Download presentation
Presentation is loading. Please wait.

1
第十三章 DNA的损伤修复

2
第一节 DNA损伤 第二节 DNA修复 第三节 基因突变

3
第一节 DNA的损伤

4
DNA存储着生物体赖以生存和繁衍的遗传信息,因此维护DNA分子的完整性对细胞至关重要。
在生物进化中突变又是与遗传相对立统一而普遍存在的现象。

5
1.自发性损伤 1.1 DNA复制中的错误 碱基配对的错误频率约为10-4-10-5;

6
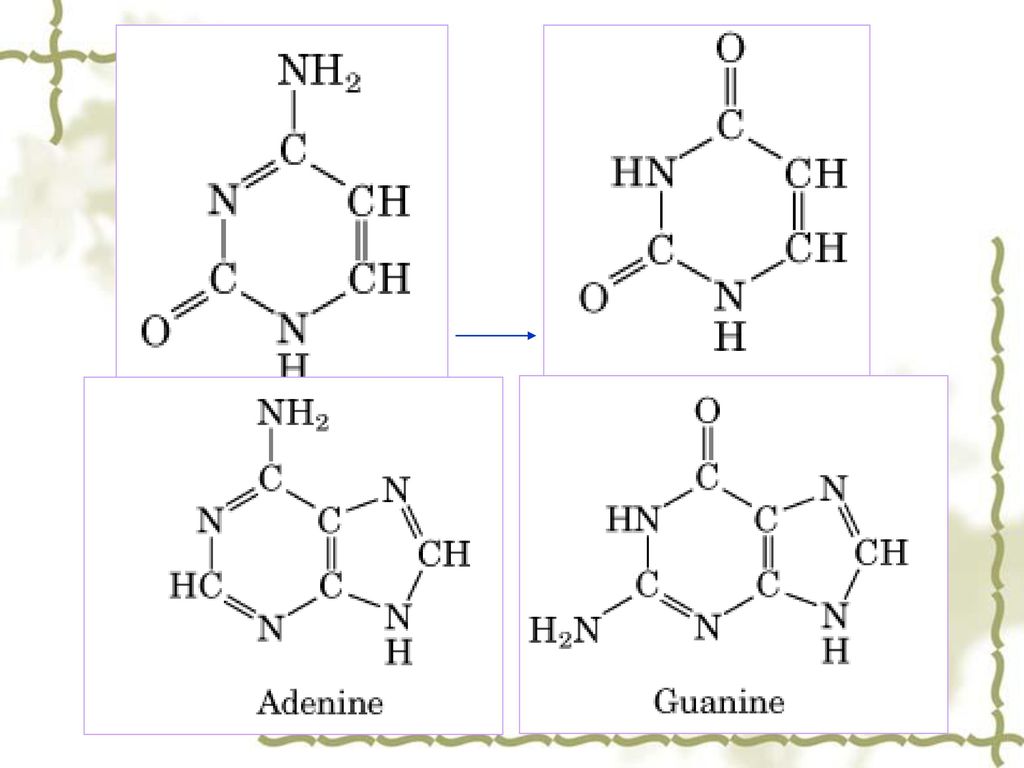
异构体间自发地相互变化形成导致下一世代中G-C配对取代A-T配对。
1.2 碱基的自发性化学变化 a. 碱基的异构互变 DNA中的4种碱基各自的异构体间自发地相互变化(例如烯醇式与酮式碱基间的互变),使碱基配对间的氢键改变。 腺嘌呤的稀有互变异体与胞嘧啶 胸腺嘧啶的稀有互变异构体与鸟嘌呤 异构体间自发地相互变化形成导致下一世代中G-C配对取代A-T配对。
,使碱基配对间的氢键改变。 腺嘌呤的稀有互变异体与胞嘧啶. 胸腺嘧啶的稀有互变异构体与鸟嘌呤. 异构体间自发地相互变化形成导致下一世代中G-C配对取代A-T配对。")
7
b.碱基的脱氨基(deamination)作用
碱基的环外氨基有时会自发脱落,从而胞嘧啶会变成尿嘧啶、腺嘌呤会变成次黄嘌呤(H)、鸟嘌呤会变成黄嘌呤(X)等。 胞嘧啶自发脱氨基的频率约为每个细胞每天190个。
、鸟嘌呤会变成黄嘌呤(X)等。 胞嘧啶自发脱氨基的频率约为每个细胞每天190个。")
8
c.脱嘌呤(depurination)与脱嘧啶
自发的水解可使嘌呤和嘧啶从DNA链的核糖磷酸骨架上脱落下来。 一个哺乳类细胞37℃,20h内DNA链自发脱落的嘌呤约1000个、嘧啶约500个, 长寿命不复制繁殖的哺乳类细胞(如神经细胞)在整个生活期间自发脱嘌呤数约为108,约占细胞DNA中总嘌呤数的3%。
在整个生活期间自发脱嘌呤数约为108,约占细胞DNA中总嘌呤数的3%。")
9
d.碱基修饰与链断裂 细胞呼吸的副产物O2、H2O2等会造成DNA损伤,能产生胸腺嘧啶乙二醇、羟甲基尿嘧啶等碱基修饰物,引起DNA单链断裂等损伤 每个哺乳类细胞每天DNA单链断裂发生的频率约为5万次。 DNA的甲基化、结构的其他变化等,这些损伤的积累可能导致老化。

10
2. 物理因素引起的DNA损伤 2.1 紫外线引起的DNA损伤
紫外线照射,同一条DNA链上相邻的嘧啶以共价键连成二聚体,相邻的两个T、或两个C、或C与T间都可以环丁基环(cyclobutane ring)连成二聚体。 图 胸腺嘧啶二聚体的形成
连成二聚体。 图 胸腺嘧啶二聚体的形成.")
11
人皮肤因受紫外线照射而形成二聚体的频率可达每小时5×104/细胞,只局限在皮肤中。
微生物受紫外线照射后,会影响其生存。 紫外线照射还能引起DNA链断裂等损伤。

12
2.2 电离辐射引起的DNA损伤 直接效应是DNA直接吸收射线能量而遭损伤,
主要是由OH-自由基引起,包括DNA链上的碱基氧化修饰、过氧化物的形成、碱基环的破坏和脱落等。 一般嘧啶比嘌呤更敏感。

13
b.脱氧核糖变化 脱氧核糖上的每个碳原子和羟基上的氢都能与OH-反应,导致脱氧核糖分解,引起DNA链断裂。 c.DNA链断裂 射线的直接和间接作用都可能使脱氧核糖破坏或磷酸二酯键断开。 单链断裂(single strand broken), 双链断裂(double strand broken)。 单链断裂发生频率为双链断裂的10-20倍,但比较容易修复;对单倍体细胞来说(如细菌)一次双链断裂就是致死事件。
。 单链断裂发生频率为双链断裂的10-20倍,但比较容易修复;对单倍体细胞来说(如细菌)一次双链断裂就是致死事件。")
14
DNA-蛋白质交联:组蛋白、染色质中的非组蛋白、调控蛋白、与复制和转录有关的酶都会与DNA共价键连接。
DNA链交联:同一条DNA链上或两条DNA链上的碱基间可以共价键结合, DNA-蛋白质交联:组蛋白、染色质中的非组蛋白、调控蛋白、与复制和转录有关的酶都会与DNA共价键连接。 3.化学因素引起的DNA损伤 突变剂或致癌剂对DNA的作用。 3.1 烷化剂对DNA的损伤 是一类亲电子的化合物,很容易与生物体中大分子的亲核位点起反应。 烷化剂的作用可使DNA发生各种类型的损伤:

15
a.碱基烷基化 烷化剂如甲基黄酸乙脂(EMS),氮芥(NM),甲基黄酸甲脂(MMS),亚硝基胍(NG)等,它们的作用是使碱基烷基化,将烷基加到DNA链中嘌呤或嘧啶的N或O上; EMS使G的第6位烷化,使T的第4位上烷化,结果产生的O-6-E-G和 O-4-E-T分别和T、G配对,导致G∶C对转换成A∶T对;T∶A对转换成C∶G

16
b.碱基脱落 烷化鸟嘌呤的糖苷键不稳定,容易脱落形成DNA上无碱基的位点,复制时可以插入任何核苷酸,造成序列的改变。 c.断链 DNA链的磷酸二酯键上的氧也容易被烷化,形成不稳定的磷酸三酯键,易在糖与磷酸间发生水解,使DNA链断裂。

17
d.交联 烷化剂有两类: 单功能基烷化剂,如甲基甲烷碘酸,只能使一个位点烷基化;
双功能基烷化剂,化学武器如氮芥、硫芥等,一些抗癌药物如环磷酰胺、苯丁酸氮芥、丝裂霉素等,某些致癌物如二乙基亚硝胺等均属此类,其两个功能基可同时使两处烷基化,结果就能造成DNA链内、DNA链间,以及DNA与蛋白质间的交联。

18
图3 氮芥引起DNA分子两条链在鸟嘌呤上的交联 (a)交联附近的总图;(b)交联部分结构图
交联附近的总图;(b)交联部分结构图")
19
3.2 碱基类似物对DNA的损伤 人工合成的一些碱基类似物,用作促突变剂或抗癌药物,如5-溴尿嘧啶(5-BU)、5-氟尿嘧啶(5-FU)、2-氨基腺嘌呤(2-AP)等。 其结构与正常的碱基相似,进入细胞能替代正常的碱基参入到DNA链中而干扰DNA复制合成。
、5-氟尿嘧啶(5-FU)、2-氨基腺嘌呤(2-AP)等。 其结构与正常的碱基相似,进入细胞能替代正常的碱基参入到DNA链中而干扰DNA复制合成。")
21
其它诱发突变的化学物质或致癌剂: 例如亚硝酸盐能使C脱氨变成U,经过复制就可使DNA上的G-C变成A-T; 羟胺能使T变成C,结果是A-T改成C-G; 黄曲霉素B也能专一攻击DNA上的碱基导致序列的变化。

22
第二节 DNA修复

23
DNA修复(DNA repairing)是细胞对DNA受损伤后的一种反应,这种反应可能使DNA结构恢复原样,重新能执行它原来的功能;
修复系统: 切除修复 回复修复 错配修复 SOS修复 重组修复 限制修饰系统-----对付外源DNA的入侵

24
1 切除修复(excision repair)
对多种DNA损伤包括碱基脱落形成的无碱基位点、嘧啶二聚体、碱基烷基化、单链断裂、交联等都能起修复作用。 这种修复方式普遍存在于各种生物细胞中,也是人体细胞主要的DNA修复机制。 需要多种酶作用,复制前进行 。 1.1 核苷酸切除修复 1.1.1 大肠杆菌核苷酸切除修复有关的酶 1) UvrABC内切酶或切除酶: 需要ATP的结合但不需要ATP水解; 是uvrA,uvrB和uvrC三个基因表达产物的复合物; 在损伤位置两侧各作一缺口(UvrB在3’端,UvrC在5’端)。
 UvrABC内切酶或切除酶: 需要ATP的结合但不需要ATP水解; 是uvrA,uvrB和uvrC三个基因表达产物的复合物; 在损伤位置两侧各作一缺口(UvrB在3’端,UvrC在5’端)。")
25
2)解链酶Ⅱ(UvrD编码): 去除UvrABC内切酶产生的寡核苷酸; 3)DNA聚合酶(DNA pol) 4)DNA连接酶(DNA Ligase) 人类核苷酸切除修复有关的基因: (XPA,XPB,XPC,XPD,XPF,XPG)缺陷 引起人类着色性干皮病。 临床表现 :严重光敏感,皮癌,视力缺陷,神经错乱。

26
1.1.2 核苷酸切除修复过程 切口

27
Uvr系统在修复各阶段中的作用: UvrA识别损伤; UvrBC在DNA上切一缺口;
UvrD解旋切口区域的DNA,并释放出切割的DNA片段。

28
1) DNA糖基化酶(DNA glycosylase):
1.2 碱基切除修复 1.2.1 碱基切除修复有关的酶 1) DNA糖基化酶(DNA glycosylase): 识别DNA中损伤的或错误的碱基;水解N-糖苷键去除碱基。 2) AP内切酶(Endonucleases) : 有3’5’外切酶活性,切除AP位点5’端的数个核苷酸。 DNA糖基化酶去除碱基后,产生一个AP位点。 AP内切酶识别此位点并在AP位点的5’端产生一个切口。 3) 脱氧核糖磷酸二酯酶(dRpase): 切除AP位点的脱氧核糖磷酸,产生一个核苷酸的缺口。 4) DNA聚合酶: 在缺口处进行修复,加入一个核苷酸。在AP位点5’端进行广泛的修复合成。 5) DNA连接酶:连接最后一个切口。
 DNA糖基化酶(DNA glycosylase): 识别DNA中损伤的或错误的碱基;水解N-糖苷键去除碱基。 2) AP内切酶(Endonucleases) : 有3’5’外切酶活性,切除AP位点5’端的数个核苷酸。 DNA糖基化酶去除碱基后,产生一个AP位点。 AP内切酶识别此位点并在AP位点的5’端产生一个切口。 3) 脱氧核糖磷酸二酯酶(dRpase): 切除AP位点的脱氧核糖磷酸,产生一个核苷酸的缺口。 4) DNA聚合酶: 在缺口处进行修复,加入一个核苷酸。在AP位点5’端进行广泛的修复合成。 5) DNA连接酶:连接最后一个切口。")
29
碱基切除修复过程 dRpase DNA糖苷酶 5´AP内切酶 切除的游离碱基 寡核苷酸切除产物 切除的 5´-脱氧核糖磷酸 5´ 3´ 5´
烷化碱基 DNA糖苷酶 5´ 3´ 切除的游离碱基 无嘌呤嘧啶位点 OH P- 5´ 3´ 5´ 3´ 5´AP内切酶 AP内切酶的 3´-5´外切酶活性 dRpase OH P 5´ 3´ OH P 5´ 3´ 3´ P 寡核苷酸切除产物 切除的 5´-脱氧核糖磷酸

30
基本步骤: ① 首先由核酸酶识别DNA的损伤位点,在损伤部位的5′侧切开磷酸二酯键。 ② 由5′→3′核酸外切酶将有损伤的DNA片段切除。 ③ 在DNA聚合酶的催化下,以完整的互补链为模板,按5′→3′方向DNA链,填补已切除的空隙。 ④ 由DNA连接酶将新合成的DNA片段与原来的DNA断链连接起来。 最终使DNA恢复原来的结构。 DNA损伤后切除修复

31
2 回复修复/直接修复(direct repair)
指无需去除碱基或核苷酸,只需一种酶经一步反应修复DNA损伤的修复机制。 较简单的修复方式,一般都能将DNA修复到原样。 2.1 烷基的转移 O6甲基鸟嘌呤甲基转移酶(MGMT),能直接将DNA链鸟嘌呤O6位上的甲基移到酶的半胱氨酸残基上而修复损伤的DNA。 这个酶的修复能力并不很强,但在低剂量烷化剂作用下此酶有修复活性。
,能直接将DNA链鸟嘌呤O6位上的甲基移到酶的半胱氨酸残基上而修复损伤的DNA。 这个酶的修复能力并不很强,但在低剂量烷化剂作用下此酶有修复活性。")
32
2.2 光修复 由细菌中的DNA光解酶(photolyase)完成:
此酶能特异性识别紫外线造成的核酸链上相邻嘧啶共价结合的二聚体,并与其结合,这步反应不需要光; 结合后如受300-600nm波长的光照射,则此酶就被激活,将二聚体分解为两个正常的嘧啶单体,然后酶从DNA链上释放,DNA恢复正常结构。

33
2.3 单链断裂的重接 2.4 碱基的直接插入 DNA单链断裂,其中一部分可仅由DNA连接酶(ligase)参与而完全修复。
2.3 单链断裂的重接 DNA单链断裂,其中一部分可仅由DNA连接酶(ligase)参与而完全修复。 双链断裂几乎不能修复。 2.4 碱基的直接插入 DNA链上嘌呤的脱落造成无嘌呤位点,能被DNA嘌呤插入酶(insertase)识别结合,在K+存在的条件下,催化游离嘌呤或脱氧嘌呤核苷插入,生成糖苷键; 所催化插入的碱基有高度专一性、与另一条链上的碱基严格配对,使DNA完全恢复。
参与而完全修复。 双链断裂几乎不能修复。 2.4 碱基的直接插入 DNA链上嘌呤的脱落造成无嘌呤位点,能被DNA嘌呤插入酶(insertase)识别结合,在K+存在的条件下,催化游离嘌呤或脱氧嘌呤核苷插入,生成糖苷键; 所催化插入的碱基有高度专一性、与另一条链上的碱基严格配对,使DNA完全恢复。 ")
34
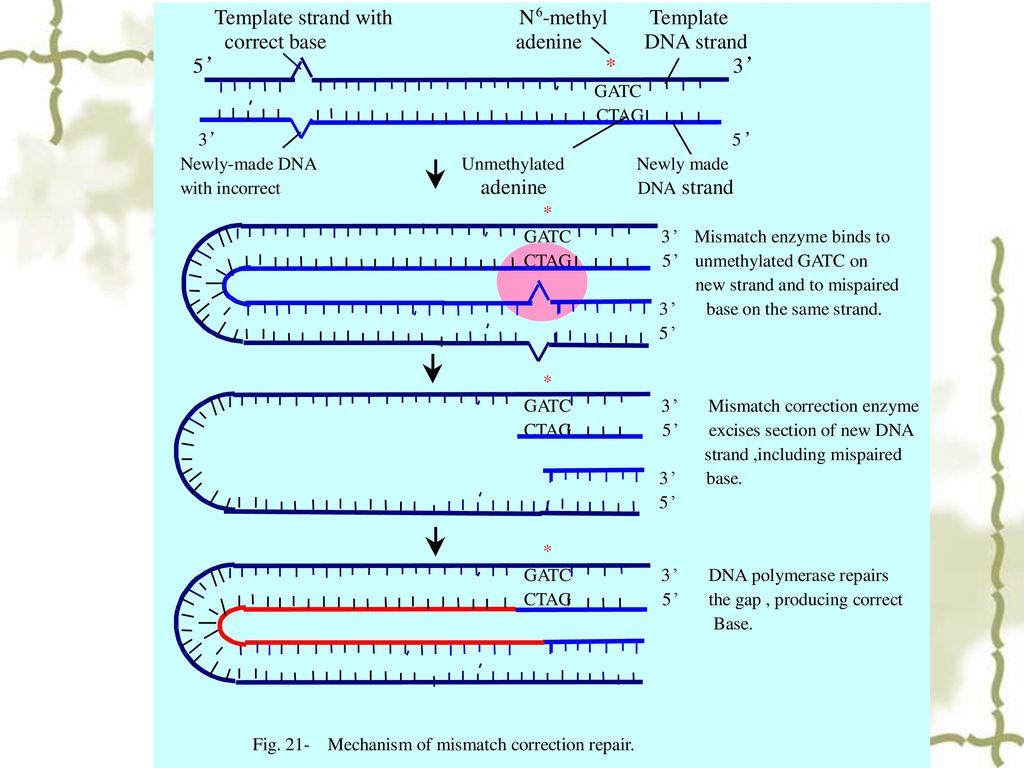
3 错配修复(mismatch repair)
一种纠正复制后子链中错配碱基的修复方式。 3.1 错配修复有关的蛋白和酶 MutS蛋白:识别错配的碱基。 MutH蛋白:内切酶,在子链未甲基化的5’-GATC-3’序列靠近鸟嘌呤的5’-端造成一个切口,使包括错配碱基在内的数百个核苷酸得以切除。 MutL蛋白:将MutH和MutS蛋白连接成复合体。 解链酶:解开DNA双链。 单链结合蛋白:稳定单链模板。 DNA聚合酶:从3’-OH填补空缺。 DNA 连接酶:连接切口。

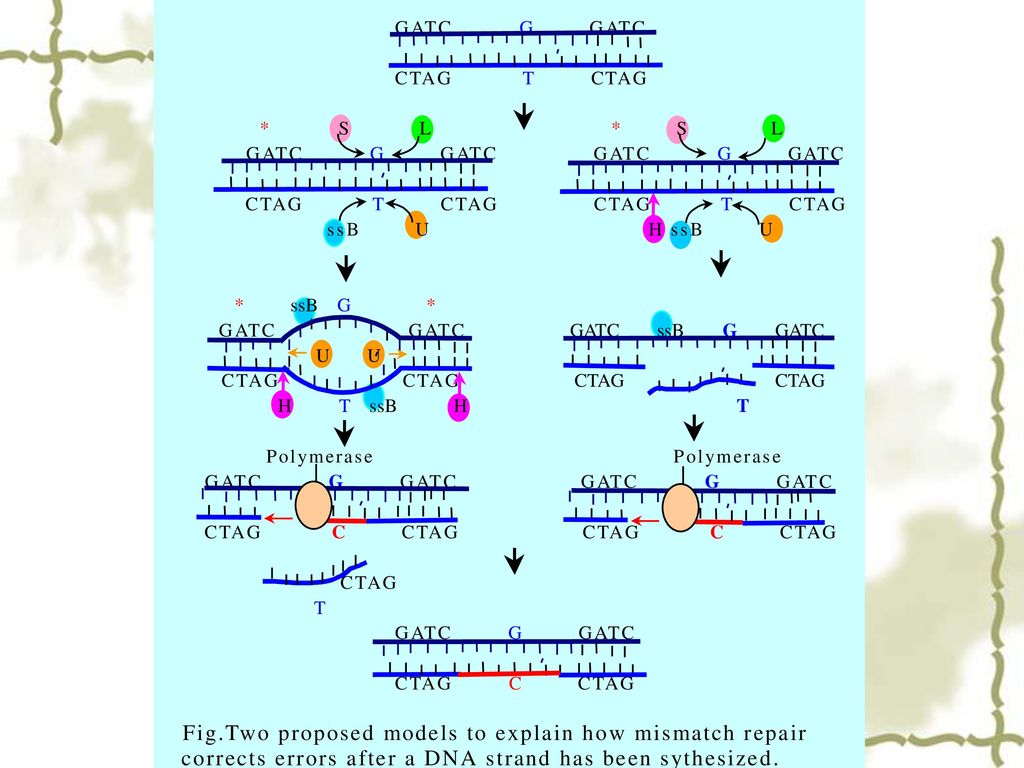
35
3.2 DNA错配修复的模型 DNA解链酶 核酸外切酶 SSB T G 切口 DNA聚合酶 DNA连接酶 dNMPs CH3 5´ 3´
G A T C 5´ 3´ C T A G T G CH3 G A T C C T AG G T 5´ 3´ CH3 G A T C C T AG G T 切口 G A T C 5´ 3´ C T A G G T CH3 DNA聚合酶 DNA连接酶 dNTPs CH3 G A T C C T AG MutH MutS MutL G T CH3 G A T C C T AG G C CH3 G A T C C T AG G T 5´ 3´

36
MutS识别错配位点,并易位到GATC位点。 MutH在GATC位点剪切非甲基化链,内切酶从GATC到错配位点降解DNA。
MutL MutS识别错配位点,并易位到GATC位点。 MutH在GATC位点剪切非甲基化链,内切酶从GATC到错配位点降解DNA。 MutS MutH

37
人类遗传性非多发性结直肠癌的病因是错配修复的基因缺陷。
3.3 错配修复的意义 修复DNA复制过程中逃避了DNA聚合酶校正作用的子链上的错配碱基。在子链合成后几分钟之内,GATC尚未甲基化,故能被MutH蛋白识别,结合并制造切口。 人类遗传性非多发性结直肠癌的病因是错配修复的基因缺陷。 mMLH1 突变谱 外显子 基因突变 蛋白产物(预测) bp缺失 外显子缺失16阅读框内 bp缺失 移码,氨基酸替换 bp插入 羧基末端延长 bp缺失 移码,不完整蛋白质
 bp缺失 外显子缺失16阅读框内 bp缺失 移码,氨基酸替换 bp插入 羧基末端延长 bp缺失 移码,不完整蛋白质.")
38
4 重组修复(recombinational repair)-复制后修复
指由基因的同源重组介导的修复过程。 在某些情况下没有互补链可以直接利用,例如在DNA复制时两条链已经分开后发生DNA损伤,采用重组修复。 作用: (1)修复DNA复制时子链的缺口。 (2)修复双链断裂,单链缺口。 (3)修复双链交联。
修复DNA复制时子链的缺口。 (2)修复双链断裂,单链缺口。 (3)修复双链交联。")
39
修复步聚: ①受损伤的DNA链复制时,产生的子代DNA在损伤的对应部位出现缺口。 ② 另一条母链DNA与有缺口的子链DNA进行重组交换,将母链DNA上相应的片段填补子链缺口处,而母链DNA出现缺口。 ③以另一条子链DNA为模板,经DNA聚合酶催化合成一新DNA片段填补母链DNA的缺口,最后由DNA连接酶连接,完成修补。 DNA重组修复方式

40
重组修复不能完全去除损伤,损伤的DNA段落仍然保留在亲代DNA链上;

41
5 SOS修复 SOS修复是指DNA受到严重损伤、细胞处于危急状态时所诱导的一种DNA修复方式, 修复结果只是能维持基因组的完整性,提高细胞的生成率,但留下的错误较多,故又称为错误倾向修复(error prone repair),细胞有较高的突变率。 (1)SOS反应能诱导多种蛋白质的合成 RecA蛋白:严重的DNA损伤产生大量单链缺口,激活RecA蛋白的水解酶活性,促进LexA阻遏蛋白的裂解和SOS反应的诱导。 LexA阻遏蛋白:调节所有SOS基因的转录。 DNA聚合酶Ⅱ:受SOS反应的诱导。能进行跨损伤复制。 (2)SOS修复是易错修复,导致突变增加。
SOS反应能诱导多种蛋白质的合成. RecA蛋白:严重的DNA损伤产生大量单链缺口,激活RecA蛋白的水解酶活性,促进LexA阻遏蛋白的裂解和SOS反应的诱导。 LexA阻遏蛋白:调节所有SOS基因的转录。 DNA聚合酶Ⅱ:受SOS反应的诱导。能进行跨损伤复制。 (2)SOS修复是易错修复,导致突变增加。")
42
SOS反应的起始: 是通过RecA的活化而引起。 诱导信号: 可能由DNA释放出的小分子组成的,或者是DNA本身某些结构变化。 RreA的激活导致LexA的基因产物的剪切。 LexA是一种小分子蛋白(22KDa),具有蛋白酶活性,是很多操纵子的阻遏物。 LexA被RecA激活后又可导致LexA自我催化它本身的裂解,这样LexA的阻遏功能失去了活性,并且同时诱导了它所结合的操纵子。

43
和SOS反应有关的基因,包括RecA,lexA,UvrA,UvrB,umuC和himA。

44
SOS修复中 lexA-recA调节子的作用机理
lexA基因 recA基因 mRNA 操纵子 可诱导基因 lexA阻遏物 作用于lexA阻遏物所控制的其他基因 非诱导状态 SOS修复中 lexA-recA调节子的作用机理 DNA损伤 诱导信号 recA 蛋白酶激活 lexA 阻遏物切断 阻遏物蓄积 蛋白酶 水平下降 信号水平 下降 DNA修复 嘧啶二聚体 活化的recA蛋白 切断的lexA阻遏物 recA蛋白 lexA阻遏物 诱导状态 lexA基因 recA基因 诱导基因

45
当DNA两条链的损伤邻近时,损伤不能被切除修复或重组修复,这时在核酸内切酶、外切酶的作用下造成损伤处的DNA链空缺,
修复过程: 当DNA两条链的损伤邻近时,损伤不能被切除修复或重组修复,这时在核酸内切酶、外切酶的作用下造成损伤处的DNA链空缺, 再由损伤诱导产生的一整套的特殊DNA聚合酶和SOS修复酶类,催化空缺部位DNA的合成, 补上去的核苷酸几乎是随机的,但仍然保持了DNA双链的完整性,使细胞得以生存。 图 SOS修复

46
6 限制-修饰系统(restriction and modificaion)
本世纪50年代初,以Arber等人对λ噬菌体在大肠杆菌不同菌株上的平板培养效应的研究为基础,发现了原核生物体内存在着寄生控制的限制(restriction)和修饰(modification)系统。
和修饰(modification)系统。")
47
结论: * E.coli K菌株和 B菌株各有自己的限制修饰系统, * E.coli C菌株没有;
♪ 菌体限制修饰系统中的限制性内切酶能将外来DNA切断, ♪ 菌体本身的DNA可受甲基化酶的保护。 两者称为限制-修饰作用 细菌借助于限制-修饰系统来区别自己DNA和外源DNA。

48
自己DNA: 限制模式相同的DNA 同株系的不同个体DNA、寄生于其中的质粒和噬菌体。 居民DNA(resident DNA):同一个细胞内的不同类型的DNA, 包括细胞DNA,质粒DNA,噬菌体DNA等。
:同一个细胞内的不同类型的DNA, 包括细胞DNA,质粒DNA,噬菌体DNA等。")
49
核酸内切限制酶的类型及其主要特性 特性 I型 II型 III型 限制和修饰活性 单一多功能的酶 分开的核酸内切酶和甲基化酶
分开的核酸内切酶和甲基化酶 具有一种共同亚基的双功能的酶 核酸内切限制酶的蛋白质结构 3种不同的亚基 单一的成份 2种不同的亚基 切割位点 在距寄主特异性位点至少700bp的地方可能随机地切割 位于寄主特异性位点或其附近 距寄主特异性位点3,端24~26bp处 甲基化作用的位点 寄主特异性的位点 识别未甲基化的序列进行核酸内切酶切割 能 能 序列特异的切割 不是 是 DNA克隆中的用处 无用 十分有用 用处不大

50
I类限制性内切酶的特点 1) I类限制性内切酶是异源多聚体,包含三个亚基R、M和S,其功能分别为限制性内切酶、甲基化酶和DNA特异位点识别。
2) I类限制性内切酶与DNA结合依赖M亚基与辅助因子S-腺苷甲硫氨酸(SAM)的相互作用。 3) I类限制性内切酶与DNA分子结合后,根据该位点的甲基化状态发挥相应的酶活性,或修饰或切割。切割位点与识别位点相隔1-5kb。 4) I类限制性内切酶的种类较少,研究的较多的有大肠杆菌的EcoK和EcoB。
 I类限制性内切酶与DNA结合依赖M亚基与辅助因子S-腺苷甲硫氨酸(SAM)的相互作用。 3) I类限制性内切酶与DNA分子结合后,根据该位点的甲基化状态发挥相应的酶活性,或修饰或切割。切割位点与识别位点相隔1-5kb。 4) I类限制性内切酶的种类较少,研究的较多的有大肠杆菌的EcoK和EcoB。")
51
I类限制性内切酶作用机制

52
II类限制性内切酶特点 II类限制性内切酶由H.O.Smith等1970年首先在流感嗜血杆菌Rd型菌株中发现。目前已有400多种II类限制性内切酶被分离,它是分子量较小的单体蛋白,识别和切割双链DNA分子时仅需要Mg2+, 识别和切割位点的序列有严格的特异性。

53
II类限制性内切酶作用机制

54
III类限制性内切酶的特点 III类限制性内切酶由R亚基和MS亚基组成,其中MS亚基具有位点识别和甲基化修饰双重活性。
该类酶与DNA识别位点的结合严格依赖ATP。修饰作用和切割作用取决于两个亚基之间的竞争。修饰作用在识别位点内进行。切割序列位于识别位点一侧若干碱基对处,无序列特异性。

55
目前对真核细胞的DNA修复的反应类型、参与修复的酶类 和修复机制了解还不多;
例如着色性干皮病(xeroderma pigmentosum)是第一个发 现的DNA修复缺陷性遗传病。
是第一个发 现的DNA修复缺陷性遗传病。")
56
第三节 基因突变

57
上述损伤会最终导致DNA分子结构的变化,这种DNA分子水平上的突变(mutation) ,是整体遗传突变的基础。
 ,是整体遗传突变的基础。")
58
1 突变类型 体细胞突变(somatic mutation):对于一个多细胞生物来说,如果突变仅发生在体细胞中,那么这种突变是不会传递给后代的。 但若突变发生在的生殖细胞中,那么这种突变就能通过配子传递给下一代,在后代个体的体细胞和生殖细胞中产生同样的突变,这种突变就叫做种系突变(germ-lime mutations)。 突变可以发生在染色水平或者基因水平。染色体结构和数目的改变叫做染色体畸变(chromosomal aberration),也称为染色体突变(chromosome mutation)。 当染色体畸变涉及到基因组中染色体套数的改变称为基因组突变(genome mutation),即整倍数改变。 发生在基因水平的突变称为基因突变(gene mutation)。 诱变剂处理所诱发的突变称为诱发突变(induced mutation),自然发生的突变是自发突变(spontaneous mutatiow)。
。 突变可以发生在染色水平或者基因水平。染色体结构和数目的改变叫做染色体畸变(chromosomal aberration),也称为染色体突变(chromosome mutation)。 当染色体畸变涉及到基因组中染色体套数的改变称为基因组突变(genome mutation),即整倍数改变。 发生在基因水平的突变称为基因突变(gene mutation)。 诱变剂处理所诱发的突变称为诱发突变(induced mutation),自然发生的突变是自发突变(spontaneous mutatiow)。")
59
2 基因突变的类型 2.1 从碱基变化分: 1)点突变(point mutation) 指DNA上单一碱基的变异。 转换(transition):嘌呤替代嘌呤(A与G之间的替代)、嘧啶替代嘧啶(C与T之间的替代); 颠换(transvertion):嘌呤变嘧啶或嘧啶变嘌呤。
:嘌呤变嘧啶或嘧啶变嘌呤。")
60
2) 缺失(deletion) 指DNA链上一个或一段核苷酸的消失。 3) 插入(insertion) 指一个或一段核苷酸插入到DNA链中。 4) 倒位或转位(transposition) 指DNA链重组使其中一段核苷酸链方向倒置、或从一处迁移到另一处。 5) 双链断裂 已如前述,对单倍体细胞一个双链断裂就是致死性事件。
 双链断裂 已如前述,对单倍体细胞一个双链断裂就是致死性事件。")
61
2.2 从突变效应分: 1)移码突变(frame shift mutaion):在为蛋白质编码的序列中如缺失及插入的核苷酸数不是3的整倍数,则发生读框移动(reading frame shift),使其后所译读的氨基酸序列全部混乱。 移码突变大多数常产生无活性产物, 有些移码突变会在基因内部形成终止密码子,从而使所生成的多肽链长度变短 。

62
移码突变

63
2)错义突变(missense mutation)是指DNA改变后mRNA中相应密码子发生改变,编码另一种氨基酸,使蛋白质中的氨基酸发生取代,可能削弱此蛋白的功能,以至影响到突变体的表型。
3)无义突变(nonsense mutation)是由DNA的碱基突变使mRNA中产生了无义密码子,翻译时在相应的位点会导致提前终止-平截或截短,产生一条不完整的多肽链,通常是没有功能的。
无义突变(nonsense mutation)是由DNA的碱基突变使mRNA中产生了无义密码子,翻译时在相应的位点会导致提前终止-平截或截短,产生一条不完整的多肽链,通常是没有功能的。")
64
校正突变:移码突变有时可由同一基因内的另一碱基的缺失或插入所抵消,但要求第二个移码突变发生在前一突变位点的下游。这样的第二个移码突变称为校正突变(suppressor mutation)。
。")
65
4)中性突变(neutral mutation)是在基因中有一对碱基对发生替换,引起的mRNA中密码子的改变,但多肽链中相应位点发生的氨基酸的取代并不影响蛋白质的功能。
5)沉默突变(silent mutation)或者同义突变:是中性突变中的一种特殊情况。即在基因中碱基对发生替换,改变了mRNA的密码子,结果蛋白质中相应位点是发生了相同氨基酸的取代,由于氨基酸的序列末发生变化,所以蛋白质仍具有野生型的功能。 6)回复突变(reverse mutation)分成两类: 正向突变(forward mutation):其突变方向是从野生型向突变型; 回复突变: 其突变方向是从突变型向野生型;回复突变可使突基因产生的无功能或有部分功能的多肽恢复部分功能或完全功能。 突变的作用还可以通过其它位点的突变而得到减少或校正,这便是前面已讨论过的抑制突变(suppressor mutation)或者校正突变。
沉默突变(silent mutation)或者同义突变:是中性突变中的一种特殊情况。即在基因中碱基对发生替换,改变了mRNA的密码子,结果蛋白质中相应位点是发生了相同氨基酸的取代,由于氨基酸的序列末发生变化,所以蛋白质仍具有野生型的功能。 6)回复突变(reverse mutation)分成两类: 正向突变(forward mutation):其突变方向是从野生型向突变型; 回复突变: 其突变方向是从突变型向野生型;回复突变可使突基因产生的无功能或有部分功能的多肽恢复部分功能或完全功能。 突变的作用还可以通过其它位点的突变而得到减少或校正,这便是前面已讨论过的抑制突变(suppressor mutation)或者校正突变。")
66
3 基因突变的后果 无影响 改变基因型(genotype)而不改变表现型(phenotype);产生遗传多态性(DNA多态性,蛋白质多态性)。 丧失某些功能;生物体结构和功能的异常引起疾病。 肿瘤 癌症发生 生殖细胞基因突变传递至后代,导致遗传性疾病的发生。 致死性; 进化 生物体获得新的性状,适应环境的能力增强。发生了有利于物种生存的结果,使生物进化。

67
4 突变和人类疾病 人类遗传性疾病已发现4000多种,其中不少与DNA修复缺陷有关,这些DNA修复缺陷的细胞表现出对辐射和致癌剂的敏感性增加。 在现代人类生活的环境中要接触很多的化学物质,如药物、食品防腐剂、化妆品、农药,用于工业的一些原料污染物等等。其中很多成份具有致癌和致突变作用。

68
名词解释 移框突变 无义突变 错义突变 增变基因 组成型表达 自己DNA 问答题: 错配修复酶的主要功能及作用机制。 核苷酸切除修复和重组修复的修复时期及简单过程。 何为RecA蛋白,其在SOS系统中有何用途? DNA损伤修复的主要类型。

70

71
A B B C 切 A A B C 切 A C B 螺旋酶 补 A B PolІ 螺旋酶 连接酶 封

74
切除修复 Exision—Repair 碱基切除修复 复制前进行 核苷酸切除修复 不易出错 UvrA, B, C gene 内切核酸酶
(Endonucleases) 外切核酸酶 (Exonuclease) DNA pol Ligase
 外切核酸酶. (Exonuclease) DNA pol. Ligase.")
Similar presentations


:核蛋白体组成成分 转移 RNA ( tRNA ):转运氨基酸 信使 RNA ( mRNA ):蛋白质合成模板 不均一核 RNA ( hnRNA ):成熟 mRNA 的前体 小核 RNA ( snRNA ):>")

















 A.细菌的遗传物质主要是DNA B.病毒的遗传物质主要是RNA>")