Download presentation
Presentation is loading. Please wait.

1
第三章 杀菌剂

2
Fungicides Bactericides Viricides Nematocides Protozoactdes Algicides
…… 防治高等寄生性种子植物的除草剂

3
Great Irish Famine , 100 万人死亡,人口减少1/4
Great Irish Famine , 100 万人死亡,人口减少1/4

4
第一节 概述 杀菌剂(Fungicide)是一类用来防治植物病害的药剂。 早期杀菌剂的含义:杀死真菌或抑制其生长的化学物质。 科学的定义:
第一节 概述 杀菌剂(Fungicide)是一类用来防治植物病害的药剂。 早期杀菌剂的含义:杀死真菌或抑制其生长的化学物质。 科学的定义: (1)直接杀死或抑制植物病原物生长发育 (2) 对病原菌无直接毒性 ①改变致病过程 ②诱导植物产生抗病性。
是一类用来防治植物病害的药剂。 早期杀菌剂的含义:杀死真菌或抑制其生长的化学物质。 科学的定义: (1)直接杀死或抑制植物病原物生长发育. (2) 对病原菌无直接毒性. ①改变致病过程. ②诱导植物产生抗病性。")
5
一、什么是杀菌剂 从字义上来看:“杀死”——“抑菌” 实际上: (1)没有直接将病原菌杀灭;
(2)抑制生长或发育,使菌丝不能伸长,停止生长。
抑制生长或发育,使菌丝不能伸长,停止生长。")
6
1. 杀菌剂作用病菌致使病菌中毒的症状 主要表现为: 菌丝生长受阻,畸形,扭曲等; 孢子不能萌发; 各种子实体、附着孢不能形成; 细胞膨胀,原生质瓦解,细胞壁破坏。

7
2. 杀菌和抑菌的区别 (1) 从中毒症状上看: 杀菌主要表现为孢子不能萌发,菌体干缩、死亡、瓦解。 抑菌则表现为菌丝生长受阻(不是死亡),药剂解除后可恢复生长。
 从中毒症状上看: 杀菌主要表现为孢子不能萌发,菌体干缩、死亡、瓦解。 抑菌则表现为菌丝生长受阻(不是死亡),药剂解除后可恢复生长。")
8
(2) 从作用机制看: 杀菌主要是影响了生物氧化——能的生成 (孢子萌发需要较多的能量) 抑菌主要是影响了生物合成 (菌丝生长耗能少)
 从作用机制看: 杀菌主要是影响了生物氧化——能的生成 (孢子萌发需要较多的能量) 抑菌主要是影响了生物合成 (菌丝生长耗能少)")
9
杀菌和抑菌二者不能截然分开,一个杀菌剂表现哪一种作用,也和下列因素有关。
(1) 药剂本身的性质 一般说来,重金属盐、有机硫类杀菌剂多表现为杀菌作用; 内吸性杀菌剂、农用抗菌素类则多表现为抑菌作用;
 药剂本身的性质. 一般说来,重金属盐、有机硫类杀菌剂多表现为杀菌作用; 内吸性杀菌剂、农用抗菌素类则多表现为抑菌作用;")
10
(2) 药剂浓度 低浓度——抑菌 5mg/L苯来特,抑制白粉菌丝生长; 高浓度——杀菌 500mg/L苯来特,影响孢子萌发。 (3) 药剂作用时间 时间短,抑菌作用 时间延长,杀菌作用
 药剂浓度 低浓度——抑菌 5mg/L苯来特,抑制白粉菌丝生长; 高浓度——杀菌 500mg/L苯来特,影响孢子萌发。 (3) 药剂作用时间 时间短,抑菌作用 时间延长,杀菌作用")
11
二、杀菌剂发展简史及开发状况 1. 简史 第一时期: 为上古时期至19世纪80年代以前(主要以元素硫为主的古代天然药物时代)
1. 简史 植物病害防治发展史就是杀菌剂的研发和应用史 第一时期: 为上古时期至19世纪80年代以前(主要以元素硫为主的古代天然药物时代) 公元前1000年,古希腊诗人荷马(Homer)在其著作中就描述了硫磺的防病作用; 公元前470年德莫克里图介绍用橄榄浸出液洒在植物上防治疫病; 约公元60年蒲林尼介绍用酒和捣碎的柏叶混合物浸种防治小麦霉病; 杀菌剂是人类历史上最古老的一类药剂,其发展史大致可分为四个重要时期。
 公元前1000年,古希腊诗人荷马(Homer)在其著作中就描述了硫磺的防病作用; 公元前470年德莫克里图介绍用橄榄浸出液洒在植物上防治疫病; 约公元60年蒲林尼介绍用酒和捣碎的柏叶混合物浸种防治小麦霉病; 杀菌剂是人类历史上最古老的一类药剂,其发展史大致可分为四个重要时期。")
12
公元900年左右,中国民间开始使用砷制剂防治蔬菜病虫害。
1705年,升汞(HgCl2)开始用于木材防腐和种子消毒。 随着植物病害病原学理论的形成,1761年Schulthess首次将硫酸铜用于防治小麦黑穗病; 1802年,William Forsyth首次研制出石灰-硫磺合剂,并应用于防治果树白粉病,此后各种元素硫和石硫合剂在欧洲和美国进一步得到应用。
开始用于木材防腐和种子消毒。 随着植物病害病原学理论的形成,1761年Schulthess首次将硫酸铜用于防治小麦黑穗病; 1802年,William Forsyth首次研制出石灰-硫磺合剂,并应用于防治果树白粉病,此后各种元素硫和石硫合剂在欧洲和美国进一步得到应用。")
13
第二时期: 19世纪80年代至20世纪30年代中期(主要以波尔多液为代表的无机合成杀菌剂时代)
1874年,法国爆发葡萄霜霉病。1882年,法国波尔多大学 Millardet发现波尔多液,开创了铜制剂时代。被称为“杀菌大王”,是一个里程碑。 1930年我国浙江省植物病虫防治所建立了中国最早的农药研究机构,研发出铜制剂、硫制剂、砷制剂和汞制剂用于植物病害防治。 早期品种:硫酸铜、王铜(碱式氯化铜)、波尔多液,存在安全性问题(药害); 当前:美国固信公司,可杀得(氢氧化铜)WP; 日本北兴化学株式会社:加瑞农(氢氧化铜、春雷霉素)WP;诺华:靠山(氧化亚铜)水分散剂。
、波尔多液,存在安全性问题(药害); 当前:美国固信公司,可杀得(氢氧化铜)WP; 日本北兴化学株式会社:加瑞农(氢氧化铜、春雷霉素)WP;诺华:靠山(氧化亚铜)水分散剂。")
14
第三时期: 20世纪30年代中期至60年代。主要是 以福美类和代森类为代表的有机合成保护性杀菌剂时代。
1934年Tisdale和Williams与Martin同时发现并各自报道了对植物相对比较安全、而对植物病害具有显著控制作用的一类有机硫保护性杀菌剂,即二硫代氨基甲酸衍生物(福美类),标志着人类进入人工合成有机杀菌剂防治植物病害的新纪元。 主要品种有:代森锰、代森锌、代森锰锌、代森铵; 该类药剂具广谱性,对大部分病害有较好的防治作用。
,标志着人类进入人工合成有机杀菌剂防治植物病害的新纪元。 主要品种有:代森锰、代森锌、代森锰锌、代森铵; 该类药剂具广谱性,对大部分病害有较好的防治作用。")
15
第四时期: 20世纪60年代后。主要以萎锈灵,苯菌灵、甲霜灵、三唑酮和嘧菌酯为代表的内吸性有机杀菌剂的涌现和广泛应用时代。
早在50年代初,就进行了以威菌磷为代表的有机内吸性杀菌剂研究 60年代中期,以萎锈灵为代表的丁烯酰胺类和以苯菌灵为代表的苯并咪唑类杀菌剂的研究成功,标志着控制植物病害的化学防治取得重大突破。随后,涌现出的200多种内吸性杀菌剂和10多种抗菌素。

16
甲菌啶、乙菌啶等氨基嘧啶类; 甲霜灵、恶霜灵等苯酰胺类; 异稻瘟净、乙磷铝、甲基立枯磷等有机磷类; 邻酰胺等苯胺类; 叶青双、拌种灵等噻唑类; 恶霉灵等恶唑类; 以三唑酮、氯苯嘧啶醇、十三吗啉等为代表的麦角甾醇生物合成抑制剂。以三唑类为代表的超高效生物活性杀菌剂的广泛应用,为实现扩大杀菌剂防治谱,降低使用剂量的目标奠定了基础。 90年代strobilurin(嗜球伞果素)类杀菌剂成为继三唑类之后杀菌剂发展史上的又一里程碑。 同时,新型的杀菌剂,如三环唑、噻瘟唑也被广泛用于稻瘟病防治。优秀的保护性杀菌剂如二甲酰亚胺类和吡咯类也研制成功。以氟吗啉为代表对卵菌病害高效的羧酸氨基化合物类杀菌剂走向市场。
类杀菌剂成为继三唑类之后杀菌剂发展史上的又一里程碑。 同时,新型的杀菌剂,如三环唑、噻瘟唑也被广泛用于稻瘟病防治。优秀的保护性杀菌剂如二甲酰亚胺类和吡咯类也研制成功。以氟吗啉为代表对卵菌病害高效的羧酸氨基化合物类杀菌剂走向市场。")
17
——杀菌剂市场规模已与杀虫剂市场基本持平(亿美元)
——果蔬类作物占据了全球杀菌剂施用的半壁江山 2011年

18
2. 研究发展状况 (1) 内吸性杀菌剂迅猛发展。1966年,萎锈灵;1969年,苯来特。…… (2) 杀菌剂作用机理研究取得飞跃发展。 (3) 新品种开发的手段发生了新变化。经验筛选——生物合理设计 (4) 病原菌抗药性问题日益严重。
 病原菌抗药性问题日益严重。")
19
三、我国病害防治中存在的主要问题 1. 药剂品种老化单一,尤其表现在卵菌病害的防治上。乙磷铝——甲霜灵 2. 病害抗药性日趋严重。
3. 制剂加工技术滞后影响药效发挥,滥用现象普遍存在。我国主要是WP悬浮率的行业标准较低。 4. 缺乏植保技术人员和推广人员,没有健全的服务体系。

20
因此不能: (1)及时准确地进行预测; (2)制定合理的防治方案。
及时准确地进行预测; (2)制定合理的防治方案。")
21
导致滥用农药现象严重,抗药性产生,药效下降(或出现药害)
(1)用药次数增多 (2)单一品种 (3)施药不及时 (4)剂量不准确 (5)喷施药液量不足
用药次数增多. (2)单一品种. (3)施药不及时. (4)剂量不准确. (5)喷施药液量不足.")
22
第二节 植物病害化学防治策略 和植物病害化学防治原理

23
消灭病害——“可忍受”共存,强调综合治理。
一、植物病害化学防治策略 消灭病害——“可忍受”共存,强调综合治理。 1. 搞好田间卫生:如铲除越冬(越夏)病菌,减少初次侵染源。 2. 合理的栽培管理 如轮栽、调节播种期和收获期,减少寄主感病期与病原物活动期之间的吻合。 3. 生物防治 4. 化学防治和抗病品种的应用
病菌,减少初次侵染源。 2. 合理的栽培管理 如轮栽、调节播种期和收获期,减少寄主感病期与病原物活动期之间的吻合。 3. 生物防治. 4. 化学防治和抗病品种的应用.")
24
此外,在化学防治中使用药剂,必须考虑: (1) 病害种类 (2) 作物:衡量经济价值 (3) 药剂的组合 (4) 时期
 病害种类 (2) 作物:衡量经济价值 (3) 药剂的组合 (4) 时期")
25
二、植物病害化学防治原理 (一) 化学保护protection 在植物得病之前,使用杀菌剂消灭病菌或防治病菌侵入,主要有两个途径: 1. 在菌源上施药 (1)处理病菌越冬场所、中间寄主或土壤。 对于一些土传病害,如棉花黄萎病等,可于秋天翻地时用杀菌剂进行土壤处理。

26
(2)处理带菌种子和苗木。 对于以种子带菌为主的病害,如小麦腥黑穗病菌,采用药剂拌种常可收到十分满意的效果。 石灰水浸种对小麦散黑穗病菌、稻瘟病菌等也很有效。
处理带菌种子和苗木。 对于以种子带菌为主的病害,如小麦腥黑穗病菌,采用药剂拌种常可收到十分满意的效果。 石灰水浸种对小麦散黑穗病菌、稻瘟病菌等也很有效。")
27
(3)处理发病中心 田间发病中心对田间未发病的植株来说,也是一种菌源。 在苗期稻瘟病田间中心施药,对于减轻此病的流行很有效果。
处理发病中心 田间发病中心对田间未发病的植株来说,也是一种菌源。 在苗期稻瘟病田间中心施药,对于减轻此病的流行很有效果。")
28
2. 在保护对象上施药 在田间生长的,尚未发病而可能被侵染的植株上施药是化学保护最有效的一条途径。 此种杀菌剂 (1)残效期较长; (2)一般以WP、胶悬剂、胶体剂为主; (3)要求全覆盖,粘着性能好。
残效期较长; (2)一般以WP、胶悬剂、胶体剂为主; (3)要求全覆盖,粘着性能好。 .")
29
(二) 化学治疗cureness 在植株感病或发病后,施用杀菌剂,是之对植物或病菌发生作用,从而改变病菌的致病过程,达到减轻或消灭病害的目的,主要有三种方法: 1. 表面化学治疗 有些病菌主要在植物表面附着,如小麦白粉病菌以吸孢或吸盘伸入寄主植物组织,施用石硫合剂,可使菌丝萎缩、脱落,可称铲除作用。用于表面化学治疗的不一定是内吸性杀菌剂。

30
2. 内部化学治疗 真正的化学治疗。 药剂进入,直接(或经转化)作用于病原菌。用于内部化学治疗的杀菌剂必须是内吸性杀菌剂。如粉锈宁防治小麦条锈病,井岗霉素防治水稻纹枯病。
作用于病原菌。用于内部化学治疗的杀菌剂必须是内吸性杀菌剂。如粉锈宁防治小麦条锈病,井岗霉素防治水稻纹枯病。")
31
3. 外部化学治疗 与表面化学治疗相似,主要用于果树病害防治。
3. 外部化学治疗 与表面化学治疗相似,主要用于果树病害防治。 在树干或大枝条树皮被侵染、发病后,先用刀将病部刮去,然后在伤口处用以杀菌剂,涂以保护剂、防水剂。这种方法在辽宁防治苹果腐烂病方面得到了大力推广。

32
(三) 化学免疫 免疫是一种生物固有的抗病能力,这种抗病性可以遗传,而化学免疫是利用化学物质使植物产生这种抗病性。
(三) 化学免疫 施用药剂以提高植物本身的抵抗能力,免于发病,即为化学免疫。这是一种间接的植物病害防治方法。 免疫是一种生物固有的抗病能力,这种抗病性可以遗传,而化学免疫是利用化学物质使植物产生这种抗病性。
 化学免疫. 施用药剂以提高植物本身的抵抗能力,免于发病,即为化学免疫。这是一种间接的植物病害防治方法。 免疫是一种生物固有的抗病能力,这种抗病性可以遗传,而化学免疫是利用化学物质使植物产生这种抗病性。")
33
植物抗病性的出现是由于植物细胞内潜在的抗性基因表达的结果,其表达可以通过生物或非生物的诱导作用来实现。
如噻二唑类杀菌剂“Bion”(生物活化剂) 10-50g/hm2, 可防治水稻稻瘟病、白粉病、霜霉病等
 10-50g/hm2, 可防治水稻稻瘟病、白粉病、霜霉病等.")
34
这类化学诱导剂,作为新型杀菌剂的研究,十分活跃,但成功的例子还不多。
应注意: 化学治疗是药剂本身或代谢物作用与病菌; 化学免疫是植物本身对病菌的抵抗,而这种抵抗是由于施用药剂后诱发植物产生抗病物质。

35
三、使用技术及问题 方法:田间喷雾、种子处理、土壤处理 1. 主要是田间喷洒作业,应注意 (1)预测预报
1. 主要是田间喷洒作业,应注意 (1)预测预报 (2)药剂:药剂的种类和浓度、喷洒时期和次数。 对症、粘着性、持久性、重复性、药害
预测预报. (2)药剂:药剂的种类和浓度、喷洒时期和次数。 对症、粘着性、持久性、重复性、药害.")
36
2. 种苗消毒 浸种:50kg水稻种子/63kg药液 拌种:0.2~0.5% 3. 土壤处理 一般10~100ppm(ai/土重15cm) (1)浇灌:2.5~5kga水/m2 (2)施于土表,翻混 (3)注射:25孔/m2; 10ml/孔;深度15-20cm
注射:25孔/m2; 10ml/孔;深度15-20cm.")
37
应遵循: 有效、经济、安全、合理的原则 (1)选用最便宜的有效品种 (2)采用较低的作用量 (3)最少的施药次数 (4)最简便的施药方法
选用最便宜的有效品种 (2)采用较低的作用量 (3)最少的施药次数 (4)最简便的施药方法")
38
第三节 杀菌剂分类 分类依据 杀菌剂类型 作用方式 保护性杀菌剂、内吸性杀菌剂、铲除性杀菌剂、植物诱导抗病激活剂 使用方法
第三节 杀菌剂分类 分类依据 杀菌剂类型 作用方式 保护性杀菌剂、内吸性杀菌剂、铲除性杀菌剂、植物诱导抗病激活剂 使用方法 土壤消毒剂、根部浇灌剂、种子处理剂、叶面喷洒剂、烟雾熏蒸剂、果实保护剂 作用机制 能量生成抑制剂 (巯基抑制剂、电子传递抑制剂、氧化磷酸化抑制剂、糖酵解抑制剂,脂肪酸β-氧化抑制剂等) 生物合成抑制剂 (细胞壁功能及其合成抑制剂、细胞膜功能及其合成抑制剂、蛋白质合成抑制剂、核酸合成抑制剂、甾醇生物合成抑制剂) 化学组成及来源 无机杀菌剂 (硫制剂、铜制剂) 有机杀菌剂 (有机汞、铜、锡;有机磷、砷;二硫代氨基甲酸类;取代苯类;苯并咪唑类,苯基酰胺类;二甲亚酰胺类;氨基甲酸酯类;取代醌类;硫氰酸类;取代甲醇类;杂环类等(吡咯苯类、三唑类、咪唑类、羧酰替苯胺类、吗啉类、苯胺嘧啶类、甲氧基丙烯酸酯类)
 生物合成抑制剂. (细胞壁功能及其合成抑制剂、细胞膜功能及其合成抑制剂、蛋白质合成抑制剂、核酸合成抑制剂、甾醇生物合成抑制剂) 化学组成及来源. 无机杀菌剂. (硫制剂、铜制剂) 有机杀菌剂. (有机汞、铜、锡;有机磷、砷;二硫代氨基甲酸类;取代苯类;苯并咪唑类,苯基酰胺类;二甲亚酰胺类;氨基甲酸酯类;取代醌类;硫氰酸类;取代甲醇类;杂环类等(吡咯苯类、三唑类、咪唑类、羧酰替苯胺类、吗啉类、苯胺嘧啶类、甲氧基丙烯酸酯类)")
39
常见杀菌剂分类汇总

40
目前全球销售的主要杀菌剂品种有160多个,超过15种化学结构类型,明确的作用机理近40种。
从应用作物分析,与除草剂、杀虫剂有所不同,谷物、水果和蔬菜、非农用药、大豆是杀菌剂的最大市场,均超过10% 。

41
2011年 十大杀菌剂品种销售额情况 从品种数量分析,十大杀菌剂品种仅占到杀菌剂品种数量的6.2%,但其销售额却超过了杀菌剂市场份额的40% 。

42
第四节 杀菌剂作用机制

43
1 琥珀酸脱氢酶 2 蛋白激酶(他渗透传感器) 3 RNA 聚合酶 4 小柱孢酮脱水酶 5 甾醇Δ14还原酶 6 非偶联剂 7 甲硫氨酸生物合成 8 蛋白激酶(渗透传感) 9 磷脂的生物合成 10 蛋白生物合成(ribosomes) 11 甾醇3-酮还原酶 12 ATP 合成酶 13 几丁质生物合成 14 二氢乳清酸脱氢酶 15 肌醇生物合成 16 其他, 未知
 11 甾醇3-酮还原酶. 12 ATP 合成酶. 13 几丁质生物合成. 14 二氢乳清酸脱氢酶. 15 肌醇生物合成. 16 其他, 未知.")
44
1 没有杀菌剂的作用机制真正的搞清楚了,这仅是大多数人可接受的观点。
2. 病原菌的致死和被抑制往往不是单方面的作用,而是多方面的综合作用。

45
对杀菌剂作用原理目前存在几种讨论方法 1. 以药剂的种类进行论述。即多作用点的(如传统保护性杀菌剂)和单一作用位点的(如内吸性杀菌剂)。
2. 以作用于菌体的不同部位进行论述。菌体细胞壁、细胞膜、线粒体、核糖体、细胞核。 3. 以对菌体的代谢影响进行研究。 (1) 对菌体呼吸的影响。 (2) 对菌体生物合成的影响。 (3) 对细胞分裂的影响。 (4) 对菌体内酶系统的影响。
 对菌体呼吸的影响。 (2) 对菌体生物合成的影响。 (3) 对细胞分裂的影响。 (4) 对菌体内酶系统的影响。")
46
一、杀菌剂对菌体细胞结构和功能的破坏 杀菌剂对细胞壁的影响 几丁质 真菌 微纤维 纤维素 细胞壁 细菌: 肽多糖 无定形物 、

47
主要有三个方面的破坏: (1)溶解和破坏菌体的细胞壁组成的部分物质 (2)抑制细胞壁附近的一些酶(如糖酶) (3)细胞壁形成受阻
几丁质受损是功能最严重的破坏,其它组分的改变也可使菌体中毒。目前应用的杀菌剂主要是阻碍菌体细胞壁的形成。 中毒症状: 真菌,孢子芽管粗糙,末端膨大或扭曲异形,菌丝过度分枝。 如:苯来特、十三吗啉、多氧酶素 细菌:原生质裸露,继而瓦解。 如:青霉素

48
抑制磷脂酰乙醇胺甲基转移酶 活性,卵磷脂(磷脂酰胆碱) 合成受阻而破坏细胞质膜结构
 合成受阻而破坏细胞质膜结构")
49
2. 破坏菌体细胞膜 细胞膜是有许多亚单位组成的: 类脂质 蛋白质 甾醇 盐类。 这些亚单位有金属桥和疏水键连接起来。 (1) 有机硫杀菌剂 与膜上亚单位连接的疏水键或金属桥结合,致使膜结构破坏,出现裂缝、孔隙、细胞膜结构失去正常生理功能。

50
(2) 含重金属元素的杀菌剂 作用与细胞上的三磷酸腺苷水解酶的-SH,从而改变膜的透性; (3)有机磷杀菌剂 抑制菌体细胞膜上的卵磷脂合成过程中的转移甲基反应,药剂作用点是这一反应的甲基转移酶。 (4) 甾醇合成抑制剂 开发非常迅速,目前已有20多个重要品种。 主要影响甾醇合成过程中鱼鲨烯形成甾醇的阶段。 包括:抑制麦角甾醇的生物合成(三唑类、咪唑类、嘧啶类,P450催化,甾醇C14上脱甲基;吗啉类作用异构酶)
 甾醇合成抑制剂. 开发非常迅速,目前已有20多个重要品种。 主要影响甾醇合成过程中鱼鲨烯形成甾醇的阶段。 包括:抑制麦角甾醇的生物合成(三唑类、咪唑类、嘧啶类,P450催化,甾醇C14上脱甲基;吗啉类作用异构酶)")
51
3.杀菌剂破坏菌体内部多种细胞器或其结构 导致深刻的、致命的菌体细胞代谢的变化。在后面的生物氧化中讨论。

52
二、杀菌剂对菌体生物氧化的影响 糖原 脂肪 蛋白质 Ⅰ 葡萄糖 脂肪酸 甘油 氨基酸 Ⅱ 乙酰CoA TCA Ⅲ 生物氧化过程

53
呼吸抑制剂作用靶标 (1)催化物质氧化降解的专化性酶 (2)或电子传递过程中的专化性载体 生物氧化过程(仿Corbett等,1984)
生物体的能量主要来源于细胞呼吸作用。杀菌剂抑制病菌呼吸作用的结果是破坏能量的生成,导致菌体死亡。 大多数传统多作用位点杀菌剂和一些现代选择性杀菌剂,它们的作用靶标正是病菌呼吸作用过程中催化物质氧化降解的专化性酶或电子传递过程中的专化性载体,属于呼吸抑制剂 。但是,传统多作用位点杀菌剂的作用靶标多为催化物质氧化降解的非特异性酶,菌体在物质降解过程中释放的能量较少,所以这些杀菌剂不仅表现活性低,而且缺乏选择性。电子传递链中的一些酶的复合物抑制剂及氧化磷酸化抑制剂往往表现很高的杀菌活性和选择性。 病原菌的不同生长发育期对能量的需要量不同 ,真菌孢子萌发要比维持菌丝生长所需要的能量多得多,因而能量供应受阻时,孢子就不能萌发,呼吸抑制剂对孢子萌发的毒力也往往显著高于菌丝生长的毒力。 由于有氧呼吸是在线粒体内进行的,所以许多对线粒体结构有破坏作用的杀菌剂,也会干扰有氧呼吸而破坏能量生成。 病原菌的生命过程需要能量,尤其是孢子萌发,更需要较多的能量供应。这些能量来自碳水化合物、脂肪和蛋白质氧化最终生成的ATP,见图生物氧化过程(仿Corbett等,1984)) 生物氧化过程(仿Corbett等,1984)
) 生物氧化过程(仿Corbett等,1984)")
54
二、杀菌剂对菌体生物氧化的影响 (1) 糖酵解 (2) 脂肪酸氧化 (3) 三羧酸循环 (4) 吸链电子转移(传递)的影响(包括末端氧化) (5) 对氧化磷酸化的影响
 糖酵解 (2) 脂肪酸氧化 (3) 三羧酸循环 (4) 吸链电子转移(传递)的影响(包括末端氧化) (5) 对氧化磷酸化的影响")
55
1. 对糖酵解的影响 糖酵解在细胞质中进行。根据目前的认识比较次要。 (1) 铜汞制剂 破坏细胞膜,使一些金属离子,主要是K离子向细胞外渗透,而菌体内糖酵解的过程中最重要的是磷酸果糖激酶的活性由K离子来活化,丙酮酸激酶也需要K离子作为辅助因子。

56
(2)硫磺:作用己糖激酶 (3)克菌丹:影响乙酰辅酶A的形成,从而影响丙酮酸脱羧作用。 (4)百菌清:菌体内参与3-磷酸甘油醛——1,3-二磷酸,甘油酸代谢过程的磷酸甘油醛脱氢酶是其作用位点。
硫磺:作用己糖激酶 (3)克菌丹:影响乙酰辅酶A的形成,从而影响丙酮酸脱羧作用。 (4)百菌清:菌体内参与3-磷酸甘油醛——1,3-二磷酸,甘油酸代谢过程的磷酸甘油醛脱氢酶是其作用位点。")
57
杀菌剂对糖酵解、有氧氧化和磷酸戊糖三条途径的影响

58
2. 对脂肪酸-β氧化的影响 脂肪酸在生物体内的降解氧化有 需氧 氧化(α-氧化) 无氧氧化(β-氧化):与杀菌剂作用有关 β-氧化过程,均需乙酰辅酶A参予。 例如:菌体内多碳长链脂肪酸,在CoA-SH帮助下,通过β位碳原子的氧化(脱氢),而使其断裂。因此,反作用于CoA-SH的杀菌剂如克菌丹、二氯萘醌和代森类显然都会阻碍菌体的脂肪酸氧化,但至今未发现转化性杀菌剂。 作用于CoA-SH,影响脂肪酸-β氧化
,而使其断裂。因此,反作用于CoA-SH的杀菌剂如克菌丹、二氯萘醌和代森类显然都会阻碍菌体的脂肪酸氧化,但至今未发现转化性杀菌剂。 作用于CoA-SH,影响脂肪酸-β氧化.")
59
脂肪酸氧化 脂肪酸氧化时先在细胞质中活化,生成酯酰辅酶A: 生成的乙酰辅酶A进入三羧酸循环

60
3. 三羧酸循环 第一阶段 第二阶段 克菌丹 硫磺、杀枯净 福美双、克菌丹、 二氯萘醌 有机硫代森类、 8-羟基喹啉、 氟代乙酸
三羧酸循环(即Krebs cycle)是所有有机物质初步代谢产物的共同氧化途径,最终产生CO2和水。 对三羧酸循环的影响,杀菌剂对三羧酸循环的影响可分为两个阶段。 (1)是对即将进入三羧酸循环之前的影响。在一些生物内丙酮酸经丙酮酸脱氢酶的作用,并以TPP为辅酶的情况下形成乙醛,乙醛再进一步与辅酶A结合形成乙酰辅酶A。在有些情况下,乙酸可以代替乙醛与辅酶A结合,同样产生乙酰辅酶A,以后则进入三羧酸循环。在进入之前,一些化合物,如对孢子形成有抑制作用的,使三羧酸循环中顺乌头酸到异柠檬酸的代谢过程受阻。 (2)是直接作用于三羧酸循环中的某个环节。这类杀菌剂品种很多。例如:作用于柠檬酸合成酶的福美双、克菌丹、二氯萘醌;作用于乌头酸酶的有机硫代森类、8-羟基喹啉、氟代乙酸;作用于α-酮戊二酸脱氢酶系的克菌丹;作用于琥珀酸脱氢酶系的硫磺、杀枯净等。 克菌丹 硫磺、杀枯净
是所有有机物质初步代谢产物的共同氧化途径,最终产生CO2和水。 对三羧酸循环的影响,杀菌剂对三羧酸循环的影响可分为两个阶段。 (1)是对即将进入三羧酸循环之前的影响。在一些生物内丙酮酸经丙酮酸脱氢酶的作用,并以TPP为辅酶的情况下形成乙醛,乙醛再进一步与辅酶A结合形成乙酰辅酶A。在有些情况下,乙酸可以代替乙醛与辅酶A结合,同样产生乙酰辅酶A,以后则进入三羧酸循环。在进入之前,一些化合物,如对孢子形成有抑制作用的,使三羧酸循环中顺乌头酸到异柠檬酸的代谢过程受阻。 (2)是直接作用于三羧酸循环中的某个环节。这类杀菌剂品种很多。例如:作用于柠檬酸合成酶的福美双、克菌丹、二氯萘醌;作用于乌头酸酶的有机硫代森类、8-羟基喹啉、氟代乙酸;作用于α-酮戊二酸脱氢酶系的克菌丹;作用于琥珀酸脱氢酶系的硫磺、杀枯净等。 克菌丹. 硫磺、杀枯净.")
61
TCA在线粒体内进行,参与TCA的酶都分布在线粒体膜、基质和液泡中。因而,对TCA的影响主要是对这些酶活性的影响。
(1)影响CoA,使其失活。福美双、克菌丹、硫磺、 二氯萘醌 (2) 与乌头酸酶鳌合:有机硫、代森类、8羟基喹啉 (3) 酮戊二酸脱氢酶:克菌丹 (4) 抑制琥珀酸、苹果酸脱氢酶:硫磺、萎锈灵 (5) 抑制延胡羧酸酶活性:含铜杀菌剂 关键酶,但并不是单一位点。
影响CoA,使其失活。福美双、克菌丹、硫磺、 二氯萘醌. (2) 与乌头酸酶鳌合:有机硫、代森类、8羟基喹啉. (3) 酮戊二酸脱氢酶:克菌丹. (4) 抑制琥珀酸、苹果酸脱氢酶:硫磺、萎锈灵. (5) 抑制延胡羧酸酶活性:含铜杀菌剂. 关键酶,但并不是单一位点。")
62
4. 呼吸链电子转移(传递)的影响(包括末端氧化)
4. 呼吸链电子转移(传递)的影响(包括末端氧化) 呼吸电子传递链(仿Corbett等,1984) 有4个关键酶复合物参与了从NADH和FADH2到O2的电子传递,并通过电子传递产生ATP。 1=NADH-辅酶Q氧化还原酶 =琥珀酸辅酶Q氧化还原酶 3=辅酶Q细胞色素c氧化还原酶 = 细胞色素a-a3(末端)氧化酶
的影响(包括末端氧化) 呼吸电子传递链(仿Corbett等,1984) 有4个关键酶复合物参与了从NADH和FADH2到O2的电子传递,并通过电子传递产生ATP。 1=NADH-辅酶Q氧化还原酶 2=琥珀酸辅酶Q氧化还原酶. 3=辅酶Q细胞色素c氧化还原酶 4= 细胞色素a-a3(末端)氧化酶.")
63
Succinate Dehydrogenase
Ubiquinone (UQ)泛醌 UQ and the inhibitor binding site (QP) second UQ binding site (QD)线粒体有外线粒体膜(OMM)、内线粒体膜(IMM)和脊组成。氧化作用的底物由特定的载体蛋白(如丙酮酸转运体,PyrT)运载到线粒体内。 乙酰辅酶A脱氢酶、丙酮酸脱氢酶和三羧酸循环的还原物通过NADH、琥珀酸-泛醌氧化还原酶(SQQ)、电子转移黄素蛋白(ETF)和电子传递黄素蛋白泛醌氧化还原酶(ETFQO)转移到电子传递链中。 NADH被复合物I再氧化,还原后的辅酶Q(UQ)通过复合物III,细胞色素c和复合物IV进行电子转移。 三个复合物“泵”出的质子氢通过膜是所产生的电化学梯度通过ATP合成酶合成ATP。 磷酸的进入和ATP/ADP在细胞溶质中的交换通过相应的磷酸载体(PC)和ATP/ADP载体调节 Ubiquinone (UQ)泛醌 cytochrome c oxidase F1F0ATP Synthase
泛醌. UQ and the inhibitor binding site (QP) second UQ binding site (QD)线粒体有外线粒体膜(OMM)、内线粒体膜(IMM)和脊组成。氧化作用的底物由特定的载体蛋白(如丙酮酸转运体,PyrT)运载到线粒体内。 乙酰辅酶A脱氢酶、丙酮酸脱氢酶和三羧酸循环的还原物通过NADH、琥珀酸-泛醌氧化还原酶(SQQ)、电子转移黄素蛋白(ETF)和电子传递黄素蛋白泛醌氧化还原酶(ETFQO)转移到电子传递链中。 NADH被复合物I再氧化,还原后的辅酶Q(UQ)通过复合物III,细胞色素c和复合物IV进行电子转移。 三个复合物 泵 出的质子氢通过膜是所产生的电化学梯度通过ATP合成酶合成ATP。 磷酸的进入和ATP/ADP在细胞溶质中的交换通过相应的磷酸载体(PC)和ATP/ADP载体调节. Ubiquinone (UQ)泛醌. cytochrome c oxidase. F1F0ATP Synthase.")
64
杀菌剂对真菌线粒体呼吸链的作用点 ATP是能量贮存“库”,可在呼吸链三个位点形成。
因此,呼吸链电子传递的干扰是杀菌剂的重要作用机理之一。敌克松、萎锈灵、敌枯双 一般认为呼吸链的复合物I、II、III、IV的四个部位都有杀菌剂的作用点。

65
5. 对氧化磷酸化的影响 这个过程是生物细胞内形成ATP的一种极为复杂的代谢过程,其机理至今也只是一般推理。 两个方面:
5. 对氧化磷酸化的影响 这个过程是生物细胞内形成ATP的一种极为复杂的代谢过程,其机理至今也只是一般推理。 两个方面: (1)化合物改变线粒体内膜透性或其他未知原因消除了内膜两侧原来形成的H+浓度和电位差,从而阻碍了能的供应。二硝基酚 (2)直接影响ATP酶的活性,如砷、汞、铜
化合物改变线粒体内膜透性或其他未知原因消除了内膜两侧原来形成的H+浓度和电位差,从而阻碍了能的供应。二硝基酚. (2)直接影响ATP酶的活性,如砷、汞、铜.")
66
三、杀菌剂对菌体细胞代谢的影响 ——干扰生物合成 主要有:对细胞壁组分合成的影响 对细胞膜上甾醇合成的影响 对核酸合成的影响
对蛋白质合成的影响

67
1. 杀菌剂对菌体核酸合成和功能的影响 (1)掺假核酸的作用。 杀菌剂与菌体内核酸碱基的化学结构相似,因而代替了核苷酸的碱基。就造成了所谓的“掺加的核酸” 苯并咪唑类药剂,如苯来特、多菌灵
掺假核酸的作用。 杀菌剂与菌体内核酸碱基的化学结构相似,因而代替了核苷酸的碱基。就造成了所谓的 掺加的核酸 苯并咪唑类药剂,如苯来特、多菌灵")
68
(2)杀菌剂与形成碱基的组分结构相似,竞争性抑制
干扰了核酸合成过程中的某一个反应,使核酸合成不能完成。 如6-氮杂尿嘧啶(6-azauracil)竞争性抑制尿嘧啶合成过程中尿嘧啶-6-甲酸核苷-5’-磷酸的脱羧作用。
竞争性抑制尿嘧啶合成过程中尿嘧啶-6-甲酸核苷-5’-磷酸的脱羧作用。")
69
(3)杀菌剂通过影响菌体叶酸而干扰核酸的合成
如敌锈钠。 (4) 影响核酸的聚合 抗生素(放线菌素D、丝裂霉素)等影响菌体内DNA的生物活性,使在DNA指令的RNA聚合酶作用下的RNA生物合成受抑制。 另外,甲霜灵,抑制核糖体rRNA的合成,使三磷酸核苷大量积累。
 影响核酸的聚合. 抗生素(放线菌素D、丝裂霉素)等影响菌体内DNA的生物活性,使在DNA指令的RNA聚合酶作用下的RNA生物合成受抑制。 另外,甲霜灵,抑制核糖体rRNA的合成,使三磷酸核苷大量积累。")
70
(5) 影响细胞分裂 药剂通过与构成纺锤丝的管蛋白的亚单位(微管蛋白)结合而阻碍管蛋白的形成,从而破坏纺锤丝的功能,使有丝分裂不能正常进行,染色体分离紊乱。如苯并咪唑类杀菌剂。 苯并咪唑类杀菌剂作用机制示意图
 影响细胞分裂 药剂通过与构成纺锤丝的管蛋白的亚单位(微管蛋白)结合而阻碍管蛋白的形成,从而破坏纺锤丝的功能,使有丝分裂不能正常进行,染色体分离紊乱。如苯并咪唑类杀菌剂。 苯并咪唑类杀菌剂作用机制示意图.")
71
2. 杀菌剂对蛋白质合成和功能的影响 蛋白质合成是在核糖体上进行,tRNA、mRNA、rRNA 药剂蛋白质合成或使蛋白质变性,其中毒现为 (1)菌体细胞内的蛋白质合成减少,含量降低,菌体生长受到明显抑制; (2)菌体内游离氨基酸增多;菌体生长明显受到抑制
菌体内游离氨基酸增多;菌体生长明显受到抑制.")
72
第一个方面:药剂对蛋白质合成的直接影响 主要是一些农用抗菌素类杀菌剂。 I 阻止f-met-tRNA与核糖体的结合
春雷霉素与稻瘟病菌核糖体的40s或30s小亚基结合,使在蛋白质合成的起始阶段阻止f-met-tRNA与核糖体的结合。 链霉素与苹果、梨、蔷薇大疫病菌核糖体30s小亚基结合(阻止结合到A位上,使f-met-tRNA阻留在P位上),并阻止多聚核糖体变为单核糖体。 四环素与油菜菌核病菌、桑萎缩病、柑桔黄龙病等类菌原体的30s小亚基结合,另外对70s、80 s小亚基均有作用,70s小亚基更敏感。
,并阻止多聚核糖体变为单核糖体。 四环素与油菜菌核病菌、桑萎缩病、柑桔黄龙病等类菌原体的30s小亚基结合,另外对70s、80 s小亚基均有作用,70s小亚基更敏感。")
73
春雷霉素和灭瘟素的作用机制 这两种农用抗菌素主要用于防治水稻稻瘟病。根据现有的研究资料,春雷霉素和灭瘟素主要是抑制了病原菌的蛋白质合成。
蛋白质合成示意图

74
II 在肽链延长阶段,破坏DNA模板功能,阻止肽链延 长,如放线菌素D
III 导致密码错误:如链霉素作用于30s小亚基上A位点,使翻译过程错认密码,合成无功能 “错误蛋白” IV 作用于终止阶段(转肽酶):稻瘟散、链霉素等作用于稻瘟病菌、细菌的60s亚基 V 整个合成过程受到抑制和促进 如Na+对植物细胞,阻止mRNA的复制、影响合成 Na+对病毒,膜透性改变,刺激了合成
:稻瘟散、链霉素等作用于稻瘟病菌、细菌的60s亚基. V 整个合成过程受到抑制和促进. 如Na+对植物细胞,阻止mRNA的复制、影响合成. Na+对病毒,膜透性改变,刺激了合成.")
75
第二个方面:药剂对蛋白质合成的间接影响 I 影响核酸合成的杀菌剂,都影响蛋白质合成 II 某些与氨基酸类似的化合物也会影响蛋白质的合成 如青霉素与多种细菌作用,其结构与细菌细胞壁的主要成分粘肽的前身化合物(丙氨酰丙氨酸)结构中一部分立体结构相似,从而抑制了蛋白质的合成。
结构中一部分立体结构相似,从而抑制了蛋白质的合成。 .")
76
III 蛋白质合成过程中某些酶的活性受到抑制
如异硫氰酯类的化合物就是有关酶的-SH辅基反应而抑制其活性。 IV 影响能量生成的杀菌剂,对菌体能量生成的阻碍,间接也使蛋白质合成受阻。 如内吸杀菌剂萎锈灵。

77
四、对菌无毒性的保护剂作用原理 主要干扰病菌寄生(调节病菌致病系统)的化合物、加强寄主防御作用的化合物。 特点:
(1)在离体条件下,很少或没有杀菌活性,但当应用到寄主植物上时可减少侵染或境地病害的严重度。
在离体条件下,很少或没有杀菌活性,但当应用到寄主植物上时可减少侵染或境地病害的严重度。")
78
(2) 活体表现: I 在寄主植物体内转变为具有毒性的化合物 II 影响真菌的致病能力 III 影响寄主的抗病性 IV 可以干扰致病原因的关键因素(真菌毒素、酶的活性、产物)和寄主抗病性的诱导剂或抑制剂
和寄主抗病性的诱导剂或抑制剂.")
79
对病原菌本身没有毒性或几乎没有毒性。 ※ 具有更高的选择性,对非靶标生物和环境安全 ※ 由于对病原菌本身无毒性,故不易产生抗药性。

80
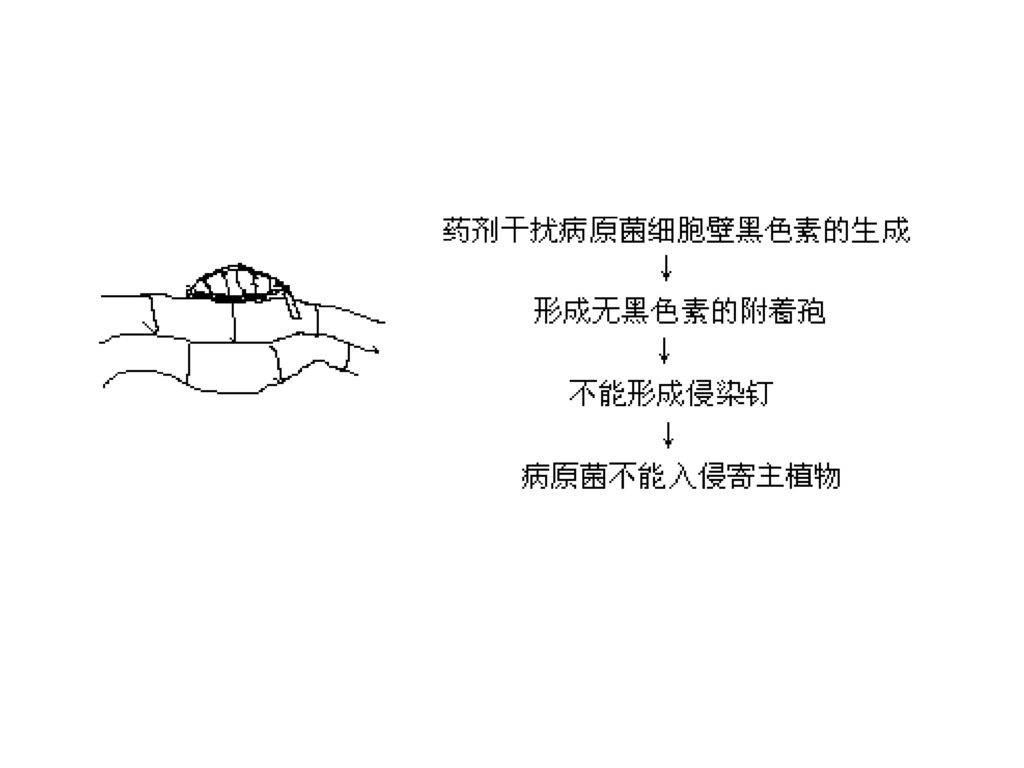
1. 黑色素生物合成抑制剂 阻碍附着孢对植物细胞壁的穿透。三环唑(第一个),防治稻瘟病
,防治稻瘟病")
81
2. 影响寄主—病原物关系的化合物 调节寄主植物抗病性
植物受病原物诱导的产生系统的抗病性能,称作系统获得性抗病性(Systemic Acguired Resistance, SAR) 除病原物外,某些化学药品亦具有类似功能,这类化学物质就是植物抗病激活剂(Plant Activator) 生物因子、物理因子或化学因子 ↓ 激发寄主受体产生防卫信号 激活或诱导植物防卫基因表达
 除病原物外,某些化学药品亦具有类似功能,这类化学物质就是植物抗病激活剂(Plant Activator) 生物因子、物理因子或化学因子. ↓ 激发寄主受体产生防卫信号. 激活或诱导植物防卫基因表达.")
82
特点: ※ 本身对病原菌没有明显的杀菌或抑菌活性,但能诱发植物自身的免疫系统,抵御病害侵袭。 ※ 一种抗病激活剂可以防治多种病害,植物一旦产生系统抗病性,其抗病性具有持久性和广谱性,甚至可以遗传。 ※ 不会对病原菌产生选择压力,不易产生抗药性。

83
例如:苯并噻二唑(BTH)类 30g/ha对小麦白粉病具有持久性抗性。
气巴-嘉基在开发磺酰脲类除草剂时发现,苯并噻二唑-7-羧酸甲酯有SAR活性 诺华公司商品名Bion 30g/ha对小麦白粉病具有持久性抗性。

84
五、以药剂的种类总结 影响生物氧化

85
1 硫代氨基甲酸酯类杀菌剂的作用机制 ① 破坏辅酶A, 代森锌 福美双

86
② 抑制以铜、铁等为辅基的酶的活性。

87
2 取代苯类杀菌剂的作用机制 百菌清的作用点是参与3-磷酸甘油醛到1,3-二磷酸甘油酸代谢过程的磷酸甘油醛脱氢酶。
2 取代苯类杀菌剂的作用机制 百菌清的作用点是参与3-磷酸甘油醛到1,3-二磷酸甘油酸代谢过程的磷酸甘油醛脱氢酶。 百菌清与该酶上的-SH基结合,生成一种复合物,从而使酶的活性丧失。

88
从3-磷酸甘油醛到1,3-二磷酸甘油酸的反应的催化机理是磷酸甘油醛脱氢酶活性位置上半胱氨酸残基的-SH基是亲核基团,它与醛基作用形成中间产物,可将羟基上的氢移至与酶紧密结合的NAD+上,从而产生NADH和高能硫酯中间产物。NADH从酶上解离,另外的 NAD+与酶活性中心结合,磷酸攻击硫酯键从而形成1,3-二磷酸甘油。 百菌清和该酶的-SH结合,抑制其活性,中断糖酵解,从而影响ATP的生成。

89
3 羧酰苯胺类杀菌剂的作用机制 如萎锈灵、氟酰胺等。 作用于呼吸电子传递链中从琥珀酸到辅酶Q之间的氧化还原体系。即复合体II。
3 羧酰苯胺类杀菌剂的作用机制 如萎锈灵、氟酰胺等。 作用于呼吸电子传递链中从琥珀酸到辅酶Q之间的氧化还原体系。即复合体II。 复合体Ⅱ是由黄酶Ⅱ(FAD)为辅酶的黄素蛋白、非血红素铁硫蛋白及其他结合蛋白组成的。 底物(琥珀酸)脱出的 2个H传递给FAD—FADH2,而Fe3+将FADH2氧化成FAD,放出2个H, Fe3+→Fe2+,这2个H又被CoQ接受,成 CoQH2
为辅酶的黄素蛋白、非血红素铁硫蛋白及其他结合蛋白组成的。 底物(琥珀酸)脱出的 2个H传递给FAD—FADH2,而Fe3+将FADH2氧化成FAD,放出2个H, Fe3+→Fe2+,这2个H又被CoQ接受,成 CoQH2.")
90
萎锈灵对复合体Ⅱ的作用(仿Lyf,1987) 萎锈灵和复合体Ⅱ的活性中心——非血红素铁硫蛋白结合,从而阻断了电子向辅酶Q的传递;
在模式图中,有两个萎锈灵分子嵌进铁硫蛋白活性中心,萎锈灵分子中的甲基和苯基锚在铁硫蛋白的疏水区,从而整个地改变了蛋白的结构。

91
4 甲氧丙烯酸酯类杀菌剂的作用机制 ICI A5504 小麦赤霉、葡萄霜霉、白粉 BAF 490F 苹果黑星、白粉、稻瘟
4 甲氧丙烯酸酯类杀菌剂的作用机制 1997年 Anke 和Oberwinkler 担子菌中先导化合物 ICI A5504 小麦赤霉、葡萄霜霉、白粉 BAF 490F 苹果黑星、白粉、稻瘟

92
作用部位:复合体III,即Cytb/Cytc1
杀菌剂和复合体III中的活性部分Cytb的Q0-中心结合,从而阻止电子从Cytb→Cytc1的转移,阻碍ATP的产生。 噁唑烷二酮类杀菌剂恶唑菌酮(famoxadone)和咪唑菌酮(fenamidone)虽然结构上和Strobilurins毫无相干,但却具有相同作用机制。 Strobilurin A是Anke和Oberwinkler(1977)从担子菌中分离的天然抗菌活性物质。近年来以Strobilurin A为先导化合物,开发出一类新型杀菌剂——甲氧丙烯酸酯类,代表品种有嘧菌酯(Azoxystrobin)、醚菌酯(Kresoxim-methy, BAS490F)、肟菌酯(Trifloxystrobin)、苯氧菌胺(Metominostrobin)和啶氧菌酯(Picoxystrobin)等[71,72]。甲氧丙烯酸酯类杀菌剂同样抑制了病原菌线粒体呼吸电子传递链中电子的传递,其作用部位是复合体Ⅲ (即细胞色素b和细胞色素C1复合体)。据研究,甲氧丙烯酸酯类杀菌剂就是通过和复合体Ⅲ中的活性部位结合而抑制了线粒体的电子传递[73]。 此外,抗菌素抗霉素A(antimycin A)和杀菌剂杀枯净 (phenazine,5-氧吩嗪)也是作用于复合体Ⅲ。
和咪唑菌酮(fenamidone)虽然结构上和Strobilurins毫无相干,但却具有相同作用机制。 Strobilurin A是Anke和Oberwinkler(1977)从担子菌中分离的天然抗菌活性物质。近年来以Strobilurin A为先导化合物,开发出一类新型杀菌剂——甲氧丙烯酸酯类,代表品种有嘧菌酯(Azoxystrobin)、醚菌酯(Kresoxim-methy, BAS490F)、肟菌酯(Trifloxystrobin)、苯氧菌胺(Metominostrobin)和啶氧菌酯(Picoxystrobin)等[71,72]。甲氧丙烯酸酯类杀菌剂同样抑制了病原菌线粒体呼吸电子传递链中电子的传递,其作用部位是复合体Ⅲ (即细胞色素b和细胞色素C1复合体)。据研究,甲氧丙烯酸酯类杀菌剂就是通过和复合体Ⅲ中的活性部位结合而抑制了线粒体的电子传递[73]。 此外,抗菌素抗霉素A(antimycin A)和杀菌剂杀枯净 (phenazine,5-氧吩嗪)也是作用于复合体Ⅲ。")
93
5 氟定胺 强有力的解偶联剂,破坏氧化磷酸化 欧洲市场,主要用于防治马铃薯晚疫病

94
影响生物合成 1 有机磷杀菌剂的作用机制 主要是抑制了卵磷脂的合成而破坏了细胞质膜的结构。

95
卵磷脂的合成必需要有磷脂酰乙醇胺甲基转移酶的参与。异稻瘟净的作用机制主要是抑制磷脂酰乙醇胺甲基转移酶的活性,阻断了卵磷脂合成
卵磷脂生物合成途径

96
异稻瘟净主要是抑制磷脂酰乙醇胺甲基转移酶活性,阻断卵磷脂合成,从而破坏了细胞膜的结构。
克瘟散主要是抑制磷酯酰乙醇胺分子向磷酯酰胆碱分子的转化。由于它在分子结构上与磷酯酰乙醇胺很相似,因此可以成为磷酯酰乙醇胺的代谢拮抗物。对禾谷镰孢菌来说,克瘟散也可以减少磷酯酰胆碱的合成。 此外,对细胞壁形成的干扰可能是此类药剂的第二作用机制。

97
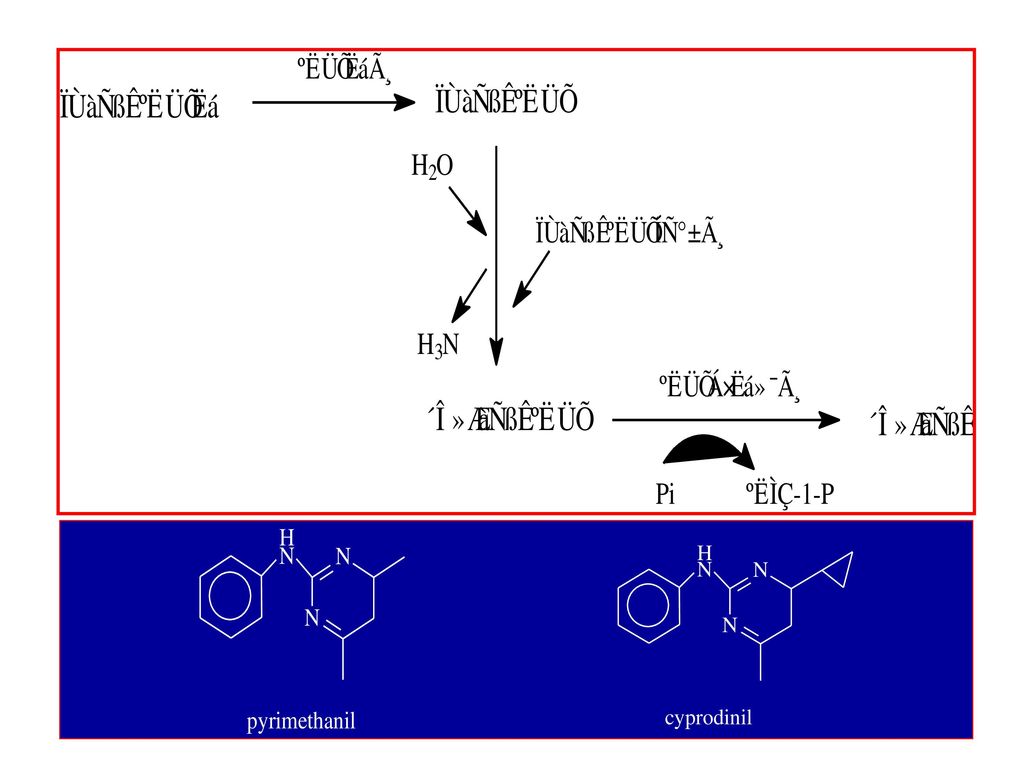
2 嘧啶胺类杀菌剂的作用机制 早期开发的嘧啶胺类杀菌剂有甲菌啶(dimethirimol)和乙菌啶(ethirimol),主要用于防治瓜类和谷物白粉病。关于乙菌啶的作用机制,Hollomoon(1979)曾指出,主要是非竞争性地抑制了腺嘌呤核苷脱氨酶的活性而影响了某些碱基及核酸的合成。 近年来又有几种新的嘧啶苯胺商品化,如嘧菌胺(mepanipyrim),pyrimethanil 和 cyprodinil,对灰葡萄孢引起的多种病害,特别是灰霉病有特效,而且与二甲酰亚胺类杀菌剂无交互抗性。
,pyrimethanil 和 cyprodinil,对灰葡萄孢引起的多种病害,特别是灰霉病有特效,而且与二甲酰亚胺类杀菌剂无交互抗性。")
99
嘧菌胺等杀菌剂的作用机制主要有两种解释:
一是抑制细胞壁降解酶的分泌。 以蚕豆褐斑病菌为试验材料的研究发现: Pyrimethanil对孢子萌发和附着孢的形成没有影响,对病原菌的早期入侵阶段几乎没有影响,但能显著地减少入侵点周围寄主细胞的死亡。 果胶酶 细胞壁降解酶 纤维素酶

100
二是干扰甲硫氨酸(蛋氨酸)的生物合成 嘧菌胺抑制了β-胱硫醚裂解酶的活性。
在寄主植物和病原菌体内,甲硫氨酸是由天冬氨酸合成的。生物合成途径如图所示 。 许多研究结果表明,Pyrimethanil、嘧菌胺抑制了甲硫氨酸生物合成途径中次末端——β-胱硫醚裂解酶(β-cyctathionase)的活性从而抑制了甲硫氨酸的合成,但详细的抑制机理还不清楚 嘧菌胺抑制了β-胱硫醚裂解酶的活性。
的活性从而抑制了甲硫氨酸的合成,但详细的抑制机理还不清楚. 嘧菌胺抑制了β-胱硫醚裂解酶的活性。")
101
3 苯基酰胺类杀菌剂的作用机制 苯基酰胺类杀菌剂至少包括4类:酰基丙氨酸类、丁内酯类、硫代丁内酯类和噁唑烷酮类,其中以酰基丙氨酸类(以甲霜灵为代表)、噁唑烷酮类(以噁霜灵为代表)最重要。这类杀菌剂广泛用于藻菌纲病害(如霜霉病)的防治。关于苯基酰胺类的作用机理,一般认为是抑制了病原菌中核酸的生物合成,主要是 RNA的合成。 细胞各类RNA,包括参与翻译过程的mRNA、rRNA和 tRNA,以及具有特殊功能的小RNA,都是以DNA为模板,在 RNA聚合酶的催化下合成的,真核生物的RNA聚合酶有好多种,分子量大约在 u,通常由4~6种亚基组成,并含有 Zn2+。利用抑制剂α-鹅膏蕈碱的抑制作用可将其分为3类:对抑制剂不敏感的RNA聚合酶A(或Ⅰ),可被低浓度抑制剂抑制的RNA聚合酶B(或Ⅱ),只被高浓度抑制剂抑制的RNA聚合酶C(或Ⅲ)。Hayes等认为,甲霜灵、噁霜灵主要是抑制了对α-鹅膏蕈碱不敏感的RNA聚合酶A,从而阻碍了rRNA前体的转录。具体的抑制机理尚不清楚。
、噁唑烷酮类(以噁霜灵为代表)最重要。这类杀菌剂广泛用于藻菌纲病害(如霜霉病)的防治。关于苯基酰胺类的作用机理,一般认为是抑制了病原菌中核酸的生物合成,主要是 RNA的合成。 细胞各类RNA,包括参与翻译过程的mRNA、rRNA和 tRNA,以及具有特殊功能的小RNA,都是以DNA为模板,在 RNA聚合酶的催化下合成的,真核生物的RNA聚合酶有好多种,分子量大约在 u,通常由4~6种亚基组成,并含有 Zn2+。利用抑制剂α-鹅膏蕈碱的抑制作用可将其分为3类:对抑制剂不敏感的RNA聚合酶A(或Ⅰ),可被低浓度抑制剂抑制的RNA聚合酶B(或Ⅱ),只被高浓度抑制剂抑制的RNA聚合酶C(或Ⅲ)。Hayes等认为,甲霜灵、噁霜灵主要是抑制了对α-鹅膏蕈碱不敏感的RNA聚合酶A,从而阻碍了rRNA前体的转录。具体的抑制机理尚不清楚。")
102
4 麦角甾醇合成抑制剂的作用机制 麦角甾醇,是细胞膜的主要组分。影响膜的通透性功能;此外,它也是一种激素前体,在有性、无性生殖过程中起到主要作用。 麦角甾醇的合成是由甲瓦龙酸开始,其中有一步反应是14位C的脱甲基反应。这步反应是MFO的细胞色素P450来完成的。三唑类杀菌剂和P450中心的Fe结合,抑制了该酶的活性,从而抑制了14C脱甲基,阻止了麦角甾醇的合成。

103
三唑类杀菌剂和P450中心的Fe结合,抑制了该酶的活性,从而抑制了14C脱甲基
吗啉类和哌啶类则主要是抑制了异构酶活性,阻断了Δ8~Δ7之间的异构化[即双键从C-8(9)移到C-7(8)]
移到C-7(8)]")
104
在脱甲基化过程中,甾醇14α-脱甲基化酶催化了氧化脱甲基化反应,使C14位置上的甲基脱去。
对这一过程的抑制是由于P450细胞色素中第6对位上的亚铁离子与甾醇合成抑制剂中氮杂茂部分的N3或N4原子形成复合物造成的。 由于它们之间形成了复合物,使其与羊毛甾醇等的正常结合受到了阻碍。 C-14脱甲基反应是由多功能氧化酶(MFO)中的细胞色素P450来完成的。三唑类杀菌剂抑制C-14脱甲基是因为杀菌剂和P450的结合部位相结合,三唑中的N原子和P450中心的Fe原子的第6位配位成一个复合物,从而抑制了细胞色素P450的脱甲基活性 (图6-30)。此外,C-22双键的引入和C-24(28)双键的加氢还原亦受到抑制[。 导致了功能性甾醇类物质的短缺和羊毛甾醇等14α-甲基甾醇类物质的过多积累。这些变化使膜的流动性发生了改变,因为14α-甲基增加了甾醇类物质的厚度从而使甾醇类物质在膜的双层结构中不能正确地装配。这可能使膜的通透性增强。这些影响最终将对真菌的生长产生抑制。
中的细胞色素P450来完成的。三唑类杀菌剂抑制C-14脱甲基是因为杀菌剂和P450的结合部位相结合,三唑中的N原子和P450中心的Fe原子的第6位配位成一个复合物,从而抑制了细胞色素P450的脱甲基活性 (图6-30)。此外,C-22双键的引入和C-24(28)双键的加氢还原亦受到抑制[。 导致了功能性甾醇类物质的短缺和羊毛甾醇等14α-甲基甾醇类物质的过多积累。这些变化使膜的流动性发生了改变,因为14α-甲基增加了甾醇类物质的厚度从而使甾醇类物质在膜的双层结构中不能正确地装配。这可能使膜的通透性增强。这些影响最终将对真菌的生长产生抑制。")
105
C-14(15)还原酶和△8(9)~△7(8)异构酶作用机制
啉类和哌啶类则主要是抑制了异构酶活性,阻断了Δ8~Δ7之间的异构化[即双键从C-8(9)移到C-7(8)]。此外,这类杀菌剂还抑制C-14(15)双键加氢还原酶的活性。Δ8~Δ7异构酶及C-14(15)还原酶,都是和碳阳离子中间态(图6-31中方括弧部分)结合才起催化作用。吗啉/哌啶杀菌剂在菌体生理pH条件下可质子化,模拟了碳阳离子中间态和这两种酶强烈结合,从而使酶失活。 C-14(15)还原酶和△8(9)~△7(8)异构酶作用机制
移到C-7(8)]。此外,这类杀菌剂还抑制C-14(15)双键加氢还原酶的活性。Δ8~Δ7异构酶及C-14(15)还原酶,都是和碳阳离子中间态(图6-31中方括弧部分)结合才起催化作用。吗啉/哌啶杀菌剂在菌体生理pH条件下可质子化,模拟了碳阳离子中间态和这两种酶强烈结合,从而使酶失活。 C-14(15)还原酶和△8(9)~△7(8)异构酶作用机制.")
106
5 苯并咪唑类杀菌剂 苯并咪唑类杀菌剂作用机制示意图
5 苯并咪唑类杀菌剂 苯并咪唑类杀菌剂苯来特、多菌灵和甲基硫菌灵等主要是影响菌体内微管的形成而影响了细胞分裂 。 微管(microtubule)是广泛存在于植物(包括病菌)细胞中的纤维状结构,直径20 ~25nm,主要含有一种蛋白质,叫做微管蛋白(tububin)。它的功能是保护细胞形状、细胞运动和细胞内物质运输,和微丝、居间纤维共同形成了立体网络,称为“微梁系统”。细胞器和膜系统都由这个网络来支架。可以说,微管是细胞的骨骼。微管除了参与合成细胞壁和在鞭毛、纤毛运动中起作用外,最主要的是在细胞分裂中起作用——微管构成了减数分裂和有丝分裂纺锤体的纤维。 微管是由微管蛋白的亚单位靠疏水键的结合聚合成多聚体,最后再形成完整的微管。 在植物体内,苯来特和硫菌灵都转化成多菌灵起作用。近年的研究表明,这类杀菌剂的主要作用机制是由于多菌灵和微管蛋白的β亚单位相结合,阻止了微管的组装,从而破坏了纺锤体的形成,影响了细胞分裂。 苯并咪唑类杀菌剂作用机制示意图
是广泛存在于植物(包括病菌)细胞中的纤维状结构,直径20 ~25nm,主要含有一种蛋白质,叫做微管蛋白(tububin)。它的功能是保护细胞形状、细胞运动和细胞内物质运输,和微丝、居间纤维共同形成了立体网络,称为 微梁系统 。细胞器和膜系统都由这个网络来支架。可以说,微管是细胞的骨骼。微管除了参与合成细胞壁和在鞭毛、纤毛运动中起作用外,最主要的是在细胞分裂中起作用——微管构成了减数分裂和有丝分裂纺锤体的纤维。 微管是由微管蛋白的亚单位靠疏水键的结合聚合成多聚体,最后再形成完整的微管。 在植物体内,苯来特和硫菌灵都转化成多菌灵起作用。近年的研究表明,这类杀菌剂的主要作用机制是由于多菌灵和微管蛋白的β亚单位相结合,阻止了微管的组装,从而破坏了纺锤体的形成,影响了细胞分裂。 苯并咪唑类杀菌剂作用机制示意图.")
107
6 三环唑和丰谷隆的作用机制 主要干扰病菌寄生(调节病菌致病系统)的化合物、加强寄主防御作用的化合物。
6 三环唑和丰谷隆的作用机制 主要干扰病菌寄生(调节病菌致病系统)的化合物、加强寄主防御作用的化合物。 在离体条件下,很少或没有杀菌活性,但当应用到寄主植物上时可减少侵染或病害的严重度。
的化合物、加强寄主防御作用的化合物。 在离体条件下,很少或没有杀菌活性,但当应用到寄主植物上时可减少侵染或病害的严重度。")
108
活体表现 I 在寄主植物体内转变为具有毒性的化合物 II 影响真菌的致病能力 III影响寄主的抗病性
IV 可以干扰致病原因的关键因素(真菌毒素、酶的活性、产物)和寄主抗病性的诱导剂或抑制剂
和寄主抗病性的诱导剂或抑制剂.")
110
黑素合成途径

111
7 苯基吡咯杀菌剂的作用机制

112
PK-Ⅲ调节甘油合成(仿Pillonel和Meyer,1997)
关于苯基吡咯杀菌剂的作用机制已有较深入的研究。当以拌种咯处理Fusarium sulphureum,使其生长受到50%抑制时,菌丝中单糖的输送受到抑制;细胞内中性多元醇如甘油和甘露醇积累。除此之外,拌种咯对细胞核分裂、呼吸氧化、几丁质合成、甾醇合成、磷酯、核酸和蛋白合成均无影响。基于这些研究结果,Jesper等认为拌种咯的初始靶标是与输送相联系的葡萄糖磷酸化的酶,特别是己糖激酶(hexokinase),然而没有找到拌种咯对己糖激酶有直接影响的证据。最近,Pillonel和Meyer研究后认为蛋白激酶PK—Ⅲ才是苯基吡咯杀菌剂的初始靶标。PK-Ⅲ调节菌体内甘油的合成,其调节机制如图6-34所示。 拌种咯和蛋白激酶PK-Ⅲ结合,抑制了它的活性,使活化的调节蛋白不失活(不被磷酸化),从而导致甘油合成失控,细胞内渗透压加大,细胞发生肿胀而死亡。 PK-Ⅲ调节甘油合成(仿Pillonel和Meyer,1997)
,然而没有找到拌种咯对己糖激酶有直接影响的证据。最近,Pillonel和Meyer研究后认为蛋白激酶PK—Ⅲ才是苯基吡咯杀菌剂的初始靶标。PK-Ⅲ调节菌体内甘油的合成,其调节机制如图6-34所示。 拌种咯和蛋白激酶PK-Ⅲ结合,抑制了它的活性,使活化的调节蛋白不失活(不被磷酸化),从而导致甘油合成失控,细胞内渗透压加大,细胞发生肿胀而死亡。 PK-Ⅲ调节甘油合成(仿Pillonel和Meyer,1997)")
113
第四节 主要杀菌剂类型和品种

114
一、 保护性杀菌剂 (一) 主要特点 1. 不进入植物体内,只沉积在作物表面 2. 只有保护作用,对已侵入和施药后长出的新枝部分不能起作用 3. 杀菌谱广,适用范围宽,防病效果稳定 4. 不易产生抗药性。

115
(二) 使用技术要点 1. 在作物未染病或染病初期即施药,若病害严重时,则效果差; 2. 施药要尽量均匀周到,尤其在叶子背面也要求有杀菌剂沉积; 3. 往往要重复施药。
 使用技术要点 1. 在作物未染病或染病初期即施药,若病害严重时,则效果差; 2. 施药要尽量均匀周到,尤其在叶子背面也要求有杀菌剂沉积; 3. 往往要重复施药。")
116
(三) 主要品种 1. 波尔多液(Bordeaux mixture) (1) 配制:等量式 CuSO4∶生石灰 :水=1∶1∶100; 第一种方法:将CuSO4倒入石灰乳中 第二种方法:二者同时倒入第三个容器中 生石灰乳和CuSO4 天蓝色悬浊液 直接使用1%(柑桔) 1.5%(倍量)(葡萄)
 1.5%(倍量)(葡萄)")
117
(1)特点:粘着力强,不易被雨水冲刷,残效期7~14天
(2)应用:适用于果树、蔬菜等经济作物的叶部病害和苗期病害,如霜霉、疫病、炭疽、幼苗猝倒病等。但对白粉病效果差(细胞壁以角质层为主),长期使用,螨类猖獗 (3) 注意问题: ① 随配随用 ② 不能加水稀释 ③ 不能以铁质容器贮放 ④ 碱性,不宜和许多农药混用 ⑤ 桃、李、白菜、小麦等易药害。
应用:适用于果树、蔬菜等经济作物的叶部病害和苗期病害,如霜霉、疫病、炭疽、幼苗猝倒病等。但对白粉病效果差(细胞壁以角质层为主),长期使用,螨类猖獗. (3) 注意问题: ① 随配随用. ② 不能加水稀释. ③ 不能以铁质容器贮放. ④ 碱性,不宜和许多农药混用. ⑤ 桃、李、白菜、小麦等易药害。")
118
(4) 作用机制:有效成分为碱式硫酸铜 CuSO4·x Cu(OH)2·y Cu(OH)2·zH2O 游离出CU++,使病原菌细胞膜的蛋白质凝固,少量进入和酶或辅酶作用,破坏CoASH(辅酶A)。 另外,还有制剂碱式硫酸铜30%悬浮剂[Cu(OH)2] 3·CuSO4·5H2O
2] 3·CuSO4·5H2O.")
119
2. 石硫合剂(lime Sulphur) (1) 配制: 生石灰 + S粉 水 加热 CaSx(主要是五硫化钙)
早在公元前约1000年古希腊人Homer在史诗中就曾谈及硫磺的防病及其性质。到19世纪已逐渐有意识地利用硫磺,1802年就有了石硫合剂的记载,后来在1833年和1888年,进一步明确了石硫合剂对白粉病的防治效果, 1850年由于硫磺的大量使用,促使了喷粉法的创立 通常以 1份生石灰,2份S粉,10份H2O熬制而成

120
(2) 特点: 强碱性,不宜和其他化学农药混用,近期内不宜和波尔多液交替使用;不宜长期存放。 (3) 应用: 主要用于防治麦类、果树、蔬菜上的白粉病、锈病、叶斑病及红蜘蛛、介壳虫等。果园冬季清园或春天前,北方3度,南方1度,消灭越冬菌源及介壳虫、虫卵;防白粉病及叶斑病0.2~0.3度为宜 (4) 注意问题: 黄瓜、大豆、番茄、隶、桃、葡萄等易发生药害。
 注意问题: 黄瓜、大豆、番茄、隶、桃、葡萄等易发生药害。")
121
(5) 作用机制 有效成分是多硫化钙,特别是CaS5,析出细微的硫,产生少量的硫化氢 可能是影响三羧酸循环,从琥珀酸 延胡索酸需琥珀酸脱氢酶系的参与,该酶的辅酶是黄酶II,S制剂影响FAD的活性

122
3. 代森锰锌 1961年开发 (1)特点:既不溶于水,也不溶于有机溶剂。 70%、80%WP,40%SC
特点:既不溶于水,也不溶于有机溶剂。 70%、80%WP,40%SC")
123
(2) 应用: ① 防治果树、蔬菜、烟草、花生等作物上的霜霉病、疫病、炭疽、叶斑、梨黑星、烟赤星。400~600×
① 防治果树、蔬菜、烟草、花生等作物上的霜霉病、疫病、炭疽、叶斑、梨黑星、烟赤星。400~600× ② 棉花拌种100kg籽,0.5kg 70%WP, 可防苗期炭疽病;小麦腥黑穗0.1~0.5%。 ③ 伴药: 甲霜灵锰锌58%WP,其中48%为代森锰锌; 杀毒矾(恶霜灵锰锌)64% WP,其中56%为代森锰锌; 霜脲锰锌72% WP,其中64%为代森锰锌。 国际上对这一类杀菌剂的使用有争议,其在生产和贮存中分解,主要产物是乙撑硫脲,会引起甲状腺肿大,并称可能有致癌作用,因此在美国等地限制使用。
64% WP,其中56%为代森锰锌; 霜脲锰锌72% WP,其中64%为代森锰锌。 国际上对这一类杀菌剂的使用有争议,其在生产和贮存中分解,主要产物是乙撑硫脲,会引起甲状腺肿大,并称可能有致癌作用,因此在美国等地限制使用。")
124
(3) 作用机制 ① 破坏辅酶A:先水解 ② 抑制以Cu和Fe为辅基的酶的活性,机理是和Cu、Fe形成络合物
 作用机制 ① 破坏辅酶A:先水解 ② 抑制以Cu和Fe为辅基的酶的活性,机理是和Cu、Fe形成络合物")
125
4. 福美双 1951年发现 (1) 特点:不溶于水,与酸易分解 (2) 应用:推广作种子处理、土壤处理和叶丛喷雾 80WP%,拌种剂较多,
4. 福美双 1951年发现 (1) 特点:不溶于水,与酸易分解 (2) 应用:推广作种子处理、土壤处理和叶丛喷雾 80WP%,拌种剂较多, ai %拌种,可治蔬菜类、蚕豆等苗期、立枯和猝倒病; ai 0.15~0.25%拌种,可防治稻苗立枯、禾谷类黑穗病; ai 3.75~5.625kg/km2处理土壤(沟施或穴施),可防治蔬菜、烟草、甜菜等苗期病害。 500~1000倍喷雾,可防治蚕豆褐斑病、瓜霜霉病和炭疽病、梨黑星、苹果斑点病等。 (3) 作用机制:同于代森锰锌。
 特点:不溶于水,与酸易分解. (2) 应用:推广作种子处理、土壤处理和叶丛喷雾. 80WP%,拌种剂较多, ai 0.125%拌种,可治蔬菜类、蚕豆等苗期、立枯和猝倒病; ai 0.15~0.25%拌种,可防治稻苗立枯、禾谷类黑穗病; ai 3.75~5.625kg/km2处理土壤(沟施或穴施),可防治蔬菜、烟草、甜菜等苗期病害。 500~1000倍喷雾,可防治蚕豆褐斑病、瓜霜霉病和炭疽病、梨黑星、苹果斑点病等。 (3) 作用机制:同于代森锰锌。")
126
5. 百菌清 (1) 特点:对酸、碱、光较稳定,广谱; (2) 应用:叶斑、霜霉病、疫病、炭疽、白粉、果腐等。 75%WP,600~700×
5. 百菌清 百菌清(chlorothalonil) (1) 特点:对酸、碱、光较稳定,广谱; (2) 应用:叶斑、霜霉病、疫病、炭疽、白粉、果腐等。 75%WP,600~700× 2.5%烟剂,200~300g/亩,封闭4h,7~10d一次;
 (1) 特点:对酸、碱、光较稳定,广谱; (2) 应用:叶斑、霜霉病、疫病、炭疽、白粉、果腐等。 75%WP,600~700× 2.5%烟剂,200~300g/亩,封闭4h,7~10d一次;")
127
(3)作用机制 从3-磷酸甘油醛生成1,3-二磷酸甘油酸的过程中,需磷酸甘油醛脱氢酶,这是百菌清的作用靶标。百菌清与该酶上的半胱氨酸-SH结合,生成络合物,抑制酶的活性。
作用机制 从3-磷酸甘油醛生成1,3-二磷酸甘油酸的过程中,需磷酸甘油醛脱氢酶,这是百菌清的作用靶标。百菌清与该酶上的半胱氨酸-SH结合,生成络合物,抑制酶的活性。")
128
6. 二甲酰亚胺类杀菌剂 (1)乙烯菌核利(农利灵) ① 生物活性:触杀性杀菌剂,对核盘菌和灰霉孢特效,低毒
6. 二甲酰亚胺类杀菌剂 (1)乙烯菌核利(农利灵) ① 生物活性:触杀性杀菌剂,对核盘菌和灰霉孢特效,低毒 ② 使用方法:发病初期1.125~1.5kg制剂+500L水/hm2,7~10d,1次。可防治灰霉病和番茄早疫、晚疫,油菜菌核病、白菜黑斑病等
乙烯菌核利(农利灵) ① 生物活性:触杀性杀菌剂,对核盘菌和灰霉孢特效,低毒. ② 使用方法:发病初期1.125~1.5kg制剂+500L水/hm2,7~10d,1次。可防治灰霉病和番茄早疫、晚疫,油菜菌核病、白菜黑斑病等.")
129
① 生物活性:接触保护性杀菌剂,弱内吸。对核盘菌和灰霉孢特效,低毒;
(2)速克灵(腐霉利) 50% WP ① 生物活性:接触保护性杀菌剂,弱内吸。对核盘菌和灰霉孢特效,低毒; ② 使用方法:发病初期250~500mg /L,喷雾。可防治灰霉病和菌核病等。
速克灵(腐霉利) 50% WP. ① 生物活性:接触保护性杀菌剂,弱内吸。对核盘菌和灰霉孢特效,低毒; ② 使用方法:发病初期250~500mg /L,喷雾。可防治灰霉病和菌核病等。")
130
① 生物活性:接触保护性杀菌剂,弱内吸。对核盘菌和灰霉孢特效,低毒
(3)咪唑霉(朴海因、异菌脲) 50% WP ① 生物活性:接触保护性杀菌剂,弱内吸。对核盘菌和灰霉孢特效,低毒 ② 使用方法:发病初期0.75kg制剂/hm2,500~1000g/kg种子拌种。 ③ 作用机理:增大线粒体膜和内质网膜上的脂质过氧化反应
咪唑霉(朴海因、异菌脲) 50% WP. ① 生物活性:接触保护性杀菌剂,弱内吸。对核盘菌和灰霉孢特效,低毒. ② 使用方法:发病初期0.75kg制剂/hm2,500~1000g/kg种子拌种。 ③ 作用机理:增大线粒体膜和内质网膜上的脂质过氧化反应.")
131
二甲酰亚胺类杀菌剂作用机制与菌体磷脂过氧化反应有关,会增大线粒体膜和内质网膜上的脂质过氧化反应;并对细胞膜和细胞壁有影响,改变膜的渗透性和功能。
除此之外,异菌脲也能抑制蛋白激酶,控制一些细胞内的信号传递。 病原菌也易对其产生抗药性,为预防抗性菌株的产生,作物全生育期,扑海因的施用次数应控制在3次以内。 腐霉利使用过程中不能与碱性药剂和有机磷药剂混用。

132
二、 内吸性杀菌剂 施于植物某一部位的杀菌剂,被植物吸收后能在植物体内传导而运输到植物体的其他部位发生作用,具有这种性质的杀菌剂称为内吸性杀菌剂。

133
(一)苯并咪唑类
苯并咪唑类")
134
该类杀菌剂的特点 (1) 剂型都是40~70%WP或SC,1000~1500×使用; (2) 广谱杀菌剂; (3) 内吸性能好; (4) 作用机理是影响了细胞分裂,药剂阻碍了微管形成; (5) 作用位点单一,容易产生抗药性。
 剂型都是40~70%WP或SC,1000~1500×使用; (2) 广谱杀菌剂; (3) 内吸性能好; (4) 作用机理是影响了细胞分裂,药剂阻碍了微管形成; (5) 作用位点单一,容易产生抗药性。")
135
1. 多菌灵 (1)特点:难溶于水及一般有机溶剂; (2)防治对象:广谱内吸,内吸性能好,能被根系或叶片吸收传导。
50%、80% WP, 1000~1500×喷雾使用 (1)特点:难溶于水及一般有机溶剂; (2)防治对象:广谱内吸,内吸性能好,能被根系或叶片吸收传导。 麦类赤霉病、稻瘟病、纹枯病、谷类黑穗病、棉花苗病 、油菜菌核病、花卉病 、贮藏病害
特点:难溶于水及一般有机溶剂; (2)防治对象:广谱内吸,内吸性能好,能被根系或叶片吸收传导。 麦类赤霉病、稻瘟病、纹枯病、谷类黑穗病、棉花苗病 、油菜菌核病、花卉病 、贮藏病害.")
136
(3)应用: 赤霉病:扬花期50%,50%WP50~60g/亩; 纹枯病:分蘖及孕穗初期,初发病施一次,1000倍,可再施一次; 红薯黑疤:移栽前稀释1000倍,浸薯或苗茎部分5min;贮存薯块2000倍喷雾 棉花苗期:50%WP按1%种子量拌种。 (4)作用机理:影响微管形成,干扰细胞分裂。
作用机理:影响微管形成,干扰细胞分裂。")
137
2. 甲基硫菌灵 (1) 特点:难溶于水,易溶于有机溶剂; (2)防治对象:麦类赤霉病、稻瘟病、油菜菌核病、 花生叶斑病、红薯黑疤病
2. 甲基硫菌灵 50% 、 70% WP (1) 特点:难溶于水,易溶于有机溶剂; (2)防治对象:麦类赤霉病、稻瘟病、油菜菌核病、 花生叶斑病、红薯黑疤病 (3) 应用 防治苹果、梨黑星、炭疽、葡萄黑疤、瓜类蔓枯病、叶霉病,70%WP,1000~1500×,5~7天一次 烟草白粉、白星、炭疽、赤星、花卉病1000× 小麦白粉、赤霉、颖枯、雪霉等 水果贮存:300~400×浸果
 特点:难溶于水,易溶于有机溶剂; (2)防治对象:麦类赤霉病、稻瘟病、油菜菌核病、 花生叶斑病、红薯黑疤病. (3) 应用. 防治苹果、梨黑星、炭疽、葡萄黑疤、瓜类蔓枯病、叶霉病,70%WP,1000~1500×,5~7天一次. 烟草白粉、白星、炭疽、赤星、花卉病1000× 小麦白粉、赤霉、颖枯、雪霉等. 水果贮存:300~400×浸果.")
138
(二)苯基酰胺类 酰苯胺类杀菌剂是1973年瑞士汽巴- 嘉基(Ciba - Geigy) 公司筛选除草剂时意外发现的一种优良的内吸性杀菌剂,其对卵菌特效。甲霜灵作为酰苯胺类杀菌剂的第一个产品, 1978年被商品化,随后系列产品得到开发。该类药剂的共同特点是低毒,选择性强,只对卵菌有效,施用后对植物有保护及治疗作用,持效期长,可达24周。代表性品种有苯霜灵、呋霜灵、甲霜灵、精甲霜灵、恶霜灵和甲呋酰胺等。大多品种在植物体内可向顶传导,甲呋酰胺则具有向顶、向基及侧向传导的活性。
 公司筛选除草剂时意外发现的一种优良的内吸性杀菌剂,其对卵菌特效。甲霜灵作为酰苯胺类杀菌剂的第一个产品, 1978年被商品化,随后系列产品得到开发。该类药剂的共同特点是低毒,选择性强,只对卵菌有效,施用后对植物有保护及治疗作用,持效期长,可达24周。代表性品种有苯霜灵、呋霜灵、甲霜灵、精甲霜灵、恶霜灵和甲呋酰胺等。大多品种在植物体内可向顶传导,甲呋酰胺则具有向顶、向基及侧向传导的活性。")
139
该类杀菌剂的特点 (1)主要用于各种疫病、霜霉病; (2)单作用位点杀菌剂,容易产生抗药性;
(3)作用机理:抑制rRNA聚合酶的活性,从而抑制了 RNA的合成,具体机理不很清楚; (4)常加工成混剂 如甲霜锰锌(10%甲+48%代) 恶霜锰锌(8%恶+56%代)
作用机理:抑制rRNA聚合酶的活性,从而抑制了 RNA的合成,具体机理不很清楚; (4)常加工成混剂. 如甲霜锰锌(10%甲+48%代) 恶霜锰锌(8%恶+56%代)")
140
甲霜灵 (Ridomil,瑞毒霉) (1) 特点:性质稳定,保护、治疗兼备; (2) 对象:霜霉、疫霉、腐霉引起的病害;
(1) 特点:性质稳定,保护、治疗兼备; (2) 对象:霜霉、疫霉、腐霉引起的病害; (3) 应用:25%WP,400~500×,5~7d一次 烟草黑茎病:移栽至7d开始,每10d一次,连续3次; 辣椒疫霉:初见发病400倍喷全株,7d一次, 500倍喷全株,200ml/株 精甲霜灵是原瑞士诺华农化有限公司开发面市的第一个旋光活性杀菌剂, 是外消旋体甲霜灵中的R体。
 特点:性质稳定,保护、治疗兼备; (2) 对象:霜霉、疫霉、腐霉引起的病害; (3) 应用:25%WP,400~500×,5~7d一次. 烟草黑茎病:移栽至7d开始,每10d一次,连续3次; 辣椒疫霉:初见发病400倍喷全株,7d一次, 500倍喷全株,200ml/株. 精甲霜灵是原瑞士诺华农化有限公司开发面市的第一个旋光活性杀菌剂, 是外消旋体甲霜灵中的R体。")
141
(三) 甾醇抑制剂 三唑类、吡唑类、嘧啶类、咪唑类、哌嗪类、吗啉类等 该类药剂的特点 (1) 有强的向定性传导和明显的熏蒸作用;
(2)杀菌谱广,除鞭毛菌、病毒外,对子囊菌、担子菌、半知菌都有一定效果; (3) 高效、使用量低,拌种0.3~0.4g/kg; (4) 药效期长,一般3~6周;
杀菌谱广,除鞭毛菌、病毒外,对子囊菌、担子菌、半知菌都有一定效果; (3) 高效、使用量低,拌种0.3~0.4g/kg; (4) 药效期长,一般3~6周;")
142
氟硅唑(flusilazole) 三唑酮(triadimefon) 烯唑醇(dinicoragole) 戊唑醇 (tebuconazole)
腈菌唑(myclobutanil) 氟硅唑(flusilazole) 苯醚甲环唑(difenoconazole) 三唑类杀菌剂是继苯并咪唑类之后,迄今为止发展最快,品种最多的一类杀菌剂,其大多品种的化学活性基团为1,2,4-三唑,也是麦角甾醇合成抑制剂中最重要的一类化合物,具有高效、广谱、持效期长等特点。该类杀菌剂的研发应该追溯到20 世纪 60 年代末, 拜耳公司和比利时詹森公司首先报道了 1- 取代唑类衍生物的杀菌活性,70 年代初,唑类化合物的高效杀菌活性逐渐引起国际农药界的高度重视。1974年,拜耳公司成功地开发出三唑酮并进行商品化,标志着三唑类杀菌剂时代的到来。 早期的三唑类杀菌剂三唑环上的支链以脂肪链为主,脂肪链上的羟基或羰基部分变成环氧基团,主要防治对象以白粉病,锈病和黑星病为主。 为了扩大防治谱,减少药剂施用量,减缓药害的发生。在上述杀菌剂研究基础上,保留了活性基团三唑环的同时,在结构中引入氟原子,硅原子和硫原子,也成为三唑类杀菌剂开发的另一个特点。近年来一些品种还增加了杂环结构。由于结构的变化,使新开发的杀菌剂不但对白粉病和锈病等具有活性,而且对灰霉病,眼纹病等也有效,由此形成了第二代三唑类杀菌剂。 抑霉唑(imazalil) 咪鲜安(prochlprag) n=10,11,12(60%~70%)或13 十三吗啉(tridemorph) 咪唑类
 氟硅唑(flusilazole) 苯醚甲环唑(difenoconazole) 三唑类杀菌剂是继苯并咪唑类之后,迄今为止发展最快,品种最多的一类杀菌剂,其大多品种的化学活性基团为1,2,4-三唑,也是麦角甾醇合成抑制剂中最重要的一类化合物,具有高效、广谱、持效期长等特点。该类杀菌剂的研发应该追溯到20 世纪 60 年代末, 拜耳公司和比利时詹森公司首先报道了 1- 取代唑类衍生物的杀菌活性,70 年代初,唑类化合物的高效杀菌活性逐渐引起国际农药界的高度重视。1974年,拜耳公司成功地开发出三唑酮并进行商品化,标志着三唑类杀菌剂时代的到来。 早期的三唑类杀菌剂三唑环上的支链以脂肪链为主,脂肪链上的羟基或羰基部分变成环氧基团,主要防治对象以白粉病,锈病和黑星病为主。 为了扩大防治谱,减少药剂施用量,减缓药害的发生。在上述杀菌剂研究基础上,保留了活性基团三唑环的同时,在结构中引入氟原子,硅原子和硫原子,也成为三唑类杀菌剂开发的另一个特点。近年来一些品种还增加了杂环结构。由于结构的变化,使新开发的杀菌剂不但对白粉病和锈病等具有活性,而且对灰霉病,眼纹病等也有效,由此形成了第二代三唑类杀菌剂。 抑霉唑(imazalil) 咪鲜安(prochlprag) n=10,11,12(60%~70%)或13. 十三吗啉(tridemorph) 咪唑类.")
143
粉锈宁:1974,Bayer公司 特点: (1)三唑类药剂对双子叶作物有明显的抑制作用。 (2)强内吸、兼有治疗和保护作用。
(3)对各种锈病、白粉病特效,对黑穗病、玉米、高梁丝黑穗病也有较好的效果。 粉锈宁:1974,Bayer公司 三唑酮(triadimefon)
对各种锈病、白粉病特效,对黑穗病、玉米、高梁丝黑穗病也有较好的效果。 粉锈宁:1974,Bayer公司. 三唑酮(triadimefon)")
144
(1) 特点:难溶于水,易溶于有机溶剂;性质稳定,持效期长;
(2) 对象:锈病、白粉病特效,对黑穗病高效; (3) 施用技术: 条锈:20%EC,1500×喷施发病中心;穗期再喷全天一次。拌种100g, 20%EC, 0.15kg,0.03% 白粉:2000×; 玉米、高梁丝黑穗:100kg,拌种玉米,50%WP,100-800g;高粱, g (4) 注意事项:药害,尤其是种衣剂。
 对象:锈病、白粉病特效,对黑穗病高效; (3) 施用技术: 条锈:20%EC,1500×喷施发病中心;穗期再喷全天一次。拌种100g, 20%EC, 0.15kg,0.03% 白粉:2000×; 玉米、高梁丝黑穗:100kg,拌种玉米,50%WP,100-800g;高粱, g (4) 注意事项:药害,尤其是种衣剂。")
145
(5) 作用机制:麦角甾醇抑制剂。 麦角甾醇,是细胞膜的主要组分。影响膜的通透性功能;此外,它也是一种激素前体,在有性、无性生殖过程中起到主要作用。 麦角甾醇的合成是由甲瓦龙酸开始,其中有一步反应是14位C的脱甲基反应。这步反应是MFO的细胞色素P450来完成的。三唑类杀菌剂和P450中心的Fe结合,抑制了该酶的活性,从而抑制了14C脱甲基,阻止了麦角甾醇的合成。

146
(四) 羧酸氨基化合物类杀菌剂 烯酰吗啉 (dimethomorph) 氟吗啉(flumorph) 双炔酰菌胺(mandipropamid)
(四) 羧酸氨基化合物类杀菌剂 烯酰吗啉 (dimethomorph) 氟吗啉(flumorph) 双炔酰菌胺(mandipropamid) 2007年先正达公司,第一个扁桃酰胺类 苯乙烯酸类 烯酰吗啉是由巴斯夫公司在上世纪80年代末开发并投入市场的杀菌剂,其对植物卵菌病害具有优异的保护和治疗作用,且与甲霜灵等苯基酰胺类杀菌剂无交互抗药性。投入市场便受到世界各国使用者的亲睐,逐步成为继甲霜灵之后用于卵菌植物病害防治的主要杀菌剂。1994年,我国沈阳化工研究院以烯酰吗啉为先导,以氟原子替代了烯酰吗啉苯环上的氯原子开发出氟吗啉。 异丙菌胺(iprovalicarb) valiphenal 2008年意大利Isagro公司,防治霜霉病 1998年拜耳公司, 第一个缬氨酸氨基甲酸盐类
 羧酸氨基化合物类杀菌剂. 烯酰吗啉. (dimethomorph) 氟吗啉(flumorph) 双炔酰菌胺(mandipropamid) 2007年先正达公司,第一个扁桃酰胺类. 苯乙烯酸类. 烯酰吗啉是由巴斯夫公司在上世纪80年代末开发并投入市场的杀菌剂,其对植物卵菌病害具有优异的保护和治疗作用,且与甲霜灵等苯基酰胺类杀菌剂无交互抗药性。投入市场便受到世界各国使用者的亲睐,逐步成为继甲霜灵之后用于卵菌植物病害防治的主要杀菌剂。1994年,我国沈阳化工研究院以烯酰吗啉为先导,以氟原子替代了烯酰吗啉苯环上的氯原子开发出氟吗啉。 异丙菌胺(iprovalicarb) valiphenal. 2008年意大利Isagro公司,防治霜霉病. 1998年拜耳公司, 第一个缬氨酸氨基甲酸盐类.")
147
特点 结构:根据化学结构的差异将CAA类杀菌剂分为三类,即苯乙烯酸类、缬氨酸氨基甲酸盐类和扁桃酸氨基化合物类。
生物活性:对植物卵菌(如霜霉属和疫霉属病菌)病害具有优异的保护和治疗作用,且与甲霜灵等苯基酰胺类杀菌剂无交互抗药性。有优异的保护、治疗和铲除效果,但均没有明显的抑制效果。 对游动孢子的释放和游动没有抑制作用,对具有细胞壁的所有阶段均具有明显的抑制作用,而对不具有细胞壁的游动孢子阶段没有影响的特性。推测可能干扰病菌细胞壁的形成,但药剂的具体靶标位点仍然不清楚。
病害具有优异的保护和治疗作用,且与甲霜灵等苯基酰胺类杀菌剂无交互抗药性。有优异的保护、治疗和铲除效果,但均没有明显的抑制效果。 对游动孢子的释放和游动没有抑制作用,对具有细胞壁的所有阶段均具有明显的抑制作用,而对不具有细胞壁的游动孢子阶段没有影响的特性。推测可能干扰病菌细胞壁的形成,但药剂的具体靶标位点仍然不清楚。")
148
(五) 甲氧丙烯酸类 (嗜球果伞素类或QoI杀菌剂 ) ICI A5504;嘧菌酯(azoxystrobin) 小麦赤霉、葡萄霜霉、白粉
20世纪60年代,捷克科学家Musilek V等人在一种蘑菇(Oudemansiella mucida)中首次发现了strobilurin A。 (嗜球果伞素类或QoI杀菌剂 ) 1992年 ICI A5504;嘧菌酯(azoxystrobin) 小麦赤霉、葡萄霜霉、白粉 醚菌酯(BASF 490F) 苹果黑星、白粉、稻瘟
中首次发现了strobilurin A。 (嗜球果伞素类或QoI杀菌剂 ) 1992年. ICI A5504;嘧菌酯(azoxystrobin) 小麦赤霉、葡萄霜霉、白粉. 醚菌酯(BASF 490F) 苹果黑星、白粉、稻瘟.")
149
肟菌酯 (trifloxystrobin) 啶氧菌酯 (picoxystrobin) 苯氧菌胺 (metominostrobin) 唑菌胺酯
2001年,拜耳,小麦叶枯病、网斑病和云纹病 1999年,拜耳 1993年,日本盐野义公司,水稻稻瘟病 唑菌胺酯 (pyraclostrobin) 烯肟菌酯 (enestroburin) 苯醚菌酯 (ZJ 0712) 2002年,巴斯夫 沈阳化工研究院 浙江化工研究院
 烯肟菌酯. (enestroburin) 苯醚菌酯. (ZJ 0712) 2002年,巴斯夫. 沈阳化工研究院. 浙江化工研究院.")
150
特点 1 结构 甲氧基丙烯酸酯类,有嘧菌酯、啶氧菌酯、烯肟菌酯、苯醚菌酯、UBF-307和嘧螨酯(杀螨剂) 。
甲氧基氨基甲酸酯类,有唑菌胺酯。 肟基乙酸酯类,有醚菌酯和肟菌酯。 肟基乙酰胺类,有苯氧菌胺、醚菌胺、肟醚菌胺和烯肟菌胺。 唑烷二酮类,有恶唑菌酮。 咪唑啉酮类,有咪唑菌酮。 肟基二恶嗪类,有氟嘧菌酯。 侧链 活性基团团 桥 strobilurins杀菌剂地活性结构(以嘧菌酯为例)
")
151
2 具有广谱、高效、安全,环境友好等特点,能同时有效地防治子囊菌门、担子菌门、半知菌门真菌和卵菌引起的病害。
3 与目前使用的杀菌剂不存在交互抗性。但药剂作用位点专单一,1996年在欧洲开始用于小麦白粉病的防治,1998年就在德国北部三个地区检测到抗性个体,抗性倍数大于500。具有高抗药性风险。 4 属于线粒体呼吸抑制剂,药剂与线粒体电子传递链中复合物Ⅲ(Cytbc1复合物)结合,阻断电子从Cytbc1复合物流向Cytc,阻止ATP合成。因为该类杀菌剂是与线粒体内膜外壁Qo位点(CoQ的氧化位点)的Cytb低势能血红素结合的抑制剂,也称为Qo位点抑制剂。
结合,阻断电子从Cytbc1复合物流向Cytc,阻止ATP合成。因为该类杀菌剂是与线粒体内膜外壁Qo位点(CoQ的氧化位点)的Cytb低势能血红素结合的抑制剂,也称为Qo位点抑制剂。")
152
醚菌酯 Stroby 50%翠贝干悬浮剂 黄瓜白粉病 g/亩( 倍) 苹果黑星病: 倍
 苹果黑星病: 倍")
153
(五) 其它类型杀菌剂 1. 黑色素生物合成抑制剂 强内吸 持效期 7-10周 浸种、浸根、喷粉 三环唑(tricyclagole)
(五) 其它类型杀菌剂 1. 黑色素生物合成抑制剂 三环唑(tricyclagole) 咯嗪酮(pyroguilon) 四氯苯酞(fthaliole) 强内吸 持效期 7-10周 浸种、浸根、喷粉
 其它类型杀菌剂. 1. 黑色素生物合成抑制剂. 三环唑(tricyclagole) 咯嗪酮(pyroguilon) 四氯苯酞(fthaliole) 强内吸. 持效期 7-10周. 浸种、浸根、喷粉.")
154
三环唑 (1)易溶于水(1.6g/L)与极性有机溶剂,对光、热稳定;
内吸性保护性杀菌剂,可迅速被根、茎、叶吸收,抗雨水冲刷,对稻瘟病由特效。 (2)使用技术:叶瘟:初发,由急性病斑时,10-20g ai/亩喷雾。穗瘟:孕穗至破口前60-80g/75kg水。喷一次即可,若天气不利,可10d再来一次。平撒毒土,20-25ai,20%wp,100g。
使用技术:叶瘟:初发,由急性病斑时,10-20g ai/亩喷雾。穗瘟:孕穗至破口前60-80g/75kg水。喷一次即可,若天气不利,可10d再来一次。平撒毒土,20-25ai,20%wp,100g。")
155
( 3) 作用机制 黑素(Velanin)是一类酚类物质,为1,8-二羟基萘聚合物,在致病性中具重要作用。稻瘟孢子在植物侵染前现形成一种附属孢的结构,然后附属孢穿透寄主表皮细胞而产生侵染。在穿透表皮前,附着孢的壁黑化(产生黑素),黑化提供刚度和结构,在穿透中起支持和集中机械力的作用。尤其附着孢下壁变硬,对寄主角质层产生膨胀力,有助于切断角质层。
是一类酚类物质,为1,8-二羟基萘聚合物,在致病性中具重要作用。稻瘟孢子在植物侵染前现形成一种附属孢的结构,然后附属孢穿透寄主表皮细胞而产生侵染。在穿透表皮前,附着孢的壁黑化(产生黑素),黑化提供刚度和结构,在穿透中起支持和集中机械力的作用。尤其附着孢下壁变硬,对寄主角质层产生膨胀力,有助于切断角质层。")
156
2. 有机磷 (1)敌瘟散 主要用于防治稻瘟病、兼治胡麻叶斑病、纹枯病等 (2)异稻瘟净 特点: 内吸有机磷杀菌剂,防/治兼备
2. 有机磷 (1)敌瘟散 主要用于防治稻瘟病、兼治胡麻叶斑病、纹枯病等 (2)异稻瘟净 特点: 内吸有机磷杀菌剂,防/治兼备 主要防稻瘟,也有杀叶螨、飞虱作用 应用:40%EC,200ml/亩喷雾 作用机制:抑制细胞膜上卵磷脂(磷脂酰胆碱)合成,从而破坏了细胞膜的结构。 有机磷杀菌剂主要品种有1965年日本Ihara公司和1968年日本组合化学公司分别开发的稻瘟净和异稻瘟净,1966年由道化学(Dow Chemical)公司开发的灭菌磷,1968年由拜耳(Bayer)公司推广的敌瘟磷(克瘟散)以及70年代由Rhone-Poulenc Agrochimie 公司开发的三乙膦酸铝(乙磷铝),80年代日本住友化学公司和英国FBC公司开发的甲基立枯磷、定菌磷等十几个个品种 硫赶磷酸酯类
敌瘟散. 主要用于防治稻瘟病、兼治胡麻叶斑病、纹枯病等. (2)异稻瘟净. 特点: 内吸有机磷杀菌剂,防/治兼备. 主要防稻瘟,也有杀叶螨、飞虱作用. 应用:40%EC,200ml/亩喷雾. 作用机制:抑制细胞膜上卵磷脂(磷脂酰胆碱)合成,从而破坏了细胞膜的结构。 有机磷杀菌剂主要品种有1965年日本Ihara公司和1968年日本组合化学公司分别开发的稻瘟净和异稻瘟净,1966年由道化学(Dow Chemical)公司开发的灭菌磷,1968年由拜耳(Bayer)公司推广的敌瘟磷(克瘟散)以及70年代由Rhone-Poulenc Agrochimie 公司开发的三乙膦酸铝(乙磷铝),80年代日本住友化学公司和英国FBC公司开发的甲基立枯磷、定菌磷等十几个个品种. 硫赶磷酸酯类.")
157
作用机制则是抑制DNA、RNA和蛋白质的生物合成,以及氧的吸收。对稻瘟病菌抑菌活性较差,而分别对白粉病菌和立枯病菌具有较高生物活性。
(3)定菌磷和甲基立枯磷 作用机制则是抑制DNA、RNA和蛋白质的生物合成,以及氧的吸收。对稻瘟病菌抑菌活性较差,而分别对白粉病菌和立枯病菌具有较高生物活性。 硫逐磷酸酯类 甲基立枯磷(tolclofos—methy1) 土壤处理、拌种、浸种或叶面喷雾 (4)威菌磷 对白粉病菌的抑制作用主要表现在影响吸器的形成过程并作用于寄主的代谢,扰乱寄主和病原间的互作。 第一个磷酰胺类的内吸性杀菌剂,它对白粉病有特效,并能兼治蚜虫和螨类。 磷酰胺类 威菌磷(triamiphos)
定菌磷和甲基立枯磷. 作用机制则是抑制DNA、RNA和蛋白质的生物合成,以及氧的吸收。对稻瘟病菌抑菌活性较差,而分别对白粉病菌和立枯病菌具有较高生物活性。 硫逐磷酸酯类. 甲基立枯磷(tolclofos—methy1) 土壤处理、拌种、浸种或叶面喷雾. (4)威菌磷. 对白粉病菌的抑制作用主要表现在影响吸器的形成过程并作用于寄主的代谢,扰乱寄主和病原间的互作。 第一个磷酰胺类的内吸性杀菌剂,它对白粉病有特效,并能兼治蚜虫和螨类。 磷酰胺类. 威菌磷(triamiphos)")
158
(5) 三乙磷酸铝(疫霉灵) 有争议:(1)诱导抗病性 (2)直接对病原菌作用 防治卵菌纲病害 产生酚类和黄酮类抗病物质
唯一的金属有机磷类杀菌剂 第一个可以防治卵菌病害的内吸性杀菌剂品种, 第一个具有向基和向顶双向传导的内吸性杀菌剂。

159
3.杀菌农用抗生素 目前,我国已登记杀菌农用抗生素13种,其中井冈霉素、农抗120、多抗霉素等8种已获正式登记,中生菌素、宁南霉素等5种获临时登记,成为我国杀菌剂市场的重要组成部分。 抗生素的研发始于1877年巴斯德关于微生物拮抗作用的发现。 1929年Fleming发现青霉素、1944年Wakesman 发现链霉素开创了抗生素的黄金时代。 重要的抗生素类型(四环素类、头孢类、氨基糖甙类及大环内酯类)均是在20世纪40~50年代开发的。
均是在20世纪40~50年代开发的。")
160
(1) 灭瘟素-S 1955年日本科学家从一株链霉菌Streptomyces griseochomagenes 2A-32J发酵液中发现灭瘟素A、B、C,其中灭瘟素A对稻瘟病有良好的防治效果,但在光照下不稳定,未能大面积推广应用。 1959年Misato等从发酵液中分离出光稳定的灭瘟素-S,但因其药害严重不增产,后改造成为灭瘟素苄氨基苯磺酸基后才减了药害,开发出第一个商品化的杀菌农用抗生素灭瘟素-S。 中国科学院微生物研究所亦于1959年从广东省花县土壤中分离得到灭瘟素产生菌,开始了灭瘟素-S的商品化生产。 主要用于防治水稻稻瘟病,具有保护兼治疗作用,可抑制病菌孢子萌发、菌丝生长及孢子形成。灭瘟素-S和稻瘟病菌原核细胞的50S核糖结合,阻断了肽基的转运和蛋白链的延长,从而抑制了蛋白质的合成。 对人畜毒性较大(大鼠急性经口LD50为55.9~56.8mg/Kg)
")
161
(2) 春雷霉素 1963年Umegawa等从春日链霉菌Streptomyces kasugaensis分离出一种抗生素,即春日霉素(kasugamycin)。 1964年中国科学院微生物研究所从江西泰和县土壤中分离的小金色放线菌Actinomyces microanueus亦产生和春日霉素相同的抗生素,命名为春雷霉素(kasugamycin)。 春雷霉素为水溶性内吸性抗生素,具有保护作用和治疗作用,对稻瘟病、甜菜叶斑病、番茄叶霉病等真菌病害及黄瓜角斑病、苹果炭疽病等细菌性病害都有良好的防治效果。
。 春雷霉素为水溶性内吸性抗生素,具有保护作用和治疗作用,对稻瘟病、甜菜叶斑病、番茄叶霉病等真菌病害及黄瓜角斑病、苹果炭疽病等细菌性病害都有良好的防治效果。")
162
(3) 多抗霉素 1965年Suzuki及其同事从Streptomyces cacaoi var.asoensis发酵产物中发现多抗霉素(polyoxin),1967年中国科学院微生物研究所亦分离得到此菌。 多抗霉素是水溶性抗生素,是具有相同肽嘧啶核苷酸骨架的14种同系物的混合物,主要为多抗霉素B和多抗霉素D。 多抗霉素是一种水溶性内吸性杀菌抗生素,具有广谱抗菌性,对多种真菌病害,特别是对由交链孢菌引起的梨黑斑病和苹果斑点落叶病及由灰葡萄孢引起的灰霉病有良好的防治效果。 多抗霉素通过干扰病原菌细胞壁(几丁质)的合成,阻止病菌产生孢子和病斑扩大。
的合成,阻止病菌产生孢子和病斑扩大。")
163
(4) 井岗霉素(有效霉素) 1972年Horii等报道的有效霉素(Validamycin)是日本科学家为水稻三大病害之一的水稻纹枯病找到一种特效抗生素。 主要为有效霉素A(VM-A),水溶性内吸杀菌剂,很容易在纹枯病菌丝中传到,引起菌丝异常分支,从而抑制其生长。 井冈霉素对立枯丝核菌并无直接杀菌作用,但井冈霉素是立枯丝核菌AG-1海藻糖酸的抑制剂,能有效地阻止纹枯病菌从菌丝的基部向顶端输送营养(如葡萄糖),从而抑制菌丝的生长和发育。 1个海藻糖 对小麦、水稻纹枯病特效 海藻糖酶 2个葡萄糖
,从而抑制菌丝的生长和发育。 1个海藻糖. 对小麦、水稻纹枯病特效. 海藻糖酶. 2个葡萄糖.")
164
(5) 米多霉素 (mildiomycin ) Harada和Kishi1978年从 Streptoverticillium rimofaciens B-98891菌株中开发出米多霉素(mildiomycin)。米多霉素不仅具有良好的保护作用,而且具有很好的治疗效果,并具有渗透性,对多种真菌病害有效,尤对白粉病高效,田间喷洒仅为5~10g/ha。
。米多霉素不仅具有良好的保护作用,而且具有很好的治疗效果,并具有渗透性,对多种真菌病害有效,尤对白粉病高效,田间喷洒仅为5~10g/ha。")
165
(6) 宁南霉素(ningnanmycin)
1997年中国科学院成都生物研究所从西昌地区分离的链霉菌Streptomyces noursei var. xichangensis发酵液中分离鉴定出抗生素宁南霉素(ningnanmycin)。 宁南霉素为水溶性胞嘧啶核苷肽型抗生素,就分子结构而言,是kangaki等1962年报道的抗肿瘤药物谷氏菌素(gougerotin)的立体,其差别在于谷氏菌素的丝氨酸为D型,而宁南霉素的丝氨酸为L型。 宁南霉素保护与治疗兼备,除对香蕉尖顶病、麦类白粉病、大豆枯腐病及某些病有较好的防治效果外,还对烟草、番茄、辣椒及瓜类病毒病有一定防治效果。
。 宁南霉素为水溶性胞嘧啶核苷肽型抗生素,就分子结构而言,是kangaki等1962年报道的抗肿瘤药物谷氏菌素(gougerotin)的立体,其差别在于谷氏菌素的丝氨酸为D型,而宁南霉素的丝氨酸为L型。 宁南霉素保护与治疗兼备,除对香蕉尖顶病、麦类白粉病、大豆枯腐病及某些病有较好的防治效果外,还对烟草、番茄、辣椒及瓜类病毒病有一定防治效果。")
166
(7) 其他农用抗生素
 其他农用抗生素")
167
杀菌农用抗生素的特点 毒性:杀菌农用抗生素对哺乳动物的毒性多数为低毒,使用中对人畜比较安全。
对环境和生态影响:农用抗生素对环境的压力较小,对非靶标生物安全。一般只含有碳、氢、氧、氮4元素,在环境中易于降解,本身又多是低毒化合物,因而不存在严重的环境污染或农产品残留性问题。大多抗生素属水溶性化合物,一般都加工成水剂、可溶性粉剂,避免了乳油等剂型中有机溶剂对环境的影响。对许多农田非靶标生物,特别对鸟类、兽类、蚯蚓、鱼类及害虫天敌等比较安全,大多数品种对农作物及有益微生物的影响亦较小。 生物活性和应用:大多产品具有内吸传导活性,可被植物组织吸收并在植物体内运转,因而兼有内吸性,保护和治疗作用。可采用根部土壤处理或叶部喷施药剂等多种方式进行药剂处理,这对于防治一些维管束病害、根部病害十分有利。但杀菌农用抗生素的作用位点单一,而且大多数是蛋白质合成抑制剂,因此靶标病原菌容易对其产生抗药性。

168
5. 植物保护活化剂 (1)特点:用药一段时间以后,植物的保卫反应才能增强; (2)使用方法:
苯并噻二唑(BTH)类 Bion 气巴-嘉基在开发磺酰脲类除草剂时发现,苯并噻二唑-7-羧酸甲酯有SAR活性. 活化酯(acibenzolar,Bion, 先正达) 50%和63%水分散颗粒剂 (1)特点:用药一段时间以后,植物的保卫反应才能增强; (2)使用方法: 小麦:2叶期,ai .2g/亩,对小麦白粉病持效期6-9周。 水稻:秧田(育种),0.01g/m2,可防治稻瘟病。 烟草:14d,1次,ai .1g/亩,霜霉病。 蔬菜:10-20d, 1次,ai g/亩,霜霉、黄斑、细菌
类 Bion. 气巴-嘉基在开发磺酰脲类除草剂时发现,苯并噻二唑-7-羧酸甲酯有SAR活性. 活化酯(acibenzolar,Bion, 先正达) 50%和63%水分散颗粒剂. (1)特点:用药一段时间以后,植物的保卫反应才能增强; (2)使用方法: 小麦:2叶期,ai .2g/亩,对小麦白粉病持效期6-9周。 水稻:秧田(育种),0.01g/m2,可防治稻瘟病。 烟草:14d,1次,ai .1g/亩,霜霉病。 蔬菜:10-20d, 1次,ai g/亩,霜霉、黄斑、细菌.")
169
烯丙异噻唑类 日本制果公司开发了赛瘟唑(probenarole) 烯丙异噻唑(PBZ)
在育秧或移栽前施于水稻植株上后,对于水稻稻瘟病具有显著的防治效果,对水稻白叶枯病也有较好的防效。 控制稻瘟40 天以上。

170
寡糖(Oligosaccharide) 真菌病原Pmg (Phytophthora megasperma f.sp.glycinea)的菌丝体壁上分离到带分枝的β-1,3和β-1,6连接组成的葡聚七糖是首次被发现的寡糖激活剂,它能明显诱导大豆子叶中植保素积累。还原端少一个葡萄糖单元的六糖类也有类似于七糖的功能。 还原端少一个葡萄糖单元的六糖类也有类似于七糖的功能。葡萄寡糖(六糖)广谱、高效,10ng寡糖激活剂应用于1g植物组织即能产生足够的植保素。在活体条件下能抑制多种病原微生物的侵染,对于番茄叶霉病、小麦白粉和西瓜枯萎病均具有良好的防治效果,并表现为提高作物抗病能力,延缓和减轻病害发生,提高农作物产量和品质等综合效能。 寡糖可诱导植保素、蛋白酶抑制剂的合成以及与病程相关PR蛋白的积累,并可使苯丙氨酸解氨酶、过氧化物酶多种防御酶系活性增强。 葡萄寡糖(六糖)
的菌丝体壁上分离到带分枝的β-1,3和β-1,6连接组成的葡聚七糖是首次被发现的寡糖激活剂,它能明显诱导大豆子叶中植保素积累。还原端少一个葡萄糖单元的六糖类也有类似于七糖的功能。 还原端少一个葡萄糖单元的六糖类也有类似于七糖的功能。葡萄寡糖(六糖)广谱、高效,10ng寡糖激活剂应用于1g植物组织即能产生足够的植保素。在活体条件下能抑制多种病原微生物的侵染,对于番茄叶霉病、小麦白粉和西瓜枯萎病均具有良好的防治效果,并表现为提高作物抗病能力,延缓和减轻病害发生,提高农作物产量和品质等综合效能。 寡糖可诱导植保素、蛋白酶抑制剂的合成以及与病程相关PR蛋白的积累,并可使苯丙氨酸解氨酶、过氧化物酶多种防御酶系活性增强。 葡萄寡糖(六糖)")
171
思考题 1、简述杀菌剂防治植物病害的主要原理。 2、什么是植物诱导抗病激活剂?请列举2个代表性的品种。
3、简述三唑类杀菌剂的作用机理及其特点;列举2个有代表性的品种。 4、简述苯并咪唑类杀菌剂的作用机制及其使用现状。 5、列举三类可防治卵菌病害的杀菌剂,并简述其作用机制。 7、简述甲氧基丙烯酸酯类杀菌剂嘧菌酯的特点及其作用机制。 8、简述波尔多液和石硫合剂的防病特点?两者是否可以混合使用?阐述原因。 9、简要分析有机磷类杀菌剂和苯并咪唑类杀菌剂为什么会淡出农药市场?

Similar presentations
:核蛋白体组成成分 转移 RNA ( tRNA ):转运氨基酸 信使 RNA ( mRNA ):蛋白质合成模板 不均一核 RNA ( hnRNA ):成熟 mRNA 的前体 小核 RNA ( snRNA ):>")
 CO.>")












受体拮抗剂 抑制性氨基酸受体受体拮抗剂 神经肽Y受体拮抗剂>")


 Glycometabolism 河北科技大学生工学院 生物化学教研组.>")

