Download presentation
Presentation is loading. Please wait.

1
第五章 发酵机制与代谢调控 微生物发酵机制是指微生物通过其代谢活动,利用基质合成人们所需要的产物的内在规律。

2
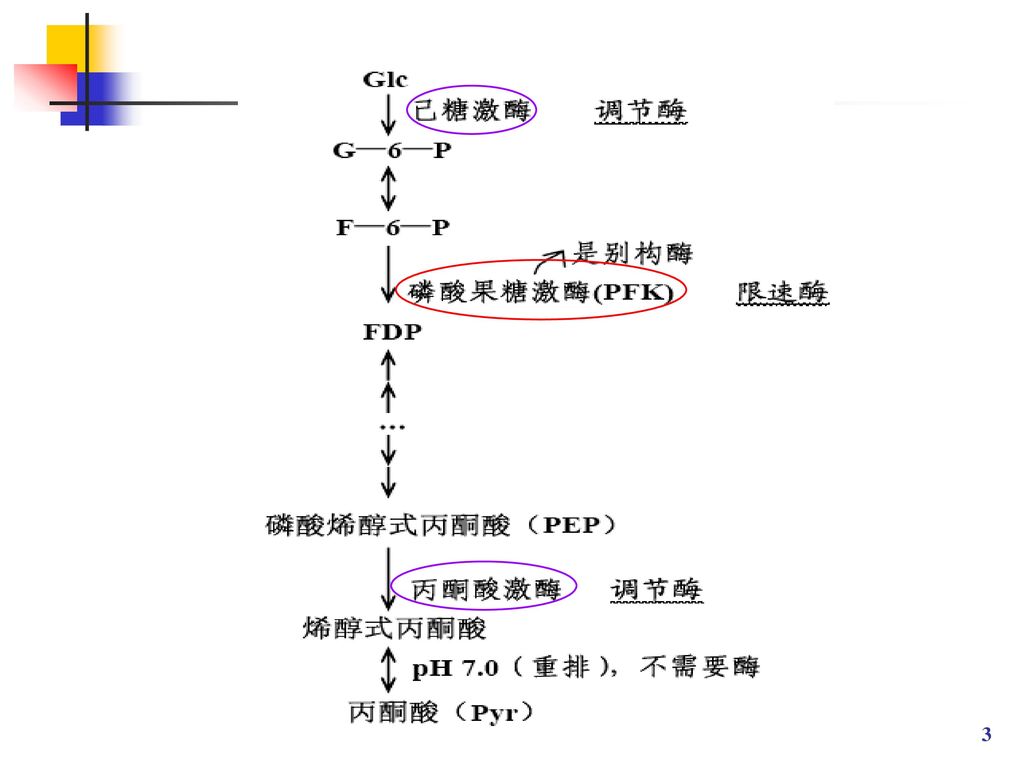
EMP途径 糖酵解(EMP)途径是葡萄糖有氧、无氧分解的共同途径。
在缺氧条件下,细胞进行无氧酵解,仅获得有限的能量以维持生命活动,丙酮酸继续进行代谢可产生酒精、乳酸、甘油及其它厌氧代谢产品。 在有氧条件下,细胞进行有氧代谢生成丙酮酸后,进入TCA循环。其发酵产品有柠檬酸、氨基酸及其它有机酸等。

4
第一节 厌氧发酵机制与代谢调控 一、酵母菌的酒精发酵(酵母的第Ⅰ型发酵) 1. 生物合成途径
 1. 生物合成途径")
5
总反应式为: 理论转化率为: 实际转化率为理论值的 95%,约 48.5% CO2产量约为酒精质量的 95.5%

6
酒精是胞内产物 渗出体外 与H2O任比例混合 酒母 水溶液中 连续发酵 pH4.5 左右 30~33℃ 60~72 h
酒母不含淀粉酶,主要含水解酶(如蔗糖酶、麦芽糖酶)和 酒化酶(指参与酒精发酵的各种酶和辅酶的总称,胞内酶)。 酒糟的利用 酒精是胞内产物 渗出体外 与H2O任比例混合
和. 酒化酶(指参与酒精发酵的各种酶和辅酶的总称,胞内酶)。 酒糟的利用. 酒精是胞内产物 渗出体外 与H2O任比例混合.")
7
CO2 溶解度较小 被动式发酵 泡沫式发酵 不利 产生的CO2应设法排除,并注意加强对随CO2逸出时被 带走酒精的捕集回收。 CO2产量约为酒精质量的 95.5%,而且纯度相当高,只需经过简单的提纯处理,便可得到几乎纯粹的CO2 。可以用来生产液体CO2 、干冰、纯碱和轻质碳酸钙等。

8
2. ▲巴斯德效应(Pasteur effect)
巴斯德在研究酵母的酒精发酵时,发现在有氧的情况下,由于进行呼吸作用,酒精产量大大下降,糖的消耗速度减慢,这种呼吸抑制发酵的作用被后人称为巴斯德效应。

9
巴斯德效应 己糖激酶被G-6-P、PEP抑制 PFK受高水平的ATP、柠檬酸、 脂肪酸所抑制, 能被AMP、ADP活化 丙酮酸激酶受高水平
的ATP抑制,被FDP激活

10
习题:酵母可以依赖Glc厌氧或有氧生长,试解释当一直处于厌氧环境中的酵母细胞暴露于空气中时,Glc的消耗速率为什么会下降?
3. 酵母菌酒精发酵中副产物的生成 主要产物是乙醇和CO2,但也伴随有40多种副产物(包括甘油和杂醇油)。副产物生成一方面耗用了糖分,同时影响了产品的质量。 酵母的第Ⅱ型发酵 酵母菌的酒精发酵又称酵母的第Ⅰ型发酵
。副产物生成一方面耗用了糖分,同时影响了产品的质量。 酵母的第Ⅱ型发酵. 酵母菌的酒精发酵又称酵母的第Ⅰ型发酵.")
11
二、细菌的酒精发酵 少数假单胞杆菌 Glc只经过4步反 应就可形成Pyr 在ED途径中生成 的2分子的丙酮酸 脱羧生成乙醛,
乙醛还原生成乙醇

12
三、乳酸发酵机制 有同型乳酸发酵和异型乳酸发酵两种类型。前者在发酵产物中只有乳酸,后者的产物中除乳酸外,还有乙醇和CO2。两者的发酵菌种不同,发酵机制也不同。 1、同型乳酸发酵 乳酸菌 德氏乳杆菌 大多数乳酸菌不具有脱羧酶 乳酸脱氢酶

13
2、异型乳酸发酵 (1) 6-磷酸葡萄糖酸途径 总反应式为: 理论转化率为: 1 mol 葡萄糖生成1 mol 乳酸和l mol 乙醇。
乳酸对糖的理论转化率是50%。 肠膜明串珠菌和葡聚糖明串珠菌等 。

14
(2)双歧途径 双歧杆菌 在这个途径中,2molGlc可以生成2mol乳酸和3mol 乙酸,乳酸对糖的转化率理论上只有50%。
双歧途径 双歧杆菌 在这个途径中,2molGlc可以生成2mol乳酸和3mol 乙酸,乳酸对糖的转化率理论上只有50%。")
15
四、甘油发酵机制 甘油 (丙三醇) 良好溶剂,广泛用于化妆品和医药行业;炸药。 1、亚硫酸盐法甘油发酵 酵母菌 酵母的第Ⅱ型发酵
 良好溶剂,广泛用于化妆品和医药行业;炸药。 1、亚硫酸盐法甘油发酵 酵母菌 酵母的第Ⅱ型发酵")
16
乙醇脱氢酶 发酵液中加入亚硫酸氢钠(NaHSO3) 亚硫酸钠加成物 ( ) ▲原理:阻遏乙醇的生物合成
 亚硫酸钠加成物 ( ) ▲原理:阻遏乙醇的生物合成")
17
实际上甘油发酵过程中总会有一些乙醇生成,因为:
1. 从能量守恒可知,要生成甘油则必须有等摩尔量的乙醛生成。即 酵母进行甘油发酵时必须依靠部分酒精发酵以获得能量(ATP和NADH + H+)。 2. 生成部分乙醇不可避免。 因为乙醛与NaHSO3复合反应效率低于89%;且酵母体内有乙醇脱氢酶存在。 总反应式为: C6H12O C3H5(OH)3 + CH3CHO + CO2 酵母的第Ⅱ型发酵
。 2. 生成部分乙醇不可避免。 因为乙醛与NaHSO3复合反应效率低于89%;且酵母体内有乙醇脱氢酶存在。 总反应式为: C6H12O6 C3H5(OH)3 + CH3CHO + CO2. 酵母的第Ⅱ型发酵.")
18
2、碱法甘油发酵 酒精酵母 酵母的第Ⅲ型发酵 如果碱性(pH值7.6以上) 两分子乙醛发生歧化反应形成各一分子的乙酸和乙醇。
酒精酵母 酵母的第Ⅲ型发酵 如果碱性(pH值7.6以上) 两分子乙醛发生歧化反应形成各一分子的乙酸和乙醇。 2C6H12O6+H2O C3H5(OH)3+CH3COOH+ C2H5OH + 2CO2 产物复杂
 两分子乙醛发生歧化反应形成各一分子的乙酸和乙醇。 2C6H12O6+H2O 2C3H5(OH)3+CH3COOH+ C2H5OH + 2CO2. 产物复杂.")
19
五、甲烷(沼气)发酵 甲烷发酵的机理是厌氧菌将碳水化合物、脂肪、蛋白质等复杂的有机物最终分解成甲烷和CO2。 三阶段 废物利用
发酵 甲烷发酵的机理是厌氧菌将碳水化合物、脂肪、蛋白质等复杂的有机物最终分解成甲烷和CO2。 三阶段 废物利用")
20
第二节 好氧发酵机制与代谢调控 一、柠檬酸发酵机制 1. 柠檬酸及其衍生物的用途 (1)食品工业:酸味剂、增溶剂、缓冲剂、抗氧化剂、
第二节 好氧发酵机制与代谢调控 一、柠檬酸发酵机制 1. 柠檬酸及其衍生物的用途 (1)食品工业:酸味剂、增溶剂、缓冲剂、抗氧化剂、 除腥脱臭剂、螯合剂等; (2)药物、化妆品; (3)工业上:去垢、无土栽培、胶粘剂等。 (4)柠檬酸盐类具有溶解度高、生理宽容性大、酸根可直接吸收和代谢。补钾、抗凝血、柠檬酸钠。 (5)柠檬酸酯(三酯等):树脂增塑剂(抗霉),化工原 料,药物等。
食品工业:酸味剂、增溶剂、缓冲剂、抗氧化剂、 除腥脱臭剂、螯合剂等; (2)药物、化妆品; (3)工业上:去垢、无土栽培、胶粘剂等。 (4)柠檬酸盐类具有溶解度高、生理宽容性大、酸根可直接吸收和代谢。补钾、抗凝血、柠檬酸钠。 (5)柠檬酸酯(三酯等):树脂增塑剂(抗霉),化工原. 料,药物等。")
21
2. 柠檬酸的生物合成途径 菌种:黑曲霉和假丝酵母 好氧发酵 发酵原料:玉米、干薯、木薯、小麦、糖蜜等。 黑曲霉偏好于无机氮源。 无机氮源被利用后,对M的pH值有影响。 T:黑曲霉生长最适33℃~37℃,柠檬酸积累32℃。 pH值:黑曲霉生长最适pH3~7,柠檬酸积累pH< 2。

22
柠檬酸的生物合成途径 EMP TCA 5 6 7

23
可见,由Glc生成柠檬酸的生物合成途径包括:EMP、TCA 循环和 CO2固定作用。
要使柠檬酸积累,必须解决两个问题: (1)设法阻断柠檬酸的进一步代谢; (2)途径被阻断后,要采取合适的策略来补充阻断点之后的物质(主要是草酰乙酸),以维持发酵的继续进行。
设法阻断柠檬酸的进一步代谢; (2)途径被阻断后,要采取合适的策略来补充阻断点之后的物质(主要是草酰乙酸),以维持发酵的继续进行。")
24
3. 柠檬酸积累的代谢调节 (1)阻断柠檬酸的进一步代谢:使顺乌头酸酶失活 Fe2+是顺乌头酸酶的激活剂 加络合剂 (2)加强 EMP途径 增强3 个关键酶尤其是PFK的活力 PFK 受高水平的柠檬酸和ATP抑制; NH4+能解除此抑制 ∴氮源用NH4+盐(铵盐) 柠檬酸发酵时要求Mn2+、Fe2+、Zn2+的含量极低
 柠檬酸发酵时要求Mn2+、Fe2+、Zn2+的含量极低.")
25
Mn2+是许多酶的组成部分。柠檬酸发酵时对Mn2+极端敏感。
Mn2+缺乏导致细胞内NH4+水平升高。 NH4+水平升高减弱了柠檬酸对EMP途径关键酶(PFK)的抑制。 (3)TCA循环的调节 增强柠檬酸合成酶的活性 (4)及时补加草酰乙酸 TCA循环被切断, 必须由另外的途径来补充草酰乙酸。
的抑制。 (3)TCA循环的调节. 增强柠檬酸合成酶的活性. (4)及时补加草酰乙酸. TCA循环被切断, 必须由另外的途径来补充草酰乙酸。")
26
草酰乙酸的来源 √ √ 主要 加强CO2固定反应 √ × × 黑曲霉中没有苹果酸酶

27
二、醋酸发酵机制 纯醋酸:化学合成法; 食醋:发酵法 原料:大米
纯醋酸:化学合成法; 食醋:发酵法 原料:大米 传统食醋多为固态发酵(需40~60 d),淋醋工艺法(又称速酿法)只需 5天。 发酵三阶段:① 原料的液化与糖化;② 酒精发酵;③ 醋酸发酵。 例如,醋杆菌发酵酒精成醋酸
,淋醋工艺法(又称速酿法)只需 5天。 发酵三阶段:① 原料的液化与糖化;② 酒精发酵;③ 醋酸发酵。 例如,醋杆菌发酵酒精成醋酸.")
28
三、谷氨酸发酵机制 细胞经济与生产经济矛盾
氨基酸发酵是典型的代谢控制发酵。由于发酵产物氨基酸都是M.B的中间代谢产物,它的积累是建立在对M.B正常代谢的抑制。也就是说,氨基酸发酵的关键是取决于能否打破M.B的正常代谢调节,人为地控制M.B的代谢。 细胞经济与生产经济矛盾

30
(一)Glu的用途 1. 味精合成前体 2. Glu可以通过转氨反应把氨基转移到酮酸上,合成人 体代谢所需的其它氨基酸。 3. 解“氨毒”
HOOC—CH2—CH2—CH—COONa·H2O NH2 2. Glu可以通过转氨反应把氨基转移到酮酸上,合成人 体代谢所需的其它氨基酸。 3. 解“氨毒” 谷氨酰胺(Gln)
")
32
(二)Glu 生物合成途径及调节机制 1. 合成途径 (1)以糖质原料(最后水解为Glc)作为碳源 (2)以乙酸作为碳源
谷氨酸的大量积累不是由于生物合成途径的特异,而是菌体代谢调节控制和细胞膜通透性的特异调节以及发酵条件的适合。

33
DCA循环 TCA循环 乳酸 由 生物合成 的代谢途径 Glc Glu

34
以乙酸为唯一碳源生物合成 的代谢途径 Glu DCA循环 TCA循环

35
注意: (1)产物为L-型 (2)需要引入NH4+(但要适量),产物一般以谷氨酸铵盐的形式存在:C5H8O4N•NH4
产物为L-型 (2)需要引入NH4+(但要适量),产物一般以谷氨酸铵盐的形式存在:C5H8O4N•NH4")
36
2. 谷氨酸生产菌 谷氨酸生产菌:棒状杆菌属、短杆菌属、小杆菌属及节 杆菌属中的细菌。 ▲谷氨酸生产菌有以下共同特征:
(1)都是需氧型微生物; (2)都是生物素缺陷型; (3)发酵中菌体发生明显的变化,同时发生细胞膜 渗透性的变化; (4)CO2固定反应酶系活力强; (5)α-酮戊二酸脱氢酶活力缺失或微弱;
都是需氧型微生物; (2)都是生物素缺陷型; (3)发酵中菌体发生明显的变化,同时发生细胞膜. 渗透性的变化; (4)CO2固定反应酶系活力强; (5)α-酮戊二酸脱氢酶活力缺失或微弱;")
37
(6)柠檬酸合成酶、顺乌头酸酶、异柠檬酸脱氢酶
以及Glu脱氢酶活力强; (7)能利用醋酸; (8)具有向环境中泄漏Glu的能力; (9)不分解利用Glu,并能耐高浓度的Glu; (10)Glu对Glu脱氢酶存在着反馈抑制和反馈阻遏。 因此,前述两种生物合成Glu的途径应为:
能利用醋酸; (8)具有向环境中泄漏Glu的能力; (9)不分解利用Glu,并能耐高浓度的Glu; (10)Glu对Glu脱氢酶存在着反馈抑制和反馈阻遏。 因此,前述两种生物合成Glu的途径应为:")
38
因为Glc抑制异柠檬酸裂解酶的活性,故异柠檬酸的分解反应微弱。
乳酸

39
DCA循环

40
3. 有关Glu代谢途径的解释 (1) 以Glc为碳源的代谢途径 ① 许多研究证明:以Glc为C源时,糖酵解经EMP和
HMP两个途径进行,但是以EMP为主(占74%)。 ② 当TCA循环有缺陷时,DCA循环起补充四碳二羧酸 的作用。 * C4二羧酸:草酰乙酸、苹果酸、延胡索酸、琥珀酸。 ③ 如果C4二羧酸完全由CO2固定反应提供:
。 ② 当TCA循环有缺陷时,DCA循环起补充四碳二羧酸. 的作用。 * C4二羧酸:草酰乙酸、苹果酸、延胡索酸、琥珀酸。 ③ 如果C4二羧酸完全由CO2固定反应提供:")
41
乳酸 DCA循环

42
总反应式为: C6H12O6 + NH3 + 3/2 O C5H9O4N + CO2 + 3 H2O 此时,Glu对Glc的理论收率为81.7% ④ 在Glu生成期,若CO2固定反应完全不起作用,C4二 羧酸完全由DCA循环提供:

43
乳酸 延胡索酸 琥珀酸

44
反应如下: 此时,Glu对Glc的理论收率仅为54.4% ∴在Glu生成期,最好没有异柠檬酸裂解反应,应关闭 DCA循环。
实际收率处于中间值(54.4%~81.7%)。CO2固定反应与DCA循环的比率对Glu的产率有影响,后者的活性越高,Glu的生成收率越低。 * 在菌体生成期,可走DCA循环。
。CO2固定反应与DCA循环的比率对Glu的产率有影响,后者的活性越高,Glu的生成收率越低。 * 在菌体生成期,可走DCA循环。")
45
所以, 由Glc生成Glu的理想途径为: 四碳二羧酸完全由CO2固定化反应提供。

46
(3) 两种途径中,细胞内α-酮戊二酸与异柠檬酸的量 需维持平衡。
(2) 以乙酸为碳源的代谢途径 DCA循环是四碳二羧酸的唯一来源。 (3) 两种途径中,细胞内α-酮戊二酸与异柠檬酸的量 需维持平衡。 反馈抑制
 以乙酸为碳源的代谢途径. DCA循环是四碳二羧酸的唯一来源。 (3) 两种途径中,细胞内α-酮戊二酸与异柠檬酸的量. 需维持平衡。 反馈抑制.")
47
4. 由Glc生物合成Glu的调节机制 生物素对糖代谢的调节
(1)α-酮戊二酸脱氢酶在Glu产生菌中先天性的缺失或微弱(尤其是在生物素缺乏的条件下),TCA循环到达α-酮戊二酸即受阻。 (2)Glu产生菌中的L- Glu脱氢酶的活力很强,α-酮戊二酸在Glu脱氢酶催化下氨基化生成Glu(需引入NH4+ )。 (3)通过控制生物素亚适量,引起代谢失调使Glu得以积累。 生物素对糖代谢的调节 生物素还影响细胞膜的通透性,后述
α-酮戊二酸脱氢酶在Glu产生菌中先天性的缺失或微弱(尤其是在生物素缺乏的条件下),TCA循环到达α-酮戊二酸即受阻。 (2)Glu产生菌中的L- Glu脱氢酶的活力很强,α-酮戊二酸在Glu脱氢酶催化下氨基化生成Glu(需引入NH4+ )。 (3)通过控制生物素亚适量,引起代谢失调使Glu得以积累。 生物素对糖代谢的调节. 生物素还影响细胞膜的通透性,后述.")
48
① 生物素对糖代谢速度的影响 生物素影响糖代谢的速度而不是EMP与HMP途径的比率。 数据:生物素充足时HMP占38%,亚适量时26%。
没有显著影响。 大量合成Glu所需要的生物素浓度比菌体生长的需要量 低,即为菌体生长需要的“亚适量”。一般5~10μg/L * 在生物素充足的条件下,Pyr趋于生成乳酸的反应。 ② 生物素对CO2固定反应的影响 生物素是丙酮酸羧化酶的辅酶,参与CO2固定反应。

49
③ 生物素对DCA循环的影响 DCA循环的关键酶是异柠檬酸裂解酶和苹果酸合成酶。 异柠檬酸裂解酶受Glc、琥珀酸阻遏,为HAC所诱导。 在以Glc为原料时,通过控制生物素亚适量,几乎看不到异柠檬酸裂解酶的活性,DCA循环基本上是封闭的。代谢流向异柠檬酸 α-酮戊二酸 Glu的方向高效率地移动。 (4)∵Glu对Glu脱氢酶存在着反馈抑制和反馈阻遏, ∴胞内产物 L-Glu需分泌出来。 细胞膜的通透性控制
∵Glu对Glu脱氢酶存在着反馈抑制和反馈阻遏, ∴胞内产物 L-Glu需分泌出来。 细胞膜的通透性控制.")
50
5. 细胞膜的通透性控制

51
Glu的分泌是由细胞膜控制的。后来,更进一步研究得到主要是受细胞膜的磷脂含量控制。
棒状杆菌的细胞膜的磷脂是由不饱和脂肪酸、甘油、磷酸和侧链组成的。所以,控制磷脂含量可以通过控制油酸、甘油或磷脂的合成来实现。

52
(1) 控制磷脂的合成 ① 生物素缺陷型 生物素参与了不饱和脂肪酸的合成,进而影响磷脂的合成。
选育生物素缺陷型,阻断生物素合成,同时限制外源性生物素供应量。 ② 添加饱和脂肪酸、表面活性剂 高级饱和脂肪酸和表面活性剂(如Tween60)对生物素有拮抗作用,通过拮抗脂肪酸的合成,导致磷脂合成不足,细胞膜结构不完整,提高了细胞膜的通透性。
对生物素有拮抗作用,通过拮抗脂肪酸的合成,导致磷脂合成不足,细胞膜结构不完整,提高了细胞膜的通透性。")
53
甘油缺陷型突变株丧失了α-磷酸甘油脱氢酶,不能合成α-磷酸甘油和磷脂。
③ 油酸缺陷型 油酸含量的多少直接影响到磷脂合成量的多少和细胞膜的渗透性。油酸缺陷型突变株丧失了自身合成油酸的能力,即丧失脂肪酸生物合成能力,必须由外界供给。 ④ 甘油缺陷型 甘油缺陷型突变株丧失了α-磷酸甘油脱氢酶,不能合成α-磷酸甘油和磷脂。

54
(2)抑制Glu生产菌细胞壁的合成 比如:添加青霉素
青霉素能抑制Glu生产菌细胞壁的生物合成,使细胞膜处于无保护状态,又由于膜内外的渗透压差,进而导致细胞膜的物理损伤,增大了Glu向胞外漏出的渗透性。

Similar presentations



 氧化分解供应能量 转化为其他物质.>")


的過濾作用 腎小管的再吸收作用 腎小管的分泌作用 尿液的組成>")





蛋白质的降解: 外源蛋白的消化 内源性蛋白的选择性降解 (2)氨基酸的分解代谢:>")








