杀虫剂喷洒在农作物上,昆虫取食农作物,杀虫剂进入口腔; b杀虫剂和毒饵拌在一起,随饵料进入昆虫口腔; c内吸性杀虫剂被植物吸收并在植物体内运转,刺吸式口器害虫取食汁液或咀嚼式口器害虫取食植物,杀虫剂进入昆虫口腔。 如果杀虫剂是离子化合物,且离子直径小于4nm,则可从质膜上的水孔通过;如果杀虫剂是亲脂性化合物,可以依其浓度级差扩散通过;如果杀虫剂是亲水化合物,可和膜上某些载体蛋白结合,依赖蛋白质分子构象的变化转入膜内;如果杀虫剂是弱电介质,则只能依赖膜两 侧的pH梯度通过。若是弱酸,中肠pH小于7,杀虫剂大多呈分子型,容易通过,中肠pH大于7,杀虫剂大多呈离子型,难以通过;相反,若是弱碱,中肠pH小于7,杀虫剂大多呈离子型,难以通过,中肠pH大于7,杀虫剂大多呈分子型,容易通过。 口腔 前肠 中肠 血液 靶标 a b c 后肠 排泄

4
昆虫消化系统示意图

5
影响毒力的因素 服食 吸收、穿透 粒子越细越好,如粉剂; 不忌避,不据食; 不引起呕吐,不腹泻 药剂在中肠能否溶解及其溶解速度;
中场是一个主要的吸收场所。 两道防线:围食膜、肠壁细胞; 中肠PH,大多大于7。如果是碱性化合物,则多为分子型,易穿透;如果是酸性化合物,则难以通过

6
2 从昆虫体壁侵入 杀虫剂从昆虫体壁侵入: 上表皮 表皮 原表皮 水泥层 蜡质层 角质精层 多元酚层 外表皮 内表皮 主要有蛋白质组成
2 从昆虫体壁侵入 杀虫剂从昆虫体壁侵入: 上表皮 表皮 原表皮 水泥层 蜡质层 角质精层 多元酚层 外表皮 内表皮 主要有蛋白质组成 主要有糖蛋白组成

7
杀虫剂附着于虫体后首先溶解于上表皮的蜡质层,然后再按杀虫剂自身的油/水分配系数而进入原表皮。
昆虫体壁是个代表油/水两相的结构 上表皮代表油相; 原表皮代表水相。 杀虫剂附着于虫体后首先溶解于上表皮的蜡质层,然后再按杀虫剂自身的油/水分配系数而进入原表皮。 离子型,或亲水性很强的杀虫剂难以溶解于蜡质层,因而难以穿透上表皮; 脂溶性很强的杀虫剂比较容易穿透上表皮,但难以穿透原表皮。 因此,杀虫剂在其分子结构上具有一个适当的亲水亲油平衡的化合物,对昆虫表皮的穿透就比较迅速。

8
影响因素 一是决取于杀虫剂自身的性质 如果杀虫剂极性太强则难溶于亲脂的上表皮;反之,则那一透过亲水的内外表皮。即既要能溶解在蜡质层,也能通过元表皮。 二是取决于昆虫体壁的性质。 表皮的硬化、薄厚、蜡质程度等等。 昆虫身体构造并非完全一致,节间膜、触角、足的基节等都是未经骨化的膜状组织,药剂易侵入。感觉器集中的部位(跗节、触角、口器)也易进入。整体而言,侵入部位越接近脑部或神经节,昆虫容易中毒。
也易进入。整体而言,侵入部位越接近脑部或神经节,昆虫容易中毒。")
9
3 杀虫剂从气门侵入 影响杀虫剂侵入气门的因素主要是昆虫气门的开闭,凡是促使昆虫气门开放的因素均有利于杀虫剂侵入,如升温、增加二氧化碳浓度或降低氧浓度均可促使气门开放利于杀虫剂侵入虫体。 气门→气管→支气管→微气管→血液→靶标

10
注意: 每一种杀虫剂不只有一种穿透方法,但每一种杀虫剂都有一种主要的穿透方式; 口服: 666>DDT>氯丹

11
每一种杀虫剂的进入方式主要决定于它的物理性质;
例如:胃毒剂(脂溶性差,不易挥发) 熏蒸剂(表面张力低,易挥发) 触杀剂(表皮穿透) 某些杀虫剂的进入方式,决定于它的特有性质 例如:除虫菊易被消化液所破坏 DDT对表皮几丁质优特殊亲和力
 熏蒸剂(表面张力低,易挥发) 触杀剂(表皮穿透) 某些杀虫剂的进入方式,决定于它的特有性质. 例如:除虫菊易被消化液所破坏. DDT对表皮几丁质优特殊亲和力.")
12
二 、杀虫剂在昆虫体内的遭遇 杀虫剂在昆虫体内的贮存 杀虫剂在昆虫体内的转移 杀虫剂在昆虫体内的代谢 杀虫剂在昆虫体内的排泄

13
1 杀虫剂在昆虫体内的贮存 昆虫脂肪体有类似哺乳动物肝脏的功能,能贮存和代谢外源物。
由于脂肪体浸浴在血液中,因此进入血液的杀虫剂很容易被脂肪体吸收,特别是亲脂性强的杀虫剂易被脂肪体吸收贮存。 这不但影响了杀虫剂实际到达靶标的有效剂量,而且形成了在虫体内大量贮存、缓慢释放的现象,这就在时间上给了昆虫以解毒的机会。

14
2 杀虫剂在昆虫体内的转移 一般认为,杀虫剂随血液转移到各组织中进行再分配,然后到达作用靶标——神经系统。
目前认为昆虫也有可能存在类似脊椎动物的血脑屏障,离子型的化合物不能通过这个屏障。 70年代以来,Philip Gerolt等对于杀虫剂在昆虫体内的转移提出了不同看法,他们认为大多数杀虫剂主要通过气管系统这一途径而进入昆虫体内;杀虫剂在虫体内的转移过程中,血液是次要的,而主要是通过昆虫表皮的侧向扩散和在真皮细胞与内表皮之间的主动运输,再经由气管系统而到达作用靶标。 在脊椎动物的脑和脊髓外围有一个血脑屏障,具有类似生物膜的结构,能选择性地让外源物通过。

15
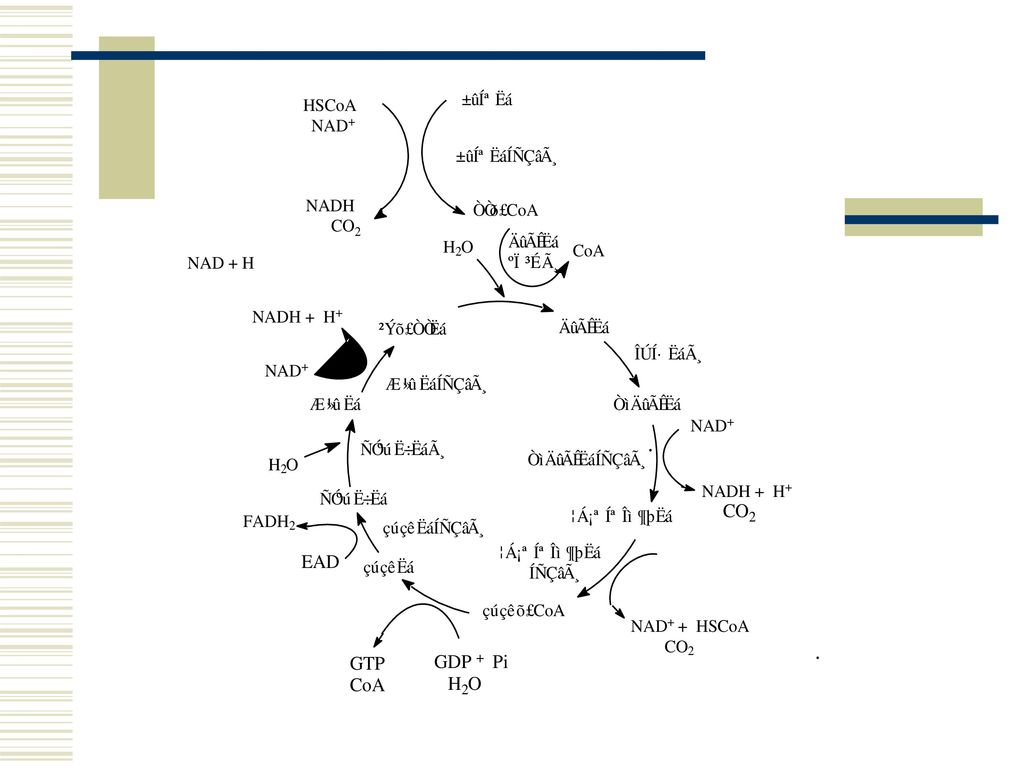
3 杀虫剂在昆虫体内的代谢 代谢大多是解毒过程,将杀虫剂转变成低毒或无毒的产物;
然而代谢也可能先将杀虫剂转变成更毒的产物,即活化,然后再解毒代谢。 杀虫剂的解毒代谢主要包括两大步骤: 第一步涉及氧化、水解及其它酶促反应,生成强极性的最终产物,这是非合成过程; 第二步是生成水溶性共轭产物,这是合成过程。 昆虫体内杀虫剂的非合成代谢中,占优势的有三种主要的酶促反应类型。即水解酶系,通过催化水解反应,分解底物(杀虫剂),如羧酸酯酶、酰胺酶、磷酸酯酶及A酯酶等;谷胱甘肽—S—转移酶系,特点是反应过程依赖还原型的谷胱甘肽,如DDT脱氯化氢酶、BHC(六六六)降解酶、脱甲基酶及脱芳基酶;微粒体氧化酶系,即多功能氧化酶系(MFO),特点是需要NADH、微粒体和氧的参与。
,如羧酸酯酶、酰胺酶、磷酸酯酶及A酯酶等;谷胱甘肽—S—转移酶系,特点是反应过程依赖还原型的谷胱甘肽,如DDT脱氯化氢酶、BHC(六六六)降解酶、脱甲基酶及脱芳基酶;微粒体氧化酶系,即多功能氧化酶系(MFO),特点是需要NADH、微粒体和氧的参与。")
16
初级代谢 (Primary metabolism)
(1)MFO酶系的氧化代谢 MFO主要由细胞色素P450、细胞色素B5、黄素蛋白—NADPH—P450还原酶、黄素蛋白—NADPH—细胞色素b5还原酶、磷酯等组成。 1958年Klingenberg和Garfinkel分别从鼠肝细胞的微粒体碎片中分离出一个复合物,具有多种生化功能,称之为多功能氧化酶;因其和CO形成的复合体在示差光谱中波长450nm处有个吸收峰,又称细胞色素P450。MFO主要存在于脊椎动物的肝、肺、肾、肠等器官中,存在于昆虫的中肠和脂肪体中。MFO主要由细胞色素P450、细胞色素B5、黄素蛋白—NADPH—P450还原酶、黄素蛋白—NADPH—细胞色素b5还原酶、磷酯等组成。
MFO酶系的氧化代谢. MFO主要由细胞色素P450、细胞色素B5、黄素蛋白—NADPH—P450还原酶、黄素蛋白—NADPH—细胞色素b5还原酶、磷酯等组成。 1958年Klingenberg和Garfinkel分别从鼠肝细胞的微粒体碎片中分离出一个复合物,具有多种生化功能,称之为多功能氧化酶;因其和CO形成的复合体在示差光谱中波长450nm处有个吸收峰,又称细胞色素P450。MFO主要存在于脊椎动物的肝、肺、肾、肠等器官中,存在于昆虫的中肠和脂肪体中。MFO主要由细胞色素P450、细胞色素B5、黄素蛋白—NADPH—P450还原酶、黄素蛋白—NADPH—细胞色素b5还原酶、磷酯等组成。")
17
MFO的氧化反应机制 细胞色素是以血红素为辅基的蛋白质,在血红素中,4个吡咯环的氮与Fe原子配位形成方形平面复合物 底物(XH)在光滑内质网上先与氧化型P450Fe3+结合,形成复合P450—Fe3+—XH。在NADPH和细胞色素还原酶催化下,由NADPH供给电子(H+留在基质中)经黄素蛋白(FAD)传递给氧化型复合物,使之还原成还原型复合物P450—Fe2+—XH。这种还原型复合物与分子氧作用,组成含氧复合物,并接受一个电子,生成P450—Fe2+—XOH,同时另一个氧与基质中的H+结合生成水。最后复合物又释出氧化产物。从图可看出,氧化的实质乃是P450在起作用。 细胞色素是以血红素为辅基的蛋白质,在血红素中,4个吡咯环的氮与Fe原子配位形成方形平面复合物,Fe的第5位、第6位(即图中的x,y)垂直于方形平面,第5位(x位)和蛋白质结合,第6位(y位)和底物结合(可和O2、CO、XH等结合)。电子交换通过Fe3+→Fe2+进行。未和底物结合时,Fe和4个吡咯环同处于一个平面内,当和底物结合时,结构改变,Fe和4个吡咯环不再处于同一平面,而是Fe高高隆起。 底物(XH)+(氧化型)P450Fe3+→(氧化型)P450—Fe3+—XH + e [NADPH供给 →黄素蛋白(FAD)] → (还原型)P450—Fe2+—XH +O2 →含氧复合物 + e →P450—Fe2+—XOH → XOH
在光滑内质网上先与氧化型P450Fe3+结合,形成复合P450—Fe3+—XH。在NADPH和细胞色素还原酶催化下,由NADPH供给电子(H+留在基质中)经黄素蛋白(FAD)传递给氧化型复合物,使之还原成还原型复合物P450—Fe2+—XH。这种还原型复合物与分子氧作用,组成含氧复合物,并接受一个电子,生成P450—Fe2+—XOH,同时另一个氧与基质中的H+结合生成水。最后复合物又释出氧化产物。从图可看出,氧化的实质乃是P450在起作用。 细胞色素是以血红素为辅基的蛋白质,在血红素中,4个吡咯环的氮与Fe原子配位形成方形平面复合物,Fe的第5位、第6位(即图中的x,y)垂直于方形平面,第5位(x位)和蛋白质结合,第6位(y位)和底物结合(可和O2、CO、XH等结合)。电子交换通过Fe3+→Fe2+进行。未和底物结合时,Fe和4个吡咯环同处于一个平面内,当和底物结合时,结构改变,Fe和4个吡咯环不再处于同一平面,而是Fe高高隆起。 底物(XH)+(氧化型)P450Fe3+→(氧化型)P450—Fe3+—XH + e [NADPH供给 →黄素蛋白(FAD)] → (还原型)P450—Fe2+—XH +O2 →含氧复合物 + e →P450—Fe2+—XOH → XOH.")
18
MFO催化的主要反应类型 羟基化反应 烷基羟基化 速灭威代谢

19
芳基羟基化 西维因代谢

20
脱去O-,S-,N-烷基中的烷基 杀虫威代谢 注意

21
实际上,脱烷基反应仍是先羟基化,生成物不稳定,又进一步分解而脱出烷基:
注意

22
酯键的氧化—— 可能是增毒反应 乐果代谢

23
环氧化—— 可能是增毒反应 艾氏剂代谢

24
(2)水解酶系的水解代谢 磷酸酯酶主要催化有机磷酸酯类杀虫剂的水解代谢
水解酶系的水解代谢 磷酸酯酶主要催化有机磷酸酯类杀虫剂的水解代谢")
25
羧酸酯酶主要催化羧酸酯的水解: 马拉硫磷 酰胺水解酶主要催化酰胺的水解:如乐果

26
(3)谷胱甘肽—S—转移酶的脱甲基代谢 谷胱甘肽—S—转移酶在有机磷杀虫剂的二甲基磷酸酯或二甲基硫代磷酸酯的解毒代谢中起重要作用。这一酶促代谢的特点是必须有谷胱甘肽的参与,结果是脱去甲基:
谷胱甘肽—S—转移酶的脱甲基代谢 谷胱甘肽—S—转移酶在有机磷杀虫剂的二甲基磷酸酯或二甲基硫代磷酸酯的解毒代谢中起重要作用。这一酶促代谢的特点是必须有谷胱甘肽的参与,结果是脱去甲基:")
27
2 次级代谢 (Secondary metabolism)
杀虫剂在昆虫体内的初级代谢产物,往往仍没有足够的水溶性,因此往往经历次级代谢,生成完全溶于水的共轭物,通过排泄系统排泄出去。 昆虫体内最重要的共轭剂是葡萄糖,此外还有谷胱甘肽、磷酸、甘氨酸、硫酸盐等,而能和共轭剂结合的次级代谢产物一般应在分子结构上具有羟基、羧基和氨基等功能基团。

28
共轭机制 和葡萄糖共轭——先生成一个活化的中间体—尿苷二磷酸葡萄糖(UDPG),然后葡萄糖从UDPG转移到共轭物上
和硫酸盐共轭—— 先生成活化中间产物3′-磷酸腺苷-5′-磷酰硫酸(PAPS)
")
29
四、 杀虫剂的排泄 杀虫剂在昆虫体内经过初级代谢和次级代谢后形成小分子的,水溶性的化合物,被马氏管(伸入在血液中)吸收后通过前后肠,到达直肠,然后随粪便排出体外。
吸收后通过前后肠,到达直肠,然后随粪便排出体外。")
30
第二节 杀虫剂作用机制有关的昆虫神经生理 一、昆虫与脊椎动物在神经结构方面有下述不同之处:
昆虫没有单独的副交感神经,而交感神经具有一定的副交感功能; 昆虫的运动神经末梢和肌纤维组成的突触,其兴奋性神经递质是谷氨酸盐,而脊椎动物则是乙酰胆碱; 昆虫的胆碱能突触(以乙酰胆碱为神经递质的突触)全部在中枢神经系统内,外周神经系统没有胆碱能突触。
全部在中枢神经系统内,外周神经系统没有胆碱能突触。")
31
脊椎动物的神经系统

32
在神经组织学和形态学方面还有以下的差别:
在昆虫中没有明显的髓鞘(myelination)存在;昆虫的神经索被覆一层“神经鞘”,它阻止了许多外源物进入鞘内,其作用好似哺乳动物的血脑屏障的作用; 昆虫的神经肌肉联结点并不像脊椎动物那样有特化的“终板”,昆虫轴突分出几个分枝,并刺激单个肌纤维,而脊椎动物的“终板”是支配一束肌纤维; 昆虫的神经系统是依靠气管系统通过扩散作用直接向神经细胞供氧,而脊椎动物是由血液向神经细胞输氧。
存在;昆虫的神经索被覆一层 神经鞘 ,它阻止了许多外源物进入鞘内,其作用好似哺乳动物的血脑屏障的作用; 昆虫的神经肌肉联结点并不像脊椎动物那样有特化的 终板 ,昆虫轴突分出几个分枝,并刺激单个肌纤维,而脊椎动物的 终板 是支配一束肌纤维; 昆虫的神经系统是依靠气管系统通过扩散作用直接向神经细胞供氧,而脊椎动物是由血液向神经细胞输氧。")
33
二、 信息的传递机制

34
信息的传递类型 轴突(axon)传导:即一个神经元内的传导 树突→细胞体→轴突→端丛
突触(Synapse)传导:神经元之间或神经元—肌肉(腺体)之间的传导。 一个神经元的端丛→另一神经元的树突
传导:神经元之间或神经元—肌肉(腺体)之间的传导。 一个神经元的端丛→另一神经元的树突.")
35
1 轴突传导 (1)静息电位的离子基础。 细胞膜两侧的电位差就是膜电位(membrane potential,Em),静息状态下的膜电位就是静息电位(resting potential)。 一个神经元就是一个细胞,轴突外面也被覆一个膜,也是个半透性膜,膜外(组织液内)有大量的Na+,少量的K+,许多阴离子,主要是Cl-及少量有机阴离子(A-);膜内(轴浆内)有大量的K+,少量的Na+,许多阴离子,主要是有机阴离子(A-)及少量Cl-。静止时(即没有兴奋传导时),K+和Cl-几乎可以自由通过,而Na+几乎完全不能通过,A-完全不能通过。 由于膜内K+浓度高,倾向外流(这是因为浓度梯度所致)。K+向膜外扩散增加了膜外的正电荷。由于异电相吸的原理,膜内的A-也有随K+外流的倾向,但由于膜的阻止,A-不得通过而靠在膜的内表面,于是形成了以膜为中心的静电场,膜内表面为负电场,膜外表面为正电场,这就是所谓“外正内负”。 (组织液内) (轴浆内)
有大量的Na+,少量的K+,许多阴离子,主要是Cl-及少量有机阴离子(A-);膜内(轴浆内)有大量的K+,少量的Na+,许多阴离子,主要是有机阴离子(A-)及少量Cl-。静止时(即没有兴奋传导时),K+和Cl-几乎可以自由通过,而Na+几乎完全不能通过,A-完全不能通过。 由于膜内K+浓度高,倾向外流(这是因为浓度梯度所致)。K+向膜外扩散增加了膜外的正电荷。由于异电相吸的原理,膜内的A-也有随K+外流的倾向,但由于膜的阻止,A-不得通过而靠在膜的内表面,于是形成了以膜为中心的静电场,膜内表面为负电场,膜外表面为正电场,这就是所谓 外正内负 。 (组织液内) (轴浆内)")
36
K+的平衡的电位(Ek) 当浓度极差使K+流出,内部的负电场使K+不能外流。当K+由于浓度极差形成的向外扩散的力与阻止K+外流的电场力相等时,K+就不再外流而达到一个动态平衡。 由于膜内K+的浓度高,即[K+]i大,因此,Ek电位是负值,即零电位以下,大约在-50V~-90mV。 式中: R----气体常数; n----阳离子的单位电荷数; T-----绝对温度; F-----法拉弟常数; [K+]o-----膜外浓度; [K+]i-----膜内浓度;
![K+的平衡的电位(Ek) 当浓度极差使K+流出,内部的负电场使K+不能外流。当K+由于浓度极差形成的向外扩散的力与阻止K+外流的电场力相等时,K+就不再外流而达到一个动态平衡。 由于膜内K+的浓度高,即[K+]i大,因此,Ek电位是负值,即零电位以下,大约在-50V~-90mV。](http://slidesplayer.com/slide/11198358/60/images/36/K%2B%E7%9A%84%E5%B9%B3%E8%A1%A1%E7%9A%84%E7%94%B5%E4%BD%8D%EF%BC%88Ek%EF%BC%89+%E5%BD%93%E6%B5%93%E5%BA%A6%E6%9E%81%E5%B7%AE%E4%BD%BFK%2B%E6%B5%81%E5%87%BA%EF%BC%8C%E5%86%85%E9%83%A8%E7%9A%84%E8%B4%9F%E7%94%B5%E5%9C%BA%E4%BD%BFK%2B%E4%B8%8D%E8%83%BD%E5%A4%96%E6%B5%81%E3%80%82%E5%BD%93K%2B%E7%94%B1%E4%BA%8E%E6%B5%93%E5%BA%A6%E6%9E%81%E5%B7%AE%E5%BD%A2%E6%88%90%E7%9A%84%E5%90%91%E5%A4%96%E6%89%A9%E6%95%A3%E7%9A%84%E5%8A%9B%E4%B8%8E%E9%98%BB%E6%AD%A2K%2B%E5%A4%96%E6%B5%81%E7%9A%84%E7%94%B5%E5%9C%BA%E5%8A%9B%E7%9B%B8%E7%AD%89%E6%97%B6%EF%BC%8CK%2B%E5%B0%B1%E4%B8%8D%E5%86%8D%E5%A4%96%E6%B5%81%E8%80%8C%E8%BE%BE%E5%88%B0%E4%B8%80%E4%B8%AA%E5%8A%A8%E6%80%81%E5%B9%B3%E8%A1%A1%E3%80%82+%E7%94%B1%E4%BA%8E%E8%86%9C%E5%86%85K%2B%E7%9A%84%E6%B5%93%E5%BA%A6%E9%AB%98%EF%BC%8C%E5%8D%B3%5BK%2B%5Di%E5%A4%A7%EF%BC%8C%E5%9B%A0%E6%AD%A4%EF%BC%8CEk%E7%94%B5%E4%BD%8D%E6%98%AF%E8%B4%9F%E5%80%BC%EF%BC%8C%E5%8D%B3%E9%9B%B6%E7%94%B5%E4%BD%8D%E4%BB%A5%E4%B8%8B%EF%BC%8C%E5%A4%A7%E7%BA%A6%E5%9C%A8-50V%EF%BD%9E-90mV%E3%80%82.jpg "式中: R----气体常数; n----阳离子的单位电荷数; T-----绝对温度; F-----法拉弟常数; [K+]o-----膜外浓度; [K+]i-----膜内浓度;")
37
Na+的平衡电位ENa 由于Na+在膜外浓度高于膜内,这一浓度级差在理论上也会使Na+内流(实际上很少通过),因此也能算出Na+的平衡电位ENa 但膜外Na+浓度高,所以ENa是正值,约为40mV。 然而事实上,当膜静止时,膜对K+是自由通透的,而Na+通透性很小,虽然Cl-也会因浓度级差从膜外扩散到膜内,增加膜外的正电荷 (负电荷少了),也参与了一部分静息膜电位的形成,但Cl-的通透性在静止时比K+小得多,因此,静息电位主要K+的外流造成的,实际上的膜电位是接近K+的平衡电位的,但由于有很少一部分的Na+内流,又使膜电位不完全等于K+电位,而是比K+电位稍高些。由于膜电位不处于零电位上,因此膜处在极化状态。
,也参与了一部分静息膜电位的形成,但Cl-的通透性在静止时比K+小得多,因此,静息电位主要K+的外流造成的,实际上的膜电位是接近K+的平衡电位的,但由于有很少一部分的Na+内流,又使膜电位不完全等于K+电位,而是比K+电位稍高些。由于膜电位不处于零电位上,因此膜处在极化状态。")
38
(2)动作电位的产生 当一个刺激作用于神经细胞时 (这个刺激可以是机械的、光的、热的等等,但都先要依赖感受器的换能机制转变为生物电流),在刺激部位上Na+的通透产生了一个突然而巨大的增加,在不到lms的时间内,膜电位就跃进到一个峰值,接近Na+平衡电位,这个膜电位向正电的上升就是动作电位的上相(rising phase),这个过程便是去极化阶段(depolarisation)
,在刺激部位上Na+的通透产生了一个突然而巨大的增加,在不到lms的时间内,膜电位就跃进到一个峰值,接近Na+平衡电位,这个膜电位向正电的上升就是动作电位的上相(rising phase),这个过程便是去极化阶段(depolarisation)")
39
(3)动作电位的传导 (a)显示出神经膜在静止时的电荷分布。
假设A点受到刺激,产生的生物电流超过了阈值电位,原来外正内负的情况在上升阶段就变成外负内正,结果在A、B之间形成一个局部电流,这个电流虽然很弱,但能引起B点去极化。这个去极化虽然很弱,但可以达到阈值电位,因此在B点自发地产生另一个动作电位,这又导致在B点及其邻近的C点之间形成一个局部电流……,这个过程全是由于邻近点上离子通透性的改变而产生去极化,去极化达到一个阈值再产生一个动作电位,每个电位都一样强,它是一个“全或无”(all or no)过程,因而兴奋传导不会减弱。
过程,因而兴奋传导不会减弱。")
40
2 突触的传导 (1)突触的构造 突触依其结构和功能不同,可分为: 电突触 化学突触两大类 和杀虫剂作用机制有关的主要是化学突触。
2 突触的传导 (1)突触的构造 突触依其结构和功能不同,可分为: 电突触 化学突触两大类 和杀虫剂作用机制有关的主要是化学突触。 突触的结构如图所示,神经元轴突末端膨大为突触体,它与下一神经元的细胞体或树突中间存在一间隙,间隙之前和之后的膜,分别称为突触前膜和突触后膜。 突触前膜中重要的结构是突触小泡,小泡中含有神经递质。不同种类的突触,小泡的大小,形状及神经递质各不相同。 突触间隙一宽的20~50nm的缝隙,其中有电子密度高的物质(为含糖基的物质)把突触前、后膜连接起来,以便于前膜释放的神经递质向后膜扩散,完成信息传递功能。 突触后膜的最大特点是其膜上有专门的神经递质受体部位以及有关的酶系。
突触的构造. 突触依其结构和功能不同,可分为: 电突触. 化学突触两大类. 和杀虫剂作用机制有关的主要是化学突触。 突触的结构如图所示,神经元轴突末端膨大为突触体,它与下一神经元的细胞体或树突中间存在一间隙,间隙之前和之后的膜,分别称为突触前膜和突触后膜。 突触前膜中重要的结构是突触小泡,小泡中含有神经递质。不同种类的突触,小泡的大小,形状及神经递质各不相同。 突触间隙一宽的20~50nm的缝隙,其中有电子密度高的物质(为含糖基的物质)把突触前、后膜连接起来,以便于前膜释放的神经递质向后膜扩散,完成信息传递功能。 突触后膜的最大特点是其膜上有专门的神经递质受体部位以及有关的酶系。")
41
(2)突触传递的特点 单向传递;有“突触延搁”,即所需时间较轴突上长,这个延搁为0.5~2ms; 最易受环境影响,最易疲劳;
需要化学介质(如乙酰胆碱、去肾上腺素、多胺、氨基于酸、谷氨酸盐等)。
。")
42
(3)突触的兴奋传递过程 当一个冲动到达前膜时,改变了膜上离子的渗透性,导致细胞外液中的Ca2+向内流动,这个Ca2+的内流,通过还不完全了解的机制增加了小泡的随机运动,并与突触前膜撞击。 撞击时,小泡中的神经递质释放,扩散通过突触间隙,与突触后膜上的受体相结合,引起后膜去极化,形成突触后电位(EPSP),突触后电位类似前述发动电位,达到Na+的活化阈值,就产生一个新的动作电位 。 为了避免由于神经递质的继续存在而引起受体的不断兴奋,在新的动作电位产生后,必须有某种机制使神经递质对受体的作用终止。在不同的突触中,这种灭活机制各不相同。在以乙酰胆碱为神经递质的突触中,乙酰胆碱酯酶及时把乙酰胆碱水解。
,突触后电位类似前述发动电位,达到Na+的活化阈值,就产生一个新的动作电位 。 为了避免由于神经递质的继续存在而引起受体的不断兴奋,在新的动作电位产生后,必须有某种机制使神经递质对受体的作用终止。在不同的突触中,这种灭活机制各不相同。在以乙酰胆碱为神经递质的突触中,乙酰胆碱酯酶及时把乙酰胆碱水解。")
43
三 乙酰胆碱酯酶(AChE) 1 AChE的结构
生物体内AChE是一大分子糖蛋白,糖基约占总量的15%。AChE按其分子特征可分为球型(对称型)及尾型(不对称型)。球型单体(G1)的分子量为70 000~80 000u,单体通过单一的链间二硫键装配成二聚体(G2),两个二聚体通过范德华力结合成四聚体(G4)。球型四聚体通过加入3股胶原样尾而成为尾型四聚体(A4)、八聚体(A8),及十二聚体(A12) 昆虫体内,AChE分子类型较简单,主要是球型二聚体(G2),其次是球型单体(G1)。
及尾型(不对称型)。球型单体(G1)的分子量为70 000~80 000u,单体通过单一的链间二硫键装配成二聚体(G2),两个二聚体通过范德华力结合成四聚体(G4)。球型四聚体通过加入3股胶原样尾而成为尾型四聚体(A4)、八聚体(A8),及十二聚体(A12) 昆虫体内,AChE分子类型较简单,主要是球型二聚体(G2),其次是球型单体(G1)。")
44
催化部位 酯动部位 这个部位主要是一个丝氨酸的羟基、酪氨酸的羟基和一个碱性基团(组氨酸的咪唑基)起作用。
一般情况下,单独的丝氨酸羟基并不能和酰基化合物反应,而要依赖邻近的组氨酸咪唑基的活化作用,底物才水解。

45
结合部位 阴离子部位:最重要部位,是酪氨酸和谷氨酸羟基起作用; 疏水部位:可以使与底物的疏水基团结合增加亲和力;
电子转移复合体:酶与底物接近时,如果一方有供电子的倾向,另一方有接受电子的倾向,二者结合,形成一个电子复合体

46
空间异构部位 远离催化部位,被占领可改变整个酶的三维结构,从而改变酶的反应性。

48
2 ACHE的催化机制 kd(解离常数)=K+1/K-1
第一步:形成酶和底物的复合体(米氏络合物)。乙酰胆碱的季氮可和酶的阴离子部位结合,两个亚甲基及3个甲基可和酶的疏水基部位结合,从而形成一个米氏络合物。络合物的稳定性可以用解离常数(kd)来表示, kd越小,说明E和AX的亲和力越大,络合物E·AX越稳定。 第二步:乙酰化反应生成乙酰化酶。用K2表示速率常数,K2越大,乙酰化反应越迅速。近年来提出在米氏络合物和乙酰化酶之间,还可能有另一个中间产物——诱导契合复合物的生成,酯动部位催化三联体中的组氨酸咪唑基吸引丝氨酸基的质子,丝氨酸基氧活化(带局部负电荷)并攻击羰基碳原子(带局部正电荷),这个碳原子本来连接3个原子,一个双键,两个单键,并处于同一平面,碳原子位于平面中心,3个键互成120度角。当氧攻击这个碳并与之结合后,原来的平面结构变为四面体结构。因此诱导契合复合物又叫做四面体中间体。中间体转化为乙酰化酶并释放出胆碱。 第三步:乙酰化酶水解成乙酸,酶复活。K3越大,说明酶的复活越迅速,组氨酸咪唑基吸引水分子的质子,产生的羟基离子再次攻击乙酰化酶的羰基碳而形成另一个四面体中间体,这个中间体水解为AChE和乙酸。 kd(解离常数)=K+1/K-1
。乙酰胆碱的季氮可和酶的阴离子部位结合,两个亚甲基及3个甲基可和酶的疏水基部位结合,从而形成一个米氏络合物。络合物的稳定性可以用解离常数(kd)来表示, kd越小,说明E和AX的亲和力越大,络合物E·AX越稳定。 第二步:乙酰化反应生成乙酰化酶。用K2表示速率常数,K2越大,乙酰化反应越迅速。近年来提出在米氏络合物和乙酰化酶之间,还可能有另一个中间产物——诱导契合复合物的生成,酯动部位催化三联体中的组氨酸咪唑基吸引丝氨酸基的质子,丝氨酸基氧活化(带局部负电荷)并攻击羰基碳原子(带局部正电荷),这个碳原子本来连接3个原子,一个双键,两个单键,并处于同一平面,碳原子位于平面中心,3个键互成120度角。当氧攻击这个碳并与之结合后,原来的平面结构变为四面体结构。因此诱导契合复合物又叫做四面体中间体。中间体转化为乙酰化酶并释放出胆碱。 第三步:乙酰化酶水解成乙酸,酶复活。K3越大,说明酶的复活越迅速,组氨酸咪唑基吸引水分子的质子,产生的羟基离子再次攻击乙酰化酶的羰基碳而形成另一个四面体中间体,这个中间体水解为AChE和乙酸。 kd(解离常数)=K+1/K-1.")
49
AChE催化机制

50
四、乙酰胆碱受体(AChR) 乙酰胆碱受体的功能是在突触部位接受由前膜释放的神经递质乙酰胆碱(ACh)后被激活,引起后膜离子通透性的改变,造成离子通道(主要是Na+通道)开放,Na+涌入膜内,膜去极化,产生动作电位,使神经兴奋继续传导。 目前,至少己发现3种乙酰胆碱受体,即烟碱样受体(N型AChR),蕈毒碱样受体(M型AChR)及蕈毒酮样受体。
,蕈毒碱样受体(M型AChR)及蕈毒酮样受体。")
51
烟碱样受体 N型受体(nicotinic receptor)
主要分布在推动物的神经-肌肉接头及植物神经节内,在中枢神经系统内,在小脑和脊髓Renshaw细胞中发现。 在昆虫中,N型受体全部在中枢神经系统内。 该受体被占领的反应为骨骼肌收缩,植物神经节兴奋。 N型受体的激动剂为烟碱(小剂量)、碳酰胆碱。N型受体的颉颃剂有α环蛇毒素、筒箭毒素、五羟季胺或六羟季胺等。
、碳酰胆碱。N型受体的颉颃剂有α环蛇毒素、筒箭毒素、五羟季胺或六羟季胺等。")
53
蕈毒碱样受体 M型受体(muscarinic receptor)
M型AChR主要分布于哺乳动物的平滑肌和各种腺体内,在中枢神经系统,主要存在于大脑皮质和纹状体内。 该受体被占领后表现出来的反应是血管舒张、肠胃收缩、瞳孔缩小、汗腺兴奋。 M型AChR的激动剂为蕈毒碱、毛果芸香碱;M型AChR的颉颃剂为阿托品、东莨菪碱等。

54
M型AChR和N型AChR的结构不同。 M型AChR被递质ACh结合后,激活腺苷酸环化酶(ACE),从而使细胞内CAMP的浓度升高,CAMP又激活蛋白激酶,蛋白激酶使离子通道蛋白磷酸化,从而调节离子通道的开闭。
,从而使细胞内CAMP的浓度升高,CAMP又激活蛋白激酶,蛋白激酶使离子通道蛋白磷酸化,从而调节离子通道的开闭。")
55
蕈毒酮样受体 (muscaronic receptor)
近年来还发现在昆虫(果蝇和家蝇)中有第3种AChR,即蕈毒酮受体。 蕈毒酮对该受体有很强的亲合性。 研究证明蕈毒酮和其受体的结合,不仅受到烟碱样药物和蕈毒碱样药物抑制,而且也为非胆碱性药物所抑制。 关于蕈毒酮样受体的分子结构,目前还知之甚少。
中有第3种AChR,即蕈毒酮受体。 蕈毒酮对该受体有很强的亲合性。 研究证明蕈毒酮和其受体的结合,不仅受到烟碱样药物和蕈毒碱样药物抑制,而且也为非胆碱性药物所抑制。 关于蕈毒酮样受体的分子结构,目前还知之甚少。")
56
第三节 杀虫剂的作用机制 杀虫剂对AChE的抑制作用 杀虫剂对AChR的抑制作用 杀虫剂对GABA受体的影响 杀虫剂对轴突传导的干扰
第三节 杀虫剂的作用机制 杀虫剂对AChE的抑制作用 杀虫剂对AChR的抑制作用 杀虫剂对GABA受体的影响 杀虫剂对轴突传导的干扰 杀虫剂对呼吸系统的干扰 杀虫剂对昆虫的其他作用

58
一、杀虫剂对AChE的抑制作用 (一)有机磷酸酯类杀虫剂的作用机制
有机磷杀虫剂的作用机制就在于其抑制了AChE的活性,使得乙酰胆碱不能及时地分解而积累,不断和受体结合,造成后膜上Na+通道长时间开放,突触后膜长期兴奋,从而影响了神经兴奋的正常传导。 但乙酰胆碱酯酶被抑制如何造成昆虫死亡,这一点尚未研究清楚。在高等动物中,AChE被抑制而影响了呼吸,造成窒息,引起死亡,而在昆虫中尚未阐明。

59
有机磷杀虫剂抑制AChE的反应同AChE分解乙酰胆碱的反应一样,也分三步:
(1)酶和抑制剂形成复合体,这一步是亲和。
酶和抑制剂形成复合体,这一步是亲和。")
60
(2)生成磷酰化酶 新键的生成和旧键的断裂是同时进行
磷酰化的机理可描述如下:酯解部位组氨酸咪唑基的1个氮原子上的质子受埋藏在AChE分子内部的天门冬氨酸游离羧基的吸引,通过共轭效应使咪唑的另一个氮原子吸引丝氨酸羟基上的质子,从而使丝氨酸活化。丝氨酸羟基氧原子上未配位电子对可与有机磷酸酯带局部正电荷的磷原子之间发生亲核取代反应,并形成共价键,酯解部位酸性基因的质子与离去基团的氧形成氢键,离去基团脱落。最后,碱性基团咪唑氮原子上的质子再交给酸性基团,于是磷酰化酶生成。

61
(3)磷酰化酶水解 这一步是复活 磷酰基完整地从磷酰化酶分子上脱落下来的反应叫做去磷酰化反应,是一种水解反应,即在水分子参与下P-O键的断裂。AChE酯解部位的组氨酸咪唑基吸引水的质子以形成氢键,使OH-基团活化,与磷酰基发生亲核取代反应,磷酰残基脱落,丝氨酸羟基氧原子从酸性基团取得一个质子。最后,碱性基团咪唑基从活化水分子得到的质子再转交给酸性基因。

62
(4)酶老化 磷酰化酶转变为另一结构,羟胺类药物不能使其恢复活性
酶老化 磷酰化酶转变为另一结构,羟胺类药物不能使其恢复活性")
63
磷酰化反应的特点 对AChE的抑制实质就是生成磷酰化酶。因此K2、K3是决定抑制速率的两个速率常数。K2小,说明磷酰化反应缓慢,昆虫中毒症状表现就慢;K3大,说明生成的磷酰化酶不稳定,很快就水解,酶复活,抑制作用解除。因此,要有强大的毒力,就要K2足够地大,K3足够地小,

64
一是利用磷原子的亲电性(磷带局部正电荷)攻击酶活性中心的丝氨酸羟基。因此,凡是可以增加磷原子正电荷的因素都可以使磷酰化反应加速;
二是亲电反应与X基团(又称”酸性基团”)的离去是同时进行的,基团分离后(即P-X键断裂)磷酰化酶才形成。因此P-X键的极性越强,键就越容易断裂。
的离去是同时进行的,基团分离后(即P-X键断裂)磷酰化酶才形成。因此P-X键的极性越强,键就越容易断裂。")
65
X基团——吸电子基团,满足1 RO2——供电子基团,满足2、3
1.要求分子中有吸电子基因,才能造磷原子的局部正电荷Pδ+,Pδ+的越大,磷酰化反应越讯速; ; 2.要求生成的磷酰化酶比较稳定,被抑制的AChE难以恢复; 3.杀虫剂本身到达作用部位前要有足够的稳定性。 X基团——吸电子基团,满足1 RO2——供电子基团,满足2、3

66
(二)氨基甲酸酯类杀虫剂的作用机制 一种看法认为,氨基甲酸酯分子与AChE的酯动部位和结合部位形成一种比较稳定的复合物,从而使AChE失去活性。但在适当条件下,这种复合物又可分解,使酶复活。因此这种抑制属于可逆的竞争性抑制。抑制过程中,氨基甲酸酯和AChE没有发生真正的化学反应。 另一种看法则认为,和有机磷酸酯类杀虫剂一样,氨基甲酸酯和AChE发生了化学反应,生成了氨基甲酰化酶,从而抑制了AChE的活性。因此这种抑制属于不可逆的竞争性抑制。

67
反应过程 Kd是E·AX的离解常数或化合物和AChE的亲和力常数,Kd越小,表示复合物越稳定;K2 是氨基甲酰化速率常数,K2越大,表示氨基甲酰化反应越快;K3是氨基甲酰化酶水解速率常数, K3越大,表示酶的复活越迅速。 吴文君认为氨基甲酸酯对 AChE抑制,既是因为复合物的形成又是因为氨基甲酰化酶的生成。但抑制AChE,使昆虫中毒的主要原因是形成了稳定的复合物,生成的氨基甲酰化酶因不稳定,仅是次要原因。

68
注意 虽然氨基甲酸酯类杀虫剂和有机磷酸酯类杀虫剂一样,对AChE的抑制过程都可用同一通式表达;
但因其Kd、 K2、 K3三个常数相差很大,因而又有重要区别:前者抑制AChE主要依赖其有很小的Kd 值,而后者则依赖其较大的K2值,特别是较小的K3值。

69
(三)中毒和治疗 中毒原因 (1)神经冲动传导受阻 (2)对后膜不断刺激 死亡:呼吸困难,产生窒息?
中毒和治疗 中毒原因 (1)神经冲动传导受阻 (2)对后膜不断刺激 死亡:呼吸困难,产生窒息?")
70
治疗 阿托品: 蕈毒碱受体拮抗剂 ACh不能结合R 影响离子通道开放 可以解除由于过度兴奋出现的症状:流泪、排尿、瞳孔收缩。
恢复酶活性

71
2-PAM类(肟类解毒剂) 结合使用 阿托品——M 解磷定——(N) AChE的复活剂 解磷定 氯磷定
作用越酶,由于些脑屏障不能进入CNS。 结合使用 阿托品——M 解磷定——(N)
")
72
二、杀虫剂对乙酰胆碱受体的影响 沙蚕毒素类杀虫剂 烟碱类杀虫剂

73
1.沙蚕毒素杀虫剂的作用机制 在症状学方面和其他杀虫剂截然不同,不表现出一般神经毒剂引起的运动失调、过度兴奋和痉挛等,而是先使昆虫活动减少,失去取食能力,继而瘫痪死亡,作用缓慢。 进入虫体后,一般要在酶的作用下代谢成沙蚕毒素起作用,如巴丹。

74
主要是作用于突触后膜乙酰胆碱受体。 电生理实验结果表明,沙蚕毒素对突触前膜的反应无明显影响,而主要是减小乃至完全阻断了兴奋性突触后电位 (EPSP),使之不能产生动作电位,神经兴奋传导中断。 实验结果还表明,沙蚕毒素是和乙酰胆碱竞争性地占领乙酰胆碱受体,使乙酰胆碱失去作用的对象,但沙蚕毒素本身是乙酰胆碱的颉颃剂,它占领N型受体后并不发生离子通透性改变,不产生动作电位。 由于受体与离子通道是耦联关系,沙蚕毒素是通过占领受体而影响离子通道还是直接作用于离子通道,尚不清楚。

75
2 烟碱类杀虫剂 植物杀虫剂烟碱及高效杀虫剂吡虫啉也是作用于突触后膜上的N型乙酰胆碱受体。
2 烟碱类杀虫剂 植物杀虫剂烟碱及高效杀虫剂吡虫啉也是作用于突触后膜上的N型乙酰胆碱受体。 烟碱可以直接和受体结合并永久性地占领受体,干扰了乙酰胆碱和其受体的结合,阻碍了神经兴奋的传导。

76
三、杀虫剂对γ-氨基丁酸(GABA)受体的影响
有机氯杀虫剂 Avermectin杀虫剂 吡唑类杀虫剂

77
1. 有机氯杀虫剂的作用机制 20世纪70年代初,W.H.Ryan等认为林丹及环戊二烯类(如狄氏剂)有机氯杀虫剂对昆虫而言,主要作用于中枢神经系统的以乙酰胆碱为神经递质的突触前膜,促进乙酰胆碱小泡过多地释放乙酰胆碱,带来的后果和有机磷杀虫剂抑制AChE的后果相似,造成突触后膜乙酰胆碱的积累。 但这些杀虫剂如何刺激小泡过多释放乙酰胆碱,其机理并不清楚。

78
20世纪80年代以来,Matsumura等,Abalis等,Casida等认为林丹、硫丹及环戊二烯类杀虫剂作用于GABA受体,抑制了氯离子的内流,导致中枢神经的兴奋和痉挛。
突触前膜释放的GABA和后膜的GABAA 受体结合后将使氯通道开放,氯离子迅速涌入膜内,使膜超极化,产生抑制性突触后电位。由于GABAA 受体上除GABA的结合位点外,还存在共它配体的结合位点,而这些位点被占领亦可影响氯通道的开闭。林丹等杀虫剂可能和GABAA 受体上的苦毒宁位点相结合,阻断氯离子的内流,从而干扰了昆虫的神经传导

79
新型杀虫剂锐劲特(Fipronil),能有效地防治土壤和作物害虫。
2. 吡唑类杀虫剂 新型杀虫剂锐劲特(Fipronil),能有效地防治土壤和作物害虫。 Fipronil也是GABAA受体的抑制剂,阻断了氯离子的内流。 3 Avermectin杀虫剂 激活GABAA型受体或GABA门控的氯通道,造成超极化效应。 Avermectin不同于林丹、硫丹及环戊二烯类杀虫剂作用的另一个是事实它们之间无交互抗性。 但阿维菌素中毒的昆虫不表现过度的兴奋症状,而是以麻痹为主要症状。但是关于引起氯离子内流的具体机制尚有不同看法:①阿维菌素刺激突触前膜,过多地释放GABA;②阿维菌素直接作用于氯通道,打开氯通道;③阿维菌素也影响了其他配体门控的氯通道
,能有效地防治土壤和作物害虫。 Fipronil也是GABAA受体的抑制剂,阻断了氯离子的内流。 3 Avermectin杀虫剂. 激活GABAA型受体或GABA门控的氯通道,造成超极化效应。 Avermectin不同于林丹、硫丹及环戊二烯类杀虫剂作用的另一个是事实它们之间无交互抗性。 但阿维菌素中毒的昆虫不表现过度的兴奋症状,而是以麻痹为主要症状。但是关于引起氯离子内流的具体机制尚有不同看法:①阿维菌素刺激突触前膜,过多地释放GABA;②阿维菌素直接作用于氯通道,打开氯通道;③阿维菌素也影响了其他配体门控的氯通道.")
80
四、杀虫剂对轴突传导的干扰 DDT的作用机制 拟除虫菊酯类杀虫剂的作用机制

81
1 、DDT的作用机制 关于DDT的作用机制,目前主要有两个学说: 钠离子通道作为DDT的靶标的学说 抑制外Ca++ —ATP酶的学说。

82
(1)钠离子通道作为靶标的学说 DDT主要作用于外周神经系统,尤其是感觉神经。
电生理的研究发现,DDT的第一个重要作用就是用DDT处理的神经,一个刺激引起了一系列的放射,即重复后放,这一重复后放阶段相应于中毒症状的兴奋期;接着产生了不规则的,甚至不需刺激也引起的放射,这相应于痉挛期;重复后放的减弱以至停止阶段相应于麻痹期。因此关健问题是解释重复后放。

83
正相没有下降到超极化水平,而负后电位延长与振幅增加了。这种负后电位延长及振幅增加,使膜电位超过了引起动作电位所要求的阈值电位,因此就自发地引起了另一个动作电位。
假如第二个动作电位也同样引起负后电位的增加并超过阈值电位,那就会再引起一个放射。实验还证实DDT处理后,引起动作电位的阈值电位逐渐下降,因此重复后放越来越容易。在痉挛期小刺激也会引起重复后放。

84
DDT处理后,负后电位为什么会延长并加强呢?采用电压钳制技术实验,结果说明,负后电位的延长与加强主要是由于DDT影响了钠电流,使之在下降阶段(灭活阶段)变得迟缓。
根据这一事实提出了Na+通道作为靶标的学说,即认为DDT影响了Na+通道,使之关闭延迟,因而延长了钠电流

85
DDT主要作用于兴奋组织(神经、肌肉)的轴突膜。
在膜上有种DDT受体,这种受体不同于其他类型的受体(如AChE一般是结合蛋白),它是膜上的一个空隙。 DDT及其类似物只要适合进入这个空隙,就会引起与之偶联的离子通道(主要是Na+通道)的改变,结果是延迟了Na+通道的关闭,形成了一个延长的、振幅加大的负后电位。 增大的负后电位超过了阈值电位,因而引起了一系列重复后放。 重复后放的结果导致产生了一种神经毒素的释放,使得昆虫最终死亡。
,它是膜上的一个空隙。 DDT及其类似物只要适合进入这个空隙,就会引起与之偶联的离子通道(主要是Na+通道)的改变,结果是延迟了Na+通道的关闭,形成了一个延长的、振幅加大的负后电位。 增大的负后电位超过了阈值电位,因而引起了一系列重复后放。 重复后放的结果导致产生了一种神经毒素的释放,使得昆虫最终死亡。")
86
(2)外Ca++—ATP酶作为靶标的学说 外Ca++—ATP酶处于细胞膜或神经膜的外表。它以ATP分解产生的能量来调节膜外的Ca++浓度。当外部Ca++浓度低时,这个酶就被活化,而当外部Ca++浓度达到一定水平时,它就停止工作或积极去除Ca++。 对DDT敏感的是外Ca++—ATP酶,DDT的作用就是抑制了这种外Ca++—ATP酶活性,导致了轴突膜外表的Ca++浓度降低,从而使得阈值电位降低,更易引起重复后放。 这个酶对DDT十分敏感,而且 在低温时更易被DDT抑制,这也可以说明DDT的负温度系数(即DDT的同一剂量对昆虫的毒力在低温时,如15℃,比在高温时,如30℃,还强大)。
。")
87
(3)DDT引起神经毒素的释放 实验证明,一切刺激(如拟除虫菊酯类、有机磷及其他有机氯杀虫剂的刺激乃至机械的刺激)都可能导致神经分泌这种毒素。 1959年Sterrburg初步分离和鉴定了这种毒素,认为是个芳香胺,而It的工作认为是L-亮氨酸及其脱羧代谢产物展异戊胺。 张宗炳等研究认为,这个毒素是酪胺。酪胺是一种不正常的神经胺(在正常情况下仅痕量存在),它和正常的神经胺起竞争作用而干扰了神经兴奋传导。在神经毒剂作用下,酪氨酸脱羧酶活性大大提高,由此形成了大量酪胺,巴丹、杀虫脒、西维因等并不产生这种神经毒素。 必须指出,这两个学说并不矛盾。对外Ca++—ATP酶的抑制可能与Na+通道的改变是相辅相成的。已知外Ca++控制着膜电位和Na+通透性之间偶联的程度,只要增加Ca++浓度就可以降低Na+的通透性。此外,外膜上高浓度的Ca++可以保护 Na+通道不被DDT“撑开”,而浓度低时,则易被“撑开”。
,它和正常的神经胺起竞争作用而干扰了神经兴奋传导。在神经毒剂作用下,酪氨酸脱羧酶活性大大提高,由此形成了大量酪胺,巴丹、杀虫脒、西维因等并不产生这种神经毒素。 必须指出,这两个学说并不矛盾。对外Ca++—ATP酶的抑制可能与Na+通道的改变是相辅相成的。已知外Ca++控制着膜电位和Na+通透性之间偶联的程度,只要增加Ca++浓度就可以降低Na+的通透性。此外,外膜上高浓度的Ca++可以保护 Na+通道不被DDT 撑开 ,而浓度低时,则易被 撑开 。")
88
2.拟除虫菊酯类杀虫剂的作用机制 拟除虫菊酯类杀虫剂的作用机制比较复杂,还有许多问题不清楚。昆虫在不同剂量作用下分别产生忌避、击倒、拒食和毒杀效果,而且不同的分子结构其作用机制也不尽相同。Cammon等(1981)按其分子中有无α-CN基将拟除虫菊酯 类杀虫剂分成两种类型:
按其分子中有无α-CN基将拟除虫菊酯 类杀虫剂分成两种类型:")
89
除虫菊酯类杀虫剂两种类型 Ⅰ型:包括天然除虫菊酯、胺菊酯、丙烯菊酯及二氯苯醚菊酯等。
这一类型的作用机制和DDT相似,外周神经系统对其最为敏感,主要作用于神经膜,改变了膜的通透性,特别是延迟了Na+通道的关闭,负后电位延长并加强,导致产生重复后放,中毒昆虫表现为高度兴奋及不协调运动。 Ⅱ型:包括溴氰菊酯、氯氰菊酯、杀灭菊酯等。 这一类型虽然也影响神经膜上Na+通道,但并不引起重复后放,反而阻断兴奋的传导,中毒昆虫不表现为高度兴奋,而是很快就产生痉挛并进入麻痹状态。

90
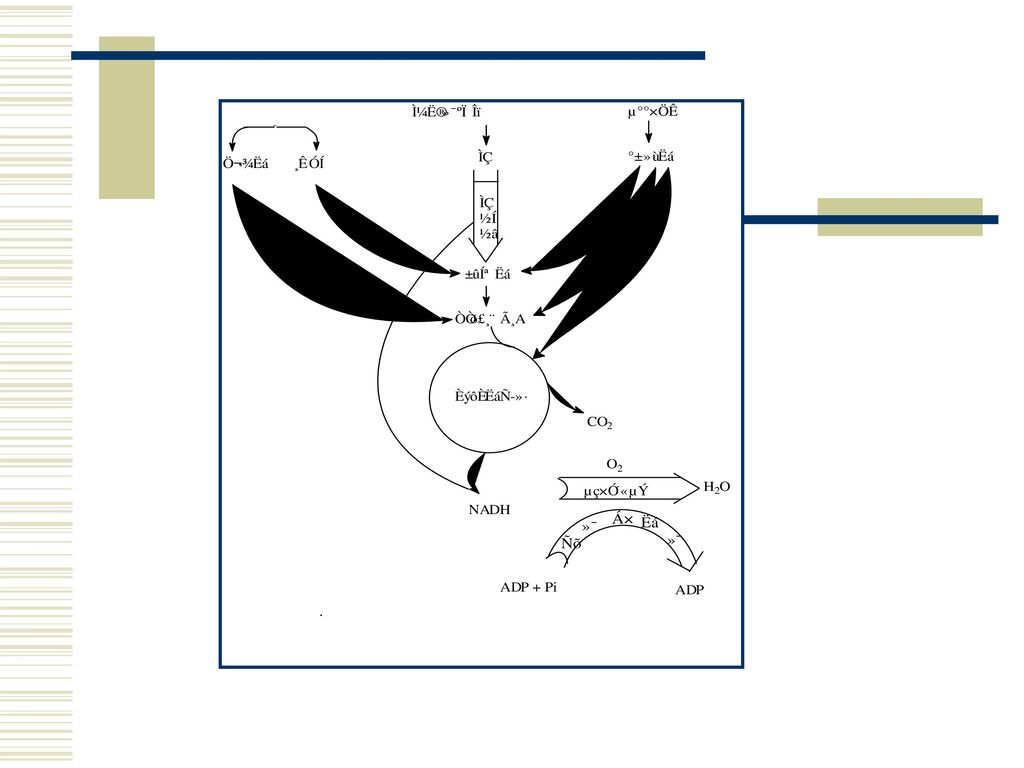
五 昆虫呼吸毒剂的作用原理 昆虫的生命过程所需的能量来自碳水化合物、脂肪和蛋白质的氧化代谢。
主要通过生成乙酰辅酶A而进入三羧循环,转入呼吸链进行氧化磷酸化,最终生成ATP。

91
昆虫细胞内的呼吸代谢过程 第一阶段:糖、脂肪、蛋白质大部分转变为乙酰辅酶A 第二阶段:从乙酰辅酶A开始的三羧酸循环
第三阶段: TCA产生的氢原子通过NAD-NADH转移给黄素蛋白及细胞色素系统,即电子转一阶段 第四阶段:电子转移的同时进行氧化磷酸化作用

94
杀虫剂对呼吸链中电子传递作用示意图

95
1 鱼藤酮的作用机制 鱼藤酮及其类似物是植物杀虫剂鱼藤、非洲山毛豆、紫穗槐等的主要杀虫有效成分。
1 鱼藤酮的作用机制 鱼藤酮及其类似物是植物杀虫剂鱼藤、非洲山毛豆、紫穗槐等的主要杀虫有效成分。 鱼藤酮有较强的触杀及胃毒作用。昆虫中毒后,中毒症状表现很快,但死亡过程却极为缓慢,往往要数天才能逐渐死亡。如家蝇,受药后约30min就可以表现中毒症状:耗氧量显著降低,心跳加速又逐渐减弱;引起神经传导和肌肉收缩受阻,麻痹、瘫痪直至死亡。

96
鱼藤酮是典型的细胞呼吸代谢抑制剂; 主要作用于呼吸链中电子转移复合体I,中断了从辅酶I到辅酶Q之间的电子传递,从而使呼吸受阻。 鱼藤酮在神经和肌肉组织中抑制呼吸,还有一部分作用是由于抑制L—谷氨酸的氧化作用。L—谷氨酸是神经组织呼吸时被氧化的氨基酸,鱼藤酮对L—谷氨酸氧化的抑制使神经机能受阻,造成昆虫麻痹和瘫痪。

97
2. 磷化氢的作用机制 磷化氢是应用最广泛的熏蒸杀虫剂。Bond等曾对磷化氢作用的症状进行了如下的描述:中毒昆虫首先表现为腿的震颤,气门关闭,氧消耗急剧下降。昆虫步态失去平衡,击倒,麻痹,身体痉挛,呼吸被抑制,心跳逐渐停止,最后昆虫死亡 。 呼吸链末端氧化酶复合体的细胞色素C氧化酶是其作用靶标

98
六、杀虫剂对昆虫的其它作用 甲脒类杀虫剂的作用机制 几丁质合成抑制剂的作用机制 昆虫保幼激素和蜕皮激素类似物作用机制

99
1、 甲脒类杀虫剂的作用机制 杀虫脒对昆虫的效应也比较特殊,对鳞翅目幼虫主要是忌避、拒食等行为的反应,对成虫有一定的触杀及忌避产卵作用,而对红蜘蛛有直接的触杀作用。 典型的神经中毒症状。 除杀虫脒、双甲脒、单甲脒等甲脒类杀虫剂外,一些咪唑啉类、唑啉类、噻唑啉类、碳二亚胺类、二嗪类杀虫杀螨剂也是OA受体的激活剂,具有和甲脒类相似的作用机制。

100
神经中毒机理 局部麻醉作用。在高剂量下,杀虫脒作用于轴突膜,主要是阻塞了Na+通道,也在一定程度上阻塞了K+通道,从而不产生动作电位,没有兴奋在轴突上传导,这就是局部麻醉作用。 对章鱼胺受体的激活作用,

101
对章鱼胺受体的激活作用 昆虫交感神经末梢和平滑肌(或腺体)构成的突触,其后膜存在章鱼胺(OA)受体。
甲脒类杀虫剂是OA受体的激活剂,甲脒类和后膜上的OA受体,主要是OA2受体相结合,引起与受体耦联的腺苷酸环化酶(ACE)活化,从而使三磷酸腺苷 (ATP)转化为环化腺苷酸(cAMP),cAMP又活化蛋白激酶。 蛋白激酶使许多种活性蛋白(包括酶及受体)磷酸化,从而产生各种生理生化效应,干扰了昆虫神经兴奋的正常传导,引起一系列昆虫行为的改变,如增加活动性,不断发抖,昆虫从植株上跌落而无法取食等。
活化,从而使三磷酸腺苷 (ATP)转化为环化腺苷酸(cAMP),cAMP又活化蛋白激酶。 蛋白激酶使许多种活性蛋白(包括酶及受体)磷酸化,从而产生各种生理生化效应,干扰了昆虫神经兴奋的正常传导,引起一系列昆虫行为的改变,如增加活动性,不断发抖,昆虫从植株上跌落而无法取食等。")
102
甲脒类杀虫剂还有双甲脒 制剂:20%螨克乳油 双甲脒又称“螨克”,主要用于农作物及家畜的蜱螨类的防治,以触杀作用为主。对鳞翅目害虫的卵也有活性。推荐用量为:棉花田用有效成分300~1 000g/hm2、柑桔园用有效成分10~60 g/hm2、高级水果园(topfruit)用有效成分40~80 g/hm2,还可用于葫芦、啤酒花等作物防治叶螨类。
用有效成分40~80 g/hm2,还可用于葫芦、啤酒花等作物防治叶螨类。")
103
2. 几丁质合成抑制剂的作用机制 以破坏昆虫表皮几丁质沉积为主要症状的IGR称为昆虫几丁质合成抑制剂,主要包括苯甲酰脲类,如除虫脲、灭幼脲、氟虫脲、定虫隆等,噻嗪酮和灭蝇胺虽然不具备苯甲酰脲结构,但其具有类似的症状,因此也归此类。

104
中毒昆虫首先表现为活动减少、取食降低,到蜕皮或变态时才表现出明显的中毒症状:旧表皮不能蜕掉或不能完全蜕掉而死亡;
形成的新表皮很薄,易裂开,体液外流; 老熟幼虫不能化蛹,或形成半幼虫一半蛹,或半蛹一半成虫畸形而死亡。

105
作用机制 大量的文献都报道苯甲酰脲类杀虫剂对害虫表皮的破坏,发现几丁质的沉积受到抑制的同时,有N一乙酰氨基葡萄糖的积累,因此称为几丁质合成抑制剂。 但关于几丁质合成被抑制的机理,只提出过几种假说。 典型的假说是苯甲酰脲类杀虫剂抑制了几丁质合成酶的活性,从而几丁质合成被抑制。但一方面没有实验证实苯甲酰脲类杀虫剂对几丁质合成酶有直接抑制作用,事实上在离体条件下,苯甲酰脲类对几丁质合成酶活性没有影响,另一方面,苯甲酰脲类中毒症状并不限于影响几丁质沉积,而是对昆虫全身性多方面的影响,包括对DNA、RNA及蛋白质合成的影响等。

106
3 昆虫保幼激素和蜕皮激素类似物的作用机制 一般认为昆虫脑接受外界环境与内在刺激后,引起脑的神经分泌细胞活动,释放脑激素。脑激素激活前胸腺分泌蜕 皮激素,激活咽侧体分泌保幼激素。 由于保幼激素的作用,使幼虫不断生长发育,保持幼虫性状; 由于蜕皮激素的作用,而引起若虫或幼虫蜕皮。两种激素的协调作用,昆虫幼虫的生长发育便完成。

107
当幼虫到最后一龄时,咽侧体停止分泌或很少分泌,而前胸腺照常分泌,因而发生变态,出现蛹或成虫
昆虫的蜕皮、变态过程,表现出幼虫、蛹或成虫的形式,就是通过上述两种激素来控制效应器官的代谢过程。如果只有蜕皮激素,被活化的效应器官就发育成成虫;如果除蜕皮激素外,尚有少量保幼激素,则发育成蛹;如果二者同时起作用,则仍发育成幼虫(龄间蜕皮)。成虫期又需要保幼激素以促进卵巢发育。
。成虫期又需要保幼激素以促进卵巢发育。")
108
保幼激素类似物 主要是抑制脂肪酸合成和脂肪酸氧化,破坏β-氧化;破坏RNA的积累,并由此抑制了与几丁质合成的有关基因的表达。
吡丙醚(蚊蝇醚)
")
109
蜕皮激素类似物 非甾醇双酰肼类RH- 5849(抑食肼)、RH-5992(米螨)主要机制是竞争性地占领蜕皮激素受体,并抑制羽化激素,从而干扰了昆虫的正常生长发育。
、RH-5992(米螨)主要机制是竞争性地占领蜕皮激素受体,并抑制羽化激素,从而干扰了昆虫的正常生长发育。")
110
第四节 有机磷杀虫剂 20世纪30年代,德国的G.Schrader就合成了一系列有机磷酸酯类化合物,并发现一些具有杀虫杀螨活性,于1937年提出下面的通式: 1941年合成第一个植物内吸杀虫剂八甲磷。 1944年合成对硫磷。 目前全世界有机磷杀虫剂品种约100余种,常用50多种。 我国农药市场上有机磷品种30余个,占我国杀虫剂品种的38%,但产量却占75%。

111
一、 类型 K.Hassall按其特点将有机磷杀虫剂分成下述几类: 短效型 内渗型 内吸型

112
1.短效型 典型代表: (phoxim) 、(dichlorvos):
基于同样的原因,在目标昆虫和非靶标生物,包括哺乳动物之间无明显的选择性。 适合于需要迅速杀死害虫而且施药后短期即将收获的作物,如茶叶,桑叶,蔬菜等害虫的防治。

113
2.内渗型 水溶性差而脂溶性强,其稳定性差异较大,但施药后在植物叶片能持效数天乃至数十天。
由于其亲脂性强,这类化合物可溶入叶片的蜡质层并作短距离扩散,有时甚至可渗到叶片背面。这种性质不仅可以减少雨水冲刷,而且对防治低位势生长植物(如草莓)叶背危害的害虫有利,因为这种情况下,叶背的害虫不会被直接喷上杀虫剂。 因而又称作持效型触杀剂(persistent contact poison)
叶背危害的害虫有利,因为这种情况下,叶背的害虫不会被直接喷上杀虫剂。 因而又称作持效型触杀剂(persistent contact poison)")
115
3.内吸型 分配系数适合于穿透植物表面蜡质层及细胞质膜,既可以从根部被吸收向顶传导,亦可被叶部吸收向下传导。
这种内吸性不仅可以减少施药后雨水的冲刷,而且可以减少对天敌的杀伤,加之合适的剂型和施药技术的配合,有利于保护和利用害虫天敌。 内吸型有机磷杀虫剂特别适合于刺吸式口器害虫如蚜虫、蓟马及植食性螨的防治。 内吸型的杀虫剂都具有很高的毒性,在我国已禁止生产和使用。

117
二、 有机磷杀虫剂的特征 化学性质不稳定。易水解,在碱性条件下易分解,因而不宜和碱性物质混合;易氧化,热分解,易于在自然环境中或动植物体内降解,在高等动物体内无累积毒性,正确使用时残留问题小,不致污染环境。 对害虫高效(毒力高于有机氯杀虫剂),广谱。作用方式多样。 化学结构变化无穷,品种多,适用范围广。
,广谱。作用方式多样。 化学结构变化无穷,品种多,适用范围广。")
118
毒性差异大。此外,对有机磷杀虫剂引起的急性中毒有特效的解毒药(解磷定和阿托品)。
除少数品种外(如敌百虫、敌敌畏对高粱、瓜类敏感),一般对农作物安全,在推荐剂量下不致发生药害。 和有机氯、特别是和拟除虫菊酯类杀虫剂相比,害虫对有机磷杀虫剂的抗药性发展缓慢。 5. 6.
,一般对农作物安全,在推荐剂量下不致发生药害。 和有机氯、特别是和拟除虫菊酯类杀虫剂相比,害虫对有机磷杀虫剂的抗药性发展缓慢。 5. 6.")
119
三、 有机磷杀虫剂研究开发动向 不对称有机磷杀虫剂 引入杂环

120
1. 不对称有机磷杀虫剂 为了对付害虫抗药性问题,更加注重以磷原子为中心的不对称有机磷杀虫剂的开发。
据研究,这些不对称化合物与以往大多数对称型品种之间较少发生交互抗性,而且引入不对称因素后,其毒性与药效均有明显改变。 毒虫畏大鼠口服LD50为130~150 mg/kg,防治稻叶蝉剂量150 g/ha。灭虫畏大鼠口服LD50为10~39 mg/kg,防治稻叶蝉剂量600~2250 g/ha。

122
特别是丙硫基不对称型硫赶磷酸酯杀虫剂的成功开发,可以说是有机磷杀虫剂发展史上的重大事件。
这类化合物不但对敏感害虫品系有良好防效,而且对抗性品系也同样表现优异防效,还明显降低了对高等动物的毒性。 典型品种如丙硫磷(prothiophos)、丙溴磷(profenofos): 丙硫磷:大鼠口服LD50为1730~1800 mg/kg,经皮LD50为4100~4170mg/kg,属低毒级,对磷翅目幼虫特效。
、丙溴磷(profenofos): 丙硫磷:大鼠口服LD50为1730~1800 mg/kg,经皮LD50为4100~4170mg/kg,属低毒级,对磷翅目幼虫特效。")
123
2 引入杂环 由于杂环往往具有很高的生物活性,因此近年来将杂环引入磷酸酯,合成了许多化合物,开发了不少新品种,显示了优异的杀虫活性。

124
四、主要品种 磷酸酯及膦酸酯 一硫代磷酸酯 二硫代磷酸酯 硫酰胺和硫代硫酰胺

125
(一)磷酸酯及膦酸酯
磷酸酯及膦酸酯")
126
敌敌畏(dichlorvos) 理化特性 室温下水中的溶解度约为10 g/L,在煤油中溶解度2~3%,能与大多数有机溶剂和气溶胶推进剂混溶。
在碱性溶液中水解更快 。

127
生物活性 高效,速效,光谱的有机磷杀虫剂,用途极为广泛,无论农业,牧业,粮仓及一般商品仓库,以及环境卫生都有应用。
具有触杀,胃毒和熏蒸作用,对咀嚼式口器害虫和刺吸式口器害虫均有良好的防治效果。敌敌畏的蒸气压较高,对害虫有极强的击倒力,对一些隐蔽性的害虫如卷夜蛾幼虫也具有良好效果。 持效期短,适用于防治棉花,果树,蔬菜,甘蔗,烟草,茶,桑等作物上的多种害虫,对蚊,蝇等卫生害虫以及空仓对米象,谷盗等有良好防治效果 敌敌畏对雌,雄大鼠急性经口LD50分别为56mg./kg和80mg./kg,急性经皮LD50分别为75mg./kg和107mg./kg,

128
制剂 50%,80% 敌敌畏乳油 28%敌敌畏油脂缓释剂 22%,30%敌敌畏烟剂

129
使用方法 80%敌敌畏乳油对水800~1500倍喷雾可防治水稻,棉花,果树,甘蔗,烟草,茶,桑等作物上的多种害虫。
例如蔬菜黄曲条跳甲,菜青虫,茶毛虫,水稻叶蝉,飞虱,斗天蛾,苹果卷叶虫,苹果巢蛾,梨星毛虫,桃小食心虫,烟青虫,甘蔗绵蚜等 80%乳油空仓杀虫防治米象,谷盗,大谷盗,长角谷盗,黑菌虫,麦蛾等仓库害虫用1000倍液喷洒,施药后密闭时间为2~3天,效果显著. 敌敌畏杀虫作用的大小与气候条件有直接关系,气温高时,杀虫效力较大 ;

130
注意事项 敌敌畏在一般浓度下对玉米,高粱易发生药害,苹果开花后喷射浓度高于1200倍者,易发生药害

131
(二)一硫代磷酸酯
一硫代磷酸酯")
132
辛硫磷(倍腈松,phoxin) 理化特性 易溶于醇,酮,芳烃,卤代烃等有机溶剂,稍溶于脂肪烃,植物油和矿物油。
辛硫磷易光解,在中性和酸性介质中易分解。原药为红棕色油状液体。

133
生物活性 广谱的有机磷杀虫剂,具有强烈的触杀和胃毒作用。
主要用于防治地下害虫,适宜于防治花生,小麦,水稻,棉花,玉米等作物的害虫,特别是甘蔗,果树,蔬菜,桑,茶等害虫,还可防治蚊,蝇,等卫生害虫及仓储害虫。特别对防治花生,大豆,小麦的蛴螬,蝼蛄有良好的效果。 辛硫磷对哺乳动物的毒性很低。对雌,雄大鼠大鼠急性经口LD50分别为2170mg/kg和1976mg/kg,雄大鼠急性经皮LD50为10200 mg/kg.

134
制剂 40%辛硫磷乳油 2.5%辛硫磷微粒剂 3.6%辛硫磷大粒剂。

135
使用方法 防治水稻二化螟,黑尾叶蝉,褐飞虱,用40%乳油对水500~800倍喷雾;防止菜青虫,以2000~5000倍液喷雾;
防治棉铃虫,红铃虫,棉蚜等以800~1000倍液喷雾; 防治蛴螬,蝼蛄采用种子处理法,小麦用40%乳油500mL价税25~50kg;玉米,高粱,大豆用40%乳油500mL,加税20kg,拌种子200kg.

136
(三)二硫代磷酸酯
二硫代磷酸酯")
137
马拉硫磷(马拉松,malathion) 理化特性 室温下微溶于水,溶解度为145 mg/L,能与多种有机溶剂互溶 对光稳定,对热稳定性差
在中性反应中稳定,但在pH7.0以上或pH5.0以下几迅速分解 不能与碱性农药混用.

138
生物活性 具有良好的触杀,胃毒作用和微弱的熏蒸作用;适用于防治水稻,高粱,蔬菜,果树等作物上的咀嚼式口器和刺吸式口器害虫,还可用来防治蚊,蝇等家庭卫生害虫,体外寄生虫和人的体虱,头虱。 雌,雄大鼠急性经口LD50分别为1751.5mg/kg和1634.5mg/kg,大鼠急性经皮LD50为4000~6150mg/kg。对蜜蜂高毒,对眼睛,皮肤有刺激性.

139
制 剂 45%马拉硫磷乳油 25%马拉硫磷油剂 70%优质马拉硫磷乳油(防虫磷) 1.2%马拉硫磷粉剂 1.8%马拉硫磷粉剂
 1.2%马拉硫磷粉剂 1.8%马拉硫磷粉剂")
140
使用方法 45%马拉硫磷乳油对水稀释2000倍喷雾可防治菜蚜; 棉蚜,棉蓟马,稀释1000倍左右防止菜青虫,棉红蜘蛛,棉椿象等;
其他害虫:棉花害虫1500~2000倍,水稻害虫2000倍,蔬菜害虫为1000~2000倍,果树害虫800~2000倍,大田作物害虫2000~3000倍。 瓜蕾和番茄幼苗对该药较敏感,不能使用高浓度药液。

141
丙硫磷(prothiofos) 理化特性 难溶于水,能与环己酮,甲苯完全互溶; 对光稳定,在酸性,碱性介质中较稳定。
 理化特性 难溶于水,能与环己酮,甲苯完全互溶; 对光稳定,在酸性,碱性介质中较稳定。")
142
生物活性 丙硫磷是广谱低毒有机磷杀虫剂,对鳞翅目幼虫有特效。具触杀和胃毒作用;
主要用于甘蓝,柑桔,烟草,菊花,樱花,和草坪等,可有效防治菜青虫,小菜蛾,甘蓝夜蛾,蚜虫,卷夜蛾,粉蚧,斜纹夜蛾,烟青虫,和美国白蛾等多种害虫。也能防治蚊蝇等卫生害虫。 丙硫磷对雄大鼠急性经口LD50分别为925~966mg/kg,急性经皮LD50为1300mg/kg.

143
制剂 使用方法 50%丙硫磷乳油 40%丙硫磷可湿性粉剂。 50%丙硫磷乳油对水稀释800~1000倍喷雾防治蔬菜害虫。

144
(四)硫酰胺和硫代硫酰胺
硫酰胺和硫代硫酰胺")
145
乙酰甲胺磷(acephate) 理化特性 易溶于水(约6.5 g/L),甲醇,乙醇,丙酮,等极性溶剂和二氯甲烷,二氯乙烷登卤代烷烃类,在苯,甲苯,二甲苯中的溶解度较小,在醚中溶解度很小 ; 低温时储藏相当稳定; 在酸性介质中很稳定,在碱性介质中易分解。

146
生物活性 内吸性广谱杀虫剂,具胃毒,触杀作用,并可杀卵;持效期长,是缓效型杀虫剂。
主要防治稻飞虱,叶蝉,蓟马,稻纵卷叶螟,棉小象鼻虫,棉铃虫,果树小食心虫,菜青虫,小菜蛾,粘虫,和各种蚜虫等等。 乙酰甲胺磷对大鼠急性经口LD50为823mg/kg 。

147
制剂 30%乙酰甲胺磷乳油; 40%乙酰甲胺磷乳油.

148
使用方法 对人畜毒性低,杀虫效果高,适合防治棉,粮,油,蔬菜,茶,桑,果树,甘蔗,烟草,牧草等作物的害虫 ;
30%乙酰甲胺磷乳油对水稀释500~1000倍喷雾防治菜青虫,小菜蛾,棉蚜,棉小造桥虫,桃小食心虫,梨小食心虫,粘虫,烟青虫等等;1000倍液喷雾防治蔬菜蚜虫;300~500倍防治稻纵卷叶螟,棉铃虫,棉红铃虫,柑桔介壳虫等 。

149
(五)含杂环的有机磷
含杂环的有机磷")
150
毒死蜱(乐斯本,chlopyrifos) 广谱杀虫,杀螨剂,具有胃毒和触杀作用,在土壤中挥发性较高;
适用于防治柑桔,棉花,玉米,苹果,梨,水稻,花生,大豆,小麦及茶树等多种作用的害虫和螨类,也可用于防治蚊,蝇等卫生害虫和家畜的体外寄生虫; 毒死蜱对大鼠急性经口LD50为63mg/kg,急性经皮LD50>2000mg/kg,对眼睛有轻度刺激,对皮肤有明显刺激。

151
制剂 40%,48%毒死蜱乳油; 40.7%乐斯本乳油; 14%毒死蜱颗粒剂。

152
使用方法 防治稻瘿蚊,柑桔潜叶蛾,小麦黏虫,桃蚜,介壳虫,桃小食心虫,茶尺蠖,小绿叶蝉,茶叶瘿螨,用40%毒死蜱乳油800~1500倍稀释液喷雾; 防治棉蚜,棉红蜘蛛,稻纵卷叶螟,茶毛虫,茶刺蛾用40%乳油1000倍液喷雾; 防治棉铃虫,棉红铃虫.小菜蛾,甜菜叶蛾,用40%乳油500~1000倍稀释液喷雾。

153
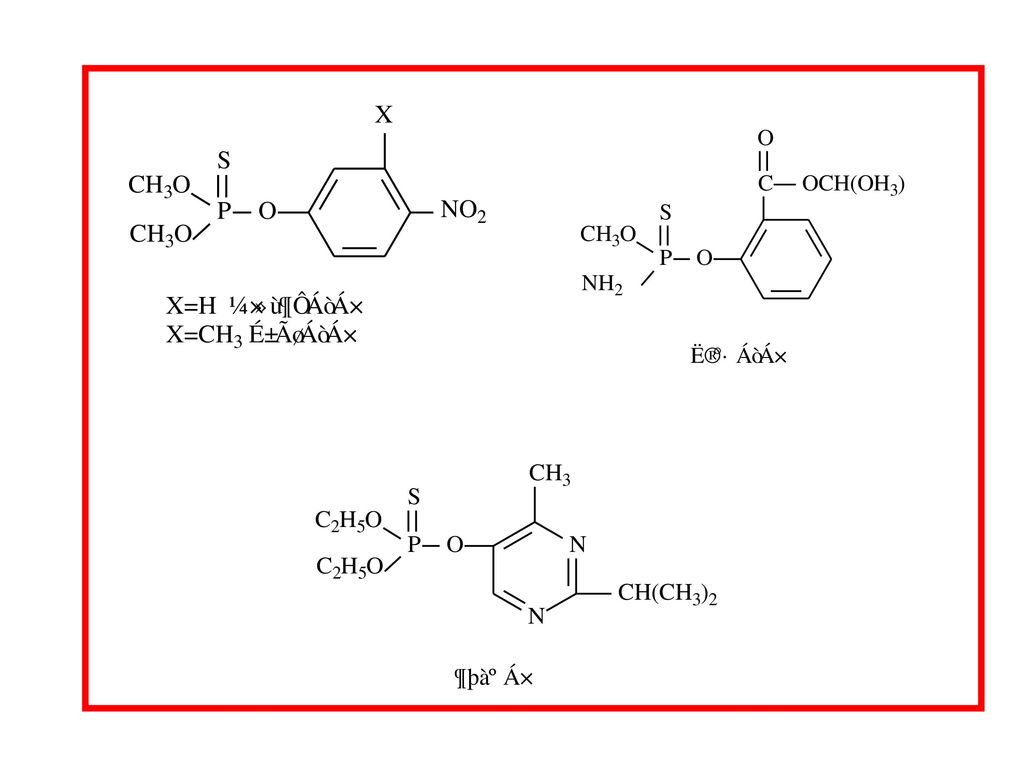
Main metabolic pathways involved in the biotransformation of parathion
Main metabolic pathways involved in the biotransformation of parathion. Reaction (1) is a bioactivating reaction, the other are detoxifying reactions. (1)Oxidative desulfuration; (2) oxidative dearylation; (3) oxidative deethylation; (4) reduction of the nitro group; (5) hydrolysis. CYP, cytochrome P450; PON1,paraoxonase.
 is a bioactivating reaction, the other are detoxifying reactions. (1)Oxidative desulfuration; (2) oxidative dearylation; (3) oxidative deethylation; (4) reduction of the nitro group; (5) hydrolysis. CYP, cytochrome P450; PON1,paraoxonase.")
154
Schematic representation of biochemical interactions between OPs and AChE. Reaction 1 leads to phosphorylated AChE. Reaction 2 is spontaneousreactivation of AChE. The rate of this reaction can be accelerated by oximes. Reaction 3 is the aging and leads to a stable, negatively charged phosphorylatedAChE. E-OH, active site of the enzyme.

155
第四节 氨基甲酸酯类杀虫剂 氨基甲酸酯类杀虫剂开发的基础是对天然毒扁豆碱(physostigmine)结构和活性的研究。
真正商品化的第一个品种乃是美国联合碳化公司1953年合成、1956年开发的甲萘威。 到20世纪60年代,氨基甲酸酯类杀虫剂的开发进入顶盛时期,几十个品种相继商品化,形成一大类杀虫剂。

156
一、 类型 按化学结构,可将氨基甲酸酯类杀虫剂分成3个类型。 1. N,N—二甲基氨基甲酸酯类 通式为

157
2. N—甲基氨基甲酸酯类 通式:

158
3. N—甲基氨基甲酸肟脂类 通式:

159
二、 氨基甲酸酯类杀虫剂的特征 大多数品种作用迅速,特效期短,选择性强。对叶蝉、飞虱、蓟马等防效好,而对螨类及介壳虫无效,一般对天敌比较安全; 大多数品种对高等动物毒性低,在生物体内及环境中易降解,但少数品种为剧毒,如克百威,涕灭威,只能加工成粒剂使用;

160
不同结构类型的品种,其生物活性和防治对象差别很大。如含萘环的甲萘威杀虫谱广,可防治棉铃虫,斜纹夜蛾,粘虫,棉蚜,棉蓟马,黄条跳甲及多种其它鳞翅目幼虫;含有苯并呋喃环的克百威,杀虫谱更广,还能杀线虫,而且具有强内吸性,但含有杂环的抗蚜威却主要用于防治除棉蚜以外的多种蚜虫; 多数对拟除虫菊酯类杀虫剂表现增效作用的增效剂,如氧化胡椒基丁醚,对氨基甲酸酯有显著的增效作用; 结构相对较简单,合成较容易,一种中间体、一套设备可同时生产多个产品。

161
三、 进展 近10多年来,最重要的进展是低毒化品种的研究取得重大突破。
三、 进展 近10多年来,最重要的进展是低毒化品种的研究取得重大突破。 对N—甲基氨基甲酸酯或N—甲基氨基甲酸肟酯类的高效高毒母体化合物的N原子上引入含硫基团或其它取代基,结果既保留了母体化合物对害虫高效的特点又降低了对哺乳动物的毒性,这类品种有丁硫克百威,硫双灭多威,丙硫克百威,棉铃威等。

163
硫双灭多威的毒性(小鼠急性口服LD50为325 mg/kg)仅为灭多威毒性(LD50为17 mg/kg)的1/18;
仅为灭多威毒性(LD50为17 mg/kg)的1/18;")
164
四、主要品种 灭多威(万灵,methomyl) 理化特性 能溶于水,丙酮,乙醇,异丙醇,甲醇,甲苯。在通常条件下稳定。在潮湿土壤中很易分解。
 理化特性 能溶于水,丙酮,乙醇,异丙醇,甲醇,甲苯。在通常条件下稳定。在潮湿土壤中很易分解。")
165
生物活性 内吸性广谱杀虫剂,通过触杀和胃毒作用杀灭害虫。可用于果树,蔬菜,棉花,苜蓿,烟草,草坪,观赏植物等。
叶面持效期短,其半衰期小于7天。对水稻螟虫,飞虱,以及果树害虫等都有很好的防治效果。 灭多威对雌、雄大鼠急性经口LD50分别为23.5mg/kg和17.0mg/kg, 兔急性经皮LD50>5000mg/kg,对皮肤无刺激作用,对眼睛有中等刺激作用。

166
制剂 使用方法 10%可湿性粉剂, 20%、90%可溶性粉剂 20%灭多威乳油 24%灭多威水剂
制剂 使用方法 10%可湿性粉剂, 20%、90%可溶性粉剂 20%灭多威乳油 24%灭多威水剂 使用剂量为有效成分为300~600mg/kghm2。 不可与碱性农药混用。

167
丁硫克百威(好安威,好年冬,carbosulfan)
理化特性 不溶于水,与丙酮,二氯甲烷,乙醇,二甲苯互溶。弱酸介质中易分解。 生物活性 克百威低毒化衍生物,杀虫谱广,有内吸性。 对大鼠急性经口LD50为209mg/kg,兔急性经皮LD50>2000mg/kg,对兔皮肤和眼睛有中等刺激作用。

168
制剂 20%好年冬乳油。 使用方法 20%好年冬乳油稀释800~1500倍喷雾防治节瓜蓟马。 蔬菜蚜虫大等,1000~1500倍防治柑桔潜叶蛾,蚜虫; 1500~2000倍防治柑桔绣壁虱。

169
第六节 拟除虫菊酯类杀虫剂 自1973年英国的Elliott发现光稳定的拟除虫菊酯在农业上推广使用后,80年代这类杀虫剂得到蓬勃发展。
第六节 拟除虫菊酯类杀虫剂 自1973年英国的Elliott发现光稳定的拟除虫菊酯在农业上推广使用后,80年代这类杀虫剂得到蓬勃发展。 目前,商品化的拟除虫菊酯杀虫剂品种近40个,占全世界杀虫剂销售额的19%,1994年达到15.6亿美元。

170
早在1800年,高加索人以除虫菊花粉防治卫生害虫。1909年,日本的Fujitami开始对其有效成分进行研究,但直到1924年瑞士化学家H
早在1800年,高加索人以除虫菊花粉防治卫生害虫。1909年,日本的Fujitami开始对其有效成分进行研究,但直到1924年瑞士化学家H.Staudinger和L.Ruzicka才提出除虫菊素Ⅰ和Ⅱ的初步分子结构,1947年才最后鉴定了结构。 现已研究清楚,天然除虫菊杀虫有效成分共6种,主要是除虫菊素Ⅰ和Ⅱ。

172
一、天然除虫菊素特点 酸部分即菊酸:菊酸I,菊酸II。 除虫菊素的化学结构有如下特征: 1.它是酯类化合物,由酸和醇两部分组成。
醇部分是3种环状醇酮:除虫菊酮醇,瓜叶除虫菊酮醇,茉莉除虫菊酮醇。

173
2.菊酸存在三元环结构,有顺反两种异构体。

174
菊酸中还有2个手性碳(C1和C3)。手性碳化合物有R、S两种空间结构,因此菊酸Ⅰ有4种异构体:
。手性碳化合物有R、S两种空间结构,因此菊酸Ⅰ有4种异构体:")
175
3.在醇部分,C4也是手性碳,亦有RS构型,它们若和菊酸Ⅰ 4个异构体结合,可形成除虫菊酯的8个异构体。
4.除虫菊素还有2个光不稳定中心,一是菊酸乙烯侧链上的偕二甲基,另一是醇部分的戊烯酮环和侧链的双键。这些化合物在光照下很快氧化而使之失去杀虫活性。

176
二、 农用光稳定型拟除虫菊酯的类型 1.菊酸和苯氧基苄醇组成的酯。

178
2.菊酸和非苯氧基苄醇组成的酯

179
3.非菊酸和苯氧基苄醇组成的酯 R为非三碳环烃基

180
4.醚类 分子结构已不是酯类,而是醚或肟醚,但仍保持菊酯类杀虫剂的生物学特性。

181
三、 拟除虫菊酯类杀虫剂的特征 广谱。 高效,速效。 对哺乳动物毒性低,使用安全,但大多数品种对蜜蜂、鱼类及天敌昆虫毒性大。
具有强大的触杀和胃毒,没有内吸作用,因此以叶面喷洒为主,很少作土壤处理或种子处理。 结构多属酯、醚,在生物体内及环境中易降解

182
四、 研究开发进展 1. 杀螨用菊酯类杀虫剂 70年代开发的拟除虫菊酯一般没有杀螨活性。
四、 研究开发进展 1. 杀螨用菊酯类杀虫剂 70年代开发的拟除虫菊酯一般没有杀螨活性。 1980年开发的甲氰菊酯,1985年开发的三氟氯氰菊酯,特别是1985年开发的联苯菊酯都有较好的杀螨活性,特别适合于棉花、果树虫、螨并发时防治,尤其是联苯菊酯,与溴氰菊酯,氯氰菊酯,氰戊菊酯等很少有交互抗性。 近年开发的杀螨菊酯(acninathrin)更是以杀螨为主,对成螨和若螨都很高效,且能兼治蚜虫、蓟马、卷叶蛾等;在结构上也很新颖,在环丙烷两侧各连有一个酯基: 自从1973年Elliott开发出第一个在农业上应用的光稳定氯菊酯(permethrin)以来,围绕防治害虫的实际需要,拟除虫菊酯类杀虫剂的研究与开发不断取得新进展。
更是以杀螨为主,对成螨和若螨都很高效,且能兼治蚜虫、蓟马、卷叶蛾等;在结构上也很新颖,在环丙烷两侧各连有一个酯基: 自从1973年Elliott开发出第一个在农业上应用的光稳定氯菊酯(permethrin)以来,围绕防治害虫的实际需要,拟除虫菊酯类杀虫剂的研究与开发不断取得新进展。")
183
1994年日本开发的halfenprox也是个菊醚类杀螨剂:

184
2. 稻田用菊酯类杀虫剂 70年代开发的菊酯类杀虫剂对鱼高毒,限制了在稻田的使用。
1986年开发的醚菊酯,其突出的优点就是对鱼类及其它水生动物毒性低,对稻田的害虫天敌也较安全,特别适合于稻田害虫防治。肟醚菊酯不单结构新颖(肟醚结构),兼有杀螨活性,而且也对鱼的毒性很低。
,兼有杀螨活性,而且也对鱼的毒性很低。")
185
3.含硅拟除虫菊酯的开发 日本住友公司80年代开发的以硅原子取代碳原子的含硅拟除虫菊酯。虽然在活性方面并未有大的突破,但在结构上开辟了一个新的领域。

186
4.其他进展 70年代开发的菊酯类杀虫剂对蜜蜂也高毒,而1983年开发的氟胺氰菊酯,在结构上属非环丙烷的拟除虫菊酯,其特点是:除杀虫外,杀螨活性也很高,对蜜蜂无毒杀、无驱避作用,除在农业上应用外,还用于蜂巢害螨的防治。 已往的菊酯类杀虫剂品种都不适合于防治地下害虫,而1987年开发的七氟菊酯,因其蒸气压较高(0.7×10-4Nm-2),主要用于土壤中地下害虫的防治。
,主要用于土壤中地下害虫的防治。")
187
五、主要品种 溴氰菊酯(deltamethrin,敌杀死,Decis) 理化特性
水中溶解度极低,易溶于丙酮,苯,二甲苯,二甲亚砜,环己酮和二恶烷等。对光照和热稳定,在酸性介质比在碱性介质中稳定。

188
生物活性 溴氰菊酯分子结构中含有三个不对称碳原子,即有八个立体异构体,其中单一右旋顺式异构体(1R,3R菊酸与S-a-氰醇合成的酯)是八个异构体中杀虫活性最高的。 溴氰菊酯具有很强的触杀作用,有一定的胃毒作用和拒避活性,无内吸及熏蒸作用。该剂是触杀活性最高的拟除虫菊酯杀虫剂,据报道其触杀毒力为DDT的100倍左右,西维因的80倍,马拉硫磷的50倍,对硫磷的40倍,生物苄芙菊酯的19倍(家蝇),氯菊酯的10倍。 昆虫对其易产生抗药性 大鼠急性经口LD50值为128.50~138.70mg/kg
,氯菊酯的10倍。 昆虫对其易产生抗药性. 大鼠急性经口LD50值为128.50~138.70mg/kg.")
189
制剂及使用 主要制剂:2.5%敌杀死乳油。 使用剂量一般为1:2 000~3 000倍。

190
氯氰菊酯(cypermethrin,灭百可,兴棉宝,)
理化特性 水溶性差,可溶于丙酮,氯仿,环己酮和二甲苯等有机溶剂。对光和热稳定,在酸性介质中比在碱性介质中稳定。

191
生物活性 氯氰菊酯及甲体氯氰菊酯均为高效,广谱具触杀和胃毒作用的杀虫剂。甲体氯氰菊酯是从氯氰菊酯八个异构体中拆分出的1R-顺式酸-S-醇酯/1S-顺式酸-R-醇酯(1:1)和1R-反式酸-S-醇酯/1S-反式酸-R-醇酯(1:1)的混合物,其药效比氯氰菊酯高约1倍。 大鼠急性经口LD50值为251~4 123mg/kg

192
制剂: 使用技术 10%氯氰菊酯(兴棉宝,灭百可,安绿宝)乳油,4.5%高效氯氰菊酯乳油等。
主要用于森林,果树,棉花和蔬菜,小麦,大豆等作物上防治鳞翅目,鞘翅目和双翅目害虫,对植食性半翅目害虫也有很好的防效,对土壤害虫有较好的持久活性。在害虫发生期采取喷雾施用,一般用制剂的1:2 000~1:3 000倍稀释液。

193
Repetitive discharge in the EMG(肌电图)
Repetitive discharge in the EMG(肌电图). Electromyogram traces from a rat before (upper record) and 70 minutes after an LD50 dose of the type II pyrethroid deltamethrin (lower record) showing abnormal repetitive discharges (*). The x-axis markers are in milliseconds, and the y-axis in millivolts.
. Electromyogram traces from a rat before (upper record) and 70 minutes after an LD50 dose of the type II pyrethroid deltamethrin (lower record) showing abnormal repetitive discharges (*). The x-axis markers are in milliseconds, and the y-axis in millivolts.")
194
Type I poisoning syndrome: allethrin, bifenthrin, bioallethrin, cismethrin, trans-fluorocyphenothrin, phenothrin, resmethrin, tetramethin, and pyrethrins. Type IIpoisoning syndrome: cyfluthrin, cyhalothrin, cypermethrin, deltamethrin, fenvalerate, and cis-fluorocyphenothrin. Mixed I/II poisoning syndrome: cyphenothrin,fenpropathrin, and flucythrinate.

195
N.B.A meta-analysis of observations made during regulatory studies by Soderlund et al. (2002) found bioallethrin, bifenthrin, permethrin, and pyrethrum to fit into the mixed category. This may have been either to misclassification by others (this table is based on Ray, 1991, plus Holton et al., 1997) or the imprecise recording of observations in the regulatory studies.
 found bioallethrin, bifenthrin, permethrin, and pyrethrum to fit into the mixed category. This may have been either to misclassification by others (this table is based on Ray, 1991, plus Holton et al., 1997) or the imprecise recording of observations in the regulatory studies..")
197
第六节 沙蚕毒素类杀虫剂 这是另一类以天然产物为模型开发成功的现代合成杀虫剂,但由于其先导化合物结构的局限性,目前开发的品种远不如有机磷类,氨基甲酸酯类及拟除虫菊酯类杀虫剂的品种多。

198
一、先导化合物 1934年日本的S.Nitta博士从生活在海滩泥地中的环节动物异足索沙蚕体内分离出一种杀虫活性物质,1962年日本学者鉴定了分子结构并全合成证实。 1964年发现沙蚕毒素对水稻螟虫有特殊的活性,同时合成开发出第一个沙蚕毒素类杀虫剂杀螟丹。至目前为止,生产上应用的仅5~6品种。

199
二、 类型 1.链状结构

200
2.环状结构

201
三、沙蚕毒素类杀虫剂的特征 沙蚕毒素类杀虫剂对害虫具有触杀、胃毒及内吸作用,杀虫谱广。
对人畜毒性低,商品化杀虫剂一般都属于低毒或中等毒性。对鱼类等水生动物毒性低,但对蜜蜂、家蚕毒性大。 作用机制独特,未发现交互抗性现象。 在动植物体内及环境中容易降解,对环境比较安全 。 有些品种对某些作物比较敏感,容易造成药害。如白菜、甘蓝等十字花科作物对杀螟丹、杀虫双敏感;豆类、棉花对杀虫环特别敏感,尤其在夏季高温季节或作物幼苗期更易造成药害。

202
四、主要品种 杀螟丹 理化特性 :水溶性很好,难溶于除醇类外的有机溶剂,在碱性条件下不稳定。
四、主要品种 杀螟丹 理化特性 :水溶性很好,难溶于除醇类外的有机溶剂,在碱性条件下不稳定。 生物活性:具有内吸、胃毒及触杀作用,有较长的持效期,对螟虫及一些鳞翅目害虫高效。对大鼠口服急性毒性LD50为250mg/kg。

203
制剂:98%巴丹可溶性粉剂,50%巴丹可溶性粉剂。
使用技术: 用于防治水稻、蔬菜及果树害虫。杀螟丹一般采取对水喷雾使用。 防治早、晚稻白穗,应在螟卵盛卵前1~2d水稻破口期施药或制成毒土撒施。对果树和蔬菜害虫,则应在害虫幼龄期喷洒防治为宜。

204
杀虫双和杀虫单 杀虫双于70年代由贵州化工研究所在研究杀螟丹合成时发现,并开发成防治水稻害虫特别是水稻螟虫的药剂。生产工艺比杀螟丹简单。防治对象与作用机理同杀螟丹。杀虫单是其一钠盐。

205
理化特性:具有很好的水溶性。在水溶液中,双、单钠盐在硫代硫酸钠的作用和空气的氧化下都转变为沙蚕毒素。
生物活性:杀虫双和杀虫单具有胃毒、触杀和内吸作用。对小鼠(雄)LD50为316mg/kg,三致试验未见异常。 制剂:18%杀虫双水剂、50%杀虫单可溶性粉剂、80%杀虫单可溶性粉剂、5%杀虫双颗粒剂等。 使用技术:对水稻螟虫、稻纵卷叶螟有特效,对许多果树及蔬菜鳞翅目害虫均有较好的防效。可采取喷雾、毒土及根区施药等方法。采取颗粒剂根区施药法,可延长持效期。
LD50为316mg/kg,三致试验未见异常。 制剂:18%杀虫双水剂、50%杀虫单可溶性粉剂、80%杀虫单可溶性粉剂、5%杀虫双颗粒剂等。 使用技术:对水稻螟虫、稻纵卷叶螟有特效,对许多果树及蔬菜鳞翅目害虫均有较好的防效。可采取喷雾、毒土及根区施药等方法。采取颗粒剂根区施药法,可延长持效期。")
206
第七节 昆虫几丁质合成抑制剂 70年代初,Van Daalen等在筛选新的除草剂时,将敌草隆去掉两个甲基,用苯甲酰基取代苯腈,合成了Du-19111

207
一、类型 以下式为先导化合物,开发出3个类型的昆虫几丁质合成抑制剂

208
1.苯甲酰脲类 ①在苯甲酰基上修饰,引入F:

209
②在芳胺环上修饰,引入CF3,OCF3, OCF2CF2等:

210
③在芳胺环对位引入取代芳(杂)氧基
氧基")
211
2.杂环类 杂环类昆虫几丁质合成抑制剂起源于对杀菌剂稻瘟灵的研究,发现稻瘟灵能影响稻飞虱若虫的蜕皮。这一现象给人以启迪:希望改变其结构,筛选出新型杀虫剂。

213
二、特征 昆虫几丁质合成抑制剂是一类高效杀虫剂,其毒力高于有机磷和氨基甲酸酯类杀虫剂。
选择性很强。对鳞翅目幼虫高效,对鞘翅目、双翅目害虫也有效。绝大多数品种对刺激式口器害虫防效甚差,但氟虫脲在常用剂量下却有良好的杀螨活性,噻嗪酮则对飞虱、叶蝉等有高活性。 对哺乳动物低毒安全,对鱼类、对害虫天敌、对蜜蜂均很安全,无残毒和环境污染之虑,可称作“生物合理杀虫剂”。 1. 。 2.这类杀虫剂3.这类杀虫剂一般以胃毒作用为主,触杀作用差,不内吸,但杂环类的噻嗪酮却是个例外,不但有强大的触杀作用,而且还有一定的熏蒸作用。 4.这类杀虫剂作用较慢,不能迅速地控制害虫危害,对鳞翅目害虫的致死时间需2~7 d,这是其主要缺点。另一缺点是对幼虫高效,但对成虫作用甚微。

214
一般以胃毒作用为主,触杀作用差,不内吸,但杂环类的噻嗪酮却是个例外,不但有强大的触杀作用,而且还有一定的熏蒸作用。
作用较慢,不能迅速地控制害虫危害,对鳞翅目害虫的致死时间需2~7 d,这是其主要缺点。另一缺点是对幼虫高效,但对成虫作用甚微。

215
三、 近年主要进展 1.苯甲酰脲类昆虫几丁质合成抑制剂的触杀作用一般都很差,这对防治不利。因此,近年来围绕如何提高触杀效果进行了许多探索。
当X=Cl时,其触杀毒力比伏虫隆提高11倍,比灭幼脲提高33倍;当X=F时,触杀毒力比伏虫隆提高4倍,比灭幼脲提高12倍。

216
2.目前商品化的这类杀虫剂都不具备内吸或内渗杀虫活性,因此近年来围绕研制内吸几丁质合成抑制剂进行了大量合成筛选。例如在定虫隆分子上引入强极性基团,
该化合物有一定水溶性,用于蔬菜灌根,上面叶片显示对小菜蛾、斜纹夜蛾的良好杀虫效果,说明它具有内吸杀虫活性,但用于田间防治蚜虫却未成功,因而至今未能商品化。

217
四、主要品种 灭幼脲(灭幼脲Ⅲ号,苏脲Ⅰ号,chlorbenzuron)
")
218
理化特性 不溶于水,易溶于二甲基甲酰胺和吡啶等有机溶剂。 遇碱和较强的酸易分解,常温下储存稳定。
25%胶悬剂的密度为1.05~1.15g/mL,白色乳状悬浮液。

219
生物活性 对鳞翅目幼虫有特效,可用于防治小麦、水稻、高梁、玉米、大豆上的粘虫、道纵卷叶螟、豆天蛾、甜菜和白菜上的甘蓝夜蛾、菜青虫,森林和果树上的松毛虫、舞毒蛾、美国白蛾、枣步曲等害虫。 可兼制某些卫生害虫,如蚊、蝇类幼虫。 持效期30d以上。 灭幼脲对大鼠急性经口LD50 >20 000mg/kg,对兔眼黏膜和皮肤无明显刺激作用。

220
制剂 使用方法 25%、50%灭幼脲悬浮剂。 防治粘虫、天幕毛虫、舞毒蛾、螟虫、甘蓝夜蛾、茶尺蠖的用量为有效成分120~150 g/hm2;
防治松毛虫的用量为有效成分150~300 g/hm2;防治菜青虫、小菜蛾只需有效成分30~75 g/hm2。

221
噻嗪酮(扑虱灵、优乐得,buprofezin)
对稻飞虱、叶蝉、温室粉虱有很强的触杀作用,也有一定熏蒸作用,有效剂量为100~200 g/ha,持效期达35~40 d, 对稻田害虫主要天敌及鱼类安全。

222
第七节 杂环类杀虫剂 杂环类杀虫剂总的特点是: 化学结构新颖,作用机制也新颖,不易和现有杀虫剂产生交互抗性;
第七节 杂环类杀虫剂 杂环类杀虫剂总的特点是: 化学结构新颖,作用机制也新颖,不易和现有杀虫剂产生交互抗性; 对害虫高效,对抗有机磷、氨基甲酸酯、拟除虫菊酯类杀虫剂的害虫种群也有很满意的防治效果; 对哺乳动物低毒或中等毒性,使用安全,对害虫天敌也安全,有利于综合治理。

223
一、含吡啶基团的杀虫剂 新烟碱类化合物的开发起源于1978年Soloway等发表的一类硝基亚甲基化合物nithiazin的研究。将nithiazin的氢化噻嗪环变成咪唑环,并在杂环上导入氯吡啶甲基,使其生物活性提高两个数量级,其中以吡虫啉(imidaclopid)为突出代表。
为突出代表。")
224
化合物 LC90 40 1.6 200 0.32 <0.32 ~0.32 8 新烟碱类化合物的杀虫活性(黑尾叶蝉 )(mg/L)
(mg/L)")
225
吡虫啉 理化特性 纯品为无色结晶,在水中的溶解度(20℃)为有效成分0.51g/L。
为有效成分0.51g/L。")
226
制 剂 5%吡虫啉可溶性粉剂 5%吡虫啉可湿性粉剂 10% 吡虫啉可湿性粉剂 5%吡虫啉乳油等。

227
生物活性及使用 对叶蝉、飞虱、粉虱、蚜虫及多种鞘翅目害虫有很高的防治效果,而且对哺乳动物及非靶标生物安全;
不仅有强大的触杀活性,而且有良好的内吸活性,加之在土壤中半衰期长达150 d,因而除可作叶面喷洒外,还可土壤施药。 吡虫啉和现有的杀虫剂无交互抗性,而且对已产生抗有机磷或拟除虫菊酯的害虫种群同样高效。

228
乙虫眯(acetamiprid) 20%莫比朗可溶性粉剂,3%莫比朗乳油。
对半翅目(尤其是蚜虫),缨翅目和鳞翅目害虫,包括抗性种群,都有很高的防治效果,既可叶面喷洒,也可作土壤处理。
,缨翅目和鳞翅目害虫,包括抗性种群,都有很高的防治效果,既可叶面喷洒,也可作土壤处理。")
229
二、三唑类杀虫剂 三唑类杀虫剂是一类很有特色的新杀虫剂,最有代表意义的是Rhom &hass 开发的唑蚜威(triaguron):
用于防治对有机磷及拟除虫菊酯类杀虫剂产生抗性的种群有很高的防治效果。有很强的内吸作用,并可双向(向顶、向基)传导,这对于防治某些蔬菜、果树的根部蚜虫十分有利。 60~90 g/ha,可维持药效10~15d。 对人畜毒性较低,对蚜虫天敌、对蜜蜂等安全。
传导,这对于防治某些蔬菜、果树的根部蚜虫十分有利。 60~90 g/ha,可维持药效10~15d。 对人畜毒性较低,对蚜虫天敌、对蜜蜂等安全。 ")
230
三、吡唑类杀虫剂 1989年法国罗纳-普朗克公司开发成功三氟甲基亚磺酰基吡唑类杀虫剂锐劲特(fipronil),1996年登记。
广谱内吸杀虫剂,对半翅目、鳞翅目、缨翅目、鞘翅目害虫高效,而且对抗有机磷、拟除虫菊酯类杀虫剂的害虫同样高效。 在水稻、棉花、玉米、甜菜等作物上,有效剂量仅50 g/ha。

231
四、吡咯类杀虫剂 将二恶吡咯霉素作为先导化合物,经改造筛选,开发出杀虫杀螨剂溴虫氰(虫螨腈、除尽)。
溴虫氰的药效和氯氰菊酯相似。田间试验结果说明,用以防治豆卫矛蚜、棉铃虫、马铃薯叶甲、潜叶蝇、甜菜夜蛾、棉红蜘蛛等,用量为125 g/ha,效果优于常规杀虫剂。

232
五、吡啶类杀虫剂 吡蚜酮作为杀虫剂出现,不但代表了一类全新的化合物,而且在作用方式上也是独树一帜的。吡蚜酮不具有“击倒”效果,对昆虫也没有直接毒性,但昆虫一接触到该化合物,立刻停止取食,而且“停食”不是由于“拒食作用”所引起的,是不可逆的。此作用方式被称为“口针穿透阻塞”(blockage of stylet penetration)。 吡蚜酮可用于防治大部分同翅目害虫,尤其是蚜科、粉虱科、叶蝉科及飞虱科害虫,适用于蔬菜、水稻、棉花、果树及多种大田作物。 吡蚜酮pymetrozine
。 吡蚜酮可用于防治大部分同翅目害虫,尤其是蚜科、粉虱科、叶蝉科及飞虱科害虫,适用于蔬菜、水稻、棉花、果树及多种大田作物。 吡蚜酮pymetrozine.")
233
第八节 其他常见杀虫剂 一、有机氯杀虫剂 1.大多数品种合成原料易得,合成工艺简单,成本低廉;
第八节 其他常见杀虫剂 一、有机氯杀虫剂 1.大多数品种合成原料易得,合成工艺简单,成本低廉; 2.杀虫谱广,残效期长,特别适合防治棉铃虫,蝗虫及地下害虫等; 3.性质稳定,不易降解,滞留时间长,易造成环境污染; 4.水溶性很差,脂溶性强,可通过食物链在动物体内富集,对人类健康带来隐患; 5. 害虫害螨容易对有机氯杀虫剂产生抗药性。

235
二、杀虫抗生素及其类似物 阿维菌素(avermectin)
")
236
三、邻苯二甲酰胺类和邻甲酰氨基苯甲酰胺类杀虫剂
邻苯二甲酰胺类和邻甲酰氨基苯甲酰胺类杀虫剂是一类作用机制独特的新颖杀虫剂,此类杀虫剂能强烈激活鱼尼丁受体(ryanodine receptor),干扰肌肉收缩活动, 鱼尼丁 ryanodine 氟虫酰胺(1st)(flubendiamide) 氯虫酰胺(chlorantraniliprole )
,干扰肌肉收缩活动, 鱼尼丁. ryanodine. 氟虫酰胺(1st)(flubendiamide) 氯虫酰胺(chlorantraniliprole )")
237
桔霉素( milbemycin) 桔霉素是一系列(6个)十六元大环内酯,其产生菌是吸水链霉菌金色亚种Streptomyces hygroscopicus var aureolacrimosus,主要用于螨类防治。 R= OH-,(OH)2P-
2P-")
238
多杀霉素( spinosad) 商品为2.5%菜喜悬浮剂 浏阳霉素( polynactins)
 商品为2.5%菜喜悬浮剂 浏阳霉素( polynactins)")
239
第九节 杀螨剂 杀螨剂(acaricide)是指用于防治蛛形纲中有害螨类的农药。
第九节 杀螨剂 杀螨剂(acaricide)是指用于防治蛛形纲中有害螨类的农药。 防治植食性螨的农用杀螨剂通常是指只杀螨不杀昆虫或以杀螨为主的药剂。 许多杀虫剂兼有杀螨活性,但不能称为杀螨剂
是指用于防治蛛形纲中有害螨类的农药。 防治植食性螨的农用杀螨剂通常是指只杀螨不杀昆虫或以杀螨为主的药剂。 许多杀虫剂兼有杀螨活性,但不能称为杀螨剂.")
240
第九节 杀螨剂 理想的杀螨剂 杀螨能力强,可杀成、若螨及卵; 较长的持效期; 化学性质相对稳定,可与其他农药混用;
第九节 杀螨剂 理想的杀螨剂 杀螨能力强,可杀成、若螨及卵; 较长的持效期; 化学性质相对稳定,可与其他农药混用; 对人畜、植物安全,对天敌、有益昆虫无毒杀作用。 早期:硫磺和无机硫制剂 二战后:有机氯 目前:高活性有机合成杀虫剂

241
一、以杀螨卵为主的杀螨剂 早期开发的硝基酚类杀螨剂,主要品种有消螨酚(dinex)、乐杀螨(binapacryl)和消螨通 (dinobuton)以及我国开发出的杀螨剂F1050等。
、乐杀螨(binapacryl)和消螨通 (dinobuton)以及我国开发出的杀螨剂F1050等。")
242
二、具有生长调节活性的杀螨剂 具有生长调节活性的杀螨剂,主要表现为对幼螨或者若螨防治效果较好,而对于成螨效果较差甚至无效。这类杀螨剂的结构一般都具有杂环结构。

243
应用技术 1、对于具有生长调节剂作用的杀螨剂,要特别注意对卵和幼螨或者若螨的控制。这类药剂对成螨无效。因此,在螨类控制上,要注意和杀成螨的药剂结合使用。 2.具有该类作用机制的不同种类的药剂,在具体作用对象的控制效果上有一定的差异,需要在具体使用过程中给予注意。 2、由于该类药剂对成螨无效,因此,在使用时要注意在卵孵化盛期和幼(若)螨期施药。喷雾时要注意均匀周到。
螨期施药。喷雾时要注意均匀周到。")
244
三、具有神经毒剂作用的杀螨剂 主要包括以DDT为先导化合物开发的一类杀螨剂、以含硫的有机氯化合物开发的一类杀螨剂、亚硫酸酯类杀螨剂以及一部分杂环类杀螨剂。 杀螨醇 早已禁止在蔬菜和茶叶上使用,并根据《斯德哥尔摩公约》(POPs公约)的要求,三氯杀螨醇将被替代。 替代方案已经基本形成。
的要求,三氯杀螨醇将被替代。 替代方案已经基本形成。 .")
246
应用技术 有机氯杀螨剂大都采用喷雾的方法施用。
乙酯杀螨醇,也可加工成烟雾剂用于防治果树、观赏植物和棉花等作物上的多种螨类的所有发育阶段的螨。 丙酯杀螨醇为非内吸性触杀杀螨剂,用于防治果树、茶叶、棉花、甜菜、蔬菜和观赏植物的多种害螨,应尽量均匀喷雾。 三氯杀螨醇除主要用于喷雾外,有时可以喷粉使用,主要用于棉花、果树、花卉等害螨的控制,对苹果某些品种易产生药害,不宜使用。

247
杀螨硫醚、氯杀螨、杀螨酯均可用于防治各种螨态的螨,具有杀卵活性,但无内吸作用。使用时,一定要考虑均匀喷雾,而且争取早期施药。
杀螨特适用于果树、棉花、黄瓜、茄子等害螨的控制,一般不可与其他药剂混合使用; 克螨特可用于防治果树、蔬菜、棉花、花卉等多种植物的害螨。在高温、高湿条件下喷洒高浓度的克螨特对某些作物的幼苗和新梢嫩叶有药害,对蔬菜及棉苗,使用倍数应不能低于3000倍;在柑橘接近成熟期时,尽量减少使用;不可与碱性农药混合使用。 联苯肼酯由于与其它药剂无交互抗性,故被用于果蔬、棉花、玉米和观赏作物防治各种螨类,特别是二斑叶螨和全爪螨,主要在螨的活动期有效。用药后3d内对靶标害螨有击倒作用,并能持续有效30d。

248
四、抑制呼吸代谢作用的杀螨剂 具有呼吸代谢抑制作用的杀螨剂大致可分为三大类。 一是早期开发的具有杀卵作用的二硝基苯类
二是20世纪70年代开发成功的有机锡类杀螨剂 三是在20世纪90年代,相继开发的一批具有良好杀螨活性的杂环类杀螨剂。

249
1、有机锡类杀螨剂 由于锡的残留作用,国外目前仅有少量生产和应用,我国曾对以上品种进行过研究和开发,国内目前仅苯丁锡还有少量生产。

250
2、具有呼吸代谢抑制作用的杂环类杀螨剂 嘧啶类 吡唑类 哒嗪酮类 萘醌类 喹唑啉类

251
使用技术 1、主要是对成螨和若螨的活性比较好,大多数品种对卵的杀伤作用比较差。该类药剂的内吸性不强,因此,喷雾时要注意均匀、周到。
2、该类药剂在施药后初期药效作用慢,故应在害螨盛发期前虫口密度较低时施用;主要用于防治柑橘、苹果、葡萄、茶及观赏植物上的瘿螨科和叶螨科类害螨,尤其对全爪螨属和叶螨属的害螨高效。 3、一般不可与波尔多液、石硫合剂等碱性农药混用。施药时要注意安全防护。

Similar presentations




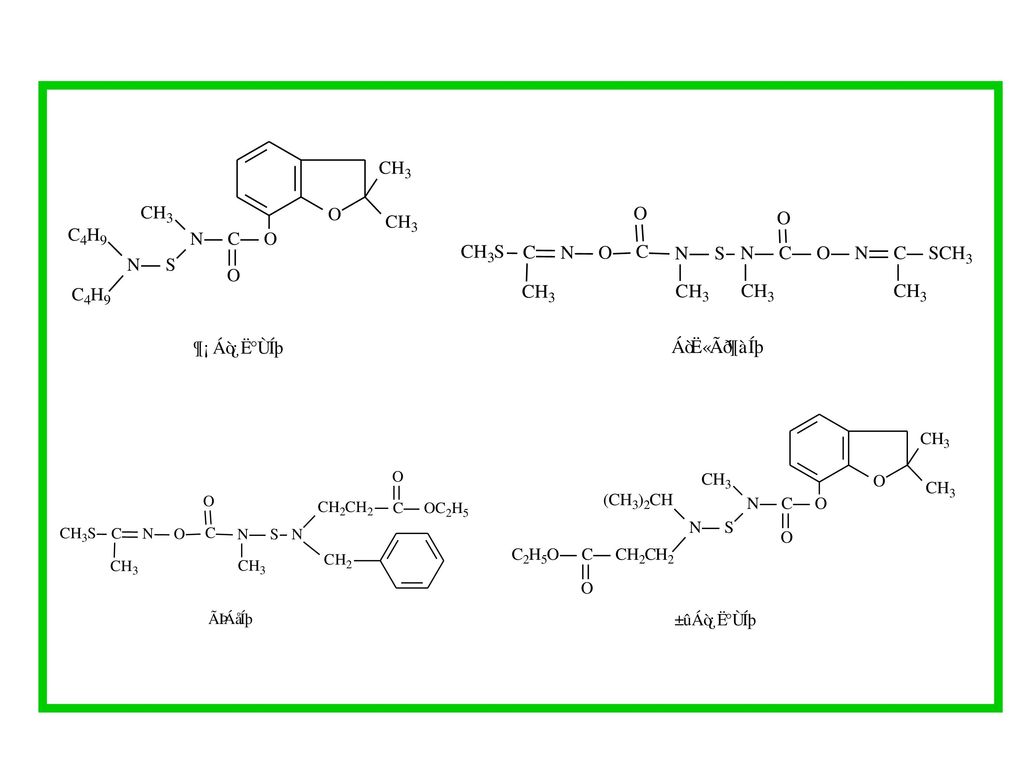




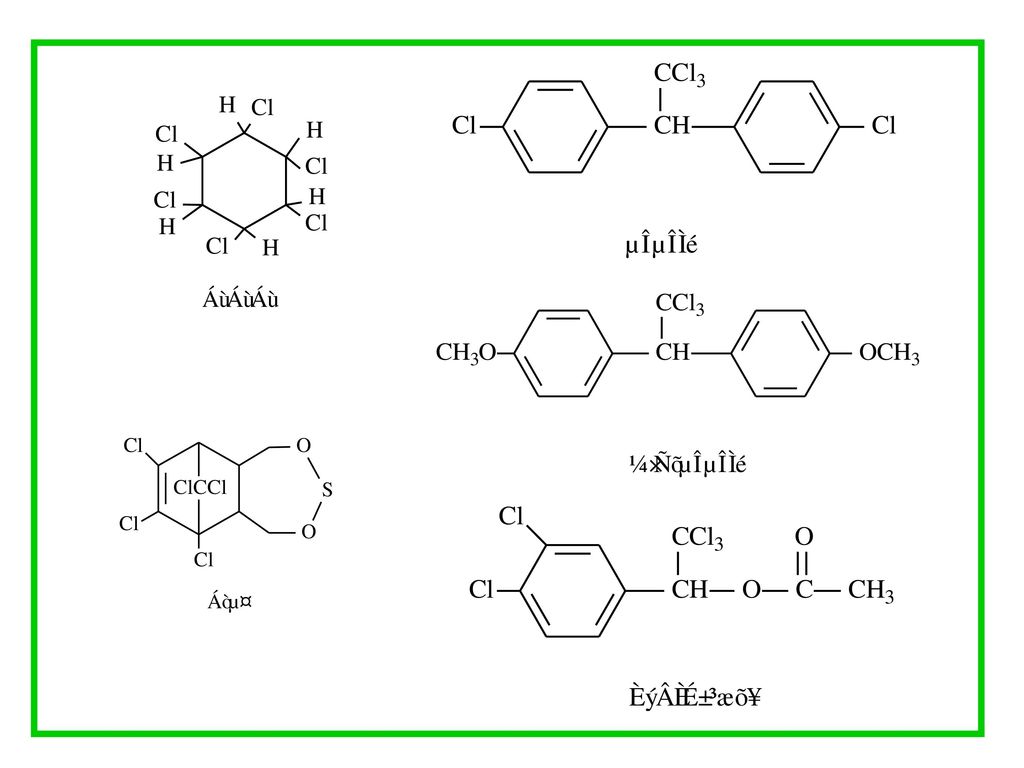

的作用消失方式.>")













分泌的化学物质进行调节,这就是激素调节。>")




