Download presentation
1
第三章 基因工程载体 第一节 克隆载体 第二节 表达载体 第三节 特殊用途载体

2
载体 (Vectors) DNA: 1)独立的一个包括启动子(promoter)、编码区(encoding region)和终止子(terminator)的基因,or 组成基因的某个元件,一般是不可以进入受体细胞的; 2)采用理化方法进入细胞后,也不容易在受体细胞内维持。所以,通过不同途径能将承载的外源DNA片段带入受体细胞,并在其中得以维持的DNA分子称为基因工程载体。
采用理化方法进入细胞后,也不容易在受体细胞内维持。所以,通过不同途径能将承载的外源DNA片段带入受体细胞,并在其中得以维持的DNA分子称为基因工程载体。")
3
载体 (Vectors) 定义:在基因工程操作中,把能携带外源DNA进入受体细胞的DNA分子叫载体。
多克隆位点(multiple cloning site) ori 复制起始点 遗传标记 pUC MCS Ampr 运送外源基因高效转入受体细胞 2、为外源基因提供复制能力或整合能力 3、为外源基因的扩增或表达提供条件
 ori. 复制起始点. 遗传标记. pUC. MCS. Ampr. 运送外源基因高效转入受体细胞. 2、为外源基因提供复制能力或整合能力. 3、为外源基因的扩增或表达提供条件.")
4
载体的功能 运送外源基因高效转入受体细胞 为外源基因提供复制能力或整合能力 为外源基因的扩增或表达提供必要的条件
目的基因能否有效转入受体细胞,并在其中维持高效表达,在很大程度上决定于载体 。

5
基因工程对载体的要求 (1)在宿主细胞内能独立复制,ori。 (2)有选择性标记 Ampr、Tetr、Kanr等。
(3)多克隆位点:外源基因插入的单一限制酶位点。 (4)分子量小,可容纳较大的外源基因片段。 (5)拷贝数多,方便外源基因在细胞内大量扩增。 外源DNA插入其中不影响载体的复制且切点是单一的,这样可将多个外源 DNA 片段插入其中。 避免基因的非控制性扩散。 (6)具有对受体细胞的可转移性。 (7)具有较好的安全性,不能任意转移。
多克隆位点:外源基因插入的单一限制酶位点。 (4)分子量小,可容纳较大的外源基因片段。 (5)拷贝数多,方便外源基因在细胞内大量扩增。 外源DNA插入其中不影响载体的复制且切点是单一的,这样可将多个外源 DNA 片段插入其中。 避免基因的非控制性扩散。 (6)具有对受体细胞的可转移性。 (7)具有较好的安全性,不能任意转移。")
6
大肠杆菌质粒载体 pBR322结构图 克隆位点 克隆位点 遗传标记基因 复制起点

7
载体 的种类 按功能分类 克隆载体 克隆一个基因或DNA片断 表达载体 用于一个基因的蛋白表达 整合载体 把一个基因插入到染色体组中
表达载体 用于一个基因的蛋白表达 整合载体 把一个基因插入到染色体组中 按来源分类 质粒载体 噬菌体载体 柯斯质粒载体 人工染色体载体 00

8
载体的种类和特征 质粒* 受体细胞 结构 插入片断 举例 E.coli 环状 < 8kb pUC18/19 , T-载体等 λ噬菌体 线状
EMBL系列, λ gt系列 丝状噬菌体及噬菌粒 < 10 kb M13mp系列 粘粒载体 35- 45kb pJB8,c2RB, pcoslEMBL, pWE15/16, pCV BAC (Bacterial Artificial Chromosome) ≈300 kb Pel oBAC系列 YAC (Yeast Artificial chromosome ) 酵母细胞 线性染色体 kb MAC (Mammalian Artificial Chromosome) 哺乳类细胞 > 1000 kb 病毒载体 动物细胞 SV40 载体,昆虫 杆状病毒载体 穿梭载体 和细菌 pSVK3质粒,PBV, Ti质粒
 ≈300 kb. Pel oBAC系列. YAC (Yeast Artificial chromosome ) 酵母细胞. 线性染色体 kb. MAC (Mammalian Artificial Chromosome) 哺乳类细胞. > 1000 kb. 病毒载体. 动物细胞. SV40 载体,昆虫. 杆状病毒载体. 穿梭载体. 和细菌. pSVK3质粒,PBV, Ti质粒.")
9
第一节 克隆载体 1. 质粒载体(plasimid vectors) 2. 噬菌体载体(phage vectors)
第一节 克隆载体 1. 质粒载体(plasimid vectors) 2. 噬菌体载体(phage vectors) 3. 柯斯质粒载体(cosmid vectors) 4. 人工染色体载体(B/Y/HAC)
 2. 噬菌体载体(phage vectors) 3. 柯斯质粒载体(cosmid vectors) 4. 人工染色体载体(B/Y/HAC)")
10
质粒的生物学特性 (1)质粒的概念 的裸露的环状双链DNA分子,比病毒更简单。
质粒是一种广泛从在于细菌细胞中染色体以外的能自主的复制 的裸露的环状双链DNA分子,比病毒更简单。 并不是寄主生长所必需的,但可以赋予寄主某些抵御外界环境因素不利影响的能力(带有抗性基因等) 。
 。")
11
(2)质粒的大小 差异很大,最小的只有1kb,只能编码中等大小的2-3种蛋白质分子,最大的达到200kb。 质粒的生存在寄主细胞中“友好”地“借居”,它可以赋予寄主一些非染色体控制的遗传性状,以利于寄主的生存。比如,对抗菌素的抗性,对重金属的抗性等。 (3)质粒的生存
质粒的生存.")
12
严紧型复制控制的质粒(stringent plasmid) (拷贝数少,为1-5个)
(4)质粒的自主复制性 质粒能利用寄主细胞的DNA复制系统进行自主复制。质粒DNA上的复制子结构决定了质粒与寄主的对应关系。根据在每个细胞中的分子数(拷贝数)多寡,质粒可分为两大复制类型: 严紧型复制控制的质粒(stringent plasmid) (拷贝数少,为1-5个) 松弛型复制控制的质粒(relaxed) (拷贝数多,可达10-200个拷贝) 因此,作为载体的质粒应该是松弛型的。
质粒的自主复制性. 质粒能利用寄主细胞的DNA复制系统进行自主复制。质粒DNA上的复制子结构决定了质粒与寄主的对应关系。根据在每个细胞中的分子数(拷贝数)多寡,质粒可分为两大复制类型: 严紧型复制控制的质粒(stringent plasmid) (拷贝数少,为1-5个) 松弛型复制控制的质粒(relaxed) (拷贝数多,可达10-200个拷贝) 因此,作为载体的质粒应该是松弛型的。")
13
(5)质粒的不相容性 两个质粒在同一宿主中不能共存的现象称质粒的不相容性。具有不相容性的质粒组成的群体称为不相容群,一般具有相同的复制子。
质粒的不相容性 两个质粒在同一宿主中不能共存的现象称质粒的不相容性。具有不相容性的质粒组成的群体称为不相容群,一般具有相同的复制子。")
14
(6)质粒的可转移性 在天然条件下,很多天然质粒都可通过细菌接合作用从一种宿主细胞内转移到另外一种宿主内,这种转移依赖于质粒上的tra基因产物。 Conjugative plasmid 接合型质粒(自我转移的 质粒):质粒可从一个细胞自发转移到另一个细胞。 Non Conjugative plasmid 非接合型质粒 (不能自我转移的质粒):由于失去控制细菌配对和自我转移的基因,质粒不能从一个细胞自发的转移到另一个细胞。 基因工程一般只能利用非接接合型质粒,保证分子操作过程中质粒在细胞中的稳定性。
:质粒可从一个细胞自发转移到另一个细胞。 Non Conjugative plasmid 非接合型质粒. (不能自我转移的质粒):由于失去控制细菌配对和自我转移的基因,质粒不能从一个细胞自发的转移到另一个细胞。 基因工程一般只能利用非接接合型质粒,保证分子操作过程中质粒在细胞中的稳定性。")
15
Tra protein from conjugative plasmid
bom site Mob gene mob mRNA Mob protein open recipient cell helper plasmid 即mob基因的产物可打开非接合质粒的bom 位点(oriT位点),借助接合质粒tra基因的产物,使非接合质粒被动迁移到受体细胞中,这种现象称为迁移作用(mobilization)。
,借助接合质粒tra基因的产物,使非接合质粒被动迁移到受体细胞中,这种现象称为迁移作用(mobilization)。")
16
质粒DNA的tra基因 E.Coli产生菌毛 宿主与受体细胞结合 遗传物在细胞之间转移 (指令) (迁移) 大肠杆菌接合(conjunction)
 (迁移) 大肠杆菌接合(conjunction)")
17
(7)携带特殊的遗传标记 野生型的质粒DNA上往往携带一个或多个遗传标记基因,这使得寄主生物产生正常生长非必需的附加性状,包括: 物质抗性 抗生素、重金属离子、毒性阴离子、有机物 物质合成 抗生素、细菌毒素、有机碱 这些标记基因对DNA重组分子的筛选具有重要意义

18
遗传标记基因 定义:在基因工程中使用与选择重组体DNA转化细胞的基因 1. 指示外源DNA分子(载体或重组分子)是否进入宿主细胞 2. 指示外源DNA分子是否插入载体分子形成了重组子 标记基因的作用
是否进入宿主细胞 2. 指示外源DNA分子是否插入载体分子形成了重组子 标记基因的作用")
19
标记基因的种类 1. 抗性标记基因(可直接用于选择转化子)
a. 抗生素抗性基因: Apr ,Tcr ,Cmr,Kanr,G418r,Hygr ,Neor b. 重金属抗性基因: Cur ,Znr ,Cdr c. 代谢抗性基因: TK,抗除草剂基因 2. 营养标记基因(可直接用于选择转化子) 主要是参与氨基酸,核苷酸及其他必需营养物合成酶类的基因, 这类基因在酵母转化中使用最频繁,如TRP1,URA3,LEU2,HIS4等。 3. 生化标记基因 其表达产物可催化某些易检测的生化反应,如lacZ, GUS,CAT 4. 噬菌斑
 主要是参与氨基酸,核苷酸及其他必需营养物合成酶类的基因, 这类基因在酵母转化中使用最频繁,如TRP1,URA3,LEU2,HIS4等。 3. 生化标记基因. 其表达产物可催化某些易检测的生化反应,如lacZ, GUS,CAT. 4. 噬菌斑.")
20
1.四环素抗性基因(Tcr) Tetracycline 可结合在核糖体30s亚基中的一种蛋白质分子上,抑制核糖体的转位过程。四环素抗性基因编码一种399 AAs蛋白质,与细菌细胞膜结合,阻止四环素分子进入细菌细胞。 2.氨苄青霉素抗性基因(Apr) Ampicillin可抑制细菌细胞膜上参与细胞壁合成酶类的活性。Apr抗性基因编码一种分泌到细菌细胞周间质的酶,催化β-内酰胺环的水解,使氨苄青霉素失活。 3. 氯霉素抗性基因(Cmr) Chlorophenicol可结合在核糖体50 S亚基上,阻止蛋白质合成。Cmr基因编码氯霉素乙酰转移酶,使氯霉素乙酰化,导致乙酰化的氯霉素不能结合在核糖体上。 4. 卡那霉素(Kanr), 新霉素(Neor)和G418抗性(G418r)基因 Kanamycin,Neomycin和G418均属脱氧链霉胺氨基葡萄糖苷类抗生素,可结合在核糖体上阻止蛋白质的合成。来自转座子Tn5和Tn903的Kanr抗性基因均可使这类抗生素磷酸化,使之不能进入细胞内。 20
 Ampicillin可抑制细菌细胞膜上参与细胞壁合成酶类的活性。Apr抗性基因编码一种分泌到细菌细胞周间质的酶,催化β-内酰胺环的水解,使氨苄青霉素失活。 3. 氯霉素抗性基因(Cmr) Chlorophenicol可结合在核糖体50 S亚基上,阻止蛋白质合成。Cmr基因编码氯霉素乙酰转移酶,使氯霉素乙酰化,导致乙酰化的氯霉素不能结合在核糖体上。 4. 卡那霉素(Kanr), 新霉素(Neor)和G418抗性(G418r)基因. Kanamycin,Neomycin和G418均属脱氧链霉胺氨基葡萄糖苷类抗生素,可结合在核糖体上阻止蛋白质的合成。来自转座子Tn5和Tn903的Kanr抗性基因均可使这类抗生素磷酸化,使之不能进入细胞内。 20.")
21
质粒的存在形式有超螺旋、开环双螺旋和线状双螺旋三种。
(8)质粒的存在形式 质粒的存在形式有超螺旋、开环双螺旋和线状双螺旋三种。 双螺旋共价闭合环(超螺旋) 线状双螺旋(两个裂口) 开环双螺旋(一个裂口)
质粒的存在形式. 质粒的存在形式有超螺旋、开环双螺旋和线状双螺旋三种。 双螺旋共价闭合环(超螺旋) 线状双螺旋(两个裂口) 开环双螺旋(一个裂口)")
22
质粒空间构型与电泳速率 同一质粒尽管分子量相同,不同的构型电泳迁移率不同: scDNA最快、l DNA次之、ocDNA最慢。 OC L SC

23
天然质粒的局限性 天然存在的野生型质粒由于分子量大、拷贝数低、单一酶切位点少、遗传标记不理想等缺陷,不能满足克隆载体的要求,因此往往需要以多种野生型质粒为基础进行人工构建。

24
质粒载体的命名原则 人工组建的质粒 人工组建的质粒的第一个字母是质粒英文名字(plasmid)的第一个字符p,用小写。p后有2个字母是大写,表示质粒的作者和实验室名称,再其后为质粒的编号。如pBR322,字母p代表质粒,BR是构建该质粒的研究人员的姓名,322代表…构建的一系质粒的编号。
的第一个字符p,用小写。p后有2个字母是大写,表示质粒的作者和实验室名称,再其后为质粒的编号。如pBR322,字母p代表质粒,BR是构建该质粒的研究人员的姓名,322代表…构建的一系质粒的编号。 .")
25
质粒载体的发展概况 第一阶段(1977年前) 天然质粒和重组质粒的利用,如pSC101, ColE1, pCR, pBR313和pBR322。 第二阶段 增大载体容量(降低载体长度),建立多克隆位点区和新的遗传标记基因。如pUC系列载体。 第三阶段 完善载体功能以满足基因工程克隆中的不同要求,如M13mp系列载体,含T3,T7,sp6启动子载体,表达型载体及各种探针型载体。

26
质粒载体的构建 质粒构建基本策略 1.加入合适的选择标记基因,如两个以上,易于选择转化体 2.增加或减少合适的酶切位点,便于重组
3.缩短长度,提高导入效率,增加装载量 4.改变复制子,变严紧为松弛,变少拷贝为多拷贝 5.根据基因工程的特殊要求加装特殊的基因元件

27
1、选择合适的出发质粒: Ori、选择标记、MCS
质粒构建原则 1、选择合适的出发质粒: Ori、选择标记、MCS 2、正确获得构建质粒载体的元件:酶切、PCR 3、组装合适的选择标记基因 4、选择合适的启动子 5、提高外源DNA的容量 灭活一些有害基因,比如与质粒移动有关的基因,或影响质粒复制的负调控基因等。 6、需要灭活初始质粒上的某些编码基因 7、达到预期目的前提下,构建过程应力求简单

28
质粒载体:作为基因工程载体,质粒至少应该具备复制的起始区、选择标记基因区、多克隆位点等部分。

29
质粒载体的分类 人工构建的质粒根据其功能和用途可分成如下几类: 高拷贝质粒 突变拷贝数控制基因 拷贝数1000-3000 扩增基因
高拷贝质粒 突变拷贝数控制基因 拷贝数 扩增基因 低拷贝质粒 来自pSC101 拷贝数小于10 表达某些毒性基因 温敏质粒 在不同温度下表现出拷贝数、整合等不同性质 测序质粒 含有测序通用引物互补序列和多酶接头polylinker 整合质粒 装有整合促进基因及位点 便于外源基因的整合 穿梭质粒 装有针对两种不同受体的复制子 便于基因克隆 表达质粒 装有强化外源基因表达的转录、翻译、纯化的元件 探针质粒 装有报告基因 便于启动子等元件的克隆筛选

30
质粒载体的分类 (1)高拷贝数的质粒载体 ColE1、pMB1、pMB9 松弛型质粒。 具有低分子量、高拷贝数的优点。
具有氯霉素扩增效应:每个细胞的拷贝数1000-3000个! 用途:适合大量增殖克隆基因、或需要表达大量的基因产物。

31
(2)低拷贝数的质粒载体 (3)温控的质粒载体 由pSC101派生来的载体。 特点是拷贝数低。pLG338、pLG339等
适合于克隆含量过高对寄主代谢有害的基因。 减少蛋白质产物对寄主细胞的毒害。 (3)温控的质粒载体 一些低拷贝基因是温度敏感型,如pBEU1、pBEU2 温度低(<37 oC),拷贝数很少; 温度增加(>40 oC),拷贝数很快增加到>1000个。
温控的质粒载体. 一些低拷贝基因是温度敏感型,如pBEU1、pBEU2. 温度低(<37 oC),拷贝数很少; 温度增加(>40 oC),拷贝数很快增加到>1000个。")
32
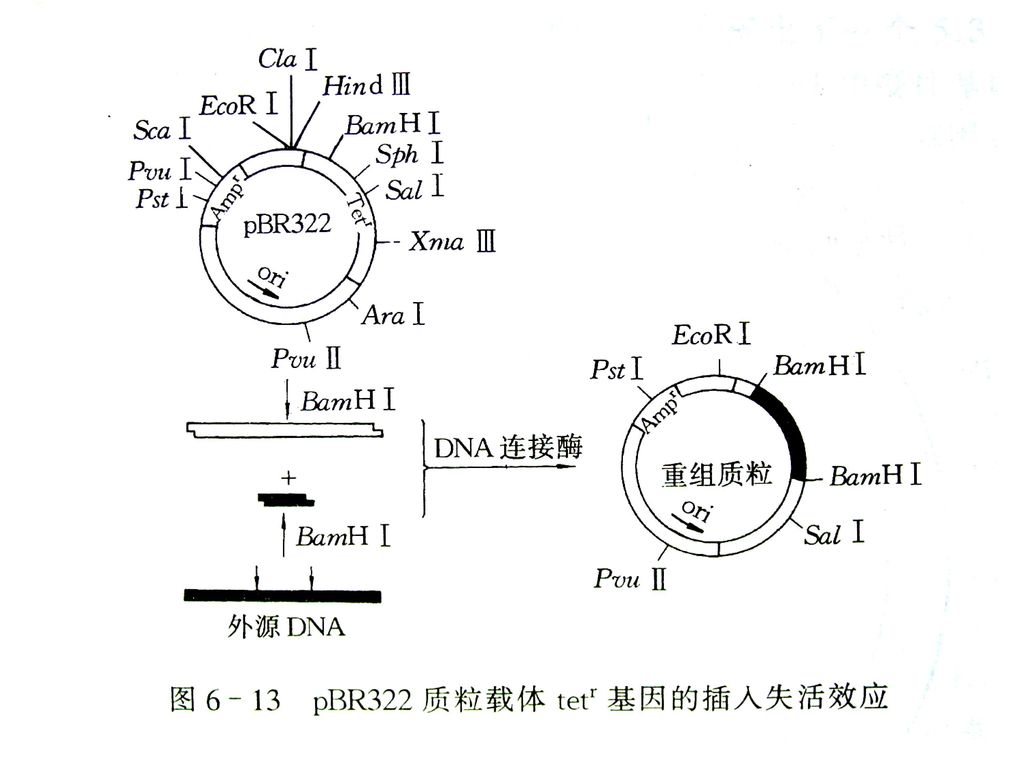
(4) 插入失活型质粒载体 如pDF41、pDF42、pBR322。 无抗生素抗性 抗生素抗性
载体的克隆位点位于其某一个选择性标记基因内部。外源DNA片段插入会导致选择记号基因(如tetr、ampr、cmr等)失活。 如pDF41、pDF42、pBR322。 外源DNA 无抗生素抗性 抗生素抗性
失活。 如pDF41、pDF42、pBR322。 外源DNA. 无抗生素抗性. 抗生素抗性.")
33
质粒载体具有直接选择记号并赋予寄主细胞相应的表型。只有带有选择标记基因的转化菌细胞才能在选择培养基上生长。
(5)正选择的质粒载体 质粒载体具有直接选择记号并赋予寄主细胞相应的表型。只有带有选择标记基因的转化菌细胞才能在选择培养基上生长。 可大大降低需要筛选的转化子的数量,从而减轻实验的工作量,提高了选择的敏感性。 通过选择具这种表型特征的转化子,便可大大降低需要筛选的转化子的数量 只有带有选择标记基因的转化菌细胞才能在选择培养基上生长。 目前通用的绝大部分质粒载体都是正选择载体。 具有直接选择记号并赋予寄主细胞相应的表型,直接选择转化后的细胞,提高了选择的敏感性。 注意启动子的性质,终止子、起始密码、终止密码的阅读正确。 并能在大肠杆菌细胞中正常转录并转译成相应蛋白质的克隆载体特称为表达载体(expression vectors)。 一种典型的大肠杆菌表达型质粒载体(图4-11)的主要组成部分,包括大肠杆菌的启动子及操纵全点序列、多克隆位点、转录及转译信号、质粒载体的复制起点及抗菌素抗性基因。
正选择的质粒载体. 质粒载体具有直接选择记号并赋予寄主细胞相应的表型。只有带有选择标记基因的转化菌细胞才能在选择培养基上生长。 可大大降低需要筛选的转化子的数量,从而减轻实验的工作量,提高了选择的敏感性。 通过选择具这种表型特征的转化子,便可大大降低需要筛选的转化子的数量. 只有带有选择标记基因的转化菌细胞才能在选择培养基上生长。 目前通用的绝大部分质粒载体都是正选择载体。 具有直接选择记号并赋予寄主细胞相应的表型,直接选择转化后的细胞,提高了选择的敏感性。 注意启动子的性质,终止子、起始密码、终止密码的阅读正确。 并能在大肠杆菌细胞中正常转录并转译成相应蛋白质的克隆载体特称为表达载体(expression vectors)。 一种典型的大肠杆菌表达型质粒载体(图4-11)的主要组成部分,包括大肠杆菌的启动子及操纵全点序列、多克隆位点、转录及转译信号、质粒载体的复制起点及抗菌素抗性基因。")
34
经典的大肠杆菌质粒载体 pSC101质粒载体 天然质粒,属严紧型低拷贝质粒。9.09 kb。四环素抗性Tetr。

35
ColE1 天然质粒,属松弛型、高拷贝质粒,6.6Kb。 有氯霉素扩增效应,每个细胞 拷贝

36
大肠杆菌素(colicin)E1和对E1免疫的基因(immE1)
选择标记 大肠杆菌素(colicin)E1和对E1免疫的基因(immE1) colicin E1能杀死不含ColE1 质粒的菌, 形成噬菌斑。 •colicin E1能杀死不含ColE1 质粒的菌,形成“噬菌斑”。 •唯一的克隆位点 EcoR I 正好位于这个基因的内部。 可以通过插入失活筛选。
E1和对E1免疫的基因(immE1) colicin E1能杀死不含ColE1 质粒的菌, 形成噬菌斑。 •colicin E1能杀死不含ColE1 质粒的菌,形成 噬菌斑 。 •唯一的克隆位点 EcoR I 正好位于这个基因的内部。 可以通过插入失活筛选。")
37
③ 不被其他细菌的colicin E1所杀死的原因
cea imm kil 结构基因 免疫基因 溶菌基因 ② 杀死不含有ColE1细菌的原因 cea + kil基因产物 产生菌对细菌素有自身免疫性 大肠杆菌所产生的细菌素称为大肠菌素(colicin),它除作用于某些型别的大肠杆菌外,还能作用于亲缘关系相近的志贺菌、沙门氏菌、克雷伯氏菌和巴氏杆菌等。 ③ 不被其他细菌的colicin E1所杀死的原因 imm基因
,它除作用于某些型别的大肠杆菌外,还能作用于亲缘关系相近的志贺菌、沙门氏菌、克雷伯氏菌和巴氏杆菌等。 ③ 不被其他细菌的colicin E1所杀死的原因. imm基因.")
38
EcoR I位于E1内部,插入外源DNA导致E1失活,使受体菌不能合成E(ColE1-),但仍表现出对E1免疫型(ImmE1+)。
唯一的克隆位点 EcoR I EcoR I位于E1内部,插入外源DNA导致E1失活,使受体菌不能合成E(ColE1-),但仍表现出对E1免疫型(ImmE1+)。 Colicin E1 外源DNA 无Colicin 用对外源colicin E1的免疫性和自身不能合成colicin E1作选择,操作非常繁杂。
,但仍表现出对E1免疫型(ImmE1+)。 Colicin E1. 外源DNA. 无Colicin. 用对外源colicin E1的免疫性和自身不能合成colicin E1作选择,操作非常繁杂。")
39
pBR322: 人工构建载体 p:质粒; BR:质粒两位主要 构建者姓氏的第一个字母; 322:实验编号 三个亲本质粒:
pSF2124 pMB1 pSC101 三个亲本质粒: pMB1:出发质粒(ColE1) pSF2124: Ampr pSC101: Tetr 把pBR322用限制性内切酶切去某片段,换上合用的表达组件,就可以构建成工作所需的新载体。4.36kb的环状双链DNA,碱基序列已经全部清楚。 许多实用的质粒载体都是在pBR322的基础上改建而成。可见其原型质粒在使用上有优点。
 pSF2124: Ampr. pSC101: Tetr. 把pBR322用限制性内切酶切去某片段,换上合用的表达组件,就可以构建成工作所需的新载体。4.36kb的环状双链DNA,碱基序列已经全部清楚。 许多实用的质粒载体都是在pBR322的基础上改建而成。可见其原型质粒在使用上有优点。")
40
3个会导致Ampr基因失活 9个导致Tetr基因失活 氨苄青霉素和四环素抗性 24个单一克隆位点。

41
pBR322的优点 ① 双抗生素抗性选择标记 抗生素抗性基因的插入失活效应是检测重组体质粒的有效方法,分两次先后选择: 没有获得载体的寄主细胞 在Amp或Tet中都死亡。 获得重组载体的寄主细胞 在Amp或Tet其中之一中死亡。

43
不含质粒载体 含有空质粒载体 含有重组质粒载体 Amp Amp+Tet

44
pBR322重组克隆的筛选 重组克隆的 “插入失活”筛选方法 pBR322—→插入在Tetr中,基因型为Tets 、Ampr——→在含有氨卞青霉素培养基上可生长,在含有四环素培养基上不生长; pBR322—→插入在Ampr中,基因型为Tetr 、Amps、——→在含有氨卞青霉素培养基上不生长,在含有四环素培养基可生长; 而在两种抗生素培养基上都生长的是非重组型。这种在一个基因位点中插入外源DNA片段,从而使该基因活性丧失的现象叫插入失活。

46
② 分子小,克隆能力大 ③ 高拷贝数 ④ 安全 ⑤具有较多的单一酶切位点(24种)
长度4363bp,易于纯化,可以携带6-8Kb的外源DNA片段。 ③ 高拷贝数 氯霉素扩增之后,每个细胞可 1000~3000copies ④ 安全 失去了转移蛋白基因mob(mobilization)。不能通过接合转移。 载体越小越好。 >10kb的DNA在纯化过程中容易断裂。缺失流动基因(mob)。这样,质粒就不会从一个细菌接触转移到 ⑤具有较多的单一酶切位点(24种)
。不能通过接合转移。 载体越小越好。 >10kb的DNA在纯化过程中容易断裂。缺失流动基因(mob)。这样,质粒就不会从一个细菌接触转移到. ⑤具有较多的单一酶切位点(24种)")
47
pBR322的缺点 保留了转移蛋白(mob)的作用位点。 能够被ColK质粒编码的mob蛋白识别,如果再有F质粒的参与,就有可能转移!
的作用位点。 能够被ColK质粒编码的mob蛋白识别,如果再有F质粒的参与,就有可能转移!")
48
PBR322的改进 ① 删除mob识别位点 (如质粒pBR327、pAT153等)。 pAT153: 从pBR322上切去HaeII片断,既除去了mob识别位点,又增加质粒的拷贝数。
。 pAT153: 从pBR322上切去HaeII片断,既除去了mob识别位点,又增加质粒的拷贝数。")
49
② 改造EcoR I 位点 pBR325: 使EcoRI 也成为插入失活型位点。 在pBR322位点上接入一段来自噬菌体PICm的HaeII酶切片断(带有氯霉素抗性基因cmlr)。 cmlr上也带一个EcoRI位点。
。 cmlr上也带一个EcoRI位点。")
50
pUC系列载体 在pBR322的基础上改造而成。属正选择载体。 pUC7、pUC8、pUC9、pUC10、pUC11、pUC18、pUC19
加入一个在5端带有10个多克隆位点的基因lacZ’

51
(1)元件来源 a .ori来自pBR322质粒的复制起点 b. 标记基因(ampr):pBR322的Ampr基因
Lacz基因编码的半乳糖苷酶是四聚体。 Lacz’基因含有lacz基因的前59个密码子,a序列,编码a肽。 c. lacz’基因:大肠杆菌lac操纵子DNA区段,编码β-半乳糖苷酶-肽链,是LacZ氨基端的一个片段.载体用于可编码半乳糖苷酶羧基端部分序列的宿主细胞。 d. MCS 多克隆位点。

52
(2)克隆位点 具有MCS(多克隆位点)区段:位于lacZ’基因的5’端。10个连续的单一限制酶切位点,但它不破坏该基因功能。
有mcs的存在,lacz’基因仍然能编码a肽。如果外源基因插入此mcs区,该基因失活。

53
x-gal:5-溴-4-氯-3-吲哚-β-D-半乳糖苷,为生色底物,半乳糖苷酶+ x-gal 蓝色
菌落蓝白选择的原理: IPTG:异丙基-β-D-硫代半乳糖苷,乳糖类似物,又称为安慰诱导物,可代替乳糖诱导乳糖操纵子结构基因的表达,即可诱导lacz’所编码的半乳糖苷酶氨基末端片段(α-肽)的合成。 x-gal:5-溴-4-氯-3-吲哚-β-D-半乳糖苷,为生色底物,半乳糖苷酶+ x-gal 蓝色 x-gal水解后呈现蓝色
的合成。 x-gal:5-溴-4-氯-3-吲哚-β-D-半乳糖苷,为生色底物,半乳糖苷酶+ x-gal. 蓝色. x-gal水解后呈现蓝色.")
54
α-互补:pUC类载体带有lacz’基因,编码半乳糖苷酶氨基端片段(α-肽),此片段与宿主细胞所编码的羧基端半乳糖苷酶实现基因内互补,形成有功能的半乳糖苷酶,称α-互补。
α-互补(α-complementation) α-互补是指 lacZ 基因上缺失近操纵基因区段的突变体与带有完整的近操纵基因区段的 β-半乳糖苷酶(β -galactosidase ,由 1024 个氨基酸组成)阴性的突变体之间实现互补。α-互补是基于在两个不同的缺陷 β-半乳糖苷酶之间可实现功能互补而建立的。大肠杆菌的乳糖 lac 操纵子中的 lacZ 基因编码 β-半乳糖苷酶,如果 lacZ 基因发生突变,则不能合成有活性的 β-半乳糖苷酶。例如, lacZ△M15 基因是缺失了编码 β-半乳糖苷酶中第 个氨基酸的 lacZ 基因,无酶学活性。对于只编码 N-端 140 个氨基酸的 lacZ 基因 (称为 lacZ') ,其产物也没有酶学活性。但这两个无酶学活性的产物混合在一起时,可恢复 β-半乳糖苷酶的活性,实现基因内互补。
 α-互补是指 lacZ 基因上缺失近操纵基因区段的突变体与带有完整的近操纵基因区段的 β-半乳糖苷酶(β -galactosidase ,由 1024 个氨基酸组成)阴性的突变体之间实现互补。α-互补是基于在两个不同的缺陷 β-半乳糖苷酶之间可实现功能互补而建立的。大肠杆菌的乳糖 lac 操纵子中的 lacZ 基因编码 β-半乳糖苷酶,如果 lacZ 基因发生突变,则不能合成有活性的 β-半乳糖苷酶。例如, lacZ△M15 基因是缺失了编码 β-半乳糖苷酶中第 个氨基酸的 lacZ 基因,无酶学活性。对于只编码 N-端 140 个氨基酸的 lacZ 基因 (称为 lacZ ) ,其产物也没有酶学活性。但这两个无酶学活性的产物混合在一起时,可恢复 β-半乳糖苷酶的活性,实现基因内互补。")
55
菌落蓝白斑选择的原理: 在基因克隆时,细菌在含有IPTG和x-gal的培养基中进行培养。 IPTG诱导质粒的lacz’基因产生α-肽,同时诱导细菌产生半乳糖苷酶的羧基端片段。两种片段形成有功能的半乳糖苷酶,从而使x-gal水解,产生蓝色物质,使非重组菌落呈现蓝色。 当外源基因插到质粒的多克隆位点后,使lacz’ 失活,不能表达α-肽,破坏了互补作用,细胞内无 有活性半乳糖苷酶,使带有重组质粒的细菌将产生 白色菌落。

56
互补显色反应

57
a 具有更小的相对分子质量和更高的拷贝数,平均每个细胞可达500-700个拷贝
PUC载体的优点: a 具有更小的相对分子质量和更高的拷贝数,平均每个细胞可达 个拷贝 b 具有MCS片段,可把具有两种不同粘性末端的外源DNA片段直接克隆到pUC类载体上。 2.6kb c 可以用组织化学方法检测重组体(蓝白斑 筛选).
.")
58
pGEM载体 总长度为2743bp 含有一个氨卞青霉素抗性编码基因和一个lacZ’编码基因
一段含有EcoR I、Sat I、Kpn I、Ava I、Sma I、BamH I、XbaI、Sall、AccI、Hinc I、Pst II、Sph I和Hind II等识别序列的多克隆位点。 此序列结构几乎与pUC18克隆载体的完全一样。

59
pGEM系列与pUC系列之间的主要差别 pGEM具有两个来自噬菌体的启动子,即T7启动子和SP6启动子,它们为RNA聚合酶的附着作用提供了特异性的识别位点。 由于这两个启动子分别位于Lac z’基因中多克隆位点区的两侧,故若在反应体系中加入纯化的识别T7或SP6启动子的RNA聚合酶,便可将已克隆的外源基因在体外转录出相应的mRNA。 质粒载体pGEM-3Z和pGEM-4Z在结构上基本相似,两者之间的差别仅仅在于SP6和T7这两个启动子的位置互换、方向相反而已。 pGEM 系列与 pUC 系列之间的主要差别是,它具有两个来自噬菌体的启动子,即 T7 启动子和 SP6 启动子,它们为 RNA 聚合酶的附着提供特异性识别位点。由于这两个启动子分别位于 lacZ‘ 基因中多克隆位点区的两侧,若在反应体系中加入纯化的 T7 或 SP6 RNA 聚合酶,便可以将已经克隆的外源基因在体外转录出相应的 mRNA 。质粒载体 pGEM-3Z 和 pGEM-4Z 在结构上基本相似,两者之间的差别仅仅在于 SP6 和 T7 这两个启动子的位置互换、方向相反而已。

60
pGEM-3Z: 多拷贝 装有多克隆位点(MCS) 正选择颜色标记 lacZ’ 装有两个噬菌体的强启动子 用于外源基因的高效表达
注意:T7和SP6启动子特异性地由噬菌体DNA编码的RNA聚合 酶所识别,因此相应的受体菌必须表达噬菌体RNA聚合酶,如: E.coli BL21(DE3)等
等.")
61
穿梭质粒载体 这种质粒分子上含有两个亲缘关系不同的复制子结构以及相应的选择性标记基因,因此能在两种不同种属的受体细胞中复制并检测,例如既能在原核生物中复制,又能在真核生物中复制的载体。 这类载体既具有细菌质粒的复制原点及选择标记基因,还有真核生物的自主复制序列(ARS)以及选择标记性状 通常穿梭载体在细菌中用于克隆,扩增克隆的基因,在酵母菌中用于基因表达分析. 穿梭载体(shuttle vector)是指含有两个亲缘关系不同的复制子,能在两种不同的生物中复制的。例如既能在原核生物中复制,又能在真核生物中复制的载体.这类载体不仅具有细菌质粒的复制原点及选择标记基因,还有真核生物的自主复制序列(ARS)以及选择标记性状,具有多克隆位点.通常穿梭载体在细菌中用于克隆,扩增克隆的基因,在酵母菌中用于基因表达分析. 61
是指含有两个亲缘关系不同的复制子,能在两种不同的生物中复制的。例如既能在原核生物中复制,又能在真核生物中复制的载体.这类载体不仅具有细菌质粒的复制原点及选择标记基因,还有真核生物的自主复制序列(ARS)以及选择标记性状,具有多克隆位点.通常穿梭载体在细菌中用于克隆,扩增克隆的基因,在酵母菌中用于基因表达分析. 61.")
62
用Taq酶的PCR产物3’端加上了一个A。根据这一特点研制出一种线性质粒,其5’端突出的T,它们之间可以连接,即TA克隆。
①克隆载体的线性化 ②克隆载体末端补平 ③克隆载体末端加T

63
pMD-T 载体



。>")
:核蛋白体组成成分 转移 RNA ( tRNA ):转运氨基酸 信使 RNA ( mRNA ):蛋白质合成模板 不均一核 RNA ( hnRNA ):成熟 mRNA 的前体 小核 RNA ( snRNA ):>")



>")

DNA、RNA及蛋白质操作技术.>")









>")

