Download presentation
Presentation is loading. Please wait.

1
三級結構: 球蛋白內也可能含有二級結構,三級結構是三度空間的構形,也可以認為絕大部分一級結構中胺基酸的支鍵(side chain)整體交互作用的結果。 X-ray繞射:解出蛋白質固態3D結構的唯一方法。
整體交互作用的結果。 X-ray繞射:解出蛋白質固態3D結構的唯一方法。")
6
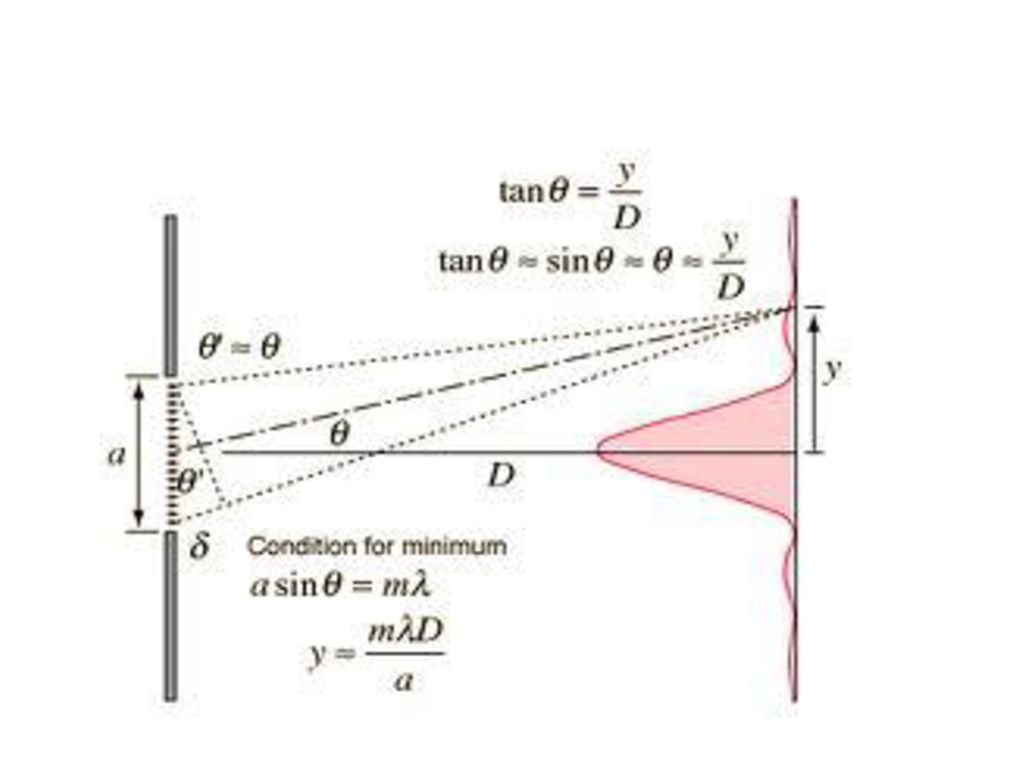
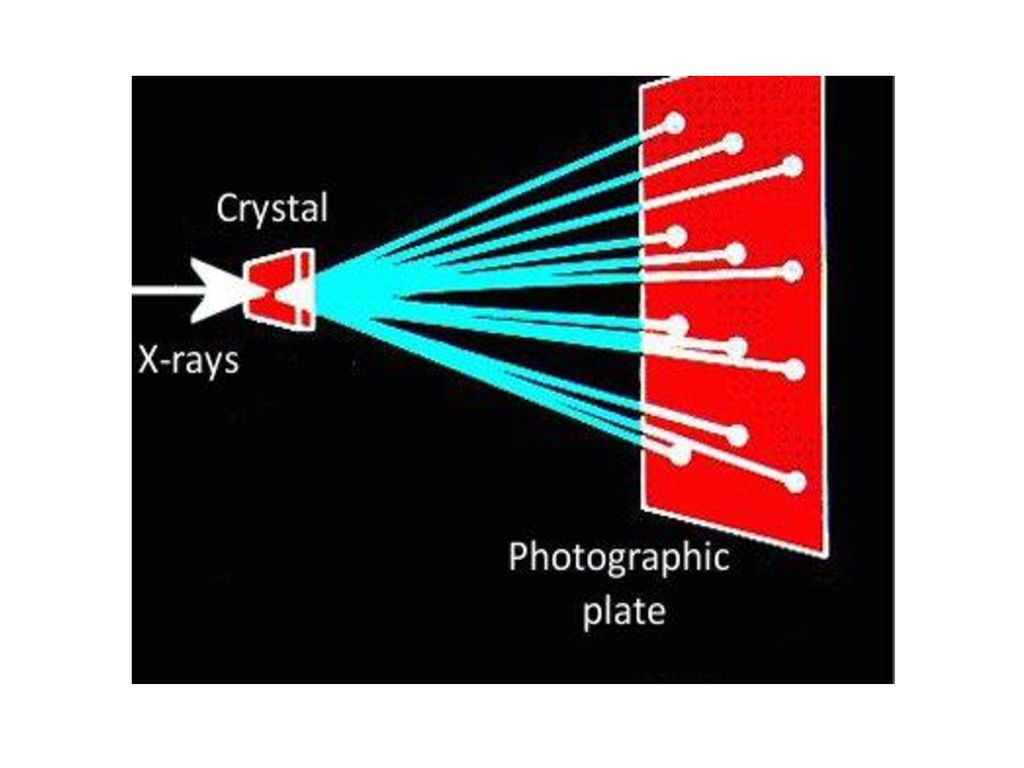
圖5.F X-射線結晶法概要圖

7
BOX 4-5 決定蛋白質三度空間結構之方法 BOX 1-2 FIGURE 1 p.136

8
BOX 4-5(續) 圖1 XX-光結晶法決定抹香鯨肌紅蛋白結構之步驟。(a) 從蛋白質晶體產生X-光繞射模式。(b) 由繞射模式中擷取的資料可用以計算蛋白質之三度空間電子密度圖譜。在此僅顯示結構中血基質之電子密度。(c) 電子密度最高的區域為原子核所在的位置,此資訊可用來拼湊出最終結構。在此,血基質結構模型已根據其電子密度圖譜加以建立。(d) 抹香鯨肌紅蛋白之完整結構,包含血基質在內(PDB ID 2MBW)。 p.136
 從蛋白質晶體產生X-光繞射模式。(b) 由繞射模式中擷取的資料可用以計算蛋白質之三度空間電子密度圖譜。在此僅顯示結構中血基質之電子密度。(c) 電子密度最高的區域為原子核所在的位置,此資訊可用來拼湊出最終結構。在此,血基質結構模型已根據其電子密度圖譜加以建立。(d) 抹香鯨肌紅蛋白之完整結構,包含血基質在內(PDB ID 2MBW)。 p.136.")
9
圖 胰凝乳蛋白(酶)(一種球狀蛋白質)之結構。
(一種球狀蛋白質)之結構。")
10
圖5.16 酵素腺苷酸激酶

11
圖5.16 酵素腺苷酸激酶

12
穩定三級結構的交互作用: 1疏水性的交互作用(hydrophobic interactions): 2靜電作用力: 3氫鍵:
4共價鍵:在蛋白合成時或合成後會形成共價鍵連結來改變polypeptide上的三級結構。影響三級結構最顯著的共價鍵為雙硫鍵(disulfide bridges)。
。")
13
圖5.22 主要的三級結構之交互作用力

14
四級結構: 許多蛋白質是由數個polypeptide chains 組成,每一個polypeptide稱次單元(subunit)。
。")
15
圖5.31 血紅素

16
圖3-23 圖3-23 在蛋白質中結構的層級。圖3-23 在蛋白質中結構的層級。一級結構是由藉由胜肽鍵與雙硫鍵連結在一起的胺基酸序列所組成,而所形成的多肽可以再被編排成二級結構的單元。例如:α 螺旋。此螺旋則是多肽摺疊成三級結構中的一部分,而三級結構則為多次單元蛋白形成四級結構中的一個次單元。此圖例為血紅蛋白。 p.95

17
變性(denaturation): 蛋白結構周遭的影響特別敏感,許多物理與化學試劑會破壞蛋白原來的構型,蛋白質的3D結構破壞稱denaturation(通常不涉及peptide bond的破壞)。
: 蛋白結構周遭的影響特別敏感,許多物理與化學試劑會破壞蛋白原來的構型,蛋白質的3D結構破壞稱denaturation(通常不涉及peptide bond的破壞)。")
18
變性的環境如下: 1強酸或強鹼 2有機溶劑 3清潔劑: 4還原劑: 5鹽濃度: 6重金屬: 7溫度改變:
8機械壓力:攪拌、研磨均可破壞蛋白結構。

19
有一個傳統著名的實例就是核糖核酸酶 A 的變性 與復性,由 Christian Anfinsen 在 1950 年代 完成。
胺基酸序列決定三級結構 某些球狀蛋白質經由加熱、極端 pH值,或變性試 劑的作用後,當環境回復到可使自然構形穩定的 條件時,可重獲其天然結構與生物活性。此過程 稱為復性(renaturation)。 有一個傳統著名的實例就是核糖核酸酶 A 的變性 與復性,由 Christian Anfinsen 在 1950 年代 完成。 p.146
。 有一個傳統著名的實例就是核糖核酸酶 A 的變性 與復性,由 Christian Anfinsen 在 1950 年代 完成。 p.146.")
20
圖4-26 圖4-26 未摺疊、變性的核糖核酸酶之復性。尿素被用來使核糖核酸酶變性,硫氫基乙醇(HOCH2CH2SH)用來還原並打斷雙硫鍵以產生 8 個半胱胺酸殘基。復性過程包含正確雙硫鍵交叉聯結之重建。 之重建。 p.146

21
圖4-27 圖4-27 模擬的摺疊途徑。絨毛蛋白(一種會與肌動蛋白結合,主要發現於小腸微絨毛中的蛋白質)36 個胺基酸殘基經由電腦模擬的摺疊途徑。此摺疊過程實際理論值約為 1 毫秒。 p.146
36 個胺基酸殘基經由電腦模擬的摺疊途徑。此摺疊過程實際理論值約為 1 毫秒。 p.146")
22
有些蛋白質可輔助其他蛋白質之摺疊 並非所有蛋白質在細胞中合成後都會自發性地 進行摺疊。許多蛋白質的摺疊是在特化蛋白質 的作用幫助下達成的。分子伴護(molecular chaperone)這群蛋白質就是負責與部分摺疊 或不正常摺疊的多肽進行交互作用,協助其走 向正確摺疊途徑,或是提供可讓摺疊進行的微 環境。 第一類蛋白質家族稱為 Hsp70。 p.148
這群蛋白質就是負責與部分摺疊 或不正常摺疊的多肽進行交互作用,協助其走 向正確摺疊途徑,或是提供可讓摺疊進行的微 環境。 第一類蛋白質家族稱為 Hsp70。 p.148.")
23
第二類分子伴護稱為伴護蛋白 (chaperonins)。 最後,有些蛋白質的摺疊途徑需要兩種催化 異構化反應的酵素參與:
最後,有些蛋白質的摺疊途徑需要兩種催化 異構化反應的酵素參與: 1. 蛋白質雙硫異構酶(PDI) 2.胜肽脯胺酸順-反異構酶(PPI) p.149
 2.胜肽脯胺酸順-反異構酶(PPI) p.149.")
24
蛋白質摺疊缺陷可能是大範圍的人類基因失調之分子 基礎
蛋白質摺疊缺陷可能是大範圍的人類基因失調之分子 基礎 很多的疾病產生來自常見的蛋白質錯誤摺疊的機制, 包括第二型糖尿病,阿滋海默症、亨丁頓舞蹈症和帕 金森氏症。 在大部分的案例中,一個可溶性蛋白質可在細胞內 被正常地分泌出來,卻在一個錯誤摺疊的狀態下被分 泌而且被轉換成不可溶的胞外澱粉狀(amyloid)蛋白 纖維。 這些疾病被通稱為澱粉樣貯積症(amyloidoses)。 p.149
蛋白 纖維。 這些疾病被通稱為澱粉樣貯積症(amyloidoses)。 p.149.")
25
圖4-31(b) 圖4-31 疾病發生的澱粉狀蛋白纖維形成。 ((b) 在阿滋海默症中,扮演重要的角色的澱粉狀-β 胜肽,是衍生自一個較大的穿膜蛋白,稱為澱粉狀-β 前驅蛋白或是 APP。這個蛋白質被發現存在於大部分的人體組織中。當它還是大型內膜蛋白的一部分時,它是一個帶有二個嵌在膜上的 α-螺旋區段的胜肽。當它外部和內部的區域被特定的蛋白酶(protease)裁切掉時,剩下來而且相當不穩定的澱粉狀-β 胜肽會離開細胞膜並且失去它的α-螺旋構形。然後它會被慢慢地組成至澱粉狀蛋白纖維中。 p.151
 在阿滋海默症中,扮演重要的角色的澱粉狀-β 胜肽,是衍生自一個較大的穿膜蛋白,稱為澱粉狀-β 前驅蛋白或是 APP。這個蛋白質被發現存在於大部分的人體組織中。當它還是大型內膜蛋白的一部分時,它是一個帶有二個嵌在膜上的 α-螺旋區段的胜肽。當它外部和內部的區域被特定的蛋白酶(protease)裁切掉時,剩下來而且相當不穩定的澱粉狀-β 胜肽會離開細胞膜並且失去它的α-螺旋構形。然後它會被慢慢地組成至澱粉狀蛋白纖維中。 p.151.")
26
圖4-31(c) 圖4-31 疾病發生的澱粉狀蛋白纖維形成。。 (c) 然後會促使典型的塊狀物形成在阿滋海默症病人的神經組織外。澱粉狀蛋白富含 β-摺板結構,其 β 股排列會垂直於澱粉狀蛋白纖維的軸線。在澱粉狀-β 胜肽中,它的結構是採一個擴張的雙層平行 β 摺板的形態。其他的可能會採左旋 β-螺旋的形態。 。 p.151
 然後會促使典型的塊狀物形成在阿滋海默症病人的神經組織外。澱粉狀蛋白富含 β-摺板結構,其 β 股排列會垂直於澱粉狀蛋白纖維的軸線。在澱粉狀-β 胜肽中,它的結構是採一個擴張的雙層平行 β 摺板的形態。其他的可能會採左旋 β-螺旋的形態。 。 p.151.")
27
BOX 4-6 摺疊錯誤造成之死亡:狂牛症 圖圖1這這是庫賈氏症患者的腦部皮質染色切片,呈現出海綿狀(液泡)退化性病變,這也是最顯著的神經組織學特徵。淡黃色的液泡位於細胞內,大多出現在神經元突觸的前方與後方。在此切片中液泡的大小變異為直徑 20至 100 μm。 p.152

28
BOX 4-6 圖2人類 PrP 球狀區塊之結構,左圖為單體,右圖為二聚體。第二個次單元體以灰色表示,來突顯當二聚體形成時,第一個次單元體之綠色 α 螺旋(改變為向下翻轉)會產生顯著的構形變化。 p.152
會產生顯著的構形變化。 p.152.")
29
4.3 蛋白質之三級與四級結構 蛋白質中所有原子的整體三度空間排列方式稱之為 蛋白質的三級結構(tertiary structure)。
有些蛋白質包含二或多個相同或不同的獨立多肽鏈 (或次單元)。這些蛋白質次單元在其三度空間複 合體中的排列方式則組成四級結構(quaternary structure)。 在探討這些較高層級的結構時,將蛋白質區分為兩 大類是個有用的作法:即纖維狀蛋白質(fibrous proteins,多肽鏈排列成股狀或片狀)與球狀蛋白質 (globular proteins,多肽鏈摺疊成球狀)。 p.127
。這些蛋白質次單元在其三度空間複 合體中的排列方式則組成四級結構(quaternary structure)。 在探討這些較高層級的結構時,將蛋白質區分為兩 大類是個有用的作法:即纖維狀蛋白質(fibrous proteins,多肽鏈排列成股狀或片狀)與球狀蛋白質 (globular proteins,多肽鏈摺疊成球狀)。 p.127.")
30
α-角質素、膠原蛋白與絲蛋白清楚地描繪出蛋白質結構與其生物功能之間的關係(表 4-2)。
纖維狀蛋白質具有特定結構性功能 α-角質素、膠原蛋白與絲蛋白清楚地描繪出蛋白質結構與其生物功能之間的關係(表 4-2)。 α-角質素 α-角質素已演化成提供力量的蛋白質。 α-角質素的螺旋是右手旋的 α 螺旋,和許多其他蛋白質中的螺旋相同。 p.127
。 α-角質素. α-角質素已演化成提供力量的蛋白質。 α-角質素的螺旋是右手旋的 α 螺旋,和許多其他蛋白質中的螺旋相同。 p.127.")
31
表4-2 p.127

32
α-keratin: 纖維蛋白中,一束束螺旋形的polypeptide被絞成一大束。如頭髮、羊毛、皮膚、角與指甲等皆是一種α-keratin的蛋白質。 這種具規則的α-helix是因為它們缺乏破壞螺旋的胺基酸如proline,但富含促成左螺旋胺基酸如ala、leu、ile、val、met與phe。 而頭髮的捲曲程度,角與指甲的硬度則是以α-keratin內含雙硫鍵的程度決定。

33
圖4-10(a) FIGURE 1-23(b) 圖4-10 毛髮之結構。(a) 毛髮 α-角質素是一種延長的 α-螺旋,在靠近胺基與羧基端處含有稍微粗一點的組成元件。成對的 α-螺旋會以左手旋的方式纏繞成雙鏈狀的纏繞線圈,然後再進一步組合成更高級數的結構,稱為原纖絲與原細纖維。每四條原細纖維(即 32 股 α-角質素)會形成一條中間纖絲。 p.128
 毛髮 α-角質素是一種延長的 α-螺旋,在靠近胺基與羧基端處含有稍微粗一點的組成元件。成對的 α-螺旋會以左手旋的方式纏繞成雙鏈狀的纏繞線圈,然後再進一步組合成更高級數的結構,稱為原纖絲與原細纖維。每四條原細纖維(即 32 股 α-角質素)會形成一條中間纖絲。 p.128.")
34
圖4-10(b) FIGURE 1-23(b) 圖4-10 毛髮之結構。((b) 一根毛髮是由許多 α-角質素纖絲排列組合而成,各種次級結構組成元件如 (a) 中所示 p.128
 FIGURE 1-23(b) 圖4-10 毛髮之結構。((b) 一根毛髮是由許多 α-角質素纖絲排列組合而成,各種次級結構組成元件如 (a) 中所示 p.128")
35
α-角質素中確實富含疏水性胺基酸殘基如:Ala、Val 、Leu、Ile、Met 與 Phe。
在 α-角質素纏繞線圈構造的個別多肽鏈具有相對簡 單的三級結構,這是受到螺旋軸心纏繞成左手超螺 旋的 α-螺旋所支配。 纖維狀蛋白質的強度可藉由多肽鏈間的共價交叉連 結加以強化。 p.128

36
BOX 永久性燙髮是種生化工程 α-角質素的延展特性及含豐富的雙硫鍵便成了燙髮的原理基礎。還原劑可以將雙硫鍵還原成兩個 Cys殘基。濕熱打斷氫鍵因而解開多肽鏈的 α-螺旋結構。經過一段時間後除去還原劑,並且添加氧化劑以建立與先前不同對 Cys 殘基間的新雙硫鍵。 p.129

37
膠原蛋白(Collagen): 脊椎動物最豐富的蛋白,是由聯接組織的細胞所合成,然後再分泌至胞外。膠原蛋白是由三個左旋的polypeptide彼此絞成大束的右旋的纖維。 主要分於牙、骨、皮膚與肌鍵。
: 脊椎動物最豐富的蛋白,是由聯接組織的細胞所合成,然後再分泌至胞外。膠原蛋白是由三個左旋的polypeptide彼此絞成大束的右旋的纖維。 主要分於牙、骨、皮膚與肌鍵。")
38
脊椎動物的膠原蛋白種類繁多。典型的膠原蛋白具有 35% Gly、11% Ala,以及 21% 的 Pro 與4-Hyp。
膠原蛋白中之胺基酸序列呈現重複性三單元(Gly-X- Y),其中 X 常是 Pro、Y則常是 4-Hyp。 唯有 Gly 殘基才能存在於個別 α 鏈間緊密連接處。 膠原蛋白細纖維(圖 4-12)是一種超分子組合,含 有三螺旋膠原蛋白分子(有時候稱為膠原單位分子) 彼此以多種不同方式組合以提供不同程度的張力。 p.129
,其中 X 常是 Pro、Y則常是 4-Hyp。 唯有 Gly 殘基才能存在於個別 α 鏈間緊密連接處。 膠原蛋白細纖維(圖 4-12)是一種超分子組合,含 有三螺旋膠原蛋白分子(有時候稱為膠原單位分子) 彼此以多種不同方式組合以提供不同程度的張力。 p.129.")
39
圖 4-12 膠原蛋白之結構。

40
圖 4-12(c) 3 條此種螺旋(分別為灰色、藍色、紫色)以右手纏繞的方式彼此互相捲繞。
圖 4-12(a) 膠原蛋白之 α 鏈具有獨特的重複性二級結構。此重複性三肽序列為 Gly-X-Pro 或 Gly-X-4-Hyp,形成每圈含 3 個胺基酸殘基之左手螺旋結構。本圖的重複序列是 Gly-Pro-4-Hyp。 圖 4-12(b) 為同一條 α 鏈之空間填充模型。 圖 4-12(c) 3 條此種螺旋(分別為灰色、藍色、紫色)以右手纏繞的方式彼此互相捲繞。 continued
 膠原蛋白之 α 鏈具有獨特的重複性二級結構。此重複性三肽序列為 Gly-X-Pro 或 Gly-X-4-Hyp,形成每圈含 3 個胺基酸殘基之左手螺旋結構。本圖的重複序列是 Gly-Pro-4-Hyp。 圖 4-12(b) 為同一條 α 鏈之空間填充模型。 圖 4-12(c) 3 條此種螺旋(分別為灰色、藍色、紫色)以右手纏繞的方式彼此互相捲繞。 continued.")
41
圖 4-12(d) 從一端觀察膠原蛋白的三股超螺旋構造,並以球-棒模型呈現。Gly 殘基以紅色表示,甘胺酸由於其支鏈最小,最適合存在於三股螺旋交會之緊密接觸位置。同樣的,為了讓大家清楚觀察到三股螺旋中支鏈之內外配置情形,所有球體大小並非以個別原子之實際凡得瓦爾半徑繪製。事實上三股超螺旋之中央並非如此圖所示是空心的,而是非常緊密組裝的。 continued

42
圖4-12 p.132

43
膠原蛋白細纖維之結構。 膠原蛋白(分子量300,000)是一種柱狀分子,長約 3,000Å,厚度則僅有15Å。其三個螺旋糾纏的 α 鏈可能具有不同的胺基酸序列,但每個皆含有約 1,000 個胺基酸殘基。膠原蛋白細纖維則由膠原蛋白分子交錯排列而成,並藉由交叉連結以增加強度
是一種柱狀分子,長約 3,000Å,厚度則僅有15Å。其三個螺旋糾纏的 α 鏈可能具有不同的胺基酸序列,但每個皆含有約 1,000 個胺基酸殘基。膠原蛋白細纖維則由膠原蛋白分子交錯排列而成,並藉由交叉連結以增加強度.")
44
為什麼水手、探險者及大學生應多吃新鮮的水果及蔬菜
BOX 4-3 為什麼水手、探險者及大學生應多吃新鮮的水果及蔬菜 圖1 脯胺酸的 Cγ-endo 構形與 4-羥基脯胺酸的 Cγ-exo 構形。 p.131

45
BOX 4-3 BOX 1-2 FIGURE 1 圖2 脯胺酸 4-羥基酶所催化的反應。(a) 正常反應,與脯胺酸羥化反應相聯結,不需要抗壞血酸參與。從 O2 來的二個氧原子以紅色表示。(b) 未聯結反應,α-酮基戊二酸進行氧化性去羧基化反應,不伴隨脯胺酸之羥化反應。抗壞血酸在此反應過程會隨著轉化為去氫抗壞血酸而消耗掉。 p.131
 正常反應,與脯胺酸羥化反應相聯結,不需要抗壞血酸參與。從 O2 來的二個氧原子以紅色表示。(b) 未聯結反應,α-酮基戊二酸進行氧化性去羧基化反應,不伴隨脯胺酸之羥化反應。抗壞血酸在此反應過程會隨著轉化為去氫抗壞血酸而消耗掉。 p.131.")
46
老化的結締組織中日漸增加的剛性與易碎性就是來自 這些膠原蛋白細纖維中共價鍵結的累積。
膠原蛋白分子的 α 鏈與膠原蛋白細纖維以不尋常的 共價鍵交錯連結:此共價鍵含有Lys 、HyLys或 His 殘基,出現在膠原蛋白中少數 X 與 Y 的位置上。這 些殘基的連結會產生特殊的胺基酸,如:去氫羥基離 胺酸正白胺酸。 老化的結締組織中日漸增加的剛性與易碎性就是來自 這些膠原蛋白細纖維中共價鍵結的累積。 p.129

47
Silk fibroin: 數種昆蟲與蜘蛛會吐絲,這種絲是β-keratin的蛋白纖維組成,polypeptide chains是反向的β-pleated sheets結構。 絲具有強的彈性,這是因為β-pleated sheets結構中,polypeptide chains分子與分子彼此間結合力不強,很容易滑動,此種特性使得絲織品很容彎曲。

48
絲蛋白由昆蟲與蜘蛛製造產生,其多肽鏈主 要是形成 β 構形。
絲蛋白由昆蟲與蜘蛛製造產生,其多肽鏈主 要是形成 β 構形。 絲蛋白富含 Ala 與 Gly殘基,使 β 摺板 得以形成緊密的組裝與 R 基團緊密的排列 (圖 4-13)。 p.132
。 p.132.")
49
圖 4-13(a) BOX 1-2 FIGURE 1 圖4-13 絲之結構。用來製造絲綢或是蜘蛛網的纖維是絲蛋白。(a) 絲蛋白是由一層層富含 Ala 與 Gly 殘基之反平行 β 摺板組合而成,從這張側視圖可以發現這些殘基之較小支鏈可供每層摺板間形成緊密的組裝。 p.133

50
球蛋白(Globular protein)
與氧結合的肌紅蛋白(myoglobin)與血紅蛋白(hemoglobin)是球蛋白研究的好範例。 兩個蛋白皆是hemoprotein,能結合heme。能結合氧分子且具可逆性,但兩個蛋白的生理角色不大一樣。
與血紅蛋白(hemoglobin)是球蛋白研究的好範例。 兩個蛋白皆是hemoprotein,能結合heme。能結合氧分子且具可逆性,但兩個蛋白的生理角色不大一樣。")
51
圖5.29 肌紅蛋白

52
圖5.28 血基質

53
Bohr effect(波爾效應): 氧氣輸送到組織的機制:
在組織細胞中,二氧化碳擴散至血液形成HCO3-與H+,pH值下降,使得H+附在hemoglobin的一些胺基酸的R基上,同時CO2也會結合至一些未帶電荷的胺基酸,增加deoxyHb結構的穩定,增加氧的釋放。

54
圖5.30 血基質的氧結合位由一個摺疊的球蛋白所形成

55
圖5.31 血紅素

56
圖5.32(a)氧化血紅素的三維立體結構
氧化血紅素的三維立體結構")
57
圖5.32(b)去氧化血紅素的三維立體結構
去氧化血紅素的三維立體結構")
58
圖5.33 血紅素異位轉移

59
圖5.34 測量血紅素和肌紅素與氧的親和力之平衡曲線

60
圖5.35 2,3-二磷三甘油酸(2,3-bisphosphoglycerate)(BPG)對於氧與血紅素間親和力的影響
(BPG)對於氧與血紅素間親和力的影響")
61
1在組織中,2,3, Bisphosphoglycerate(BPG)在調節hemoglobin的功能上扮演重要的角色。缺乏BPG時,hemoglobin對氧有非常高的親合力,當H+與CO2增加時BPG結合hemoglobin有助於穩定去氧的hemoglobin(deoxyHb)。 2在肺部情形正好相反,高濃度的氧與hemoglobin趨使deoxyHb 成oxyHb,導致H+、CO2與BPG從oxyHb釋出。H+與HCO3-結合形成carbonic acid(H2CO3),然後分離成CO2與H2O釋出。
,然後分離成CO2與H2O釋出。")
62
次單元間的作用常常受到一些結合的配位基(ligand)影響,在異化作用(allostery)中,經由配位基的結合可以控制蛋白的功能。
一個配位基結合至蛋白一個特異的位置會啟動蛋白構形的改變,這種作用稱allosteric transitions。引起此種作用的配位基稱effectors或modulators。

63
表 3-2 p.86

64
表 3-3 p.86

65
蛋白質毒物 霍亂:簡介歷史 生化恐怖主義及炭疽熱 肉毒桿菌素 蛇毒

66
圖5.A 霍亂毒素的 B-次單位

67
表 5-1 表5-1 p.164

68
蛋白質與配位基間的互補性交互作用: 免疫系統與免疫球蛋白
蛋白質與配位基間的互補性交互作用: 免疫系統與免疫球蛋白 一系列特化的細胞和蛋白質為免疫反應的主要特徵 免疫能力的運作需要多種白血球(leukocytes)的參與,包括巨噬細胞(macrophages)和淋巴球(lymphocytes) 免疫反應是由兩種互補的系統組成:體液免疫和細胞免疫系統。體液免疫系統(humoral immune system,拉丁文 humor 意指「液體」)對付細菌感染和細胞外的病毒(存在於體液中的病毒),和應付侵入生物體內的蛋白質。 p.178
的參與,包括巨噬細胞(macrophages)和淋巴球(lymphocytes) 免疫反應是由兩種互補的系統組成:體液免疫和細胞免疫系統。體液免疫系統(humoral immune system,拉丁文 humor 意指「液體」)對付細菌感染和細胞外的病毒(存在於體液中的病毒),和應付侵入生物體內的蛋白質。 p.178.")
69
細胞免疫系統(cellular immune system)則是破壞被病毒感染的宿主細胞以及寄生蟲與外來的組織。
體液免疫的中心是可溶性蛋白質稱為抗體(antibodies)或免疫球蛋白(immunoglobulins;Ig) 免疫球蛋白占血液蛋白質的 20%,是由 B 淋巴球(B lymphocytes)或稱為 B 細胞(B cells)所製造的。 參與細胞免疫反應的主要細胞是細胞毒性 T 細胞(cytotoxic T cells;TC cells,又稱為殺手 T 細胞),是 T 淋巴球(T lymphocytes)或稱為 T 細胞(Tcells)的一種。 p.178
或免疫球蛋白(immunoglobulins;Ig) 免疫球蛋白占血液蛋白質的 20%,是由 B 淋巴球(B lymphocytes)或稱為 B 細胞(B cells)所製造的。 參與細胞免疫反應的主要細胞是細胞毒性 T 細胞(cytotoxic T cells;TC cells,又稱為殺手 T 細胞),是 T 淋巴球(T lymphocytes)或稱為 T 細胞(Tcells)的一種。 p.178.")
70
分子量小於5,000 的分子通常不具有抗原性。然而,當小分子以共價鍵的方式連接上大的蛋白質,這樣的形式則有可能誘發出免疫反應。
每個抗體或是 T 細胞受體只能與抗原中一特定的分子結構結合,這個部位稱為抗原決定簇(antigenic determinant)或抗原決定位(epitope)。 分子量小於5,000 的分子通常不具有抗原性。然而,當小分子以共價鍵的方式連接上大的蛋白質,這樣的形式則有可能誘發出免疫反應。 此小分子稱為半抗原(haptens)。 p.179
或抗原決定位(epitope)。 分子量小於5,000 的分子通常不具有抗原性。然而,當小分子以共價鍵的方式連接上大的蛋白質,這樣的形式則有可能誘發出免疫反應。 此小分子稱為半抗原(haptens)。 p.179.")
71
表 5-2 TABLE 5-2 p.179

72
免疫球蛋白 G(Immunoglobulin G;IgG)是主要的抗體種類,同時也是血清中含量最多的蛋白質之一。
抗體具有兩個相同的抗原結合部位 免疫球蛋白 G(Immunoglobulin G;IgG)是主要的抗體種類,同時也是血清中含量最多的蛋白質之一。 免疫球蛋白可被木瓜酶切割成兩片段:基底片段稱為 Fc(crystallized fragment),因為通常較容易結晶而得名;兩片分岔片段稱為 Fab(antigenbinding fragments),是抗原結合片段。每一個 Fab 的分岔片段只有一個抗原結合部位。 p.179
是主要的抗體種類,同時也是血清中含量最多的蛋白質之一。 免疫球蛋白可被木瓜酶切割成兩片段:基底片段稱為 Fc(crystallized fragment),因為通常較容易結晶而得名;兩片分岔片段稱為 Fab(antigenbinding fragments),是抗原結合片段。每一個 Fab 的分岔片段只有一個抗原結合部位。 p.179.")
73
圖 5-21 (a) 圖 5-21免疫球蛋白 G (IgG)圖 5-21 免疫球蛋白 G (IgG)。免疫球蛋白 G (IgG)。(a) IgG 是由一對重鏈與一對輕鏈結合成 Y 字型的分子。抗原結合部位是由輕鏈可變區(VL)與重鏈可變區(VH)組合而成。木瓜酶可在扭轉點將 IgG 分割成 Fab 片段與 FC 片段。IgG 在 FC 的部分含有醣類連結〔顯示於 (b)〕。 p.180
圖 5-21 免疫球蛋白 G (IgG)。免疫球蛋白 G (IgG)。(a) IgG 是由一對重鏈與一對輕鏈結合成 Y 字型的分子。抗原結合部位是由輕鏈可變區(VL)與重鏈可變區(VH)組合而成。木瓜酶可在扭轉點將 IgG 分割成 Fab 片段與 FC 片段。IgG 在 FC 的部分含有醣類連結〔顯示於 (b)〕。 p.180.")
74
圖 5-21 (b) 圖 5-21 免疫球蛋白 G (IgG).免疫球蛋白 G (IgG)。免疫球蛋白 G (IgG)。(a) IgG 是由一對重鏈與一對輕鏈結合成 Y 字型的分子。抗原結合部位是由輕鏈可變區(VL)與重鏈可變區(VH)組合而成。木瓜酶可在扭轉點將 IgG 分割成 Fab 片段與 FC 片段。IgG 在 FC 的部分含有醣類連結〔顯示於 (b)〕。 p.180
.免疫球蛋白 G (IgG)。免疫球蛋白 G (IgG)。(a) IgG 是由一對重鏈與一對輕鏈結合成 Y 字型的分子。抗原結合部位是由輕鏈可變區(VL)與重鏈可變區(VH)組合而成。木瓜酶可在扭轉點將 IgG 分割成 Fab 片段與 FC 片段。IgG 在 FC 的部分含有醣類連結〔顯示於 (b)〕。 p.180.")
76
分液收集器

77
3.3 蛋白質的操作與分析 在一種蛋白質的性質與活性能明確決定前,製備出高純度的蛋白質是絕對必須的。蛋白質的來源一般是組織或微生物細胞。
3.3 蛋白質的操作與分析 蛋白質可分離與純化 在一種蛋白質的性質與活性能明確決定前,製備出高純度的蛋白質是絕對必須的。蛋白質的來源一般是組織或微生物細胞。 蛋白質純化的第一步驟就是將這些細胞打破,使其蛋白質釋出至溶液中,此部分即稱為粗萃取物(crudeextract)。 一般粗萃取物會先以基於蛋白質大小或電荷差異為基礎的處理方法加以分離,稱為分劃(fractions)或分部分離(fractionation)。 p.87
。 一般粗萃取物會先以基於蛋白質大小或電荷差異為基礎的處理方法加以分離,稱為分劃(fractions)或分部分離(fractionation)。 p.87.")
78
透析(dialysis)就是一種利用蛋白質大分子性質而將之交換溶劑的方法。
功能最強大的分劃方法是管柱層析法(columnchromatography)。它是利用蛋白質之大小、電荷、結合能力與其他性質之差異加以分離(圖 3-16)。 離子交換層析法(ion-exchange chromatography)是利用在特定的 pH 值下,蛋白質淨電荷之正負與帶電量的差異來進行純化。 管柱基質是一個具有帶電基團的合成聚合物(樹酯);帶陰離子的基團稱為陽離子交換劑(cation exchangers),而帶陽離子的基團稱陰離子交換劑(anion exchangers)。 p.88
。它是利用蛋白質之大小、電荷、結合能力與其他性質之差異加以分離(圖 3-16)。 離子交換層析法(ion-exchange chromatography)是利用在特定的 pH 值下,蛋白質淨電荷之正負與帶電量的差異來進行純化。 管柱基質是一個具有帶電基團的合成聚合物(樹酯);帶陰離子的基團稱為陽離子交換劑(cation exchangers),而帶陽離子的基團稱陰離子交換劑(anion exchangers)。 p.88.")
79
色層分析法原理 層析系統的兩個主要組成為 固定相 (stationary phase) 及 流動相 (mobile phase),二者各有不同的極性或非極性強度;樣本分子因其自身極性的強弱,與此二相之親和力不同。 與固定相親和力大者,易留滯原地; 與流動相親和力大者,易隨流動相移動,因而達成分離的目的。
 及 流動相 (mobile phase),二者各有不同的極性或非極性強度;樣本分子因其自身極性的強弱,與此二相之親和力不同。 與固定相親和力大者,易留滯原地; 與流動相親和力大者,易隨流動相移動,因而達成分離的目的。")
80
在陽離子交換管柱層析法(cation-exchange chromatography)中(圖3-17a),固相基質是帶負電荷基團。
大小-排除層析法(size-exclusion chromatography),也稱為膠體過濾法(gel filtration,圖 3-17b),是利用蛋白質大小之差異進行分離。 親和性層析法(affinity chromatography)則利用蛋白質結合親和力之差異加以分離(圖 3-17c)。 p.90
,也稱為膠體過濾法(gel filtration,圖 3-17b),是利用蛋白質大小之差異進行分離。 親和性層析法(affinity chromatography)則利用蛋白質結合親和力之差異加以分離(圖 3-17c)。 p.90.")
81
膠體過濾法 膠体過濾 屬 partition 層析法,流動相為溶離緩衝液,固定相為膠体。 溶質 (樣本蛋白質) 根據其 分子量 的大小,決定分佈在這兩相的比例。 分子量大的不易進入膠球,隨流動相溶離;分子量小的,則易竄入膠球內的固定相,而被延滯流出膠柱。
 根據其 分子量 的大小,決定分佈在這兩相的比例。 分子量大的不易進入膠球,隨流動相溶離;分子量小的,則易竄入膠球內的固定相,而被延滯流出膠柱。")
83
圖 3-17(b) 圖3-17 蛋白質純化常用的三種管柱層析方法。 。(b) 大小-排除層析法(也稱為膠體過濾法或是分子篩過濾法)是利用蛋白質大小之差異進行分離。 p.89
 圖3-17 蛋白質純化常用的三種管柱層析方法。 。(b) 大小-排除層析法(也稱為膠體過濾法或是分子篩過濾法)是利用蛋白質大小之差異進行分離。 p.89")
84
圖5.D 利用膠體過濾管柱層析來測定分子量

85
離子交換法 乃利用分子的帶電性質進行分離,分離效果好且具多樣性,是重要而應用極廣的純化方法。
是一種 吸附式(adsorption )層析法,流動相為溶離緩衝液,固定相為介質擔体表面的帶電基團。 樣本中各種離子,與介質表面帶電基團間的親和力強弱不同,吸附上去之後,可使用不同離子濃度的緩衝液,分別溶離出這些成分
層析法,流動相為溶離緩衝液,固定相為介質擔体表面的帶電基團。 樣本中各種離子,與介質表面帶電基團間的親和力強弱不同,吸附上去之後,可使用不同離子濃度的緩衝液,分別溶離出這些成分")
87
圖 3-17(a) 圖3-17 蛋白質純化常用的三種管柱層析方法。(a) 離子交換層析法是利用蛋白質在特定 pH 值時之靜電荷差異進行分離。
 圖3-17 蛋白質純化常用的三種管柱層析方法。(a) 離子交換層析法是利用蛋白質在特定 pH 值時之靜電荷差異進行分離。")
88
蛋白質與配位基之間的結合,很少能像上述的公式那樣簡單,其中的交互作用深受蛋白質結構的影響,且結合時通常伴隨著構形的變化。
蛋白質結構影響配位基的結合 蛋白質與配位基之間的結合,很少能像上述的公式那樣簡單,其中的交互作用深受蛋白質結構的影響,且結合時通常伴隨著構形的變化。 舉例來說,當血基質包埋於肌紅蛋白結構中時,血基質與不同配位基之間的專一性會有所改變(圖 5-5a)。 p.164
。 p.164.")
89
親和層析法 (Affinity chromatography)
基本原理 所謂的親和力,乃生物大分子和其配體(ligand)之間形成可逆結合的能力,如酵素和它的受質(substrate)、抗體和抗原、激素和受體(receptor)。親和層析就是根據這種親和力發展出來的純化方法,例如將酶的substrate接在固體支持物上,再用此支持物填裝管柱,當含有這種酶的樣品溶液通過管柱時,酶便被吸附在管柱上,其它的蛋白質及雜質不被吸附,全部從層析柱流出,最後使用適當的緩衝液,將欲分離的酶從柱中洗出來,經過這些步驟便能得到純度高的酶。
之間形成可逆結合的能力,如酵素和它的受質(substrate)、抗體和抗原、激素和受體(receptor)。親和層析就是根據這種親和力發展出來的純化方法,例如將酶的substrate接在固體支持物上,再用此支持物填裝管柱,當含有這種酶的樣品溶液通過管柱時,酶便被吸附在管柱上,其它的蛋白質及雜質不被吸附,全部從層析柱流出,最後使用適當的緩衝液,將欲分離的酶從柱中洗出來,經過這些步驟便能得到純度高的酶。")
90
圖 3-17(c) 圖3-17 蛋白質純化常用的三種管柱層析方法。 (c) 親和性層析法是利用蛋白質與固定相基質上連接之特殊配位基間結合專一性能力之差異進行分離。在內文中將對此方法做進一步詳細介紹。 p.89

91
最新改良的層析法是高效能液相層析法(highperformance liquid chromatography;HPLC)。此方法利用高壓幫浦,搭配填充可抵抗高壓流動下造成之碎裂力的高品質層析介質,以提高蛋白質分子在管柱中移動的速度。 p.90

92
表 3-5 p.91

93
圖3-18(a) FIGURE 1-13 圖3-18 電泳。((a) 將不同樣品注入聚丙烯醯胺膠體上方之樣品槽中,通以電流後蛋白質樣品將進入膠體中。膠體不但將輕微的溫度梯度引起之對流流動降至最小,而且可以確保蛋白質的移動完全是由電場所驅動。 p.91

94
圖3-18(b) FIGURE 1-13 p.91
 FIGURE 1-13 p.91")
95
圖3-18 電泳。 b) 電泳完成後,可利用如Coomassie blue 等呈色劑(與蛋白質結合但不與膠體結合)加以處理,使蛋白質色帶呈色而能以肉眼觀察。每一個色帶均代表一種不同的蛋白質(或蛋白質次單元);小分子蛋白質在膠體中泳動之速度較大分子快,因此會出現於較接近膠體底部處。圖中所示為大腸桿菌 RecA 蛋白(將在第 25 章介紹)的純化。由於 RecA 蛋白的基因已經被選殖於質體中(見第 9 章),因此它的蛋白表現(即蛋白質的合成)是可以被控制的。第一行顯示的為一組標準樣品蛋白(已知分子量)作為分子量標識。下兩行顯示在大腸桿菌誘導合成 RecA 蛋白的前後結果。第四行顯示出在粗細胞萃取物中含有的蛋白。接下來幾行(從左至右)表示蛋白質經過各種純化步驟後的結果,從最右行可以看到最終純化出的蛋白為單一條多肽鏈(分子量約 38,000)。
 電泳完成後,可利用如Coomassie blue 等呈色劑(與蛋白質結合但不與膠體結合)加以處理,使蛋白質色帶呈色而能以肉眼觀察。每一個色帶均代表一種不同的蛋白質(或蛋白質次單元);小分子蛋白質在膠體中泳動之速度較大分子快,因此會出現於較接近膠體底部處。圖中所示為大腸桿菌 RecA 蛋白(將在第 25 章介紹)的純化。由於 RecA 蛋白的基因已經被選殖於質體中(見第 9 章),因此它的蛋白表現(即蛋白質的合成)是可以被控制的。第一行顯示的為一組標準樣品蛋白(已知分子量)作為分子量標識。下兩行顯示在大腸桿菌誘導合成 RecA 蛋白的前後結果。第四行顯示出在粗細胞萃取物中含有的蛋白。接下來幾行(從左至右)表示蛋白質經過各種純化步驟後的結果,從最右行可以看到最終純化出的蛋白為單一條多肽鏈(分子量約 38,000)。")
96
(b) 電泳完成後,可利用如Coomassie blue 等呈色劑(與蛋白質結合但不與膠體結合)加以處理,使蛋白質色帶呈色而能以肉眼觀察。每一個色帶均代表一種不同的蛋白質(或蛋白質次單元);小分子蛋白質在膠體中泳動之速度較大分子快,因此會出現於較接近膠體底部處。圖中所示為大腸桿菌 RecA 蛋白(將在第 25 章介紹)的純化。由於 RecA 蛋白的基因已經被選殖於質體中(見第 9 章),因此它的蛋白表現(即蛋白質的合成)是可以被控制的。第一行顯示的為一組標準樣品蛋白(已知分子量)作為分子量標識。下兩行顯示在大腸桿菌誘導合成 RecA 蛋白的前後結果。第四行顯示出在粗細胞萃取物中含有的蛋白。接下來幾行(從左至右)表示蛋白質經過各種純化步驟後的結果,從最右行可以看到最終純化出的蛋白為單一條多肽鏈(分子量約 38,000)。
 電泳完成後,可利用如Coomassie blue 等呈色劑(與蛋白質結合但不與膠體結合)加以處理,使蛋白質色帶呈色而能以肉眼觀察。每一個色帶均代表一種不同的蛋白質(或蛋白質次單元);小分子蛋白質在膠體中泳動之速度較大分子快,因此會出現於較接近膠體底部處。圖中所示為大腸桿菌 RecA 蛋白(將在第 25 章介紹)的純化。由於 RecA 蛋白的基因已經被選殖於質體中(見第 9 章),因此它的蛋白表現(即蛋白質的合成)是可以被控制的。第一行顯示的為一組標準樣品蛋白(已知分子量)作為分子量標識。下兩行顯示在大腸桿菌誘導合成 RecA 蛋白的前後結果。第四行顯示出在粗細胞萃取物中含有的蛋白。接下來幾行(從左至右)表示蛋白質經過各種純化步驟後的結果,從最右行可以看到最終純化出的蛋白為單一條多肽鏈(分子量約 38,000)。")
97
一般用來概估蛋白質分子量與純度之電泳法會使用到清潔劑十二烷基硫酸鈉(sodium dodecyl sulfate;SDS)。
在與經同一電泳膠體分離之已知分子量蛋白質標準品比較後,任一未知蛋白質均可由其在膠體上所在之位置計算出其概估之分子量(圖 3-19)。 p.92
。 p.92.")
98
SDS-PAGE電泳 分離蛋白質最常用的電法技術
Sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE)
")
99
圖3-19 FIGURE 1-13 p.92

100
圖3-19 估算待測蛋白質之分子量。蛋白質在 SDS 聚丙烯醯胺膠體電泳中之泳動率與其分子量大小有關。(a) 已知分子量之蛋白質標準品經電泳分離如第一行所示,這些樣品蛋白質可用來估算未知蛋白質之分子量(第二行)。(b) 以分子量之對數值對相對電泳泳動率作圖可得到一線性關係,如此即可在圖中讀取未知蛋白質之分子量。
 已知分子量之蛋白質標準品經電泳分離如第一行所示,這些樣品蛋白質可用來估算未知蛋白質之分子量(第二行)。(b) 以分子量之對數值對相對電泳泳動率作圖可得到一線性關係,如此即可在圖中讀取未知蛋白質之分子量。")
101
等電焦集法(isoelectric focusing)是一種用以計算蛋白質等電點(pI)的電泳方法(圖 3-20)。
當置入蛋白質混合物進行電泳分析時,每一種蛋白質均會泳動到恰等於本身等電點之 pH 值所在(表 3-6)。 將等電焦集法與 SDS 電泳組合而成之實驗流程稱為二維電泳(two-dimensional electrophoresis)。此方法用於分析複雜蛋白質混合物時可大幅提高其解析度(圖 3-21)。 p.92
。 將等電焦集法與 SDS 電泳組合而成之實驗流程稱為二維電泳(two-dimensional electrophoresis)。此方法用於分析複雜蛋白質混合物時可大幅提高其解析度(圖 3-21)。 p.92.")
102
圖3-20 FIGURE 1-13 p.93

103
圖3-20 等電焦集法。利用蛋白質之等電點差異進行分離。先添加適當兩性電解質以製備穩定的 pH 梯度之膠體,待測蛋白質混合樣品則置入膠體中之樣品槽,通以電流後各種蛋白質則進入膠體並開始緩慢移動;當移動到與其 pI 值相同之pH 值才停止。記得!當 pH = pI 時,蛋白質的淨電荷為零。

104
表 3-6 p.93

105
圖3-21 FIGURE 1-13 圖3-21 二維電泳。(a) 蛋白質樣品先以柱狀之等電焦集法進行第一次分離,爾後將此柱狀膠體水平置於平板狀膠體上進行 SDS 聚丙烯醯胺膠體電泳分析。完成後所得到之膠體,水平方向是依蛋白質之不同等電點進行分離,垂直方向則依蛋白質分子量大小差異進行分離。(b) 以二維電泳技術可以解析出超過 1,000 種大腸桿菌中之蛋白質。 p.93
 蛋白質樣品先以柱狀之等電焦集法進行第一次分離,爾後將此柱狀膠體水平置於平板狀膠體上進行 SDS 聚丙烯醯胺膠體電泳分析。完成後所得到之膠體,水平方向是依蛋白質之不同等電點進行分離,垂直方向則依蛋白質分子量大小差異進行分離。(b) 以二維電泳技術可以解析出超過 1,000 種大腸桿菌中之蛋白質。 p.93.")
106
圖3-21 二維電泳。(a) 蛋白質樣品先以柱狀之等電焦集法進行第一次分離,爾後將此柱狀膠體水平置於平板狀膠體上進行 SDS 聚丙烯醯胺膠體電泳分析。完成後所得到之膠體,水平方向是依蛋白質之不同等電點進行分離,垂直方向則依蛋白質分子量大小差異進行分離。(b) 以二維電泳技術可以解析出超過 1,000 種大腸桿菌中之蛋白質。
 蛋白質樣品先以柱狀之等電焦集法進行第一次分離,爾後將此柱狀膠體水平置於平板狀膠體上進行 SDS 聚丙烯醯胺膠體電泳分析。完成後所得到之膠體,水平方向是依蛋白質之不同等電點進行分離,垂直方向則依蛋白質分子量大小差異進行分離。(b) 以二維電泳技術可以解析出超過 1,000 種大腸桿菌中之蛋白質。")
107
BOX 3-1 分子 吸收光:比爾定律 當結晶物質溶解使得熵增加
許多生物分子會在特定波長吸收光,如色胺酸會在波長 280 nm 吸收光(圖 3-6)。 在特定波長下,入射光被溶液吸收的比例會與吸收層的厚度(即光徑長)與吸光物種之濃度有關(圖 1)。這兩種關係可組合為比爾定律: log(I0/I)所表示的即為吸光率(absorbance),命名為 A。 莫耳消光係數會因吸光化合物之本質、溶劑、波長,甚至 pH 值(若吸光物種與吸光性質不同的另一種游離態達平衡時)而異。 p.79
。 在特定波長下,入射光被溶液吸收的比例會與吸收層的厚度(即光徑長)與吸光物種之濃度有關(圖 1)。這兩種關係可組合為比爾定律: log(I0/I)所表示的即為吸光率(absorbance),命名為 A。 莫耳消光係數會因吸光化合物之本質、溶劑、波長,甚至 pH 值(若吸光物種與吸光性質不同的另一種游離態達平衡時)而異。 p.79.")
108
BOX 1-2 FIGURE 1 BOX 3-1 分子 吸收光:比爾定律 (續)
圖1 分光光度計之主要構成要素。光源發出涵蓋廣泛光譜之光,單色儀選擇並傳導特定波長之光。單色光通過光徑長為 l 之比色管中的樣品溶液,並以與吸光物種濃度成比例之方式被樣品吸收。穿透光之強度則可由偵測器測得。 p.79

Similar presentations















 12 (表面因子) 7 (促凝血酶原激酶原) (3,因子 III) 11>")





>")
>")

