Download presentation
1
分子肿瘤学 第六节 细胞信号转导与肿瘤

2
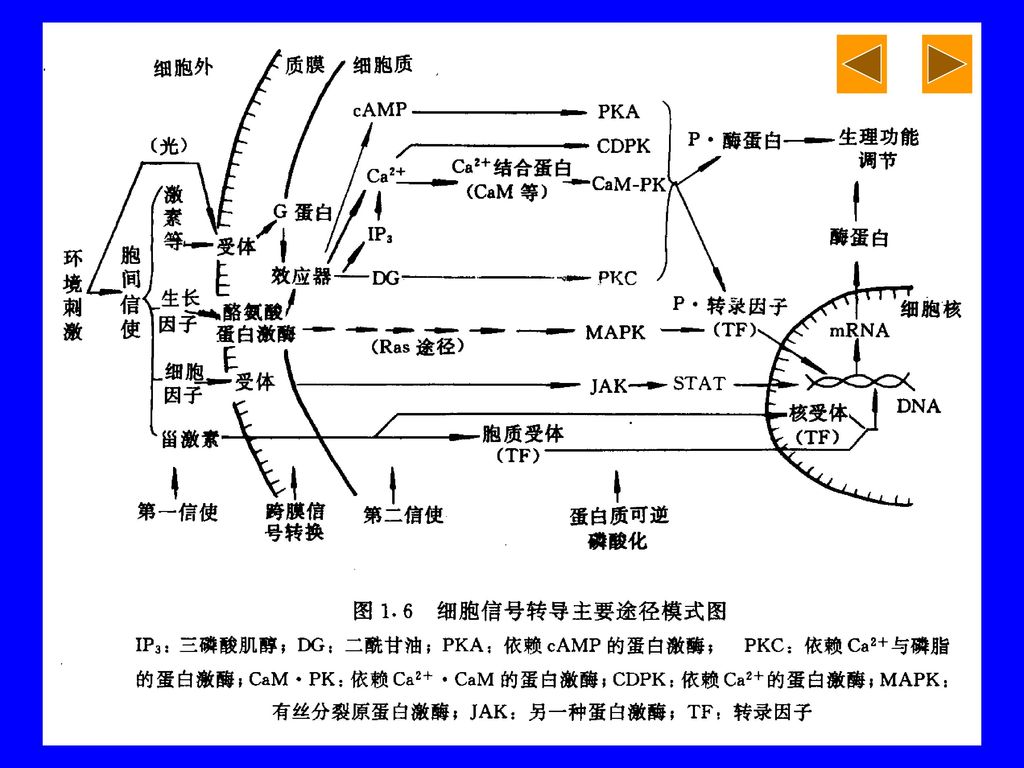
各类信号通过细胞膜和细胞内信使分子引起一系列生物化学反应,引起细胞基因表达改变,继而产生相应的生理病理效应。
外部信息传入细胞内并引起细胞应答反应的过程称为信号转导(signal transduction). 各类信号通过细胞膜和细胞内信使分子引起一系列生物化学反应,引起细胞基因表达改变,继而产生相应的生理病理效应。 细胞信号转导过程发生障碍或异常,可导致细胞生长、分化、代谢和生物学行为的异常,引起各种疾病,乃至肿瘤。
. 各类信号通过细胞膜和细胞内信使分子引起一系列生物化学反应,引起细胞基因表达改变,继而产生相应的生理病理效应。 细胞信号转导过程发生障碍或异常,可导致细胞生长、分化、代谢和生物学行为的异常,引起各种疾病,乃至肿瘤。")
3
一、信号转导异常与肿瘤概略

5
(一)促细胞增值的信号过强 1、细胞外信号分子与肿瘤 细胞外信号分子如生长因子产生过的,使细胞增值的信号转导过强,如:
(1)血小板源生长因子(platelet-derived growth factor,PDGF):某些肿瘤如肉瘤、神经胶质瘤产生和释放大量的PDGF,并表达其受体,使细胞脱离外来生长信号的调节,成为“自我刺激”生长的细胞 (2)血管内皮生长因子(vascular endothelial growth factor,VEGF) VEGF的高表达对肿瘤血管生成非常重要 (3)成纤维细胞生长因子 (fibroblast growth factor,FGF) FGF家族通过“自分泌”(Auocrine effect)机制激活受体酪氨酸激酶来参与细胞的恶性转化
血小板源生长因子(platelet-derived growth factor,PDGF):某些肿瘤如肉瘤、神经胶质瘤产生和释放大量的PDGF,并表达其受体,使细胞脱离外来生长信号的调节,成为 自我刺激 生长的细胞. (2)血管内皮生长因子(vascular endothelial growth factor,VEGF) VEGF的高表达对肿瘤血管生成非常重要. (3)成纤维细胞生长因子 (fibroblast growth factor,FGF) FGF家族通过 自分泌 (Auocrine effect)机制激活受体酪氨酸激酶来参与细胞的恶性转化.")
6
2、受体改变与肿瘤 (1)某些生长因子受体表达异常增多 如多种肿瘤组织中发现有编码EGFR的原癌基因c-erb-B的扩增及EGFR的过度表达; (2)突变使受体组成型激活 如多种肿瘤组织中证实有RTK的组成型激活

7
3、细胞内信号转导蛋白的改变 如小G蛋白(Smgs)在信号转导中起重要作用,Ras蛋白的基因突变使Ras自身GTP酶活性下降,造成Ras-Raf-MEK-ERK通路的过度激活,导致细胞的过度增殖与肿瘤发生。
在信号转导中起重要作用,Ras蛋白的基因突变使Ras自身GTP酶活性下降,造成Ras-Raf-MEK-ERK通路的过度激活,导致细胞的过度增殖与肿瘤发生。")
8
(二)抑制细胞增值的信号转导过弱 生长抑制因子受体减少、丧失 受体后信号转导通路异常 细胞的生长负调控机制减弱或丧失
抑制细胞增值的信号转导过弱 生长抑制因子受体减少、丧失 受体后信号转导通路异常 细胞的生长负调控机制减弱或丧失")
9
二、转录因子与肿瘤 一个细胞能在特定时间中多水平调控蛋白质表达,包括RNA转录、RNA加工、RNA翻译,RNA降解及翻译后水平调节,其中最为重要的是转录调控,即调控一个基因何时以及如何被转录。转录调控异常是肿瘤发病机制的重要环节。

10
(一)转录因子的几个基本问题 1、转录因子的修饰:转录因子的磷酸化是最重要的蛋白质修饰方式,蛋白质磷酸化可影响转录因子功能的多个方面
( 1)、核定位的调节:胞浆中的转录因子与抑制亚基结合,磷酸化使核定位功能区暴露,如NF-kB与IkB的结合 (2)、DNA结合活性的调节 (3)、转录活性的调节
、核定位的调节:胞浆中的转录因子与抑制亚基结合,磷酸化使核定位功能区暴露,如NF-kB与IkB的结合. (2)、DNA结合活性的调节. (3)、转录活性的调节.")
11
2、转录因子的活化方式 (2)诱导性活化:在明显外在因素影响下,细胞内某种转录因子的表达和活化显著不同于非诱导状态
(1)组成性活化( constitutive activation):在没有明显外在因素影响下,细胞中所呈现出的转录因子的活化状态 (2)诱导性活化:在明显外在因素影响下,细胞内某种转录因子的表达和活化显著不同于非诱导状态 (3)组成性激活突变(constitutively activated mutation某些信号转导蛋白在突变后获得了自 发激活和持续性激活的能力。
组成性活化( constitutive activation):在没有明显外在因素影响下,细胞中所呈现出的转录因子的活化状态. (2)诱导性活化:在明显外在因素影响下,细胞内某种转录因子的表达和活化显著不同于非诱导状态. (3)组成性激活突变(constitutively activated mutation某些信号转导蛋白在突变后获得了自 发激活和持续性激活的能力。")
12
(4)显性负性作用(dominant negative effect):某些信号转导蛋白突变后不仅自身无功能,还能抑制或阻断野生型信号转导蛋白的 作用。这种作用被称为显性负性作用。具有显性负性作用的突变被称为显性负性突变(dominant negative mutant)。
显性负性作用(dominant negative effect):某些信号转导蛋白突变后不仅自身无功能,还能抑制或阻断野生型信号转导蛋白的 作用。这种作用被称为显性负性作用。具有显性负性作用的突变被称为显性负性突变(dominant negative mutant)。")
13
NF-kB家族与肿瘤 NF-kB家族广泛调控一系列基因的表达,尤其是免疫反应、炎症反应、与病毒有关的癌基因、抑癌基因的表达。在多种肿瘤组织中NF-κB是组成性活化的, NF-kB的异常活化通过干扰细胞周期、细胞凋亡等途径在肿瘤的发生和发展中起重要作用。目前的研究表明,NF-κB参与了癌症的恶性转化,NF-κB还可能通过促进肿瘤血管生成、侵袭、转移来促进肿瘤的演进,在肿瘤的抗药机理中也起作用。

14
(一)NFκB家族及信号通路 1、NFκB家族
NFκB是由一系列同源或异源Rel家族成员形成的二聚体转录因子的总称,在哺乳动物细胞中已发现Rel/NF-kB家族的5个成员:p65(RelA)、RelB、c-Rel、p50(NF-kB1)、p52(NF-kB2)。 NFκB家族中,异源二聚物p50/RelA(p50/p65)表达最丰富,通常把p50/p65二聚体称为NFκB。
、RelB、c-Rel、p50(NF-kB1)、p52(NF-kB2)。 NFκB家族中,异源二聚物p50/RelA(p50/p65)表达最丰富,通常把p50/p65二聚体称为NFκB。")
15
2、NFκB的结构特点 NFκB 5个成员都有一个高度保守的Rel同源结构源(Rel homology domain,RHD),内含DNA结合区、蛋白二聚体化区、NFκB的抑制蛋白(IκB)结合区及核定位信号。其中p50,p52分别来源于前体蛋白p105,p100,它们的C端包含锚蛋白重复序列;而RelA,RelB及c-Rel的C端含有反式激活区域。 NFκB在DNA的特异性结合位点称κB位点,其核心结合序列为GGGACTTCC, NFκB家族成员的κB位点略有差异。
,内含DNA结合区、蛋白二聚体化区、NFκB的抑制蛋白(IκB)结合区及核定位信号。其中p50,p52分别来源于前体蛋白p105,p100,它们的C端包含锚蛋白重复序列;而RelA,RelB及c-Rel的C端含有反式激活区域。 NFκB在DNA的特异性结合位点称κB位点,其核心结合序列为GGGACTTCC, NFκB家族成员的κB位点略有差异。")
16
3、 NFκB 活化 (1)NFκB活化通路 静息状态下, NFκB在胞质中以同源或异源二聚体的形式与抑制蛋白I kB结合,呈无活性状态。在外界因素如脂多糖(LPS)、白介素1(IL1)、肿瘤坏死因子(TNFα)的刺激下,受体与配体结合,进而激活NFκB 诱导性激酶(NIK),进而激活IκB激酶(IKKα与 IKKβ)复合物,引起IκB蛋白特异丝氨酸位点 的磷酸化( IKKα Ser32 、Ser36 ,IKKβ Ser19 、Ser23) ,磷酸化IκB从三聚体中解离下来并泛素化降解,暴露p50亚基的核定位序列及p65亚基的DNA结合位点,使NFκB活化可以从胞浆移位至细胞核与DNA特异位点相结合,参与转录进程。
、白介素1(IL1)、肿瘤坏死因子(TNFα)的刺激下,受体与配体结合,进而激活NFκB 诱导性激酶(NIK),进而激活IκB激酶(IKKα与 IKKβ)复合物,引起IκB蛋白特异丝氨酸位点 的磷酸化( IKKα Ser32 、Ser36 ,IKKβ Ser19 、Ser23) ,磷酸化IκB从三聚体中解离下来并泛素化降解,暴露p50亚基的核定位序列及p65亚基的DNA结合位点,使NFκB活化可以从胞浆移位至细胞核与DNA特异位点相结合,参与转录进程。")
17
(2)NFκB的活化的反馈调节 能够上调IκBα的mRNA水平,新合成的IκBα进入胞浆与游离的NFκB结合并使之失活,细胞通过这种自身的反馈调节途径调节NFκB的活性以维持细胞的正常生理功能。
NFκB的活化的反馈调节 能够上调IκBα的mRNA水平,新合成的IκBα进入胞浆与游离的NFκB结合并使之失活,细胞通过这种自身的反馈调节途径调节NFκB的活性以维持细胞的正常生理功能。")
20
(二) NFκB信号转导通路的异常与肿瘤的发生与发展
大量研究表明,IKK/ I kB /NF-κB信号转导通路的异常可以促进肿瘤的发生发展.许多炎症因子、致癌剂、促癌剂和肿瘤微环境都可以激活NF-κB.NF-κB蛋白本身和其调控的蛋白与肿瘤的发生、增殖、抗凋亡、侵袭、血管生成和转移有关。在多种肿瘤中NF-κB都处于持续性激活状态。

21
1、癌基因介导的恶性转化及肿瘤的演进需要NF-κB的参与
Ras-转化过的细胞通过H-Ras激活NF-κB来抑制凋亡;HTLV-1TAX使得大鼠成纤维母细胞恶变也需要NF-κB的激活;癌基因激活后BCR-ABL溶合蛋白形成也需要NF-κB的激活;NF-κB的激活可能是细胞及病毒癌基因产物发挥功能的重要基础。支持这一观点的实验有:抑制了NF-κB就会抑制了许多造血系和实体瘤细胞系的生长和存活,促进凋亡。

22
2、 NF-κB活化与细胞周期异常 (1) CyclinD1 在多种肿瘤中过表达。CyclinD1启动子含有3个κB位点, NF-κB活化具有促进CyclinD1表达及G1/S期转换的功能。外界致瘤因素导致NF-κB活化继而调节CyclinD1的过表达,缩短G1期,促进细胞转化。 (2)CyclinD1启动子区还有两个AP-1结合位点,3个SP1结合位点,这为从Cross-talk角度调节CyclinD1表达,导致细胞周期紊乱提供了分子基础。
CyclinD1启动子区还有两个AP-1结合位点,3个SP1结合位点,这为从Cross-talk角度调节CyclinD1表达,导致细胞周期紊乱提供了分子基础。")
23
3、 NF-κB活化、细胞凋亡与肿瘤 细胞凋亡对机体自稳态的维持和肿瘤发生的抑制起着重要作用。NF-κB的一个重要功能是参与正常细胞凋亡的调控。 NF-κB是凋亡通路的核心调控成员,一方面受到死亡受体信号的调控,另一方面以反式激活的方式调节下游的抗凋亡基因。通过抑制细胞凋亡, NF-κB促进肿瘤的发生发展

24
(1)EB病毒、HCV 病毒等可激活NF-κB相关的信号途径,抑制Fas/TNF等介导的凋亡 (2) NF-κB 调控Survivin的表达
Survivin的启动子区具有多个转录因子结合位点,包括NF-κB、AP-1。 NF-κB使Survivin过表达,促进其核移位,使细胞Rb磷酸化增加,促进细胞S期转化,促进细胞增殖;也可通过抑制caspase3活化而抑制细胞凋亡 Survivin为靶点的抗癌治疗有可能成为肿瘤治疗的一个新的突破。

25
4、 NF -κB途径在淋巴细胞恶 性肿瘤中的作用
(1)NF -κB途径是白血病细胞增殖过程中的一个非常重要的途径 , 在 ALL和 ATL中 ,NF -κB在免疫和炎症反应中起重要作用 ,通过调节编码炎症因子基因和诱导酶 ,如诱导型一氧化氮合酶 (iNOS)和环氧合酶 - 2(COX - 2),NF-κB也参与癌症的发生 ,发展和药物抵抗。 (2)NF -κB的蛋白酶体抑制剂 PS - 341或 Bay 可以抑制 NF -κB从而抑制 ATL细胞的生长和诱导凋亡。
NF -κB途径是白血病细胞增殖过程中的一个非常重要的途径 , 在 ALL和 ATL中 ,NF -κB在免疫和炎症反应中起重要作用 ,通过调节编码炎症因子基因和诱导酶 ,如诱导型一氧化氮合酶 (iNOS)和环氧合酶 - 2(COX - 2),NF-κB也参与癌症的发生 ,发展和药物抵抗。 (2)NF -κB的蛋白酶体抑制剂 PS - 341或 Bay 可以抑制 NF -κB从而抑制 ATL细胞的生长和诱导凋亡。")
26
(3)在多发性骨髓瘤 (MM)中也有 NF -κB的激活 ,通过 PS - 341对 NF -κB的抑制和 NF -κB的抑制剂 InBa的表达 ,它使MM细胞对传统的化疗药物如 (阿霉素 )更敏感。
(4)冬凌草素甲 ,一种天然的二萜化合物,通过抑制 NF-κB和下调 Bcr- 2家族蛋白诱导恶性淋巴细胞的生长抑制 ,促使细胞凋亡。 (5)NF -κB是对 ATL和 MM等淋巴细胞恶 性肿瘤靶向治疗的一个靶点。
冬凌草素甲 ,一种天然的二萜化合物,通过抑制 NF-κB和下调 Bcr- 2家族蛋白诱导恶性淋巴细胞的生长抑制 ,促使细胞凋亡。 (5)NF -κB是对 ATL和 MM等淋巴细胞恶 性肿瘤靶向治疗的一个靶点。")
27
4、 NF –κB是维持LSC生存的要素 在维持LSC生存的要素中NF –κB和PI3k通路是很重要。在AML LSC中, NF –κB是持续激活的,抑制NF –κB的药物MG-132能诱导LSC凋亡,却不能影响正常HSC,因为正常HSC在没有丝裂原的刺激下, NF –κB是不激活的

28
(三) NF –κB及其他转录因子研究常用的一项方法-EMSA
凝胶迁移或电泳迁移率实验(EMSA) : (EMSA-electrophoretic mobility shift assay)是一种研究DNA结合蛋白和其相关的DNA结合序列相互作用的技术,可用于定性和定量分析。这一技术最初用于研究DNA结合蛋白,目前已用于研究RNA结合蛋白和特定的RNA序列的相互作用。 通常将纯化的蛋白和细胞粗提液和32P同位素标记的DNA或RNA探针一同保温,在非变性的聚丙烯凝胶电泳上,分离复合物和非结合的探针。DNA-复合物或RNA-复合物比非结合的探针移动得慢。同位素标记的探针依研究的结合蛋白的不同,可是双链或者是单链。当检测 如转录调控因子一类的DNA结合蛋白,可用纯化蛋白,部分纯化蛋白,或核细胞抽提液。在检测RNA结合蛋白时,依据目的RNA结合蛋白的位置,可用纯化或部分纯化的蛋白,也可用核或胞质细胞抽提液。竞争实验中采用含蛋白结合序列的DNA或RNA片段和寡核苷酸片段(特异), 和其它非相关的片段(非特异),来确定DNA或RNA结合蛋白的特异性。在竞争的特异和非特异片段的存在下,依据复合物的特点和强度来确定特异结合。
 : (EMSA-electrophoretic mobility shift assay)是一种研究DNA结合蛋白和其相关的DNA结合序列相互作用的技术,可用于定性和定量分析。这一技术最初用于研究DNA结合蛋白,目前已用于研究RNA结合蛋白和特定的RNA序列的相互作用。 通常将纯化的蛋白和细胞粗提液和32P同位素标记的DNA或RNA探针一同保温,在非变性的聚丙烯凝胶电泳上,分离复合物和非结合的探针。DNA-复合物或RNA-复合物比非结合的探针移动得慢。同位素标记的探针依研究的结合蛋白的不同,可是双链或者是单链。当检测 如转录调控因子一类的DNA结合蛋白,可用纯化蛋白,部分纯化蛋白,或核细胞抽提液。在检测RNA结合蛋白时,依据目的RNA结合蛋白的位置,可用纯化或部分纯化的蛋白,也可用核或胞质细胞抽提液。竞争实验中采用含蛋白结合序列的DNA或RNA片段和寡核苷酸片段(特异), 和其它非相关的片段(非特异),来确定DNA或RNA结合蛋白的特异性。在竞争的特异和非特异片段的存在下,依据复合物的特点和强度来确定特异结合。")
31
Ets转录因子家族在肿瘤发生中的作用 Ets基因最早发现于美国Frederick的国家癌症研究所分子肿瘤学实验室.通过研究禽类逆转录病毒-E26,研究者发现了一系列拥有与E26高度保守的同源序列的基因,根据E26 (E-twenty six)的缩写而将该基因命名为Ets. 该基因的共同特点是含有高度保守的DNA结合域,能够和特定序列结合调控靶基因的表达和功能.通过调节细胞的增生、分化、凋亡及上皮-间充质间的相互作用,Ets参与许多生理和病理过程. 大量研究发现,Ets在哺乳动物的发育和肿瘤的侵袭转移中发挥重要的调控作用。
的缩写而将该基因命名为Ets. 该基因的共同特点是含有高度保守的DNA结合域,能够和特定序列结合调控靶基因的表达和功能.通过调节细胞的增生、分化、凋亡及上皮-间充质间的相互作用,Ets参与许多生理和病理过程. 大量研究发现,Ets在哺乳动物的发育和肿瘤的侵袭转移中发挥重要的调控作用。")
32
(一)Ets转录因子家族 人类Ets家族基因在进化上高度保守,其共同点是均含有一个大约85个氨基酸的DNA结合域,也就是位于C-端的Ets区. 该区富含精氨酸和赖氨酸残基,为螺旋-转角-螺旋基序,可识别并结合富含嘌呤的DNA核心序列A/CGGAA/T,这一序列存在于许多基因的5’-侧翼调节区,有反式活化功能.目前Ets转录因子家族包括30多个成员,根据ETS结构域构象和位置不同,Ets家族分为多个亚家族。

33
亚族 组成 Ets Ets1 Ets2 EGR Erg2 Fli-1 FevELG GABPa ETS4 TEL(ETV6) PEA3 E1AF ERM ETV1 ERB1 TCFs E1k-1 Sapla NET/ERP/Sap Elf Elf-1 NERF1b MEF Spi PU.1 Spi-BERF ERF PE-1ESX ESX/ESE-1
 PEA3 E1AF ERM ETV1 ERB1. TCFs E1k-1 Sapla NET/ERP/Sap. Elf Elf-1 NERF1b MEF. Spi PU.1 Spi-BERF ERF PE-1ESX ESX/ESE-1.")
34
(二)Ets转录因子家族成员在胚胎发育中的调节作用
Ets转录因子家族成员在胚胎发育中的调节作用")
35
(三)Ets转录因子家族在肿瘤发生 中的调节作用
Ets基因在整个进化过程中是一个保守基因,其正常功能发生改变时,可导致分化和发育异常及肿瘤发生,在某些类型的肿瘤中,有许多与肿瘤相关的Ets突变发生,一个易位染色体将Ets基因片段与一个不相关的基因融合,导致了嵌合癌蛋白的表达,该类蛋白分为两类,第一类Ets蛋白可使他们的癌基因效应作为转录因子,来下调Ets调控的基因,这对细胞周期的调控十分重要. 第二类Ets嵌合肿瘤蛋白,通过永久的激活某些信号转导途径或抑制一个关键转录因子的活性来促进癌基因

36
1、Ets家族的重要成员Ets-1的表达与胰腺癌、甲状腺癌、乳腺癌、胃癌、子宫内膜癌、口腔鳞癌、肝细胞性肝癌(HCC)的发生、转移显著相关.
2、Erg基因在髓细胞白血病、Ewing’肉瘤、儿童长骨肿瘤中发生重排. 3、发生浸润的肿瘤组织基质细胞中存在Ets1和MMP-1(Matrix Metalloproteinase 1)、MP-3、MMP-7共表达,并且随肿瘤的浸润转移,这些基因表达明显上调,而在良性和非浸润的肿瘤中,就很少检测到Ets1
、MP-3、MMP-7共表达,并且随肿瘤的浸润转移,这些基因表达明显上调,而在良性和非浸润的肿瘤中,就很少检测到Ets1.")
37
4、血管生成是多阶段过程,其中Ets参与了内皮细胞的增殖和迁移过程。阻断Ets1表达可抑制血管上生成
5、Ets也通过调节癌基因或抑癌基因的表达来参与肿瘤的发生,。如Met(控制侵袭性生长的癌基因)、N-Myc等
、N-Myc等.")
38
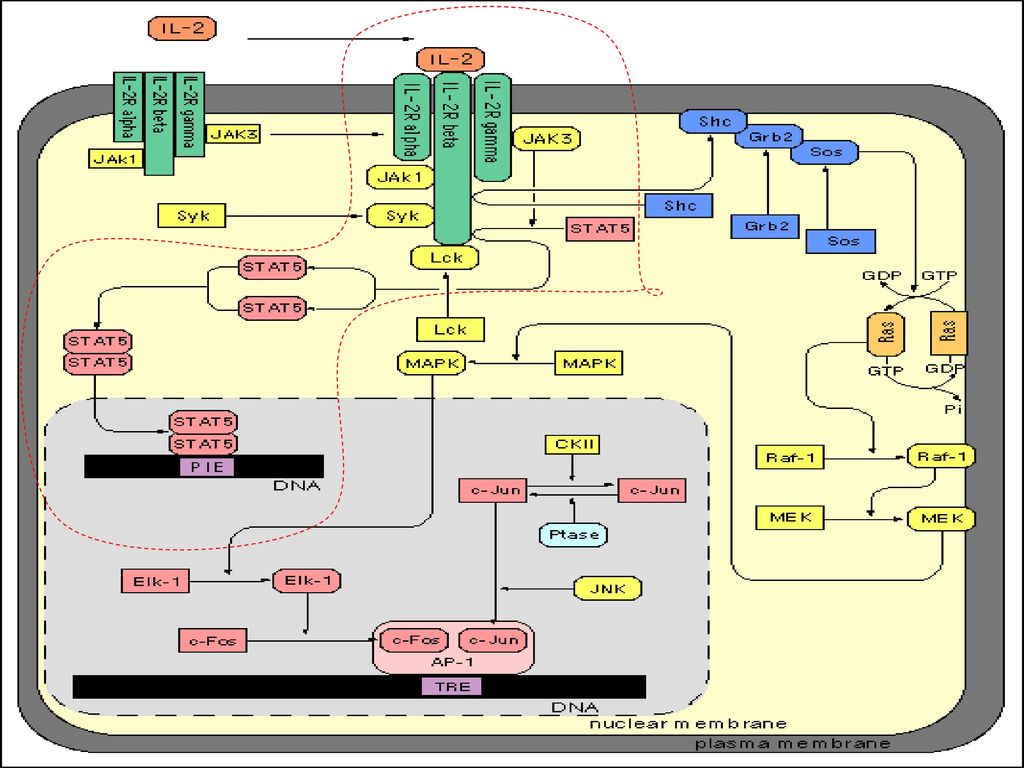
JAK-STAT传导途径与肿瘤 JAK可与多种细胞因子受体结合 ,并选择性地激活其下游底物 —STAT ,使之转位到核 ,与核内特异的 DNA 调节元件结合而指导转录。这一信号传递途径称之为JAK-STAT途径。 JAK-STAT传导途径的异常与肿瘤的发生发展关系密切。

39
(一)JAK家族 (1)JAK是 just another kinase 或 janus kinase 的简称 ,是一种酪氨酸激酶。 JAK家族目前共发现四种:JAK1 , JAK2 , JAK3和 Tyk2 。JAK家族的特征是在 C端有 2 个功能区 ,即催化功能区和激酶相关功能区 ,而其N端在JAK与受体蛋白偶联的过程中能发挥调节作用 。JAK几乎在所有的细胞中表达 ,根据细胞受体结构的不同 ,JAK以不同的方式与之结合。
JAK是 just another kinase 或 janus kinase 的简称 ,是一种酪氨酸激酶。 JAK家族目前共发现四种:JAK1 , JAK2 , JAK3和 Tyk2 。JAK家族的特征是在 C端有 2 个功能区 ,即催化功能区和激酶相关功能区 ,而其N端在JAK与受体蛋白偶联的过程中能发挥调节作用 。JAK几乎在所有的细胞中表达 ,根据细胞受体结构的不同 ,JAK以不同的方式与之结合。")
40
JAK1 与 IFNα、IFNβ、GM-CSF、IL22等 受体 JAK2 与GM-CSF、Epo、IL23等受体
JAK3与 IL22、IL24、IL21等受体 Tyk2 与IFNα等受体 均可发生偶联反应。由于调控造血的细胞因子受体缺乏酪氨酸激酶区域 ,因此JAK参与的酪氨酸磷酸化就显得尤为重要 , 其中 JAK2与造血因子的关系似乎最为密切。

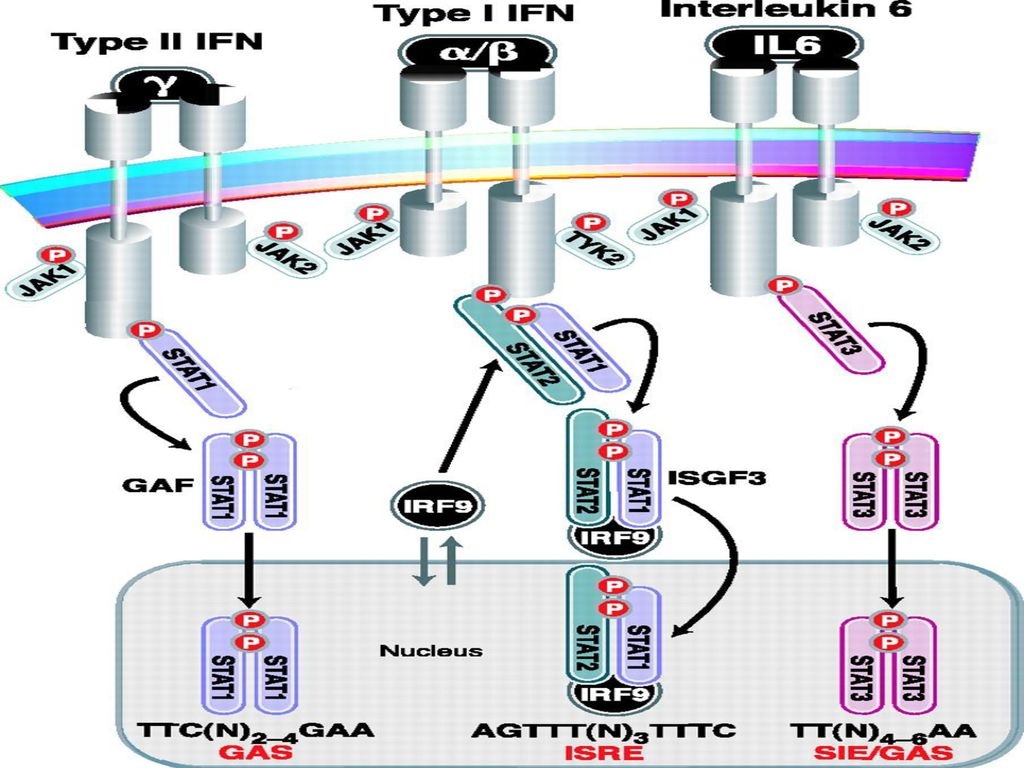
41
(二)STAT家族 STAT (signal transducers and activators of transcription, 信号转导子和转录活化子), 是在研究 IFN对基因表达的调控中被认识的 ,是一种能与 DNA 结合的独特蛋白家族。该家族有六个成员 : STAT1(STAT1α/β) , STAT2 STAT3 STAT4 STAT5(α/β) STAT6
 , STAT2. STAT3. STAT4. STAT5(α/β) STAT6.")
42
(三)JAK-STAT途径的激活 细胞因子的受体本身如果不具有酪氨酸激酶的作用 ,则在被活化后 ,即具备了激活与其相连接的一种或几种JAK的能力。JAK几乎在所有的细胞中表达 ,一般在靠近膜的部位。它被活化后 ,能介导胞浆中 STAT的 SH2 功能区的 C端酪氨酸残基磷酸化。STAT被磷酸化后 ,即脱离它所结合的受体(在受体活化过程中 ,STAT与受体发生短暂结合) ,以同源或异源二聚体的形式游向并定位于细胞核 ,与特异的DNA元件结合并激活靶基因的转录 ,这就是JAK-STAT途径的激活。
 ,以同源或异源二聚体的形式游向并定位于细胞核 ,与特异的DNA元件结合并激活靶基因的转录 ,这就是JAK-STAT途径的激活。")
43
JAK属非受体酪氨酸激酶,底物为STAT,即信号转导子和转录激活子。
JAK/STAT Pathway

46
(四) JAK-STAT传导途径与肿瘤的发生发展
多种肿瘤细胞系和实体肿瘤中有STATs的 活化,尤其是STAT3,目前倾向认为是一 种癌基因。 在多种人类血液疾病包括白血病和淋巴瘤 中有特定的JAK、STAT活化 STAT3组成性活化可能在具有激活的生长 因子受体信号途径的肿瘤发生发展中起关 键作用

47
subsequent JAK activation trigger a cascade of signaling
JAK proteins,are stringently maintained in inactive complexes . Cytokine-mediated receptor activation and subsequent JAK activation trigger a cascade of signaling events, which control survival, proliferation and differen- tiation of different cell types. During the past several years, several JAK mutations have been identified that encode constitutively active or hyperactive JAK proteins, which have crucial roles in hematopoietic malignancies, especially myeloproliferative neoplasms.

48
When expressed in cells and bound to membrane
cytokine receptors, mutant JAK proteins are no longer maintained in an inactive state, and they persistently activate STAT proteins . This activity promotes oncogenic transformation and uncontrolled blood cell production. The role of JAK–STAT mutations in cancer has only recently begun to be explored, and it is possible that mutations in components of this pathway are present in a wide variety of cancers. -

49
Mutations in JAKs and disease association
JAKs Substitution Disease association JAK2 (human) V617F MPDs K539L (and the region F537–E543) PV T875N Acute megakaryoblastic myeloid leukemia Deletion of IREED Acute lymphoblastic leukemia (position 682–686) JAK3 (human) A572V Acute megakaryoblastic myeloid leukemia V722I Acute megakaryoblastic myeloid leukemia P132T Acute megakaryoblastic myeloid leukemia JAK1 (human) G871E Uterine leiomyosarcomas [45]
 V617F MPDs. K539L (and the region F537–E543) PV. T875N Acute megakaryoblastic myeloid leukemia. Deletion of IREED Acute lymphoblastic leukemia. (position 682–686) JAK3 (human) A572V Acute megakaryoblastic myeloid leukemia. V722I Acute megakaryoblastic myeloid leukemia. P132T Acute megakaryoblastic myeloid leukemia. JAK1 (human) G871E Uterine leiomyosarcomas [45]")
51
可能机制: 1、STAT活化与肿瘤细胞凋亡异常: STAT3 / STAT5可通过抑制凋亡来介导细胞恶性演进;
STAT3/Bcl-xL信号通路在凋亡中发挥重要作用 2、STAT活化与肿瘤细胞周期异常: CyclinD1与 CDK4和 CDK6结合有助于G1/S期进程,在 CyclinD1调控区的-481位有STAT5的结合位点,提示STAT5 通过介导CyclinD1表达刺激细胞增殖导致癌变。 Cyclin B及A是G2/M期转换过程中的调控蛋白, Cyclin B起 关键作用,STAT5还参与了Cyclin B的调控。 此外, STAT3还能调控CDKI(如p21等)
")
52
3、STAT活化与肿瘤细胞侵袭与转移 肿瘤细胞侵袭与转移包括细胞的黏附、运动、细胞外基质降解、 血管生成等一系列病理生理过程。 STAT3可通过调节EGFR(成纤维生长因子受体)和PI3K(磷脂酰肌醇(-3)激酶)使β-catenin(连环蛋白)磷酸化后从细胞膜释放,从而使细胞间黏附能力下降。 在黑色素瘤细胞中, STAT3可活化金属蛋白酶-2(MMP2),降解细胞外基质。 VEGF(血管内皮生长因子)是JAK-STAT信号途径下游一个重要的调控基因,是最重要的血管生长因子。 STAT3可直接作用于其启动子,上调其表达
,降解细胞外基质。 VEGF(血管内皮生长因子)是JAK-STAT信号途径下游一个重要的调控基因,是最重要的血管生长因子。 STAT3可直接作用于其启动子,上调其表达.")
53
4、STAT活化与肿瘤细胞免疫逃逸: STAT3通过促进DC细胞的分化成熟及参与其异常分化,导致T细胞免疫识别障碍,从而使肿瘤细胞获得免疫耐受。

54
转录因子研究展望 蛋白质组学(proteomics)时代的到来,为研究带来了挑战和机遇,信号转导蛋白质组学,简称信号组学(signalomics)将突破传统信号传导研究的局限性 目前有2000多个未知功能基因编码的转录因子,阐明其功能及与肿瘤发生发展的关系,将为以转录因子为靶点的肿瘤治疗提供线索 注重研究致瘤因素导致的转录因子翻译后修饰(通常为磷酸化修饰)所赋予转录因子的新功能 在肿瘤研究中,通过转录因子,将信号转导与细胞周期研究有机结合是目前的持续热点
所赋予转录因子的新功能. 在肿瘤研究中,通过转录因子,将信号转导与细胞周期研究有机结合是目前的持续热点.")






的作用而发生一系列质的 改变,形成一种异常的增生。这种异常 增生一旦形成,即使外界至病因素停止 作用,也还能继续自然发展,这说明肿 瘤是整体性疾病的一种局部表现。>")
 谢喜秀 1040800114.>")

染色体结构不同; ( 2 )原核生物具有正调控和负调控并重的特点,真核 生物目前已知的主要是正调控; ( 3 )原核生物的转录和翻译是相偶联的,真核生物的.>")
是血管外科常见病 和多发病,其发病率为 15 % ~20 %,占全部下肢静 脉疾病的 70% 。 因持续性的静脉高压,可导致下肢水肿、疼痛、 静脉曲张、皮肤色素改变甚至溃疡形成,严重影 响患者的生活质量。 2016-8-13.>")
 (一)概述: ﹡ 细胞原癌基因 启动子插入 染色体移位 活化 点突变 基因扩增 癌基因 转录合成 生长因子 生长因子受体 癌基因产物(蛋白) 传导因子 核蛋白 转化蛋白 细胞恶性转化.>")












 白血病 第三組 組員名單:洪世烜 4A (組長) 徐德享 4A 黃嘉偉 4A090132>")
