Download presentation
1
第三章 植物的光合作用 教师:李侠 学院:生命科学学院 《植物物生理学》

2
第一节 光合作用的重要性 一、光合作用概念 (一)碳素营养 (二)碳素同化作用 (三)光合作用(photosynthesis)
第一节 光合作用的重要性 一、光合作用概念 (一)碳素营养 (二)碳素同化作用 (三)光合作用(photosynthesis) 绿色植物利用光能把CO2和水合成有机物,同时释放氧气的过程。 CO2+2H2O* 光 绿色植物(CH2O)+ O2*+ H2O
碳素营养. (二)碳素同化作用. (三)光合作用(photosynthesis) 绿色植物利用光能把CO2和水合成有机物,同时释放氧气的过程。 CO2+2H2O* 光 绿色植物(CH2O)+ O2*+ H2O.")
3
二、光合作用的意义 (一)无机物变为有机物 约合成5千亿吨/年 (二)太阳能转变为可贮存的化学能 转化3.2×1021J/y的日光
(三)维持大气中O2和CO2的相对平衡 释放出5.35千亿吨氧气/年 (四)指导科学研究
维持大气中O2和CO2的相对平衡. 释放出5.35千亿吨氧气/年. (四)指导科学研究.")
4
第二节 叶绿体和光合色素

5
一、叶绿体发育、基本结构和成分 (一)叶绿体的发育、形态及分布 1.发育 2.形态 由前质体发育而来。
叶绿体(chloroplast)是光合作用最重要的细胞器。它分布在叶肉细胞的细胞质中。 (一)叶绿体的发育、形态及分布 1.发育 由前质体发育而来。 在光照下合成叶绿素,使前质体发育成叶绿体。 2.形态 扁平椭圆形 一个叶肉细胞中约有10至数百个叶绿体,其长3~6μm,厚2~3μm。
是光合作用最重要的细胞器。它分布在叶肉细胞的细胞质中。 (一)叶绿体的发育、形态及分布. 1.发育. 由前质体发育而来。 在光照下合成叶绿素,使前质体发育成叶绿体。 2.形态. 扁平椭圆形. 一个叶肉细胞中约有10至数百个叶绿体,其长3~6μm,厚2~3μm。")
6
水稻叶绿体 玉米叶绿体

7
3.分布 4.运动 叶肉细胞中的叶绿体较多分布在与空气接触的质膜旁, 有利于叶绿体同外界进行气体交换。 随原生质环流运动
随光照的方向和强度而运动

8
侧视图 叶绿体 俯视图 棉叶栅栏细胞 叶绿体随光照的方向和强度而运动

9
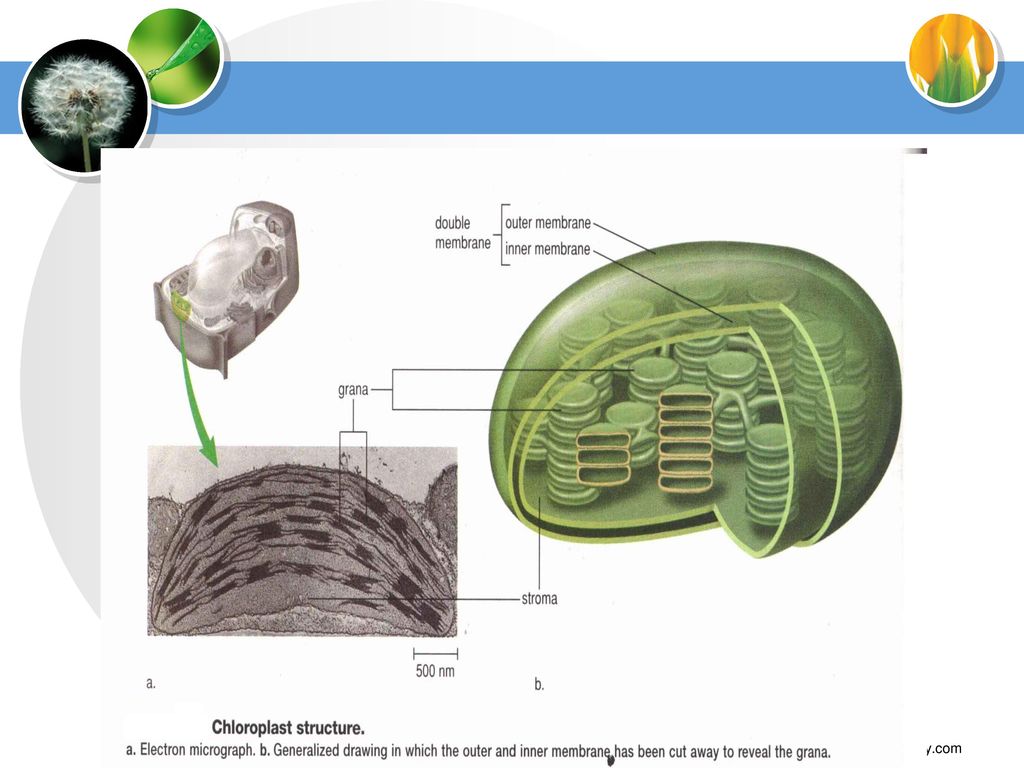
(二) 叶绿体的基本结构
 叶绿体的基本结构")
10
1.被膜 由两层单位膜组成,两膜间距5~10nm。被膜上无叶绿素, 外膜:非选择性膜 。 内膜:选择透性膜。
主要功能是控制物质的进出,维持光合作用的微环境。

12
2.基质(间质)及内含物 3.类囊体(片层) 被膜以内的基础物质; 成分:可溶性蛋白质,其它代谢活跃物质; 内含物:淀粉粒,质体小球。
基质类囊体:伸展在基质中彼此不重叠; 基粒类囊体:可自身或与基质类囊体重叠,组成基粒。

13
(三)叶绿体的成分
叶绿体的成分")
14
二、光合色素的化学特性 (一)叶绿素 叶绿素a 叶绿素b
叶绿素 叶绿素a 叶绿素b")
15
叶绿醇 卟啉环

16
(二)类胡萝卜素(carotenoid)
3 (紫罗兰酮环) 环己烯 橙黄色 黄色 (二)类胡萝卜素(carotenoid) 胡萝卜素(C40H56)呈橙黄色,有α、β、γ三种同分异构体; 叶黄素(C40H56O2)是由胡萝卜素衍生的醇类
 环己烯. 橙黄色. 黄色. (二)类胡萝卜素(carotenoid) 胡萝卜素(C40H56)呈橙黄色,有α、β、γ三种同分异构体; 叶黄素(C40H56O2)是由胡萝卜素衍生的醇类.")
17
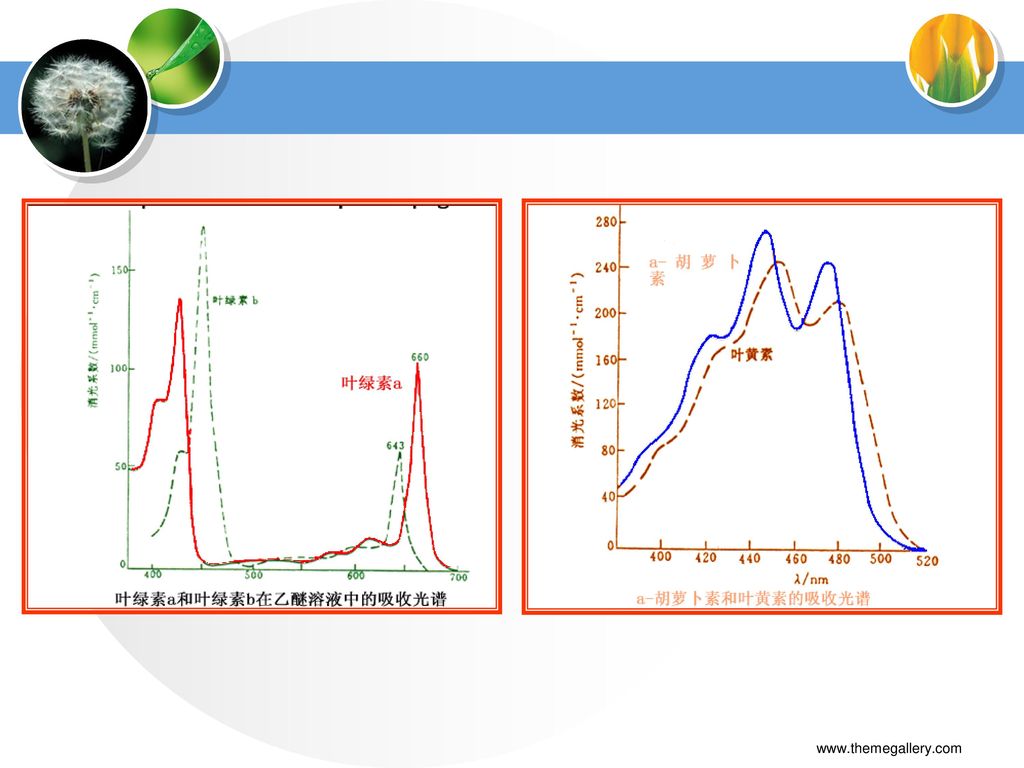
三、光合色素的光学特性 (一)吸收光谱 1.叶绿素吸收光谱 2.类胡萝卜素吸收光谱
两个强吸收峰区:640~660nm的红光, 430~450nm的蓝紫光 叶绿素a在红光区的吸收峰比叶绿素b的高,蓝紫光区的吸收峰则比叶绿素b的低。 2.类胡萝卜素吸收光谱 类胡萝卜素吸收带在400~500nm的蓝紫光区 基本不吸收黄光,从而呈现黄色。

19
(二)荧光现象 1.概念 2.形成机制
荧光现象 1.概念 2.形成机制")
20
为什么叶绿素发出的荧光是红色的? 为什么植物的叶片观察不到荧光现象?

21
(三)磷光现象 1.概念 2.特点
磷光现象 1.概念 2.特点")
22
四、叶绿素的形成(一)生物合成
生物合成")
23
(二) 植物叶色 1.正常叶片中 2.影响叶片中色素数量的因素 叶绿素和类胡萝卜素的分子比例 约为3:1;
叶绿素a与叶绿素b的分子比例也约为3:1; 叶黄素与胡萝卜素约为2:1。 2.影响叶片中色素数量的因素 植物种类 叶片老嫩 生育期 季节

24
一般叶片中叶绿素与类胡萝卜素的比值约为3∶1,所以正常的叶子总呈现绿色。秋天或在不良的环境中,叶片中的叶绿素较易降解,数量减少,而类胡萝卜素比较稳定,所以叶片呈现黄色。

25
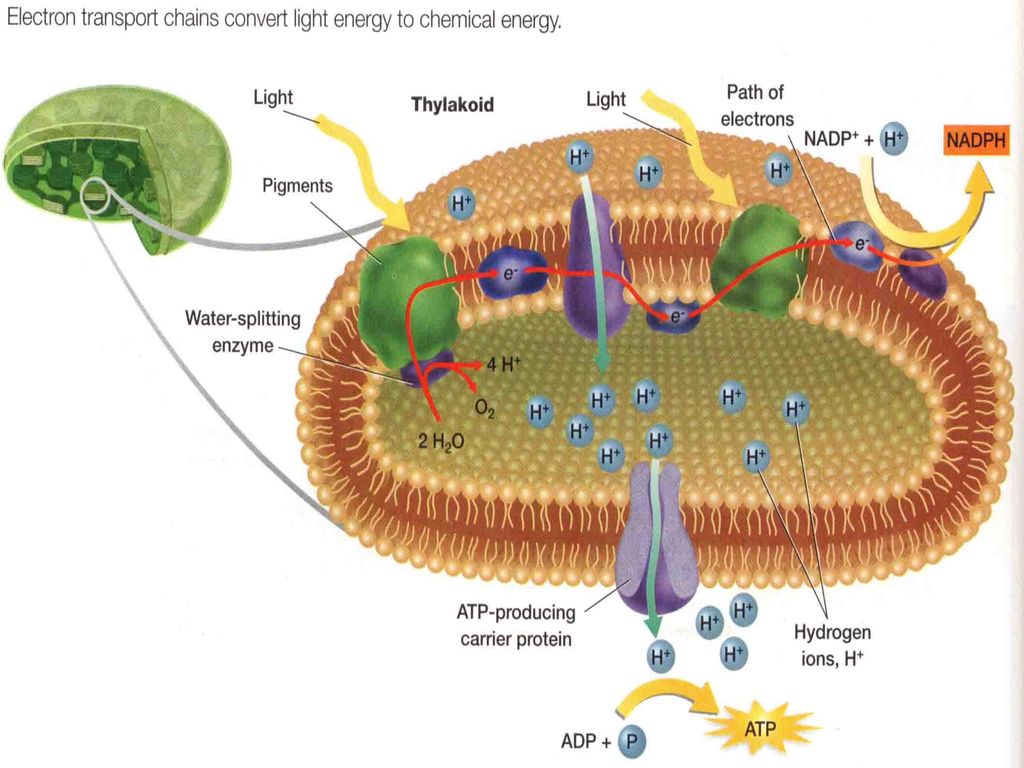
第三节 光合作用过程(Ⅰ):光的吸收 一、光反应和碳反应
:光的吸收 一、光反应和碳反应")
26
表1 光合作用中各种能量转变情况 能量转变 光能 (光化学反应) 活跃的化学能 稳定的化学能 是否需光 光 光或暗
能量转变 光能 (光化学反应) 活跃的化学能 稳定的化学能 是否需光 光 光或暗 贮能物质 量子 电子 ATP、NADPH 碳水化合物等 转变过程 原初反应 电子传递 光合磷酸化 碳同化 时间跨度(s) 反应部位 类囊体膜 叶绿体基质
 活跃的化学能 稳定的化学能. 是否需光 光 光或暗. 贮能物质 量子 电子 ATP、NADPH2 碳水化合物等. 转变过程 原初反应 电子传递 光合磷酸化 碳同化. 时间跨度(s) 反应部位 类囊体膜 叶绿体基质.")
27
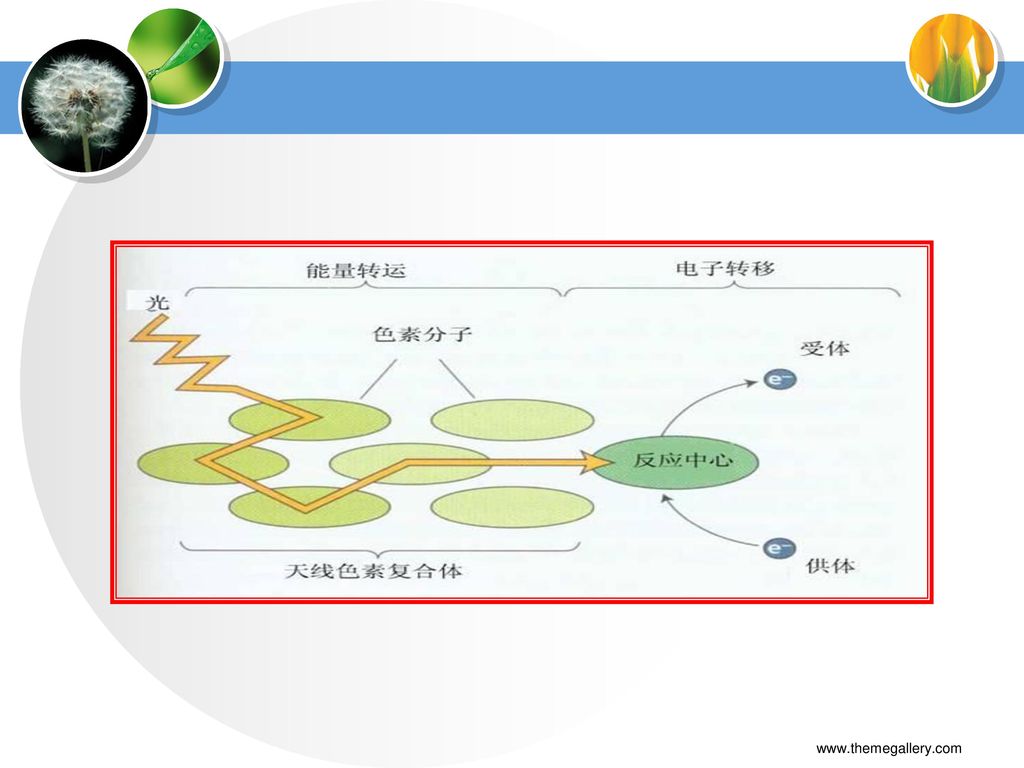
二、原初反应 (一)概念 (二)组成 指从光合色素分子被光激发,到引起第一个光化学反应为止的过程。 聚光色素系统+反应中心=光合单位

30
1.聚光色素(天线色素) 概念:没有光化学活性,只有收集光能的作用 组成:大部分chla 和全部chlb、胡萝卜素、叶黄素
功能:吸收传递光能,方式有激子传递和共振传递

31
D·〔P+·A-〕·A1 D+·〔P·A〕·A1-
2.反应中心 概念:将光能转变为化学能的膜蛋白复合体 组成:反应中心色素分子、原初电子受体、原初电子供体、次级电子供体和受体等 反应式: D·〔P+·A-〕·A D+·〔P·A〕·A1-

32
第四节 光合作用过程(Ⅱ): 电子传递与光合磷酸化
一、光系统(PSⅠ和PSⅡ) 红降现象和双光增益效应
 红降现象和双光增益效应.")
33
二、光合电子传递体及其功能 PSⅠ、PSⅡ、细胞色素b6f、质体醌、质体蓝素、Fd和FNR

34
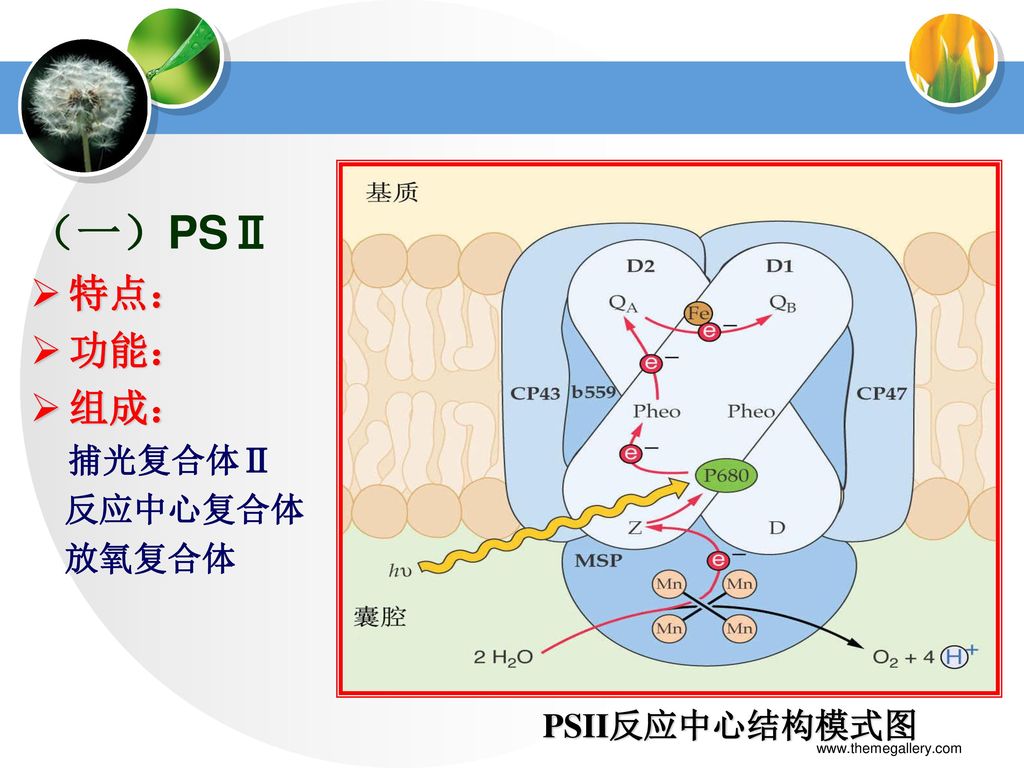
(一)PSⅡ 特点: 功能: 组成: 捕光复合体Ⅱ 反应中心复合体 放氧复合体 PSII反应中心结构模式图
35
(二)质醌(PQ) 特点: 作用: 机制: 也叫质体醌,是PSⅡ反应中心的末端电子受体;
脂溶性分子,能在类囊体膜中自由移动,转运电子与质子; 含量高,称“PQ库”。 作用: 传递电子、建立类囊体膜内外建立质子梯度 机制:

36
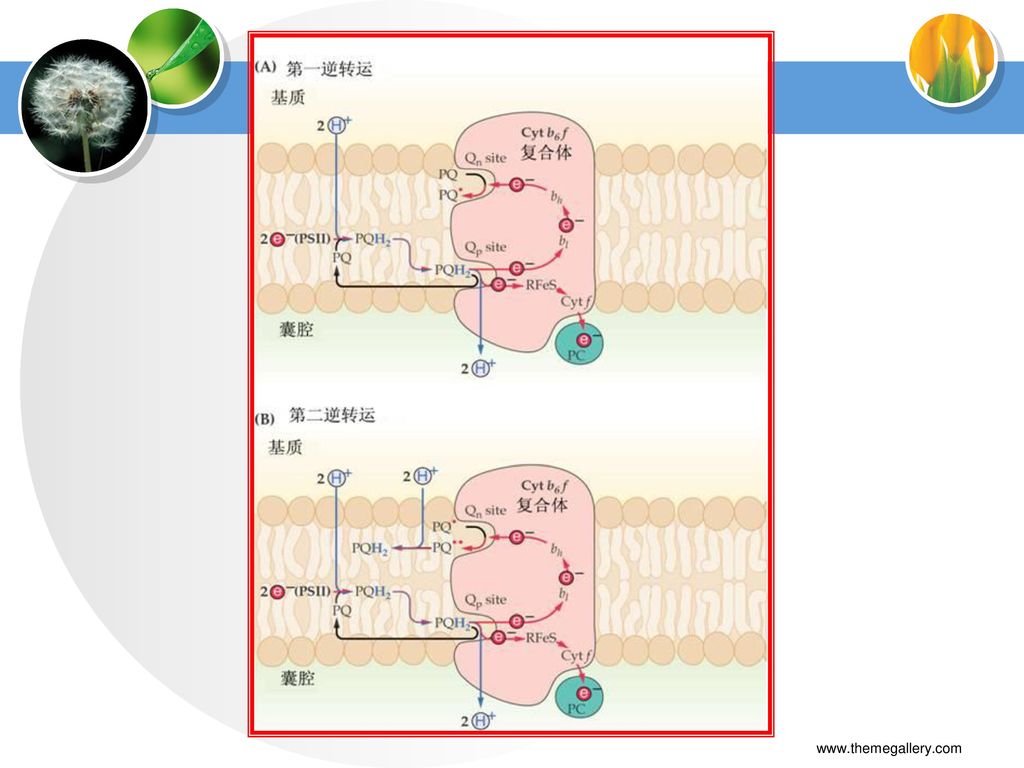
(三)Cytb6/f复合体 特点: 功能: 组成: 催化PQH2的氧化; PC的还原; 把质子从类囊体膜外间质中跨膜转移到膜内腔中
Cytf、Cytb 、Rieske (铁-硫蛋白)、2个醌氧还部分。
、2个醌氧还部分。")
38
(四)质体蓝素(PC) 特点: 位于类囊体膜内侧表面的含铜的蛋白质 功能: 将电子传给PSⅠ
质体蓝素(PC) 特点: 位于类囊体膜内侧表面的含铜的蛋白质 功能: 将电子传给PSⅠ")
39
(五) PSⅠ复合体 特点: 功能: 组成: 捕光复合体Ⅱ 反应中心色素P700 电子受体
 PSⅠ复合体 特点: 功能: 组成: 捕光复合体Ⅱ 反应中心色素P700 电子受体")
40
(六)铁氧还蛋白(Fd)和 铁氧还蛋白-NADP+还原酶(FNR) 特点: 功能: 反应式: 存在类囊体膜表面的蛋白质;
FNR 依靠核黄素的氧化还原来传递H+。 反应式: 2Fd还原+NADP++ H+ FNR Fd氧化 + NADPH

41
(七)光合电子传递方式 1. 光合链概念 在类囊体膜上的PSⅡ、PSI之间几种排列紧密的电子传递体 完成电子传递的总轨迹 2
(七)光合电子传递方式 1.光合链概念 在类囊体膜上的PSⅡ、PSI之间几种排列紧密的电子传递体 完成电子传递的总轨迹 2.光合电子传递的“Z”方案
光合电子传递方式 1.光合链概念 在类囊体膜上的PSⅡ、PSI之间几种排列紧密的电子传递体 完成电子传递的总轨迹 2.光合电子传递的 Z 方案.")
43
3. 光合电子传递的类型 (1)非环式电子传递 H2O→ PSⅡ→PQ→Cyt b6/f→PC→PSⅠ→Fd→FNR→ NADP+
4个e-,2个H2O,1个O2,2个NADP+,8个光量子,8个H+

44
(2)环式电子传递 PSⅠ→Fd→PQ→Cytb6/f→PC→PSⅠ 环式电子传递不发生H2O的氧化,也不形成NADPH,
有H+的跨膜运输,可产生ATP, 每传递一个电子需要吸收一个光量子。

45
(3)假环式电子传递 H2O→PSⅡ→PQ→Cytb6/f→PC→ PSⅠ→Fd →O2
O2 - + O H2 SOD 2H2O2 + O2

46
三、光合磷酸化 (一)概念 (二)机制 1. 光合磷酸化与电子传递的关系--偶联 2. 机理 化学渗透学说 中间产物学说 变构学说
在光合作用中由光驱动并贮存在跨类囊体膜的质子梯度的能量把ADP与Pi合成ATP的过程 (二)机制 1. 光合磷酸化与电子传递的关系--偶联 2. 机理 化学渗透学说 中间产物学说 变构学说
机制. 1. 光合磷酸化与电子传递的关系--偶联. 2. 机理. 化学渗透学说. 中间产物学说. 变构学说.")
47
ATP酶

49
(三)光合磷酸化的类型 1. 非环式光合磷酸化 与非环式电子传递偶联产生ATP的反应。
2NADP++2ADP+2Pi+2H2O 光 2NADPH+2ATP+O2 生成ATP、NADPH和氧。 含有基粒片层的放氧生物所特有,在光合磷酸化中占主要地位。

50
2. 环式光合磷酸化 与环式电子传递偶联产生ATP的反应。 ADP + Pi 光 ATP+ H2O
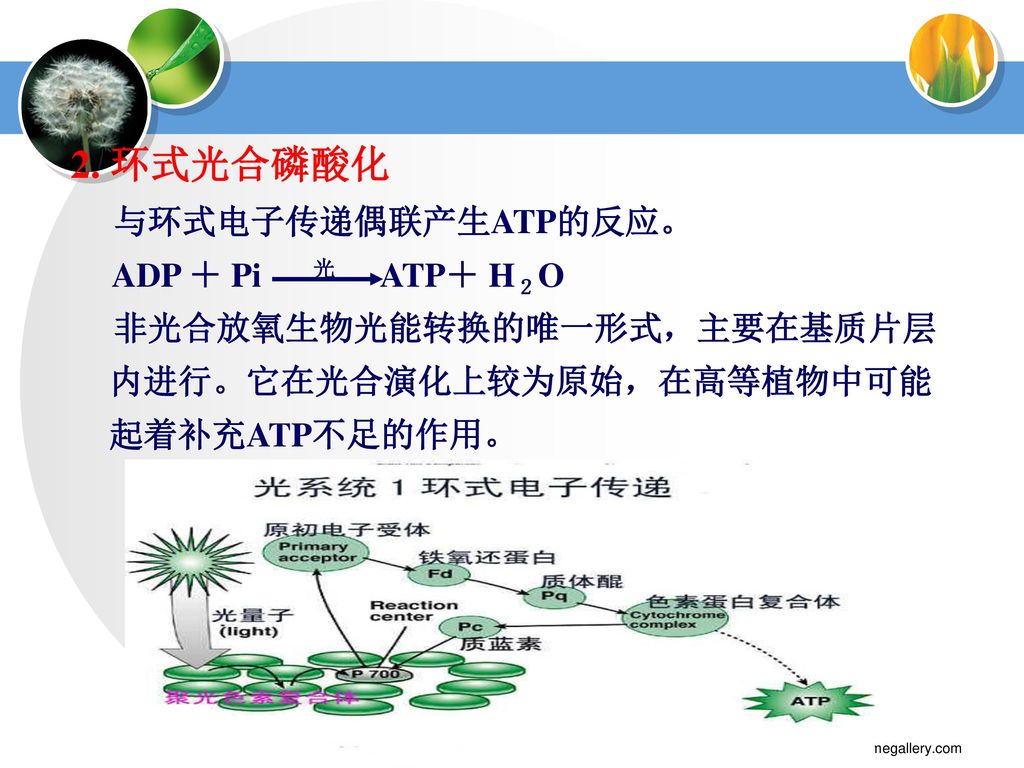
51
第五节 光合作用过程(Ⅲ):碳同化 概念: 途径:
植物利用光反应中形成的NADPH和ATP将CO2转化成稳定的碳水化合物的过程,称为CO2同化或碳同化。 途径: C3途径 C4途径 CAM(景天科酸代谢)途径。
途径。")
52
(一) C3途径的反应过程 一、卡尔文循环—C3 途径
 C3途径的反应过程 一、卡尔文循环—C3 途径")
53
(一)羧化阶段 3RuBP+3CO2+3H2O Rubisco 6PGA + 6H+
羧化阶段 3RuBP+3CO2+3H2O Rubisco 6PGA + 6H+")
54
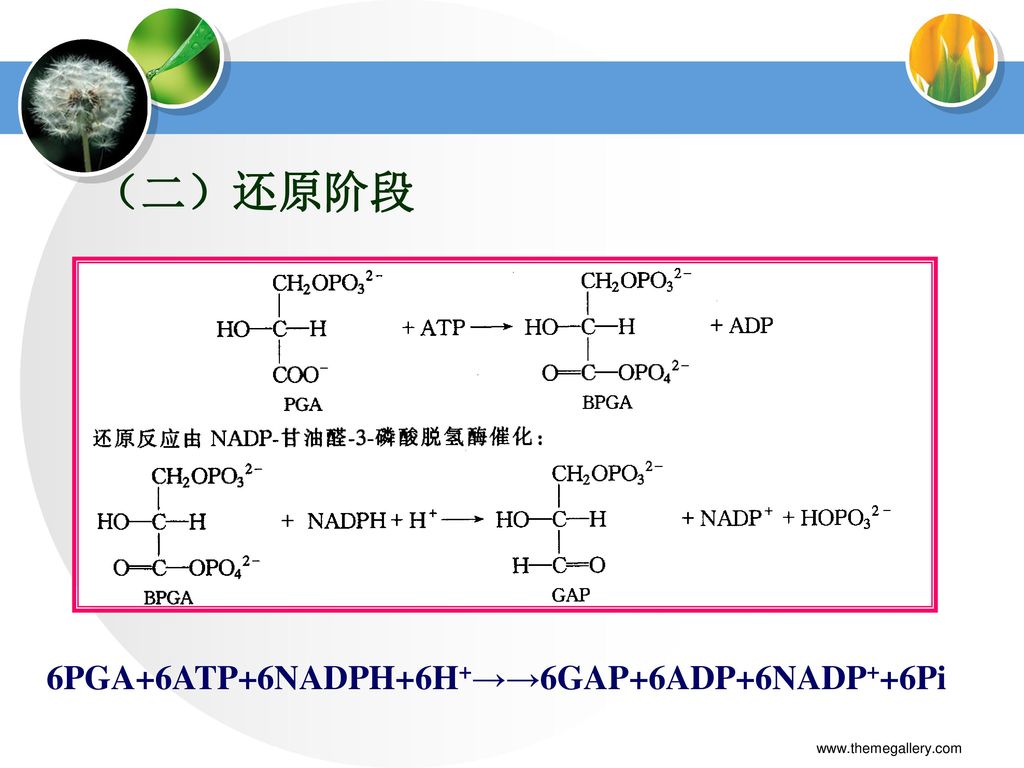
(二)还原阶段 6PGA+6ATP+6NADPH+6H+→→6GAP+6ADP+6NADP++6Pi
55
(三)再生阶段 5GAP+3ATP+2H2O→→→3RuBP+3ADP+2Pi+3H+
再生阶段 5GAP+3ATP+2H2O→→→3RuBP+3ADP+2Pi+3H+")
56
C3途径的总反应式 3CO2+5H2O+9ATP+6NADPH→GAP+9ADP+8Pi+6NADP++3H+
每同化一个CO2需消耗3个ATP和2个NADPH,可输出1个磷酸丙糖(GAP或DHAP) ; C3途径在叶绿体的基质中进行,形成的磷酸丙糖可运出叶绿体,在细胞质中合成蔗糖或参与其它反应; 形成的磷酸己糖则留在叶绿体中转化成淀粉而被临时贮藏; C3途径是光合碳代谢中最基本的循环,是所有放氧光合生物所共有的同化CO2的途径。
 ; C3途径在叶绿体的基质中进行,形成的磷酸丙糖可运出叶绿体,在细胞质中合成蔗糖或参与其它反应; 形成的磷酸己糖则留在叶绿体中转化成淀粉而被临时贮藏; C3途径是光合碳代谢中最基本的循环,是所有放氧光合生物所共有的同化CO2的途径。")
57
羧化 还原 再生

58
二、C4 途 径 (一) C4途径的发现 1954年,哈奇(M.D.Hatch)等人发现甘蔗叶片中有与C3途径不同的光合最初产物;
1965年,美国夏威夷科思谢克(H.P.Kortschak)等人报道,甘蔗叶中14C标记物首先出现于C4二羧酸; 于70年代初,澳大利亚的哈奇和斯莱克(C.R.Slack)重提出了C4-双羧酸途径,简称C4途径,也称C4光合碳同化循环,或叫Hatch-Slack途径。
等人报道,甘蔗叶中14C标记物首先出现于C4二羧酸; 于70年代初,澳大利亚的哈奇和斯莱克(C.R.Slack)重提出了C4-双羧酸途径,简称C4途径,也称C4光合碳同化循环,或叫Hatch-Slack途径。")
59
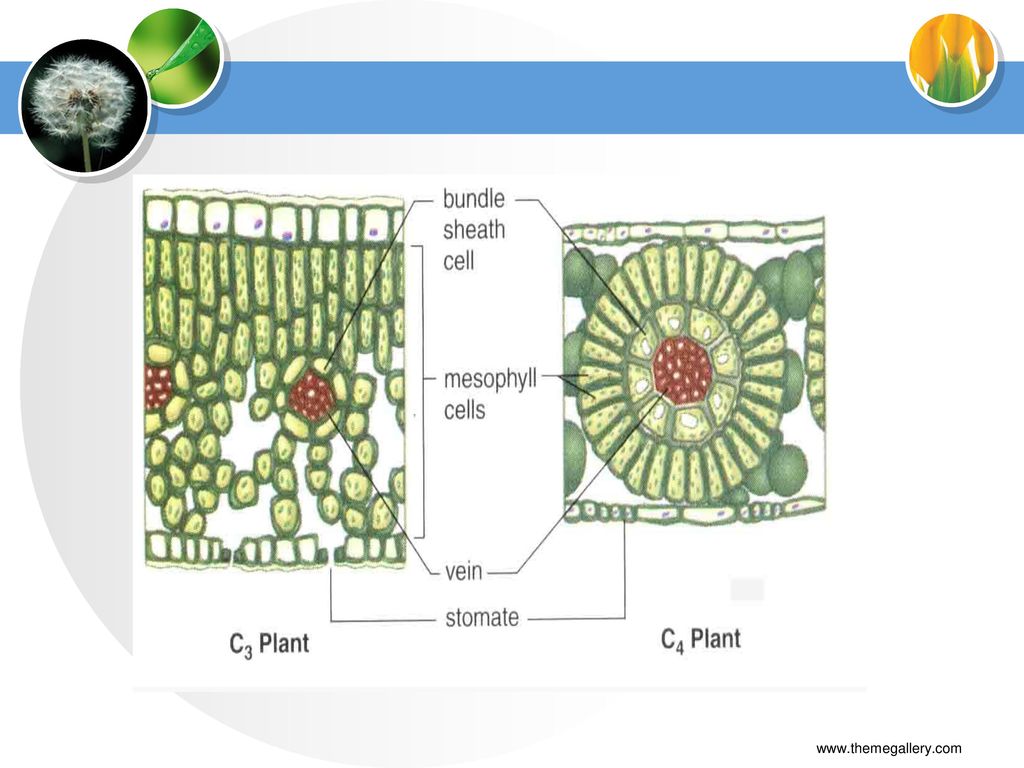
(二) C4植物结构特点 至今已知,被子植物中有20多个科约近2000种植物按C4途径固定CO2,这些植物被称为C4植物(C4 plant)。 玉 米 高 梁 苋菜 甘 蔗 粟

61
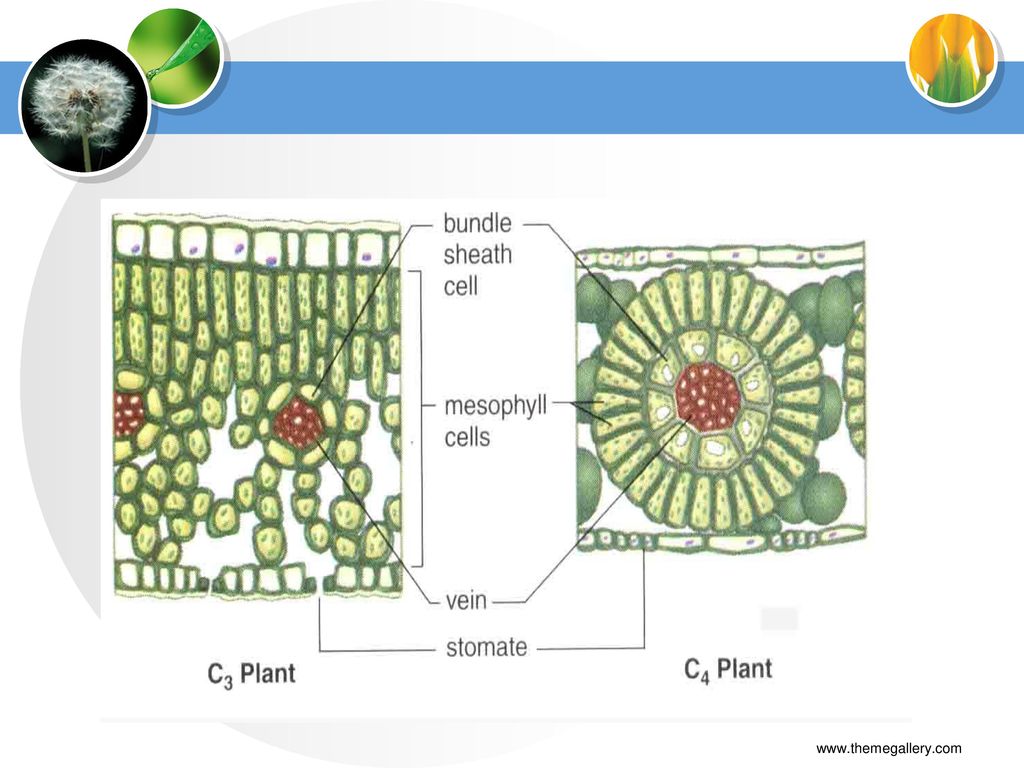
C4植物结构特点 栅栏组织与海绵组织分化不明显,叶片两侧颜色差异小; 有两类光合细胞:叶肉细胞和维管束鞘细胞(BSC);
“花环”(Kranz,德语)结构; BSC与相邻叶肉细胞间壁中纹孔多,胞间连丝丰富; 两类光合细胞中含有不同的酶类。
结构; BSC与相邻叶肉细胞间壁中纹孔多,胞间连丝丰富; 两类光合细胞中含有不同的酶类。")
62
(三)C4途径的反应过程
C4途径的反应过程")
63
1.羧化阶段

64
2.还原或转氨阶段 OAA被还原成苹果酸或经转氨作用形成天冬氨酸。

65
3.脱羧与还原 四碳双羧酸在维管束鞘中脱羧后变成丙酮酸或丙氨酸。 4.底物再生阶段

66
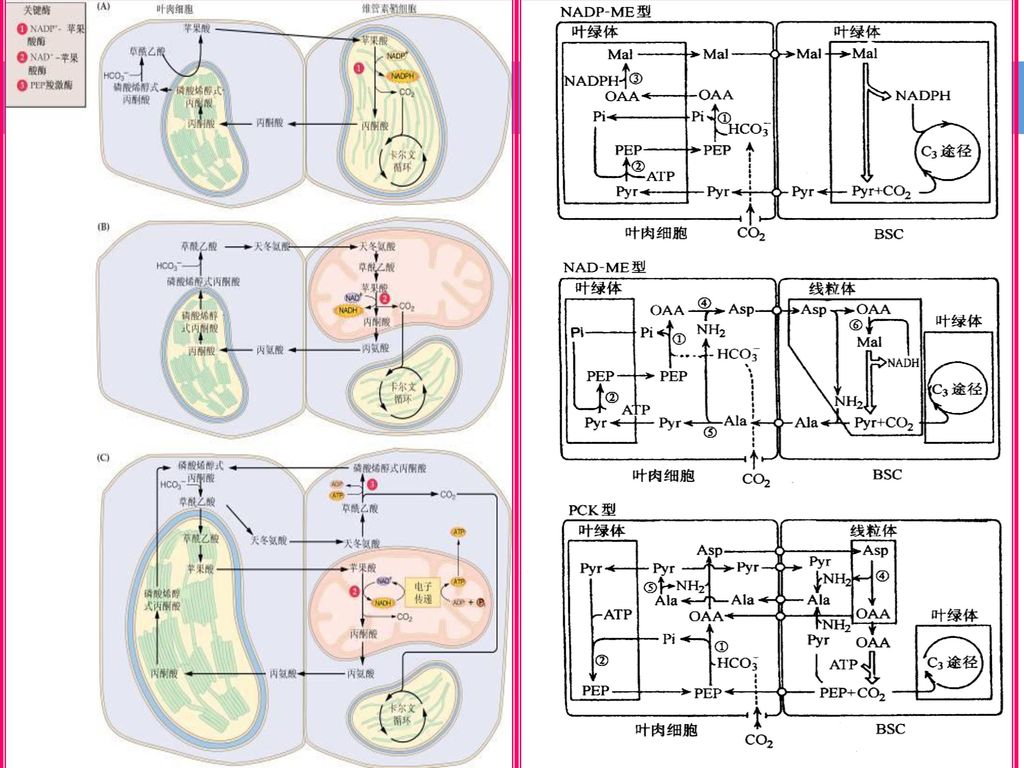
(四)C4途径的类型 根据植物所形成的初期C4二羧酸的种类以及脱羧反应参与的酶类,可把C4途径分为三种亚类型: 1
(四)C4途径的类型 根据植物所形成的初期C4二羧酸的种类以及脱羧反应参与的酶类,可把C4途径分为三种亚类型: 1. 依赖NADP的苹果酸酶的苹果酸型(NADP-ME型); 2. 依赖NAD的苹果酸酶的天冬氨酸型(NAD-ME型); 3. 具有PEP羧激酶的天冬氨酸型(PCK型)。
C4途径的类型 根据植物所形成的初期C4二羧酸的种类以及脱羧反应参与的酶类,可把C4途径分为三种亚类型: 1. 依赖NADP的苹果酸酶的苹果酸型(NADP-ME型); 2. 依赖NAD的苹果酸酶的天冬氨酸型(NAD-ME型); 3. 具有PEP羧激酶的天冬氨酸型(PCK型)。")
68
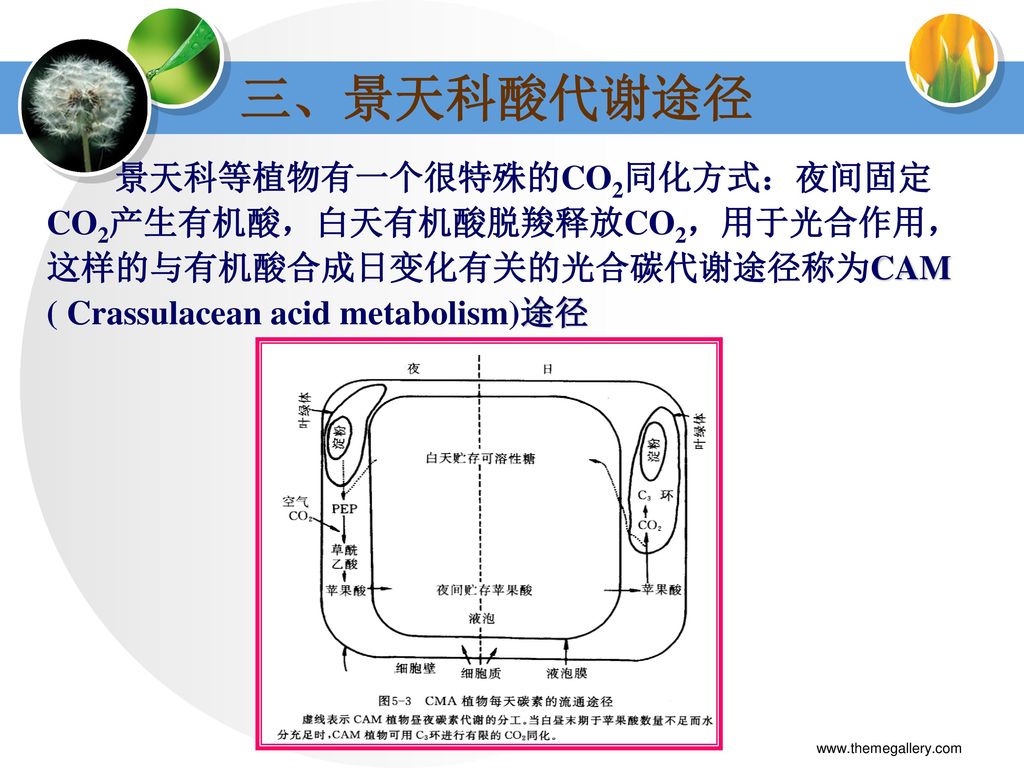
三、景天科酸代谢途径 景天科等植物有一个很特殊的CO2同化方式:夜间固定CO2产生有机酸,白天有机酸脱羧释放CO2,用于光合作用,这样的与有机酸合成日变化有关的光合碳代谢途径称为CAM ( Crassulacean acid metabolism)途径
69
CAM最早是在景天科植物中发现的,主要分布在景天科、仙人掌科、兰科、凤梨科、大戟科、番杏科、百合科、石蒜科等植物中;

70
剑麻 龙舌兰 落地生根 芦荟 宝绿 绯牡丹 昙花

71
鸡冠掌 静夜 红司 锦晃星

72
CAM植物-瓦松属 瓦松属1

73
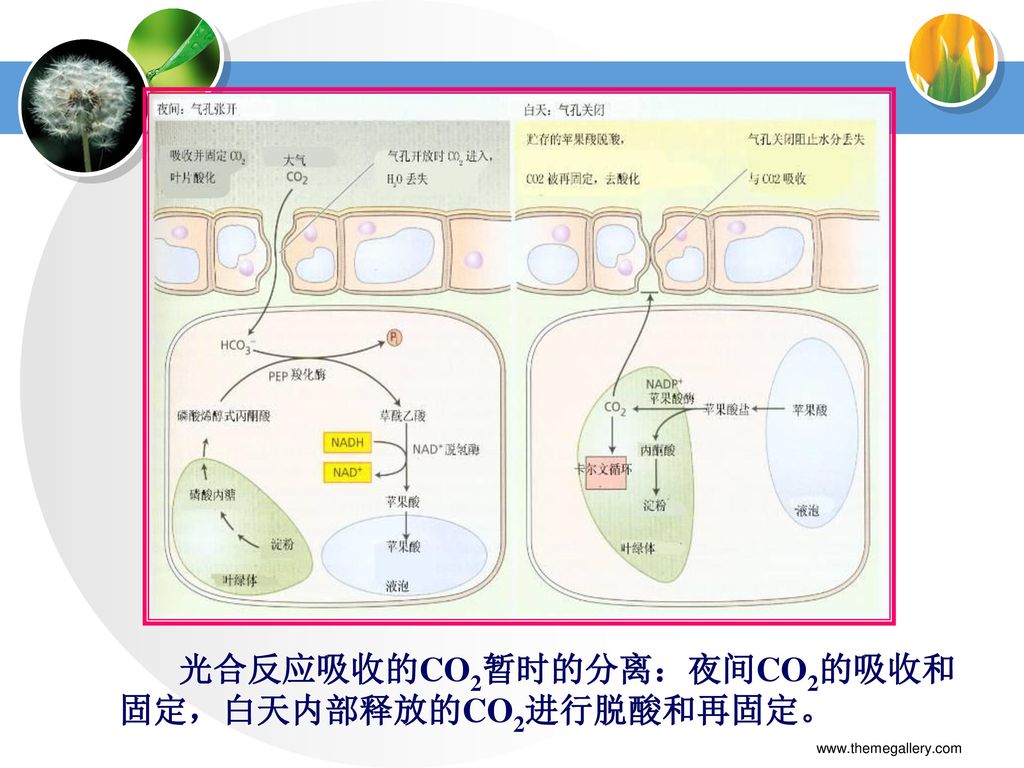
光合反应吸收的CO2暂时的分离:夜间CO2的吸收和固定,白天内部释放的CO2进行脱酸和再固定。
74
12.51 景天酸科代谢途径(CAM)是光合作用适应干旱环境的一种进化形式。CAM植物开放的气孔使进入。PEP羧化酶(1)参与CO2形成四碳有机酸草酰乙酸,草酰乙酸可以被苹果酸脱氢酶(2)还原成苹果酸。夜间在液泡中。在光下CAM植物关闭气孔以阻止水分流失。贮存的苹果酸被NADP+-苹果酸酶(3)脱羧,释放CO2通过卡文循环转变碳氢化合物。

75
第六节 光 呼 吸 (一)光呼吸的发现 1920年瓦伯格在用小球藻做实验时发现,O2对光合作用有抑制作用,这种现象被称为瓦伯格效应(Warburg effect)。这实际上是氧促进光呼吸的缘故。

76
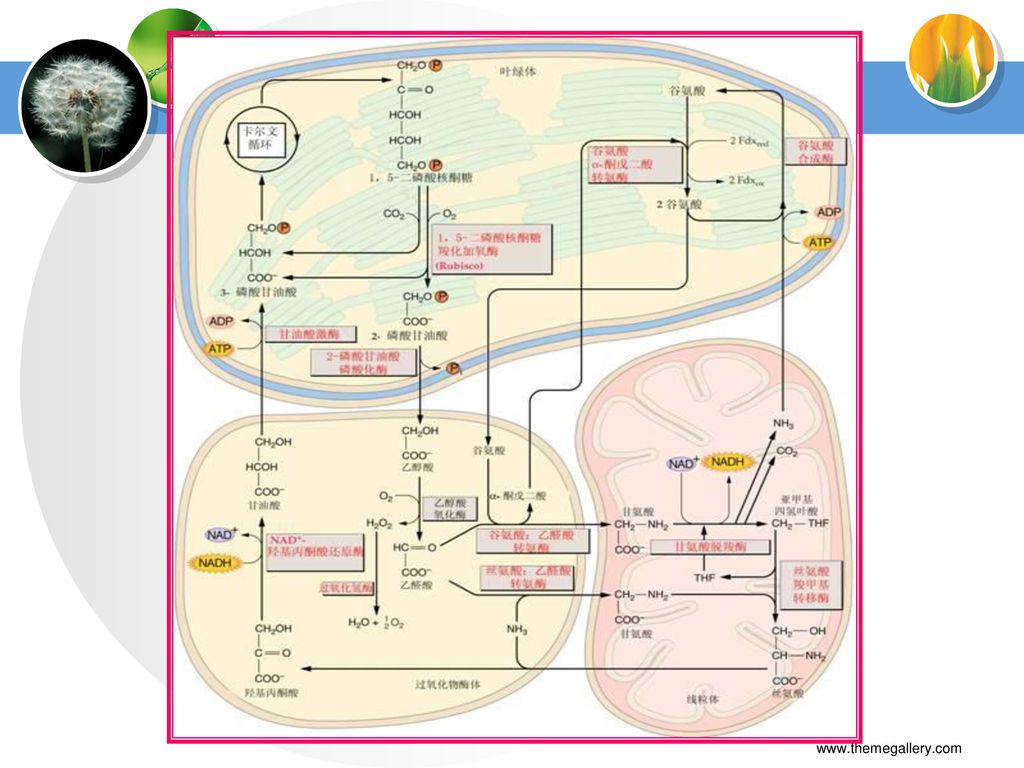
(二)光呼吸的概念 植物的绿色细胞在光照下有吸收氧气,释放CO2的反应。因为光呼吸底物乙醇酸和其氧化产物乙醛酸,以及后者经转氨作用形成的甘氨酸皆为C2化合物,因此光呼吸途径又称为C2光呼吸碳氧化循环,简称C2循环。 (三)光呼吸的生化途径 乙醇酸代谢要经过三种细胞器: 叶绿体 过氧化体 线粒体

78
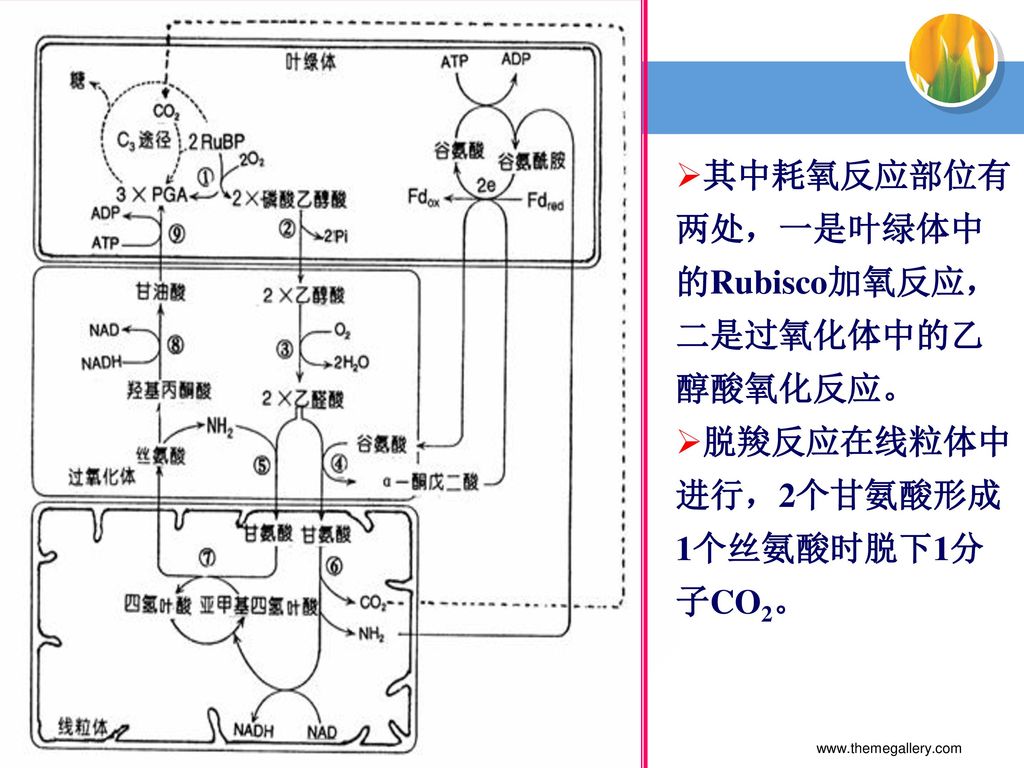
其中耗氧反应部位有两处,一是叶绿体中的Rubisco加氧反应,二是过氧化体中的乙醇酸氧化反应。
79
光呼吸的总反应方程式 RuBP+15O2+11H2O+34ATP+15NADPH+10Fdred
CO2+34ADP+36Pi+15NADP++10Fdox+9H+

80
(四)光合作用和光呼吸的联系 均由Rubisco开始; 光合作用提供ATP和NADPH,光呼吸消耗 ATP和FdxRED;
C3循环中底物CO2是C2循环的产物,C2循环的底物O2是C3循环的产物。

81
(五)光呼吸生理意义: 1.回收碳素 通过C2碳氧化环可回收乙醇酸中3/4的碳;
2.维持C3光合碳还原循环的运转 在叶片气孔关闭或外界CO2浓度低时,光呼吸释放的CO2能被C3途径再利用,以维持光合碳还原循环的运转。 3.防止强光对光合机构的破坏作用 光呼吸却可消耗同化力与高能电子,降低O2-·的形成,从而保护叶绿体,免除或减少强光对光合机构的破坏。 4.消除乙醇酸对细胞有毒害 光呼吸则能消除乙醇酸,使细胞免遭毒害。

82
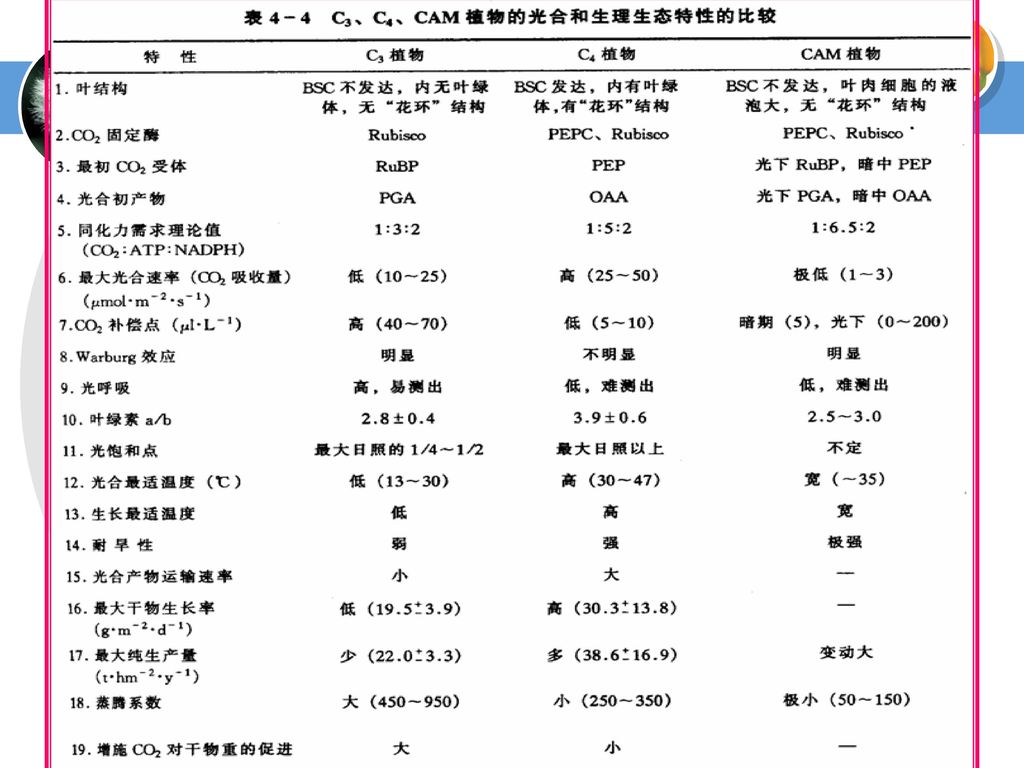
三、C3、C4与CAM植物的光合特性比较 (一)C4植物结构特点 栅栏组织与海绵组织分化不明显,叶片两侧颜色差异小;
有两类光合细胞:叶肉细胞和维管束鞘细胞(BSC); “花环”(Kranz,德语)结构; BSC与相邻叶肉细胞间壁中纹孔多,胞间连丝丰富; 两类光合细胞中含有不同的酶类。
; 花环 (Kranz,德语)结构; BSC与相邻叶肉细胞间壁中纹孔多,胞间连丝丰富; 两类光合细胞中含有不同的酶类。")
84
(二)光合特性 在高温、强光、干旱和低CO2条件下,C4植物显示出高光合效率。 C4植物具较高光合速率的因素有:
1.C4植物的叶肉细胞中的PEPC对底物HCO-3的亲和力极高,细胞中的HCO3-浓度一般不成为PEPC固定CO2的限制因素; 2.C4植物由于有“CO2泵”浓缩CO2的机制,使得BSC中有高浓度的CO2,从而促进Rubisco的羧化反应,降低了光呼吸,且光呼吸释放的CO2又易被再固定; 3.鞘细胞中的光合产物可就近运入维管束,从而避免了光合产物累积对光合作用可能产生的抑制作用。

85
4. 高光强又可推动电子传递与光合磷酸化,产生更多的同化力,以满足C4植物PCA循环对ATP的额外需求;
但是C4植物同化CO2消耗的能量比C3植物多,也可以说这个“CO2泵”是要由ATP来开动的,故在光强及温度较低的情况下,其光合效率还低于C3植物。可见C4途径是植物光合碳同化对热带环境的一种适应方式。

87
第六节 影响光合作用的因素 一、光合速率 (一)概念 1.光合速率 单位时间单位叶面积吸收CO2量或放出O2量,或积累干物质量。
2.表观光合速率或净光合速率 通常测定光合速率时没有把呼吸作用(光、暗呼吸)以及呼吸释放的CO2被光合作用再固定等因素考虑在内,所测结果; 3.总光合速率或真正光合速率 表观光合速率加上光、暗呼吸速率
以及呼吸释放的CO2被光合作用再固定等因素考虑在内,所测结果; 3.总光合速率或真正光合速率. 表观光合速率加上光、暗呼吸速率.")
88
(二)测定方法 CO2+ H2O 光 叶绿体 (CH2O) + O2 CO2吸收量 干物质积累量 O2释放量
μmol CO2·m-2·s mgDW·dm-2·h-1 μmolO2·m-2·s-1 1μmol·m-2·s-1=1.58mg·dm-2·h-1

89
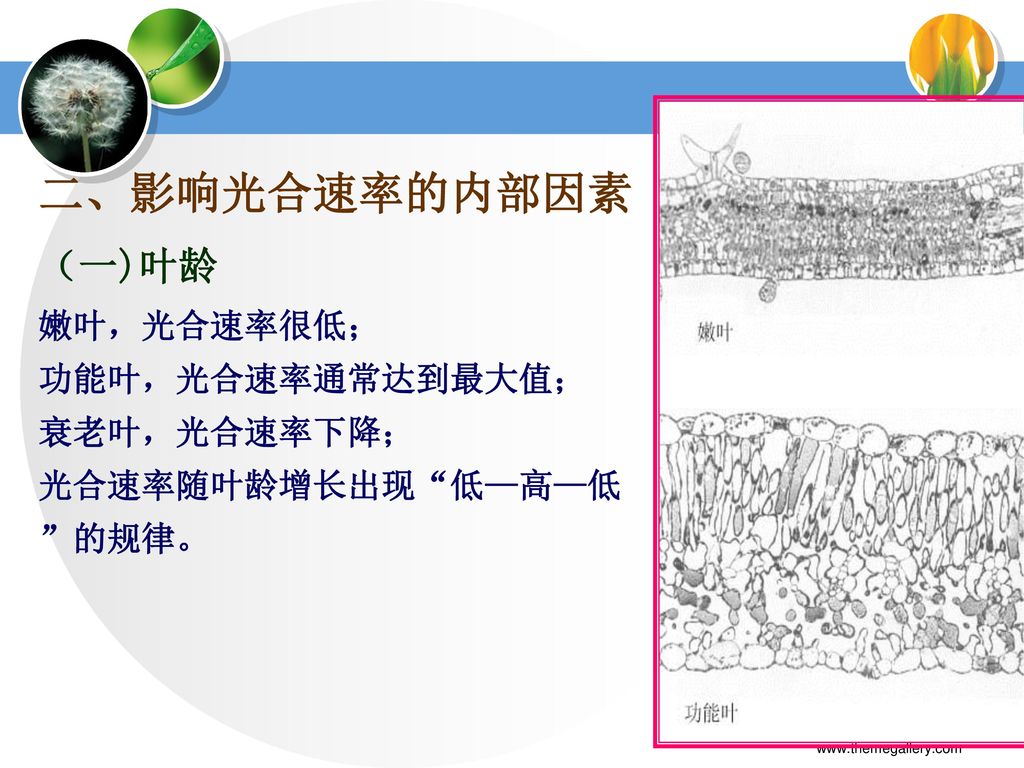
二、影响光合速率的内部因素 (一)叶龄 嫩叶,光合速率很低; 功能叶,光合速率通常达到最大值; 衰老叶,光合速率下降; 光合速率随叶龄增长出现“低—高—低”的规律。
90
(二)叶的结构 (三)植物不同生育期 营养生长期最强 C4植物的叶片光合速率通常要大于C3植物,这与C4植物叶片具有花环结构等特性有关。
(三)植物不同生育期 营养生长期最强
植物不同生育期 营养生长期最强.")
91
(四)光合产物的输出 光合产物(蔗糖)从叶片中输出的速率会影响叶片的光合速率。 光合产物积累到一定的水平后会影响光合速率的原因:
(1)反馈抑制。 (2)淀粉粒的影响。 例如,摘去花、果、顶芽等,邻近叶的光合速率? 摘除其他叶片,只留一张叶片与所有花果,留下叶的光合速率? 对苹果等枝条环割,使环割上方枝条上的叶片光合速率?
反馈抑制。 (2)淀粉粒的影响。 例如,摘去花、果、顶芽等,邻近叶的光合速率 摘除其他叶片,只留一张叶片与所有花果,留下叶的光合速率 对苹果等枝条环割,使环割上方枝条上的叶片光合速率")
92
三、影响光合速率的外部因素 (一)光照 直接制约着光合速率的高低 光照因素中有光强、光质与光照时间
光照 直接制约着光合速率的高低 光照因素中有光强、光质与光照时间")
93
在低光强区,光合速率随光强的增强而呈比例地增加(直线A); 当超过一定光强,光合速率增加就会转慢(曲线B);
1. 光强 在低光强区,光合速率随光强的增强而呈比例地增加(直线A); 当超过一定光强,光合速率增加就会转慢(曲线B); 当达到某一光强,光合速率不再增加,呈光饱和现象 (直线C)。 光补偿点:当到达某一光强时,叶片的光合速率等于呼吸速率,表观光合速率为零时的光强。 光饱和点:开始达到光合速率最大值时的光强。 C4植物>C3植物
; 当超过一定光强,光合速率增加就会转慢(曲线B); 当达到某一光强,光合速率不再增加,呈光饱和现象 (直线C)。 光补偿点:当到达某一光强时,叶片的光合速率等于呼吸速率,表观光合速率为零时的光强。 光饱和点:开始达到光合速率最大值时的光强。 C4植物>C3植物.")
94
不同植物的光补偿点和光饱和点也有很大的差异: 光补偿点高的植物一般光饱和点也高, 草本植物的光补偿点与光饱和点通常要高于木本植物;
阳生植物的光补偿点与光饱和点要高于阴生植物; C4植物的光饱和点要高于C3植物。

95
图 不同光波下光合速率 实线为26种草本植物的平均值;
虚线为7种木本植物的平均值。 2.光质 在太阳幅射中,只有可见光部分才能被光合作用利用。 用不同波长的可见光照射植物叶片,测定到的光合速率不一样。 光合作用的作用光谱与叶绿体色素的吸收光谱大体吻合。在600~680nm红光区,光合速率有一大的峰值,在435nm左右的蓝光区又有一小的峰值。

96
大树底下无丰草? 树木的叶片吸收红光和蓝光较多,故透过树冠的光线中绿光较多,由于绿光是光合作用的低效光,因而会使树冠下生长的本来就光照不足的植物利用光能的效率更低,“大树底下无丰草”就是这个道理。

97
图 30 叶片光合速率对细胞间隙CO2浓度响应示意图
CO2补偿点(图中C点); CO2饱和点(图中S点) 。 图 30 叶片光合速率对细胞间隙CO2浓度响应示意图 曲线上四个点对应浓度分别为CO2补偿点(C),空气浓度下细胞间隙的CO2浓度(n),与空气浓度相同的细胞间隙CO2浓度(350μl·L-1左右)和CO2饱和点(S)。Pm为最大光合速率;CE为比例阶段曲线斜率,代表羧化效率;OA光下叶片向无CO2气体中的释放速率,可代表光呼吸速率。
; CO2饱和点(图中S点) 。 图 30 叶片光合速率对细胞间隙CO2浓度响应示意图. 曲线上四个点对应浓度分别为CO2补偿点(C),空气浓度下细胞间隙的CO2浓度(n),与空气浓度相同的细胞间隙CO2浓度(350μl·L-1左右)和CO2饱和点(S)。Pm为最大光合速率;CE为比例阶段曲线斜率,代表羧化效率;OA光下叶片向无CO2气体中的释放速率,可代表光呼吸速率。")
98
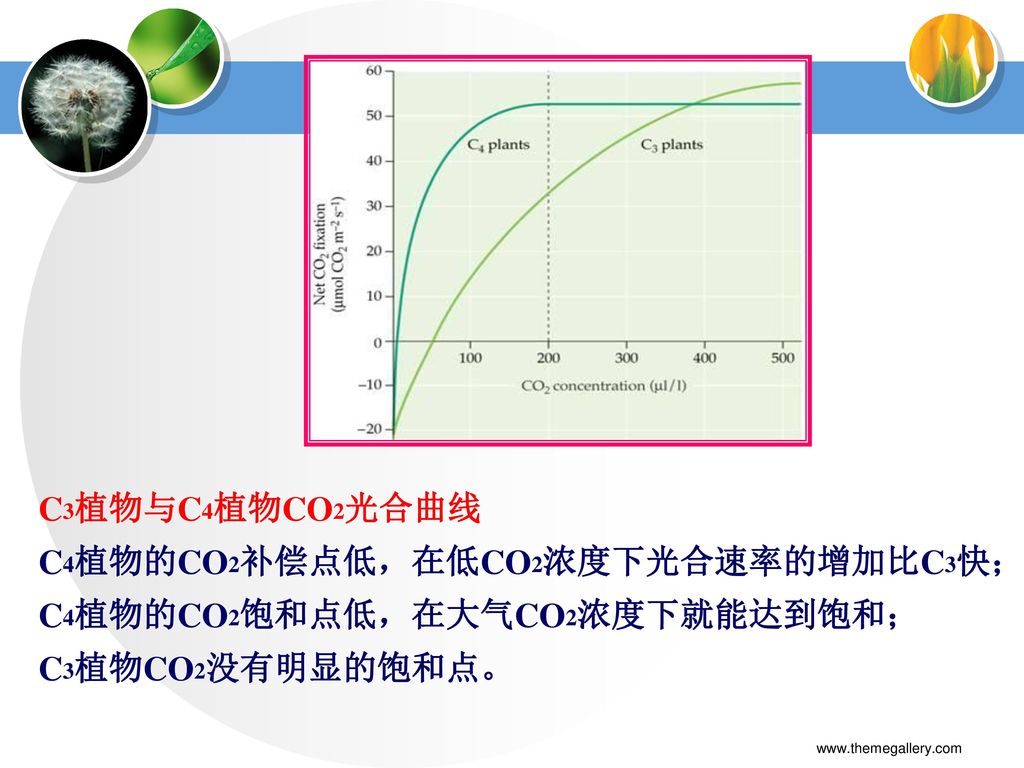
C4植物的CO2补偿点低,在低CO2浓度下光合速率的增加比C3快; C4植物的CO2饱和点低,在大气CO2浓度下就能达到饱和;
99
在不通风的温室、大棚和光合作用旺盛的作物冠层内的CO2浓度可降至200μl·L-1左右。由于光合作用对CO2的消耗以及存在CO2扩散阻力,因而叶绿体基质中的CO2浓度很低,接近CO2补偿点。因此,加强通风或设法增施CO2能显著提高作物的光合速率,这对C3植物尤为明显。

100
(三)温度
温度")
101
(三)温度 光合作用的温度三基点: 最低温度(冷限):该低温下表观光合速率为零(0℃) ) 最高温度(热限):该高温下表观光合速率为零(45℃) 最适温度:能使光合速率达到最高的温度

103
(四)水分 水分对光合作用的影响以间接影响较大 干旱,气孔关闭; 缺水,淀粉分解,糖积累。
水分 水分对光合作用的影响以间接影响较大 干旱,气孔关闭; 缺水,淀粉分解,糖积累。")
104
(五)矿质营养 矿质营养在光合作用中的功能:
1.叶绿体结构的组成成分 如N、P、S、Mg是叶绿体中构成叶绿素、蛋白质、核酸以及片层膜不可缺少的成分。 2.电子传递体的重要成分 如PC中含Cu,Fe-S中心、Cytb、Cytf和Fd中都含Fe,放氧复合体不可缺少Mn2+和Cl-。 3.磷酸基团的重要作用 构成同化力的ATP和NADPH,光合碳还原循环中所有的中间产物,合成淀粉的前体ADPG,以及合成蔗糖的前体UDPG,这些化合物中都含有磷酸基团。 4.活化或调节因子 如Rubisco,FBPase等酶的活化需要Mg2+;Fe、Cu、Mn、Zn参与叶绿素的合成;K+和Ca2+调节气孔开闭;K和P促进光合产物的转化与运输等。

105
(六)光合速率的日变化 随光强日变化呈单峰曲线 如果白天云量变化不定,则光合速率会随光强的变化而变化。
当光照强烈、气温过高时,光合速率日变化呈双峰曲线,大峰在上午,小峰在下午,中午前后,光合速率下降,呈现“午睡”现象,且这种现象随土壤含水量的降低而加剧(图35)。
。")
106
第八节 植物对光能的利用 —、植物的光能利用率概念 指植物光合作用所累积的有机物所含的能量,占照射在单位地面上的日光能量的比率。
第八节 植物对光能的利用 —、植物的光能利用率概念 指植物光合作用所累积的有机物所含的能量,占照射在单位地面上的日光能量的比率。

107
二、提高光能利用率的途径 (一)延长光合时间 (二)增加光合面积 (三)提高光合速率 1.提高复种指数 2.补充人工光照 1.合理密植
2.改变株型 (三)提高光合速率 1.增加CO2浓度 2.降低光呼吸
提高光合速率. 1.增加CO2浓度. 2.降低光呼吸.")
108
谢谢 教师:李侠 学院:农学院 《植物物生理学》



















.>")




 2、血糖平衡的调节(B)>")