Download presentation
1
第六讲 基因组进化 的分子基础

2
基因组进化的分子基础: 突 变 重 组 转 座

3
♦ 自发性损伤(复制中的损伤、碱基的自发性化学改变、 自发脱碱基、 细胞的代谢产物对DNA的损伤)
突变 1.1 突变的机制 ♦ 自发性损伤(复制中的损伤、碱基的自发性化学改变、 自发脱碱基、 细胞的代谢产物对DNA的损伤) ♦ 物理因素引起的损伤(电离辐射、紫外线、热诱变等) ♦ 化学因素引起的损伤(烷化剂、碱基类似物、嵌入试剂等) 碱基的自发性化学改变----互变异构、自发脱氨基 细胞的代谢产物对DNA的损伤---O2-和H2O2等产生的氧自由基 电离辐射---直接引起碱基、戊糖等导致链的断裂、交联 氧化损伤、烷基化损伤、大的加合物损伤 Transition : Purine or pyrimidine is replaced by the other AG T C Transversion: a purine is replaced by a pyrimidine or vice verse A T or C T A or G G T or C C A or G
 ♦ 物理因素引起的损伤(电离辐射、紫外线、热诱变等) ♦ 化学因素引起的损伤(烷化剂、碱基类似物、嵌入试剂等) 碱基的自发性化学改变----互变异构、自发脱氨基 细胞的代谢产物对DNA的损伤---O2-和H2O2等产生的氧自由基. 电离辐射---直接引起碱基、戊糖等导致链的断裂、交联. 氧化损伤、烷基化损伤、大的加合物损伤. Transition : Purine or pyrimidine is replaced by the other AG T C. Transversion: a purine is replaced by a pyrimidine or vice verse A T or C T A or G. G T or C C A or G.")
4
1.1.1 自发性损伤 错配突变 纯化学的碱基配对差错率为:5%~10% 为维持基因组的稳定性,DNA 的复制必须增加几个数量级,提高DNA复制的精确性有2种方法: ♪ 渗入碱基的筛选 ♪ 错配碱基的校正

5
误导渗入 碱基异构式引起DNA复制过程的错误 -----自发突变 碱基异构式: A(amino 氨基) A(imino 亚氨基)
C(a) C(i) G(keto 酮式) G(enol 烯醇式) G(k) G(e,i) T(keto) T(enol-2’) or T(enol-4’)
 C(i) G(keto 酮式) G(enol 烯醇式) G(k) G(e,i) T(keto) T(enol-2’) or T(enol-4’)")
6
碱基异构式引起DNA复制的错配 G(k) C(a) 正确配对 A(a) T(k) 错误配对 G(k) T(e) A(a) C(i)
A(i, anti) A(a, syn) A(i, anti) G(k, syn) G(e,i, anti) G(k, syn) G(e,i, anti) A(a, syn) A(i) C(a) G(e) T(k)
 A(a, syn) A(i, anti) G(k, syn) G(e,i, anti) G(k, syn) G(e,i, anti) A(a, syn) A(i) C(a) G(e) T(k)")
7
碱基异构式引起DNA的错配突变 A(a) G(k) A(a) T(k) C(i) C(i) C(a) A(i, anti)
A(a, anti) T(k, anti) C(a, anti) G(k, syn) G(k, syn) G(k, anti)
 T(k, anti) C(a, anti) G(k, syn) G(k, syn) G(k, anti)")
8
滑序复制

9
1.1.2 化学因素引起的损伤 碱基类似物(Base analog) 5-溴尿嘧啶(BrU) 2-氨基嘌呤(AP或2-AP)
5-Bromine Uracil 2-氨基嘌呤(AP或2-AP) 2-Amino purine O Br NH2
 2-Amino purine. O. Br. NH2.")
10
: G : A 酮式5-BrU的渗入 烯醇式enol 第一轮复制 酮式到稀醇式的转变 第二轮复制 酮式Keto A·TG·C 转变
AGCTTCCTA TCGAAGGAT AGCTBCCTA 酮式5-BrU的渗入 烯醇式enol Br OH H O : G AGCTBCCTA TCGAGGGAT AGCTTCCTA TCGAAGGAT 第一轮复制 酮式到稀醇式的转变 Br 酮式Keto H O AGCTBCCTA TCGAAGGAT AGCTCCCTA TCGAGGGAT 第二轮复制 : A A·TG·C 转变 5-BrU 烯醇式渗入为 G·CA·T 转变

11
碱基的化学修饰导致突变 又称化学突变剂: 亚硝酸(nitrous acid HNO2) 羟氨(hydroxylamine HA)
甲磺酸乙酯(ethyl mathanesulfonate EMS) N-甲基-N’-硝基-N-亚硝基胍 (N-mathyl-N’-nitro-N-nitrosoguanidion NNG)
 N-甲基-N’-硝基-N-亚硝基胍. (N-mathyl-N’-nitro-N-nitrosoguanidion NNG)")
12
NH2OH (Hydroxylamine HA 羟胺)
HNH H O C(a) HA HNH H O N HO HN H O N HO C(i) A(a)
 HA. HNH. H. O. N. HO. HN. H. O. N. HO. C(i) A(a)")
13
嵌合剂的质突变作用 吖啶橙 (Acridine Orange AO) 扁平染料分子 溴化乙锭 (Ethidium Bromide EB )
分子插入 -A TTTCG - -T AAAGC- TAO T -ATTTCG - -TAAAGC- AO EB -ATTTTTCG - -TAAAAAGC- 吖啶橙插入导致碱基的缺失突变 结合前面错义、移框的抑制突变-----因此遗传学上鉴定突变型(突变类型)可用分别使用产生错义、移框突变的突变剂二次处理,看其回复突变的有无 -AT EB TTTTCG- -TA -ATX’TTTTCG- -TAX AAAAGC- AAAAGC- X 结果产生---移框突变
可用分别使用产生错义、移框突变的突变剂二次处理,看其回复突变的有无. -AT EB TTTTCG- -TA. -ATX’TTTTCG- -TAX AAAAGC- AAAAGC- X. 结果产生---移框突变.")
14
物理因素引起的损伤 ♪ 紫外线的致突变作用 ---嘧啶二聚体 (TT dimer ) ∧ U.V. …C T T A…
 ∧ U.V. …C T T A…")
15
脱氨氧化 U.V. C U A(a) H2O H+ + OH- C(a) C(i) A(a) 脱嘌呤 造成的突变:碱基替代、缺失、重复、移框

16
同义突变:没有改变产物氨基酸序列的密码子 错义突变:碱基序列的改变引起了氨基酸序列的改变 (中性突变、渗漏突变)
1.2 突变的效应 ◙ 突变对基因组的影响 同义突变:没有改变产物氨基酸序列的密码子 错义突变:碱基序列的改变引起了氨基酸序列的改变 (中性突变、渗漏突变) 无义突变:碱基的改变使代表某种氨基酸的密码子变 为蛋白合成的终止密码子 连读突变:与终止突变正好相反,终止密码子变成指 令某一氨基酸的密码子,使翻译继续进行 GAA(E) → GAG(E) Glu E---代表Glu 有些错义突变严重影响到蛋白质活性甚至完全无活性,若该基因为必需的-----致死突变 渗漏突变---表型介于野生型与完全突变型之间 中性突变---突变基本不影响蛋白质活性,不表现明显的性状变化
 无义突变:碱基的改变使代表某种氨基酸的密码子变 为蛋白合成的终止密码子. 连读突变:与终止突变正好相反,终止密码子变成指. 令某一氨基酸的密码子,使翻译继续进行. GAA(E) → GAG(E) Glu E---代表Glu. 有些错义突变严重影响到蛋白质活性甚至完全无活性,若该基因为必需的-----致死突变. 渗漏突变---表型介于野生型与完全突变型之间. 中性突变---突变基本不影响蛋白质活性,不表现明显的性状变化.")
18
◙ 突变对多细胞生物的影响 ♫ 多细胞生物的细胞有2种类型: 体细胞 --不参与时代间的遗传事件 种质细胞--负责将遗传物质传递给下一代
♫ 多细胞突变产生的影响: 体细胞-- 仅限其本身、不会影响后代及进化; 即使死亡,也有同类型的体细胞存在; 引起细胞无限增值的突变,使细胞分裂失去控制,产生肿瘤; 种质细胞--当代不会对个体表型产生重要影响,局限于很小的器官; 可以传递给下一代,使子代个体所有细胞都含有从亲代继承的突变;

19
显性导致遗传病,如Marfan综合症,产生异常的结缔组织蛋白原纤维蛋白;
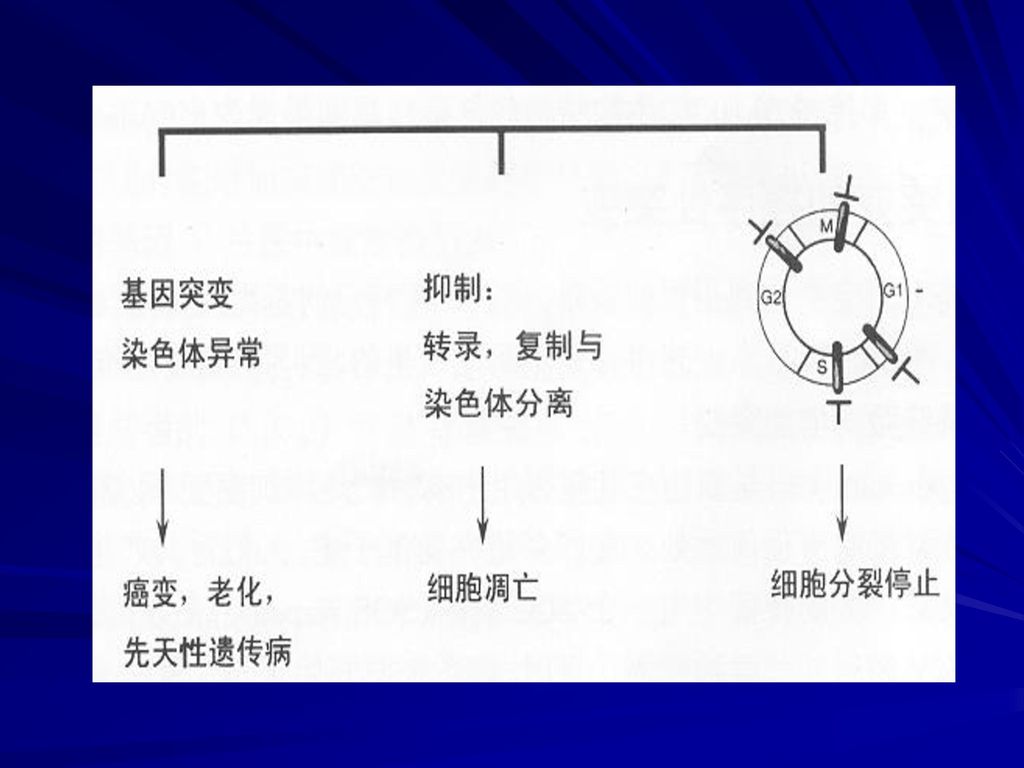
♫ 多细胞生物的突变效应分为2类: 功能丧失:使蛋白质的活性降低或丧失; 显性、隐姓; 显性导致遗传病,如Marfan综合症,产生异常的结缔组织蛋白原纤维蛋白; 功能增益:突变提供一种异常蛋白质活性; 一般为显性; 多发生在调控区,如使1个或多个基因在错误的组织中表达,导致细胞功能紊乱,或控制细胞周期的一 个或几个基因的过量表达,使细胞分裂失控引发癌症;

21
◙ 突变率与生物的复杂性 生物进化的基本动力:突变 如果突变率太高--基因组处于不稳定状态,不利于进化;
现存生物,包括低等生物和高等生物基因组的自发突变率约为10-9----这是各种因素综合作用的结果; 每个基因都有积累突变的风险,而大多数突变都是有害的,因此生物含有的基因数越多,发生突变的几率越大,由此判断平均突变率为生物的复杂性设定了一个上限; 群体遗传学家估计,根据DNA复制的忠实性,哺乳动物含有基因数不超过60 000。

22
1.3 超突变和程序性突变 SOS修复系统倾向增加突变 SOS repair 是一种错误倾向性极强的修复机制
是进化中形成的“ 丧失某些信息而存活总比死亡好一些” 的措施 (正常状态下,SOS是关闭的)
")
23
SOS系统诱导,错误潜伏的复制超越二聚体而进行
SOS 修复--无模板指导的DNA复制 大剂量的紫外线照射,大量的二聚体产生 SOS系统诱导,错误潜伏的复制超越二聚体而进行 SOS修复酶只有在细胞受到损伤时才存在(正常细胞中不存在), 机制--a、SOS系统以某种方式对polⅢ进行修饰(改变校对亚基功能)b、由polⅡ负责超越创伤 复制结果大量的没有被错配修复系统和切除修复系统纠正的错误碱基导致突变--错误倾向性----存活下来总比死亡好 错误碱基
, 机制--a、SOS系统以某种方式对polⅢ进行修饰(改变校对亚基功能)b、由polⅡ负责超越创伤. 复制结果大量的没有被错配修复系统和切除修复系统纠正的错误碱基导致突变--错误倾向性----存活下来总比死亡好. 错误碱基.")
24
SOS 修复只是SOS反应的一部分 RecA在SOS反应 反应中起核心作用 RecA与LexA组成调控环路 DNA 损伤
细菌在UV、丝裂霉素C、烷化剂等作用下造成DNA的损伤,抑制DNA的复制----此时细胞产生一系列的表型变化---对损伤DNA修复能力迅速增强、诱变率的提高、细胞分裂停止以及λ原噬菌体的释放-----统称为SOS反应 LexA protein represses many genes, including repair functions, recA and lexA,. Activation of RecA leads to proteolytic cleavage of LexA and induces all of these genes. UV引起DNA损伤的Signal--- (S.S. DNA tail or 5Nt S.S.DNA) SOS
 SOS.")
25
RecA-P; 三种功能 a、 DNA 重组活性 b、 与S.S. DNA结合活性 c、 少数蛋白的proteinase活性 当DNA正常复制时 (无复制受阻,无DNA损伤, 无TT dimer) RecA-p不表现proteinase活性

26
当DNA复制受阻/ DNA damaged 细胞内原少量表达的RecA-p 与S.S, DNA结合 激活RecA-p的proteinase活性 修复损伤 LexA-p降解 SOS open RecA-p高效表达 当DNA复制度过难关后 RecA-p很快消失 LexA gene on SOS off

27
免疫球蛋白基因V片段中突变的引入 免疫球蛋白多样性产生的过程: ♫ 免疫球蛋白重链和轻链基因的V,D,J,C片段重组连接;

28
大肠杆菌的适应性突变 大肠杆菌(乳糖操纵子发生移码突变) 在只有乳糖的培养基上培养 恢复野生性
发现正常生长的细胞(乳糖操纵子发生第二次突变) 引发激烈争论:”环境影响生物的表型”, ”生物对环境作出响应发生程序性突变”
 引发激烈争论: 环境影响生物的表型 , 生物对环境作出响应发生程序性突变")
29
1.4 DNA修复 DNA的修复系统: ♫ 碱基切除修复 将受损的核苷酸碱基周围一段核苷酸切除,然后通过DNA多聚酶重新合成
♫ 核苷酸切除修复 与碱基修复系统类似,只是切除的受损DNA范围更大,涉及更多极端损伤的类型 ♫ 错配修复 ♫ 重组修复

30
碱基切除修复

31
核苷酸切除修复 复制前进行 核苷酸切除修复 不易出错 UvrA, B, C gene 内切核酸酶 (Endonucleases) 外切核酸酶
碱基切除修复 复制前进行 不易出错 核苷酸切除修复 UvrA, B, C gene 内切核酸酶 (Endonucleases) 外切核酸酶 (Exonuclease) DNA pol Ligase
 外切核酸酶. (Exonuclease) DNA pol. Ligase.")
33
错配修复 ♪ 校正校正活性所漏校的碱基, 使复制的保真性提高102~103倍 错配修复系统(MRS Mismatch Repair
♪ 校正校正活性所漏校的碱基, 使复制的保真性提高102~103倍 错配修复系统(MRS Mismatch Repair System) DNA mismatch A----- C--- 错配碱基的修复会使复制的保真性提高102~103倍 DNApol (ξ= 10-8) 经第二次校正ξ= 10-11
 DNA mismatch A C--- 错配碱基的修复会使复制的保真性提高102~103倍. DNApol (ξ= 10-8) 经第二次校正ξ=")
34
♪ 错配修复系统组成(Mismatch repair system)
dam gene DNA腺嘌呤甲基化酶(m6A甲基化酶) DNA polymerase Helicase SSB 外切核酸酶 (Ⅰ和Ⅶ) 连接酶 MCE (mismatch correct enzyme) 3 subunits mutH, L, S 扫描新生链中错配碱基 识别新生链中非 m6A 的GATC序列 酶切含错配碱基的新生DNA区段
 DNA polymerase. Helicase SSB 外切核酸酶 (Ⅰ和Ⅶ) 连接酶. MCE (mismatch correct enzyme) 3 subunits mutH, L, S. 扫描新生链中错配碱基. 识别新生链中非 m6A 的GATC序列. 酶切含错配碱基的新生DNA区段.")
35
♪ 修复过程 a、MutH/MutS 扫描识别错配 碱基和邻近的GATC序列 切点--甲基化GATC中 G的5’侧 甲基化程度的差异
DNA helicase II, SSB, exonuclease I去除包括错 配碱基的片段 Immediate methylation of the A in GATC of the parental strand but not of the daughter strand right after replication Expensive to keep the accuracy DNA polymerase III 和 DNA ligase 填充缺口 昂贵的代价用于保证DNA的准确性

36
重组修复 后复制修复、E.coli的挽回系统 该系统存在的实验证据 E.coli 存活% U.V 计量 w.t. UvrA+ RecA+

37
★ Rec-A. gene 以某种方式参与DNA损伤修复
目前知道 ♫ Uvr系统负责切除二聚体 ♫ Rec系统负责消除没有被切除的二聚体 可能造成的后果

38
复制过程越过二聚体而在相应新链上留下缺口
★ 修复时期的证明 E.coli (Rec-A, uvr-a-) D.S. DNA S.S. DNA U.V 复制 提取 变性 TT TT TT TT AA AA 变性 TT TT AA AA 复制过程越过二聚体而在相应新链上留下缺口 ★二聚体后起始
 D.S. DNA. S.S. DNA. U.V. 复制. 提取 变性. TT TT. TT TT. AA AA. 变性. TT TT. AA AA. 复制过程越过二聚体而在相应新链上留下缺口. ★二聚体后起始.")
39
重组修复 (链转移修复) 复制后修复 二聚体后起始 容易出错 RecA, DNApolymerae ligase RecA
重组修复后的损伤位点可 由其它机制进一步修复 聚合酶、连接酶

40
DNA修复和人类疾病

41
1.5 DNA单链的非对称性进化 大肠杆菌DNA复制的差错率107个核苷酸1个错误;
2条新合成的子链中差错率分布不一致, 延滞链复制的差错率是引导链的20倍; 造成子链差错率非均一性的主要原因: ♫ DNA双链复制的非对称性; 延滞链的复制总比先行链慢一拍,先行链在复制前只有很短的DNA双链区解链,但延滞链却要求很长的一段单链暴露; 延滞连采取冈畸模型复制,大肠杆菌基因组每隔2Kb起始一次引物合成,DNA多聚酶Ⅰ的碱基选择活性及较读能力均比DNA多聚酶Ⅲ差 ♫ 转录的非对称性; 转录时,非转录链则保持短暂单链暴露状态,增加了碱基突变的可能.,已知单链状态的胞嘧啶脱氨基的比例高于双链DNA上百倍,转录状态使非转录链脱氨基比例增加4倍;

42
2.重组 同源重组 Holliday模型 “亲本链” “重组体”

43
3‘ 5‘ 切割 Meselson-Radding模型 单链入侵模型(链转移模型) 5’ 置换 侵入 Loop切除 同化 异构化 分支迁移
 5’ 置换 侵入 Loop切除 同化 异构化 分支迁移")
44
位点专一性重组 大肠杆菌的位点专一 性重组(同源重组): ◘ att位点为 POP’(240bp) BOB’(23bp)
◘ 整合过程需要λ整合酶 (integrase Int)(λ编码) 和寄主的整合宿主因子IHF (integration host factor) 共同作用
(λ编码) 和寄主的整合宿主因子IHF. (integration host factor) 共同作用.")
45
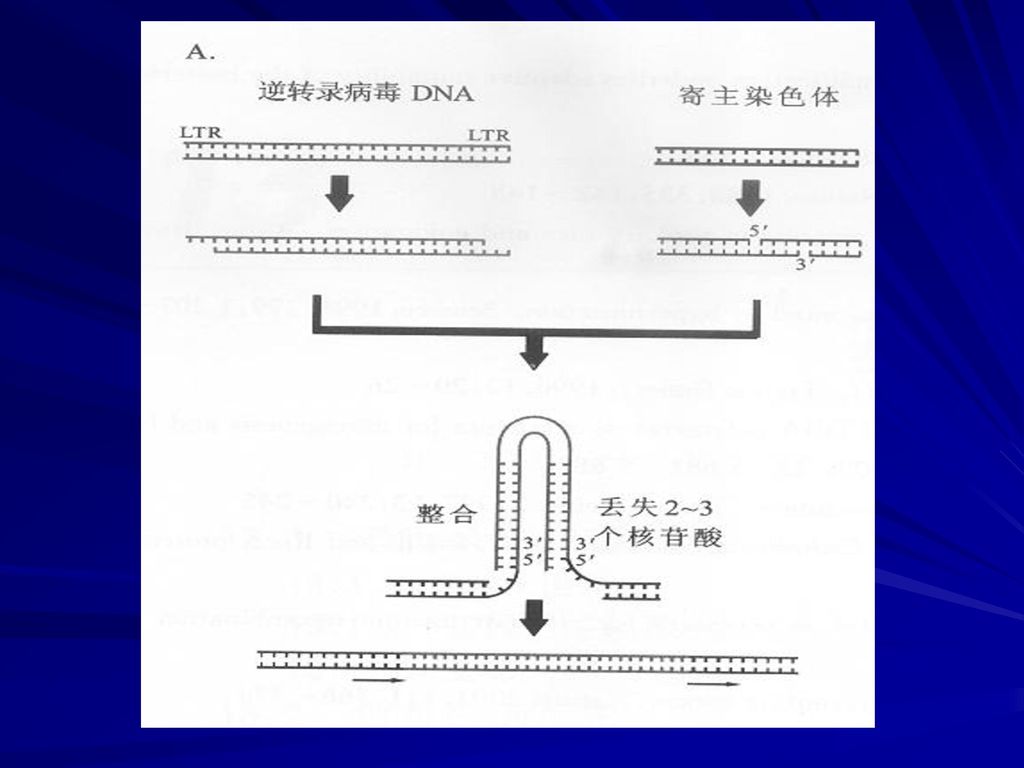
免疫球蛋白基因的重排(双链断裂重接模型):
:")
46
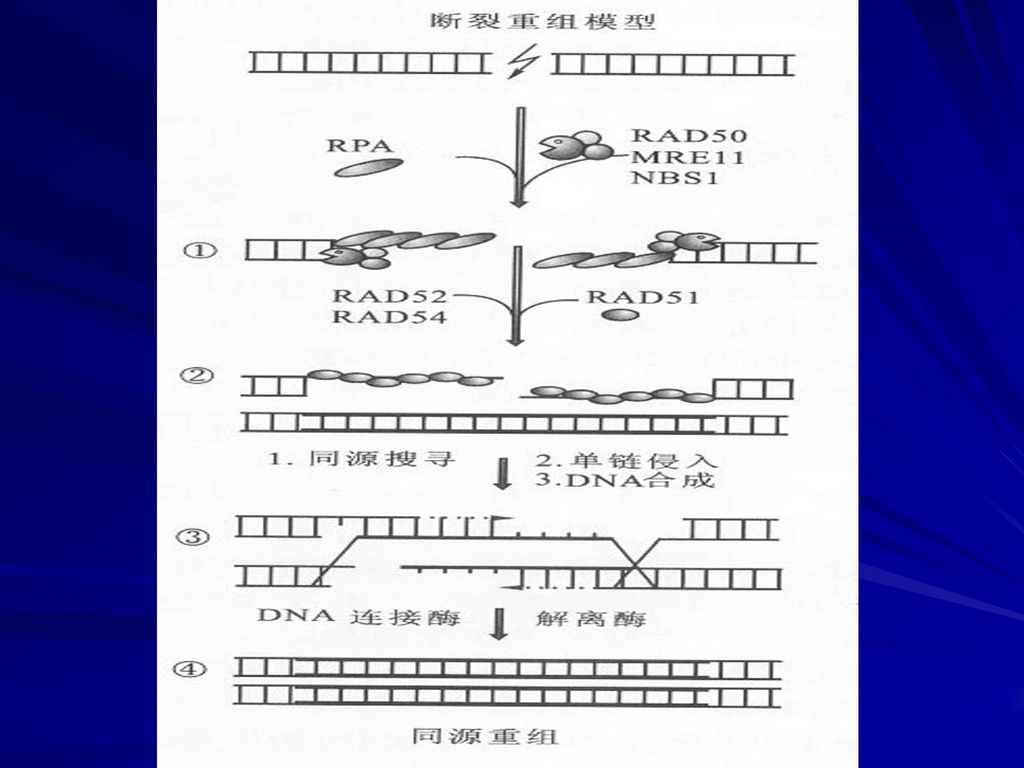
双链断裂重组模型

48
3. 转座 转座: 是基因进化的一种重要方式,它不是重组,但利用了重组的过程,它是一段DNA或其拷贝从基因组的一个位置转移到另一位置,并在插入位点两测产生1对很短的正向重复序列 根据转座机制可分为2个大的范畴: DNA转座子 以DNA区段作为转座成分 又可分为复制转座和保守转座 逆转录转座子 以RNA为中介

49
DNA转座 ♫复制转座

50
♫ 保守转座 杂种分子形成后 细菌中,转座子切除后留下的空缺不能修复,供体分子降解 真核生物中,空缺由双链断裂重组修复

51
逆转录转座

53
本章主要内容 ♫ 基因组进化的分子基础 ♫ 突变的分类及作用效应 ♫ 同源重组的简单过程及Holliday结构形成的分子机制 ♫ 转座子的分类及作用机理





:核蛋白体组成成分 转移 RNA ( tRNA ):转运氨基酸 信使 RNA ( mRNA ):蛋白质合成模板 不均一核 RNA ( hnRNA ):成熟 mRNA 的前体 小核 RNA ( snRNA ):>")





>")






 A.染色体 B.质体 C.线粒体 D.核糖体>")



 A.细菌的遗传物质主要是DNA B.病毒的遗传物质主要是RNA>")
