Download presentation
1
第十三章 DNA的复制和修复 生物体的遗传信息储存在DNA中,并通过DNA的复制由亲代传给子代。
在子代的生长发育中遗传信息自DNA转录给RNA,然后翻译成蛋白质以执行各种生命功能,使后代表现出与亲代相似的遗传性状。 基因性状:one gene-one enzyme hypothesis.

2
1958年,F.Crick提出中心法则: (1)以原DNA分子为模板,合成出相同DNA分子的过程。
(2)以某一段DNA分子为模板,合成出与其序列对应的RNA分子的过程。 (3)以mRNA为模板,根据三联密码规则,合成对应蛋白质的过程。 中心法则揭示了生物体内遗传信息的传递方向。
以某一段DNA分子为模板,合成出与其序列对应的RNA分子的过程。 (3)以mRNA为模板,根据三联密码规则,合成对应蛋白质的过程。 中心法则揭示了生物体内遗传信息的传递方向。")
3
DNA生物合成有两种方式:DNA复制和反转录

4
P321 图191 Watson和Crick提出的DNA双螺旋复制模型
1953年,Watson和Crick在提出DNA双螺旋结构模型时就推测DNA可能按照半保留机制进行自我复制。 P321 图191 Watson和Crick提出的DNA双螺旋复制模型 在复制过程中,首先亲代双链解开,然后每条链作为模板,在其上合成互补的子代链,结果新形成的两个子代DNA与亲代DNA分子的碱基顺序完全一样,而且每个子代DNA分子中有一条链完全来自亲代DNA,另一条是新合成的。 1958年,Meselson和Stahl用15N标记E.coli. DNA,用密度梯度离心试验证明了DNA的复制是半保留复制。 P322 图 DNA的半保留复制。

6
1963年,Cairns用放射自显影法,在显微镜下首次观察到完整的正在复制的E. coli. 染色体DNA。
P323 图 19-3 3H-脱氧胸苷标记E.coli. DNA ,经过将近两代时间,用溶菌酶消化细胞壁,将E.coli. DNA转至膜上,干燥,压感光胶片,3H放出β粒子,还原银,在光学显微镜下观察。用这种方法证明了大肠杆菌染色体DNA是一个环状分子,并以半保留的形式进行复制。

7
DNA的半保留复制可以说明DNA在代谢上的稳定性。经过多代复制,DNA的多核苷酸链仍可以保持完整,并存在于后代而不被分解掉。

8
二、 复制起点、单位和方向 1、 复制起点 DNA复制的调控是在起始阶段进行的,一旦复制起始,它就会继续下去直到整个复制子完成复制。
二、 复制起点、单位和方向 DNA复制的调控是在起始阶段进行的,一旦复制起始,它就会继续下去直到整个复制子完成复制。 1、 复制起点 复制起点是以一条链为模板起始DNA合成的一段序列。有时,两条链的复制起点并不在同一点上(不对称复制,如D环复制)。 在一个完整的细胞周期中,每一个复制起点只使用一次,完成一次复制过程。 多数生物的复制起点,都是DNA呼吸作用强烈(甲醛变性实验)的区段,即经常开放的区段,富含A.T。
。 在一个完整的细胞周期中,每一个复制起点只使用一次,完成一次复制过程。 多数生物的复制起点,都是DNA呼吸作用强烈(甲醛变性实验)的区段,即经常开放的区段,富含A.T。")
9
★环状DNA复制起点的gene mapping P325 图19-6

10
★复制起点的克隆和功能分析——重组质粒转化法
大肠杆菌的复制起点oriC区1Kb的重组质粒在转化子中的复制行为与其染色体一样,受到严密控制,每个细胞只有1-2个拷贝,用核酸外切酶缩短oriC克隆片段的大小,最后得到245bp的基本功能区,携带它的质粒依然能够自我复制,拷贝数可以增加到20以上,这说明发动复制的序列在245bp的基本功能区,而决定拷贝数的序列在基本功能区之外和1Kb之间。 鼠伤寒沙门氏菌的起点位于一段296bp的DNA片段上,与大肠杆菌的复制起始区有86%同源性,而且有些亲缘关系较远的细菌,其复制起点在大肠杆菌中亦能起作用。因此,复制起始区的结构可能是很保守的。 起始序列含有一系列对称的反向重复和某些短的成簇的保守序列。

11
2、 复制单位 复制子(Replicon):Genome能独立进行复制的单位,每个复制子都含有一个复制起点。 原核生物的染色体和质粒、真核生物的细胞器DNA都是环状双链分子,它们都是单复制子,都在一个固定的起点开始复制,复制方向大多数是双向的,少数是单向复制。多数是对称复制(θ形复制),少数是不对称复制(一条链复制后才进行另一条链的复制,如D-环复制)。 真核生物的染色体DNA是线形双链分子,含有许多复制起点,因此是多复制子,每个复制子约有 Kbp。人体细胞平均每个染色体含有1000个复制子。 病毒DNA多种多样,环状或线形,双链或单链,但都是单复制子。
,少数是不对称复制(一条链复制后才进行另一条链的复制,如D-环复制)。 真核生物的染色体DNA是线形双链分子,含有许多复制起点,因此是多复制子,每个复制子约有 Kbp。人体细胞平均每个染色体含有1000个复制子。 病毒DNA多种多样,环状或线形,双链或单链,但都是单复制子。")
13
3、 复制方向 大多数是双向的(等速进行或异速进行),形成两个复制叉,少数是单向复制,形成一个复制叉。
,形成两个复制叉,少数是单向复制,形成一个复制叉。")
14
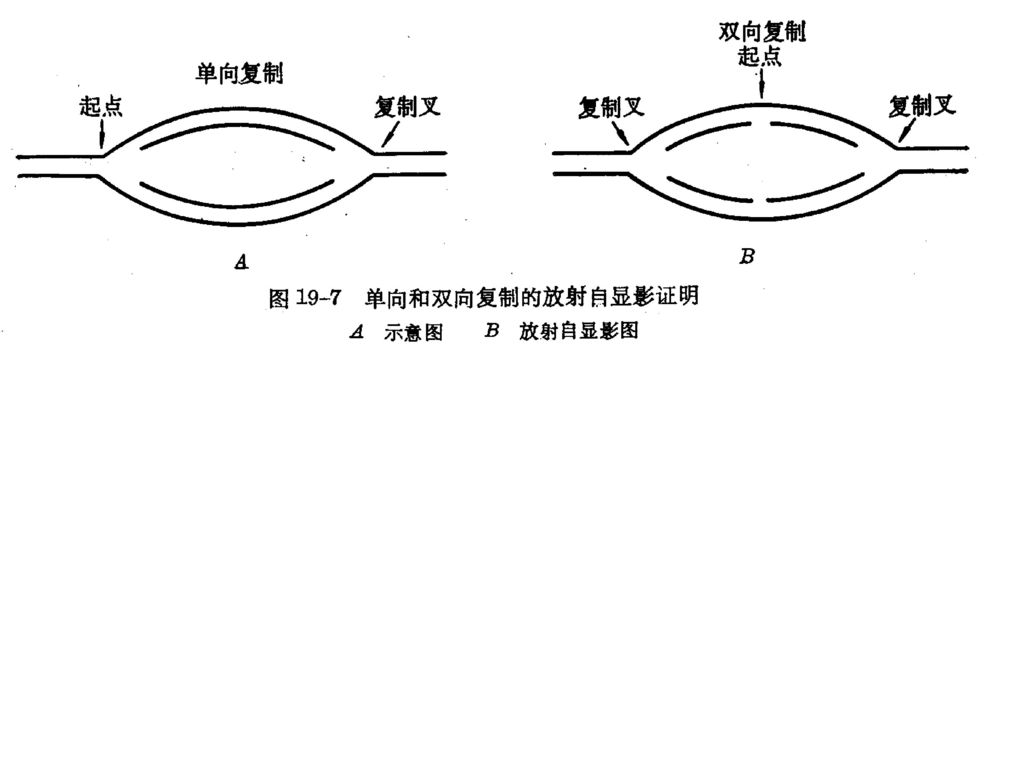
★用放射自显影实验判断DNA的复制方向及速度
P325 图 单向和双向复制的放射自显影证明 低放射性3H-脱氧胸苷 高放射性3H-脱氧胸苷 a. 单向 b. 双向等速 c. 双向异速 E.coli.的一个温度敏感株,在42℃时,能使DNA在完成复制后,不再开始新的复制过程,而在25℃时复制功又能能恢复。

16
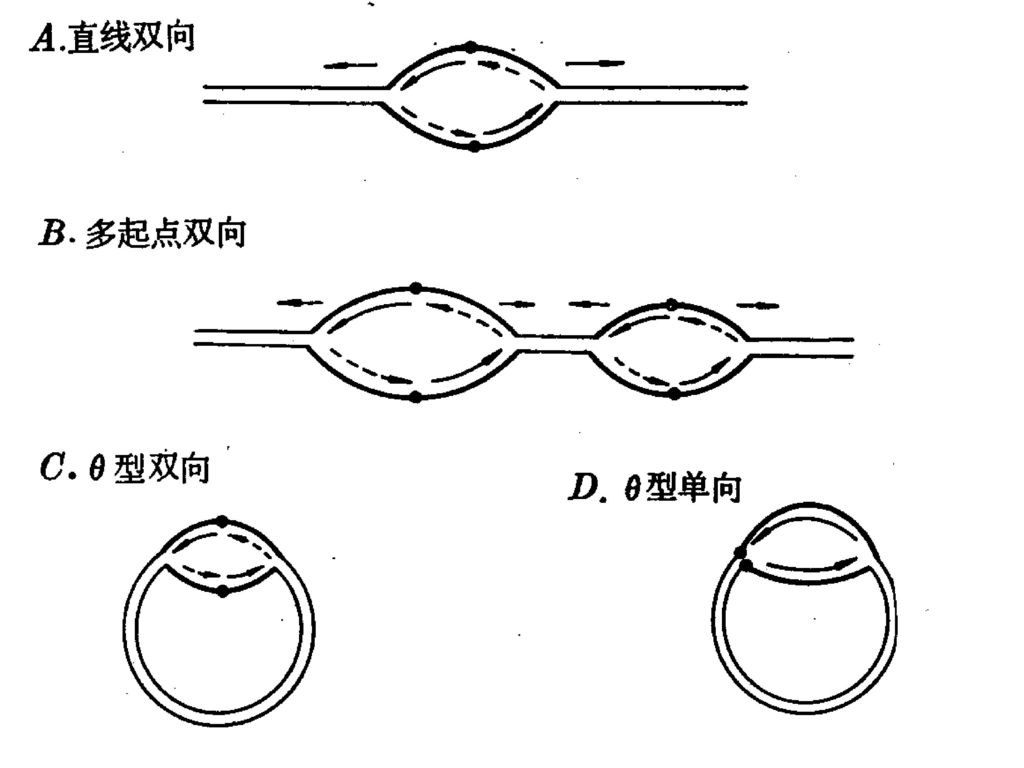
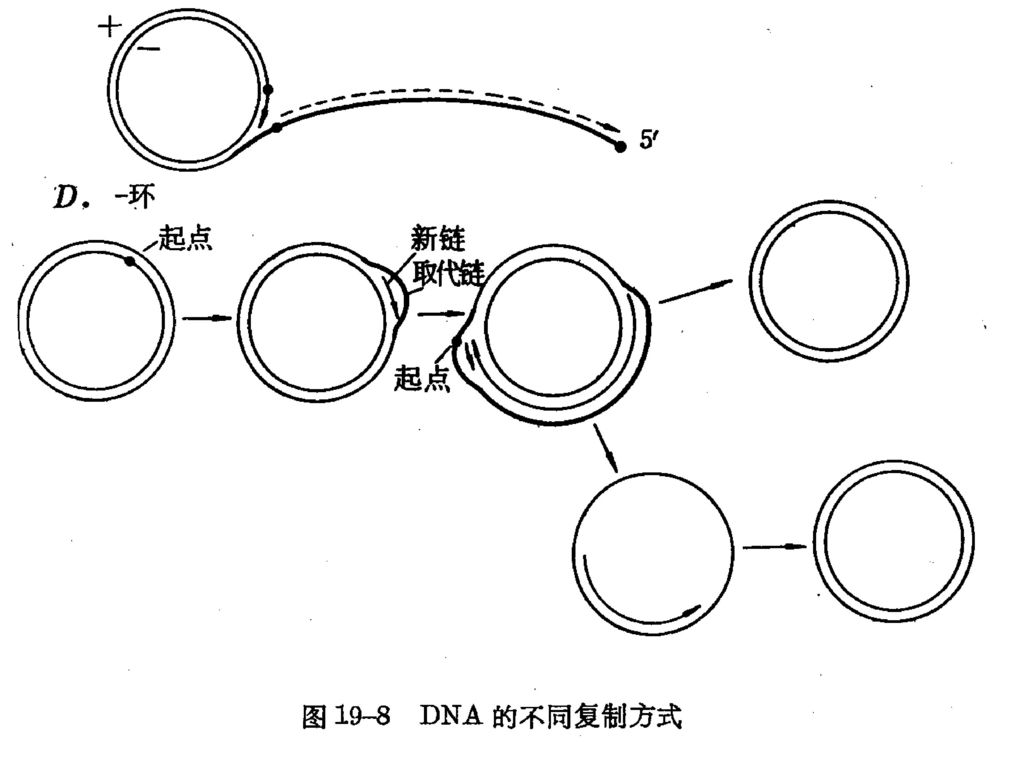
4、 DNA的几种复制方式 (1)、 直线双向复制 (2)、θ型复制: (3)、 滚环复制: (4)、 D环复制(取代环复制):
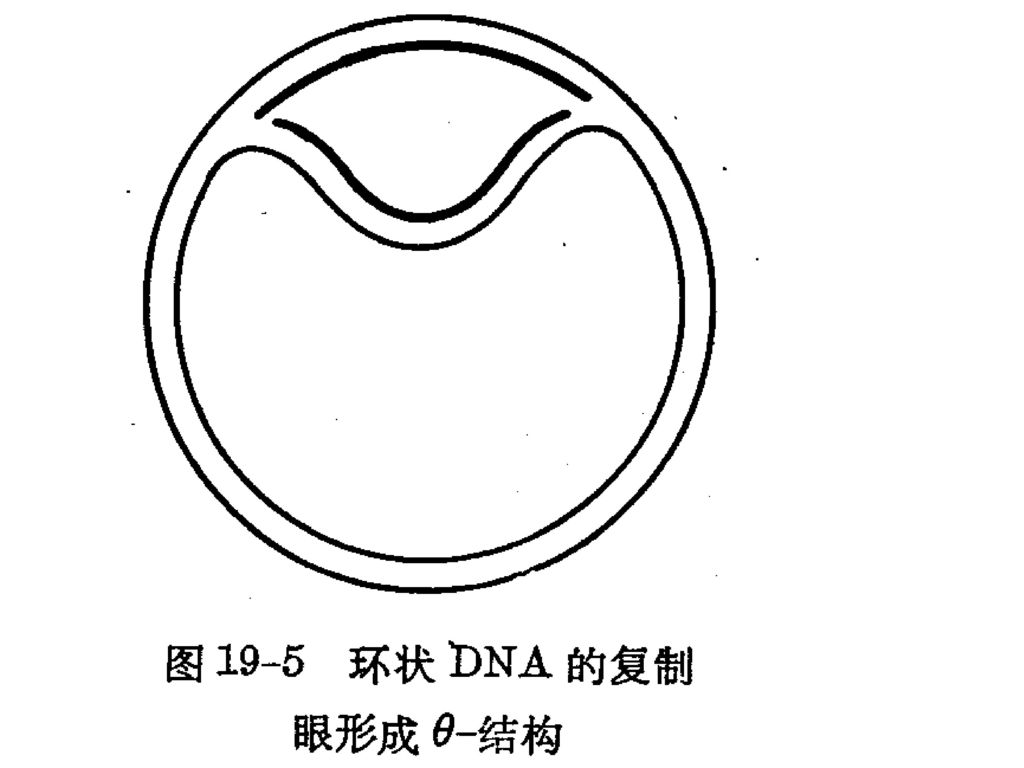
(1)、 直线双向复制 单点双向,T7 多点双向,真核染色体DNA (2)、θ型复制: 环状双链DNA,对称复制,单向或双向(E .coli.) (3)、 滚环复制: 环状单链DNA,Φx174 (4)、 D环复制(取代环复制): 不对称复制,线粒体、叶绿体DNA。 两条链的复制起点不在同一点上,一条链先复制,另一条链保持单链而被取代:当一条链复制到一定程度时才暴露出另一条链的复制起点,另一条链才开始复制。
、 直线双向复制. 单点双向,T7. 多点双向,真核染色体DNA. (2)、θ型复制: 环状双链DNA,对称复制,单向或双向(E .coli.) (3)、 滚环复制: 环状单链DNA,Φx174. (4)、 D环复制(取代环复制): 不对称复制,线粒体、叶绿体DNA。 两条链的复制起点不在同一点上,一条链先复制,另一条链保持单链而被取代:当一条链复制到一定程度时才暴露出另一条链的复制起点,另一条链才开始复制。")
17
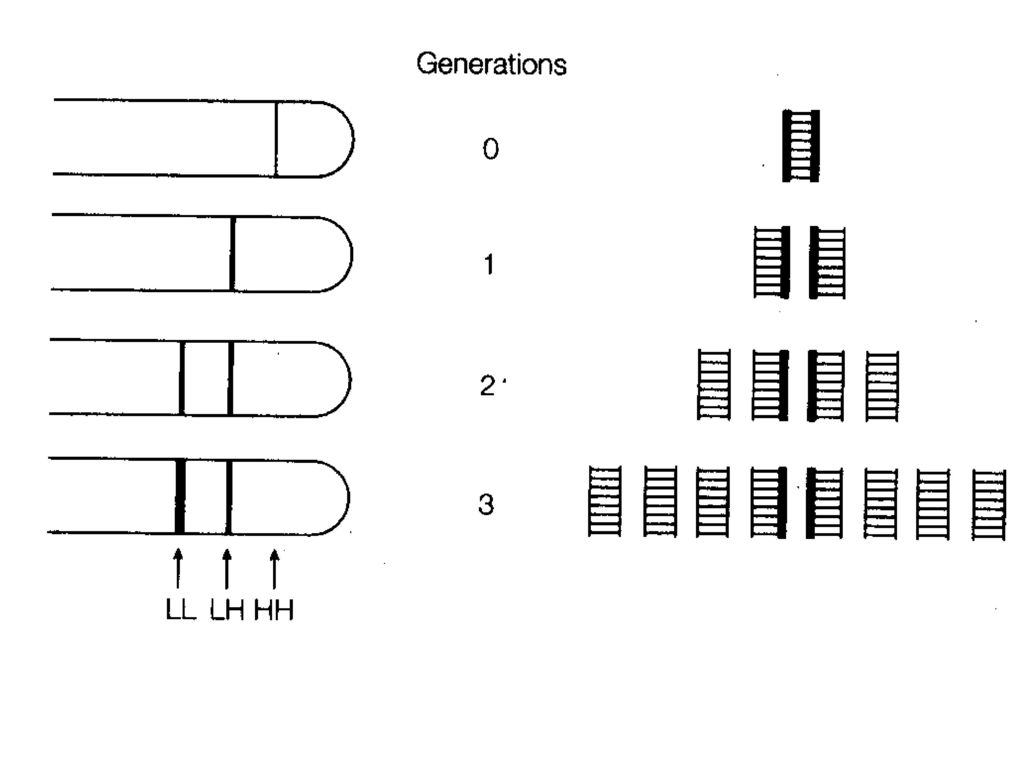
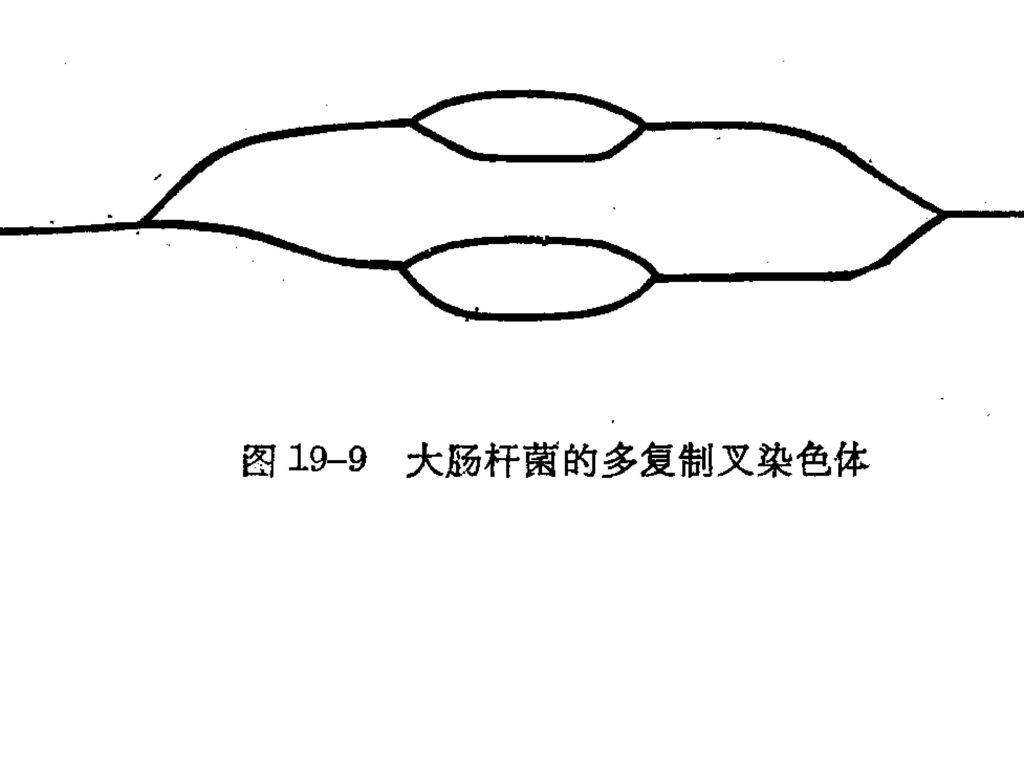
(5)、 多复制叉复制: 第一轮复制尚未完成,复制起点就开始第二轮的复制。
(5)、 多复制叉复制: 第一轮复制尚未完成,复制起点就开始第二轮的复制。 DNA复制时复制叉前进的速率比较恒定,DNA复制的速率实际上取决于起始频率。 E.coli复制叉移动的速率约50Kb/min,复制一代约需40分钟[4.2X106/(50KbX2)=42]。富营养时,可采取多复制叉复制方式,20min复制一代。 真核复制叉前进的速率约 bp/min,采用多复制子方式,真核染色体复制一代要6-8小时。
、 多复制叉复制: 第一轮复制尚未完成,复制起点就开始第二轮的复制。 DNA复制时复制叉前进的速率比较恒定,DNA复制的速率实际上取决于起始频率。 E.coli复制叉移动的速率约50Kb/min,复制一代约需40分钟[4.2X106/(50KbX2)=42]。富营养时,可采取多复制叉复制方式,20min复制一代。 真核复制叉前进的速率约 bp/min,采用多复制子方式,真核染色体复制一代要6-8小时。")
21
目前已发现30多种酶及蛋白质因子参与DNA复制 (一) DNA聚合酶和DNA的聚合反应 1、 DNA聚合反应必备的条件
⑵ DNA模板(反转录时用RNA模板) ⑶引物 (DNA、RNA或蛋白质) ⑷ 4种dNTP ⑸ Mg2+
 ⑶引物 (DNA、RNA或蛋白质) ⑷ 4种dNTP. ⑸ Mg2+")
22
2、 聚合反应过程及特点 总反应式: n1dATP DNA pol . dAMP
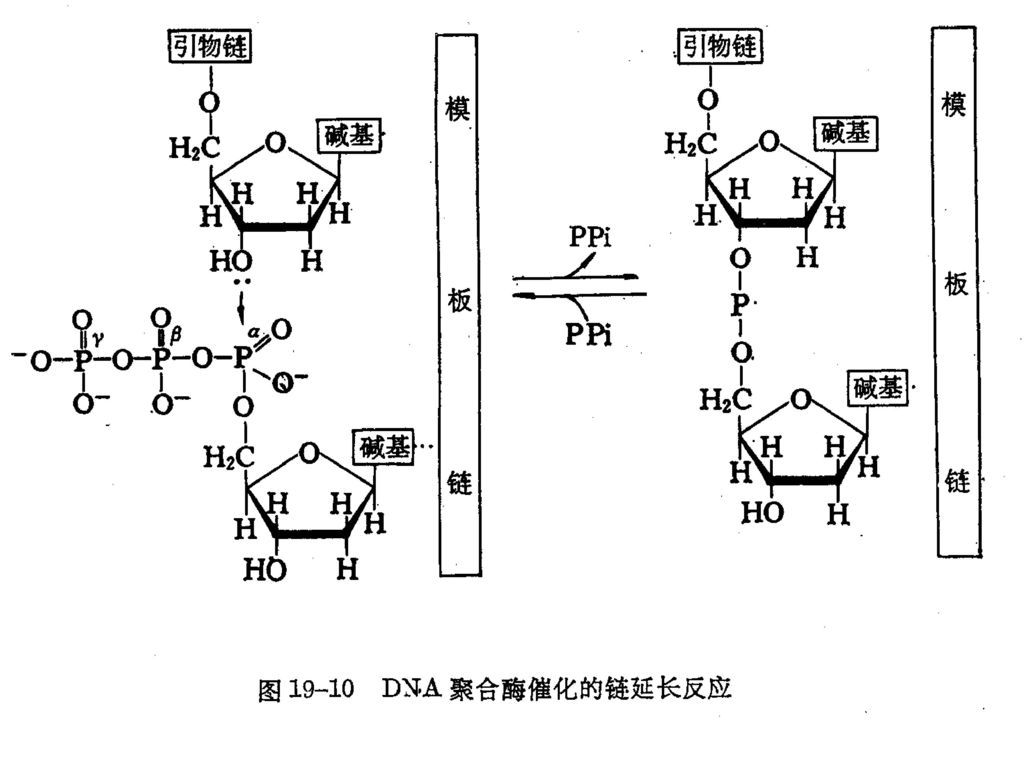
2、 聚合反应过程及特点 总反应式: n1dATP DNA pol dAMP n2dGTP +DNA dGMP DNA+(n1+n2+n3+n4)PPi n3dCTP Mg dCMP n4dTTP dTMP P329 图 P330图19-11 在链的延长过程中,链的游离3,-羟基,对进入的脱氧核糖核苷三磷酸α磷原子发生亲核攻击,生成3,.5,-磷酸二酯键,并脱下焦磷酸。
PPi. n3dCTP Mg2+ dCMP. n4dTTP dTMP. P329 图19-10 P330图 在链的延长过程中,链的游离3,-羟基,对进入的脱氧核糖核苷三磷酸α磷原子发生亲核攻击,生成3,.5,-磷酸二酯键,并脱下焦磷酸。")
24
DNA聚合酶的反应特点: ⑴ 以4种dNTP为底物 ⑵ 反应需要接受模板的指导,不能催化游离的dNTP的聚合。
⑶ 反应需有引物3,-羟基存在 ⑷ 链生长方向5, → 3, ⑸ 产物DNA的性质与模板相同

25
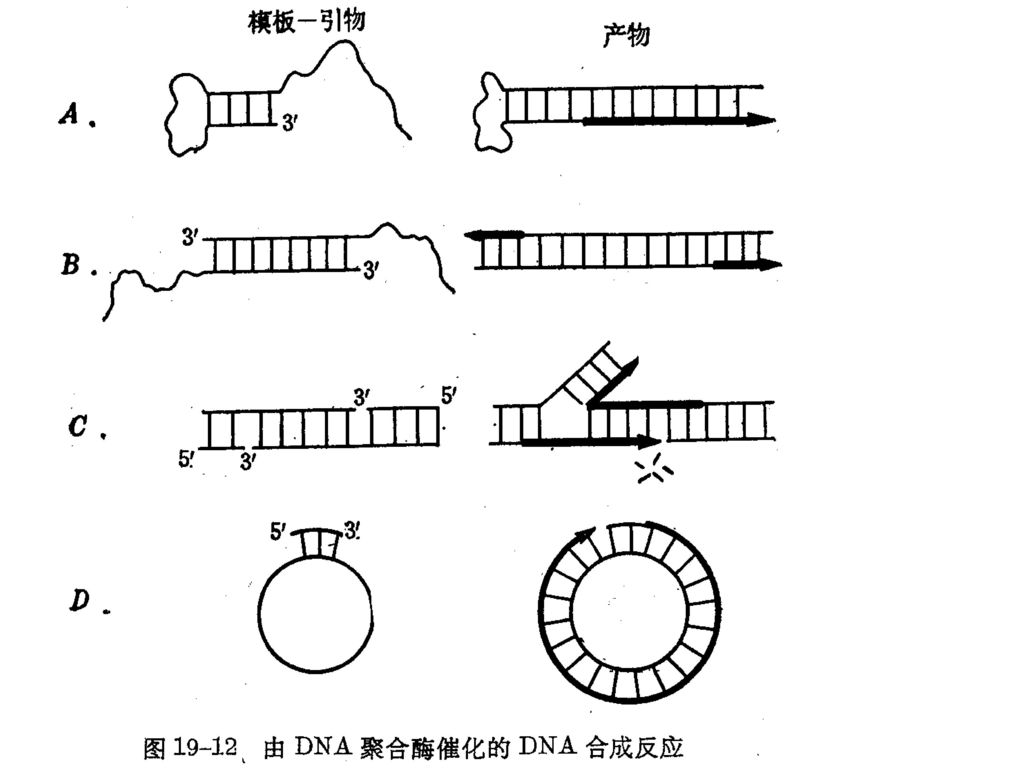
3、 由DNA聚合酶催化的几种DNA聚合类型 P331图19-12

27
4、 E.coli DNA聚合酶 (1)、E.coli. DNA pol. I(Kornberg酶,400 copy/cell)
单体酶,分子量109Kd,含一个Zn2+,每个细胞中含400个DNA pol.Ⅰ 催化活性: 5, → 3, 聚合活性 3, → 5, 外切活性 5, → 3, 外切活性 用蛋白水解酶将DNA pol.Ⅰ部分水解可得: 大片段(Klenow),75Kd,活性:5, → 3,聚合活性、3, → 5,外切活性。 小片段,36Kd,活性:5, → 3,外切活性(只作用于双链DNA的碱基配对部分,切除修复)。 Klenow片段的用途: a 补齐DNA 3,隐缩未端 b. 标记DNA片段未端 c.cDNA合成第二链 d.d DNA测序
,75Kd,活性:5, → 3,聚合活性、3, → 5,外切活性。 小片段,36Kd,活性:5, → 3,外切活性(只作用于双链DNA的碱基配对部分,切除修复)。 Klenow片段的用途: a 补齐DNA 3,隐缩未端. b. 标记DNA片段未端. c.cDNA合成第二链. d.d DNA测序.")
28
(2)、 E.coli. DNA Pol.Ⅱ(100 copy/cell)
单体酶,分子量120Kd 催化活性:5,→ 3,聚合(活性很低) 3,→ 5,外切 可能在DNA的修复中起某中作用。 (3)、 E.coli.DNA pol.Ⅲ(复制酶,10-20 copy/cell) 寡聚酶,全酶由10种共22个亚基组成,α、ε和θ三种亚基组成核心酶。 P334表10-3 DNA pol.Ⅲ是合成新链DNA主要的酶,又称复制酶(Replicase) Pol.Ⅲ的5,→3,外切酶活性只作用于单链DNA。
 3,→ 5,外切. 可能在DNA的修复中起某中作用。 (3)、 E.coli.DNA pol.Ⅲ(复制酶,10-20 copy/cell) 寡聚酶,全酶由10种共22个亚基组成,α、ε和θ三种亚基组成核心酶。 P334表10-3. DNA pol.Ⅲ是合成新链DNA主要的酶,又称复制酶(Replicase) Pol.Ⅲ的5,→3,外切酶活性只作用于单链DNA。")
29
P334 表19-2 E.coli三种DNA聚合酶的性质比较
★DNA聚合酶有6个结合位点 ⑴ 模板DNA结合位点 ⑵ 引物结合位点 ⑶ 引物3,-OH位点、反应位点 ⑷ 底物dNTP结合位点 ⑸ 5, → 3, 外切位点(pol.Ⅱ没有) ⑹ 3, → 5, 外切位点(校正)
 ⑹ 3, → 5, 外切位点(校正)")
30
5、 真核生物DNA聚合酶 P334 表19-4 真核生物DNA聚合酶 真核DNA聚合酶一般不具备外切活力,可能由另外的酶在DNA复制中起校正功能。 ⑴ DNA聚合酶α,多亚基,功能与E.coli. pol.Ⅲ类似,是真核DNA复制酶。 ⑵ DNA聚合酶β,主要在DNA损伤的修复中起作用。 ⑶ DNA聚合酶γ,从线粒体得到,可能与线粒体DNA的复制有关。 ⑷ DNA聚合酶δ,特点:有3, → 5,外切活力

31
(二) 引物酶或RNA聚合酶(引发酶) 细胞内,DNA的复制需要引物(DNA或RNA),引物酶或RNA聚合酶可合成6-10个碱基的RNA引物。 ★DNA复制为什么要用RNA引物?(为什么DNA聚合酶要用引物,RNA聚合酶不需要引物?) P338 ⑴从模板复制最初几个核酸时,碱基堆集力和氢键都较弱,易发生错配 ⑵新复制的最初几个核苷酸,没有与模板形成稳定双链,DNA聚合酶的5,→3,校对功能难发挥作用。

32
(三)、 解螺旋酶 大肠杆菌的解螺旋酶Ⅰ、Ⅱ、Ⅲ与rep蛋白共同作用,将DNA两条链解开。 解螺旋酶I、II、III沿着模板链的5’→3’方向随着复制叉的前进而移动,而rep蛋白则在另一条模板链上沿3’→5’方向移动。
、 解螺旋酶 大肠杆菌的解螺旋酶Ⅰ、Ⅱ、Ⅲ与rep蛋白共同作用,将DNA两条链解开。 解螺旋酶I、II、III沿着模板链的5’→3’方向随着复制叉的前进而移动,而rep蛋白则在另一条模板链上沿3’→5’方向移动。")
33
(四) DNA旋转酶 属DNA拓扑异构酶Ⅱ,可引入负超螺旋,消除复制叉前进时带来的扭曲张力。 拓扑异构酶分两类:I和II,广泛存在于原核生物和真核生物。 拓扑异构酶I使DNA的一条链发生断裂和再连接,反应无须供给能量,主要集中在活性转录区,与转录有关。 拓扑异构酶Ⅱ使DNA的两条链同时断裂和再连接,当它引入超螺旋时需要由ATP供给能量。分布在染色质骨架蛋白和核基质部,与复制有关。

34
(五) 单链DNA结合蛋白(SSB) 复制叉上的解螺旋酶,沿双链DNA前进,产生单链区,大量的单链DNA结合蛋白与单链区结合,阻止复性和保护单链DNA不被核酸酶降解。
 单链DNA结合蛋白(SSB) 复制叉上的解螺旋酶,沿双链DNA前进,产生单链区,大量的单链DNA结合蛋白与单链区结合,阻止复性和保护单链DNA不被核酸酶降解。")
35
(六) DNA连接酶(ligase) (七) DNA复制的拓扑结构 P338-339 连接双链DNA上的切口。
大肠杆菌连接酶只能在模板上连接DNA缺口。T4DNA ligase即可连接粘性末端的DNA,又可连接平齐末端的双链DNA。 E.coli.和其它细菌的DNA ligase以NAD为能源,动物细胞和噬菌体DNA ligase以ATP为能源。 (七) DNA复制的拓扑结构 P
 DNA复制的拓扑结构. P")
36
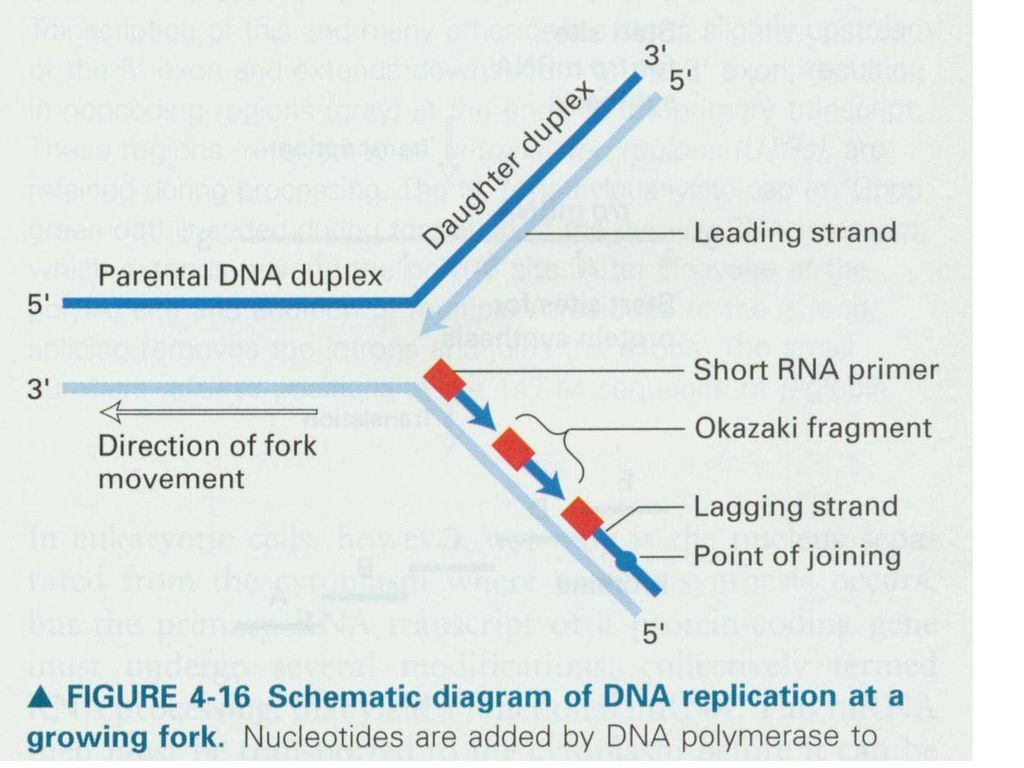
四、 DNA的半不连续复制 P336 图19-15 DNA的半不连续复制 DNA聚合酶催化的方向是5,→3,。 前导链: 滞后链:
1968年,发现冈崎片段。长度: 细菌:1Kb-2Kb,相当于一个顺反子的大小。 真核: bp,约等于一个核小体DNA的长度。

38
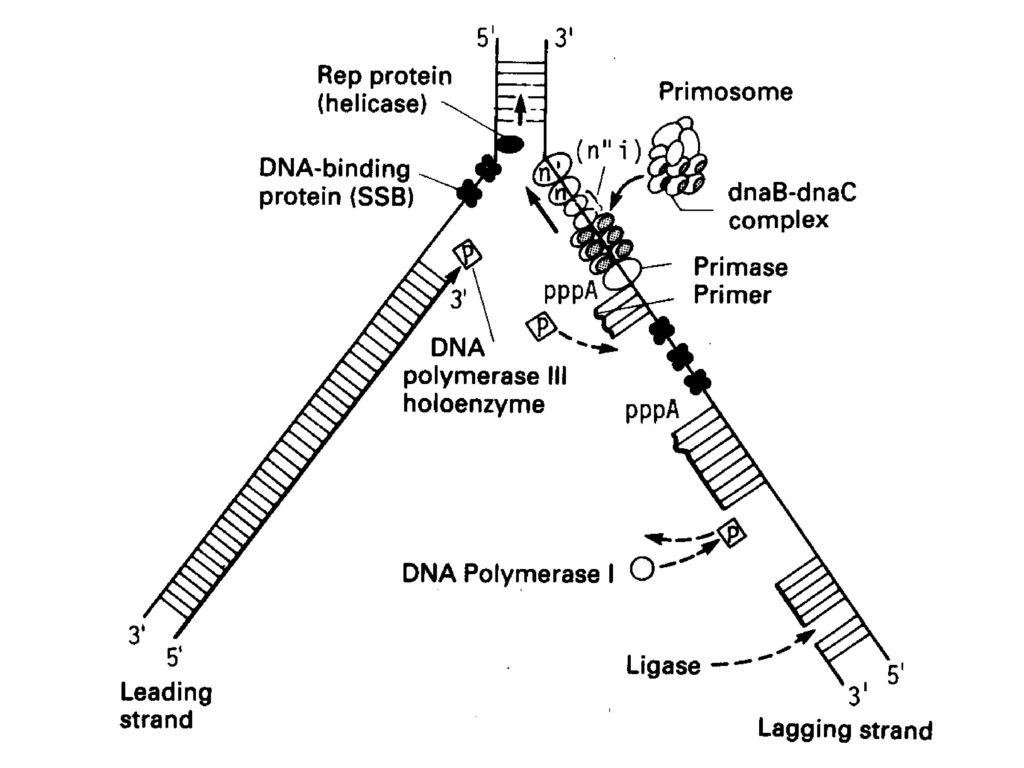
五、 DNA复制过程(E.coli.) P342 图 大肠杆菌的复制体结构示意图
 P342 图19-17 大肠杆菌的复制体结构示意图")
40
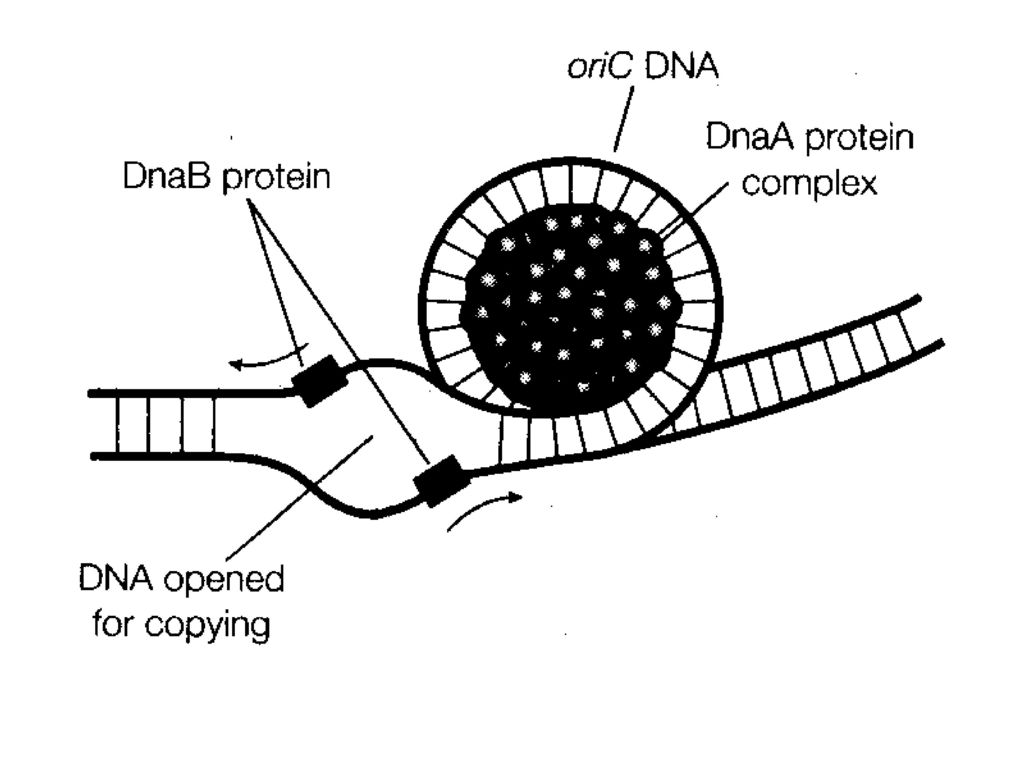
1、 复制的起始 引发:当DNA的双螺旋解开后,合成RNA引物的过程。 引发体:引物合成酶与各种蛋白质因子(dnaB、dnaC、n、n'n''I)构成的复合体,负责RNA引物的合成。 引发体沿着模板链5’→3’方向移动(与冈崎片段合成的方向正好相反,而与复制叉移动的方向相同),移到一定位置上即可引发RNA引物的合成。 E.coli.DNA复制原点ori C,由245bp组成,三组13bp重复序列(近5,端处),四组9 bp重复序列(另一端处)。 图
,移到一定位置上即可引发RNA引物的合成。 E.coli.DNA复制原点ori C,由245bp组成,三组13bp重复序列(近5,端处),四组9 bp重复序列(另一端处)。 图.")
41
★大肠杆菌复制原点起始复制所需蛋白质: DNaA 在原点处打开双螺旋 DNaB 使DNA解旋 DNaC DNaB结合在原点所需 Hu 刺激起始 引物酶(DNaG) 合成RNA引物 SSB 结合单链DNA RNA聚合酶 促进DNaA活性 旋转酶 松驰DNA扭曲应力 20个DnaA结合在四组9bp重复区,形成起始复合物,DNA环绕此复合物。 三组13bp重复区依次变性,产生开放型复合物。 DnaB(在DnaC协助下)与开放复合物结合,进一步解链。
 合成RNA引物. SSB 结合单链DNA. RNA聚合酶 促进DNaA活性. 旋转酶 松驰DNA扭曲应力. 20个DnaA结合在四组9bp重复区,形成起始复合物,DNA环绕此复合物。 三组13bp重复区依次变性,产生开放型复合物。 DnaB(在DnaC协助下)与开放复合物结合,进一步解链。")
43
2、 DNA链的延长反应 前导链只需要一个RNA引物,后随链的每一个冈崎片段都需要一个RNA引物,链的延长反应由DNA pol.Ⅲ催化。 复制体:在DNA合成的生长点(既复制叉上)分布着许多与复制有关的酶和辅助因子,它们在DNA的模板链形成离散的复合物,彼此配合进行高度精确的复制,称为复制体。 复制体沿着复制叉方向前进就合成DNA。
分布着许多与复制有关的酶和辅助因子,它们在DNA的模板链形成离散的复合物,彼此配合进行高度精确的复制,称为复制体。 复制体沿着复制叉方向前进就合成DNA。")
44
3、 RNA引物的切除及缺口补齐 DNA polⅠ的5, → 3,外切活力,切除RNA引物。 DNApolⅠ的5, → 3,合成活性补齐缺口。 4、 DNA切口的连接 DNA ligase,动物、真核由ATP供能,原核由NAD供能。 5、 DNA合成的终止 环状DNA、线性DNA,复制叉相遇即终止。

45
★ 小结: ⑴ DNA解螺旋酶解开双链DNA。 ⑵ SSB结合于DNA单链。 ⑶ DNA旋转酶引入负超螺旋,消除复制叉前进时带来的扭曲张力。 ⑷ DNA引物酶(在引发体中)合成RNA引物。 ⑸ DNA pol.Ⅲ在两条新生链上合成DNA。 ⑹ DNA polⅠ切除RNA引物,并补上DNA。 ⑺ DNA ligase连接一个冈崎片段。 DNA复制过程中,聚合酶对dTTP和dUTP的分辨能力高,有少量dUTP掺入DNA链中,此时,U-糖苷酶、AP内切酶、DNA polⅠ、DNA ligase共同作用,切除尿嘧啶,接上正确的碱基。

46
六、 真核生物DNA的复制 P343 1、 复制起点和单位 真核生物染色体DNA是多复制子,有多个复制起点,可以多点起始,分段进行复制。每个复制子大多在 bp之间,比细菌染色体DNA(单复制子)小得多。 ★试验证据:5-氟脱氧胞苷标记

47
真核生物DNA复制叉移动的速度此原核的慢,如哺乳动物复制叉移动的速度每分钟1-3Kb,细菌每分钟5Kb。

48
2、 复制过程中组蛋白的装配 核小体的结构(200bp左右)
2、 复制过程中组蛋白的装配 核小体的结构(200bp左右) 在真核生物的复制子上,亲代染色体的核小体被逐个打开,组蛋白以完整的八聚体形式直接转移到子代DNA的前导链上,新合成的组蛋白与后随链组装成核小体。因此,DNA的复制是半保留的,而组蛋白则是全保留的。 ★试验证据:环己酮亚胺抑制组蛋白合成,电子显微镜下观察
 在真核生物的复制子上,亲代染色体的核小体被逐个打开,组蛋白以完整的八聚体形式直接转移到子代DNA的前导链上,新合成的组蛋白与后随链组装成核小体。因此,DNA的复制是半保留的,而组蛋白则是全保留的。 ★试验证据:环己酮亚胺抑制组蛋白合成,电子显微镜下观察.")
49
3、 真核生物DNA复制的终止 端粒:一段DNA序列与蛋白质形成的一种复合体,是真核细胞染色体末端所特有的结构。 功能: ⑴保证线性DNA的完整复制 ⑵保护染色体末端 ⑶决定细胞寿命,胚系细胞含端粒酶,体细胞不表达端粒酶。

50
端粒(telomeres)分布于线性真核染色体未端。酵母端粒约100bp的重复序列,形式为:5,(TxGy)n3,(AxCy) n,x和y一般为1—4。
端粒末端的重复序列,通过端粒酶(telomerase)将其加到染色体末端。 端粒酶含有RNA和蛋白质(起DNA聚合酶的作用)两种组分,RNA分子约159b,含有多个CyAx重复序列,RNA分子用作端粒TxGy链合成的模板。端粒酶是一种反转录酶,它只合成与酶自身的RNA模板互补的DNA片段。 人类体细胞的端粒长度,随个体年龄增加而逐渐缩短。细胞每分裂一次,端粒缩短50-200bp,短至1-4Kbp时,细胞就停止分裂。若能重建端粒,则细胞可以永远分裂。恶性肿瘤细胞端酶表达多。
将其加到染色体末端。 端粒酶含有RNA和蛋白质(起DNA聚合酶的作用)两种组分,RNA分子约159b,含有多个CyAx重复序列,RNA分子用作端粒TxGy链合成的模板。端粒酶是一种反转录酶,它只合成与酶自身的RNA模板互补的DNA片段。 人类体细胞的端粒长度,随个体年龄增加而逐渐缩短。细胞每分裂一次,端粒缩短50-200bp,短至1-4Kbp时,细胞就停止分裂。若能重建端粒,则细胞可以永远分裂。恶性肿瘤细胞端酶表达多。")
51
⑴杂交 图 ⑵聚合 ⑶转位再杂交 ⑷进一步聚合 ⑸非标准GG配对

52
七、 DNA复制的调控 八、 DNA复制的真实性 《杨岐生》P144 生物体DNA复制具有高度真实性,复制 碱基对,只有一个错误碱基。 碱基对的自由能通常在4-13KJ/mol,这样的自由能相当于平均参入100个核苷酸就可能出现一次错配,仅靠Watson-Crick双螺旋的碱基配对原则,突变率将高达10-2 。

53
1、 DNA聚合酶对碱基的选择作用 酶的被动论:不同的核苷酸在聚合位点停留时间不同,正确的dNTP能长时间停留,而参与聚合。DNA聚合酶能依照模板的核苷酸,选择正确的dNTP掺入引物末端。 酶积极参与理论:DNA聚合酶对正确与错误的核苷酸,不仅亲和性不同,而且将它们插入DNA引物端的速度也不同。 动力学校正阅读:在新的磷酸二酯键未形成时,dNTP结合在酶与模板—引物复合物的聚合位点上,DNA聚合酶能识别正确与错误的dNTP。 DNA聚合酶对底物的识别作用,DNA聚合酶有两种底物,一种是DNA模板—引物,另一种是dNTP。 DNA聚合酶先识别DNA模板和引物的3,未端,再识别底物dNTP,是一种有序的识别过程。

54
2、 3,→5,外切活性的校正阅读 E. coli. DNA pol.Ⅰ和pol.Ⅲ有3,→5,外切活性,可删除错误插入的核苷酸。 缺失3, →5,外切活性的E. coli. DNA pol.Ⅰ,催化DNA合成时,出现错误的几率增高5-50倍。因此,3,→5,外切活性可以使DNA复制的真实性,提高1-2个数量级。 图

55
3、 影响DNA合成真实性的因素 ⑴高浓度NMP(如3,-AMP, 5,-GMP) NMP竞争酶的dNTP结合位点,抑制3,→5,外切活性。 ⑵某一种dNTP浓度银高,可使引物3,末端离开外切活性中心。 ⑶dNTP 一般与二价阳离子结合成活化形式,Mg2+为主要的二价阳离子。当用其它二价阳离子(如Mn2+)代替Mg2+时,会改变酶的主体结构,影响聚合活性和3,→3,外切活性。
代替Mg2+时,会改变酶的主体结构,影响聚合活性和3,→3,外切活性。")
56
4、 为什么用RNA引物 ⑴从模板复制最初几个核酸时,碱基堆集力和氢键都较弱,易发生错配 ⑵新复制的最初几个核苷酸,没有与模板形成稳定双链,DNA聚合酶的5,→3,校对功能难发挥作用。

57
第二节 DNA的损伤及修复 DNA的损伤,《罗纪盛》P428 一些物理化学因子如紫外线、电离辐射和化学诱变剂均可引起DNA损伤,破坏其结构与功能。然而在一定条件下,生物机体能使这种损伤得到修复。 紫外线可使DNA分子中同一条链上两个相邻的胸腺嘧啶碱基之间形成二聚体(TT),两个T以共价键形成环丁烷结构。CT、CC间也可形成少量二聚体(CT、CC),使复制、转录受阻。 P346图19-22 细胞内具有一系列起修复作用的酶系统,可以除去DNA上的损伤,恢复DNA的双螺旋结构。目前已知有4种酶修复系统:光复活、切除修复、重组修复、SOS反应诱导的修复,后三种不需要光,又称为暗修复。
,两个T以共价键形成环丁烷结构。CT、CC间也可形成少量二聚体(CT、CC),使复制、转录受阻。 P346图 细胞内具有一系列起修复作用的酶系统,可以除去DNA上的损伤,恢复DNA的双螺旋结构。目前已知有4种酶修复系统:光复活、切除修复、重组修复、SOS反应诱导的修复,后三种不需要光,又称为暗修复。")
58
一、 光复活 1949年已发现光复活现象,可见光(最有效400nm)可激活光复活酶,此酶能分解由于紫外线形成的嘧啶二聚体。高等哺乳动物没有此酶。 P347 图19-23 紫外线损伤的光复活过程
可激活光复活酶,此酶能分解由于紫外线形成的嘧啶二聚体。高等哺乳动物没有此酶。 P347 图19-23 紫外线损伤的光复活过程")
59
A 形成嘧啶二聚体 B. 光复合酶结合于损伤部位 C 酶被可见光激活 D. 修复后释放酶

60
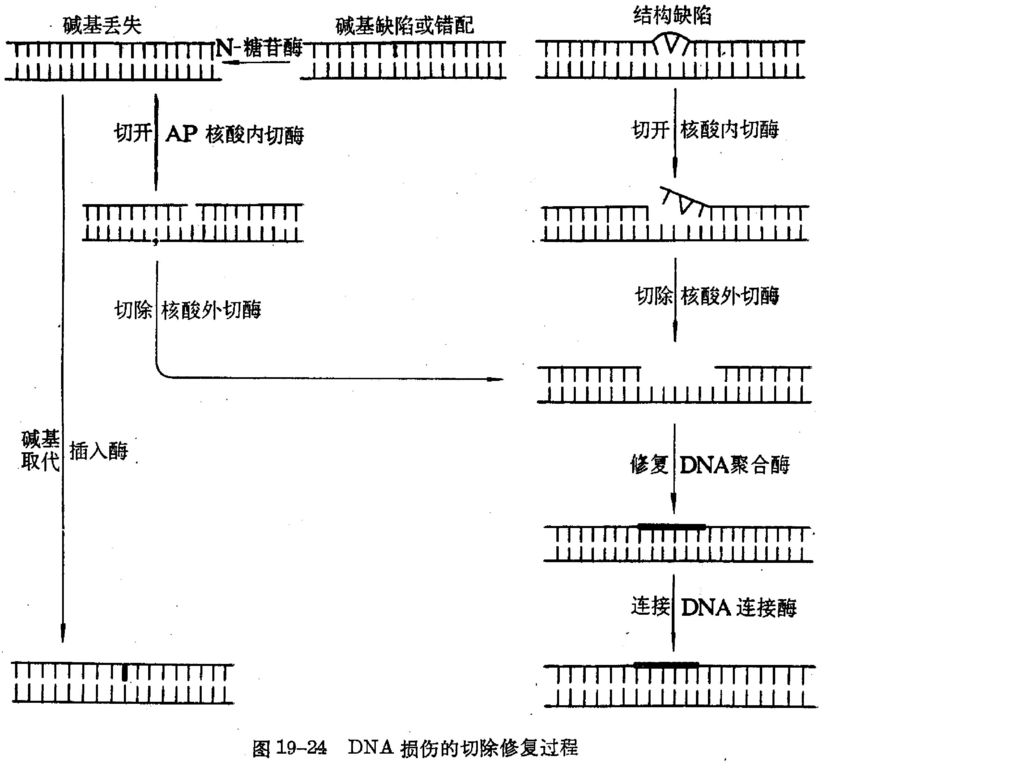
二、 切除修复 P348 图 DNA损伤的切除修复过程 在一系列酶的作用下,将DNA分子中受损伤部分切除,并以完整的那一条链为模板,合成出切去部分,DNA恢复正常结构。 I、结构缺陷的修复: (1)核酸内切酶识别DNA损伤部位,在其附近将其切开。 (2)核酸外切酶切除损伤的DNA。 (3)DNA聚合酶修复。 (4)DNA连接酶连接。 图
核酸内切酶识别DNA损伤部位,在其附近将其切开。 (2)核酸外切酶切除损伤的DNA。 (3)DNA聚合酶修复。 (4)DNA连接酶连接。 图.")
61
II、无嘌呤无嘧啶——碱基缺陷或错配——脱碱基(N-糖苷酶):
甲基磺酸甲酯可使鸟嘌呤第7位氮原子烷基化,活化β—糖苷键,造成脱嘌呤作用;酸也能使DNA脱嘌呤。 DNA复制时,DNA聚合酶对dTTP和dUTP分辨力不高,有少量dUTP掺入DNA链。细胞中的尿嘧啶-N-糖苷酶可以切掉尿嘧啶。腺嘌呤脱氨形成次黄嘌呤时也可以被次黄嘌呤-N-糖苷酶切掉次黄嘌呤。 对于无嘌呤无嘧啶的损伤有两种修复方法: (1) AP核酸内切酶切开,核酸外切酶切除,DNA聚合酶修复,DNA连接酶连接。 (2) 插入酶插入正确碱基
 AP核酸内切酶切开,核酸外切酶切除,DNA聚合酶修复,DNA连接酶连接。 (2) 插入酶插入正确碱基.")
63
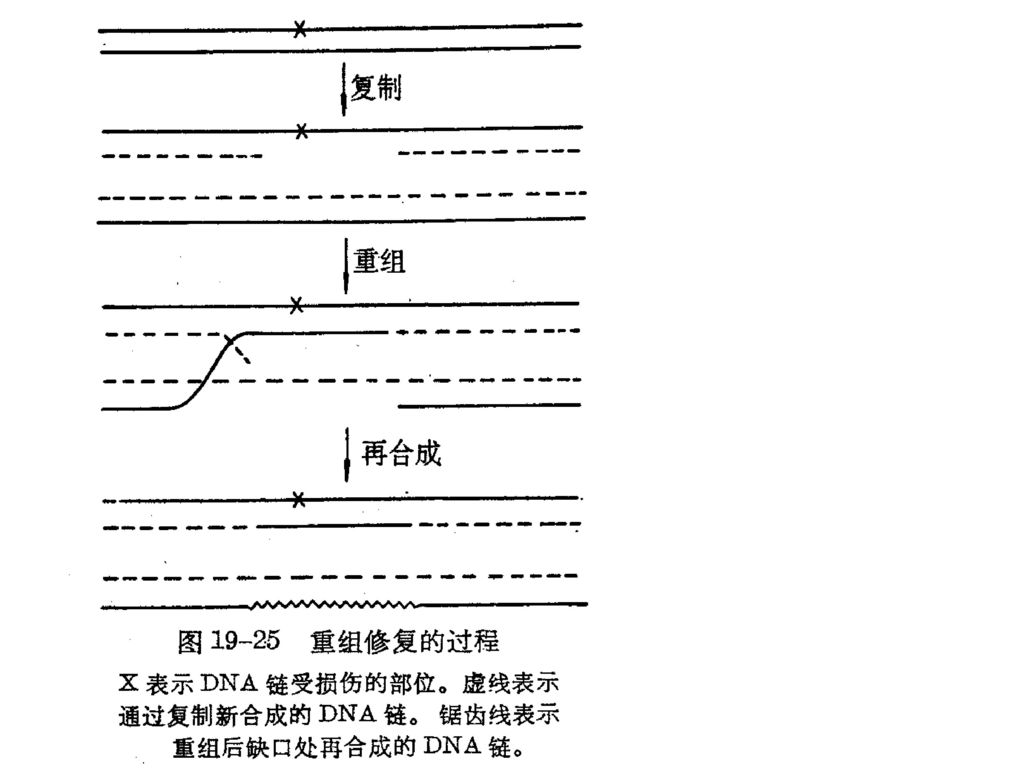
三、 重组修复 P349图19—25重组修复的过程 切除修复发生在DNA复制之前,而当DNA发动复制时尚未修复的损伤部位,可以先复制,再重组修复。 在重组修复过程中,DNA链的损伤并未除去。 重组修复至少需要4种酶组分。 重组基因recA编码一种分子量为40000的蛋白质,它具有交换DNA链的活力。RecA蛋白被认为在DNA重组和重组修复中均起关键作用。 recB、recC基因分别编码核酸外切酶V的两个亚基。 此外,修复合成还需要DNA聚合酶和连接酶。

65
四、 诱导修复和应急反应(SOS反应) 诱导修复是细胞DNA受到严重损伤或DNA复制系统受到抑制的紧急情况下,为求得生存而出现的一系列诱导性修复。 SOS反应诱导的修复系统包括避免差错的修复(无差错修复)和倾向差错的修复。 避免差错的修复:SOS反应能诱导光复活切除修复和重组修复中某些关键酶和蛋白质的产生,从而加强光复活切除修复和重组修复的能力,这属于避免差错的修复。 倾向差错的修复:SOS反应还能诱导产生缺乏校对功能的DNA聚合酶,它能在DNA损伤部位进行复制而避免了死亡,可是却带来了高的突变率,这属于倾向差错的修复。

66
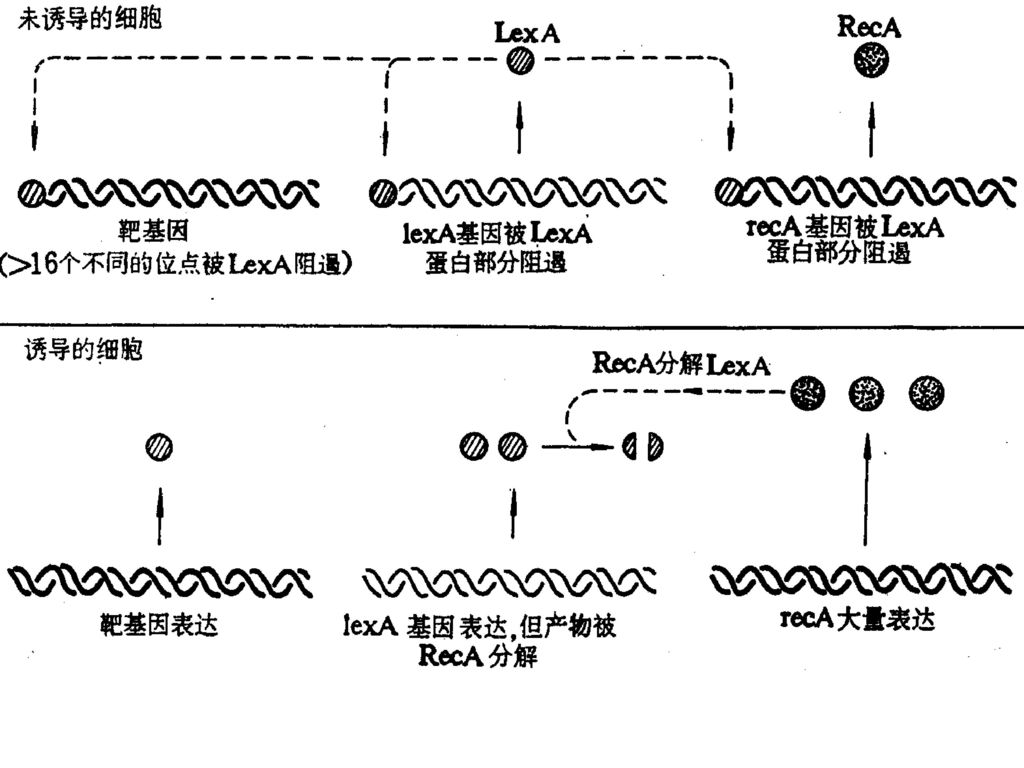
SOS反应是由RecA蛋白和LexA阻遏物相互作用引起的。RecA蛋白不仅在同源重组中起重要作用,而且它也是SOS反应的最初发动因子。在有单链DNA和ATP存在时,RecA蛋白被激活而表现出蛋白水解酶的活力,它能分解λ噬菌体的阻遏蛋白和LexA蛋白。LexA蛋白(22Kd)许多基因的阻遏物,当它被RecA的蛋白水解酶分解后就可以使一系列基因得到表达其中包括紫外线损伤的修复基因uvrA、uvrB、uvrC(分别编码核酸内切酶的亚基)以及recA和lexA基因本身,还有单链结合蛋白基因ssb,与λ噬菌体DNA整合有关的基因himA、与诱变作用有关的基因umuDC,与细胞分裂有关的基因sulA,ruv,和lon,以及一些功能不清楚的基因dinA,B,D,F等。
许多基因的阻遏物,当它被RecA的蛋白水解酶分解后就可以使一系列基因得到表达其中包括紫外线损伤的修复基因uvrA、uvrB、uvrC(分别编码核酸内切酶的亚基)以及recA和lexA基因本身,还有单链结合蛋白基因ssb,与λ噬菌体DNA整合有关的基因himA、与诱变作用有关的基因umuDC,与细胞分裂有关的基因sulA,ruv,和lon,以及一些功能不清楚的基因dinA,B,D,F等。")
68
SOS反应广泛存在于原核生物和真核生物,它是生物在极为不利的环境中求得生存的一种基本功能。
然而癌变有可能也是通过SOS反应造成的,因为能引起SOS反应的作用剂通常都具有致癌作用,如X-射线,紫外线,烷化剂,黄曲霉素等,而某些不能致癌的诱变剂并不引起SOS反应,如5-溴尿嘧啶。目前,有关致癌物的一些简便检测方法就是根据SOS反应原理而设计的,既测定细菌的SOS反应。

69
第三节 RNA指导的DNA合成(反转录) 反转录(reverse transcription):以RNA为模板,合成DNA。与通常转录过程中遗传信息流从DNA到RNA的方向相反。 1970年,Temin 和Baltimore分别从致癌RNA病毒(劳氏肉瘤病毒和鼠白血病病毒)中发现发反转录酶。 致癌RNA病毒是一大类能引起鸟类、哺乳类等动物白血病、肉瘤以及其它肿瘤的病毒。这类病毒侵染细胞后并不引起细胞死亡,却可以使细胞发生恶性转化。经过改造后可以作为基因治疗的载体。 放线菌素D(抑制以DNA为模板的反应,复制和转录)能抑制致癌RNA病毒的复制,可见致癌RNA病毒的复制过程必然涉及DNA。 Bader 用嘌呤霉素(puromycin)来抑制静止细胞蛋白质的合成,发现这种细胞仍能感染劳氏肉瘤病毒(RSV),证实反转录酶是由反转录病毒带入细胞的,而不是感染后在宿主细胞中新合成的。
中发现发反转录酶。 致癌RNA病毒是一大类能引起鸟类、哺乳类等动物白血病、肉瘤以及其它肿瘤的病毒。这类病毒侵染细胞后并不引起细胞死亡,却可以使细胞发生恶性转化。经过改造后可以作为基因治疗的载体。 放线菌素D(抑制以DNA为模板的反应,复制和转录)能抑制致癌RNA病毒的复制,可见致癌RNA病毒的复制过程必然涉及DNA。 Bader 用嘌呤霉素(puromycin)来抑制静止细胞蛋白质的合成,发现这种细胞仍能感染劳氏肉瘤病毒(RSV),证实反转录酶是由反转录病毒带入细胞的,而不是感染后在宿主细胞中新合成的。")
70
一、 反转录酶 由一个α亚基和一个β亚基组成,含有Zn2+,具有三种酶活力。
一、 反转录酶 由一个α亚基和一个β亚基组成,含有Zn2+,具有三种酶活力。 (1)RNA指导的DNA聚合酶活力(以RNA为模板,合成一条互补的DNA,形成RNA—DNA杂种分子)。 (2)RNase H酶活力,水解RNA—DNA杂种分子中的RNA,可沿3’→5’和5’→3’两个方向起外切酶作用。 (3)DNA指导的DNA聚合酶活力。 模板:RNA或DNA 以自身病毒类型的RNA为模板时,该酶的反转录活力最大,但是带有适当引物的任何种类的RNA都能作为合成DNA的模板。 引物:RNA或DNA 底物:dNTP 二价阳离子:Mg2+或Mn2+ 真核mRNA3’端有polyA,加入oligo dT后,可以作为反转录酶的模板,合成cDNA。
RNA指导的DNA聚合酶活力(以RNA为模板,合成一条互补的DNA,形成RNA—DNA杂种分子)。 (2)RNase H酶活力,水解RNA—DNA杂种分子中的RNA,可沿3’→5’和5’→3’两个方向起外切酶作用。 (3)DNA指导的DNA聚合酶活力。 模板:RNA或DNA. 以自身病毒类型的RNA为模板时,该酶的反转录活力最大,但是带有适当引物的任何种类的RNA都能作为合成DNA的模板。 引物:RNA或DNA. 底物:dNTP. 二价阳离子:Mg2+或Mn2+ 真核mRNA3’端有polyA,加入oligo dT后,可以作为反转录酶的模板,合成cDNA。")
71
二、 病毒RNA的反转录过程 所有已知的致癌RNA病毒都含有反转录酶,因此被称为反转录病毒(retrovirus),反转录病毒的复制需要经过一个DNA中间体(前病毒)。 1、 反转录病毒的基因组结构 P353 图19-27 (1) 反转录病毒基因组通常由两条相同的(+)RNA链组成。5’端附近区域以氢键结合在一起,全长7-10Kb。 (2) 每一条RNA链的两端具有相同的序列,形成正向重复序列。 (3) 5’端有帽子结构,3’端有polyA,与真核mRNA相似。 (4) 5’端带有1分子的宿主tRNA,作为反转录时的引物。某些鸟类反转录病毒携带的是tRNAtrp,鼠类是tRNApro
 反转录病毒基因组通常由两条相同的(+)RNA链组成。5’端附近区域以氢键结合在一起,全长7-10Kb。 (2) 每一条RNA链的两端具有相同的序列,形成正向重复序列。 (3) 5’端有帽子结构,3’端有polyA,与真核mRNA相似。 (4) 5’端带有1分子的宿主tRNA,作为反转录时的引物。某些鸟类反转录病毒携带的是tRNAtrp,鼠类是tRNApro.")
72
2、 反转录过程。 当致癌RNA病毒侵染宿主细胞时,病毒RNA及反转录酶一起进入宿主细胞,病毒自身带入的反转录酶使RNA反转录成双链DNA。 (1) 以病毒(+)RNA为模板,合成互补的(-)DNA。 (2) 切除RNA—DNA杂种分子中的RNA。 (3) 以(-)DNA链为模板,合成(+)DNA链,最后形成两端带有LTR(长末端重复序列)的双链DNA。 反转录病毒只有整合到宿主染色体DNA后才能被转录,转录产物经拼接可以产生不同的病毒mRNA。LTR(长末端重复序列)对前病毒DNA整合到宿主染色体DNA以及整合后的转录均起着重要作用。 反转录病毒合成的过程: 图 缺口的模板(基因组)RNA,在U3旁生成一个正链DNA的合成RNA的引物,而其余的模板RNA被降解。 正链DNA合成开始,复制。
 切除RNA—DNA杂种分子中的RNA。 (3) 以(-)DNA链为模板,合成(+)DNA链,最后形成两端带有LTR(长末端重复序列)的双链DNA。 反转录病毒只有整合到宿主染色体DNA后才能被转录,转录产物经拼接可以产生不同的病毒mRNA。LTR(长末端重复序列)对前病毒DNA整合到宿主染色体DNA以及整合后的转录均起着重要作用。 反转录病毒合成的过程: 图. 缺口的模板(基因组)RNA,在U3旁生成一个正链DNA的合成RNA的引物,而其余的模板RNA被降解。 正链DNA合成开始,复制。")
73
3、 反转录病毒的生活周期 P354 图19-29 (1) 病毒粒子侵染细胞,病毒RNA和反转录酶一起进入细胞。 (2) RNA被反转录成双链DNA(前病毒),环化,进入细胞核。 (3) 反转录病毒的DNA整合到宿主染色体DNA中。 (4) 前病毒DNA进行复制,转录出功能基因、基因组RNA和病毒蛋白。 (5) 基因组RNA和病毒蛋白在胞质中组装成新病毒粒子,转移到质膜,通过出芽方式释放新病毒粒子。
 反转录病毒的DNA整合到宿主染色体DNA中。 (4) 前病毒DNA进行复制,转录出功能基因、基因组RNA和病毒蛋白。 (5) 基因组RNA和病毒蛋白在胞质中组装成新病毒粒子,转移到质膜,通过出芽方式释放新病毒粒子。")
74
三、 反转录的生物学意义。 1.反转录酶存在于所有致癌RNA病毒中,它的存在与RNA病毒引起细胞恶性转化有关。 2.艾滋病毒(AIDS) 人类免疫缺陷病毒(HIV),也是一种反转录病毒,主要感染T4淋巴细胞和B淋巴细胞。病毒粒子直径100nm,球状,粒子外包被两层脂质质膜,膜上有糖蛋白(gp120、gp41),另有两层衣壳蛋白p24、p18。 HIV基因组由两条单链正链RNA组成,每个链长9.7kb,RNA 5,端有帽子结构,3’端有PolyA,链上结合有反转录酶。
,也是一种反转录病毒,主要感染T4淋巴细胞和B淋巴细胞。病毒粒子直径100nm,球状,粒子外包被两层脂质质膜,膜上有糖蛋白(gp120、gp41),另有两层衣壳蛋白p24、p18。 HIV基因组由两条单链正链RNA组成,每个链长9.7kb,RNA 5,端有帽子结构,3’端有PolyA,链上结合有反转录酶。")
75
3.乙肝病毒 大蛋白、中蛋白、主蛋白(表面抗原) 核心抗原 双链环状DNA 乙肝病毒与反转录病毒的区别: P355 4、真核生物正常细胞内也存在反转录过程 真核生物的谈色体基因组中存在为数众多的逆假基因和逆基因。 逆假基因:无启动子和内含子,但有polyA的残基,推测是由mRNA反转录后整合到基因组中去的。 逆基因:具有启动子和转录功能,无内含子。可能是由于mRNA反转录后刚好整合到启动子的下游处,或者是带启动子的RNA序列反转录后整合到基因组中

76
第四节 DNA合成技术 一、 cDNA合成 1、 cDNA文库的构建 cDNA:以mRNA为模板,用反转录酶合成第一链,去除mRNA,合成第二链。 cDNA文库是获得真核结构基因的最好方法,成熟的mRNA无内含子。

77
(1)、 真核mRNA的分离纯化 特点:含量少,不均一,表达具有发育阶段性和组织特异性。 总RNA: rRNA ~85% mRNA ~5% tRNA及其它小分子RNA 10~15% 不均一:在1~5%的mRNA中,有10000—30000种mRNA。 分离、纯化: 用Oligo dT纤维素柱(亲和层析法),加入总RNA,高盐洗脱,先流出非mRNA,降低盐浓度,加入Oligo dA竞争,可洗出mRNA 混合物。 免疫法可分离特定的mRNA。
,加入总RNA,高盐洗脱,先流出非mRNA,降低盐浓度,加入Oligo dA竞争,可洗出mRNA 混合物。 免疫法可分离特定的mRNA。")
78
(2)、 cDNA合成(反转录酶) A. 自身引物法(S1核酸酶降解法) 图 Oligo(dT)15-18个核苷酸
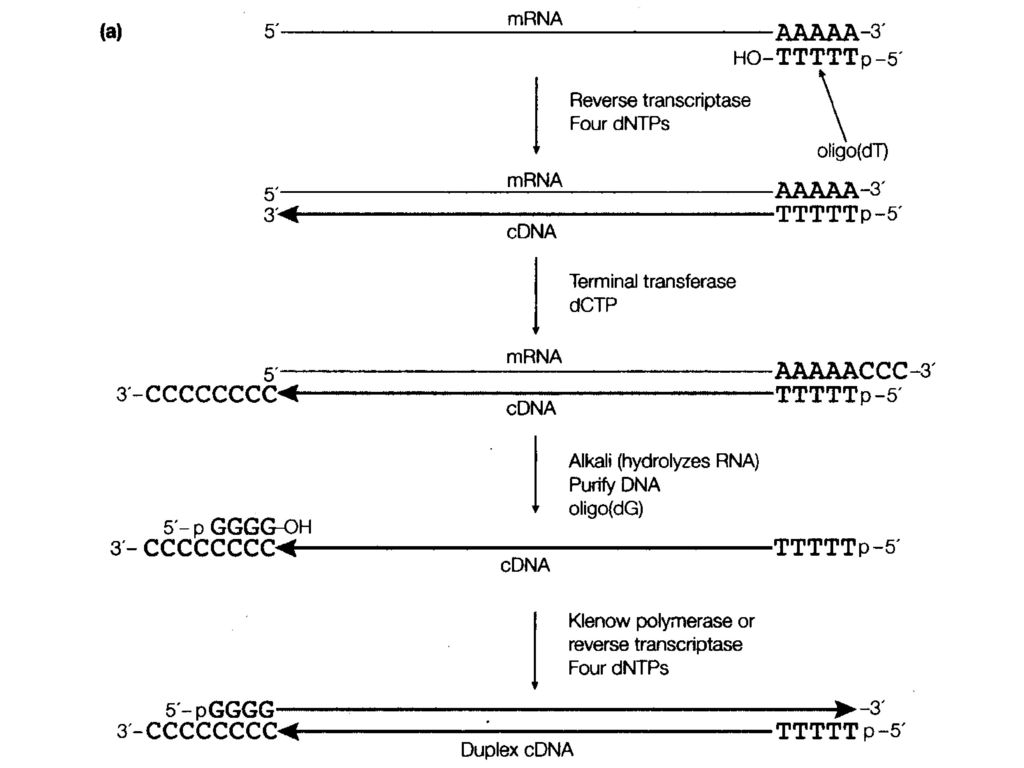
mRNA5’端序列有丢失。 B. 取代合成法(较常用) Oliyo(dT)与mRNA3’端AAA杂交作为引物,合成第一条DNA链。 RNaseH在mRNA上产生多个切口。 DNA pol.Ⅰ切口平移,DNA ligase连接,合成出第二条DNA链。 T4DNA pol.切去端头的RNA-DNA杂交链。 C. 引物合成法 可以合成全长cDNA,mRNA的5’端不丢失。
 Oliyo(dT)与mRNA3’端AAA杂交作为引物,合成第一条DNA链。 RNaseH在mRNA上产生多个切口。 DNA pol.Ⅰ切口平移,DNA ligase连接,合成出第二条DNA链。 T4DNA pol.切去端头的RNA-DNA杂交链。 C. 引物合成法. 可以合成全长cDNA,mRNA的5’端不丢失。")
79
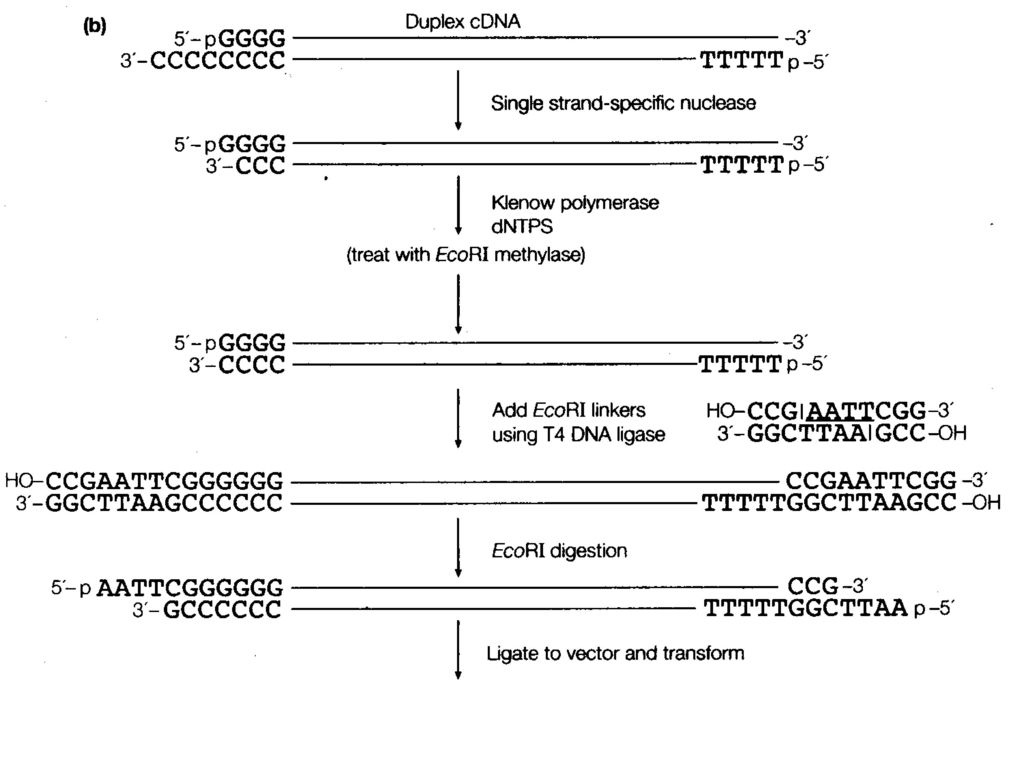
D、末端转移酶法

82
(3)、 cDNA与载体连接 (4)、 重组体的转化 (5)、 扩增、保存 2、 获取特定mRNA的cDNA (1)免疫法分离特定的mRNA (2)PCR法
PCR法.")
83
二、 PCR技术(聚合酶链式反应) Polymerase chain Reaction 以目的基因或DNA片段为模板,在引物介导及Taq DNA聚合酶催化下,在体外用核苷酸大量合成目的基因或DNA片段。 它能快速、专一地扩增所希望得到的目的基因或DNA片段。

84
1、 反应物 (1)模板 单、双链DNA或cDNA都可以作为PCR的模板,若以RNA为起始材料,则须经反转录,获得第一条cDNA后才能用于PCR。 (2)Taq DNA聚合酶 DNA聚合酶是进行PCR扩增的关键,从水生栖热菌(Thermus aquaticns VT-1)分离出来。 Taq DNA聚合酶有很好的热稳定性,92.5℃处理130min,仍保留50%的酶活性,在74℃活性最高,错误掺入核苷酸的比率为1/7500。 (3)引物 引物是决定PCR结果的关键,它由寡核苷酸组成,15-30个b (4)核苷酸dNTP dNTP的浓度50-200umul/L。 (5)镁离子Mg2+
分离出来。 Taq DNA聚合酶有很好的热稳定性,92.5℃处理130min,仍保留50%的酶活性,在74℃活性最高,错误掺入核苷酸的比率为1/7500。 (3)引物. 引物是决定PCR结果的关键,它由寡核苷酸组成,15-30个b. (4)核苷酸dNTP. dNTP的浓度50-200umul/L。 (5)镁离子Mg2+")
85
2、 PCR原理 图 变性 95℃ 复性 55℃ 延伸 72℃


 ” 。 他的体内没有任何免疫系统, 没有任何抵御细菌、病毒的能力。>")
 RNA 的提取与制备 1. 工业用 RNA 的提取 ( 1 ) RNA 及其工业来源 通常在细菌中 RNA 占 5 % -25 %,在酵母中占 2.7 %~ 15 %, 在霉菌中占 0.7%~28%,>")
 發現遺傳因子。 1. 基因的不同等位基因解釋了諸多的遺傳性狀。 2. 對每一種性狀而言,一種生物體遺傳有兩個等位基因, 每一個等位基因得自於一方親代.>")


![学案6 基础实验 [题型剖析] 教材中基础实验的考查是近几年试题命制的一个趋势,本类试题主要考查考生对相关实验的目的、原理、方法、操作步骤和相关技能的掌握情况。主要分为显微观察类、物质鉴定类、探究设计类、调查模拟类。 [突破策略] 首先要将教材中的实验分类归纳;然后对教材中每个实验的目的、原理、材料、步骤及注意事项等进行比较记忆,要认真领悟每个实验的设计意图,并从其中提炼总结实验方法和技术。](/60/11174577/big_thumb.jpg "学案6 基础实验 [题型剖析] 教材中基础实验的考查是近几年试题命制的一个趋势,本类试题主要考查考生对相关实验的目的、原理、方法、操作步骤和相关技能的掌握情况。主要分为显微观察类、物质鉴定类、探究设计类、调查模拟类。 [突破策略] 首先要将教材中的实验分类归纳;然后对教材中每个实验的目的、原理、材料、步骤及注意事项等进行比较记忆,要认真领悟每个实验的设计意图,并从其中提炼总结实验方法和技术。>")


>")


 染色體:細胞核上(細胞未分裂前稱為染色質)>")




 一、蛋白质的结构(p378)>")


