Download presentation
Presentation is loading. Please wait.

1
第五章 蛋白质的三维结构

2
主要内容 稳定蛋白质三维结构的作用力 多肽主链折叠的空间限制 蛋白质的二级结构 纤维状蛋白质 超二级结构和结构域 球状蛋白质与三级结构
第五章 蛋白质的三维结构 主要内容 稳定蛋白质三维结构的作用力 多肽主链折叠的空间限制 蛋白质的二级结构 纤维状蛋白质 超二级结构和结构域 球状蛋白质与三级结构 蛋白质折叠与结构预测 亚基缔合与四级结构

3
第一节 稳定蛋白质三维结构的作用力 1、维持三维结构的作用力 a 离子键 b 氢键 c 疏水相互作用 d 范德华力 e 二硫键

4
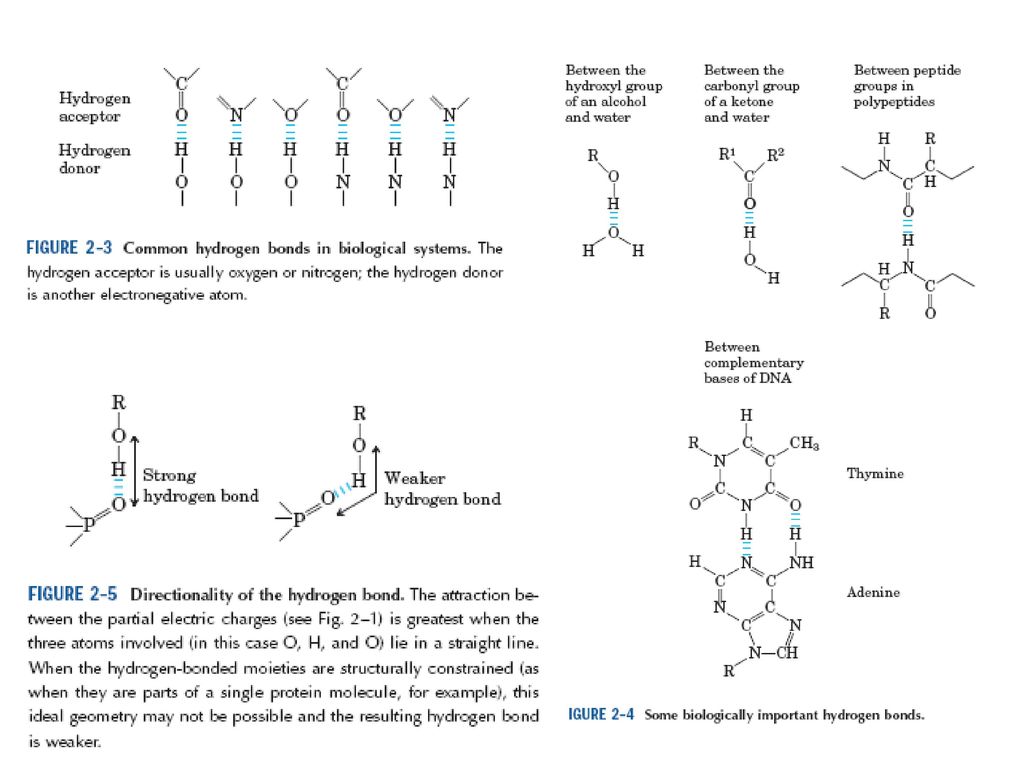
(一)氢键 氢键的贡献是协同蛋白质的折叠和帮助稳定球蛋白的天然构象。
第一节 稳定蛋白质三维结构的作用力 (一)氢键 氢键的贡献是协同蛋白质的折叠和帮助稳定球蛋白的天然构象。 多肽链骨架的羰基和酰胺基之间,常常形成氢键使肽链形成α-螺旋和β-折叠结构。 在多肽链骨架和水之间,多肽链骨架和极性侧链之间,两个极性侧链之间以及极性侧链和水之间也可以形成氢键。 大多数氢键都是N-H……O类型的。
氢键. 氢键的贡献是协同蛋白质的折叠和帮助稳定球蛋白的天然构象。 多肽链骨架的羰基和酰胺基之间,常常形成氢键使肽链形成α-螺旋和β-折叠结构。 在多肽链骨架和水之间,多肽链骨架和极性侧链之间,两个极性侧链之间以及极性侧链和水之间也可以形成氢键。 大多数氢键都是N-H……O类型的。")
6
(二)范德华力 广义范德华力包括定向效应、诱导效应和分散效应等。
第一节 稳定蛋白质三维结构的作用力 (二)范德华力 广义范德华力包括定向效应、诱导效应和分散效应等。 范德华力包括吸引力和斥力两种相互作用,范德华力只有当两个非键合原子之间处于一定距离时才能达到最大。 虽然范德华力相对来说比较弱,但由于范德华力相互作用数量大,并且具有加和性,因此范德华力是一种不可忽视的作用力。
范德华力. 广义范德华力包括定向效应、诱导效应和分散效应等。 范德华力包括吸引力和斥力两种相互作用,范德华力只有当两个非键合原子之间处于一定距离时才能达到最大。 虽然范德华力相对来说比较弱,但由于范德华力相互作用数量大,并且具有加和性,因此范德华力是一种不可忽视的作用力。")
7
(三)疏水作用 蛋白质中的疏水基团彼此靠近、聚集以避开水的现象称之疏水相互作用(hydrophobic interaction)。
第一节 稳定蛋白质三维结构的作用力 (三)疏水作用 蛋白质中的疏水基团彼此靠近、聚集以避开水的现象称之疏水相互作用(hydrophobic interaction)。 因为水分子彼此之间的相互作用要比水与其它非极性分子的作用更强烈,非极性侧链避开水聚集被压迫到蛋白质分子内部,而大多数极性侧链在蛋白质表面维持着与水的接触。 疏水相互作用在维持蛋白质构象中起着主要的作用,也是使蛋白质多肽链进行折叠的主要驱动力。
疏水作用. 蛋白质中的疏水基团彼此靠近、聚集以避开水的现象称之疏水相互作用(hydrophobic interaction)。 因为水分子彼此之间的相互作用要比水与其它非极性分子的作用更强烈,非极性侧链避开水聚集被压迫到蛋白质分子内部,而大多数极性侧链在蛋白质表面维持着与水的接触。 疏水相互作用在维持蛋白质构象中起着主要的作用,也是使蛋白质多肽链进行折叠的主要驱动力。")
8
第一节 稳定蛋白质三维结构的作用力 水分子中氧原子的4个轨道是不等性sp3杂化

9
第一节 稳定蛋白质三维结构的作用力 极性化合物在水中的溶解是熵增加的过程

10
第一节 稳定蛋白质三维结构的作用力 水分子在非极性化合物周围形成笼型结构,有序性增加,熵减少。热力学不利!

11
第一节 稳定蛋白质三维结构的作用力 在疏水相互作用下,蛋白质疏水氨基酸折叠在蛋白质内部,而亲水氨基酸残基暴露在外。

12
(四)离子键 离子键是带有相反电荷的侧链之间的离子相互作用。
第一节 稳定蛋白质三维结构的作用力 (四)离子键 离子键是带有相反电荷的侧链之间的离子相互作用。 离子化的侧链一般都出现在球蛋白的表面,与水分子形成水化层,对于整个球蛋白的稳定性的贡献是最小的。 荷电的侧链也在蛋白质内部出现,一般与其它基团形成强氢键。
离子键. 离子键是带有相反电荷的侧链之间的离子相互作用。 离子化的侧链一般都出现在球蛋白的表面,与水分子形成水化层,对于整个球蛋白的稳定性的贡献是最小的。 荷电的侧链也在蛋白质内部出现,一般与其它基团形成强氢键。")
13
(五)二硫键 二硫键的形成并不决定多肽链的折叠。
第一节 稳定蛋白质三维结构的作用力 (五)二硫键 二硫键的形成并不决定多肽链的折叠。 二硫键有助于某些球蛋白的天然构象的稳定,二硫键有时存在于由细胞分泌的蛋白质中,当这样的蛋白质离开细胞内环境时,由于有二硫键的存在,可使得蛋白质对去折叠以及降解不那么敏感,而维持蛋白质的稳定。
二硫键. 二硫键的形成并不决定多肽链的折叠。 二硫键有助于某些球蛋白的天然构象的稳定,二硫键有时存在于由细胞分泌的蛋白质中,当这样的蛋白质离开细胞内环境时,由于有二硫键的存在,可使得蛋白质对去折叠以及降解不那么敏感,而维持蛋白质的稳定。")
14
2、多肽主链折叠的空间限制 2.1 肽键 一个多肽链的骨架是由通过肽键连接的重复单位N—Cα—C组成的。
第二节多肽主链折叠的空间限制 2、多肽主链折叠的空间限制 2.1 肽键 一个多肽链的骨架是由通过肽键连接的重复单位N—Cα—C组成的。 参与肽键形成的2个原子和另外4个取代成员:羰基氧原子、酰胺氢原子以及2个相邻的α-碳原子,形成一个肽单位

15
酰胺氮上的孤对电子与相邻羰基之间具有共振作用,形成共振杂化体,稳定性高。

16
肽键的特点: 1.肽键具有部分双键性质: C-N单键键长0.149nm C=N双键键长0.127nm 肽键键长0.132nm
第二节多肽主链折叠的空间限制 肽键的特点: 1.肽键具有部分双键性质: C-N单键键长0.149nm C=N双键键长0.127nm 肽键键长0.132nm 2.肽键不能自由旋转,但N-C和C-C键都可以自由旋转。 3.组成肽键的四个原子和与之相连的两个碳原子(C)都处于同一个平面内,此刚性结构的平面叫肽平面或酰胺平面(amide plane)。 4.几乎所有的肽单位处于反式构象,只有约6%的Pro残基处于顺势构象。
都处于同一个平面内,此刚性结构的平面叫肽平面或酰胺平面(amide plane)。 4.几乎所有的肽单位处于反式构象,只有约6%的Pro残基处于顺势构象。")
17
第二节多肽主链折叠的空间限制 2.2、多肽主链折叠的空间限制 一个蛋白质的构象取决于肽单位绕N-Cα和Cα-C键的旋转,这些键连接着多肽链中的刚性的肽单位。绕N-Cα键旋转的二面角称φ,绕Cα-C键旋转的称ψ。

18
当φ或ψ旋转键所在的酰胺平面的取向二等分H-Cα-R平面,且该旋转键两侧的主链处于顺式构型时,规定φ=0°或ψ=0°
第二节多肽主链折叠的空间限制 当φ或ψ旋转键所在的酰胺平面的取向二等分H-Cα-R平面,且该旋转键两侧的主链处于顺式构型时,规定φ=0°或ψ=0°

19
从Cα沿键轴方向观察,顺时针旋转的φ或ψ角度为正值,逆时针旋转的为负值。
第二节多肽主链折叠的空间限制 从Cα沿键轴方向观察,顺时针旋转的φ或ψ角度为正值,逆时针旋转的为负值。 二面角所规定的构象能否存在,主要取决于两个相邻肽单位中非共价键合原子之间的接近有无障碍。

20
第二节多肽主链折叠的空间限制 拉氏构象图 拉氏构象图上的每一点对应于一对二面角( φ或ψ ),代表一个Cα的两个相邻肽单位的构象。如果将一个蛋白质多肽链上所有C α的成对二面角都画在图上,那么蛋白质的主链构象将清楚的表现在图上。 图中,阴影部分为允许区,空白部分为非允许区。

21
第三节 蛋白质的二级结构 3、蛋白质的二级结构 蛋白质的二级结构是指肽链的主链在空间的排列,或规则的几何走向、旋转及折叠。它只涉及肽链主链的构象及链内或链间形成的氢键。 常见的二级结构元件主要有-螺旋、-折叠、-转角、无规卷曲。

22
3.1 α-螺旋 α-螺旋是蛋白质中最常见,最典型、含量最丰富的二级结构元件。
第三节 蛋白质的二级结构 3.1 α-螺旋 α-螺旋是蛋白质中最常见,最典型、含量最丰富的二级结构元件。 1950年,L.Pauling预测出能够稳定存在的α-螺旋结构

23
第三节 蛋白质的二级结构 α-螺旋的结构特点 多肽链中的各个肽平面围绕同一轴旋转,形成螺旋结构,螺旋一周,沿轴上升的距离即螺距为0.54nm,含3.6个氨基酸残基;每个残基绕轴旋转100°,沿轴上升0.15nm;

24
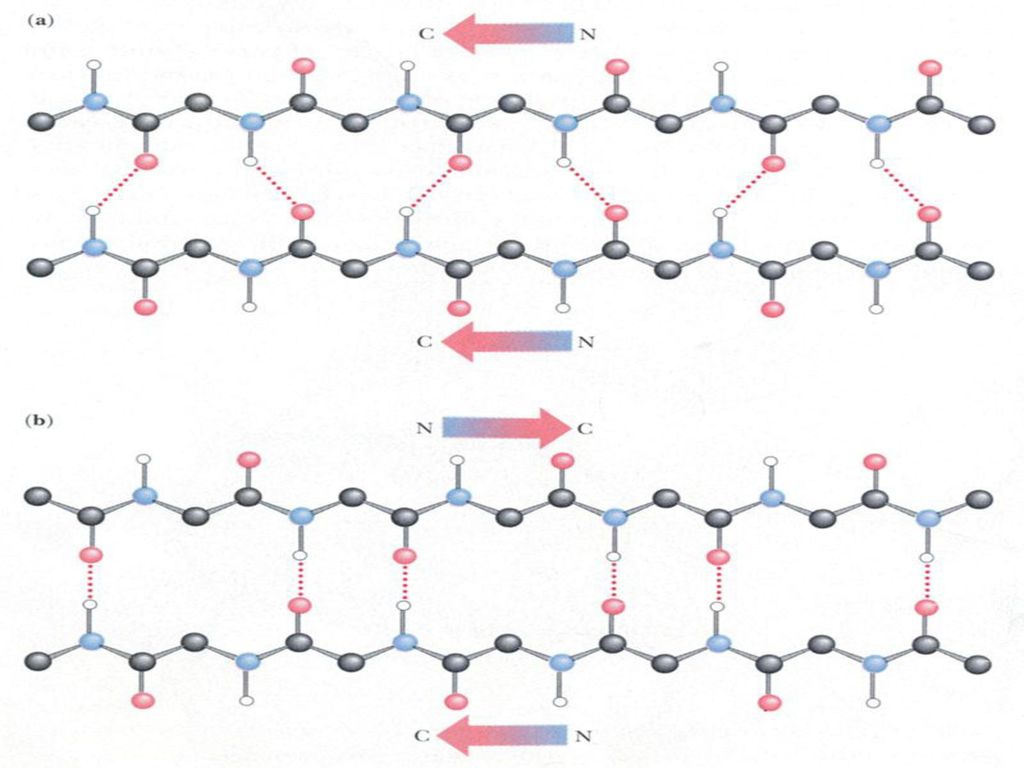
蛋白质中的-螺旋几乎都是右手螺旋。 -螺旋折叠过程中存在协同性。
第三节 蛋白质的二级结构 肽链内形成氢键,氢键的取向几乎与轴平行,每个氨基酸残基的C=O氧与其后第四个氨基酸残基的N-H氢形成氢键。由氢键封闭的环是13元环。(因此,-螺旋也称3.613-螺旋) 蛋白质中的-螺旋几乎都是右手螺旋。 -螺旋折叠过程中存在协同性。
 蛋白质中的-螺旋几乎都是右手螺旋。 -螺旋折叠过程中存在协同性。")
25
第三节 蛋白质的二级结构 Make fists of your two hands with thumbs outstretched and pointing straight up. Looking at your right hand, think of a helix spiraling up your right thumb in the direction in which the other four fingers are curled as shown (counterclockwise). The resulting helix is right-handed. Your left hand will demonstrate a lefthanded helix, which rotates in the clockwise direction as it spirals up your thumb. a simple method for determining whether a helical structure is right-handed or left-handed.
. The resulting helix is right-handed. Your left hand will demonstrate a lefthanded helix, which rotates in the clockwise direction as it spirals up your thumb. a simple method for determining whether a helical structure is right-handed or left-handed.")
26
影响α-螺旋形成的因素 -螺旋的形成及其稳定性与其氨基酸组成和顺序有关 PolyAla生理条件下自发卷曲,形成-螺旋(R小,不带电荷)
第三节 蛋白质的二级结构 影响α-螺旋形成的因素 -螺旋的形成及其稳定性与其氨基酸组成和顺序有关 PolyAla生理条件下自发卷曲,形成-螺旋(R小,不带电荷) polyLys、PolyIle生理条件下不能形成-螺旋(带电荷、R较大)。 多肽链中只要存在Pro, -螺旋即被中断。(刚性环、不能形成氢键)
 polyLys、PolyIle生理条件下不能形成-螺旋(带电荷、R较大)。 多肽链中只要存在Pro, -螺旋即被中断。(刚性环、不能形成氢键)")
27
第三节 蛋白质的二级结构 3.2、 -折叠 -折叠也称-结构或-构象,它是蛋白质中第二种最常见的二级结构。

28
第三节 蛋白质的二级结构 -折叠特点 -折叠结构的氢键主要是由两条肽链之间形成的;也可以在同一肽链的不同部分之间形成。几乎所有肽键都参与链间氢键的交联,氢键与链的长轴接近垂直。 -折叠有两种类型:一种为平行式,即所有肽链的N-端都在同一边。另一种为反平行式,即相邻两条肽链的方向相反。

30
3.3、 -转角 肽链在某处回折180°所形成的结构,由四个氨基酸残基组成;
第三节 蛋白质的二级结构 3.3、 -转角 肽链在某处回折180°所形成的结构,由四个氨基酸残基组成; 弯曲处的第一个氨基酸残基的 -C=O 和第四个残基的 –N-H 之间形成氢键,形成一个比较稳定的环状结构。 -转角有两种主要类型,它们之间的差别只是中央肽基旋转了180°。

31
-转角的特定构象取决于它的组成氨基酸。 由于甘氨酸缺少侧链,在-转角中能够很好的调整其它残基的空间阻碍,因此是立体化学上最合适的氨基酸。
第三节 蛋白质的二级结构 -转角的特定构象取决于它的组成氨基酸。 由于甘氨酸缺少侧链,在-转角中能够很好的调整其它残基的空间阻碍,因此是立体化学上最合适的氨基酸。 脯氨酸具有环状的结构和固定的φ角,因此,一定程度上迫使-转角形成,促进多肽链自身回折。这有助于反平行-折叠片的形成。

32
第三节 蛋白质的二级结构 3.4、无规卷曲 泛指那些不能被归入明确的二级结构如折叠片或螺旋的多肽片段。 酶的功能部位常包含此构象。

33
第四节 纤维状蛋白质 4、纤维状蛋白质 纤维状蛋白质(fibrous protein)广泛地分布于脊椎和无脊椎动物体内,它是动物体的基本支架和外表保护成分,占脊椎动物体内蛋白质总量的一半或一半以上。 这类蛋白质外形呈纤维状或细棒状,分子是有规则的线型结构,这与其多肽链的有规则二级结构有关,而有规则的线型二级结构是它们的氨基酸顺序的规则性反映。

34
纤维状蛋白质的类型 纤维状蛋白质可分为不溶性(硬蛋白)和可溶性两类,前者有角蛋白、胶原蛋白和弹性蛋白等;
第四节 纤维状蛋白质 纤维状蛋白质的类型 纤维状蛋白质可分为不溶性(硬蛋白)和可溶性两类,前者有角蛋白、胶原蛋白和弹性蛋白等; 后者有肌球蛋白和纤维蛋白原等,但不包括微管(microtubule)和肌动蛋白细丝(actin filament),它们是球状蛋白质的长向聚集体(aggregate)。
和可溶性两类,前者有角蛋白、胶原蛋白和弹性蛋白等; 后者有肌球蛋白和纤维蛋白原等,但不包括微管(microtubule)和肌动蛋白细丝(actin filament),它们是球状蛋白质的长向聚集体(aggregate)。")
35
-角蛋白 角蛋白广泛存在于动物的皮肤及皮肤的衍生物,如毛发、甲、角、鳞和羽等,属于结构蛋白。角蛋白中主要的是-角蛋白。
第四节 纤维状蛋白质 -角蛋白 角蛋白广泛存在于动物的皮肤及皮肤的衍生物,如毛发、甲、角、鳞和羽等,属于结构蛋白。角蛋白中主要的是-角蛋白。 -角蛋白主要由-螺旋构象的多肽链组成。一般是由三条右手-螺旋肽链形成一个原纤维(向左缠绕),原纤维的肽链之间有二硫键交联以维持其稳定性 例如毛的纤维是由多个原纤维平行排列,并由氢键和二硫键作为交联键将它们聚集成不溶性的蛋白质。 -角蛋白的伸缩性能很好,当-角蛋白被过度拉伸时,则氢键被破坏而不能复原。此时-角蛋白转变成-折叠结构,称为-角蛋白。
,原纤维的肽链之间有二硫键交联以维持其稳定性. 例如毛的纤维是由多个原纤维平行排列,并由氢键和二硫键作为交联键将它们聚集成不溶性的蛋白质。 -角蛋白的伸缩性能很好,当-角蛋白被过度拉伸时,则氢键被破坏而不能复原。此时-角蛋白转变成-折叠结构,称为-角蛋白。")
36
第四节 纤维状蛋白质 毛发的结构 一根毛发周围是一层鳞状细胞,中间为皮层细胞。皮层细胞横截面直径约为20m。在这些细胞中,大纤维沿轴向排列。所以一根毛发具有高度有序的结构。 毛发性能就决定于—螺旋结构以及这样的组织方式

37
第四节 纤维状蛋白质 -角蛋白 丝心蛋白(fibroin):这是蚕丝和蜘蛛丝的一种蛋白质。丝心蛋白具有抗张强度高,质地柔软的特性,但不能拉伸。丝心蛋白是典型的反平行式-折叠片,多肽链取锯齿状折叠构象。
:这是蚕丝和蜘蛛丝的一种蛋白质。丝心蛋白具有抗张强度高,质地柔软的特性,但不能拉伸。丝心蛋白是典型的反平行式-折叠片,多肽链取锯齿状折叠构象。 .")
38
第四节 纤维状蛋白质

39
丝蛋白是由伸展的肽链沿纤维轴平行排列成反向-折叠结构。丝蛋白的肽链通常是由小侧链的Gly,Ser,Ala组成,每隔一个残基就是Gly。
第四节 纤维状蛋白质 丝蛋白是由伸展的肽链沿纤维轴平行排列成反向-折叠结构。丝蛋白的肽链通常是由小侧链的Gly,Ser,Ala组成,每隔一个残基就是Gly。

40
第四节 纤维状蛋白质 胶原蛋白 天然的胶原是一个由3条具有左手螺旋的链相互缠绕形成右手超螺旋的分子。胶原的超螺旋结构是靠链间氢键以及螺旋和超螺旋的反向盘绕维持其稳定性的。一个典型的胶原分子长300nm、直径为1.5nm。在每一条左手螺旋的胶原链内,每一圈螺旋需要3个氨基酸残基,螺距为0.94nm,即每一个氨基酸残基轴向距离为0.31nm。胶原中含有在其他蛋白中很少出现的羟脯氨酸(Hyp),序列-Gly-Pro-Hyp-常出现在胶原分子中。
,序列-Gly-Pro-Hyp-常出现在胶原分子中。")
41
第四节 纤维状蛋白质

42
第五节 超二级结构和结构域 5、超二级结构和结构域 超二级结构指蛋白质中相邻的二级结构单位(-螺旋或-折叠或-转角)组合在一起,彼此相互作用,形成有规则的、在空间上能够辨认的二级结构组合体。 超二级结构的基本类型:、 、

43
结构域:多肽链在二级结构或超二级结构的基础上形成三级结构的局部折叠区,它是独立的紧密球状实体。
第五节 超二级结构和结构域 结构域:多肽链在二级结构或超二级结构的基础上形成三级结构的局部折叠区,它是独立的紧密球状实体。

44
结构域有时也称功能域。一般说,功能域是蛋白质分子中能独立存在的功能单位。功能域可以是一个结构域,也可以是两个或以上结构域组成。
第五节 超二级结构和结构域 结构域有时也称功能域。一般说,功能域是蛋白质分子中能独立存在的功能单位。功能域可以是一个结构域,也可以是两个或以上结构域组成。 结构域之间通常只有一段柔性的肽链连接,形成所谓铰链区,使结构域容易发生相对运动。

45
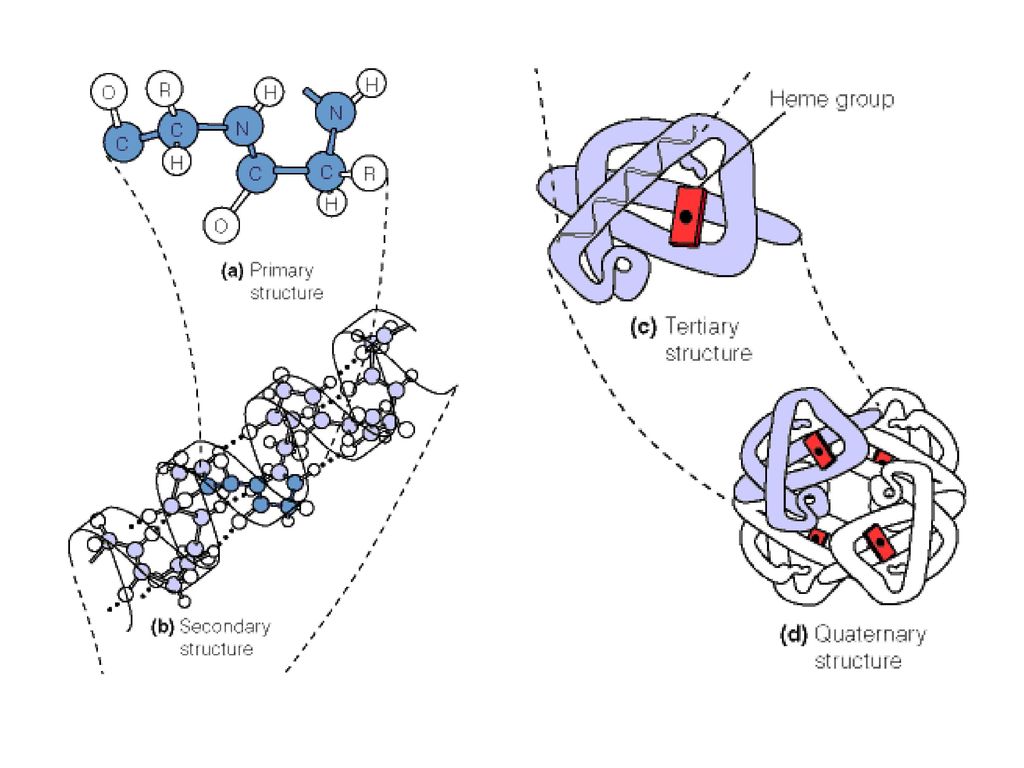
第六节 蛋白质的三级结构 6、蛋白质的三级结构 蛋白质的三级结构(Tertiary Structure)是指多肽链在二级结构、超二级结构、结构域的基础上,进一步盘绕、折叠形成的包括主链和侧链构象在内的特征三维结构。
是指多肽链在二级结构、超二级结构、结构域的基础上,进一步盘绕、折叠形成的包括主链和侧链构象在内的特征三维结构。 .")
46
球状蛋白质 球状蛋白质可根据它们的结构域类型分为4大类: 1、全α-结构蛋白质 2、α,β-结构蛋白质 3、全β-结构蛋白质
第六节 蛋白质的三级结构 球状蛋白质 球状蛋白质可根据它们的结构域类型分为4大类: 1、全α-结构蛋白质 2、α,β-结构蛋白质 3、全β-结构蛋白质 4、富含金属或二硫键蛋白质

47
第六节 蛋白质的三级结构 全α-结构蛋白质

48
第六节 蛋白质的三级结构 α,β-结构蛋白质

49
第六节 蛋白质的三级结构 全β-结构蛋白质

50
富含金属或二硫键蛋白质

51
第六节 蛋白质的三级结构 Motifs是蛋白质结构分类的基础

52
虽然发现的新蛋白质的数目逐年递增,但发现的新的folds的量却在显著减少。

53
第六节 蛋白质的三级结构 Protein tertiary structure is more reliably conserved than primary sequence. Proteins with significant primary sequence similarity, and/or with demonstrably similar structure and function, are said to be in the same protein family. Two or more families with little primary sequence similarity sometimes make use of the same major structural motif and have functional similarities; these families are grouped as superfamilies.

54
第六节 蛋白质的三级结构 Structural motifs become especially important in defining protein families and superfamilies. Improved classification and comparison systems for proteins lead inevitably to the elucidation of new functional relationships. Given the central role of proteins in living systems, these structural comparisons can help illuminate every aspect of biochemistry, from the evolution of individual proteins to the evolutionary history of complete metabolic pathways.

55
球状蛋白质三维结构的特征 1、球状蛋白质分子含有多种二级结构元件。 2、球状蛋白质三维结构具有明显的折叠层次。
第六节 蛋白质的三级结构 球状蛋白质三维结构的特征 1、球状蛋白质分子含有多种二级结构元件。 2、球状蛋白质三维结构具有明显的折叠层次。 3、球状蛋白质是紧密的球状或椭球状实体。 4、球状蛋白质疏水侧链埋藏在分子内部,亲水侧链暴露在分子表面。 5、球状蛋白质分子表面有空穴,这种空穴常是结合底物、效应物等配体并行使生物功能的活性部位。

56
第六节 蛋白质的三级结构 抗体的抗原结合部位

57
7、蛋白质的折叠和结构预测 蛋白质精致的三维结构是怎样获得的?蛋白质的三维结构与一维的氨基酸序列信息之间是何关系?
第七节 蛋白质的折叠和结构预测 7、蛋白质的折叠和结构预测 蛋白质精致的三维结构是怎样获得的?蛋白质的三维结构与一维的氨基酸序列信息之间是何关系? 二十世纪五十年代,Christian Anfinsen 关于核糖核酸酶变性和复性的经典实验揭示了氨基酸序列与蛋白质三维结构之间的关系。

58
第七节 蛋白质的折叠和结构预测 7.1 蛋白质的变性与复性 天然蛋白质分子受到某些物理因素如热、紫外线照射、高压和表面张力等或化学因素如有机溶剂、脲、胍、酸、碱等的影响时,生物活性丧失,溶解度降低,不对称性增高以及其他的物理化学常数发生改变,这种过程称为蛋白质变性。

59
蛋白质变性的实质是蛋白质分子中的次级键被破坏,引起天然构象解体。变性不涉及共价键(肽键)的破裂,一级结构仍保持完好。
第七节 蛋白质的折叠和结构预测 蛋白质变性的实质是蛋白质分子中的次级键被破坏,引起天然构象解体。变性不涉及共价键(肽键)的破裂,一级结构仍保持完好。 变性因素除去后,变性蛋白又可重新恢复到天然构象,这一现象称为蛋白质的复性。
的破裂,一级结构仍保持完好。 变性因素除去后,变性蛋白又可重新恢复到天然构象,这一现象称为蛋白质的复性。")
60
第七节 蛋白质的折叠和结构预测 7.2 核糖核酸酶的变性和复性试验 变性:加8mol/L尿素或6mol/L盐酸胍和巯基乙醇(还原二硫键),酶变性,紧密结构伸展成松散的无规卷曲构象。 复性:将尿素等变性剂和巯基乙醇用透析法除去,酶活性又可恢复,最后达原活性的95~100%。
,酶变性,紧密结构伸展成松散的无规卷曲构象。 复性:将尿素等变性剂和巯基乙醇用透析法除去,酶活性又可恢复,最后达原活性的95~100%。 .")
61
实验:酶变性后,首先透析除去巯基乙醇,Cys氧化后,除去尿素,酶活性只恢复到原活性的1%。
第七节 蛋白质的折叠和结构预测 结论:氨基酸序列决定三级结构。 问题:二硫键的形成是否决定三级结构? 实验:酶变性后,首先透析除去巯基乙醇,Cys氧化后,除去尿素,酶活性只恢复到原活性的1%。 结论:二硫键只能在多肽链折叠成正确的结构之后形成。

62
蛋白质的变性和复性是一个协调过程,在所加变性剂很窄的浓度范围内,或很窄的pH或温度区间内突然发生。
第七节 蛋白质的折叠和结构预测 蛋白质的变性和复性是一个协调过程,在所加变性剂很窄的浓度范围内,或很窄的pH或温度区间内突然发生。

63
第七节 蛋白质的折叠和结构预测 7.3、蛋白质的折叠可能是通过中间体的累进稳定性(Progressive stabilization of intermediates)而非随机组合 Consider a small protein with 100 residues. If each residue can assume three different conformations, the total number of structures would be 3100, which is equal to 5 × If it takes s to convert one structure into another, the total search time would be 5 × 1047 × s, which is equal to 5 × 1034 s, or 1.6 × 1027 years.

64
the power of cumulative selection
第七节 蛋白质的折叠和结构预测 the power of cumulative selection 1040 of keystrokes to about 2800

65
第七节 蛋白质的折叠和结构预测 7.4、分子伴侣(Chaperone) 伸展或部分折叠的蛋白质在高浓度时倾向于聚集,在体内蛋白质能在很稠的介质中高效发生的原因是因为分子伴侣的参与。 分子伴侣包括大量不同的蛋白质家族,这些蛋白质的功能包括帮助蛋白质的折叠和装配,稳定去折叠的蛋白质,对蛋白质去折叠以便转运过膜或进入蛋白降解途径等。

66
分子伴侣的特性 分子伴侣通常与去折叠的或部分折叠的蛋白质相互作用。 他们可以促进蛋白质正确折叠或稳定非天然蛋白构象。
第七节 蛋白质的折叠和结构预测 分子伴侣的特性 分子伴侣通常与去折叠的或部分折叠的蛋白质相互作用。 他们可以促进蛋白质正确折叠或稳定非天然蛋白构象。 它们不与天然蛋白相互作用,也不会成为折叠后蛋白结构的一个组成部分。 分子伴侣对底物的特异性不同。 分子伴侣协助蛋白质折叠的过程中需要与ATP的水解偶联。 在细胞胁迫(cell stress)的情况下,分子伴侣的表达通常会增加。
的情况下,分子伴侣的表达通常会增加。")
67
Families Of Molecular Chaperones
第七节 蛋白质的折叠和结构预测 Families Of Molecular Chaperones Small heat shock proteins (hsp25) [holders] protect against cellular stress prevent aggregation in the lens (cataract) Hsp60 system (cpn60, GroEL) ATPase [(un)folders] protein folding Hsp70 system (DnaK, BiP) ATPase [(un)folders] stabilization of extended chains membrane translocation regulation of the heat shock response
 [holders] protect against cellular stress. prevent aggregation in the lens (cataract) Hsp60 system (cpn60, GroEL) ATPase [(un)folders] protein folding. Hsp70 system (DnaK, BiP) ATPase [(un)folders] stabilization of extended chains. membrane translocation. regulation of the heat shock response.")
68
第七节 蛋白质的折叠和结构预测 Hsp90 [holder] binding and stabilization/regulation of steroid receptors, protein kinases Buffer for genetic variation? Hsp100 (Clp) ATPase [unfolder] thermotolerance, proteolysis, resolubilization of aggregates Calnexin, calreticulin glycoprotein maturation in the ER quality control Folding catalysts: PDI, PPI [folders] Prosequences: alpha-lytic protease, subtilisin (intramolecular chaperones) [folders]
![第七节 蛋白质的折叠和结构预测 Hsp90 [holder] binding and stabilization/regulation of steroid receptors, protein kinases.](http://slidesplayer.com/slide/11444700/61/images/68/%E7%AC%AC%E4%B8%83%E8%8A%82+%E8%9B%8B%E7%99%BD%E8%B4%A8%E7%9A%84%E6%8A%98%E5%8F%A0%E5%92%8C%E7%BB%93%E6%9E%84%E9%A2%84%E6%B5%8B+Hsp90+%5Bholder%5D+binding+and+stabilization%2Fregulation+of+steroid+receptors%2C+protein+kinases..jpg "Buffer for genetic variation Hsp100 (Clp) ATPase [unfolder] thermotolerance, proteolysis, resolubilization of aggregates. Calnexin, calreticulin. glycoprotein maturation in the ER. quality control. Folding catalysts: PDI, PPI [folders] Prosequences: alpha-lytic protease, subtilisin (intramolecular chaperones) [folders]")
69
第七节 蛋白质的折叠和结构预测

70
第七节 蛋白质的折叠和结构预测 蛋白质结构预测 根据分子生物学一个中心原理:顺序规定构象,活性依靠结构,蛋白质结构是可预测的。全新蛋白质设计和蛋白质工程更需要蛋白质结构预测。

71
第七节 蛋白质的折叠和结构预测 氨基酸残基在二级结构中的相对出现频率 Amino acid α helix β sheet Turn Ala
1.29 0.90 0.78 Cys 1.11 0.74 0.80 Leu 1.30 1.02 0.59 Met 1.47 0.97 0.39 Glu 1.44 0.75 1.00 Gln 1.27 His 1.22 1.08 0.69 Lys 1.23 0.77 0.96 Val 0.91 1.49 0.47 Ile 1.45 0.51 Phe 1.07 1.32 0.58 Tyr 0.72 1.25 1.05 Trp 0.99 1.14 Thr 0.82 1.21 1.03 Gly 0.56 0.92 1.64 Ser 0.95 1.33 Asp 1.04 1.41 Asn 0.76 1.28 Pro 0.52 0.64 1.91 Arg 0.88 氨基酸残基在二级结构中的相对出现频率

72
第七节 蛋白质的折叠和结构预测 The α helix can be regarded as the default conformation. Branching at the β-carbon atom, as in valine, threonine, and isoleucine, tends to destabilize α helices because of steric clashes. These residues are readily accommodated in β strands, in which their side chains project out of the plane containing the main chain. Serine, aspartate, and asparagine tend to disrupt α helices because their side chains contain hydrogen-bond donors or acceptors in close proximity to the main chain, where they compete for main-chain NH and CO groups. Proline tends to disrupt both α helices and β strands because it lacks an NH group and because its ring structure restricts its φvalue to near -60 degrees. Glycine readily fits into all structures and for that reason does not favor helix formation in particular.

73
第七节 蛋白质的折叠和结构预测 关于二级结构预测的经验规则 α螺旋:相邻6个aa中,若4个倾向于形成螺旋,则被认为是螺旋核。螺旋向两端延伸到4个残基的Pa<1.0为止; 折叠片预测:相邻5个残基中有3个倾向于形成折叠片,则认为是折叠核。折叠向两端延伸至4个残基的Pb<1.0; β转角:Pt>1.0,且>Pa和Pb

74
8、蛋白质的四级结构 四级结构是指由两 个或两个以上具有三级结构的亚基按一定方式聚合而成的特定构象的蛋白质分子。
第八节 蛋白质的四级结构 8、蛋白质的四级结构 四级结构是指由两 个或两个以上具有三级结构的亚基按一定方式聚合而成的特定构象的蛋白质分子。 通常亚基只有一条多肽链,但有的亚基由两条或多条多肽链组成,这些多肽链间多以二硫键相连。亚基单独存在时无生物学活性。 亦可以说,具有四级结构的蛋白属于寡聚蛋白。(寡聚蛋白中亚基可以是相同的或不相同的,亚基相同时,分子具有对称性) 实质:蛋白质的四级结构实际上研究亚基之间的相互作用、空间排布及亚基接触部位的空间布局。
 实质:蛋白质的四级结构实际上研究亚基之间的相互作用、空间排布及亚基接触部位的空间布局。")
76
维持蛋白质四级结构的主要作用力 在蛋白质四级结构中,亚基之间的作用力主要包括:氢键、离子键、范德华力和疏水键。
第八节 蛋白质的四级结构 维持蛋白质四级结构的主要作用力 在蛋白质四级结构中,亚基之间的作用力主要包括:氢键、离子键、范德华力和疏水键。 即亚基之间通过次级键彼此缔合在一起(二硫键?) 疏水键是最主要的作用力。
 疏水键是最主要的作用力。")
77
第八节 蛋白质的四级结构 四级结构对称性 是四级结构蛋白主要性质之一,有旋转对称轴,如二聚体,四聚体。

78
四级缔合在结构和功能上具有优越性 可增强结构稳定性; 提高遗传经济性和效率,使编码所需DNA减少; 可使催化基团汇集在一起;
第八节 蛋白质的四级结构 四级缔合在结构和功能上具有优越性 可增强结构稳定性; 提高遗传经济性和效率,使编码所需DNA减少; 可使催化基团汇集在一起; 具有协同性和别构效应。

79
具有别构效应的蛋白质为别构蛋白质,酶称为别构酶,具有调节代谢的功能。
第八节 蛋白质的四级结构 别构效应:多亚基蛋白质一般具有多个结合部位,结合在蛋白质分子的特定部位上的配体对该分子和其他部位所产生的影响(如改变亲和力或催化能力)称为别构效应。 具有别构效应的蛋白质为别构蛋白质,酶称为别构酶,具有调节代谢的功能。
称为别构效应。 具有别构效应的蛋白质为别构蛋白质,酶称为别构酶,具有调节代谢的功能。")
80
第八节 蛋白质的四级结构

Similar presentations


:核蛋白体组成成分 转移 RNA ( tRNA ):转运氨基酸 信使 RNA ( mRNA ):蛋白质合成模板 不均一核 RNA ( hnRNA ):成熟 mRNA 的前体 小核 RNA ( snRNA ):>")







.>")

蛋白质的降解: 外源蛋白的消化 内源性蛋白的选择性降解 (2)氨基酸的分解代谢:>")






 蛋 白 质 的 一 级 结 构 (primary structure) 指 它 的 氨 基 酸 序 列 。 首 N 端( “H”) 末 C 端(“OH”) 其 中 的 氨 基 酸 称 氨 基 酸.>")