Download presentation
Presentation is loading. Please wait.

1
第5章 基因工程载体 Vectors in Gene Engineering
高等学校生物工程、生物科学及生物技术专业教材 基因工程 Gene Engineering 彭银祥等编著 第5章 基因工程载体 Vectors in Gene Engineering 华中科技大学出版社2008年2月第二次印刷

2
2 根据功能,把载体分为: 克隆载体和表达载体
1 载体 (Vectors) 在基因工程操作中,外源DNA片段运送进宿主细胞进行扩增或表达的运载工具称为载体,它也是一个DNA分子。 2 根据功能,把载体分为: 克隆载体和表达载体
 在基因工程操作中,外源DNA片段运送进宿主细胞进行扩增或表达的运载工具称为载体,它也是一个DNA分子。 2 根据功能,把载体分为: 克隆载体和表达载体.")
3
3. 载体的种类 基因工程中常用的载体有5类: 质粒(plasmid) 单链DNA噬菌体M13 噬菌体的衍生物 柯斯质粒(cosmid)
动物病毒(virus)
")
4
4. 基因工程载体的必备条件 (1)复制子,在宿主细胞内能独立复制; (2)有选择性标记,有一个或多个鉴于检测的遗传标记;
4. 基因工程载体的必备条件 (1)复制子,在宿主细胞内能独立复制; (2)有选择性标记,有一个或多个鉴于检测的遗传标记; (3)有一段多克隆位点。位于非必需区内的DNA序列内含有多个单一的酶切位点;外源DNA插入其中不影响载体的复制。 (4)分子量小,拷贝数多。具有较高的遗传稳定性; (5)容易从宿主细胞中分离纯化。
复制子,在宿主细胞内能独立复制; (2)有选择性标记,有一个或多个鉴于检测的遗传标记; (3)有一段多克隆位点。位于非必需区内的DNA序列内含有多个单一的酶切位点;外源DNA插入其中不影响载体的复制。 (4)分子量小,拷贝数多。具有较高的遗传稳定性; (5)容易从宿主细胞中分离纯化。")
5
5.1 质粒载体 质粒(plasmid) 是独立于染色体以外的能自主复制的双链闭合环状DNA分子。一般为1-200kb。
5.1 质粒载体 质粒(plasmid) 是独立于染色体以外的能自主复制的双链闭合环状DNA分子。一般为1-200kb。 存在于细菌、霉菌、蓝藻、酵母等细胞中。
 是独立于染色体以外的能自主复制的双链闭合环状DNA分子。一般为1-200kb。 存在于细菌、霉菌、蓝藻、酵母等细胞中。")
6
大肠杆菌的质粒

7
5.1.1 质粒的一般生物学特性 (1)分子小: 1—200 kb (2)编码基因少: 2—3个中等大小的蛋白质。
质粒的一般生物学特性 (1)分子小: 1—200 kb (2)编码基因少: 2—3个中等大小的蛋白质。 如抗菌素抗性、代谢特征等,赋予细菌一些额外的特性(非必须)。 (3)环形状: 双链环状DNA。 (酵母的“杀伤质粒”是RNA)。
分子小: 1—200 kb. (2)编码基因少: 2—3个中等大小的蛋白质。 如抗菌素抗性、代谢特征等,赋予细菌一些额外的特性(非必须)。 (3)环形状: 双链环状DNA。 (酵母的 杀伤质粒 是RNA)。")
8
(4)质粒的空间构型: ① 共价闭合环状DNA(cccDNA) Covalent close circular DNA 呈超螺旋(SC)(super coil)
质粒的空间构型: ① 共价闭合环状DNA(cccDNA) Covalent close circular DNA 呈超螺旋(SC)(super coil)")
9
② 开环DNA( open circular, ocDNA)
一条链上有一至数个缺口。 ③ 线形DNA ( linear ,lDNA)
")
10
(5)质粒空间构型与电泳速率 同一质粒尽管分子量相同,不同的构型电泳迁移率不同: scDNA最快、l DNA次之、ocDNA最慢。 SC OC L
质粒空间构型与电泳速率 同一质粒尽管分子量相同,不同的构型电泳迁移率不同: scDNA最快、l DNA次之、ocDNA最慢。 SC OC L")
11
2. 质粒的类型 在大肠杆菌中的质粒,可以分为: 接合型质粒: 能自我转移 非接合型质粒:不能自我转移

12
1. 接合型质粒: 又叫自我转移型质粒。 除了带有自我复制所必需的遗传信息外还带有一套控制细菌配对和质粒接合转移的基因。 如:F质粒(性质粒、或F因子): 甚至能使寄主染色体上的基因随其一道转移到原先不存在该质粒的受体菌中。 不符合基因工程的安全要求。

13
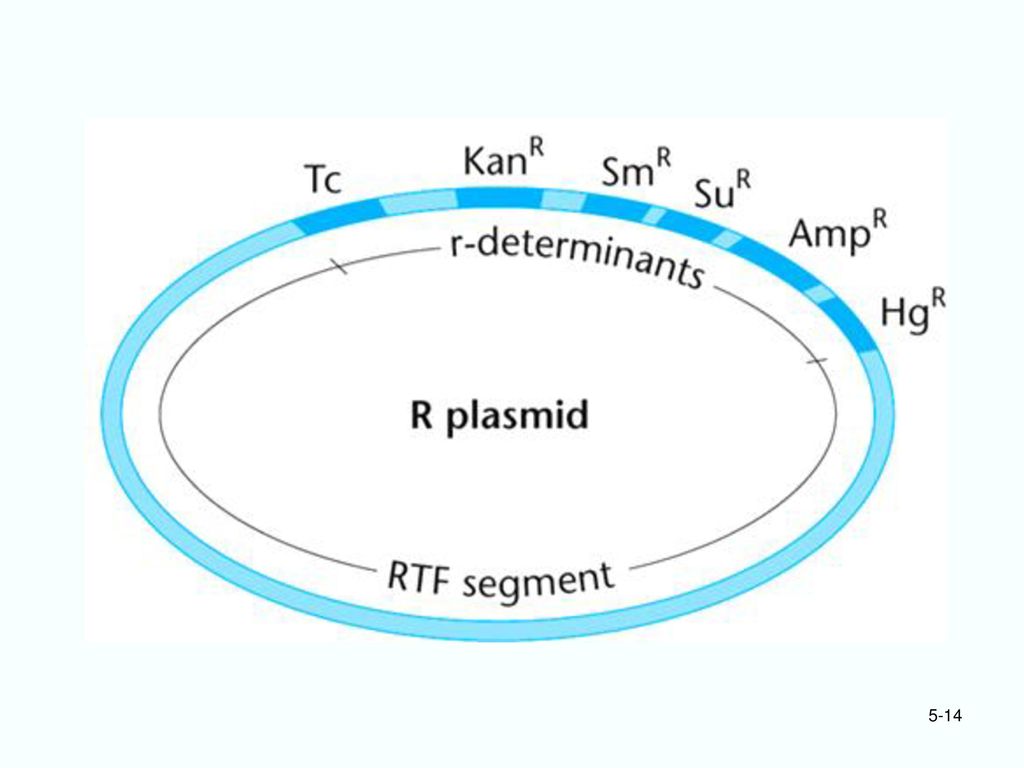
2. 非接合型质粒: 虽然带有自我复制所必需的遗传信息,但失去了控制细菌配对和质粒接合转移的基因,因此不能从一个细胞转移到另一个细胞。 符合基因工程的安全要求。 (1)R质粒(抗性质粒): 带有一种或数种抗生素抗性基因,使寄主获得同样的抗生素抗性性状(resistance)。
R质粒(抗性质粒): 带有一种或数种抗生素抗性基因,使寄主获得同样的抗生素抗性性状(resistance)。 .")
15
(2)Col质粒: 带有控制大肠杆菌素(colicin)合成的基因。 大肠杆菌素对不带Col质粒的大肠杆菌有毒。 Col质粒或R质粒如果带有转移基因,也可以成为接合型质粒。
Col质粒: 带有控制大肠杆菌素(colicin)合成的基因。 大肠杆菌素对不带Col质粒的大肠杆菌有毒。 Col质粒或R质粒如果带有转移基因,也可以成为接合型质粒。")
16
1. 严紧型质粒(stringent plasmid)
质粒的复制类型 1. 严紧型质粒(stringent plasmid) 拷贝数少,只有1—3份拷贝。 (接合型质粒分子量大,一般属严紧型DNA polymerase III )。 2. 松弛型质粒(relaxed plasmid) 拷贝数多,有10—60份拷贝。 (非接合型质粒分子量小,一般属松弛型 DNA polymerase I )。
 拷贝数少,只有1—3份拷贝。 (接合型质粒分子量大,一般属严紧型DNA polymerase III )。 2. 松弛型质粒(relaxed plasmid) 拷贝数多,有10—60份拷贝。 (非接合型质粒分子量小,一般属松弛型. DNA polymerase I )。")
17
5.1.2 构建质粒克隆载体的基本策略 (1)去掉非必需的DNA区,保留质粒必需区,降低分子量; (2)减少质粒本身内切酶位点的数目;
构建质粒克隆载体的基本策略 (1)去掉非必需的DNA区,保留质粒必需区,降低分子量; (2)减少质粒本身内切酶位点的数目; (3)加入易检出的筛选标记; (4)关于质粒安全性的改造,不发生重组和转移,不能离体扩增; (5)改造或增加表达基因的调控序列;如启动子、增强子等。
去掉非必需的DNA区,保留质粒必需区,降低分子量; (2)减少质粒本身内切酶位点的数目; (3)加入易检出的筛选标记; (4)关于质粒安全性的改造,不发生重组和转移,不能离体扩增; (5)改造或增加表达基因的调控序列;如启动子、增强子等。")
18
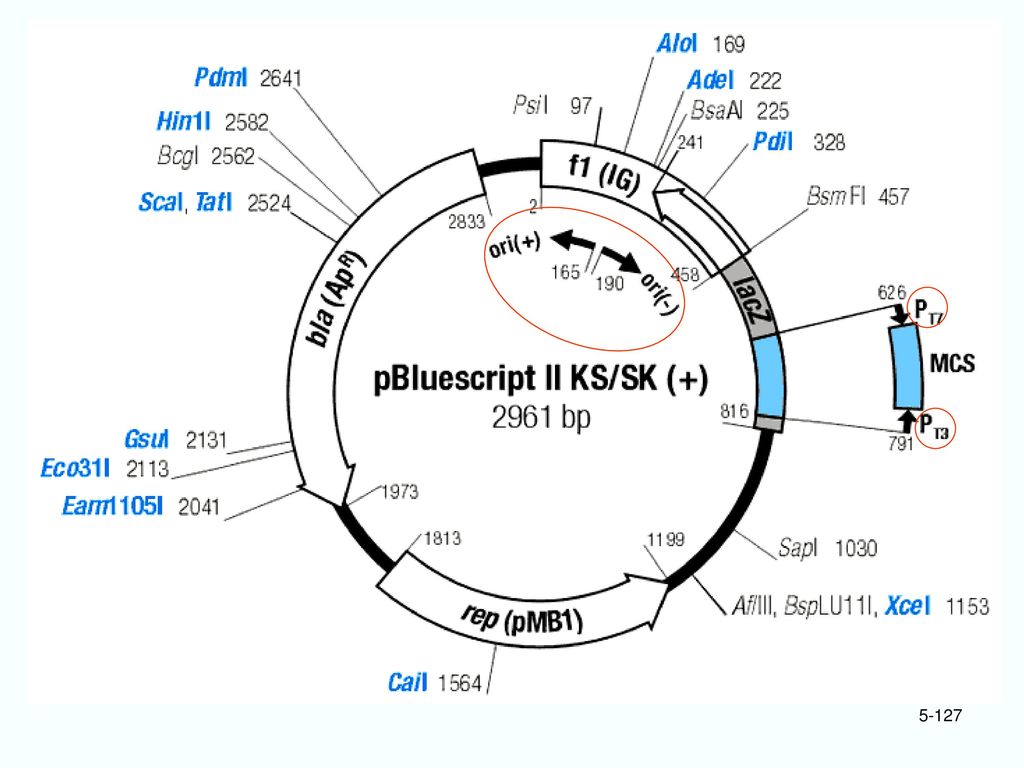
5.1.3 质粒克隆载体的构建 1. pBR322: F. Bolivar和R.L. Rodriguez人工构建载体。 (1)元件来源
质粒克隆载体的构建 1. pBR322: F. Bolivar和R.L. Rodriguez人工构建载体。 (1)元件来源 ① 复制起点 ori:pMB1系列(来源于ColE1)的高拷贝型复制起点 ② Ampr基因:pSP2124质粒的Ampr基因 ③ Tetr基因: pSC101的Tet r 基因。
元件来源. ① 复制起点 ori:pMB1系列(来源于ColE1)的高拷贝型复制起点. ② Ampr基因:pSP2124质粒的Ampr基因. ③ Tetr基因: pSC101的Tet r 基因。")
19
(2)长度 4363bp (3)选择标记 氨苄青霉素和四环素抗性。 (4)克隆位点 24个克隆位点。 其中9个会导致Tetr基因失活(如BamH I、Hind Ⅲ、Sal I); 3个会导致Ampr基因失活(Sca I、PvuI、Pst I)。
; 3个会导致Ampr基因失活(Sca I、PvuI、Pst I)。 .")
20
4363bp

21
(5)pBR322的优点 ① 双抗菌素抗性选择标记 插入失活,分两次先后选择: 没有获得载体的寄主细胞 在Amp或Tet中都死亡。 获得载体的寄主细胞 在Amp或Tet其中之一中死亡。
pBR322的优点 ① 双抗菌素抗性选择标记 插入失活,分两次先后选择: 没有获得载体的寄主细胞 在Amp或Tet中都死亡。 获得载体的寄主细胞 在Amp或Tet其中之一中死亡。")
22
外源基因BamH I 外源基因Pst I Amp中存活 Tet中存活 但在Tet中死亡 但在Amp中死亡 ?

23
② 分子小,克隆能力大 载体越小越好。 >10kb的DNA在纯化过程中容易断裂。 ③ 高拷贝数 氯霉素扩增之后,每个细胞可达1000~3000copy ④ 安全 失去了转移蛋白基因mob(mobilization)。不能通过接合转移。
。不能通过接合转移。 .")
24
(6)pBR322的缺点 保留了转移蛋白(mob)的作用位点。 能够被ColK质粒编码的mob蛋白识别,如果再有F质粒的参与,就有可能转移!
pBR322的缺点 保留了转移蛋白(mob)的作用位点。 能够被ColK质粒编码的mob蛋白识别,如果再有F质粒的参与,就有可能转移!")
25
(7)pBR322的改进 ① 删除mob识别位点 (如质粒pBR327、pAT153等)。 pAT153: 从pBR322上切去HaeII片断,既除去了mob识别位点,又增加质粒的拷贝数。
pBR322的改进 ① 删除mob识别位点 (如质粒pBR327、pAT153等)。 pAT153: 从pBR322上切去HaeII片断,既除去了mob识别位点,又增加质粒的拷贝数。")
26
② 改造EcoR I 位点 pBR325: 使EcoRI 也成为插入失活型位点。 在pBR322位点上接入一段来自噬菌体PICm的HaeII酶切片断(带有氯霉素抗性基因cmlr)。 cmlr上也带一个EcoRI位点。
。 cmlr上也带一个EcoRI位点。")
27
pUC7、pUC8、pUC9、pUC10、pUC11、pUC18、pUC19
University of California的J. Messing和J. Vieria于1978年,在pBR322的基础上改造而成。属正选择载体。 pUC7、pUC8、pUC9、pUC10、pUC11、pUC18、pUC19

28
(1)元件来源 ① 复制起点: pBR322的 ori ② Ampr 基因: pBR322的Ampr基因 ③ lacZ的启动子: 来自于大肠杆菌 ④ lacZ’基因: 大肠杆菌LacZ的-肽链序列, 是LacZ 的氨基端片断。

29
(2)长度:约2.7kb (3)克隆位点: 10个连续的单一限制酶切位点,位于lacZ’基因的5’端。
长度:约2.7kb (3)克隆位点: 10个连续的单一限制酶切位点,位于lacZ’基因的5’端。")
30
β-galactosidase coding sequence (26aa)
pUC18/19 EcoRI SacI KpnI SmalI BamHI XbaI SalI PstI SphI Hind III ATGACCATGATTACGAATTCGAGCTCGGTACCCGGGGATCCTCTAGAGTCGACCTGCAGGCATGCAAGCTTGGCACTG TACTGGTACTAATGCTTAAGCTCGAGCCATGGGCCCCTAGGAGATCTCAGCTGGACGTCCGTAGCTTCGAACCGTGAC β-galactosidase coding sequence (26aa)
")
31
Ampicillin 抗性和 lacZ的肽互补(蓝白斑)相结合。
(4)选择标记 Ampicillin 抗性和 lacZ的肽互补(蓝白斑)相结合。 蓝白斑选择原理: ① Xgal (5-Bromo-4-chloro-3-indolyl--D-galactoside)
选择标记. Ampicillin 抗性和 lacZ的肽互补(蓝白斑)相结合。 蓝白斑选择原理: ① Xgal. (5-Bromo-4-chloro-3-indolyl--D-galactoside)")
32
-半乳糖苷酶能把无色的化合物Xgal分解成半乳糖和一个深蓝色的物质5-溴-4-氯靛蓝。

33
C端大部分 C端大部分 C端大部分 C端大部分
③ lacZ的肽互补 1)-肽( lacZ’ ): -半乳糖苷酶N端的一段氨基酸片断(11-41氨基酸)。 N端的11-41aa C端大部分 N端的11-41aa C端大部分 4聚体 N端的11-41aa C端大部分 N端的11-41aa C端大部分 lacZ只有在4聚体的状态下才有功能.
-肽( lacZ’ ): -半乳糖苷酶N端的一段氨基酸片断(11-41氨基酸)。 N端的11-41aa. C端大部分. N端的11-41aa. C端大部分. 4聚体. N端的11-41aa. C端大部分. N端的11-41aa. C端大部分. lacZ只有在4聚体的状态下才有功能.")
34
2)受体菌lacZ突变(lacZ∆M15) 受体菌基因组的-半乳糖苷酶基因的氨基端有缺失(缺失肽),不能形成4聚体的活性酶,不能分解Xgal 受体菌株:JM系列、TG1、TG2、XL1-blue、XS127、XS101、KK2186、MV1184、DH5a

35
3)载体lacZ’与互补 pUC质粒载体上的lacZ’ 编码肽与这个缺失突变的-半乳糖苷酶“互补”,使它能形成4聚体。又能分解Xgal。产生蓝色物质。 N端的11-41aa C端大部分 受体菌lacZ∆ pUC lacZ’

36
pUC载体上LacZ’的5‘端有一段多克隆位点(MCS)区,本身虽不干扰LacZ’的合成,但插入外源基因就会阻止LacZ’的合成。不能互补。
4)互补的插入失活 pUC载体上LacZ’的5‘端有一段多克隆位点(MCS)区,本身虽不干扰LacZ’的合成,但插入外源基因就会阻止LacZ’的合成。不能互补。 lacZ’ 5’ 3’ MCS lacZ’ 5’ 3’ 外源DNA 肽 肽移码突变 互补 不互补
互补的插入失活. pUC载体上LacZ’的5‘端有一段多克隆位点(MCS)区,本身虽不干扰LacZ’的合成,但插入外源基因就会阻止LacZ’的合成。不能互补。 lacZ’ 5’ 3’ MCS. lacZ’ 5’ 3’ 外源DNA. 肽. 肽移码突变. 互补. 不互补.")
37
④ IPTG的诱导作用 IPTG是乳糖的类似物。能诱导lac操纵子的启动转录,使受体菌基因组中的lacZ 的C端部分和载体的lacZ’肽都表达。从而互补。 但载体MCS上插入外源DNA后,不能产生肽! IPTG

38
IPTG诱导的结果: MCS无插入时,互补,蓝菌斑。 MCS有插入时,不互补,白菌斑。 通过看培养皿上的菌斑的颜色就能直接知道是否有DNA插入。

39
无质粒的 受体菌 有蓝色菌斑生长 有白色菌斑生长 -半乳糖苷酶部分缺失,不能分解Xgal;无抗菌素抗性 在含抗菌素和Xgal的培养基上培养
无菌斑生长 -半乳糖苷酶的缺失被载体产物互补,能分解Xgal。且有抗菌素抗性 在含抗菌素和Xgal的培养基上培养 有蓝色菌斑生长 质粒转化的受体菌 载体产物失活,不能互补-半乳糖苷酶的缺失。不能分解Xgal,但有抗菌素抗性 带外源DNA插入的质粒转化的受体菌 在含抗菌素和Xgal的培养基上培养 有白色菌斑生长

41
(5)pUC系列载体的优点 ① 更小的分子量: 如pUC18为2682bp,pUC8为2750bp。 ② 选择方便 Xgal显色、抗菌素双重直接选择。 ③ 克隆便利 具有多克隆位点(MCS),使有两个不同粘性末端的外源DNA方便地插入。 ④ 测序方便

42
pUC的MCS与M13噬菌体载体的MCS完全相同,便于把外源DNA转移到M13载体上测序。
M13mp18的多克隆位点

43
3. pUC的衍生质粒载体 1. pGEM-3Z 可被T7和SP6的RNA聚合酶识别转录。 lacZ’ Ampr T7启动子 MCS
由pUC派生而来。与pUC的主要区别是在MCS的两侧分别加了一个噬菌体启动子T7和SP6。 可被T7和SP6的RNA聚合酶识别转录。 lacZ’ Ampr T7启动子 MCS SP6启动子 ori

44
pGEM-3Z的特点: ① 如果加入纯化的T7或SP6 RNA聚合酶,在试管里就可以转录mRNA ② 外源基因正接反接都可以转录。 ③ MCS与pUC18的完全一样。 2. pGEM-4Z: 与pGEM-3Z相同,只是T7启动子和SP6启动子的位置互换。

45
T2 Genomic DNA single linear DNA (about 182,000 bp)
")
46
5.2 噬菌体载体 1. 噬菌体的一般特性 噬菌体是一类细菌病毒(Bacteriophage)。 结构:
1. 噬菌体的一般特性 噬菌体是一类细菌病毒(Bacteriophage)。 结构: 蛋白质外壳内包裹着DNA(双链、单链、线性、环状等)。
。 结构: 蛋白质外壳内包裹着DNA(双链、单链、线性、环状等)。")
47
2. 噬菌体DNA分子的特点 (1)长度为48502 bp; (2)双链线性DNA; (3)但在两端有cos位点,可以环化。 cos位点(cohensive-end site): DNA两端各有12bp的粘性末端,粘性末端形成的双链区域称为cos位点。
长度为48502 bp; (2)双链线性DNA; (3)但在两端有cos位点,可以环化。 cos位点(cohensive-end site): DNA两端各有12bp的粘性末端,粘性末端形成的双链区域称为cos位点。 .")
48
DNA进入细菌体内后,可形成双链环状DNA复制型(RF DNA)。
。")
49
噬菌体和其DNA

50
3. 噬菌体的基因组特点 DNA上有至少61个基因,有一半是必须的,与自身的活动有关,成簇排列。
其他约1/3是非必须基因(位于尾部合成至阻遏之间的区段)。 19.6kb 头尾蛋白合成 12-24kb 重组 整合剪切 9-11kb DNA复制 溶原
。 19.6kb 头尾蛋白合成 kb 重组 整合剪切. 9-11kb DNA复制 溶原.")
51
4. 噬菌体的生活周期 分为两种: 1)溶源化周期: 原噬菌体(prophage):
感染细菌后,DNA环化,将自己的DNA整合到细菌的染色体DNA中。形成这一过程称为溶源化(lysogenization)。 原噬菌体(prophage): 整合到细菌染色体的噬菌体DNA称为原噬菌体,噬菌体DNA随细菌的染色体复制而复制。
。 原噬菌体(prophage): 整合到细菌染色体的噬菌体DNA称为原噬菌体,噬菌体DNA随细菌的染色体复制而复制。")
53
2) 溶菌周期: 感染细菌后立即在菌体内复制和合成蛋白质外壳,重新组装成噬菌体颗粒,并导致细菌细胞解体,释放出大量子代噬菌体。
 溶菌周期: 感染细菌后立即在菌体内复制和合成蛋白质外壳,重新组装成噬菌体颗粒,并导致细菌细胞解体,释放出大量子代噬菌体。")
56
3) DNA的复制 模型 (1) 型复制 噬菌体感染细菌的早期:双链进行型复制。
 DNA的复制 模型 (1) 型复制 噬菌体感染细菌的早期:双链进行型复制。")
57
(2)滚环复制 感染的晚期:启动滚环复制机制。 形成多联体DAN分子。
滚环复制 感染的晚期:启动滚环复制机制。 形成多联体DAN分子。")
58
这种多联体分子必须在cos位点被切断成单体分子才能被包装起来。

59
噬菌体DNA上本身有5个 EcoR I 和7 个Hind III切点!
5. 噬菌体载体的构建 (1)构建原理: 删除噬菌体的非必需区,留出插入空间。 (2)构建过程 ①在这个非必须区内制造限制酶切点 ②引进某些突变表型,作为选择标记 ③突变某些基因,使它成为安全载体 ④删除DNA必须区段上常用的限制酶切点 噬菌体DNA上本身有5个 EcoR I 和7 个Hind III切点!
构建原理: 删除噬菌体的非必需区,留出插入空间。 (2)构建过程. ①在这个非必须区内制造限制酶切点. ②引进某些突变表型,作为选择标记. ③突变某些基因,使它成为安全载体. ④删除DNA必须区段上常用的限制酶切点. 噬菌体DNA上本身有5个 EcoR I 和7 个Hind III切点!")
60
① 插入型载体(insertion vectors):
(3)人工构建的噬菌体载体有两种类型: ① 插入型载体(insertion vectors): 如 gt10、gt11、BV2、NM540、NM1590、NM607 1)免疫功能失活型: 插入位点位于载体合成活性阻遏物区域(cI基因)内。 cI gt10 EcoR I
人工构建的噬菌体载体有两种类型: ① 插入型载体(insertion vectors): 如 gt10、gt11、BV2、NM540、NM1590、NM607. 1)免疫功能失活型: 插入位点位于载体合成活性阻遏物区域(cI基因)内。 cI. gt10. EcoR I.")
61
如gt10载体的两个克隆位点(EcoR I 和Hind III)都是位于cI基因内部。
选择标记: 没有外源DNA插入的载体感染受体菌形成混浊噬菌斑。 插入导致载体不能合成阻遏物,载体DNA不能进入溶源期,受体菌全部裂解,形成清晰的噬菌斑。 如gt10载体的两个克隆位点(EcoR I 和Hind III)都是位于cI基因内部。
都是位于cI基因内部。")
62
在基因组中引入了LacZ’序列。(其上含有一个EcoR I 克隆位点)。
2)-半乳糖苷酶失活型载体: 在基因组中引入了LacZ’序列。(其上含有一个EcoR I 克隆位点)。 LacZ’ Charon16A EcoR I 选择标记: 感染LacZ突变的大肠杆菌,经IPTG的诱导,利用Xgal的显色反应作选择标记。
-半乳糖苷酶失活型载体: 在基因组中引入了LacZ’序列。(其上含有一个EcoR I 克隆位点)。 LacZ’ Charon16A. EcoR I. 选择标记: 感染LacZ突变的大肠杆菌,经IPTG的诱导,利用Xgal的显色反应作选择标记。")
63
② 替换型载体(substitution vectors)
两个多克隆位点区以反向重复形式分别位于DNA的非必须区两端。 MCS MCS 可置换区 用酶切后,可以将中间的区段切下来,与用同样酶切成的外源DNA片断置换。 如: EMBL3、Charon40等。

64
EMBL3的可置换片断内部还有Sal I酶切位点。
BamH I EcoR I Sal I BamH I EcoR I Sal I Sal I 左臂 可置换区 右臂 EMBL3 以便于进一步消化中间片断,防止自我连接。

65
Charon40的可置换片断是由DNA短片重复构成,片断之间有Hae I 切点。
MCS MCS 左臂 可置换区 右臂 Charon40 在克隆的时候,可以用Hae I 酶进一步将可置换片断切成小片断,以防止重新与载体连接。

66
(4)载体的筛选标记 ① 置换型载体 可取代片断中如果包含LacZ’,可用Xgal显色作筛选标记。 ② 插入型载体 根据插入所引起的表型突变。
载体的筛选标记 ① 置换型载体 可取代片断中如果包含LacZ’,可用Xgal显色作筛选标记。 ② 插入型载体 根据插入所引起的表型突变。")
67
载体克隆策略 置换型载体

68
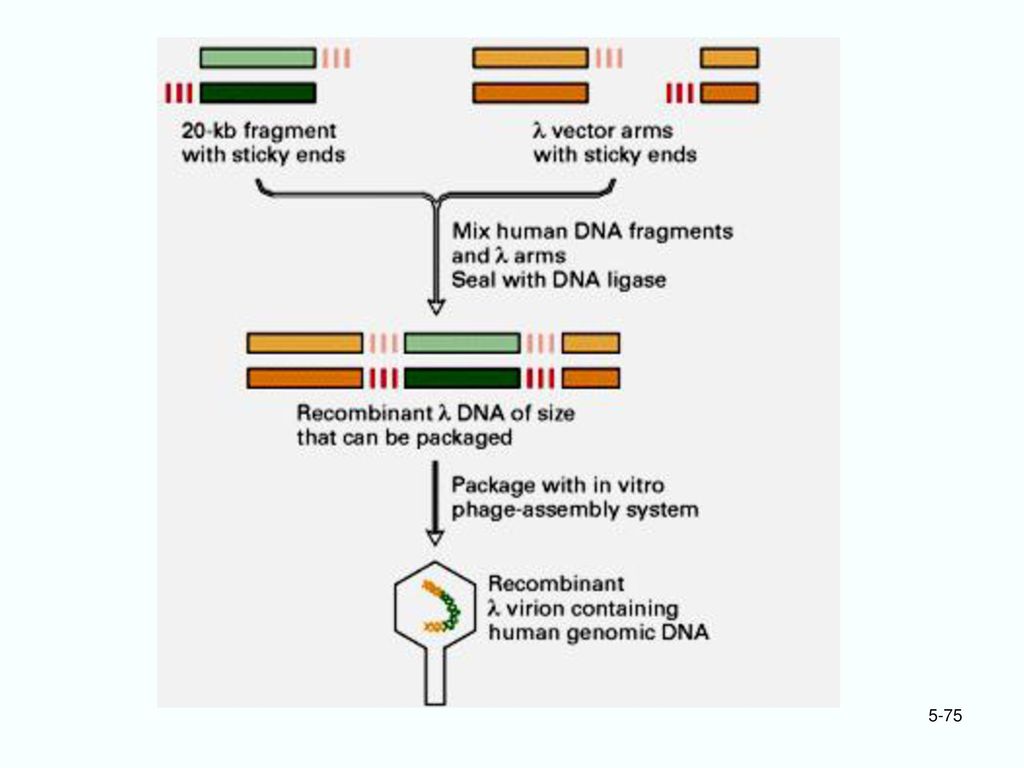
6. 噬菌体的体外包装 DNA象质粒载体那样直接转化细菌时效率远比质粒低。 必须利用噬菌体外壳的作用! (1)体外包装: 在试管中与噬菌体的头部和尾部蛋白人工装配成噬菌体颗粒,才能高效地感染细菌,把重组DNA注入受体菌中。
体外包装: 在试管中与噬菌体的头部和尾部蛋白人工装配成噬菌体颗粒,才能高效地感染细菌,把重组DNA注入受体菌中。 .")
70
(2)体外包装过程 ① 噬菌体外壳蛋白的合成 E 基因 的E 基因编码头部蛋白的主要成分(占头部总蛋白的70%)。 D基因 的D基因的产物也是头部蛋白,但主要与 DNA 进入头部和头部的成熟有关(只占20%)。
。 .")
71
只合成尾部蛋白和包装识别蛋白 提取尾部蛋白 E基因突变 体外包装成活性噬菌体 合成头部蛋白和尾部蛋白,但不能聚合和包装DNA 提取头部和尾部蛋白 D基因突变 重组的载体DNA

72
(3)体外包装存在的问题: ① 包装错误: 既能包装用于生产头部蛋白的内源性原DNA,也能包装重组的DNA载体。 ② 重组: 外源的重组DNA载体被诱发与内源性原DNA发生重组。
体外包装存在的问题: ① 包装错误: 既能包装用于生产头部蛋白的内源性原DNA,也能包装重组的DNA载体。 ② 重组: 外源的重组DNA载体被诱发与内源性原DNA发生重组。")
73
③ 体外包装的容量限制: 必须在正常野生型容量的75%—105% (36—51kb)之间。 既不能超过正常野生型容量的5%; 又不能低于正常野生型容量的75% 野生型DNA的必须区是28kb,所以载体的最大克隆容量是23kb。(实际上为15kb)。
之间。 既不能超过正常野生型容量的5%; 又不能低于正常野生型容量的75% 野生型DNA的必须区是28kb,所以载体的最大克隆容量是23kb。(实际上为15kb)。")
74
(4)DNA载体的优点 比一般的质粒载体的容量大的多。 用在真核生物基因组文库的建立。 BamHI Sau3A
DNA载体的优点 比一般的质粒载体的容量大的多。 用在真核生物基因组文库的建立。 BamHI Sau3A")
76
噬菌体感染细菌形成的噬菌斑

77
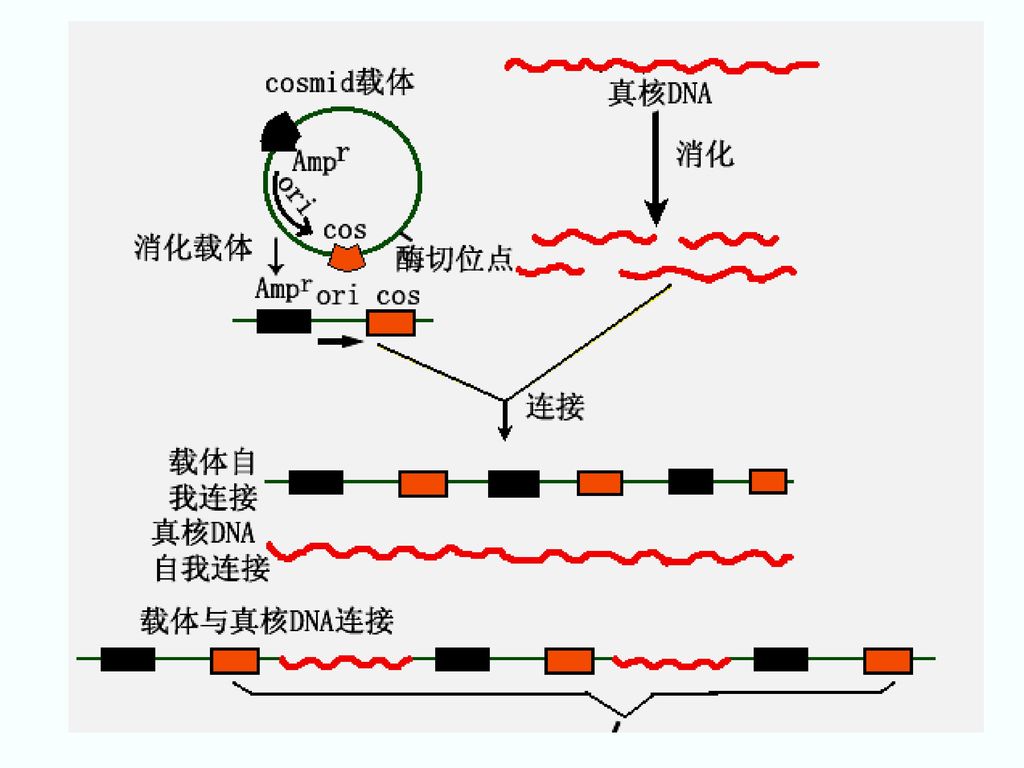
cosmid载体(粘粒) 1978年J. Collins和B. Hohn等人发展出cosmid vector(cos site-carrying plasmid) (1)大小 5—7kb (2)组成 DNA: cos序列和控制包装的的序列。 质粒的复制子 抗药性基因 几个限制性酶的单一位点 pBR322:
组成. DNA: cos序列和控制包装的的序列。 质粒的复制子. 抗药性基因. 几个限制性酶的单一位点. pBR322:")
78
pHC79

79
(3)cosmid vector的特点 ①具有噬菌体的特性; ②具有质粒载体的特性;
在寄主细胞内形成环化DNA(但不能形成新的噬菌体颗粒而溶菌 )。 克隆外源DNA后,可以体外包装成噬菌体颗粒。 ②具有质粒载体的特性; 大多带有pMB1或ColE1的复制子,能象质粒一样复制。
。 克隆外源DNA后,可以体外包装成噬菌体颗粒。 ②具有质粒载体的特性; 大多带有pMB1或ColE1的复制子,能象质粒一样复制。")
80
③方便的选择; 有抗菌素抗性基因,和插入失活的克隆位点。 ④具有高容量的克隆能力; 45kb (最少不能低于30kb)。 51kb-5kb=45kb ⑤具有与含同源序列的质粒进行重组的能力;
。 51kb-5kb=45kb ⑤具有与含同源序列的质粒进行重组的能力;")
81
部分cosmid vector c2XB pHC79 pHS262 pJC74 pJC75-58 pJC74m pJC720 Cosmid
载体 复制子 大小(kb) 选择标记 克隆位点 克隆能力(kb) c2XB pMB1 6.8 Ampr, Kanr BamHI, ClaI, EcoRI, HindIII, PstI, SmaI 32~45 pHC79 6.4 Ampr , Tetr EcoRI, HindIII, SalI, BamHI, PstI, CalI 29~46 pHS262 ColE1 2.8 BamHI, EcoRI, HincII 34~50 pJC74 15.8 Ampr EcoRI, BamHI, BglII, SalI 21~37 pJC75-58 11.4 EcoRI, BamHI, BglII 16~42 pJC74m 21 BamHI 16~32 pJC720 7.1 Elimm, Rifr HindIII, XmaI 11~28
 选择标记. 克隆位点. 克隆能力(kb) c2XB. pMB Ampr, Kanr. BamHI, ClaI, EcoRI, HindIII, PstI, SmaI. 32~45. pHC Ampr , Tetr. EcoRI, HindIII, SalI, BamHI, PstI, CalI. 29~46. pHS262. ColE BamHI, EcoRI, HincII. 34~50. pJC Ampr. EcoRI, BamHI, BglII, SalI. 21~37. pJC EcoRI, BamHI, BglII. 16~42. pJC74m. 21. BamHI. 16~32. pJC Elimm, Rifr. HindIII, XmaI. 11~28.")
82
Cosmid 载体 复制子 大小(kb) 选择标记 克隆位点 克隆能力(kb) pJC81 pMB1 7.1 Ampr, Tetr KpnI,BamHI,HindIII,SalI 30~46 pJB8 ColE1 5.4 Ampr BamHI,HindIII,SalI 31~47 MuA-3 4.8 PstI,EcoRI,BalI,PvuI, PvuII 32~48 MuA-10 EcoRI,BalI,PvuI,PvuI pTL5 5.6 BglII,BalI,HpaI pMF7 EcoRI,SalI

83
(4)cosmid cloning 应用cosmid载体在大肠杆菌中克隆大片端的真核基因组DNA技术,叫“柯斯克隆”(cosmid cloning)。 ①理论依据: 包装识别。 cos位点: “多联体”复制: 噬菌体的生命周期中,会产生数百个DNA通过cos位点连接的“多联体”分子。

84
Ter体系: 在包装的时候, 噬菌体具有位点特异的末端酶(terminase)体系(Ter体系),识别cos位点,把多联体切成单个DNA长度。 包装限制: 两个cos位点之间,必须保持38—45kb的DNA,Ter体系才能识别。
体系(Ter体系),识别cos位点,把多联体切成单个DNA长度。 包装限制: 两个cos位点之间,必须保持38—45kb的DNA,Ter体系才能识别。")
85
(5)cosmid克隆的一般过程 ①用特定的核酸内切酶消化真核生物DNA。 ②用同样的内切酶消化cosmid载体。 ③连接 产物中将有一定比例的分子是两端各有一个cos位点、长度40kb左右的真核DNA与载体的连接物。

86
④ Ter体系识别并切割 切割合适长度的杂种DNA连接物,并包装入噬菌体头部。 ⑤感染大肠杆菌 注入到细菌体内的杂种DNA分子环化,并按质粒的方式复制。

89
大质粒!

90
(7)cosmid载体克隆的缺点 ①载体片断自我连接。 ②外源DNA片断自我连接。 ③多个本来不在一起的外源DNA片断连接起来同时插入载体。 克服载体自体连接的改良方案: 用碱性磷酸酶除去线性质粒片断5’端的磷酸,可防止载体自体连接。
cosmid载体克隆的缺点 ①载体片断自我连接。 ②外源DNA片断自我连接。 ③多个本来不在一起的外源DNA片断连接起来同时插入载体。 克服载体自体连接的改良方案: 用碱性磷酸酶除去线性质粒片断5’端的磷酸,可防止载体自体连接。")
91
(7)cosmid载体的改良 ① Ish-Horowice—Burke改良方案 pJB8 cosmid载体 1981年,D. Ish-Horowice和J.F. Burke 。 1)突出的特点: 在BamH I位点两侧各有一个EcoR I切点。 使克隆在BamH I上的外源DNA可被EcoR I切下来。

92
2)Ish-Horowice—Burke方案克隆过程
SalI HindIII HindIII SalI T4 ligase

93
2)Ish-Horowice—Burke方案的缺点
a. 需要脱磷酸化处理。 防止载体自我连接 b. 电泳纯化载体两臂。 第二次酶切后。 c. 载体两臂的最终得率少的可怜!

94
② Bates---Swift 改良方案 c2XB cosmid载体 1983年P.F. Bates和R.A. Swift 1)特点: a. 具有两个cos位点。 b. 一个SmaI切点把两个cos位点分开。 SmaI平端的连接效率低,阻止了载体臂的自我连接,增加了外源DNA的连接效率。 c. BamHI克隆位点。

95
Bates---Swift 克隆过程:

96
5.2.3 M13 噬菌体克隆载体 (2)DNA长度 (3)DNA提纯 (1) M13 噬菌体的生物特性
+单链 DNA, 6407 bp。 (3)DNA提纯 RF dsDNA在寄主细胞内以高拷贝形式存在。 成熟的噬菌体里只包装有 + DNA,也容易提取。
DNA提纯. RF dsDNA在寄主细胞内以高拷贝形式存在。 成熟的噬菌体里只包装有 + DNA,也容易提取。")
97
M13序列

98
+DNA +DNA +DNA -DNA +DNA mRNA M13外壳蛋白
M13通过雄性细菌的F性须注入其+DNA +DNA 复制 形成RF dsDNA 200个 RF DNA +DNA -DNA 转录 指导合成 +DNA mRNA 翻译 组装成子代M13 M13外壳蛋白 每个细胞可以放出约1000个M13颗粒!

99
(5)没有包装限制 M13的包装过程: M13基因V编码单链DNA结合蛋白 RF dsDNA指导合成的+DNA DNA-蛋白质复合物 转移到寄主的细胞膜 基因V蛋白脱落 M13外壳蛋白 包装 溢出细胞膜 +DNA

100
3. M13载体的构建: (1)克隆区域的选定 ① 基因间隔区(intergenic region, IG区) M13基因组中只有基因II与基因IV之间存在一段507bp的基因间隔区。 ② 酶切位点 IG区内只有一个 BsuI 切点。 其余9个BsuI限制性位点分布在其它部位。

101
M13 J. Messing证明,IG区存在M13的复制起点,但可以插入外源DNA而不影响M13噬菌体的活力。

102
(2)加入酶切位点 ① 在IG区内加入单一内切酶位点 第一个M13载体:M13mp1: 在IG区内插入一个大肠杆菌的LacZ’(-肽序列)。 利用-肽序列中的三个单一酶切位点(Bgl II、Ava II 和 Pvu I)。
。 利用-肽序列中的三个单一酶切位点(Bgl II、Ava II 和 Pvu I)。 .")
103
M13 RF E.coli lac基因的HindII片断(lacI、lacP、lacO、lacZ’) BsuI不完全消化 各种长度的线性片断 (其中应有只在IG上切开的线性全长M13) 连接 M13mp1 JM101宿主 只有在IG区插入lacZ’才能存活并在Xgal上出现互补的蓝噬菌斑

104
(3)M13载体系列 ① M13载体系列的命名 M13mpn n代表系列数字 ② 对M13mp1的改进 加上常用的酶切位点。 M13mp2: B. Gronenborn和J. Messing1978年把LacZ’ 5’端的第13个核苷酸G突变成A,产生了一个EcoR I切点。

105
5’ATGACCATGATTACGGATTCA-
M13mp1 用N-甲基-N-亚硝基脲 把G甲基化 Me 5’ATGACCATGATTACGGATTCA- 转染E.coli。mG与T配对,复制两次后 M13mp1 5’ATGACCATGATTACGAATTCA- EcoRI切点 用EcoRI切 选择能切开的 线性分子 M13mp2 再环化

106
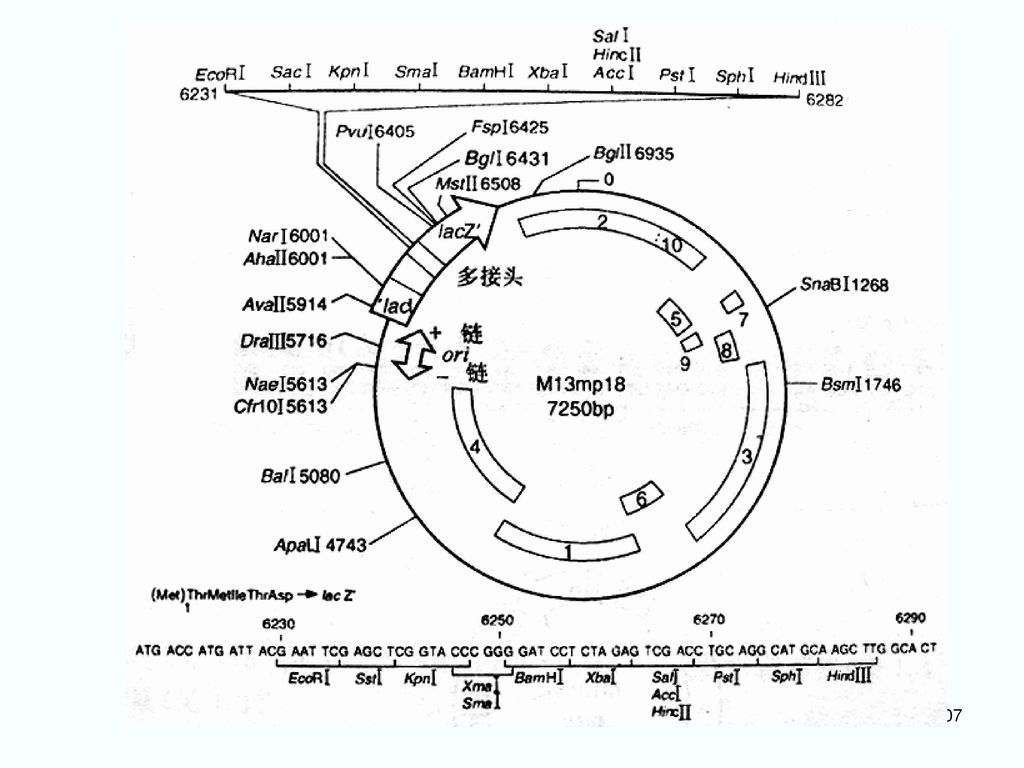
③ 对M13mp2的进一步改进 成为M13mp系列载体: 在M13mp2的LacZ’的5’端加上一段人工合成的多克隆位点(MCS)。 M13mp7、M13mp8、M13mp9、M13mp10、M13mp11、M13mp18、M13mp19 对应有相同MCS的pUC系列载体: pUC7、pUC8、pUC9、pUC10、pUC11、pUC18、pUC19

109
M13mp18的多克隆位点 与pUC18相同

110
4. M13系列载体的优点 (1)有MCS,便于克隆不同的酶切片段 (2) Xgal显色反应,可供直接选择 (3)无包装限制,克隆能力大 (4)可以克隆双链DNA分子中的每一条链 子代M13噬菌体中包含的是单连+DNA。
无包装限制,克隆能力大. (4)可以克隆双链DNA分子中的每一条链. 子代M13噬菌体中包含的是单连+DNA。 .")
111
MCS + M13 vector - 不同的酶切 + + - - 插入 不同的酶切 非编码链 外源DNA 编码链

112
M13 RF - + 外源DNA - + - + 转染E. coli + + 成熟的子代M13中

113
5. M13载体的缺点 ① 插入外源DNA后,遗传稳定性显著下降 ② 实际克隆能力小于1500bp。

114
噬菌粒载体(phagemid vectors)
由质粒载体与单链噬菌体载体的复制起点结合而成的新型载体系列。 MCS lacZ’ lacI 噬菌体ori 质粒ori Ampr

115
(1)噬菌粒载体的特点 ①分子量小 约3000bp(比M13小) ②克隆能力大 能插入10kb的外源DNA。 ③两种复制形式 既具有质粒的复制起点,又具有噬菌体的复制起点。 既能在大肠杆菌中以质粒的形式双链复制,又能在噬菌体内进行单链复制。

116
(2)常见的噬菌粒载体 辅助噬菌体用来复制和包装噬菌粒载体。 噬菌粒载体 质粒部分 单链噬菌体部分 辅助噬菌体 大肠杆菌寄主 pEMBL8
pUC8 f1 IR1 71/18 pRSA101 VX M13 M13变异株 XS127,XS101 pUC118/pUC119 pUC18/ pUC19 M13K07 MV1184 pBS pUC XL1-Blue 辅助噬菌体用来复制和包装噬菌粒载体。

117
(3)pUC118/pUC119 ① 构成 1)M13的基因间隔区(IG) 带有M13复制起点。 2)pUC18/pUC19质粒载体: 质粒复制起点 Ampr lacZ’ MCS
pUC118/pUC119 ① 构成 1)M13的基因间隔区(IG) 带有M13复制起点。 2)pUC18/pUC19质粒载体: 质粒复制起点 Ampr lacZ’ MCS")
118
pUC118/119 MCS lacZ’ lacI M13 ori pUC18/pUC19 ori 3.2kb Ampr

119
② pUC118/119的复制模式 两种不同的复制模式 1)双链质粒复制模式 受pUC本身的复制起点(来源于ColE1)的控制。 每个细胞能达到500个拷贝。
双链质粒复制模式 受pUC本身的复制起点(来源于ColE1)的控制。 每个细胞能达到500个拷贝。")
120
来源于M13的复制起点被辅助噬菌体的基因II产物控制。
2)单链滚环噬菌体复制模式 当寄主细胞被辅助噬菌体M13感染后。 来源于M13的复制起点被辅助噬菌体的基因II产物控制。 复制蛋白 包装 辅助M13 外壳蛋白
单链滚环噬菌体复制模式. 当寄主细胞被辅助噬菌体M13感染后。 来源于M13的复制起点被辅助噬菌体的基因II产物控制。 复制蛋白. 包装. 辅助M13. 外壳蛋白.")
121
③ 噬菌粒载体的包装 如M13K07等。 1)辅助噬菌体: 自身的复制起点发生了突变,复制效率低下,但感染细菌后能表达出外壳蛋白和复制蛋白,帮助噬菌粒载体复制。 2)包装: 噬菌体 外壳蛋白和 复制蛋白 辅助噬菌体 噬菌粒载体 ssDNA

122
(4)pBluescript 噬菌粒载体(pBS)
Stratagene公司发展的一类噬菌粒载体。 ① 组成 由pUC质粒载体、f1噬菌体的复制起点和T3、T7噬菌体的启动子组成。 ② 特点 1)定向体外转录 MCS两侧分别加上T3和T7噬菌体的启动子。加入适当的噬菌体RNA聚合酶,就可以定向体外转录。
定向体外转录. MCS两侧分别加上T3和T7噬菌体的启动子。加入适当的噬菌体RNA聚合酶,就可以定向体外转录。")
123
2)两种复制模式 既能以质粒的形式复制,又能以噬菌体的形式复制包装。 3)选择方便 有Ampr和lacZ’,可以进行Amp抗性选择和Xgal-IPTG蓝白斑选择。 4)插入方便 18个单一酶切位点的MCS

124
1)pBluescript SK/KS(+/-)区分:
pBluescript SK(+/-)/pBluescript KS(+/-) 1)pBluescript SK/KS(+/-)区分: SK:表示lacZ’的转录方向是沿MCS上的 SacI KpnI KS:反向( KpnI SacI ,MCS相反) +(f1+):单链复制起始方向背离 lacZ’,能回收lacZ’的编码链(+) -(f1-):能回收lacZ’的非编码链(-)
/pBluescript KS(+/-) 1)pBluescript SK/KS(+/-)区分: SK:表示lacZ’的转录方向是沿MCS上的. SacI KpnI. KS:反向( KpnI SacI ,MCS相反) +(f1+):单链复制起始方向背离. lacZ’,能回收lacZ’的编码链(+) -(f1-):能回收lacZ’的非编码链(-)")
125
pBS SK+

126
pBS KS-

129
PCR产物克隆载体—— T 载体 pCR系列载体 Invitrogen公司开发的线性噬菌粒载体。 ① 结构 pUC的质粒部分、fi噬菌体ori、Kanr和Ampr抗性。 ② 特点 MCS的中部已经切开,各有一个3’端突出的T。 PCR产物往往在3’端突出一个A,所以能与这个载体直接连接。

130
pCR2.1

131
pCR2.1的MCS 克隆的PCR产物能被两侧的EcoRI切下来回收。

132
Promega 公司的T载体系列 pGEM-T vector

133
pTarget vector G418抗性 可在真核生物表达 外显子捕捉

134
5.3 大分子DNA克隆载体 5.3.1 细菌人工染色体(BAC) 1. F质粒的特点:
H. Shizuya等1992年在大肠杆菌F因子的基础上构建的。 1. F质粒的特点: (1)单拷贝复制。 (2)克隆容量可达300kb。 (3)比较稳定。每细胞只有一个拷贝,不会重组。 (4)便于提纯;像提取质粒一样直接提纯。
单拷贝复制。 (2)克隆容量可达300kb。 (3)比较稳定。每细胞只有一个拷贝,不会重组。 (4)便于提纯;像提取质粒一样直接提纯。")
135
2. BAC的组成结构 (1)OriS和repE控制质粒的单向复制。 (2)parA和parB维持低水平质粒拷贝数。
(3)CosN是噬菌体末端酶专一性切割位点。 (4)loxP是P1 Cre蛋白作用位点。 (5)HindIII和BamHI是插入位点。 (6)两侧的NotI可用于切下外源DNA。 (7)氯霉素抗性基因CMR
CosN是噬菌体末端酶专一性切割位点。 (4)loxP是P1 Cre蛋白作用位点。 (5)HindIII和BamHI是插入位点。 (6)两侧的NotI可用于切下外源DNA。 (7)氯霉素抗性基因CMR.")
137
5.3.2 酵母人工染色体 (yeast artificial chromosome, YAC)
酵母人工染色体 (yeast artificial chromosome, YAC) A.W.Murray 1983年形成基础理论, D.T.Burke等1987年变为现实。 1. YAC的特点: (1)只能在酵母细胞中扩增。 (2)克隆容量大,为230kb-1700kb。 (3)构建原理,按染色体结构构建。
 A.W.Murray 1983年形成基础理论, D.T.Burke等1987年变为现实。 1. YAC的特点: (1)只能在酵母细胞中扩增。 (2)克隆容量大,为230kb-1700kb。 (3)构建原理,按染色体结构构建。")
138
2. 染色体复制和遗传的三个基本组件: (1)DNA复制起始点(ARS) (2)着丝粒(centromere,cen) (3)两个端粒(telomeres,tel) TEL CEN ARS
DNA复制起始点(ARS) (2)着丝粒(centromere,cen) (3)两个端粒(telomeres,tel) TEL CEN ARS")
139
3. YAC的组成结构 (1)着丝粒区(CEN) 酵母染色体着丝粒区的保守序列: 78-86bp I II III 由三个区组成 A A
GTCACGTG T TGTTTCTGNTTTCCGAAA 78-86bp I II III 由三个区组成

140
酵母第3、4、6、11号染色体的着丝粒区序列:

141
(2)端粒(TEL) 两个端粒序列Tel。 酵母端粒保守序列是(G4T2)n 重复序列。 人是TTAGGG重复序列; 四膜虫是 GGGTTG重复序列。 (3)复制起点(ORI) 约100bp的自主复制序列(ARS)。 (真核生物中只有酵母菌有ARS)
")
142
(4)克隆位点 位于 SUP4 基因内部。 SUP4酶失活的酵母菌落呈红色;不失活的菌落是白色。 插入失活选择:
与 YAC 载体配套的宿主酵母菌(如AB1380)的胸腺嘧啶合成基因带有一个赭石突变 ade 2-1。有这个突变的酵母菌在基本培养基上形成红色菌落,当带有赭石突变抑制基因 sup4 的载体存在于细胞中时,就抑制 ade 2-1基因的突变效应,形成白色菌落。利用这一菌落颜色转变的现象,来筛选载体中含有外源 DNA 片段插入的重组子。
的胸腺嘧啶合成基因带有一个赭石突变 ade 2-1。有这个突变的酵母菌在基本培养基上形成红色菌落,当带有赭石突变抑制基因 sup4 的载体存在于细胞中时,就抑制 ade 2-1基因的突变效应,形成白色菌落。利用这一菌落颜色转变的现象,来筛选载体中含有外源 DNA 片段插入的重组子。")
143
(5)在酵母中的标记基因 TRP1、URA3(分别位于两臂)。 (6)在细菌中操作 细菌的复制起点 ori和抗生素标记Ampr
在酵母中的标记基因 TRP1、URA3(分别位于两臂)。 (6)在细菌中操作 细菌的复制起点 ori和抗生素标记Ampr")
144
4. YAC克隆外源DNA

145
5. YAC转化过程 (1)宿主酵母菌: AB1380 trp- 和 ura- (2)选择方式 cen trp1 ura3
只有同时得到YAC的两个臂,才能在基本培养基(不加TRP和URA)上生长。 cen trp1 ura3
上生长。 cen. trp1. ura3.")
147
6. YAC的缺点: (1)存在插入子的稳定性问题。 (2)同一酵母细胞内多个YAC引起交换 (3)转化效率低。 (4)难以制备纯的YAC-DNA。
存在插入子的稳定性问题。 (2)同一酵母细胞内多个YAC引起交换 (3)转化效率低。 (4)难以制备纯的YAC-DNA。")
148
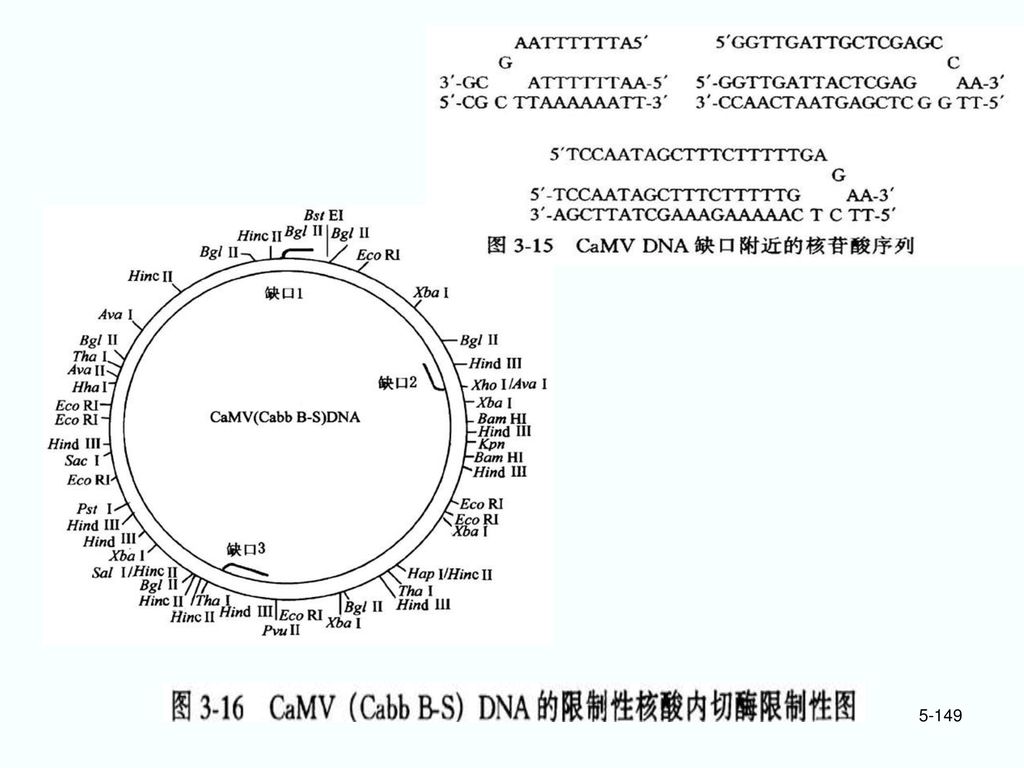
5.4 病毒载体 5.41 CaMV克隆载体 花椰菜花叶病病毒(CaMV)环状双链DNA分子的植物病毒。
5.4 病毒载体 5.41 CaMV克隆载体 花椰菜花叶病病毒(CaMV)环状双链DNA分子的植物病毒。 1. 基因:CaMV DNA分子大小:~8kb 2. 结构:在病毒颗粒中呈开环形式,负链有一缺口,正链有两缺口。进入植物细胞后单链部分被酶解,缺口自行闭合,成为环状DNA分子。
环状双链DNA分子的植物病毒。 1. 基因:CaMV DNA分子大小:~8kb. 2. 结构:在病毒颗粒中呈开环形式,负链有一缺口,正链有两缺口。进入植物细胞后单链部分被酶解,缺口自行闭合,成为环状DNA分子。")
150
CaMV基因区 8个阅读框(ORF)和3个间隔区(IR1~3) IR1- ORF VI与ORF VII之间 IR2- ORF VII与ORF I之间 IR3- ORF V与ORF VI之间 IR1 和IR3区各有一个启动子结构,分别启动同方向转录35S RNA和19S RNA,而两者的终止子都在IR1区。 35S启动子——组成型强启动子。组装了35S启动子的外源基因可在植物细胞高效表达,在蓝藻细胞内也有效表达。

151
CaMV的增殖和感染 1、增殖途径: Ca MV颗粒→蚜虫→病毒DNA进入植物细胞核→ 转录19S RNA→翻译 35S RNA 翻译 反转录 “-”链DNA →复制出“+”链 2、Ca MV-DNA可转染植物细胞 在ORF II和ORF VII区插入外源DNA后仍能转染植物细胞 包装成新的 Ca MV颗粒

152
3. 构建Ca MV克隆载体的基本策略和途径 设计思路:CaMV-DNA能侵入植物细胞而将目的基因导入,并且清除CaMV对植物的致病性基因。
(2)构建混合型克隆载体 CaMV-DNA →合适的限制性酶切→组入Ti的T-DNA区→完成构建 (3)构建CaMV35S启动子融合基因克隆载体系统 将35S启动子与目的基因融合,高效表达目的基因。
构建混合型克隆载体. CaMV-DNA →合适的限制性酶切→组入Ti的T-DNA区→完成构建. (3)构建CaMV35S启动子融合基因克隆载体系统. 将35S启动子与目的基因融合,高效表达目的基因。")
154
5.4.2 烟草花叶病毒(TMV)克隆载体 基因组:烟草花叶病毒(TMV)的是一种单链的正义RNA,6395nt。
特点:至少编码四种多肽:两种复制酶亚单位,130kD和180kD;一种30kD外壳蛋白和一种与细胞间运动有关的重要蛋白。 外壳蛋白表达量高,寄主范围广,能在整株植物上快速扩散 TMV载体构建策略

155
构建TMV载体的策略 表面展示 插入型 替换型 运动蛋白缺失突变型 互补型 外壳蛋白缺失突变型 复制酶缺失突变型

156
5.4.3 SV40 克隆载体 1. SV40基因组:双链环状DNA:5243bp 早期转录区:一个基因(2个蛋白质 t,T)
晚期转录区:先转录出一个多顺反子mRNA VP1 mRNA VP2 mRNA VP3 mRNA VP3比VP2N端少1/3 AUG 位于VP2和 VP3内

157
T - 抗原和t-抗原mRNA前体的不同剪接方式
UAA 内含子1 内含子2 前体mRNA 剪接方式1:去除内含子1 剪接方式2:去除内含子2 T-mRNA T- 抗原 t-抗原 分布于核内:100 KD 分布于胞浆内:18 KD t -mRNA

158
1.SV40载体 SV40病毒的基因组是环状双链DNA,大小仅为5243bp,与质粒大小相似,适于基因操作.
SV40在感染敏感宿主细胞后,其基因组DNA进入细胞核,首先启动早期基因转录,在细胞质中表达出早期基因t抗原和T抗原,当这2种蛋白质积累到一定程度时DNA开始复制,同时开始驱动病毒晚期基因转录,翻译产生VP1,VP2和VP3等病毒衣壳蛋白,并开始进行病毒粒子的装配.当病毒粒子数量积累到一定量时,细胞裂解释放出子代病毒粒子.

159
外源DNA是直接插入在缺陷性的病毒基因组中. 根据置换位置不同分为: ①早期置换型 (2)重组的病毒-质粒载体
● SV40病毒不能包装大于SV40基因组的DNA分子,因此,外源基因只能通过取代病毒本身的DNA片段进行克隆. ● 以SV40病毒为基础的克隆载体主要有2种不同类型: (1)置换行重组病毒载体 外源DNA是直接插入在缺陷性的病毒基因组中. 根据置换位置不同分为: ①早期置换型 ② 晚期置换型 (2)重组的病毒-质粒载体 载体只保留复制序列,无衣壳蛋白,不能产生病毒粒子,但可进行复制并产生很多拷贝,所以依然可以高水平表达外源基因.
置换行重组病毒载体. 外源DNA是直接插入在缺陷性的病毒基因组中. 根据置换位置不同分为: ①早期置换型. ② 晚期置换型. (2)重组的病毒-质粒载体. 载体只保留复制序列,无衣壳蛋白,不能产生病毒粒子,但可进行复制并产生很多拷贝,所以依然可以高水平表达外源基因.")
160
2.SV40-DNA复制与增殖 3. 构建策略和途径 猿猴细胞——受纳细胞 嚙齿动物(小鼠、仓鼠)——非受纳细胞,导致细胞转化
人:1~2%细胞产生病毒颗粒,不会插入染色体,相对安全 3. 构建策略和途径 包装严格,不能包装大于SV40-DNA的分子,只能构建取代型载体或SV40-DNA与质粒DNA的重组型载体 (1)取代型载体 被取代的DNA序列不能超过SV40-DNA的30% 1)晚期转录区取代型:需辅助载体共同转染受体细胞 2)早期转录区取代型:辅助细胞系 (将SV40早期转录区DNA序列整合到敏感细胞基因组上)
取代型载体. 被取代的DNA序列不能超过SV40-DNA的30% 1)晚期转录区取代型:需辅助载体共同转染受体细胞. 2)早期转录区取代型:辅助细胞系. (将SV40早期转录区DNA序列整合到敏感细胞基因组上)")
161
质粒载体的衍生物,利用SV40元件和质粒载体的元件构建的适合在真核细胞表达的载体。
(2)穿梭质粒载体 质粒载体的衍生物,利用SV40元件和质粒载体的元件构建的适合在真核细胞表达的载体。 病毒 -质粒重组克隆载体(穿梭质粒) :含SV40完整早期转录区和DNA复制起始点,及质粒DNA的复制起始点和选择性标记。
穿梭质粒载体. 质粒载体的衍生物,利用SV40元件和质粒载体的元件构建的适合在真核细胞表达的载体。 病毒 -质粒重组克隆载体(穿梭质粒) :含SV40完整早期转录区和DNA复制起始点,及质粒DNA的复制起始点和选择性标记。")
162
反转录病毒载体 1.反转录病毒(ritrovirus) 属于正链RNA病毒,显著的特点是具有2倍体基因组。 两个相同的RNA分子在5’端附近经氢键连接形成70S RNA,这种结构可能在逆转录过程中起调节作用。
 属于正链RNA病毒,显著的特点是具有2倍体基因组。 两个相同的RNA分子在5’端附近经氢键连接形成70S RNA,这种结构可能在逆转录过程中起调节作用。")
163
反转录病毒结构

164
Influenza virus HIV

165
Moloney murine leukemia virus (Moloney MLV)
2. 反转录病毒的基因组结构 类似于真核细胞mRNA 3’ cap U3 R U5 + gag pol env U3 R U5 polyA 5’ LTR LTR Moloney murine leukemia virus (Moloney MLV) LTR: 长末端重复序列。在病毒基因组整合中至关重要。整合时,整合酶在U5-U3连接处切开双链DNA,随机插入到宿主基因组里。 LTR本身还具有启动子和polyA加尾信号的功能。
 LTR: 长末端重复序列。在病毒基因组整合中至关重要。整合时,整合酶在U5-U3连接处切开双链DNA,随机插入到宿主基因组里。 LTR本身还具有启动子和polyA加尾信号的功能。")
166
5’ 3’ U3 R U5 + gag pol env U3 R U5 LTR LTR U5、U3:5’段或3’端独特序列。
cap U3 R U5 + gag pol env U3 R U5 polyA LTR LTR U5、U3:5’段或3’端独特序列。 +:组装时必需的非编码序列。 env:外壳蛋白。 pol:反转录酶和整合酶。 gag:核心蛋白。

167
3. 反转录病毒载体构建 (1)删除pol、env和gag基因。 (2)插入标记基因(如neor)。 (3)外源基因和启动子插入到+下游。
5’LTR + 启动子 外源基因 neor 3’LTR

168
4.反转录病毒载体的包装 (1)包装细胞系的构建 反转录病毒DNA转化细胞的效率很低,必须包装成病毒颗粒转染细胞。 用两个缺失了+的反转录病毒染色体转化细胞,以制造病毒外壳蛋白。 5’LTR gag 3’LTR 5’LTR pol env 3’LTR

169
5’LTR gag 3’LTR 5’LTR pol env 3’LTR
包装细胞 5’LTR gag 3’LTR 5’LTR pol env 3’LTR 病毒外壳蛋白

170
转入的载体DNA上带有+,转录成的载体RNA就会被包装细胞中原有的病毒外壳蛋白识别包装成重组病毒颗粒。
5’LTR + 外源基因 neor 3’LTR 高效转染靶细胞

171
5. 反转录病毒载体的优点 (1)将DNA插入宿主基因组的随机位点上。 (2)侵染范围广、感染率高。 (3)整合的原病毒在宿主基因中比较稳定, 拷贝数目较低。 (4)包装的外源DNA可达10kb。 (5)反转录病毒感染哺乳动物细胞,对宿主 细胞没有毒性。 反转录病毒一般只感染处在分裂状态的细胞。
反转录病毒感染哺乳动物细胞,对宿主. 细胞没有毒性。 反转录病毒一般只感染处在分裂状态的细胞。")
172
腺病毒载体(adenovirus) 1. 腺病毒的基因组 线状双链DNA病毒,基因组中有14个基因。 共同特点是都带有倒置的末端重复(ITR),在病毒的复制过程中起非常重要的作用。
 1. 腺病毒的基因组 线状双链DNA病毒,基因组中有14个基因。 共同特点是都带有倒置的末端重复(ITR),在病毒的复制过程中起非常重要的作用。")
173
2. 腺病毒的优点 (1)比较安全,无致病、致癌、致畸作用。 (2)宿主范围广 不仅能感染有分裂能力的细胞,也能感染不能分裂的细胞(如神经细胞)。 (3)可以在呼吸道和肠道中繁殖, 可以通过口服或喷雾的方式感染。 (4)载体中的插入片断可达7.5kb (有包装容量限制)。 (5)腺病毒容易制备、纯化。 (6)基因组结构和功能了解得清楚。
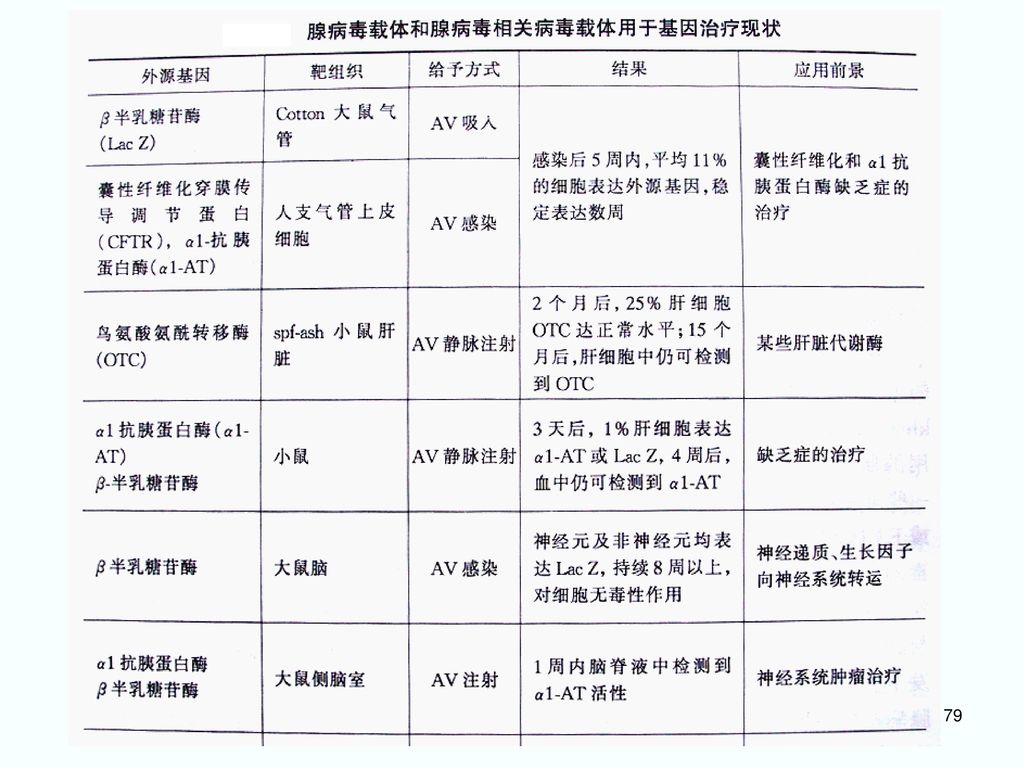
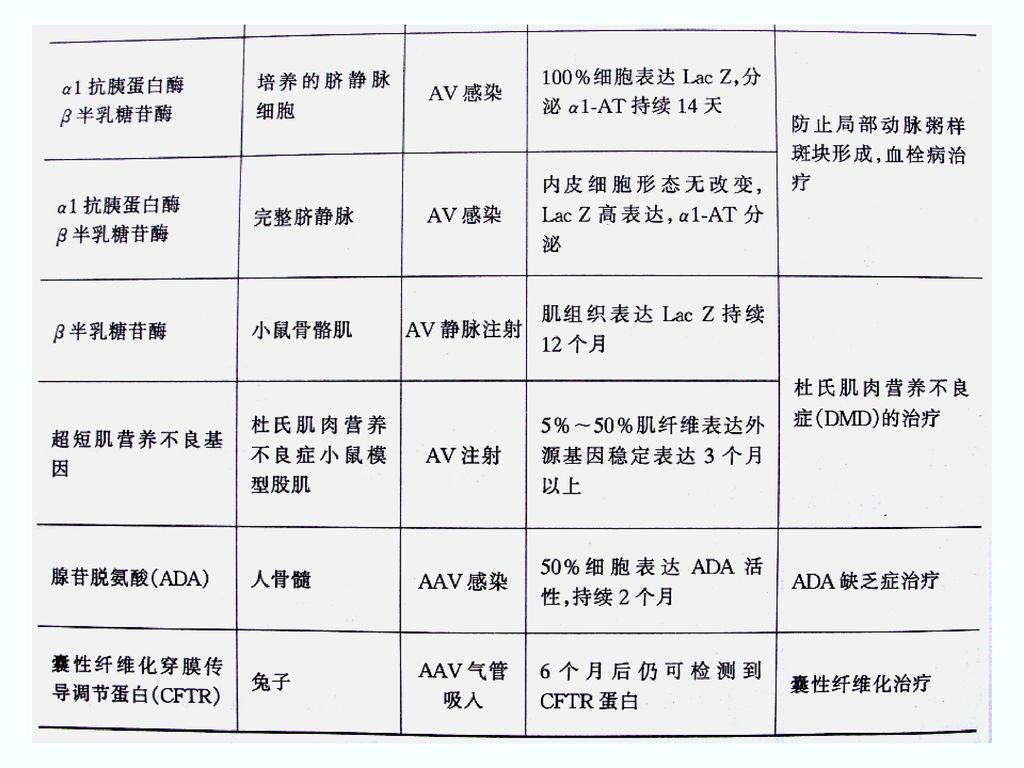
载体中的插入片断可达7.5kb. (有包装容量限制)。 (5)腺病毒容易制备、纯化。 (6)基因组结构和功能了解得清楚。")
174
3. 腺病毒载体 是目前最常用的基因治疗载体之一。 (1)腺病毒载体的构建 在人腺病毒C亚群的Ad2 和Ad5 的基础上构建。 E1区为病毒复制必需区,缺失则成为复制缺陷型载体。 E3区是病毒复制非必需区,可用于外源基因的插入。

175
(1)腺病毒载体的构建 用同源重组的方法插入外源基因。 ①删除腺病毒DNA一些非必需区 如E1或E3区,增加插入能力。 E1或E3缺失病毒。
ITR E1 E3

176
②构建质粒载体 含有E1或E3区两侧的同源序列。 在同源序列之间插入外源基因和选择标记基因。

177
③把质粒切成线性 ④共转染细胞 质粒与病毒DNA(缺失E1或E3) 在细胞中发生同源重组,把外源DNA加到病毒基因组中,并包装成重组病毒颗粒。
 在细胞中发生同源重组,把外源DNA加到病毒基因组中,并包装成重组病毒颗粒。")
178
4. 重组腺病毒DNA在细胞中的生存 (1)以游离的附加体形式存在于细胞中 不能整合到细胞基因组中。基因表达时间短。 (2)E1是腺病毒繁殖的必需区 缺失E1的重组腺病毒必须在已经被腺病毒(辅助病毒)感染的细胞中繁殖。 (3)E3区对腺病毒的繁殖不是必需的 缺失E3的重组腺病毒不需要辅助病毒。
E3区对腺病毒的繁殖不是必需的. 缺失E3的重组腺病毒不需要辅助病毒。")
181
5. 腺病毒载体的优点 ①腺病毒粒子相对稳定,病毒基因组重排频率低,外源基因片段插入片段在病毒复制几个周期后仍可保持不变,易于用重组DNA技术操作; ②安全性较好,腺病毒无需整合进宿主细胞基因组中,目的基因在宿主细胞基因组外游离状态下表达,整合突变致癌可能性小,基因毒性低; ③腺病毒基因组较大(36 kb) ,绝大多数基因组(约35 kb)均能被外源基因取代,插入大片段外源性基因的潜力大; ④有较大的宿主范围,可以感染肝细胞、血管内皮细胞等多种人体细胞,对于受体细胞是否处于分裂期要求不严格; ⑤腺病毒载体容易将外源基因直接转移到靶细胞中,并有效表达成活性蛋白。
 ,绝大多数基因组(约35 kb)均能被外源基因取代,插入大片段外源性基因的潜力大; ④有较大的宿主范围,可以感染肝细胞、血管内皮细胞等多种人体细胞,对于受体细胞是否处于分裂期要求不严格; ⑤腺病毒载体容易将外源基因直接转移到靶细胞中,并有效表达成活性蛋白。")
182
6. 腺病毒载体应用于基因治疗 在美国近三分之一的基因治疗临床试验中采用腺病毒载体,其中5型腺病毒的使用最为广泛。具有基因治疗的安全性和有效性。 根据美国国家卫生院的生物技术活动办公室统计,在过去的20年里,已进行了将近144例使用腺病毒载体的基因治疗临床试验,近1/5引入了腺病毒载体。 2004年1月20日,我国拥有自主知识产权的重组腺病毒p53注射液(深圳赛百诺基因技术有限公司)获得国家食品药品监督管理局的生产文号。
获得国家食品药品监督管理局的生产文号。")
183
双链线性DNA病毒,基因组大小在180kb左右,其中有30kb左右为非必需区,可以被约25kb的外源DNA片段取代而不影响病毒的复制与增殖.

184
5.4.7 痘苗病毒载体(Vaccinia virus)
1. 痘病毒概况 为最大的一类DNA病毒,结构复杂。 病毒粒呈砖形或椭圆形,大小(300~450)×(170~260)纳米,有核心、侧体和包膜,核心含有与蛋白结合的病毒DNA。 病毒粒中有30种以上的结构蛋白和几种酶,核心蛋白中含依赖于DNA的RNA多聚酶。 病毒在细胞质内增殖,形成包涵体,病毒粒,由细胞裂解而释放。
×(170~260)纳米,有核心、侧体和包膜,核心含有与蛋白结合的病毒DNA。 病毒粒中有30种以上的结构蛋白和几种酶,核心蛋白中含依赖于DNA的RNA多聚酶。 病毒在细胞质内增殖,形成包涵体,病毒粒,由细胞裂解而释放。")
185
2.痘苗病毒的基因组 双链线性DNA病毒, 基因组大小在180kb左右,其中有30kb左右为非必需区,可以被约25kb的外源DNA片段取代而不影响病毒的复制与增殖。 在每个DNA 分子末端有一个发夹环结构。

186
3. 痘病毒分类 根据脊椎动物和昆虫宿主范围,将痘病毒科分为两个亚科:脊椎动物痘病毒亚科和昆虫痘病毒亚科。 迄今为止,还未见任何昆虫痘病毒载体的报道。 脊椎动物痘病毒亚科包括8个属: 禽痘病毒属、羊痘病毒属、野兔痘病毒属、拟软体动物痘病毒属、正痘病毒属、副痘病毒属、猪痘病毒属和亚塔痘病毒属。

187
4. 痘病毒载体的特点: *容易培养,相当稳定,能在大多数哺乳动物细胞中复制繁殖。 *由于其基因组很大,因此只能通过先将目的基因克隆到转移载体上,然后将充足转移载体与野生性痘苗病毒DNA工转染哺乳动物细胞,在细胞内发生同源重组而获得重组痘苗病毒。 *高效,能同时预防集中病原微生物引起的传染病。

188
5. 痘病毒载体及其应用 痘病毒是研究最早最成功的载体病毒之一 (1) 痘病毒分子生物学领域的研究; (2) 体外生产和蛋白质功能化研究;
5. 痘病毒载体及其应用 痘病毒是研究最早最成功的载体病毒之一 (1) 痘病毒分子生物学领域的研究; (2) 体外生产和蛋白质功能化研究; (3) 作为活的疫苗或工具用于疫苗研究。 1982年,研究人员将外源基因插入痘苗病毒的TK基因中,首次成功地构建了在哺乳动物细胞中表达外源基因的重组痘苗病毒。
 痘病毒分子生物学领域的研究; (2) 体外生产和蛋白质功能化研究; (3) 作为活的疫苗或工具用于疫苗研究。 1982年,研究人员将外源基因插入痘苗病毒的TK基因中,首次成功地构建了在哺乳动物细胞中表达外源基因的重组痘苗病毒。")
189
5.5 基因打靶载体(Targeting vector)
1. 基因打靶(gene targeting) 通过DNA同源重组,使细胞特定的内源基因被破坏而造成其功能丧失或在基因组特定的位点插入外源基因。 基因敲出(knock-out) 基因敲入(knock-in)
 通过DNA同源重组,使细胞特定的内源基因被破坏而造成其功能丧失或在基因组特定的位点插入外源基因。 基因敲出(knock-out) 基因敲入(knock-in)")
190
2.基因打靶载体的要求 定点整合 1. 要求能整合到染色体上特定的位置。 2. 整合的位置尽量选在基因组内编码非必需产物的地方。 3. 必需整合在基因组中可以转录的区域。

191
新霉素(neomycin)磷酸转移酶基因
3. 基因打靶载体组成 1. 两段同源DNA序列(HB1和HB2) 与靶位点两端的区域同源 2. 目的基因 3. 编码抗G418的基因(neor) 新霉素(neomycin)磷酸转移酶基因 4. 胸苷激酶基因(HSV-tk1和HSV-tk2) 分别来自I型和II型单纯疱疹病毒(HSV)。 载体 tk1 HB1 neor 目的基因 HB2 tk2 载体
 与靶位点两端的区域同源. 2. 目的基因. 3. 编码抗G418的基因(neor) 新霉素(neomycin)磷酸转移酶基因. 4. 胸苷激酶基因(HSV-tk1和HSV-tk2) 分别来自I型和II型单纯疱疹病毒(HSV)。 载体. tk1. HB1. neor. 目的基因. HB2. tk2. 载体.")
192
tk1 HB1 neor HB2 tk2 HB1 HB2 tk1 HB1 neor HB2 tk2
4. 载体的整合方式 1. 非特异整合——非同源重组 载体 tk1 HB1 neor 目的基因 HB2 tk2 载体 染色体DNA HB1 染色体DNA HB2 染色体DNA 染色体 tk1 HB1 neor 目的基因 HB2 tk2 染色体 两个tk基因至少有一个可能整合到基因组里。细胞被tk基因杀死。

193
tk1 HB1 neor HB2 tk2 HB1 HB2 HB1 neor HB2
2.特异位点整合——同源重组 载体 tk1 HB1 neor 目的基因 HB2 tk2 载体 染色体DNA HB1 染色体DNA HB2 染色体DNA 染色体DNA HB1 neor 目的基因 HB2 染色体DNA 只有目的基因和neor基因整合进去,tk基因不会整合。细胞在G418中存活。

194
五、转染细胞的选择 正-负选择法: 1. 正选择(positive selection): 用G418进行选择。 没有整合进neor基因的细胞都被杀死。 2. 负选择(negative selection): 用9-鸟嘌呤(ganciclovir,GCV)。 表达胸苷激酶(tk)的细胞都被杀死(Tk能把GCV转化成有毒的化合物)。
。 表达胸苷激酶(tk)的细胞都被杀死(Tk能把GCV转化成有毒的化合物)。")
195
Thank You!

Similar presentations













:核蛋白体组成成分 转移 RNA ( tRNA ):转运氨基酸 信使 RNA ( mRNA ):蛋白质合成模板 不均一核 RNA ( hnRNA ):成熟 mRNA 的前体 小核 RNA ( snRNA ):>")


>")
DNA、RNA及蛋白质操作技术.>")

>")







 A.细菌的遗传物质主要是DNA B.病毒的遗传物质主要是RNA>")



