Download presentation
1
第十二章 植物對內在和外在訊息的反應 (Plant Response to Internal and External Signals)
植物科學研究所 陳益明老師

2
一、 緒言 本章討論植物的形態與生理如何受到複雜的環境因子及內在訊息(signals)而改變原來的生命現象,這些內在的訊息包括化學信息(chemical messengers)—荷爾蒙(hormones)及光及溫度等外在信息。
而改變原來的生命現象,這些內在的訊息包括化學信息(chemical messengers)—荷爾蒙(hormones)及光及溫度等外在信息。")
3
二、 訊息傳遞與植物的感應 (Signal transduction and plant responses)
")
4
二、 訊息傳遞與植物的感應 訊息的傳遞途徑需聯結內在及外在環境訊息才能產生細胞的反應(signal-transduction pathway link internal and environmental signals to cellular responses): 1. 白化幼苗(etiolated seedlings)或在暗中長芽的馬鈴薯照光後變綠色。
或在暗中長芽的馬鈴薯照光後變綠色。")
6
二、 訊息傳遞與植物的感應 2.植物細胞接受各種訊息後將引發一連串的信息傳遞途(signal transduction pathway)
一級訊息(primary messenger ): 植物荷爾蒙(plant hormones),光,溫度等。 二級訊息(secondary messenger): Ca+2 ,cyclic GMP,cyclic AMP等。
: 植物荷爾蒙(plant hormones),光,溫度等。 二級訊息(secondary messenger): Ca+2 ,cyclic GMP,cyclic AMP等。")
7
Review of a general model for signal-transduction pathways

9
三、 光接受器 (photoreceptors)
")
10
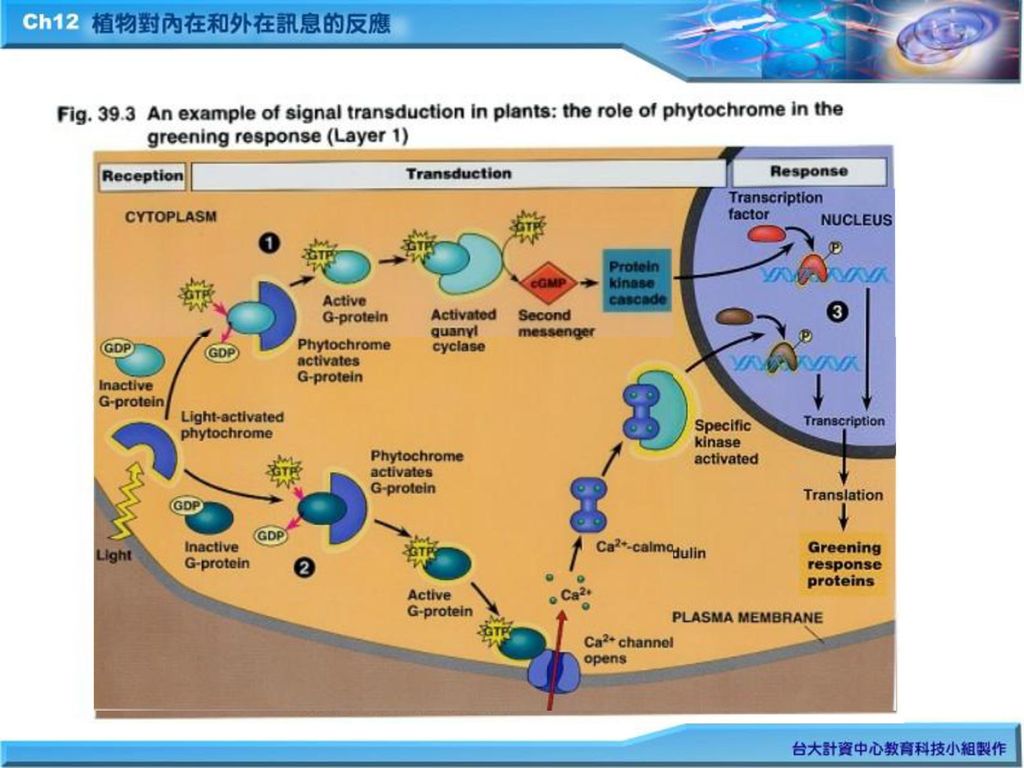
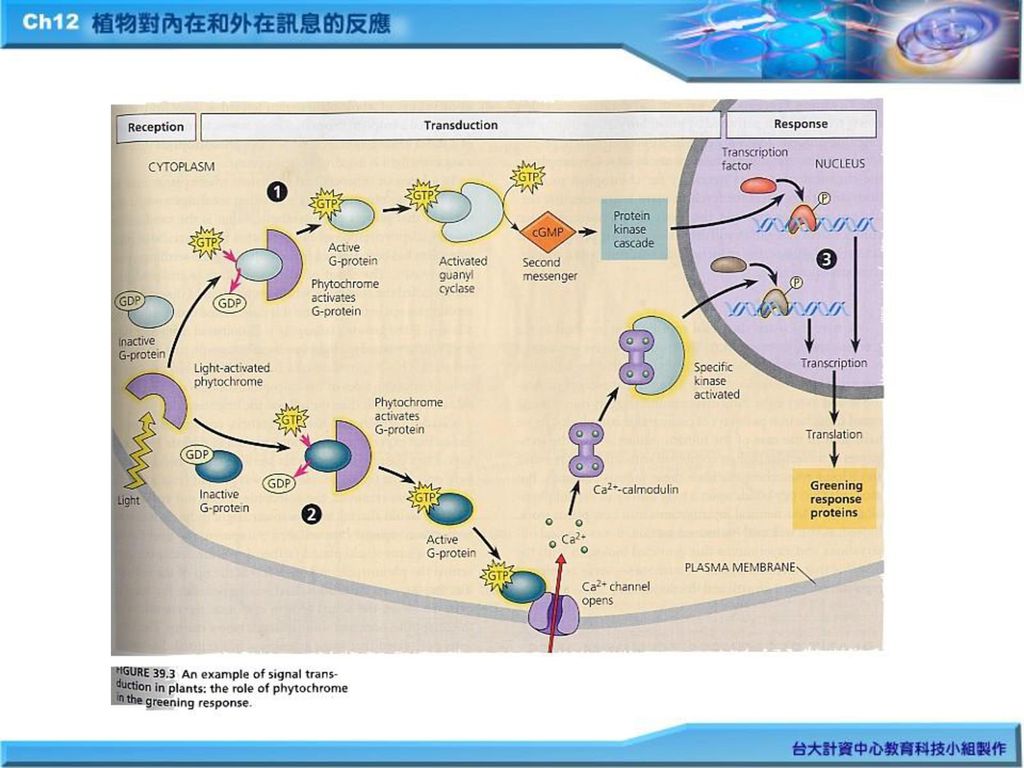
三、 光接受器 光訊息先被植物色原或稱光敏素(phytochrome)等接收後,才產生一系列訊息傳遞途徑。G-protein與cAMP充當二級訊息,它活化細胞膜上的蛋白質激酶(protein kinase),藉骨牌效應(cascade)而活化一系列生化反應(Fig.11.11);另一途徑則形成Ca+2 -calmodulin complex,再活化某一特殊的protein kinase。以上二者最後皆導致植物綠化(greening)所需之基因表現(the expression of genes)。
等接收後,才產生一系列訊息傳遞途徑。G-protein與cAMP充當二級訊息,它活化細胞膜上的蛋白質激酶(protein kinase),藉骨牌效應(cascade)而活化一系列生化反應(Fig.11.11);另一途徑則形成Ca+2 -calmodulin complex,再活化某一特殊的protein kinase。以上二者最後皆導致植物綠化(greening)所需之基因表現(the expression of genes)。")
12
四、 蛋白質的轉譯後的修飾(Post-translational modification of protein)
")
13
四、 蛋白質的轉譯後的修飾 生物細胞內蛋白質的磷酸化作(phosphorylation)及去磷酸化作用 (dephosphorylation)是活化細胞代謝或關閉(switch-off)代謝的主要機制。前者以protein kinase 將磷酸基(~PO4)基加入某一蛋白質(Fig.11.11)而活化某一生化代謝。protein phosphatase 則從某一特殊之蛋白質的磷酸基去除,此二者代謝後細胞的活性保持平衡。
及去磷酸化作用 (dephosphorylation)是活化細胞代謝或關閉(switch-off)代謝的主要機制。前者以protein kinase 將磷酸基(~PO4)基加入某一蛋白質(Fig.11.11)而活化某一生化代謝。protein phosphatase 則從某一特殊之蛋白質的磷酸基去除,此二者代謝後細胞的活性保持平衡。")
14
五、 植物對荷爾蒙的反應(Plant response to hormones)
")
15
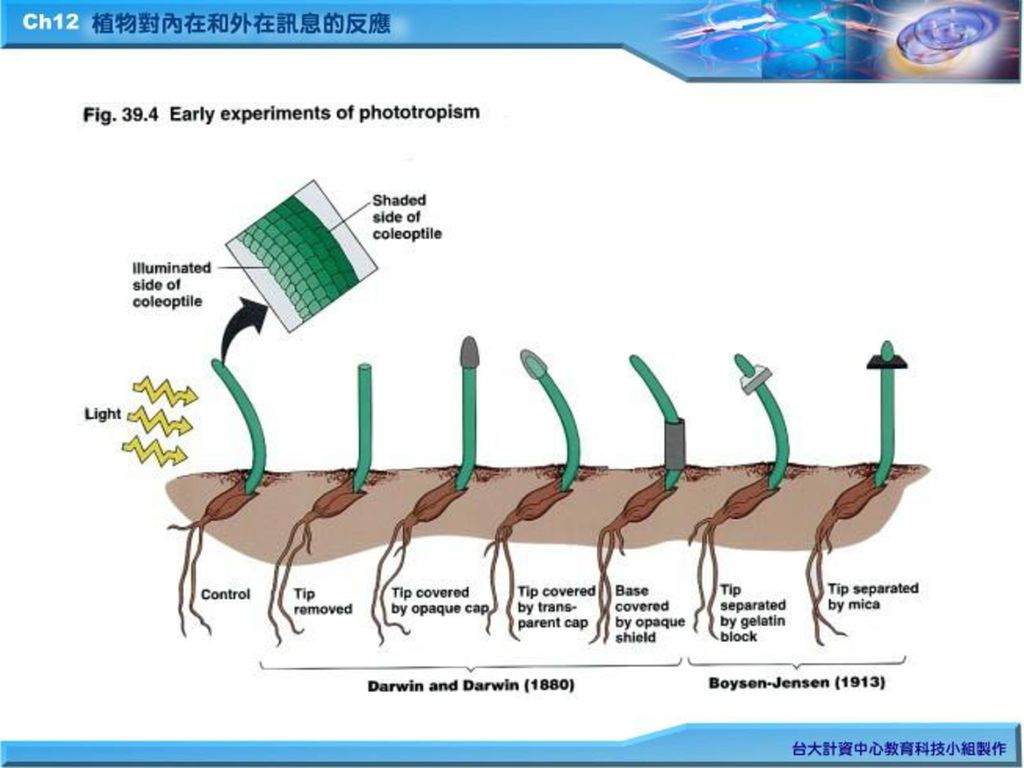
Charles Darwin 及其兒子Francis 研究植物何能向光彎曲生長而導致生長素(auxin)的發現。
五、 植物對荷爾蒙的反應 (一)植物荷爾蒙的發現: Charles Darwin 及其兒子Francis 研究植物何能向光彎曲生長而導致生長素(auxin)的發現。 1.向光性(phototropism)的早期發現。 1881年合寫一本” The power of movement on plant”
植物荷爾蒙的發現: Charles Darwin 及其兒子Francis 研究植物何能向光彎曲生長而導致生長素(auxin)的發現。 1.向光性(phototropism)的早期發現。 1881年合寫一本 The power of movement. on plant")
17
五、 植物對荷爾蒙的反應 2. Peter Boysen-Jensen 及E.W.Wen(1926)的實驗(Fig.39.5)。 3. K. Thimann次分離出生長素(1930’s)。
的實驗(Fig.39.5)。 3. K. Thimann次分離出生長素(1930’s)。")
19
五、 植物對荷爾蒙的反應 (二)植物荷爾蒙扮演協調植物生長和發育, 並接收環境刺激產生感應(response)。 至今已知的生長素,依照其名稱、分子結構,來源及主要生理功能可分為六大類,參考Table 39.1)。
植物荷爾蒙扮演協調植物生長和發育, 並接收環境刺激產生感應(response)。 至今已知的生長素,依照其名稱、分子結構,來源及主要生理功能可分為六大類,參考Table 39.1)。")
20
五、 植物對荷爾蒙的反應 生長素(auxin) 細胞分裂素(cytokinins) 徒長素或吉貝素(gibberellins) 離層素(abscisic acid) 乙烯(ethylene) 芥子類固醇(brassinosteroid)
")
22
(三)生長素的生理功能(physiological function):
五、 植物對荷爾蒙的反應 (三)生長素的生理功能(physiological function): 1.促進細胞延長(cell elongation) 1)在10-8至10 –3 M濃度下是一般生理作用的適當濃度。 2)具有極性運送(polar transport)的特性(Fig.39.6)。
生長素的生理功能(physiological function): 1.促進細胞延長(cell elongation) 1)在10-8至10 –3 M濃度下是一般生理作用的適當濃度。 2)具有極性運送(polar transport)的特性(Fig.39.6)。")
23
Polar auxin transport:
a chemiosmotic model

24
五、 植物對荷爾蒙的反應 3)酸性生長假說(acid growth hypotheis)(Fig.39.7)。
酸性生長假說(acid growth hypotheis)(Fig.39.7)。")
25
五、 植物對荷爾蒙的反應 2.其它生理作用: 1)促進維管束形成層的細胞分裂 2)長不定根(adeventitious roots) 3)促進果實發育,無種子的水果 4) 除草劑(herbicides)
 除草劑(herbicides) .")
26
五、 植物對荷爾蒙的反應 (四) 細胞分裂素(cytokinins) 1.F.Skoog and C.O.Miller 首次由青魚(sperm)的DNA經分解後分離出kinetin。 2.控制細胞分裂與分化 3.控制頂芽優勢(apical dominance) 4.防止葉片等器官之老化(aging)
 4.防止葉片等器官之老化(aging)")
28
(五)吉貝素、徒長素、激勃素(gibberellins),簡稱Gas
五、 植物對荷爾蒙的反應 (五)吉貝素、徒長素、激勃素(gibberellins),簡稱Gas 1.1926年日本人E.Kurosawa 在台大農場發現水稻徒長病(bakanae disease),它是由Gibberella屬病菌感染引起的(Fig.39.9)。
吉貝素、徒長素、激勃素(gibberellins),簡稱Gas 年日本人E.Kurosawa 在台大農場發現水稻徒長病(bakanae disease),它是由Gibberella屬病菌感染引起的(Fig.39.9)。")
29
五、 植物對荷爾蒙的反應 2.至今共有八十餘種不同的GAs被發現,每一種都具有促進遺傳性矮種植物莖的延長(stem elongation)的特性(Fig.39.10)。
的特性(Fig.39.10)。")
31
五、 植物對荷爾蒙的反應 3.果實的生長:Thompson 無子葡萄(Fig.39.11)。 4.種子萌芽:GAs促進α- amylase等水解酵素基因之表現,分解種子貯藏養分。
。 4.種子萌芽:GAs促進α- amylase等水解酵素基因之表現,分解種子貯藏養分。")
33
(六)離層素(abscissic acid 簡稱ABA)
五、 植物對荷爾蒙的反應 (六)離層素(abscissic acid 簡稱ABA) 1.促進植物生長,延緩或休眠,屬逆境荷爾蒙(stress hormones) 2.促使種子及芽等之休眠 (dormancy)
離層素(abscissic acid 簡稱ABA) 1.促進植物生長,延緩或休眠,屬逆境荷爾蒙(stress hormones) 2.促使種子及芽等之休眠 (dormancy)")
34
五、 植物對荷爾蒙的反應 (七) 乙烯(ethylene) 1.乙烯(CH2=CH2) 2.抑制細胞及根部的延長,並抑制腋芽的生長
3.使植物失去地心引力等三種反應(triple response)(Fig.39.13) 4.促進植物老化與死亡 5.促進果實成熟(ripening) 6.促進葉片脫落(abscission)
(Fig.39.13) 4.促進植物老化與死亡. 5.促進果實成熟(ripening) 6.促進葉片脫落(abscission)")
36
五、 植物對荷爾蒙的反應 (八)突變株(mutant)植物的分析有助於荷爾蒙生合成及其功能的分子生物學研究。 1.番茄果實不成熟的突變株,是因為乙烯的接受器(receptor)基因發生突變,對乙烯信息無法接受,導致果實無法進行成熟之一系列生化變化。例如,乙烯triple response突變株。
突變株(mutant)植物的分析有助於荷爾蒙生合成及其功能的分子生物學研究。 1.番茄果實不成熟的突變株,是因為乙烯的接受器(receptor)基因發生突變,對乙烯信息無法接受,導致果實無法進行成熟之一系列生化變化。例如,乙烯triple response突變株。")
39
2.對上述五類植物荷爾蒙的了解,由分析突變株可發現其它種類的荷爾蒙,例如:
五、 植物對荷爾蒙的反應 2.對上述五類植物荷爾蒙的了解,由分析突變株可發現其它種類的荷爾蒙,例如: oligosaccharins: 由纖維素與果膠質之分解物分離出之寡醣類,它與植物對病原菌引起的防禦,調節植物生長、細胞分化及花的發育等有關。

40
五、 植物對荷爾蒙的反應 brassinosteroids: 構造與膽固醇及性荷爾蒙相似,正常生長及發育必要的。阿拉伯芥的此種突變株,整株植物會變矮小,但外加此化合物後可恢復。

41
五、 植物對荷爾蒙的反應 (九) 植物荷爾蒙信息及環境刺激會經由細胞一系列的信息傳遞途徑(signal-transduction pathway簡稱STP),最後導致細胞的反應(celluar response)(Fig.39.3) 1.

43
六、 植物對光的反應 (Plant response to light)
")
44
六、 植物對光的反應 (一) 光形態發生(phtomorphogenesis)
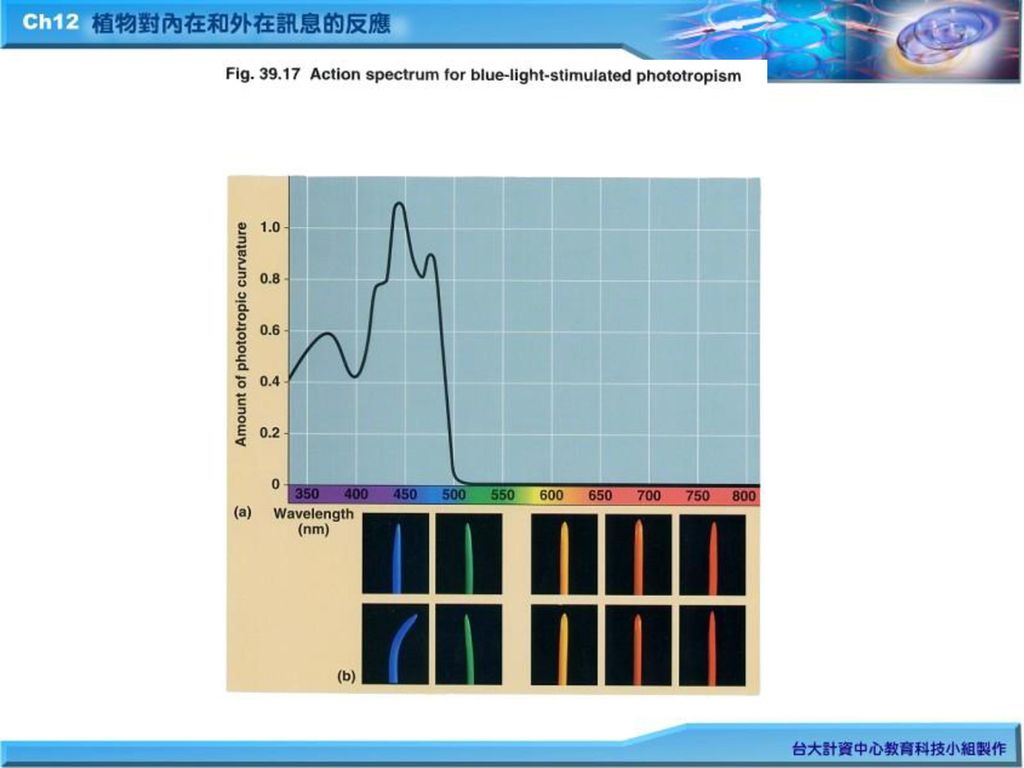
1.光對植物除引發光合作用(photosynthesis)利用太陽能把CO2固定 ,形成醣類外,光另一重要現象是影響植物的生長與發育(development)。不同的光譜作用於植物會產生不同的生長與發育現象。 2. 作用光譜(action spectrum)(Fig.39.17)
利用太陽能把CO2固定 ,形成醣類外,光另一重要現象是影響植物的生長與發育(development)。不同的光譜作用於植物會產生不同的生長與發育現象。 2. 作用光譜(action spectrum)(Fig.39.17)")
46
六、 植物對光的反應 (二)光形態發生的光接收物(Photoreceptors in phtomorphogenesis)
1.cryptochromes: 向光性(phototropism)的作用光譜是藍光(blue-light) (Fig.39.17)。早期發現藍光的phtoreceptors稱為cryptochromes。
的作用光譜是藍光(blue-light) (Fig.39.17)。早期發現藍光的phtoreceptors稱為cryptochromes。")
47
六、 植物對光的反應 在1990’s,植物分子生物學家分析阿拉伯芥突變株得知cryptochromes至少可分三種不同類型之色素:
cryptochrome (for inhibition of hypocotyl enlongation); phototropin (for phototropism), zeaxanthin (for stomatal opening):a carotenoid-based phtoreceptor。
; phototropin (for phototropism), zeaxanthin (for stomatal opening):a carotenoid-based phtoreceptor。")
48
六、 植物對光的反應 2.植物色原或光敏素(phytochromes):
(1) The phytochromes switch and seed germination(Fig.39.18) Red light(660nm)increases lettuce(萵苣)seed germination far-red light (730nm)inhibits lettuce seed germination
 The phytochromes switch and seed germination(Fig.39.18) Red light(660nm)increases lettuce(萵苣)seed germination. far-red light (730nm)inhibits lettuce seed germination.")
49
六、 植物對光的反應 (2) Phtochrome之物理化學性質:
a.The two phtoreversible variations of phtochromes(Fig.30.20) Pr(red aborbing),Pfr(far-red aborbing) Red light Pr Pfr Far-red light
 Pr(red aborbing),Pfr(far-red aborbing) Red light. Pr Pfr. Far-red light.")
51
六、 植物對光的反應 b.Structure of a phtochrome
phtochrome 是由兩個次單元之(subunit),每一單位的分子量為120Kd所組成的。在蛋白質的N-端接一個chromophore,它具有〝phtoreceptor activity〞 ,蛋白質之C端則有〝kinase activity〞(Fig.39.19)。
,每一單位的分子量為120Kd所組成的。在蛋白質的N-端接一個chromophore,它具有〝phtoreceptor activity〞 ,蛋白質之C端則有〝kinase activity〞(Fig.39.19)。")
53
六、 植物對光的反應 c.Phtochrome: a molecular switching mechanism:
Pfr form of the pigment switch on physiological and developmental responses in the plant.

54
六、 植物對光的反應 (3) The phytochrome switch and shade avoidance
Pr及Pfr二種形式的動態平衡,使植物能適應不同光條件而生存。以森林樹種之〝遮蔭〞〝shade avoidance〞現象為例,具有生態意義。

55
七、 生物時鐘控制植物及其他生物的概日韻律 (Biological clocks control circadian rythms in plants and other organisms)
")
56
七、生物時鐘控制植物及其他生物的概日韻律
(一) 以24小時為頻律的生理週期稱為概日韻律,此係來自內部的時針或某些環境的循環(例如:地球自轉,每天有晝夜)的反應,以豆科葉片為例(Fig.39.21)。
 以24小時為頻律的生理週期稱為概日韻律,此係來自內部的時針或某些環境的循環(例如:地球自轉,每天有晝夜)的反應,以豆科葉片為例(Fig.39.21)。")
58
七、生物時鐘控制植物及其他生物的概日韻律
(二) 若植物置於恆定的環境,全天的韻律會偏離24h週期(在21-27h之間)。 (三) 內源性的時鐘是什麼? The mechanism may be involve turning cellular responses on and off by means of the Pf Pfr switch
 若植物置於恆定的環境,全天的韻律會偏離24h週期(在21-27h之間)。 (三) 內源性的時鐘是什麼? The mechanism may be involve turning cellular responses on and off by means of the. Pf Pfr switch.")
59
八、 光週期(phtoperioodism)是使許多植物對季節的變化的感應產生同步化
是使許多植物對季節的變化的感應產生同步化")
60
八、光週期是使許多植物對季節的變化的感應產生同步化
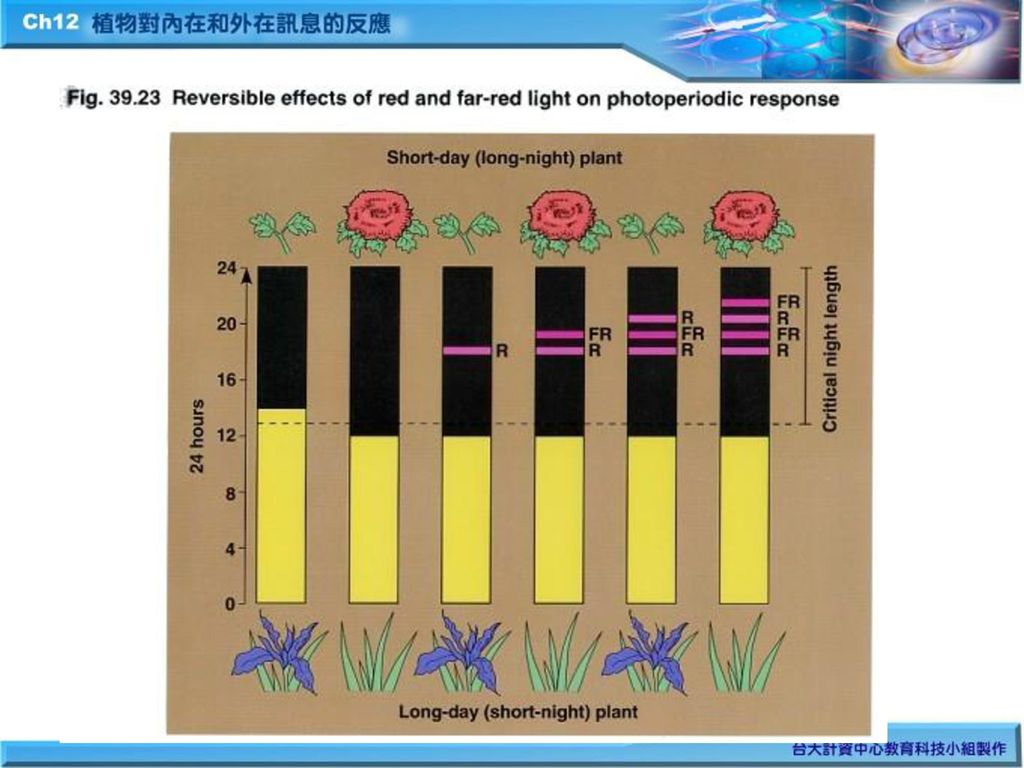
植物對光照及暗期的週期信息而引發開花的生理反應稱為光週期。 (一) 光週期與開花的控制(Fig.39.22) 1. 長日照植物(long- day plants) 2. 短日照植物(short-day plants) 3. 中性日照植物( day-neutral plants) 4. 臨界夜長(critical night lengh)
 光週期與開花的控制(Fig.39.22) 1. 長日照植物(long- day plants) 2. 短日照植物(short-day plants) 3. 中性日照植物( day-neutral plants) 4. 臨界夜長(critical night lengh)")
63
八、光週期是使許多植物對季節的變化的感應產生同步化
(二) 光週期數目與植物開花 (三) 是否有開花荷爾蒙? 以植物嫁接實驗(grafting experiment)為例(Fig.39.24) (四) 莖頂分生組織由營養生長轉變為生殖生長,前述之organ-identity genes原理
 光週期數目與植物開花. (三) 是否有開花荷爾蒙? 以植物嫁接實驗(grafting experiment)為例(Fig.39.24) (四) 莖頂分生組織由營養生長轉變為生殖生長,前述之organ-identity genes原理.")
65
九、 植物對各種環境逆境的感應 (Plant response to environmental stress)
")
66
九、植物對各種環境逆境的感應 植物經由發育與生理的控制系統(control systems),使植物能適應環境逆境造成之衝擊。
,使植物能適應環境逆境造成之衝擊。")
67
九、植物對各種環境逆境的感應 非生物性逆境(abiotic stresses)
1. 缺水反應(responses to water deficit) 2. 缺氧反應(responses to oxygen deprivation) 3. 鹽分逆境(salt stress) 4. 熱逆境(heat stress) 熱休克蛋白(heat shock proteins) 5. 低溫逆境(cold stress)
 2. 缺氧反應(responses to oxygen deprivation) 3. 鹽分逆境(salt stress) 4. 熱逆境(heat stress) 熱休克蛋白(heat shock proteins) 5. 低溫逆境(cold stress)")
69
十、植物運動(plant movements)可充當研究控制系統的模式
可充當研究控制系統的模式")
70
十、植物運動可充當研究控制系統的模式 植物對環境的刺激引起的反應,這些可經由發育上及生理上機理(physiological mechanisms)產生有韻律的運動,例如:向性(tropisms)及膨壓運動(turgor movements)。 向性(tropism)是植物器官趨向或背向刺激物的方向生長。
是植物器官趨向或背向刺激物的方向生長。")
71
十、植物運動可充當研究控制系統的模式 1.向光性(phototropism)
2.向地性(geotropism):根向地性之平衡石假說(Stalolith hypothesis)(Fig.39.25)
:根向地性之平衡石假說(Stalolith hypothesis)(Fig.39.25)")
73
十、植物運動可充當研究控制系統的模式 3.向觸性(thigmotropism)
1)植物之攀爬器官在碰到東西,其接觸會引起捲繞反應,因接觸而引生長速度不同的反應,簡稱為觸性。
植物之攀爬器官在碰到東西,其接觸會引起捲繞反應,因接觸而引生長速度不同的反應,簡稱為觸性。")
74
十、植物運動可充當研究控制系統的模式 2) 向觸形態發育(thigmomorphogenesis):由機械性刺激(例摩擦及風力等)引發發育的反應稱之。此因長期間的機械性刺激造成乙烯生成量增加所致。
 向觸形態發育(thigmomorphogenesis):由機械性刺激(例摩擦及風力等)引發發育的反應稱之。此因長期間的機械性刺激造成乙烯生成量增加所致。")
75
十、植物運動可充當研究控制系統的模式 3) Arabidopsis經接觸刺激會改變基因之表現,影響植物之生長(Fig.39.26)。
 Arabidopsis經接觸刺激會改變基因之表現,影響植物之生長(Fig.39.26)。")
77
十、植物運動可充當研究控制系統的模式 4)含羞草(Mimosa pudica)經觸摸,引起葉部motor organ之motor cells 膨壓的改變而下垂(Fig.39.27)。
含羞草(Mimosa pudica)經觸摸,引起葉部motor organ之motor cells 膨壓的改變而下垂(Fig.39.27)。")
79
十一、植物防禦:對草食性動物及對病原菌的反應 (Plant defense: Responses to herbivores and pathogen)
")
80
十一、植物防禦: 對草食性動物及對病原菌的反應
(一) 草食性動物,例如昆蟲咬傷葉片後因受傷(wounding)或唾腺(saliva)的化學物等刺激物而產生信息傳遞途徑,最後產生有毒物質,例如canavanine(arginine類似物),使昆蟲致死。
 草食性動物,例如昆蟲咬傷葉片後因受傷(wounding)或唾腺(saliva)的化學物等刺激物而產生信息傳遞途徑,最後產生有毒物質,例如canavanine(arginine類似物),使昆蟲致死。")
81
十一、植物防禦: 對草食性動物及對病原菌的反應
(二) 對病原菌的防禦,植物外表(表皮及周皮)是植物的第一層防禦系統,此層並非對病原菌或病毒不可穿過的,但當病原體進入植物體內,植物會啟發化學反應做第二道防線,受感染的植物會產生植物性殺菌素(phytoalexins)的化合物,用以摧毀或抑制病原體的生長。
 對病原菌的防禦,植物外表(表皮及周皮)是植物的第一層防禦系統,此層並非對病原菌或病毒不可穿過的,但當病原體進入植物體內,植物會啟發化學反應做第二道防線,受感染的植物會產生植物性殺菌素(phytoalexins)的化合物,用以摧毀或抑制病原體的生長。")
82
十一、植物防禦: 對草食性動物及對病原菌的反應
1. 植物病原菌之間依據gene for gene的辯認而產生抗病的能力(Fig.39.30)。
。")
84
十一、植物防禦: 對草食性動物及對病原菌的反應
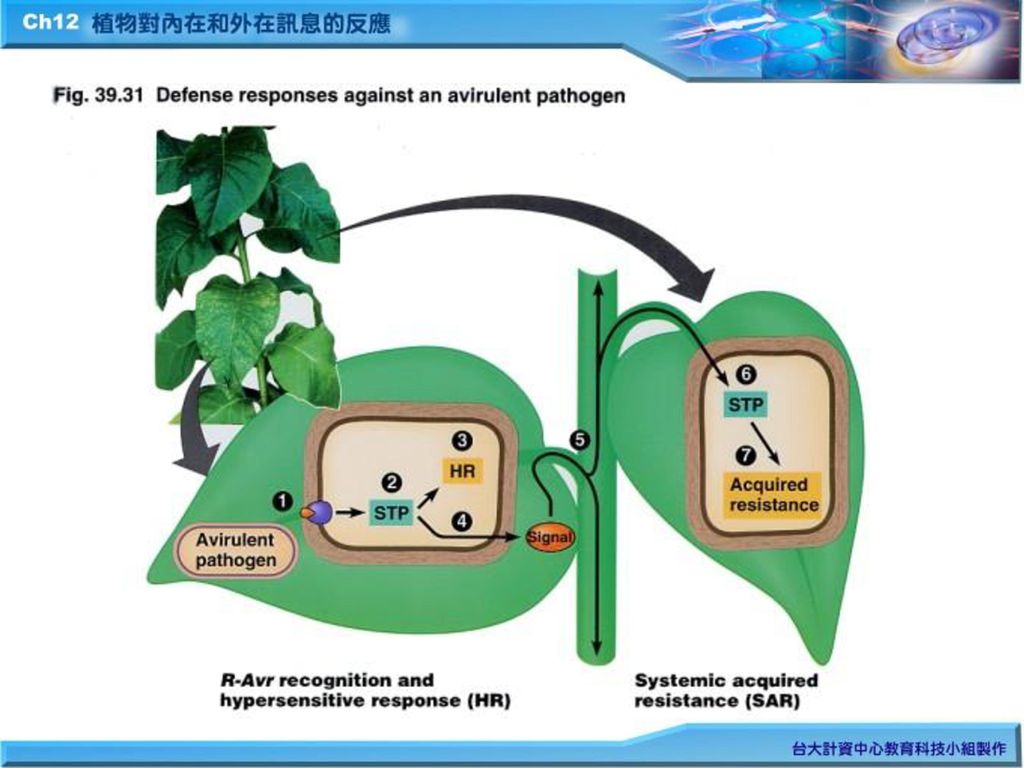
2. 過敏反應(hypersensitive response(HR):致病性病原菌感染非抗病性植物後,植物會由被感染的受傷細胞中釋放出分子信息,並經一序列signal transduction pathway(STP)而導致產生抗微生物化合物--植物性殺菌素(phytoalexins)及PR(pathogenesis-related proteins)感染亦促進細胞壁分子的橫向連接及木質素的累積,減少病原菌感染的漫延。 a. 過敏反應(hypersensitive response) b.自體獲得抗性(systemic acquired resistance)
:致病性病原菌感染非抗病性植物後,植物會由被感染的受傷細胞中釋放出分子信息,並經一序列signal transduction pathway(STP)而導致產生抗微生物化合物--植物性殺菌素(phytoalexins)及PR(pathogenesis-related proteins)感染亦促進細胞壁分子的橫向連接及木質素的累積,減少病原菌感染的漫延。 a. 過敏反應(hypersensitive response) b.自體獲得抗性(systemic acquired resistance)")
86
十二、參考資料 1 .Campbell, NA and Reece, JB(2002) Biology(6thed), Chap.39 pp publishing by Benjamin Cumming. 2.Taiz L. and Zeiger E. (2002) Plant Physiology. Published by Sinauer Associates Inc. Chap
 Plant Physiology. Published by Sinauer Associates Inc. Chap")





















 光及其生态作用 一、光在地球表面上的分布规律 紫外线(< 380nm ) 9% 、可见光( 380 ~ 760nm ) 45% 、 红外线(> 760nm ) 46% 。 1 ,在空间上的分布 纬度 海拔高度 太阳高度角 坡向 水体中 植被.>")


.>")




>")






細胞衰老(senescence) (2)基因體的不穩定(genomic instability)>")



